sea-surface temperatures and palaeoenvironments of dolichosaurs and early mosasaurs
TRANSCRIPT

Introduction
The aim of the present paper is to provide a comparativeanalysis of the depositional environments in which some earlybasal mosasaurs and dolichosaurs are found, specifically ‘EinYabrud in the Middle East and localities in the Eagle Ford Shalein Texas (Fig. 1). Comparing these sites, which occur in differentregional geological settings and in distinct depositionalenvironments, elucidates and constrains ecological parametersin the origin and distribution of mosasaurs and dolichosaurs.The influence of sea-surface temperature as a factor controllinghabitat range of modern marine lizards and snakes, and theobserved distribution of fossil small-bodied marine squamatesin the context of sea-surface temperature models for theCenomanian and Turonian stages are then compared.
In the Dallas-Fort Worth area (north-central Texas), thefossils on which this study focuses are Dallasaurus turneri,
Russellosaurus coheni and specimens of Coniasaurus crassidens(B.A. Bell et al., 1982; G.L. Bell & Polcyn, 2005; Polcyn & G.L.Bell, 2005) in the Shuler Museum of Paleontology at SouthernMethodist University (SMU). At ‘Ein Yabrud, we are most con-cerned with Haasiasaurus gittelmani (Polcyn et al., 1999, 2003)and the legged snakes Haasiophis terrasanctus (Tchernov et al.,2000) and Pachyrhachis problematicus (Polcyn et al., 2005).Haasiasaurus is a basal mosasaur (Polcyn & Bell, 2005).Coniasaurus is a dolichosaur and is a likely close sister tomosasaurs (Haber & Polcyn, 2005). Dolichosaurs, like mosasaurs,were marine. Dolichosaurs and early mosasaurs are knownfrom the Western Interior Seaway of North America to theeastern Mediterranean region during the Cenomanian andTuronian stages (Jacobs et al., 2005). Haasiasaurus at 98 Mais the oldest known well-documented mosasaur.
Plesiopedal mosasaurs such as Haasiasaurus, which bear anessentially terrestrial limb condition, appeared around
Sea-surface temperatures and palaeoenvironments of dolichosaurs and early mosasaurs
L.L. Jacobs1,*, M.J. Polcyn1, L.H. Taylor2 & K. Ferguson1
1 Department of Geological Sciences, Southern Methodist University, Dallas, Texas 75275, USA.
2 Department of Earth Sciences, Denver Museum of Nature and Science, 2001 Colorado Boulevard, Denver, Colorado 80205-5798, USA.
* Corresponding author. Email: [email protected]
Manuscript received: December 2004; accepted: May 2005
Abstract
The north-central Texas Cretaceous section and its contained fossils, as compared to sections at ‘Ein Yabrud in the eastern Mediterranean region,
demonstrate that dolichosaurs and primitive mosasaurs inhabited marine environments at least from the intertidal zone to <100 m in depth. The
small body size of dolichosaurs and primitive mosasaurs, and the association of Haasiasaurus with marine snakes at ‘Ein Yabrud, suggest similar
temperature requirements to modern sea snakes and marine iguanas (Amblyrhynchus cristatus), specifically sea-surface temperatures between
approximately 20° and 30° C. The Cenomanian and Turonian stages are characterised by widespread shallow seas with relatively high sea-surface
temperatures extending to at least 45° N and 65° S latitude. The distribution of dolichosaurs, mosasaurs and snakes during this interval contradicts
palaeoclimate models that predict high (>30° C) equatorial sea-surface temperatures and a steep latitudinal temperature gradient in the Northern
Hemisphere.
Keywords: Cretaceous, mosasaurs, Middle East, sea snakes, Texas
269Netherlands Journal of Geosciences — Geologie en Mijnbouw | 84 – 3 | 2005
Netherlands Journal of Geosciences — Geologie en Mijnbouw | 84 – 3 | 269 - 281 | 2005

northeastern Gondwana as it fragmented in the Cenomanianagainst the backdrop of rising sea levels and relatively highsea-surface temperatures. They are represented in North Americasoon after under the same general sea level and temperatureregimes. Therefore, it is useful to evaluate the role of habitatunder these environmental constraints as contributing factorsto the early evolution of the group. The later record ofmosasaurs spans the entire Upper Cretaceous when they exhibita broad range of adaptations and generally larger adult bodysize that reflects progressive optimisation towards a fullymarine existence. Derived mosasaurs, best known from theConiacian through the Maastrichtian stages, can be dividedinto four distinct clades, the Mosasaurinae, Tylosaurinae,Plioplatecarpinae and Halisauromorpha. By the mid-Turonian
(92 Ma), basal members of the four clades were already presentand thus imply some level of selection was at play in the earlyevolution of these groups prior to 92 Ma (G.L. Bell & Polcyn,2005; Polcyn & G.L. Bell, 2005).
Material and methods
We collected lithologic specimens and isotopic samples inTexas in conjunction with the stratigraphic study presentedby Jacobs et al. (2005), which provides the geological contextand chronological control for this study. During fieldwork at‘Ein Yabrud in December 1999 we recorded elevations and geo-graphic locations using GPS instrumentation. Locality data areon file at the Shuler Museum of Paleontology (SMU). Lithologicsamples were examined using thin section petrography andscanning electron microscopy. Descriptions are provided on-linein ‘Supplemental material’. Stable isotope determinations followmethods presented in on-line supplemental material thataccompanies Jacobs et al. (2005). Energy dispersive X-rayanalyses were performed on some concretions in the SMUElectron Microscopy Laboratory.
The carbonate classification system used here is fromDunham (1962). Although this system has been modifiedslightly since its origin (Embry & Klovan, 1971), the funda-mental terminology remains unchanged. It consists of fourmajor categories defined by a decrease in lime mud content,and therefore, interpretively by a corresponding increase inenergy level: mudstone, wackestone, packstone and grainstone.Dunham’s (1962) term ‘mudstone’ refers to what Folk (1962)and others referred to as ‘micrite’. In this context, the termmudstone has no clastic connotation and does not refer toshale content; it is composed entirely of carbonate.
Results
Depositional environment of the Eagle Ford Shale in north-central Texas
The Eagle Ford Shale (Jacobs et al., 2005; Polcyn & Bell, 2005,fig. 1) has been recognised as a formation or alternatively asa group, but its hierarchical level is of little concern here. TheEagle Ford Shale can be divided into the basal sandy TarrantMember, overlain by the Britton Shale, followed by the KampRanch Limestone, and culminated by the Arcadia Park Shale.The top of the Eagle Ford is at the base of the Austin Chalkwhere it unconformably overlies the Arcadia Park Shale of theEagle Ford. The unconformity is represented by a hardground.Within the Britton and the Arcadia Park, occasional sandstonestringers and detrital limestones composed of Inoceramusprisms occur; however, these pinch out and cannot be recog-nised across the study area. They should not be confused withthe Kamp Ranch Limestone, which is also a detrital limestone,as will be discussed below. Septarian nodules are common in
Netherlands Journal of Geosciences — Geologie en Mijnbouw | 84 – 3 | 2005270
Fig. 1. Cenomanian and Turonian dolichosaur and mosasaur localities.
Southern North Sea Basin: 1: England and France (Anglo-Paris Basin;
Rage, 1989; Caldwell, 1999; Caldwell & Cooper, 1999; 2: NW Germany
(Diedrich, 1997, 1999). Eastern Mediterranean: 3: Slovenia (Carroll &
DeBraga, 1992); 4: Hvar and Lesina, Croatia (Carroll & De Braga, 1992);
5: Hajula, Hakel and Al Nammoura, Lebanon (Dal Sasso & Renesto, 1999;
Dalla Vecchia & Venturini, 1999); 6: ‘Ein Yabrud (Polcyn et al., 1999);
7: Besokty II, Kazakhstan (Averianov, 2001); 8: Goulmima, Morocco
(Bardet et al., 2003); 9: Iembi, Angola (Antunes, 1964); 10: Yagua,
Columbia (Páramo, 1994). USA: 11: Big Bend, Texas (Bell & VonLoh,
1998); 12: Dallas area (Jacobs et al., 2005); 13: Kansas; 14: South
Dakota and Wyoming (Bell & VonLoh, 1998; VonLoh & Bell, 1998). Buchy
et al. (2005) report a plesiopedal mosasaur from Nuevo León, Mexico,
near Big Bend. For a discussion of the age and correlation of these
localities see Jacobs et al. (2005). Map after Hay et al. (1999).
the Eagle Ford, often forming around ammonites. Sideriteconcretions in the Britton Shale form irregular layers containingsome complete and many fragmentary fossils. The concretionspreserve original lamination in slightly compacted mudstonewith discontinuous stylolites representing insoluble residuesresulting from pressure dissolution. The internal structure ofthe siderite concretions does not present a series of concentriclayers about a nucleus, but rather the siderite concretionsappear to have formed soon after, perhaps nearly simultaneouswith mud deposition. δ13C analysis (Table 1) of one sideriteconcretion yielded a value of 3.3‰, which, if reflective oforiginal depositional composition, is consistent with expectedCenomanian values of pelagic marine carbonates (Jacobs etal., 2005, fig. 2).
Britton and Arcadia Park shales represent deposits laid downin a shallow-water environment, below wave base. Some biotur-bation is present, including Ophiomorpha burrows, indicatingactivity in normal marine water well within the photic zone.The low δ18O value of the Britton siderite concretion, if primary,might be accounted for by the influx of meteoric water into ashallow sea, although salinity must have remained within theappropriate range to support the marine fauna. Most of thefossil fragments in the Britton and Arcadia Park are clastsindicative of origin in relatively high energy, and because theyare deposited in shale, they represent higher-energy materialtransported to a lower-energy environment.
