scanning electron microscopy in paleoanthropological research
TRANSCRIPT

Science, Technology and Education of Microscopy: an Overview
420
Scanning Electron Microscopy In
Paleoanthropological Research
Alejandro Romero* and Joaquín De Juan
Department of Biotechnology, Faculty of Science, University of Alicante,
Apdo. Correos 99, Alicante 03080, Spain
Keywords: paleoanthropology, image processing, fossil bone, dental microwear.
Abstract. Knowledge about the biology of ancient human populations has increased significantly thanks to new research technologies. In this way, paleobiology has gradually become an interdisciplinary field of study where progress in other sciences have been successfully applied. However, the traditional use of macroscopic and light microscopic techniques are insufficient in the analysis of specific biodynamic problems studied in paleoanthropology. Needless to say, the use of Scanning Electron Microscopy (SEM) has gradually been applied to several paleoanthropological materials. In this study, we review the most relevant aspects in modern paleobiology by using SEM. We pay special attention to the conceptual changes that affect the classical hypothesis about the feeding behavior of fossil hominids and ancient human groups when SEM analysis is applied. Introduction Our knowledge about the biological aspects of ancient human populations has increased significantly with the introduction of new research methods. After a transitional period when most publications were simply reviews of Physical Anthropology, a new stage began where data from the fossil bones records was reevaluated and correlated with new data on human evolution [1-2]. In this way, paleobiological research has gradually turned into an interdisciplinary field where progress in other disciplines has been successfully applied in order to obtain joint results. Macroscopic and even light microscopic techniques have been shown to be insufficient in the analysis of specific biodynamic problems and artefactual changes observed in paleoanthropological material. Needless to say, Scanning Electron Microscopy (SEM) has been gradually introduced into research on several paleoanthropological materials [3-7]. Much interest has been shown towards the use of SEM on this type of material because of its high-resolution power and depth of focus. Therefore, SEM is now a basic tool in some aspects of paleoanthropological research although it does also present some limitations in several types of application. From a technical point of view, the electron beam of the SEM scans the surface of the specimen to be analyzed. This enables us to view to study very small areas when we use a high magnification in order to obtain an electronic map of the sample. In the same way, the quality of the micrograph is linked to several technical aspects such as the use of dry and non-porous surfaces, the use of small specimens, etc. The use of SEM in paleoanthropological studies allows researchers to carry out new analysis techniques which focus on the study of small areas related to one particular area of study. The aim of many of the studies that use SEM is to analyze casts from "in vivo" and "in vitro" specimens [8-11] that have been coated with conductive materials (e.g. 15-20 nm of gold-

Science, Technology and Education of Microscopy: an Overview
421
palladium). By using a cast, the study can be carried out without damaging the original material from the museum. Another alternative is the use of a SEM equipped with an environmental chamber (WET mode), which is useful for examining the original specimens [12-14] without coating them. This method is very important because, in some cases, it is necessary to analyze the original paleoanthropological specimens in order to observe the effects produced by the geochemical environment where the bone fossil is found. On the other hand, the SEMs equipped with an x-ray microanalyzer unit can also offer qualitative data about the chemical elements present in the specimens [15-16] (Fig. 1A-B). This is particularly useful in the study of postmortem microstructural changes in forensic science or in paleoanthropological research [17-18]. Finally, the spectacular advances in computer technology and image processing techniques have provided more precise information about the teeth and fossil bones.
Fig. 1. Post mortem changes in the cortical fraction of human femur (ss. X-XI AD): (A) geochemical environment (top) and micro-organisms parasitation (down). BSE image at 500x. (B) X-ray microanalysis results (x3500) (see area fig.1A). Here, we have reviewed the most relevant aspects of modern paleobiology using SEM. We focus on the conceptual changes that affect the classical hypothesis about the feeding behavior of fossil hominids and ancient human groups when SEM analysis is applied. Toothmarks and Cutmarks on bone surface using SEM There are many publications about the late Pleistocene and early Holocene period, with important data about the fauna found in the hominid group's habitat. The belief that Australopithecus was a predator [19-20], together with the discovery of pools of animal bones near stone artifacts, has been the center of studies carried out on the earliest hominids and their habitats. In fact, these studies have focused on interpreting the significance of these places and economic activities related to finding meat for food. This way of obtaining protein resources has been considered as cause and effect of adaptive and evaluative changes on the brain [21]. However, fossil specimens, which may have often been hidden for several million years, can undergo changes that hinder an accurate interpretation. It is not always clear if the bones found were associated with hominid foodstuffs. This is why research looks for found definitive evidence on the superficial bone marks [22]. The use of SEM to analyze the marks on the bone surface, appears to be a very useful analysis tool. In fact, one of the main purposes of applying SEM is to establish criteria that allows marks caused by several agents to be distinguished: carnivore teeth, stone tool action and other agents [23-25]. In this way, it is possible to establish the microscopical specific pattern of the bone marks and its relationship with an actor (hominid or carnivore) from signals produced on the bone by teeth, stone flakes, cutting edge, or hammerstone (effector) [26]. This criteria is extremely important for interpreting, for example, the possible predatory behavior of hominids when trying to distinguish bone fractures that result from several different actions (hunting, taphonomic processes,

