scopolamine amnesia of passive avoidance: a deficit of information acquisition
TRANSCRIPT

BEHAVIORAL AND NEURAL BIOLOGY 50, 255--274 (1988)
Scopolamine Amnesia of Passive Avoidance: A Deficit of Information Acquisition
DOUGLAS K. RUSH 1
Cassella AG, CNS Pharmacology, Frankfurt, West Germany
Despite its increasing use as an animal model of memory deficit in human dementia, relatively few studies have attempted to assess the memory processes involved in the anticholinergic-induced impairment of passive avoidance retention. In the present experiments, the influence of scopolamine administered prior to or immediately following training on 24-h retention of step-through passive avoidance was studied in NMRI mice. In low doses (0.3-3.0 mg/kg ip) pretraining admin- istration ( - 5 min) of scopolamine induced a very strong amnesia. Post-training scopolamine induced a significant effect only at the highest dose tested (30 mg/kg). In a retention test of longer than normal duration (600 vs 180 s), which resulted in a more favorable comparison value in the control group, an intermediate post-training dose (10 mg/kg) induced a small effect which approached significance; a finding which may account for conflicting reports in the literature concerning the ability of scopolamine to induce a post-training deficit. The pretraining effect does not appear to have been solely the result of state-dependent learning; scopolamine (3 mg/kg) administered before both the training and test sessions induced a deficit of approximately the same magnitude as that found when administered before training or before testing only. The results indicate that scopolamine can induce a small post-trial effect, presumably through an influence on consolidation processes. The much larger effect of pretrial scopolamine, however, indicates a primary influence on processes related to information ac- quisition. Together with findings from the literature, the present experiments suggest that scopolamine-induced amnesia partially, but not completely, models the memory deficits of human dementia. © 1988 Academic Press, Inc.
Although several neurochemical systems have been implicated in Alz- heimer's disease, a hypofunctional cholinergic system is thought to be responsible for the cognitive, primarily memory, impairments found in this the most common form of dementia (Bartus, Dean, Beer, & Lippa, 1982; Coyle, Price, & DeLong, 1983; Gottfries, 1985). Attempts to establish animal models of Alzheimer's disease have utilized two general approaches which have focused on this cholinergic deficit.
1 The assistance of Kornelia Streit in collecting and analyzing the data is gratefully acknowledged. Reprint requests should be addressed to the author's present address: Hoechst-Roussel Pharmaceuticals, Inc., Rt. 202-206 North, Somerville, NJ 08876.
255 0163-1047/88 $3.00
Copyright © 1988 by Academic Press, Inc. All rights of reproduction in any form reserved.

256 DOUGLAS K. RUSH
In reproducing the neuronal degeneration reported to occur in the disease state (Whitehouse, Price, Struble, Clark, Coyle, & DeLong, 1982), excitotoxic amino acids have been used to lesion forebrain cholinergic structures (e.g., Dunnett, 1985). Although inducing a more specific lesion than can be obtained electrolytically, destruction of these forebrain nuclei does not result in neurotransmitter-specific damage, as these structures contain both cholinergic and noncholinergic neurons (Woolf, Hernit, & Butcher, 1986). The proposed cholinergic neurotoxin AF64A (Vickroy, Watson, Leventer, Roeske, Hanin, & Yamamura, 1985) would greatly improve this modeling approach if questions concerning its specificity can be resolved (Jarrard, Kant, Meyerhoff, & Levy, 1984).
The second approach utilizes application of anticholinergic compounds, primarily muscarinic antagonists such as scopolamine (e.g., Schindler, Rush, & Fielding, 1984), and occasionally the high-affinity choline uptake blocker hemicholinium-3 (Ridley, Barratt, & Baker, 1984) to induce deficits in learning and memory paradigms. Although the deleterious influence of anticholinergics on memory has been known for many years (see Essman, 1983; Spencer & Lal, 1983, for recent reviews), no concensus has emerged as to which of the processes encompassed by the term memory--acquisition, encoding, storage, or retrieval is affected.
An approach shown to be useful in separating effects on various aspects of memory processing involves comparison of the influence of experimental manipulations conducted before as opposed to after a learning experience (Gold & Zornetzer, 1983; McGaugh, 1983). A retention deficit induced by a post-training manipulation, such as drug treatment, can be interpreted as the result of an effect on processes related to memory formation. Experimental manipulations performed prior to the learning experience or at the time of memory retention testing could influence performance, i.e., the expression of memory, through effects on motor activity, mo- tivation, or sensory capacity, rather than memory formation itself (Heise, 1981).
As is true for other research efforts in the neurobiology of memory (see Gold, 1986), most studies which have examined the effects of post- training administration of anticholinergics have utilized passive avoidance tasks. Bammer's (1982) review of the pharmacology of passive avoidance retention cited 10 studies in which post-training administration of atropine or scopolamine was utilized; impairment was found in half of the studies, no effect was found in the other half. Two of the studies in which impairment was found utilized intracerebral injection into either the hip- pocampus (Wiener & Messer, 1973) or the caudate nucleus (Prado-Alcalft, Cruz-Morales, & L6pez-Miro, 1980; see Giordano & Prado-Alcal~i, 1986, for more recent findings). In the other three studies by Glick and colleagues (Glick & Greenstein, 1972; Glick & Zimmerberg, 1971, 1972), scopolamine