Based on the facies relationships between the WoodbineFormation and the Eagle Ford Shale in the western part of thestudy area, the proximity of the shoreline to relevant outcropsof the Eagle Ford at least in the lower part of the section is inthe order of kilometres. Outcrops containing the Acanthocerasamphibolum Zone in the Tarrant Member of the Eagle Ford onthe seaward side (now in Tarrant County), and an occurrencein the Lewisville shelf-strandplain system of the WoodbineFormation (Kennedy & Cobban, 1990) in Grayson County,
presents a time line. These localities are separated by ~160 kmin a line running close to and approximating the western north-south shoreline in this region. During deposition of the upperportion of the Eagle Ford Shale the northern shoreline remainedconstrained by the Ouachita Uplift but the western shoreline
271Netherlands Journal of Geosciences — Geologie en Mijnbouw | 84 – 3 | 2005
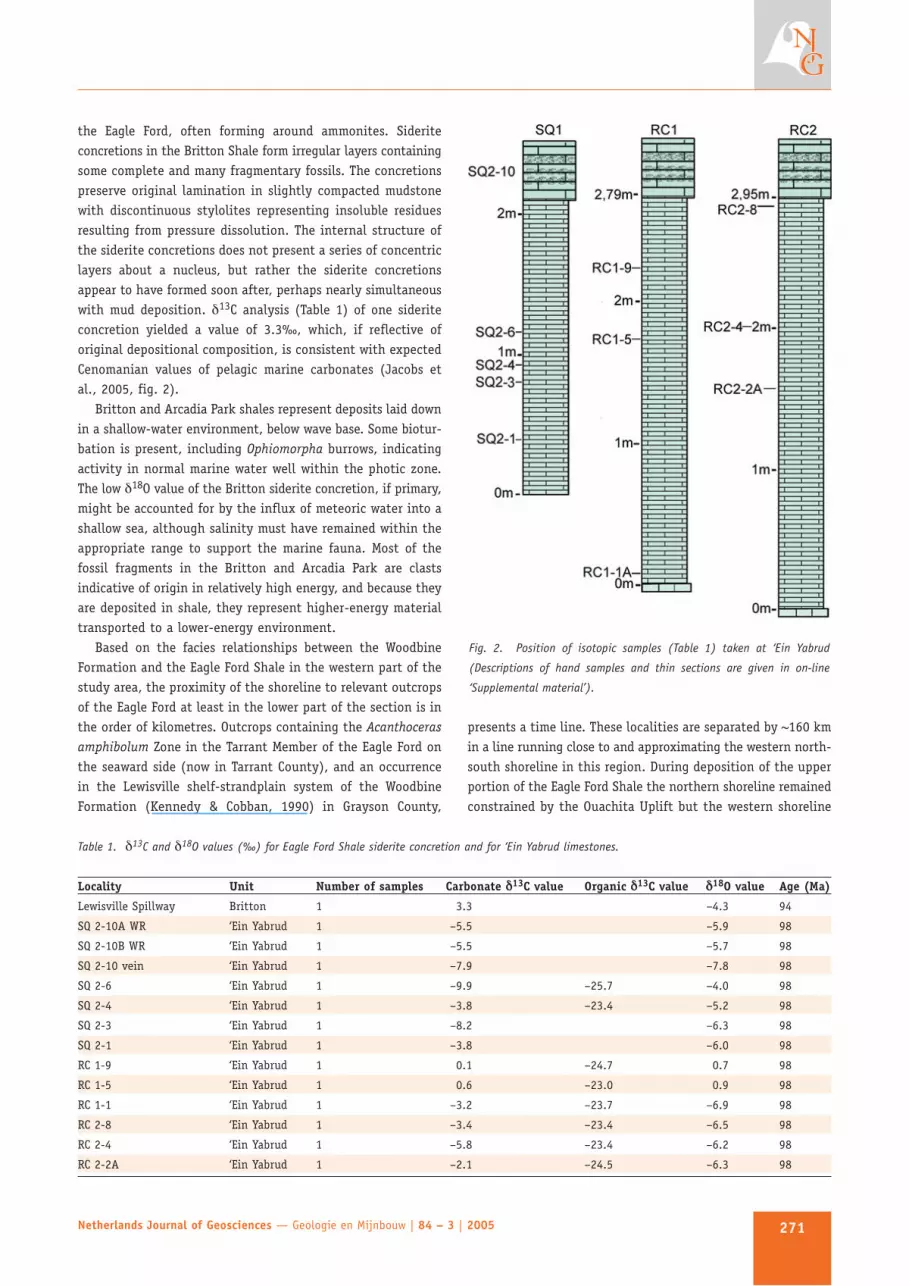
Fig. 2. Position of isotopic samples (Table 1) taken at ‘Ein Yabrud
(Descriptions of hand samples and thin sections are given in on-line
‘Supplemental material’).
Table 1. δ13C and δ18O values (‰) for Eagle Ford Shale siderite concretion and for ‘Ein Yabrud limestones.
Locality Unit Number of samples Carbonate δδ13C value Organic δδ13C value δδ18O value Age (Ma)
Lewisville Spillway Britton 1 3.3 –4.3 94
SQ 2-10A WR ‘Ein Yabrud 1 –5.5 –5.9 98
SQ 2-10B WR ‘Ein Yabrud 1 –5.5 –5.7 98
SQ 2-10 vein ‘Ein Yabrud 1 –7.9 –7.8 98
SQ 2-6 ‘Ein Yabrud 1 –9.9 –25.7 –4.0 98
SQ 2-4 ‘Ein Yabrud 1 –3.8 –23.4 –5.2 98
SQ 2-3 ‘Ein Yabrud 1 –8.2 –6.3 98
SQ 2-1 ‘Ein Yabrud 1 –3.8 –6.0 98
RC 1-9 ‘Ein Yabrud 1 0.1 –24.7 0.7 98
RC 1-5 ‘Ein Yabrud 1 0.6 –23.0 0.9 98
RC 1-1 ‘Ein Yabrud 1 –3.2 –23.7 –6.9 98
RC 2-8 ‘Ein Yabrud 1 –3.4 –23.4 –6.5 98
RC 2-4 ‘Ein Yabrud 1 –5.8 –23.4 –6.2 98
RC 2-2A ‘Ein Yabrud 1 –2.1 –24.5 –6.3 98

is less tightly constrained. Published shoreline reconstructionsof the Western Interior Seaway in the study area are imprecisebecause the relevant sedimentary units are missing andpresumed eroded away from northwestern Texas and thePanhandle (Elder & Kirkland, 1994). However, Sageman &Arthur (1994) presented a palaeobathymetric map of theWestern Interior Seaway based primarily upon sedimentaryfeatures. The study area is represented on their map as being‘proximal offshore noncalcareous mudrocks’, which at the scaleof the map is correct. The depth at which that facies wasdeposited is reported to be 50 - 100 m.
The most distinctive unit within the Eagle Ford Shale is theKamp Ranch Limestone. It is a detrital limestone sandwichedbetween the Arcadia Park Shale and the Britton Shale. TheKamp Ranch is important in this study because it serves as astratigraphic marker, contains both fragmentary coniasaurs andmosasaurs, has implications for sea level, and represents a dis-crete and small amount of time confined to the Collignoniceraswoollgari Zone. We traced the Kamp Ranch Limestone over adistance of ~90 km, north to south (Denton, Dallas and Elliscounties), in the study area, correlating isolated outcrops bylithology and by consistent homotaxial position approximately30 metres below the Eagle Ford-Austin contact. Betweennorthern and southern outcrops, a water well log from theSouthern Methodist University campus reported by Shuler(1918, prior to the naming of the Kamp Ranch) shows a metre-thick ‘lime rock’ at the appropriate level to be the Kamp RanchLimestone.
The Kamp Ranch Limestone usually is no more than a metrethick. Its base is in sharp but irregular and bioturbatedcontact with the underlying Britton Shale. The upper contactwith the Arcadia Park is sometimes sharp and bioturbated, butoften shows an increasing percentage of shale stringers versusthin limestone beds. Cross bedding is associated with asym-metrical ripple marks, which trend in diverse directions andsometimes interfere with each other (Reid, 1952).
Kamp Ranch lithology is consistent across the study areaalthough the internal architecture varies somewhat and thereis a range in the abundance of larger shell clasts, as opposedto Inoceramus laths, from place to place. In general, the KampRanch Limestone consists of a series of thin flaggy packstonesto grainstones interbedded with shale stringers. Thin stylolyticmudstone beds are present. The stylolytic mudstone wasdeposited in a low-energy environment. The overwhelminglypredominant clasts in the Kamp Ranch Limestone are calcitelaths derived from Inoceramus shells (85 - 89%; Reid, 1952).These Inoceramus prisms are in general not rounded and donot appear to show a consistent preferred orientation.
The samples of Kamp Ranch analysed by Reid (1952) showedonly 1 - 3% quartz. Collignoniceras woollgari is abundant asimpressions parallel to bedding planes with the aragonitic shellmaterial almost always completely dissolved. Minor amountsof glauconite (as identified in this study), pyrite, biotite
and limonite are present. Glauconite peloids may representgastropod faecal material. Fish bone fragments and shark teethcan be quite common and concentrated on bedding surfaces,indicative of winnowing. Carbonized wood fragments, the largest(>1 cm) of which are rounded, are present within the KampRanch. δ13C analyses of wood from the Kamp Ranch arepresented in Jacobs et al. (2005).