Science, Technology and Education of Microscopy: an Overview
422
cannibalism, etc.). In other words, this is a vital key to understanding the behavior of early primates. SEM provides substantial evidence of anthropophagy in human bones in the Spain Lower Pleistocene [27-28] or France Neolithic periods [29]. Toothmarks form a U-shape in the cross-section which has a wide breadth when observed with SEM. However, cut marks have a different morphology. In fact, when a stone tool is used for making a cutting or slicing action, the cut mark is V-shaped and has narrower breadth in the longitudinal microstriations (Fig. 2A). On the other hand, more information about the origin of cutmarks is provided by its orientation and "Hertzinian cones" [30-31]. In fact, the orientations make it possible to determine several different actions on the bone surface such as carcass processing, handedness, etc. Recently, opto-electronic scanning methods [32] have enabled a bone surface topography map (3D-surface model) to be created which introduces new methods in the electron microscopy study of fossil bone surfaces. SEM analysis in Teeth Because of the extreme durability of enamel, teeth are the most common, adn often only, fragments of fossil primates to be found during excavations. Furthermore, teeth are a very good indicator of the different aspects of hominid fossil life such as: masticatory and non-masticatory uses, age at death or pathological process. The use of SEM in paleoanthropology has acquired a growing interest and, in most cases, is the only methodological tool for this type of study. Grooves on fossil hominid teeth In fossil dental fragments of ancient humans, as well as masticatory marks, there also appear others that suggest a non-masticatory use and probably result from an exogenous influence. These influences can be intentional or unintentional [33] and reflect behaviors derived from particular habits and lifestyles. Several research studies have paid special attention to interproximal grooves in human and fossil hominid skeletal series. Manipulation of the interproximal teeth spaces using flexible (e.g. bone and wood awls, vegetal and leader fibres, vegetables) or inflexible tools is known as the “toothpicking hypothesis”. If this hypothesis is valid, it should be possible to observe this type of mark with the same gross and microscopic appearance in different specimens. Macroscopically, the concave groove is located at or near the enamel-cementum line either on the mesial or distal tooth surface. These marks are mainly observed in molars (M) or pre-molars (Pm) and their dimension ranges from 1 to 4mm in diameter. SEM-studies show an oval outline with a round concavity that ends abruptly in the lingual direction. This suggests that interproximal abrasion was repeatedly manipulated with a relatively hard object not associated with feeding and shows multiple fine parallel scratches in a buccolingual direction. This confirms that the scratches are of an artificial origin [34] (Fig. 2B). The SEM study of these grooves in a hominid tooth from the Olduvai Gorge (Tanzania, Africa) [35], suggests the earliest use of a toothpick by our Plio-Pleistocene ancestors. However, these habits have also been recorded in teeth from Middle and early Upper Pleistocene hominids [36-38] and in Holocene bioarchaeological samples [39-44]. Evidence of previous tooth use in hominids When using SEM, some authors have observed special marks in tooth samples on the labial surfaces of the maxillary central incisors (I1s) in Middle-Late Pleistocene fossil hominids from Europe and the Levant region. These marks are microscopic scratches resulting from food ingestion and the use of the frontal teeth as a grasping tool to hold the food (meat) during the cutting process. In fact, these marks represent the expression of stone knife abrasion behaviors. The direction of these striations can provide evidence of preferential right-handed “stuff-and-cut” behaviors [45].