SCOPOLAMINE AMNESIA OF PASSIVE AVOIDANCE 257
was injected peripherally. These latter experiments are discussed in more detail below.
Very few studies have compared the effects of pre- and post-training administration using identical experimental parameters. The available data suggest that post-training treatment sometimes but not always induces an amnesia and that the size of pretraining effects is much larger than that induced by post-training treatment. In a study utilizing an active avoidance task (Flood & Cherkin, 1986), the effects of pre- and post- training administration appear to have been approximately the same, but a larger dose was used post-training (1.0 mg/kg) than pretraining (0.1 mg/kg) to induce impairment. Where comparisons have been made between pre- and post-training administration in passive avoidance paradigms, only Glick and colleagues have demonstrated an impairment of retention following post-training administration.
Gliek and Greenstein (1972) and Glick and Zimmerberg (1971, 1972) reported that post-training peripheral administration of scopolamine in the relatively high doses of 10.0 and 20.0 mg/kg (lower doses were not tested) impaired retention of passive avoidance. In comparison to pre- training administration of scopolamine, the effects were rather small. Utilizing a maximum cut-off latency of 600 s, the effects of post-training administration were significant, although this would not have been the case had a cut-off latency of 300 s been used, as pointed out by Glick and Zimmerberg (1971) and Bammer (1982). Wiener and Messer (1973) in discussing these findings, commented that a retention latency of more than 300 s following post-trial administration can hardly be considered indicative of a strong amnesia when compared to latencies of less than 100 s induced by pretraining administration or latencies of 20 s in untreated control animals.
Three other studies which have compared the effects of pre- and post- training administration of cholinergic antagonists (Bohdaneck9 & Jarvik, 1967; Calhoun & Smith, 1968; Dilts & Berry, 1967) have found impairment following pretraining but no effect following post-training treatment. Boh- daneck57 and Jarvik (1967) studied only one dose of scopolamine (1.0 mg/kg), which according to the results of Glick and colleagues cited above may have been too low to induce a post-training effect. In the studies of Calhoun and Smith (1968) and Dilts and Berry (1967), a variety of doses ranging from 0.03 to 20.0 mg/kg was employed, a range which included the doses (10.0 and 20.0 mg/kg) found by Glick and colleagues to induce a post-training effect. The failure by Calhoun and Smith (1968) and Dilts and Berry (1967) to find a post-training effect may have been the result of the rather weak learning engendered by their training con- ditions. In both studies, the untreated control group had a median retention latency of less than 50 s, a baseline from which it may have been difficult

258 DOUGLAS K. RUSH
to induce a demonstrable amnesia if post-training administration of sco- polamine produces only a weak effect, as suggested by the results of Glick and Greenstein (1972) and Glick and Zimmerberg (1971, 1972).
A number of studies (reviewed in Bammer, 1982) indicate that pretraining scopolamine treatment impairs retention at doses between 1 and 3 mg/kg. The studies reviewed above suggest that larger doses are required to induce post-training impairment. Furthermore, there is some question as to the size of the pre- as opposed to post-training effects, which has implications concerning the involvement of the cholinergic system in memory processes. Smaller effects of post-training administration would suggest that the cholinergic system modifies memory primarily through an influence on processes operating at the time of information acquisition rather than following the learning experience.
The present experiments were conducted to add to those few studies in the literature in which pre- and post-training anticholinergic treatments have been compared. Given an adequate baseline of learning in control subjects for demonstrating small effects, we were interested in determining whether a post-training effect could be found with smaller doses than those used by Glick and Greenstein (1972) and Glick and Zimmerberg (1971, 1972), and whether the pre- and post-training effects were of comparable size. Such information could prove useful both for an un- derstanding of the neurobiology of memory and in the establishment of valid animal models of human memory disorder.
GENERAL METHODS
Male NMRI mice (Charles River-Wiga, Sulzfield, West Germany) weighing 24-30 g served as subjects following a 1-week adaptation period in our colony housing (12 h light cycle from 0600 to 1800, ad lib food and water).
Training and testing of passive avoidance were conducted in two identical black and white Plexiglas V-shaped cages consisting of dark (20 cm long) and light (40 cm long) compartments. An opening (5 x 5 cm) in the wall separating the two halves of the cage could be closed with a sliding guillotine door. A grid floor (5 cm wide) consisting of 2-mm stainless- steel rods spaced 1 cm apart extended throughout the length of the cage.
Following random assignment to experimental groups of 14 or 15, subjects were administered one training trial on Day 1 and were tested for acquisition of passive avoidance 24 h later, between the hours of 0900 and 1500. On the training day, mice were placed into the lit (100- W bulb) side of the test cage facing away from the dark compartment. After entering the dark compartment (response latency was recorded), the door separating the two sides was gently closed and a footshock (! mA) of 1 s duration was administered. In the retention test 24 h following training, the latency to first enter the dark compartment and both the

SCOPOLAMINE AMNESIA OF PASSIVE AVOIDANCE 259
duration and frequency of dark compartment entries were measured for 3 min with a microcomputer (Rush, 1986a). Those subjects not entering the dark side were assigned a latency of 180 s and duration and frequency scores of 0. Only rarely are parameters other than latency reported in the experimental literature on passive avoidance. By measuring several parameters, the possibility of finding relatively small effects, as was desired in the present studies, may be increased.
Data Analysis
Due to skewed distributions and inhomogeneity of variance, data were analyzed with a nonparametric multiple comparison procedure (Zwick and Marascuilo, 1984). The Mann-Whitney-Wilcoxon statistic was cal- culated using pairwise ranking of groups and a correction for ties (Gibbons, 1976). As the comparisons were planned, no omnibus tests were conducted. p < .01 was required for significance. This resulted in control of Type I experimental error rate approximating that achieved with the Dunn- Bonferroni procedure in which an overall significance level of .05 would be divided equally among four to five comparisons for each parameter (latency, duration, and frequency) within each experiment. Utilizing the same p value for all comparisons has the advantage, however, of keeping the power per comparison equal (given approximately equal sample sizes) across experiments with varying numbers of contrasts, which was the case in the present series of studies.
In addition to the Wilcoxon statistic, Fisher's exact test of the distribution of mice with latencies equal to as opposed to less than the maximum measured latency was calculated, p < .01 was required for significance.
EXPERIMENT 1
This experiment was conducted to demonstrate the degree to which the cholinergic antagonist scopolamine induces an amnesia under our experimental conditions.
Methods
The experiment comprised five groups of animals, four of which were administered footshock during training. The other group received no footshock (the shock leads to the grid floor were disconnected) and served as a no learning control group for assessing the size of the induced amnestic effects.
All groups were injected ip 5 min prior to training. Two groups, the no footshock control group and another group which received footshock, were injected with saline. The three remaining groups were administered one of three doses of scopolamine HBr (0.3, 1.0, or 3.0 mg/kg). Doses were calculated as the free base, dissolved in saline (0.9% NaC1), and injected in volumes of 10 ml/kg.