As the Kamp Ranch Limestone is primarily a detrital lime-stone unit between two siliciclastic shales, it clearly representsa change in depositional regime from strata above and belowit. Detrital Inoceramus calcite prisms occur in poorly sortedcross-bedded layers, indicating movement in a higher-energyenvironment than the shale. In that higher-energy environ-ment, and probably with the aid of biological agents prior tobeing subjected to higher-energy currents, Inoceramus shellswere reduced to small fragments, most to their individualcalcite laths that make up the shell. As the currents waned,the Inoceramus clasts that formed the Kamp Ranch Limestonecame to rest in a slightly lower-energy environment than thatin which they were derived. Because the laths are not signifi-cantly rounded, however, they were likely derived from arelatively close source. The low percentage of quartz indicatesreduced terrigenous input and an offshore source area for theinoceramids. Thin micritic mudstone beds associated withpoorly sorted Inoceramus packstone indicate fluctuations indepositional energy. The uniform thickness of the Kamp Ranch,its regional extent, and its continuity suggest that it is not awedge deposit. Nor is it a turbidite or other deep-water deposit.Its proximity to shore and lack of hardground features suggestlowstand rather than highstand deposition. However, thepresence of pre-existing fauna and lack of reef- or mound-building organisms suggest that the Kamp Ranch deposits werenot a response to shoaling.
Because Inoceramus-prism limestones similar to the KampRanch but thinner and less continuous are found throughoutthe Eagle Ford, the processes that produced such beds musthave been recurrent. Higher-energy deposits coupled with low-energy mudstones and with shale stringers, poor sorting,reduced terrigenous input, comminuted shells and currentbedding, all taken together, suggest storm deposits. Acceptingthis, the lateral persistence and greater thickness of the KampRanch compared to other Inoceramus-prism limestones in theEagle Ford would suggest storms either of greater magnitudeor greater frequency than those occurring during other intervalsof Eagle Ford deposition. We have no reason to suspect thateither was the case. Therefore, a minor drop in sea level, aswas presented by Haq et al. (1988) in their sea level curve, inwhich storm events of normal size for the Turonian were moreconsistently recorded in the Eagle Ford sedimentary record,seems more plausible. We interpret the Kamp Ranch Limestoneas storm deposits formed during a short period of lowered sealevel on the shelf of the southeastern part of the WesternInterior Seaway. This lowering of sea level occurred at ~92 Ma,
Netherlands Journal of Geosciences — Geologie en Mijnbouw | 84 – 3 | 2005272

during deposition of the Collignoniceras woollgari Zone, duringan interval when Coniasaurus had already inhabited the regionfor 4 million years (Jacobs et al., 2005), and when earlymosasaurs have their first records in North America.
We disagree with the assessment of Hancock & Walaszczyk(2004) that the Kamp Ranch Limestone was deposited duringa highstand. The Kamp Ranch is not a hardground as would beexpected in such a case. In fact, a hardground is present atthe top of the Eagle Ford below the Austin Chalk. Moreover,while a carbonate hardground often has fossils preserved ingrowth position, the Kamp Ranch Limestone consists, in largepart, of shell prisms deposited as we have described. It lackscomplete fossils in growth position. Henderson (2004) discussedInoceramus coquinas in the Albian Toolebuc Formation inAustralia, which he hypothesised to have formed in responseto fluctuating bacterial productivity on a deep, anoxic seafloor. Even if the hypothesis is correct in that case, there aresubstantial differences between the geological contexts andoccurrences of the Kamp Ranch Limestone and the Toolebuccoquinas indicating that the Toolebuc model does not work forthe Kamp Ranch.
Depositional environment of ‘Ein Yabrud in the eastern Mediterranean region
In the Middle East, in the West Bank, the locality of ‘Ein Yabrud(Fig. 1) produced, among other spectacular vertebrates, theplesiopedal mosasaur Haasiasaurus gittelmani (Polcyn et al.,1999, 2003). The locality was originally placed in the Bet MeirFormation (Chalifa & Tchernov, 1982). Later, Chalifa (1985)placed it in the Amminadav Formation. Lewy (1990) consideredthe Bet Meir and Moza formations to be the lower unit of whathe termed a biosedimentary cycle; the Amminadav Formationis the upper unit of the cycle. The lower unit of a biosedimentarycycle, according to Lewy (1990), is more variable lithologicallyand more fossiliferous than the upper unit. In the case of ‘EinYabrud, the sediment resembles dolostone of the AmminadavFormation, but it is of limited extent and may well be a localfacies of the lower unit (Z. Lewy, pers. comm.), which wouldplace it in the Bet Meir Formation. This is not an issue that wecan resolve in this paper, nor is it fundamental to our purpose.However, in a regional sense, the distribution of facies isimportant for understanding ‘Ein Yabrud and other Cenomanianand Turonian localities of the eastern Mediterranean. The age of‘Ein Yabrud is early Cenomanian (~98 Ma; Jacobs et al., 2005).
The vertebrate-bearing unit in the area of ‘Ein Yabrud isexposed in a series of small, now abandoned, flagstone quarriesand in a road cut for Highway 449, approximately 20 km northof Jerusalem. We did not visit all quarries, but did measuretwo sections in the road cut and one in the abandoned quarry(now used as a rubbish pit) to the east of the road. The thick-ness of the ‘Ein Yabrud unit as measured in section RC 2, whichhas both the base and the top exposed, is 2.95 m (Fig. 2).
Little is known about the provenance of the original fossilcollections; however, one of the famous ‘Ein Yabrud limbedsnakes is said to have come from the quarry where we measureda section (SQ section in Fig. 2; E. Tchernov, pers. comm.). Wecollected the fish Serriventor sp. at 1 m below the top of theunit as measured in SQ section and an unidentified fish 97 cmabove the base of the unit in section RC 2. A fish vertebralfragment was found in sample SQ 2-1 approximately 55 cmbelow the Serriventor.
The two fish mentioned above, to our knowledge, are theonly reasonably complete vertebrate specimens that can betied to a precise geographic and stratigraphic position. Theoriginal collections were amassed by Georg Haas of The HebrewUniversity of Jerusalem through the purchase of specimensfrom quarry workers. While there is no reason to question thatearlier collections came from the ‘Ein Yabrud quarries and fromthis specific 3 m unit, taphonomic and stratigraphic informationregarding associations and successions of specimens can haveno greater resolution.
Several hundred specimens from ‘Ein Yabrud are cataloguedinto the collections of The Hebrew University of Jerusalem.Macrofossils are rare, but diverse and extremely well preserved.Soft tissue is evident in some specimens, especially sharks andrays. Marine plants are represented by calcareous algae, andterrestrial plants are represented by a gymnosperm (E. Tchernov,pers. comm.). Invertebrates include ammonites, echinoids,starfish, bivalves and crustaceans, but none is abundant. Themost abundant vertebrates are actinopterygian fish (Gayet,1980; Chalifa, 1985, 1989a - c). The vidalamiine amiid Pachyamialatimaxillaris (Chalifa & Tchernov, 1982) may have beenanadromous (Grande & Bemis, 1998).
Amniotes are rare, some more so than others, but includepelomedusid turtles (Haas, 1978a, b; de Broin & Werner, 1998),a mosasaur (Polcyn et al., 1999), snakes (Tchernov et al., 2000;Polcyn et al., 2005) and a pterosaur. As a group, pelomedusidshave a record suggesting that while they are primarily fresh-water they ventured into saltwater environments (Moody,1997). Haasiasaurus gittelmani, as a plesiopedal mosasaur, isnot eliminated from being totally or mostly marine, althoughits ability for terrestrial progression is not eliminated eitherand, in our opinion, is likely. Every snake can swim, but onlya few are adapted for a fully marine existence.
Of the two snake species (known from a total of threespecimens) from ‘Ein Yabrud, both have osteologically complete,albeit small, hind legs, the function of which is unknown. Inour opinion, the diminutive legs are most likely for intraspe-cific behaviour and certainly not for locomotion. Haasiophisterrasanctus has no obvious adaptations for aquatic or marinelife. Pachyrhachis problematicus, a second species of ‘Ein Yabrudsnake, has pachyostotic ribs, which have been interpreted asan adaptation for marine life. No living marine snake haspachyostotic bones. However, a tooth plate of a marine pyc-nodont fish was found within the ribcage of Pachyrhachis and
273Netherlands Journal of Geosciences — Geologie en Mijnbouw | 84 – 3 | 2005
was thought to represent stomach contents (Haas, 1979). Ifeither of those interpretations is correct, then it appears thatPachyrhachis, at least, was a marine snake.
The only unquestionably obligate terrestrial vertebrateknown from ‘Ein Yabrud is a pterosaur (recognising that volantforms are included among terrestrial vertebrates), butpterosaurs are known to soar over the sea as well as land.Nothing in the vertebrates would suggest an adaptation tohypersaline conditions but several of the taxa, including theelasmobranchs, the amiid, the pelomedusid and the squamatesmight have been tolerant of a range of salinity from marine tofreshwater conditions. Extant sea snakes are more tolerant offreshwater than they appear to be of hypersaline waters(Dunson & Ehlert, 1971).
The ‘Ein Yabrud fossiliferous unit consists of thinly laminatedmudstones. It has a sharp contact with an underlying massivemudstone unit. It is overlain by a massive mudstone similar tothat underlying it. Algal laminae are common. Birds-eyestructures indicate dewatering of the lime mud and the lack ofevaporites precludes supratidal deposition. Diagenesis ofsamples includes sparry calcite in voids and fractures andminor dolomitisation. Some, but not all, thin sections revealrecrystallisation. A symmetrically ripple-marked hand samplewas found as float in the Snake Quarry. Although we do notknow its exact provenance, it appears most likely that it wasderived from the ‘Ein Yabrud unit because the bottom of thesection is not exposed in the quarry, the top of the section istruncated above the unit, and the overlying unit is massivelimestone. Symmetrical ripples indicate oscillating currentflow, such as occurs in an intertidal environment.
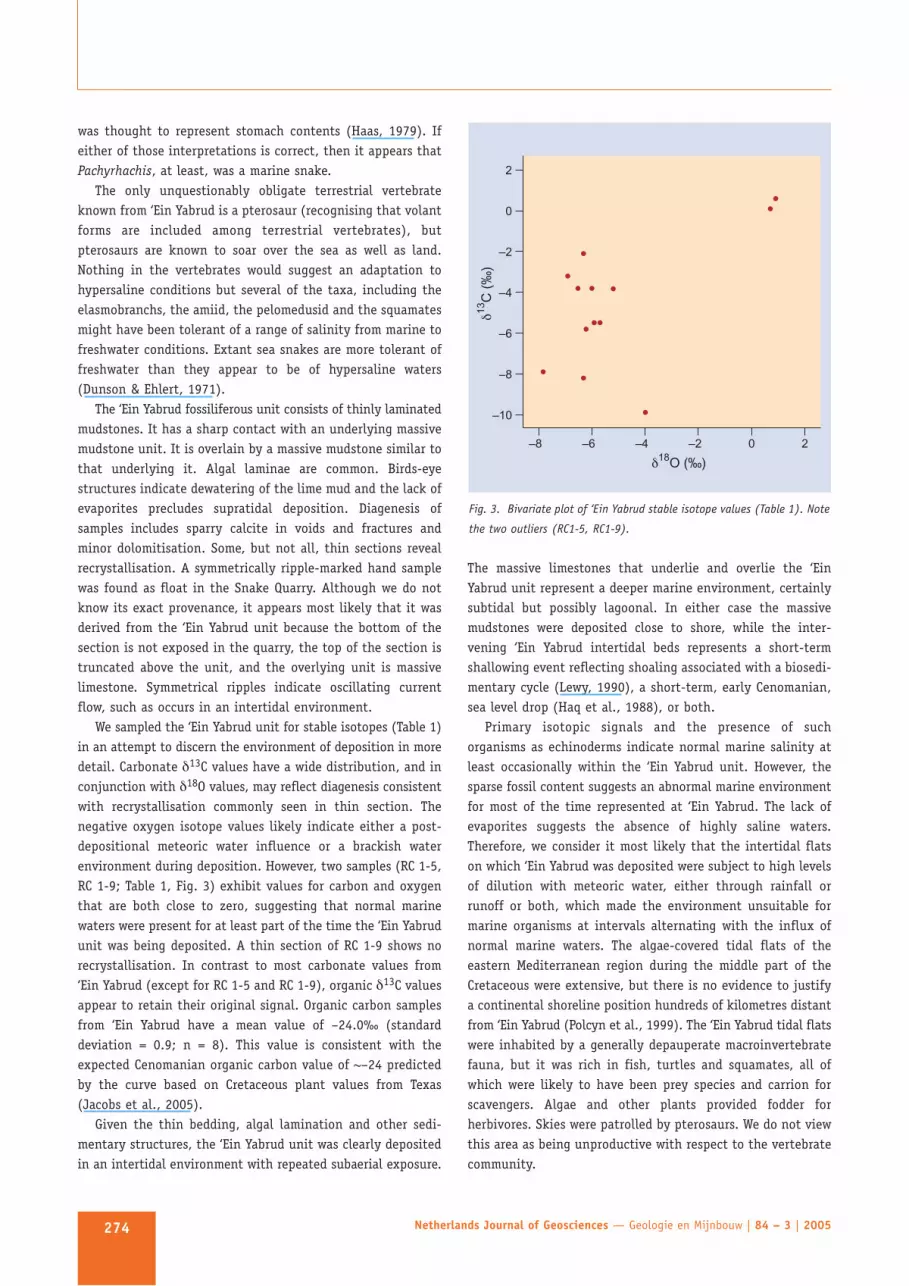
We sampled the ‘Ein Yabrud unit for stable isotopes (Table 1)in an attempt to discern the environment of deposition in moredetail. Carbonate δ13C values have a wide distribution, and inconjunction with δ18O values, may reflect diagenesis consistentwith recrystallisation commonly seen in thin section. Thenegative oxygen isotope values likely indicate either a post-depositional meteoric water influence or a brackish waterenvironment during deposition. However, two samples (RC 1-5,RC 1-9; Table 1, Fig. 3) exhibit values for carbon and oxygenthat are both close to zero, suggesting that normal marinewaters were present for at least part of the time the ‘Ein Yabrudunit was being deposited. A thin section of RC 1-9 shows norecrystallisation. In contrast to most carbonate values from‘Ein Yabrud (except for RC 1-5 and RC 1-9), organic δ13C valuesappear to retain their original signal. Organic carbon samplesfrom ‘Ein Yabrud have a mean value of –24.0‰ (standarddeviation = 0.9; n = 8). This value is consistent with theexpected Cenomanian organic carbon value of ~–24 predictedby the curve based on Cretaceous plant values from Texas(Jacobs et al., 2005).
Given the thin bedding, algal lamination and other sedi-mentary structures, the ‘Ein Yabrud unit was clearly depositedin an intertidal environment with repeated subaerial exposure.
The massive limestones that underlie and overlie the ‘EinYabrud unit represent a deeper marine environment, certainlysubtidal but possibly lagoonal. In either case the massivemudstones were deposited close to shore, while the inter-vening ‘Ein Yabrud intertidal beds represents a short-termshallowing event reflecting shoaling associated with a biosedi-mentary cycle (Lewy, 1990), a short-term, early Cenomanian,sea level drop (Haq et al., 1988), or both.
Primary isotopic signals and the presence of suchorganisms as echinoderms indicate normal marine salinity atleast occasionally within the ‘Ein Yabrud unit. However, thesparse fossil content suggests an abnormal marine environmentfor most of the time represented at ‘Ein Yabrud. The lack ofevaporites suggests the absence of highly saline waters.Therefore, we consider it most likely that the intertidal flatson which ‘Ein Yabrud was deposited were subject to high levelsof dilution with meteoric water, either through rainfall orrunoff or both, which made the environment unsuitable formarine organisms at intervals alternating with the influx ofnormal marine waters. The algae-covered tidal flats of theeastern Mediterranean region during the middle part of theCretaceous were extensive, but there is no evidence to justifya continental shoreline position hundreds of kilometres distantfrom ‘Ein Yabrud (Polcyn et al., 1999). The ‘Ein Yabrud tidal flatswere inhabited by a generally depauperate macroinvertebratefauna, but it was rich in fish, turtles and squamates, all ofwhich were likely to have been prey species and carrion forscavengers. Algae and other plants provided fodder forherbivores. Skies were patrolled by pterosaurs. We do not viewthis area as being unproductive with respect to the vertebratecommunity.
Netherlands Journal of Geosciences — Geologie en Mijnbouw | 84 – 3 | 2005274
2
0
–2
–4
–6
–8
–10
–8 –6 –4 –2 0 2
δ13C
(‰
)
δ18O (‰)
Fig. 3. Bivariate plot of ‘Ein Yabrud stable isotope values (Table 1). Note
the two outliers (RC1-5, RC1-9).

Geographic distribution of dolichosaurs and early mosasaurs
The age and correlation of dolichosaur- and early mosasaur-producing sites are considered by Jacobs et al. (2005). Here wedescribe their distribution from an ecological perspective.During the Cenomanian and Turonian stages both dolichosaursand plesiopedal mosasaurs were widespread longitudinally fromNorth America to the eastern Mediterranean region. Few datapoints are available within this time interval, so details of theorigins and dispersal of these marine squamates are mostlyunknown. Nevertheless, the sedimentary regimes at ‘Ein Yabrudand in the Eagle Ford Shale clearly represent a number ofdistinct depositional environments indicating that dolichosaursand plesiopedal mosasaurs were tolerant (at least) of intertidalto marine shelf habitats, assuming that the environments ofdeposition reflect the broad ecological tolerances of the animals.Further, there is no evidence to suggest that these organismswere deep divers. On the contrary, if their distribution inintertidal to shallow marine shelf is an accurate indication,they were not.
Globally, so far as the Cenomanian and Turonian recordpermits interpretation (Fig. 1), the environmental constraintson dolichosaurs and early mosasaurs appear similar. Withinthe Eagle Ford in Texas, the dolichosaur Coniasaurus crassidensis known from the Tarrant Member (Conlinoceras tarrantenseZone, 95.78±0.61 Ma) into the Austin Chalk (Coniacian, <85 Ma).An outstanding associated specimen was recovered from asiderite concretion in the Britton Shale (Sciponoceras gracileZone, ~94 Ma) by Lloyd Hill. Mosasaurs are unknown in the Tarrant Member and Britton Shale, but fragments ofRussellosaurus coheni occur in the Kamp Ranch. The typespecimens of the mosasaurs Russellosaurus coheni andDallasaurus turneri are from the Arcadia Park Shale within 20cm of the Kamp Ranch Limestone (Collignoniceras woollgariZone, ~92 Ma). Elsewhere within the Western Interior Seaway,dolichosaurs and early mosasaurs are known from Big BendNational Park, Texas, which is approximately their westernmostknown record, to South Dakota, which at ~45° N is the mostnortherly known occurrence.