260 DOUGLAS K. RUSH
Results and Discussion
The results for all five groups are presented in Table 1. The administration of footshock (no scopolamine treatment) resulted in strong learning of passive avoidance; 12 of the 15 subjects did not enter the dark compartment within the 180-s test session. This distribution of scores was significantly different (Fisher's exact test) than in the no footshock group, where no mice had latencies of 180 s. Mann-Whitney comparisons of the group not administered footshock with the shock group indicated that retention was significantly better as measured by all three parameters.
Scopolamine administered prior to training induced a very strong dose- related amnesia. The effects of all three doses on all three parameters were significant; significant decreases in latency were accompanied by significant increases in both the duration and frequency of dark com- partment entries. The number of subjects with a latency of 180 s was significantly decreased by all three doses (Fisher's test).
Mann-Whitney comparisons of the no footshock group with the groups administered scopolamine indicated significant differences for all three doses on the latency parameter and at the lowest dose on the duration parameter. Marginally significant effects on duration at the 1 mg/kg dose and on frequency at the two lowest doses were also found. The no footshock and 3 mg/kg scopolamine groups did not differ on the duration and frequency parameters and Fisher's exact test indicated no difference between these groups in the distribution of subjects with latency scores equal to as opposed to less than the lS0-s maximum. In summary, the severity of the amnesia at the highest dose equaled the level of performance of animals that did not learn the task, i.e., the amnesia was almost complete.
In replication of several other studies (see Bammer, 1982, for a review), pretraining scopolamine induced a very strong impairment of passive avoidance retention. This impairment does not appear to have been the result of any debilitating changes induced by the administration of sco- polamine, as measured by the latency to enter the dark compartment during training; a comparison of the median training latencies of the group injected with saline which received footshock with each of the other groups indicated no significant differences (Table 1). Furthermore, the induced amnesia is of central origin since we have previously shown (Rush, 1986b) that methyl-scopolamine, which poorly penetrates the blood- brain barrier, has no effect on retention.
EXPERIMENT 2
Experiment 2 was conducted to provide a direct comparison of the size of pre- and post-training effects of scopolamine on the retention of passive avoidance utilizing identical experimental conditions.

SCOPOLAMINE AMNESIA OF PASSIVE AVOIDANCE 261
Methods
Five groups of animals were used. All groups were administered foot- shock to induce learning and were injected two times, once 5 min prior to and once within 30 s following the training experience. The control group for assessing the strength of the induced amnesia was injected twice with saline. Each of the other groups received one of two doses of scopolamine (0.3 or 3.0 mg/kg) either 5 min before or within 30 s immediately following training; the other injection was of saline.
Results and Discussion
The degree of memory retention shown by the footshock control group in this experiment was less than that shown by the equivalent group in experiment 1, although the groups did not differ significantly. This was most clearly manifested by the number of mice with the maximum latency of 180 s; 12 versus 7 in experiments 1 and 2, respectively. Despite this variability, the results of experiment 1 were replicated; the pretraining administration of scopolamine resulted in a very strong amnesia. Except for the Mann-Whitney comparison of the frequency parameter (p < .02) and the Fisher's test (p < .04) at the 0.3 mg/kg dose, all other effects were significant (Table 2).
In contrast to the effects of pretraining administration, the effects of post-training scopolamine failed to reach significance. The effects on latency for both doses (p < .05) and on the duration measure at the 3.0 mg/kg dose (p < .03) approached, but did not reach, the probability level (p < .01) required for significance in this study. Results of Fisher's exact test of the distribution of latency scores supported the results of the tests of central tendency; neither of the drug groups differed from the control group (p > .40).
The pattern of results indicates that pretraining administration of sco- polamine induces a much stronger amnesia than does post-training ad- ministration. Direct statistical comparison of the pre- versus post-training groups provided further evidence for this interpretation. The frequency of dark compartment entries was significantly greater in subjects admin- istered 3 mg/kg of the drug prior to training than in the group receiving the same dose after training. Effects on the duration parameter at the 0.3 mg/kg dose (p < .03) approached significance. Comparison of the distribution of latency scores with Fishers's test indicated no effects.
The difference in the effects of pre- as opposed to post-training sco- polamine on retention does not appear to have been the result of general debilitating effects in the groups administered scopolamine before training. As in experiment 1, there were no differences between the groups in latency to enter the dark compartment in the training phase (Table 2).

TA
BL
E 1
t,~
P
retr
aini
ng S
copo
lam
ine
(-5
ra
in)
Str
ongl
y Im
pair
s P
assi
ve A
void
ance
Ret
enti
on
va
Med
ian
(int
erqu
arti
le r
ange
, IQ
R)
Tra
inin
g R
eten
tion
tes
t m
g/kg
la
tenc
y (s
) L
aten
cy (
s)
Dur
atio
n (s
) F
requ
ency
N
=
180
s
No
Foo
tsho
ck
12 (
5)
10 (
3) ~
94
(10)
" 5
(1)"
0"
F
oots
hock
17
(13
) 18
0 (0
) 0
(0)
0 (0
) 12
F
oots
hock
+
scop
olam
ine
0.3
18 (
5)
e 51
(80
) b'"
30(4
2) b'
d 3
(3) b
'~
4 h
1.0
17 (
7y
41 (
43) ..
.. 67
(30
) ""
4 (2
) "'"
2 ~
3.0
23 (
13y
36 (
8)"
" 82
(21
)"
5 (2
)"
0"
Not
e. N
=
15 f
or a
ll g
roup
s. C
ompa
riso
ns w
ith
the
foot
shoc
k gr
oup:
~p
< .0
01,
op <
.0
1. C
ompa
riso
ns w
ith
the
No
Foo
tsho
ck g
roup
: "p
<
.001
, ap
<
.01,
ep
< .0
5.
TA
BL
E 2
P
ost-
trai
ning
Sco
pola
min
e (w
ithi
n 30
s)
Mil
dly
Impa
irs
Pas
sive
Avo
idan
ce R
eten
tion
©
Med
ian
(IQ
R)
Tra
inin
g R
eten
tion
tes
t m
g/kg
la
tenc
y (s
) L
aten
cy (
s)
Dur
atio
n (s
) F
requ
ency
N
=
180
s
Con
trol
15
(5)
16
6 (2
7)
11 (
21)
1 (1
) 7
Bef
ore
0.3
21 (
4)
25 (
13)"
12
0 (2
9)"
2 (1
)"
1 c
3.0
17 (
7)
21 (
13)"
95
(30
)"
4 (2
) b
0 b
Aft
er
0.3
15 (
4)
82 (
78)"
28
(57
)"
2 (1
) 4
3.0
12 (
6)
104
(79)
''~
48 (
63)"
I
(1) a
4
Not
e. N
=
15 f
or a
ll g
roup
s ex
cept
the
0.3
mg/
kg A
fter
gro
up w
here
N
= 14
. C
ompa
riso
ns w
ith
the
Con
trol
gro
up:
"p <
.0
01,
hp <
.0
1, '
p <
.05.
Bef
ore
vs A
fter
com
pari
sons
at
the
sam
e do
se:
dp <
.0
1, U
p <
.05.