In the eastern Mediterranean, from ‘Ein Yabrud and Lebanonto Croatia and the Balkan Peninsula, the environment appearsto have been part of an extensive carbonate platform system.Southern Europe was assembled from blocks of this carbonateplatform system, tectonically carved out of northeasternGondwana. The portion of the platform not welded to centralEurope and destined to become southern Europe remainedattached to the African component of Gondwana untilapproximately the Cenomanian or later (Polcyn et al., 1999).
The easternmost occurrence of dolichosaurs is Besokty II,Kazakhstan (Averianov, 2001). In western Europe (France,England and Germany), dolichosaurs are known from chalksand associated sediments (Jacobs et al., 2005). Other Turonian
basal Russellosaurina in addition to Russellosaurus cohenifrom Texas include Tethysaurus nopcsai, a plesiopedal formfrom Morocco, and Yaguarasaurus columbianus from Columbia.Yaguarasaurus is found at about the same palaeolongitude asthe type locality of Russellosaurus. If the Late Turonian plio-platecarpine Angolasaurus bocagei from Africa is taken intoaccount, early mosasaurs have a Southern Hemisphere recordat nearly 30° S. The total range of these Cenomanian andTuronian species spans approximately 120° of longitude asreconstructed, and from 45° N latitude to 30° S latitude. Theirbroad latitudinal distribution begs the question of sea-surfacetemperatures during the Cenomanian and Turonian stages.
Temperature tolerance of marine squamates and sea-surface temperature models
Dolichosaurs and early mosasaurs are small animals withbody weight from <1 kg to about 20 kg or larger. Because oftheir relatively small size, their surface area/volume ratio is aconsideration for thermoregulation, especially as water hasthermal properties that quickly cause heating or coolingthrough convection. Few modern analogues exist because fewliving squamates are marine, most notably Galápagos marineiguanas (Amblyrhynchus cristatus), which are inhabitants ofthe intertidal zone, and a variety of sea snakes. These groupsare of appropriate body size to be considered as analogues, atleast for the purpose of considering temperature tolerances.
Body mass of marine iguanas reaches 6 kg (Wikelski & Wrege,2000). They feed predominately to exclusively on intertidal tosubtidal macroalgae and spend a great deal of time foraging inthe water (Dawson et al., 1977; Wikelski & Wrege, 2000). Wikelskiet al. (1997) reported sea-surface temperatures around two setsof the Galápagos Islands (Genovesa and Santa Fe islands) thatrange between ~21° C and 30° C. These temperatures, as willbe seen below, are consistent with those tolerated by marinesnakes that spend little or no time on land. Marine iguanas,however, do spend time on land during which they often mustadjust body temperature by behavioural thermoregulation toprevent overheating (White, 1973). Because dolichosaurs andearly mosasaurs are found at a distance some kilometres fromshore, it is unlikely they required the terrestrial behaviouralmechanisms exhibited by marine iguanas except in thosecircumstances when they found themselves on shore, butunlike marine iguanas, they were able to exist in a fully aquaticmode for extended periods.
Sea snakes include the only extant obligate marinesquamates. They are primarily tropical in distribution. Theyellow-bellied sea snake, Pelamis platurus, inhabits waters thatremain at or above a monthly average of 20° C for each monthof the year (Dunson, 1975a; Dunson & Ehlert, 1971). Thus, thebreeding range of the species is defined by the 20° C isothermsfor the austral and boreal winters (Dunson, 1975b). Its lethalupper temperature limit is 33° C or above (Graham et al., 1971;
275Netherlands Journal of Geosciences — Geologie en Mijnbouw | 84 – 3 | 2005

Dunson & Ehlert, 1971), although sea snakes are rarelyobserved at temperatures approaching their lethal limit. Seasnakes swimming at surface temperatures up to 31° C probablymust regulate temperature behaviourally by diving below thesurface to filter the sun and to escape high surface temperatures(Dunson & Ehlert, 1971). Sea-surface temperatures above 30° Care limited in geographic extent and in duration. Monthly sea-surface averages above 30° C are not sustained on an annualbasis. These observations are particularly applicable to the ‘EinYabrud legged snakes, which co-occur with the mosasaurHaasiasaurus.
As a first approximation based on modern marine squamates,we hypothesise that Cenomanian and Turonian small-bodiedmarine squamates were normally restricted to waters no coolerthan 20° C and no warmer than 30° C because those are theconsistent limits for modern marine sea snakes and marineiguanas. Their lethal limits are beyond their optimal limits,but organisms do not normally prefer temperatures at theirlethal limits. It is biologically implausible that comparably-sized marine squamates (such as dolichosaurs and plesiopedalmosasaurs) would inhabit waters that would be lethal orunsuitable for modern sea snakes. Using those assumptions,we can now compare the distribution pattern of Cenomanianand Turonian marine squamates with modern distributions,with predictions derived from general circulation models andwith measured isotopic ocean surface palaeotemperaturevalues.
Two anomalies of distribution are immediately observable.First, sea snakes do not now occur in the Mediterranean Seabecause of temperature limitations at the low end of therange, possibly reinforced by high salinity and high summersurface temperatures in the Red Sea (Dunson, 1975b). Thus weconclude that the Mediterranean region had higher sea-surface temperatures in the Cenomanian and Turonian thannow, assuming that the limbed snakes of the Cenomanian inthe eastern Mediterranean region had ecological constraintssimilar to those of modern sea snakes. The configuration ofshallow sea on a carbonate platform at that time may in alllikelihood have contributed to higher sea-surface temperaturesduring the Cretaceous. Second, sea snakes do not extend to45° N latitude today. Although that was also the case forsnakes in the Cenomanian and Turonian so far as the recordshows, dolichosaurs and mosasaurs did extend to that latitude.If their temperature tolerances were similar to those of marinesnakes or marine iguanas, which we assume, then the surfacetemperature of the Western Interior Seaway at 45° N was onaverage between 20° C and 30° C. The depth of water may havebeen a contributing factor in maintaining high sea-surfacetemperature in that case as well.
Conversely, the distribution of dolichosaurs and mosasaursin the Cenomanian and Turonian includes the low-tropicallatitudes, which receive greater insolation than high latitudesand would be expected to have the highest sea-surface
temperatures. The localities of ‘Ein Yabrud, Goulmima andYaguara all fall at low latitudes (Fig. 1), and therefore, becauserelatively small mosasaurs are found there, suggest an uppersea-surface temperature limit of 30° C. This result may standin contrast to δ18O palaeotemperature calculations on plank-tonic foraminifera from Deep Sea Drilling Project (DSDP) Site144, north of Suriname in the Atlantic Ocean but at a palaeo-latiude of ~2 - 5° N, close to that of Yaguara. Norris et al. (2002)estimated sea-surface temperatures at that site to be ~33 - 34° C± 2° C. This estimate, in our opinion, is too high.
Bice et al. (2003) fitted δ18O sea-surface temperature esti-mates from the Southern Hemisphere high latitudes (60 - 65° S;DSDP Site 511, Falkland Plateau and James Ross Island) andfrom DSDP Site 144 during the Turonian to latitudinaltemperature gradient curves derived from atmospheric generalcirculation models with varying CO2 concentrations tosimulate greenhouse warming (Bice & Norris, 2002). The modelcurves are asymmetrical (Fig. 4), with high temperatures inthe southern high latitudes and a steeper gradient in theNorthern Hemisphere. Maximum equatorial temperature is~34° C at most CO2 concentrations. The δ18O palaeotemperaturedata of higher latitudes best fit the models with 6500 ppm and4500 ppm CO2, which is an order of magnitude higher thancurrent levels. Using marine squamate temperature tolerancedata, we would suggest that the maximum equatorial temper-ature in the models is too high and that the NorthernHemisphere gradient is too steep.
Discussion and conclusions
Our interpretation assumes certain sea-surface temperaturepreferences for dolichosaurs and plesiopedal mosasaurs. Wereject as implausible a scenario in which those small-bodiedmarine squamates would have inhabited waters with sea-surface temperatures beyond the normal tolerance of seasnakes. Biological attributes inferred for fossils, in this casesquamates, are useful in evaluating and testing models thatmanipulate CO2 to simulate global palaeotemperature distri-bution. Isotopic palaeotemperature determinations are alsovaluable in testing the models; however, even though δ18Ovalues are objectively determined, the temperature values theyrepresent are more subjective and require certain assumptionsregarding isotopic composition of the Cretaceous ocean. Arobust model must be realistic with respect to its constraints,and at the very least, inconsistencies should be evaluated.
The δ18O palaeotemperatures reported by Bice & Norris(2002) and Bice et al. (2003) reflect open-ocean sea-surfacevalues. The marine squamate localities on which we focus areintertidal or shelf, rather than deep open ocean. We wouldexpect therefore that in general temperature estimates basedon squamate tolerances in shallow water would be higher, notlower than sea-surface temperature estimates for deep open-ocean, other factors being equal. Our interpretations of sea-
Netherlands Journal of Geosciences — Geologie en Mijnbouw | 84 – 3 | 2005276

surface temperature are based on lumping Cenomanian andTuronian distribution records and have no greater resolutionthan the length of that interval in toto, although the temper-ature interpretation for each specific locality based on thepresence of small-bodied squamates is valid within the limitsof the underlying assumptions.