SCOPOLAMINE AMNESIA OF PASSIVE AVOIDANCE 263
EXPERIMENT 3
As discussed under Introduction, Glick and Greenstein (1972) and Glick and Zimmerberg (1971, 1972) may have found impairment of retention in their studies due to the use of a 600-s cut-off and high doses of scopolamine. The present experiment replicated these experimental con- ditions in an attempt to duplicate the impaired retention they reported.
Methods
Subjects in the three groups of this experiment were injected within 30 s immediately following training. The control group received the saline vehicle; the experimental groups were administered either 10 or 30 mg/kg scopolamine. As in experiments 1 and 2, the latency to enter the dark compartment and the duration and frequency of dark compartment entries during the retention test session were recorded. In addition, the latency of those animals not entering the dark compartment within 180 s was measured up to a maximum of 600 s, as in the studies of Glick and colleagues (Glick & Greenstein, 1972; Glick & Zimmerberg, 1971, 1972).
Results and Discussion
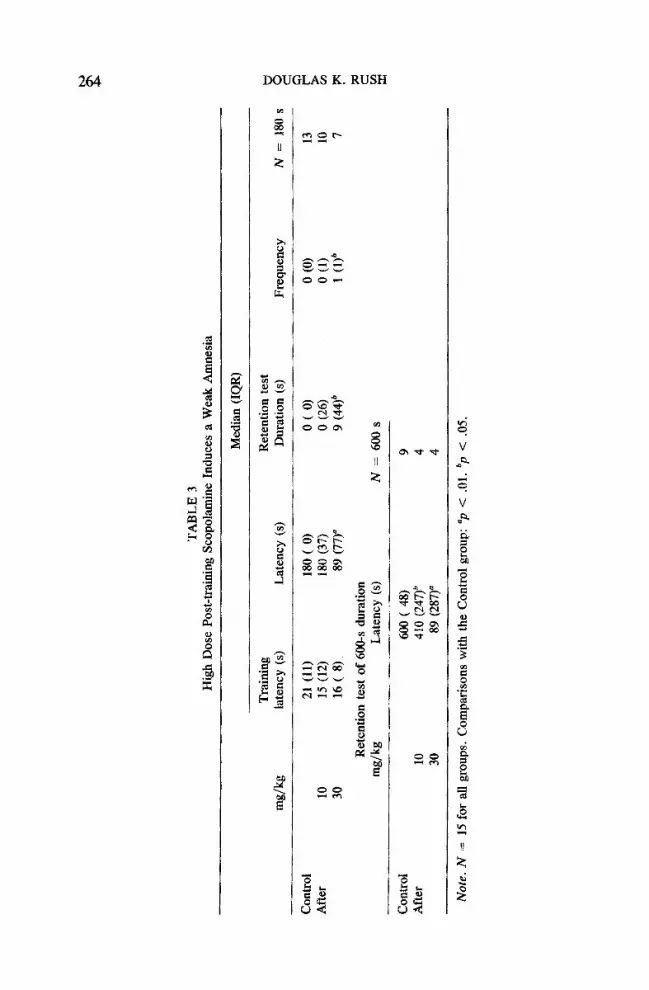
The learning exhibited by the footshock control group was stronger in this experiment than in experiment 2, though not significantly so, and comparable to that in experiment 1. In replication of Glick and Greenstein (1972) and Glick and Zimmerberg (197 l, 1972), a high dose of scopolamine was able to induce a significant retention impairment (Table 3). Utilizing measurements made in our normal 3-rain test, the group administered a dose of 30 mg/kg showed a significantly shorter latency and increases in both duration (p < .02) and frequency (p < .03) measurements that approached significance. In addition, the relatively nonpowerful Fisher's test indicated a modest effect (p < .06) on the distribution of latency scores at the 30 mg/kg dose. Utilizing a 600-s cut-off, analysis of the latency data (the only parameter measured up to a maximum of 10 min) indicated a dose-related retention impairment; the impairment induced with 30 mg/kg was significant and the shorter latency observed in the 10 mg/kg group approached significance (p < .03). Fisher's test results were nonsignificant for both groups. As in experiments 1 and 2, latencies to enter the dark compartment during training did not differ between the the groups.
EXPERIMENT 4
The results of experiments 1-3 demonstrate clear-cut and large effects of pretraining as compared to post-training administration of scopolamine. The interpretation offered for this differential effect assumes that only
b~
TA
BL
E 3
H
igh
Dos
e Po
st-t
rain
ing
Sco
pola
min
e In
duce
s a
Wea
k A
mne
sia
Med
ian
(IQ
R)
Tra
inin
g R
eten
tion
tes
t m
g/kg
la
tenc
y (s
) L
aten
cy (
s)
Dur
atio
n (s
) F
requ
ency
N
=
180
s
Con
trol
A
fter
21
(11
) 18
0 (0
) 0
(0)
0 (0
) 13
~.
10
15
(12
) 18
0 (3
7)
0 (2
6)
0 (1
) 10
30
16
(8
) 89
(77
)"
9 (4
4) b
1 (1
) h
7 .~
Ret
enti
on t
est
of 6
00-s
dur
atio
n m
g/kg
L
aten
cy (
s)
N
= 60
0 s
Con
trol
60
0 (4
8)
9 A
fter
10
41
0 (2
47) b
4
30
89 (
287)
" 4
No
te.
N
=
15 f
or a
ll g
roup
s. C
ompa
riso
ns w
ith
the
Con
trol
gro
up:
ap <
.0
1. b
p <
.05.