Body size and its correlated surface area/volume ratio areextremely important for all vertebrates, but especially in thiscase where small body size, relatively large surface area andthe ease of convective heat transfer in an aqueous medium aremajor controls on the distribution of the organisms. Later andmore derived mosasaurs reach quite large sizes and were betteradapted to a more whale-like marine existence. That is not thecase for dolichosaurs, which weighed less than a kg, or evenfor plesiopedal mosasaurs, which weighed much more than thatamount but did not reach the large size of later mosasaurs.
Had we only the intertidal record of the Middle East, wewould have interpreted dolichosaurs and plesiopedal mosasaursto have had a lifestyle more like Galápagos marine iguanas,being amphibious or perhaps more strictly shore dwellers. Ifthat were the case we would reason that they could controlbody temperature behaviourally, even if the water were a fewdegrees hotter than expected. However, when the Eagle Fordrecord is considered, these squamates are found in the basalsandy Tarrant Member, in the Britton Shale, in the shallowerand storm-generated Kamp Ranch Limestone and in the over-lying Arcadia Park Shale, up to a maximum depth of <100 m.In addition, dolichosaurs are found in the normal marinedeposits of the southern North Sea Basin in England, Franceand Germany. Thus, the habitats of these squamates, ifaccurately reflected by the environments of deposition inwhich they are preserved, are not limited to coastal orintertidal zones, but extend through shallow shelf depositsinto clean ocean water rich in carbonate-producing plankton.They are found repeatedly in deposits that were formedkilometres offshore, indicating that, even if amphibious, theyspent significant amounts of time in water, subject to thermalstress brought about by convective heat loss or gain.
In summary, small-bodied dolichosaurs and early mosasaursof the Cenomanian and Turonian stages inhabited a range ofenvironments from intertidal to shallow marine so far asknown. They were tolerant of brackish as well as normalmarine salinities. Their wide latitudinal distribution supportsgeneral circulation models that predict low temperaturegradients from equator towards poles, but it is not consistentwith unusually high equatorial sea-surface temperature. Bycomparison with living sea snakes and marine iguanas, theywere limited in their range to waters with surface temper-atures between 20° and 30° C.
277Netherlands Journal of Geosciences — Geologie en Mijnbouw | 84 – 3 | 2005
Fig. 4. Cloud containing all Cenomanian and Turonian latitudinal sea-
surface temperature distributions derived from GENESIS general
circulation models with CO2 values between 1120 and 7500 ppm (shaded
area) and δ18O palaeothermometry of planktonic foraminifera (Bice and
Norris, 2002; Bice et al., 2003; Norris et al., 2002). Latitude from south
(S) to north (N) on horizontal axis. Deep Sea Drilling Project (DSDP) core
sites are indicated by black arrows and labeled by number above
horizontal axis. The vertical range in δ18O temperature values at
individual DSDP sites reflects determinations of various foraminiferal
species within a sample of given age (solid triangle = late Albian, open
triangle = Cenomanian, shaded circle = Turonian). The horizontal black
lines at 20° C and 30° indicate the postulated normal temperature range
of dolichosaurs and early plesiopedal mosasaurs (see text). The dashed
vertical line at 45° N latitude indicates the northern limit of dolichosaurs
and early mosasaurs as their distribution is understood at this time. The
results of GENESIS experiments vary with CO2 concentration. We have
chosen two (1120 ppm and 7500 ppm) to mark the limits of our
comparison, and in that sense our selection of specific models is arbitrary.
The model utilizing 1120 ppm CO2 (lower border of shaded area) has
tropical sea-surface temperatures at the extreme acceptable upper tem-
perature limit for dolichosaurs and early mosasaurs (30° C) but predicts
sea-surface temperature at 45° N that are cooler than would be expected
based on those vertebrates (~15° C). The model specifying 7500 ppm CO2
(upper border of shaded area) predicts warm sea-surface temperatures at
high latitudes but also predicts warmer tropical temperatures than could
likely be tolerated by dolichosaurs and early mosasaurs (~33° C). Other
model values resulting from varying CO2 concentration fall within the
shaded area between the curves for 1120 and 7500 ppm (upper and lower
borders of shaded area) and present similar paradoxes. Note that
temperature estimates from δ18O palaeothermometry are transformed
values with underlying assumptions of ice volumes and ocean water 18O
concentrations that add uncertainty to temperature calculations.
Nevertheless, within the uncertainties of palaeothermometry and the
temperature tolerances of small-bodied squamates, tropical temperatures
near 30° C and low temperature gradients towards the poles is possible.

Acknowledgements
This work was facilitated by the help of a great manycolleagues including Gary Byrd, Roger Farish, Robert Gregory,Annat Haber, Bonnie Jacobs, Ze’ev Lewy, John Maurice, KentNewman, Ian Richards, Neil Tabor, Diana Vineyard and DaleWinkler. Aaran Pan and other students at SMU are appreciatedfor their help in the field and stimulating questions. RoyBeavers and the SMU Scanning Electron Microscopy Laboratory(NSF Grant EAR-0240251) provided energy dispersive X-rayanalyses. We thank the Institute for the Study of Earth andMan and the Stable Isotope Laboratory at SMU for use offacilities and financial support. Fieldwork in the Middle Eastwas supported by the National Geographic Society. Thin sectionswere prepared by Tulsa Sections, Inc., courtesy of StandardGeological Services, Inc., Littleton, Colorado. Special thanksare due to Anne S. Schulp and John W.M. Jagt for organisingthe First Mosasaur Meeting in Maastricht (May 2004), and toSteven Spurger of the Lewisville Independent School DistrictOutdoor Learning Center for access to a classic exposure ofEagle Ford Shale. We dedicate this contribution to our goodfriends now departed, Lloyd Hill, who taught us so much aboutthe stratigraphy and palaeontology of north-central Texas, andEitan Tchernov, who was our colleague and mentor in Israel.
References
Antunes, M.T., 1964. O Neocretácico e o Cenozóico do litoral de Angola. Junta
de Investigações do Ultramar (Lisboa), 257 pp.
Averianov, A.O., 2001. The first find of a dolichosaur (Squamata, Dolichosauridae)
in Central Asia. Paleontological Journal 35: 525-527.
Bardet, N., Pereda Suberbiola, X. & Jalil, N.-E., 2003. A new mosasauroid
(Squamata) from the Late Cretaceous (Turonian) of Morocco. Comptes
Rendus Palevol 2: 607-616.
Bell, B.A., Murry, P.A. & Osten, L.W., 1982. Coniasaurus Owen, 1850 from North
America. Journal of Paleontology 56: 520-524.
Bell, G.L., Jr. & Polcyn, M.J., 2005. Dallasaurus turneri, a new primitive
mosasauroid from the Middle Turonian of Texas and comments on the
polyphyly of Mosasauridae (Squamata). In: Schulp, A.S. & Jagt, J.W.M.
(eds): Proceedings of the First Mosasaur Meeting. Netherlands Journal of
Geosciences 84: 174-194.
Bell, G.L. Jr. & VonLoh, J.P., 1998. New records of Turonian mosasauroids from
the western United States. In: Martin, J.E., Hogenson, J.W. & Benton, R.C.
(eds): Partners preserving our past, planning our future. Dakoterra 5: 15-28.
Bice, K.L., Huber, B.T. & Norris, R.D., 2003. Extreme polar warmth during the
Cretaceous greenhouse? Paradox of the late Turonian δ18O record at Deep
Sea Drilling Project Site 511. Paleoceanography 18: 1031, doi:10.1029/
2002PA000848, 3003.
Bice, K.L. & Norris, R.D., 2002. Possible atmospheric CO2 extremes of the
middle Cretaceous (late Albian-Turonian). Paleoceanography 17: 1070,
doi:10.1029/2002PA000778, 2002.
Broin, F. de & Werner, C., 1998. New Late Cretaceous turtles from the Western
Desert, Egypt. Annales de Paléontologie 84: 131-214.
Buchy, M.-C., Smith, K.T., Frey, E., Stinnesbeck, W., González González, A.H.,
Ifrim, C., López-Oliva, J.G. & Porras-Muzquis, H., 2005. Annotated
catalogue of marine squamates (Reptilia) from the Upper Cretaceous of
northeastern Mexico. In: Schulp, A.S. & Jagt, J.W.M. (eds): Proceedings of
the First Mosasaur Meeting. Netherlands Journal of Geosciences 84: 195-205.
Caldwell, M.W., 1999. Description and phylogenetic relationships of a new
species of Coniasaurus Owen, 1850 (Squamata). Journal of Vertebrate
Paleontology 19: 438-455.
Caldwell, M.W. & Cooper, J.A., 1999. Redescription, palaeobiogeography and
palaeoecology of Coniasaurus crassidens Owen, 1850 (Squamata) from the
Lower Chalk (Cretaceous, Cenomanian) of SE England. Zoological Journal of
the Linnean Society 127: 423-452.
Carroll, R.L. & DeBraga, M., 1992. Aigialosaurs: mid-Cretaceous varanoid lizards.
Journal of Vertebrate Paleontology 12: 66-86.
Chalifa, Y., 1985. Saurorhamphus judeaensis (Salmoniformes: Enchodontidae),
a new longirostrine fish from the Cretaceous (Cenomanian) of ’Ein Yabrud,
near Jerusalem. Journal of Vertebrate Paleontology 5: 181-193.
Chalifa, Y., 1989a. Two new species of longirostrine fishes from the early
Cenomanian (Late Cretaceous) of Ein Yabrud, Israel, with comments on the
phylogeny of the Dercetidae. Journal of Vertebrate Paleontology 9: 324-328.
Chalifa, Y., 1989b. New species of Enchodus (Pisces: Enchodontoidei) from the
lower Cenomanian of Ein-Yabrud, Israel. Journal of Paleontology 63: 356-364.
Chalifa, Y., 1989c. Yabrudichthys and Serrilepis, two new genera of enchodontids
(Teleostei) from lower Cenomanian beds of ‘Ein-Yabrud, Israel. Israel Journal
of Zoology 36: 11-38.
Chalifa, Y. & Tchernov, E., 1982. Pachyamia latimaxillaris, a new genus and
species (Actinopterygii: Amiidae) from the Cenomanian of Jerusalem.
Journal of Vertebrate Paleontology 2: 269-285.
Dal Sasso, C. & Renesto, S., 1999. Aquatic varanoid reptiles from the Cenomanian
(Upper Cretaceous) lithographic limestones of Lebanon. Rivista del Museo
civico di Scienze naturali ‘E. Caffi’, Bergamo 20: 63-69.
Dalla Vecchia, F.M. & Venturini, S., 1999. The middle Cenomanian Lagerstätte
of al Nammoura (Kesrouâne Caza, N. Lebanon). Rivista del Museo civico di
Scienze naturali ‘E. Caffi’, Bergamo 20: 75-77.
Dawson, W.R., Bartholomew, G.A. & Bennett, A.F., 1977. A reappraisal of the
aquatic specializations of the Galapagos marine iguana (Amblyrhynchus
cristatus). Evolution 31: 891-897.
Diedrich, C., 1997. Ein Dentale von Coniasaurus crassidens Owen (Varanoidea)
aus dem Ober-Cenoman von Halle/Westf. (NW-Deutschland). Geologie und
Paläontologie in Westfalen 47: 43-51.
Diedrich, C., 1999. Erster Nachweiss von Dolichosaurus longicollis Owen
(Varanoidea) aus dem Ober-Cenoman von Halle/Westf. (NW-Deutschland).
Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1999/6: 372-384.
Dunham, R.J., 1962. Classification of carbonate rocks according to depositional
texture. In: Ham, W.E. (ed.): Classification of carbonate rocks. American
Association of Petroleum Geologists Memoir 1: 108-121.
Dunson, W.A., 1975a. Adaptations of sea snakes. In: Dunson, W.A. (ed.): The
biology of sea snakes. University Park Press (Baltimore, Maryland): 3-19.
Dunson, W.A., 1975b. Sea snakes and the sea level canal controversy. In:
Dunson, W.A. (ed.): The biology of sea snakes. University Park Press
(Baltimore, Maryland): 517-524.
Netherlands Journal of Geosciences — Geologie en Mijnbouw | 84 – 3 | 2005278

Dunson, W.A. & Ehlert, G.W., 1971. Effects of temperature, salinity, and surface
water flow on distribution of the sea snake Pelamis. Limnology and
Oceanography 16: 845-853.
Elder, W.P. & Kirkland, J.I., 1994. Cretaceous paleogeography of the southern
western interior region. In: Caputo, M.V., Peterson, J.A. & Franczyk, K.J.
(eds): Mesozoic systems of the Rocky Mountain Region, USA. Rocky
Mountain Section SEPM (Denver): 415-440.
Embry, A.F. & Klovan, J.V., 1971. A late Devonian reef tract on northeastern
Banks Island, Northwest Territories, Canada. Petroleum Geology Bulletin 19:
730-781.
Folk, R.L., 1962. Spectral subdivision of limestone types. In: Ham, W.E. (ed.):
Classification of carbonate rocks. American Association of Petroleum
Geologists Memoir, 1: 62-84.
Gayet, M., 1980. Recherches sur l’ichthyofaune cénomanienne de Monts de Judée:
les ‘Acanthopterygiens’. Annales de Paléontologie (Vertébrés) 66: 75-128.
Grande, L. & Bemis, W.E., 1998. A comprehensive phylogenetic study of amiid
fishes (Amiidae) based on comparative skeletal anatomy. An empirical
search for interconnected patterns of natural history. Society of Vertebrate
Paleontology Memoir 4: 1-690.
Haas, G., 1978a. A Cretaceous pleurodire turtle from the surroundings of
Jerusalem. Israel Journal of Zoology 27: 20-33.
Haas, G., 1978b. A new turtle of the genus Podocnemis from the lower
Cenomanian of ‘Ein Yabrud. Israel Journal of Zoology 27: 169-175.
Haas, G., 1979. On a new snakelike reptile from the lower Cenomanian of ‘Ein
Yabrud, near Jerusalem. Bulletin du Muséum national d’Histoire naturelle
(4)1: 51-64.
Haber, A. & Polcyn, M.J., 2005. A new marine varanoid from the Cenomanian
of the Middle East. In: Schulp, A.S. & Jagt, J.W.M. (eds): Proceedings of the
First Mosasaur Meeting. Netherlands Journal of Geosciences 84: 247-255.
Hancock, J.M. & Walaszczyk, I., 2004. Mid-Turonian to Coniacian changes of
sea level around Dallas, Texas. Cretaceous Research 25: 459-471.
Haq, B.U., Hardenbol, J. & Vail, P.R., 1988. Mesozoic and Cenozoic chrono-
stratigraphy and cycles of sea-level change. Society of Economic
Paleontologists and Mineralogists, Special Publication 42: 71-108.
Hay, W.W., DeConto, R.M., Wold, C.N., Wilson, K.M., Voigt, S., Schulz, M.,
Wold, A.R., Dullo, W.-C., Ronov, A.B., Balukhovsky, A.N. & Söding, E.,
1999. Alternative global Cretaceous paleogeography. In: Barrera, E. &
Johnson, C.C. (eds): Evolution of the Cretaceous Ocean-Climate System.
Geological Society of America, Special Paper 332: 1-47.
Henderson, R.A., 2004. A Mid-Cretaceous association of shell beds and organic-
rich shale: bivalve exploitation of a nutrient-rich, anoxic sea-floor
environment. Palaios 19: 156-169.
Jacobs, L.L., Ferguson, K., Polcyn, M.J. & Rennison, C., 2005. Cretaceous δ18O
stratigraphy and the age of dolichosaurs and early mosasaurs. In: Schulp,
A.S. & Jagt, J.W.M. (eds): Proceedings of the First Mosasaur Meeting.
Netherlands Journal of Geosciences 84: 257-268.
Kennedy, W.J. & Cobban, W.A., 1990. Cenomanian ammonite faunas from the
Woodbine Formation and lower part of the Eagle Ford Group, Texas.
Palaeontology 33: 75-154.
Lewy, Z., 1990. Transgressions, regressions and relative sea level changes on the
Cretaceous shelf of Israel and adjacent countries. A critical evaluation of
Cretaceous global sea level correlations. Paleoceanography 5: 619-637.
Moody, R.T., 1997. The paleogeography of marine and coastal turtles of the
North Atlantic and trans-Sahara regions. In: Callaway, J.M. & Nicholls, E.L.
(eds): Ancient marine reptiles. Academic Press (San Diego): 259-278.
Páramo, M.E., 1994. Posición sistemática de un reptil marino con base en los
restos fosiles encontrados en capas del Cretácico superior en Yaguara
(Huila). Revista de la Academia Colombiana de Ciencias exactas, fisicas y
naturales 19: 63-80.
Norris, R.D., Bice, K.L., Magno, E.A. & Wilson, P.A., 2002. Jiggling the
tropical thermostat in the Cretaceous hothouse. Geology 30: 299-302.
Polcyn, M.J. &Bell, G.L., 2005. Russellosaurus coheni n. gen., n. sp., a 92
million-year-old mosasaur from Texas (USA), and the definition of the
parafamily Russellosaurina. In: Schulp, A.S. & Jagt, J.W.M. (eds): Proceedings
of the First Mosasaur Meeting. Netherlands Journal of Geosciences 84:
321-333.
Polcyn, M.J., Jacobs, L.L. & Haber, A., 2005. A morphological model and CT
assessment of the skull of Pachyrhachis problematicus (Squamata:
Serpentes): a 98 million-year-old snake with legs from the Middle East.
Palaeontologica Electronica; http://palaeo-electronica.org/2005_1.
Polcyn, M.J., Tchernov, E. & Jacobs, L.L., 1999. The Cretaceous biogeography
of the eastern Mediterranean with a description of a new basal mosasauroid
from ‘Ein Yabrud, Israel. In: Tomida, Y., Rich, T.H. & Vickers-Rich, P. (eds):
Proceedings of the Second Gondwanan Dinosaur Symposium. National
Science Museum Tokyo, Monographs 15: 259-290.
Polcyn, M.J., Tchernov, E. & Jacobs, L.L., 2003. Haasiasaurus gen. nov., a new
generic name for the basal mosasauroid Haasia Polcyn et al., 1999. Journal
of Vertebrate Paleontology 23: 476.
Rage, J.-C., 1989. Le plus ancien lézard varanoïde de France. Bulletin de la
Société d’Étude scientifiques d’Anjou 13: 19-26.
Reid, W.T., 1952. Clastic limestone in the upper Eagle Ford Shale, Dallas County,
Texas. Field and Laboratory 20: 111-122.
Sageman, B.B. & Arthur, M.A., 1994. Early Turonian paleogeographic/paleo-
bathymetric map, Western Interior, U.S. In: Caputo, M.V., Peterson, J.A. &
Franczyk, K.J. (eds): Mesozoic systems of the Rocky Mountain Region, USA.
Rocky Mountain Section SEPM (Denver): 457-469.
Shuler, E.W., 1918. The geology of Dallas County. University of Texas Bulletin
1818: 1-54.