SCOPOLAMINE AMNESIA OF PASSIVE AVOIDANCE 265
processes operative at the time of drug administration are responsible for the induced amnesia. An alternative explanation, however, could account for the pretraining administration induced effect, namely, state- dependent learning (Overton, 1984).
Although state-dependent effects of anticholinergics have been dem- onstrated, for example, on active avoidance (Overton, 1966), only two studies have utilized the standard 2 x 2 design used to assess such effects in passive avoidance paradigms. Glick and Greenstein (1972) were able to demonstrate state-dependent forgetting: scopolamine administered either before training or before retention testing induced an amnesia; when administered before both training and testing no deficit was found. In contrast, Calhoun and Smith (1968) found effects under all three conditions. These two studies also differed, as discussed under Introduction, in finding (Glick & Greenstein, 1972) and not finding (Calhoun & Smith, 1968) an effect of post-training scopolamine. Given the availability of only these two conflicting studies in which state dependency and post- training effects were examined under the same conditions, experiment 4 was conducted.
Methods
Utilizing the standard state-dependent design in which subjects were injected twice, the effects of a single dose (3 mg/kg) of scopolamine administered before training ( - 5 min), before the retention test ( - 5 min), or before both training and test were investigated. All groups, including a control group, received the saline vehicle when not administered scopolamine.
Results and Discussion
The learning exhibited by the footshock group administered saline was weaker in this experiment than in experiments 1 and 3 and comparable to that of experiment 2; Mann-Whitney and Fisher tests were nonsignificant, however. Despite the variability in the control group values from experiment to experiment, the results of the scopolamine treatments were consistent with those obtained in the previous experiments. As in experiments ! and 2, pretraining scopolamine produced a very strong amnesia; the effects on all three parameters were significant (Mann-Whitney tests) and the number of subjects with a latency equal to 180 s was reduced (Fisher's test). When administered prior to the test session only or prior to both training and test, scopolamine also produced a significant amnesia; Mann-Whitney tests indicated significant effects on all measured parameters.
To assess the degree of amnesia produced, the three groups administered scopolamine were compared to the no footshock group. Only the frequency

b.~
TA
BL
E 4
P
rete
st S
copo
lam
ine
(-5
m
in )
Str
ongl
y D
isru
pts
Ret
enti
on
Per
form
ance
Med
ian
(IQ
R)
Tra
inin
g R
eten
tio
n t
est
late
ncy
(s)
Lat
ency
(s)
D
urat
ion
(s)
Fre
qu
ency
N
=
180
s
~7
©
No
Fo
ots
ho
ck
15 (
6)
7 (3
) °
90 (
27) a
6
(1)"
0 c
F
oots
hock
13
(3)
17
0 (3
4)
6 (2
3)
1 (2
) 6
Sco
pola
min
e (3
mg
/kg
) be
fore
T
rain
ing
14 (
5)
8 (1
4)"
86(3
5) ~
3
(2) b
0'
T
est
13 (
4)
9 (5
) a
64 (
8)
~'1
9 (2
) ...
..
0 ~
Tra
inin
g an
d T
est
15 (
4)
14 (
9)
a 64
(14
) b's
9
(2) "
'h
0 C
t"
Not
e. N
=
15 f
or a
ll g
roup
s ex
cept
the
Bef
ore
Tra
in a
nd T
est
grou
p w
here
N
= 14
. C
om
par
iso
ns
wit
h th
e F
oo
tsh
ock
gro
up:
"p <
.0
01,
bp <
.0
1, '
)9 <
.0
5. C
om
par
iso
ns
wit
h th
e N
o F
oo
tsh
ock
gro
up:
ap <
.0
01,
"p <
.0
1, I
p <
.05.
Co
mp
aris
on
s w
ith
the
Bef
ore
Tra
inin
g gr
oup:
~p
< .0
01,
hp <
.0
1.

SCOPOLAMINE AMNESIA OF PASSIVE AVOIDANCE 267
parameter in the group administered scopolamine before the test session reached the probability level required for significance (p < .01). In the groups administered scopolamine before the test or before both training and test, the probability levels on the duration parameter approached (p's < .05) but did not reach significance. These findings suggest that the degree of amnesia was strongest in the group administered scopolamine before training, since none of the three parameters for this group differed from the no footshock group.
Comparisons among the three groups administered scopolamine, how- ever, suggest that pretest administration of this drug produced the largest effects; the frequency of dark compartment entries was significantly higher in the two groups administered scopolamine before the retention test, which did not differ from each other. No differences were found on the other two parameters, latency or duration, nor in the distribution of latencies equal to or less than 180 s (Fisher's test) among the three groups.
In summary, administration of scopolamine prior to both training and testing produced an impairment at least as large as, if not larger than, that found when the compound was administered only prior to training. This result rules out an interpretation of the deficits found in the groups administered the drug either before training or before the test as due solely to state-dependent learning. On the other hand, state-dependent processes cannot be totally excluded; according to Overton (1984), the pattern of results suggests a possible confounding of effects on memory processes (acquisition/encoding and retrieval) with effects on drug-induced state-dependent processes.
GENERAL DISCUSSION
The results of the present experiments demonstrate, in corroboration with previous findings (Glick & Greenstein, 1972; Glick & Zimmerberg, 1971, 1972), that the post-training administration of scopolamine can induce a retention deficit in a passive avoidance paradigm. This effect, however, is of smaller magnitude than that which can be induced by pretraining administration and a relatively high dose of scopolamine is necessary to induce it. The results also show that a retention test of long duration may enable the demonstration of a small magnitude post-training effect by providing a better control group comparison value. With a maximum latency of 180 s, there was no effect of a dose of 10 mg/kg (experiment 3); with a cut-off of 600-s, the effect of this dose approached significance.
Glick and Zimmerberg (1972) have reported analogous findings; post- training scopolamine impaired retention with high (350/xA) but not with low (60 or 230 txA) footshock. The failure to induce amnesia with low

268 DOUGLAS K. RUSH
footshock was probably the result of an inadequate baseline from which to demonstrate a small effect; the retention latencies of the control groups increased from 254 to 600 s with increasing shock intensity. As mentioned under Introduction, a failure in some studies (Bohdaneck~ & Jarvik, 1967; Calhoun & Smith, 1968) to find a post-training effect with a high dose of scopolamine was probably the result of this floor effect of control latencies which were too low to permit demonstration of a deficit.
The finding that pretest scopolamine strongly impaired retention test performance (experiment 4) corroborates several previously published studies (Bure~ov~, Buret, Bohadneck~,, & Weiss, 1964; Calhoun & Smith, 1968; Glick & Greenstein, 1972), and suggests that scopolamine impaired the retrieval of the previously learned information. As discussed under Introduction, this interpretation is open to the criticism that the presence of the drug during testing influenced performance, i.e., the expression of memory, through effects on motor activity, motivation, or sensory capacity. In fact, the higher frequency of dark compartment entries in the groups administered scopolamine before the retention test than in the group receiving the drug prior to training suggests an effect on motor activity.
Numerous studies (e.g., Bushnell, 1987) have demonstrated the motor activity enhancing effects of anticholinergics. In our own studies (Jablonka and Rush, 1987), a dose of 3 mg/kg scopolamine induces a maximal increase in arm changes measured in a symmetrical Y-maze in the same species of mouse as used in the present experiments. The increased frequency of dark compartment entries seen in experiment 4, in those groups administered this same dose of scopolamine prior to retention testing, may have resulted from an induced hyperactivity. If this inter- pretation is correct, it would be expected tht other compounds which increase motor activity, e.g., amphetamine, might also induce deficits in passive avoidance when administered before retention testing. Both pub- lished studies (see Bammer, 1982, for a review) and our own unpublished experiments, however, indicate little, if any, impairing effect of am- phetamine on retention test performance. Given the paucity of data in the literature on the effects of anticholinergics on parameters other than latency in passive avoidance paradigms, future studies might well focus on this issue.
Regardless of the correct interpretation of the impairment produced by pretest scopolamine, the results of experiment 4 indicate that the deficit induced by administration of scopolamine prior to training is not solely the result of state-dependent learning; effects on memory are at least partially responsible for the retention deficit. As post-training sco- polamine exerts only a small influence on retention, the primary effect of pretraining scopolamine must be on acquisition processes.