Tchernov, E., Rieppel, O., Zaher, H., Polcyn, M.J. & Jacobs, L.L., 2000. A fossil
snake with limbs. Science 287: 2010-2012.
Von Loh, J.P. & Bell, G.L. Jr., 1998. Fossil reptiles from the Late Cretaceous
Greenhorn Formation (Late Cenomanian-Middle Turonian) of the Black Hills
region, South Dakota. In: Martin, J.E., Hogenson, J.W. & Benton, R.C. (eds):
Partners preserving our past, planning our future. Dakoterra 5: 29-38.
White, F.N., 1973. Temperature and the Galapagos marine iguana: insights into
reptilian thermoregulation. Comparative Biochemistry and Physiology 45A:
503-513.
Wikelski, M., Carrillo, V. & Trillmich, F., 1997. Energy limits to body size in a
grazing reptile, the Galapagos marine iguana. Ecology 78: 2204-2217.
Wikelski, M. & Wrege, P.H., 2000. Niche expansion, body size, and survival in
Galápagos marine iguanas. Oecologia 124: 107-115.
279Netherlands Journal of Geosciences — Geologie en Mijnbouw | 84 – 3 | 2005

Appendix 1
This appendix consists of notes from observation of handspecimens and thin sections of rock samples of the Eagle FordShale, Kamp Ranch Limestone and ‘Ein Yabrud Limestone.Locality co-ordinates are on file in the Shuler Museum ofPaleontology, Southern Methodist University (Dallas, Texas).
Eagle Ford Shale
Britton Shale concretions. Not formed about a single fossil but,rather, preserve a laminated, fossiliferous environment. Thesiderite appears to be an overall, perhaps simultaneous,replacement rather than forming as a series of layers about acentral nucleus. The original shale deposition is represented inthe concretions, which do not reflect limestone environments.
LEF-3: Contains red-brown, presumably siderite, material, withbrachiopod and/or mollusc fragments and foraminifera.Inoceramus is absent from these samples. Sparry calcite andlime mud fill the fossil-related intramouldic and shelterporosity. Laminated shale, orientation unknown. Laminae are3 - 7+ mm thick. Lamina 1 is slightly compacted mudstonewith discontinuous stylolites. Fossils are rare and consist ofonly tiny fragments. Horizontally oriented non-sideritic intra-clasts present, but not common. Laminae 2 and 3 are in tran-sitional, with lamina 2 consisting of abundant non-sideriticintraclasts and flow structures. Lamina 3 is darker, moresideritic, and with horizontally oriented large fossil frag-ments. Shell fragments in this lamina reach 12 mm in length.
Laminae represent deposits made, presumably, in a shallow-water environment, below wave base. The fossil fragmentsindicate origin in relatively high energy. The materialpreserved in the concretions, therefore, represents high-energy material transported to a lower-energy environment.
EF-Concretion – This consists entirely of sideritic mudstonewith a single fossil fragment 20+ mm in length. Intraclastsare present, but not dominant. Similar to lamina 1 from theLEF-3 sample and represents a similar depositionalenvironment. Some bioturbation is present, but indicateslittle except normal marine water within the photic zone.
Kamp Ranch Limestone. Sample 1 (1382 and Eagle Ford Drive):this consists of Inoceramus shell fragments. Some lime mud ispresent, but the major layers are mostly shell fragments.Contains two ammonite impressions, presumably those ofCollignoniceras woollgari.
EFD 1382 (two samples) – Sample 1. This brown limestone is astylolitic mudstone – This limestone was deposited in alow-energy environment. It was later diagenetically alteredby pressure dissolution. The stylolites represent insolubleresidues resulting from the dissolution.
Sample 2. The brown and black sample is composed entirelyof limestone; shale is absent in this sample. The bottom totop stratification of one specimen is as follows:A–25 mm thick – Light brown fossiliferous packstone/
grainstone with brachiopod and Inoceramus shell frag-ments, and globorotaliid- and textulariid-type fora-minifera. Minute rounded sparry fragments appear to befragments of individual Inoceramus laths. Minor roundedglauconite peloids. Wood fragments are present as well.– High-energy environment, possible storm deposit. Woodsourced from a terrestrial environment. Glauconitepeloids may represents gastropod faecal material. If so,it represents a shallow-water environment of origin.
B–11 mm thick – Dark grey fossiliferous packstone with clearhard sparry spheroids, possibly rounded Inoceramus lathfragments. Brachiopod shell fragments and spines arepresent, as are wood fragments. Pyrite replacement offossils and some mudstone visible. High-energy environ-ment, possible storm deposit. Wood sourced from aterrestrial environment.
C– 10 mm thick – Same as unit AD– 9 mm thick – Same as unit BE– 14 mm thick – Same as units A and C, but more orange-
brown than brown.
‘Ein Yabrud
‘Ein Yabrud samples were taken from two outcrops. RC samplesare from a road cut through the crest of an anticline. SQsamples are from the quarry area, now a rubbish dump, thatproduced one of the famous ‘Ein Yabrud snakes with legs.Samples marked with asterisks are those from which thinsections were made.
SQ 2
The chert at top of SQ 2 section represents simple replacementof limestone, some of which contained worm tubes. Thenodule was not formed about a single organism, nor does itcontain sponge spicules.
**SQ 2-10 – Massive mudstone, dolomitic. Subtidal or abnormalmarine lagoon.
SQ 2-9 – Peloidal mudstone, dolomitic. Fractures filled withsparry calcite.
SQ 2-8 – Bioturbated mudstone with brachiopod and otherfossil fragments. Intertidal.
**SQ 2-7 – Bioturbated thinly laminated mudstone, with somedisturbed bedding, possibly due to desiccation. Intertidal.
SQ 2-6 – Interbedded laminae of mudstone and chert. Wormtubes fill some layers. Intertidal.
SQ 2-5 – Thinly laminated mudstone with algal laminae andstromatolites. Intertidal.
Netherlands Journal of Geosciences — Geologie en Mijnbouw | 84 – 3 | 2005280

SQ 2-4 – Thinly laminated mudstone with algal laminae.Predominantly recrystallised. Intertidal.
SQ 2-3 – Chalky powder, but with occasional larger piecesindicating thinly laminated mudstone. Intertidal.
SQ 2-2 – Chalky laminated mudstone.SQ 2-1 – Thinly laminated mudstone with algal laminae. It
represents an intertidal environment, fish vertebra present.Ripple marked hand sample in Snake Quarry. Found as float.
The ripple marks are symmetrical, indicating a back-and-forth water movement, typical of intertidal environments.
RC 1
**RC 1-9 – Thinly laminated mudstone with possibledolomitization of some laminae. Algal laminae present.Voids and fractures filled with sparry calcite. One largefracture is lined with a brown, presumably iron-rich,mineral and filled with sparry calcite. Intertidal.
RC 1-8 – Similar to RC 1-7, but lacking chert. Intertidal.RC 1-7 – Thinly laminated mudstone with chert nodules.
Porosity and fractures filled with sparry calcite; some poreslined with sparry calcite. Birds-eye structures present.Incompletely formed chert nodules show that the chert isreplacing sparry calcite. Some recrystallisation. Intertidal.
RC 1-6 – Thinly laminated mudstone with disturbed layers,possibly resulting from desiccation. Voids and fracturesfilled with sparry calcite. Intertidal.
**RC 1-5 – Similar to RC 1-4. Thinly laminated mudstone withalgal laminae. Voids and fractures filled with sparry calcite.Birds-eye structures present. Intertidal.
RC 1-4 – Chalky, peloidal mudstone with algal laminae. Voidsand fractures filled with sparry calcite. Birds-eye structurespresent. Some recrystallisation. Intertidal.
RC 1-3 – Chalky powder only. Mudstone. RC 1-2 – Chalky mudstone, thinly laminated with sparry
calcite filling void space. Fossils not observed. Intertidal.RC 1-1 – Thinly laminated mudstones with algal laminae.
Birds-eye structures present. Voids and fractures filled withsparry calcite. Peloids present, but fossils absent. Peloidalwackestone. Intertidal.
RC 2
**RC 2-9 – Massive mudstone, slightly fossiliferous. Subtidal.RC 2-8 – Thinly laminated mudstone. Irregular contacts
between laminae, some with minute load casts. Intertidal.**RC 2-7 – Base of sample is thinly laminated fossiliferous
mudstone, but most of the sample consists of bioturbatedfossiliferous mudstone. Worm tubes present. Foraminiferapresent. Intertidal.
RC 2-6 – Nearly all recrystallised to sparry calcite, but withghost laminae. May have been laminated mudstone,probably indicating intertidal environment.
RC 2-5 – Chalky powder, but with evidence of chertreplacement and worm tubes as seen in RC 2-3. Intertidal.
**RC 2-4 – Thinly laminated mudstone with algal laminae.Birds-eye structures. Wispy laminae present, indicatingcompaction. Intertidal.
RC 2-3 – Chalky powder with some larger pieces. Thinlylaminated mudstone with chert nodules. Tubes, possiblyworm tubes, present. Intertidal.
**RC 2-2 – Thinly laminated mudstone with algal laminae.Birds-eye structures. Voids filled with sparry calcite.Stylolites present, indicating compaction and pressuredissolution. Intertidal.
RC 2-2A – Thinly laminated mudstone with algal laminae.Fractures filled with sparry calcite.
**RC 2-1 (base of section) – Massive mudstone, slightlyfossiliferous. Subtidal or abnormal marine lagoon.
281Netherlands Journal of Geosciences — Geologie en Mijnbouw | 84 – 3 | 2005