SCOPOLAMINE AMNESIA OF PASSIVE AVOIDANCE 269
One other aspect of the present experiments supports the contention that scopolamine induces a deficit primarily via an effect on acquisition processes. The use of a 5-min resorption time for pretraining scopolamine resulted in a time difference of approximately 6 min between the pre- and post-training conditions. We have examined the effects of pretraining resorption times up to 60 min and have found no differences in the degree of impairment produced. The results of experiments 2 and 3, however, show that a shift of 6 min in resorption time from before training to after training dramatically alters the degree of impairment produced. The dif- ferential effects on retention, therefore, do not seem to be the result of pharmacokinetic factors, but rather of the processes differentially operating at the time of, as opposed to after, acquisition.
Several lines of evidence, in addition to studies of anticholinergic impairment of passive avoidance, do, however, support an involvement of the cholinergic system in post-training processes which influence re- tention performance. For example, impairment of retention in learning tasks other than passive avoidance, such as active avoidance, has been demonstrated following both post-training intracerebroventricular (Flood, Landry, & Jarvik, 1981) and peripheral (Flood & Cherkin, 1986) admin- istration of scopolamine. Antagonism of pretraining scopolamine-induced amnesia with post-training pharmacological treatments (Flood & Cherkin, 1986; Glick & Zimmerberg, 1972; Rush, 1986b) suggests that part of the pretraining effect of anticholinergics may be the result of muscarinic receptor blockade which occurs after the learning experience. In support of the pharmacological specificity of the effects of cholinergic antagonists, improvement of both active avoidance (Flood, Smith, & Cherkin, 1985) and passive avoidance with cholinergic agonists has also been shown (Baratti, Huygens, Mifio, Merlo, & Gardella, 1979; Gower, 1987; Har- outunian, Barnes, & Davis, 1985).
Only one of these studies, however, has provided a direct comparison of the size of pre- versus post-training effects. Glick and Zimmerberg (1972) found no difference in the ability of 1 mg/kg physostigimine ad- ministered either before or after training to antagonize an amnesia induced with 10 mg/kg scopolamine administered prior to training. Such com- parisons are necessary to determine the relative contribution of cholinergic stimulation (or blockade) on acquisition as opposed to postacquisition processes affecting memory retention. This study appears to be the only one in the literature in which pre- and post-training administration of cholinergic agonists on retention has been directly compared.
A greater number of comparisons has been carried out with cholinergic antagonists. The present experiments and those in the literature show that pretraining amnestic effects are much larger than those induced by post-training treatment (see Introduction), which suggests that the primary, though not sole, effect of anticholinergics is on processes operating during

270 DOUGLAS K. RUSH
the learning experience. Two recent studies employing spatial learning tasks lend further support to the results of investigations of retention of passive avoidance; in both, an impairment of learning with pretraining but not post-training anticholinergic treatment was found (Beatty & Bierley, 1986; Hagan, Tweedie, & Morris, 1986).
Studies in humans have also provided evidence that the primary, although perhaps not exclusive, effect of anticholinergics is on acquisition processes. In all of these studies, retention testing was conducted shortly after the administration of anticholinergics. Typically, acquisition of the to-be- remembered information also occurred after drug administration; that is, both learning and retrieval occurred under drug influence. In only three studies (Ghoneim & Mewaldt, 1975; Petersen, 1977; Mewaldt & Ghoneim, 1979) was retention of information (lists of words) acquired prior to drug administration examined; scopolamine had no effect on retrieval. These and other experiments, however, have shown profound effects on ac- quisition (see Beatty, Butters, & Janowsky, 1986; Caine, Weingartner, Ludlow, Cudahy, & Wehry, 1981, for references). As in the passive avoidance paradigm, these findings suggest that pretraining treatment induces a much stronger deficit than does treatment following training, although the experimental protocol used in these human studies differs from those conducted with animals. In animal experiments retention testing normally occurs at least 24 h after training, when the drug pre- sumably no longer is present to influence performance. Studies in humans utilizing the equivalent of this post-training paradigm have not been conducted.
Exactly which processes operative at the time of acquisition are adversely influenced by anticholinergics is not clear. Studies with animals have implicated effects on attention (Cheal, 1981) and discrimination (Milar & Dykstra, 1985). An adverse effect on short-term memory as measured, for example, in the radial maze (Eckerman, Gordon, Edwards, MacPhail, & Gage, 1980) has also been found although studies in operant short- term memory tasks have yielded conflicting results; some studies offer support for a specific effect on memory (e.g., Bartus, 1978; Dunnett, 1985), whereas others have attributed impaired performance to a generalized disruption of discrimination processes (e.g., Spencer, Pontecorvo, & Heise, 1985). Recent work in humans (Dunne & Hartley, 1986; Richardson, Frith, Scott, Crow, & Cunningham-Owens, 1984) has pointed to effects on encoding processes secondary to effects on discrimination or attention. Procedures for studying the means by which animals encode information into long-term storage are at present not available, although the results in short-term memory paradigms suggest a possible influence on a rehearsal process.
If the effects of scopolamine do mimic, at leat in part, the cognitive deficits of dementia, then the results of the present experiments suggest

SCOPOLAMINE AMNESIA OF PASSIVE AVOIDANCE 271
that the primary impairment in demented patients is not of memory storage, but rather of acquisition or encoding processes involving attention, discrimination, or short-term memory. Direct comparisons of aged humans or Alzheimer patients with normal subjects administered scopolamine (Beatty et al., 1986; Drachman & Leavitt, 1974; Fuld, 1984) indicate that anticholinergic medication, while inducing memory impairment, does not fully mimic the pattern of deficits found in dementia. Beatty et al. (1986), for example, found retrieval deficits, as assessed in a test of verbal fluency, in their sample of Alzheimer patients but no effect of scopolamine on this measure of cognitive performance.
The relevance of scopolamine-induced memory impairment in animals as a model of human dementia has also been recently questioned (Flood & Cherkin, 1986). A variety of compounds which affect various neuro- transmitter systems were all able to antagonize a scopolamine-induced active avoidance retention deficit, suggesting pharmacological nonspecificity of the antagonistic effects. Certainly this criticism has implications for substances which have been shown to counteract scopolamine-induced retention deficits, such as the opiate antagonist naloxone (Rush, 1986b) and the nootropics aniracetam (Cumin, Bandle, Gamzu, & Haefely, 1982; Pontecorvo & Evans, 1985) and tenilsetam (Schindler et al., 1984), but which are not known to influence the cholinergic system directly, for example by binding to muscarinic receptors.
Given the involvement of several neurochemical systems in learning and memory processes (Gold & Zornetzer, 1983), it is perhaps to be expected that interactions of these systems can take place in experimental models of memory deficit. Likewise it seems not unlikely that neuro- chemical pathways in addition to cholinergic ones are responsible for the complex symptoms of Alzheimer's disease (Gottfries, 1985). Further studies, especially comparative ones, with both animals and normal and clinical human populations are necessary to delineate the similarities and differences between naturally occurring and chemically induced memory disorder.
REFERENCES Bammer, G. (1982). Pharmacological investigations of neurotransmitter involvement in
passive avoidance responding: A review and some new results. Neuroscience & Bio- behavioral Reviews, 6, 247-296.
Baratti, C. M., Huygens, P., Mifio, J., Merlo, A., & Gardella, J. (1979). Memory facilitation with posttrial injection of oxotremorine and physostigmine in mice. Psychopharmacology, 64, 85-88.
Bartus, R. T. 0978). Evidence for a direct cholinergic involvement in the scopolamine- induced amnesia in monkeys: Effects of concurrent administration of physostigmine and methylphenidate with scopolamine. Pharamacology Biochemistry & Behavior, 9, 833-836.
Bartus, R. T., Dean, R. L., Beer, B., & Lippa, A. S. (1982). The cholinergic hypothesis of geriatric memory dysfunction. Science, 217, 408-417.

272 DOUGLAS K. RUSH
Beatty, W. W., Bierley, R. A. (1986). Scopolamine impairs encoding and retrieval of spatial working memory in rats. Physiological Psychology, 14, 82-86.
Beatty, W. W., Butters, N., & Janowsky, D. S. (1986). Patterns of memory failure after scopolamine treatment: Implications for cholinergic hypotheses of dementia. Behavioral and Neural Biology, 45, 196-211.
Bohdaneck~), Z., & Jarvik, M. E. (1967). Impairment of one-trial passive avoidance learning in mice by scopolamine, scopolamine methylbromide, and physostigmine. International Journal of Neuropharmacology, 6, 217-222.
Bure~ov~t, O., Buret, J., Bohdaneck~, Z., & Weiss, T. (1964). Effect of atropine on learning, extinction, retention and retrieval in rats. Psychopharmacologia 5, 255-263.
Bushnell, P. J. (1987). Effects of scopolamine on locomotor activity and metabolic rate in mice. Pharamacology Biochemistry & Behavior, 26, 195-198.
Caine, E. D., Weingartner, H., Ludlow, C. L., Cudahy, E. A., & Wehry, S. (1981). Qualitative analysis of scopolamine-induced amnesia. Psychopharmacology, 74, 74- 80.
Calhoun, W. H., & Smith, A. A. (1968). Effects of scopolamine on acquisition of passive avoidance. Psychoparmacologia, 13, 201-209.
Cheal, M. L. (1981). Scopolamine disrupts maintenance of attention rather than retrieval processes. Behavioral and Neurol Biology, 33, 163-187.
Coyle, J. T., Price, D. L., & DeLong, M. R. (1983). Alzheimer's disease: A disorder of cortical choliuergic innervation. Science, 219, 1184-1190.
Cumin, R., Bandle, E. F., Gamzu, E., & Haefely, W. E. (1982). Effects of the novel compound aniracetam (Ro 13-5057) upon impaired learning and memory in rodents. Psychopharmacology, 78, 104-I11.
Dilts, S. L., & Berry, C. A. (1967). Effect of cholinergic drugs on passive avoidance in the mouse. Journal of Pharmacology and Experimental Therapeutics, 158, 279-285.
Drachman, D. A., & Leavitt, J. (1974). Human memory and the cholinergic system: A relationship to aging? Archives of Neurology, 30, 113-121.
Dunne, M. P., & Hartley, L. R. (1986). Scopolamine and the control of attention in humans. Psychopharmacology, 89, 94-97.
Dunnett, S. B. (1985). Comparative effects of cholinergic drugs and lesions of nucleus basalis or fimbria-fornix on delayed matching in rats. Psychopharmacology, 87, 357- 363.
Eckerman, D. A., Gordon, W. A., Edwards, J. D., MacPhail, R. C., & Gage, M. I. (1980). Effects of scopolamine, pentobarbital, and amphetamine on radial arm maze performance in the rat. Pharmacology Biochemistry & Behavior, 12, 595-602.
Essman, W. B. (1983). Clinical pharmacology of learning and memory. Jamaica, New York: Spectrum.
Flood, J. F., & Cherkin, A. (1986). Scopolamine effects on memory retention in mice: A model of dementia? Behavioral and Neural Biology, 45, 169-184.
Flood, J. F., Landry, D. W., & Jarvik, M. E. (1981). Cholinergic receptor interactions and their effects on long-term memory processing. Brain Research, 215, 177-185.
Flood, J. F., Smith, G. E., & Cherkin, A. (1985). Memory enhancement: Supra-additive effect of subcutaneous cholinergic drug combinations in mice. Psychopharmacology, 86, 61-67.
Fuld, P. A. (1984). Test profile of cholinergic dysfunction and of Alzheimer-type dementia. Journal of Clinical Neuropsychology, 6, 380-392.
Ghoneim, M. M., & Mewaldt, S. P. (1975). Effects of diazepam and scopolamine on storage, retrieval and organizational processes in memory. Psychopharmacologia, 44, 257-262.
Gibbons, J. D. (1976). Nonparametric methods for quantitative analysis. New York: Holt, Rinehart and Winston.
Giordano, M., & Prado-Alcal{t, R. A. (1986). Retrograde amnesia induced by post-trial

SCOPOLAMINE AMNESIA OF PASSIVE AVOIDANCE 273
injection of atropine into the caudate-putamen. Protective effect of the negative reinforcer. Pharmacology, Biochemistry & Behavior, 24, 905-909.
Glick, S. D., & Greenstein, S. (1972). Differential effects of scopolamine and mecamylamine on passive avoidance behavior. Life Sciences, 11, 169-179.
Glick, S. D., & Zimmerberg, B. (1971). Comparative learning impairment and amnesia by scopolamine, phencyclidine, and ketamine. Psychonomic Science, 25, 165-166.
Glick, S. D., & Zimmerberg, B. (1972). Amnesic effects of scopolamine. Behavioral Biology, 7, 245-254.
Gold, P. E. (1986). The use of avoidance training in studies of modulation of memory storage. Behavioral and Neural Biology, 46, 87-98.
Gold, P. E., & Zornetzer, S. F. (1983). The mnemon and its juices: Neuromodulation of memory processes. Behavioral and Neural Biology, 38, 151-189.
Gottfries, C. G. (1985). Alzheimer's disease and senile dementia: Biochemical characteristics and aspects of treatment. Psychopharmacology, 86, 245-252.
Gower, A. J. (1987). Enhancement by secoverine and physostigmine of retention of passive avoidance response in mice. Psychopharmacology, 91, 326-329.
Hagan, J. J., Tweedie, F., & Morris, R. G. M. (1986). Lack of task specificity and absence of posttraining effects of atropine on learning. Behavioral Neuroscience, 10G 483- 493.
Haroutunian, V., Barnes, E., & Davis, K. L. (1985). Cholinergic modulation of memory in rats. Psychopharmacology, 87, 266-271.
Heise, G. A. (1981). Learning and memory facilitators: Experimental definition and current status. Trends in Pharmacological Sciences, 2, 158-160.
Jablonka, B, & Rush, D. K. (1987). The interaction of scopolamine and oxotremorine on motor activity and central muscarinic receptors. Nauyn-Schmiedeberg's Archives of Pharmacology, 335, R97.
Jarrard, L. E., Kant, G. J., Meyerhoff, J. L., & Levy, A. (1984). Behavioral and neurochemical effects of intraventricular AF64A administration in rats. Pharmacology Biochemistry & Behavior, 21, 273-280.
McGaugh, J. L. (1983). Preserving the presence of the past. Hormonal influences on memory storage. American Psychologist, 38, 161-174.
Mewaldt, S. P., & Ghoneim, M. M. (1979). The effects and interactions of scopolamine, physostigmine and methamphetamine on human memory. Pharmacology Biochemistry & Behavior, 10, 205-210.
Milar, K. S., & Dykstra, L. A. (1985). The effects of morphine and scopolamine on auditory discrimination in squirrel monkeys. Psychopharmacology, 85, 148-150.
Overton, D. A. (1966). State-dependent learning produced by depressant and atropine-like drugs. Psychopharmacologia, 10, 6-31.
Overton, D. A. (1984). State dependent learning and drug discrimination. In L. L. Iversen, S. D. Iversen, & S. H. Snyder (Eds.), Handbook ofpsychopharmacology (Vol. 18, pp. 59-127). New York: Plenum.
Petersen, R. C. (1977). Scopolamine induced learning failures in man. Psychopharmacology, 52, 283-289.
Pontecorvo, M. J., & Evans, H. L. (1985). Effects of aniracetam on delayed matching- to-sample performance of monkeys and pigeons. Pharmacology Biochemistry & Behavior, 22, 745-752.
Prado-Alcal~, R. A., Cruz-Morales, S. E., & L6pez-Miro, F. A. (1980). Differential effects of cholinergic blockade of anterior and posterior caudate nucleus on avoidance behaviors. Neuroscience Letters, 18, 339-345.
Richardson, J. T. E., Frith, C. D., Scott, E., Crow, T. J., & Cunningham-Owens, D. (1984). The effects of intravenous diazepam and hyoscine upon recognition memory. Behavioural Brain Research, 14, 193-199.

274 DOUGLAS K. RUSH
Ridley, R. M., Barratt, N. G., & Baker, H. F. (1984). Cholinergic learning deficits in the marmoset produced by scopolamine and 1CV hemicholinium. Psychopharmacology, 83, 340-345.
Rush, D. K. (1986a). Apple II assembly language routines for digital data acquisition and contingency control. Behavior Research Methods, Instruments, & Computers, 18, 32- 35.
Rush, D. K. (1986b). Reversal of scopolamine-induced amnesia of passive avoidance by pre- and post-training naloxone. Psychopharmacology, 89, 296-300.
Schindler, U., Rush, D. K., & Fielding, S. (1984). Nootropic drugs: Animal models for studying effects on cognition. Drug Development Research, 4~ 567-576.
Spencer, D. G., & Lal, H. (1983). Effects of anticholinergic drugs on learning and memory. Drug Development Research, 3, 489-502.
Spencer, D. G., Pontecorvo, M. J., & Heise, G. A. (1985). Central cholinergic involvement in working memory: Effects of scopolamine on continuous nonmatching and discrim- ination performance in the rat. Behavioral Neuroscience, 99, 1049-1065.
Vickroy, T. W., Watson, M., Leventer, S. M., Roeske, W. R,, Hanin, I., & Yamamura, H. I. (1985). Regional differences in ethylcholine mustard aziridinium ion (AF64A)- induced deficits in presynaptic cholinergic markers for the rat central nervous system. Journal of Pharmacology and Experimental Therapeutics, 235, 577-582.
Whitehouse, P. J., Price, D. L., Struble, R. G., Clark, A. W., Coyle, J. T., & DeLong, M. R. (1982). Alzheimer's disease and senile dementia: Loss of neurons in the basal forebrain. Science, 215, 1237-1239.
Wiener, N. I., & Messer, J. (1973). Scopolamine-induced impairment of long-term retention in rats. Behavioral Biology, 9, 227-234.
Woolf, N. J., Hernit, M. C., & Butcher, L. L. (1986). Cholinergic and non-cholinergic projections from the rat basal forebrain revealed by combined choline acetyltransferase and Phaseolus vulgaris leucoagglutinin immunohistochemistry. Neuroscience Letters, 66, 281-286.
Zwick, R., & Marascuilo, L. A. (1984). Selection of pairwise multiple comparison procedures for parametric and nonparametric analysis of variance models. Psychological Bulletin, 95, 148-155.