Science, Technology and Education of Microscopy: an Overview
423
Fig. 2. A. Three cutmarks on ovicaprid humerus from a Roman period site (s. I AD). Bar: 0.5mm. B. Micrograph taken on interproximal groove of mesial surface (M1) from Eneolithic human period (see parallel striations running buccolingually. Top: occlusal, left: lingual side of the tooth. Bar: 100µm. C. Micrograph taken just beyond the incisal edge near the midsagittal plane on tooth enamel(I1) from a Bronze Age human skull (see feature in V-section). Bar: 50 µm. In a sample of Middle Pleistocene hominids a microscopic pattern of scratches caused by the cutting process has been observed [46] and similar results have been noted in the Kaprina Neanderthal [47]. This kind of SEM-microscopic feature presents a particular preferential orientation and different width in comparison with marks caused by dust or endogenous silicates or grit in the foodstuffs. On the other hand, these scratches resemble to bone marks produced by stone tools [48, 23] (Fig. 2C). However, some results in modern people suggest that there is no relationship between handedness and a preference for right oblique striations [49], although it probably offers important information about survival techniques [50]. Most of these features can only be observed using high magnifications in a SEM analysis and are their characteristics are then related to feeding behaviors or toothbrush marks. SEM implications for development and structure evaluation In paleoanthropological research dental attrition is used to date the age of past peoples based on the assumption that a close relationship exists between chronological age and dental attrition severity. However, anthropologists have observed some limitations to these marks as being signs of aging due to dental wear. In fact, these marks can be effected by the influence of several factors (individual, gender, cultural, feeding behavior, more or less abrasive diet, etc.). For this reason, a closer relationship needs to be established between the physical data on other bone marks, more statistical rigor in the use of life tables [51] and the application of different methods of data validation [52]. But what can microscopic evaluation tell us about these aspects? Incremental markings and growth layers in enamel are particularly clear and this may reflect the speed with which enamel matrix initially mineralizes after secretion by ameloblasts [53]. These markings (cross striations and striae of Retzius) are visible microscopically. On the other hand, during the enamel tooth development, there are marks or perikymata that reflect its growth (Fig. 3A). The perikymata have been used to determine crown formation times to estimate an age at death in modern humans and fossil hominids [54-57]. These indicators are found where the Retzius striae joins the enamel surface during crown dental formation. Finally, SEM analysis of tooth enamel ultra-structural features reveal varying prism divisions. The electron microscopic examination of enamel thickness allows various mammals and Hominoidea [58-59] to be classified into specific phylogenetic and taxonomic groups. Different morphological studies of gross enamel prisms on teeth of extinct Pongidae and hominids have revealed varying structures [60-62] (Fig. 3B).

Science, Technology and Education of Microscopy: an Overview
424
Fig. 3. A. Enamel bands (perikymata). Bar: 100 µm. B. Micrograph of enamel prism pattern in Homo sp. sp. (Neolithic period). Bar: 25µm. Dental wear attrition The loss of dental substance during mastication is due to attrition and abrasion. The existence of varying degrees is due either to the strength and intensity of mastication or to the type of intrinsic and extrinsic particles (grit, ash, phytoliths) ingested by each individual. The macroscopic study of dental wear has been used by paleoanthropologists to analyze the degree of abrasivity exerted by the diet in large samples of ancient human groups [63-65]. However, as has been explained above, there are several factors that may interfere in the interpretation of qualitative studies of dental wear in determining age. One example is the relationship between abrasivity and diet. It is for this reason that qualitative analyses must be combined with microscopic studies, using quantitative image processing techniques in the study of occlusal topography. More recently, a new methodology has been developed to examine the shape of worn teeth. This methodology involves using Geographic Information Systems (GIS) for the study of occlusal surface topographic models. In this way, a digital elevation model is generated for each cusp in order to obtain 3-D landscapes [66-67] that can be used to map the occlusal surfaces of human and non-human primate molars. Dental microwear analysis Dental microwear analysis of the enamel, is the most thorough technique for classifying the physical properties of our ancestors and fossil primates’ diets. With this methodology, it is possible to analyse the intergroupal variability as regards feeding behaviors. It was at the end of the 1970´s that SEM was introduced to study the degree of abrasion of the diet on the dental enamel of humans, non-human primates and faunal remains to test dietary hypotheses on extinct hominoid teeth. The dental enamel surfaces present microscopic features (pitted surfaces or parallel striations) due to different physical consistency of the food during the masticatory action. These features reveal different patterns in grazers, browsers, carnivores, frugivores, scavengers or bone eaters [68-72]. The application of SEM in paleoanthropological research has shown that the robust Australopithecus is not only a grass seed eater and bone crushing scavenger. Dental microwear shows that Australopithecus follow a frugivore pattern similar to mandrills, chimpanzees and orangutans [73]. Slight scratches in gracile forms of Australopithecines prove that they consume softer foods than robust forms, which show a more pitted pattern [74]. The use of image analysis in paleanthropology marks a highly important achievement and permits statistical and qualitative data to be compared when analyzing the results of different specimens. This methodology provides a better definition of tooth and enamel models and micrographic quantitative morphological criteria (scratches are linear features with a discernible angle of orientation, by contrast, pit features are approximately equal in length and breadth and to observe

Science, Technology and Education of Microscopy: an Overview
425
pit-to-striation frequencies, feature orientation, length and width) [75-76]. On the other hand, there are other variables to be taken into account, such as micrographs, electron-type (backscatterers – BSEs - or secondary – SE -), accelerating voltage (Kv), working distance (WD) or image resolution (Fig.4A-F, Table 1). There is extensive literature about dietary in differences humans, non-humans and fossil primates, from the quantification of microwear pit or scratch percentages where more folivorous taxa had relatively more scratches than frugivores [77-79]. However, the proportions of pits or scratches may vary significantly between surfaces (occlusal and non-occlusal) or in different areas on the same surface (shearing, grinding or crushing) (Fig.4A, Fig.5A-E, Table 2). Several software packages have been applied in paleoantropological studies in order to analyze the variables. However, there are specific packages for semi-automated image analysis which quantify dental microwear [83-84]. With these programs, it is possible to obtain metrical information from one single SEM micrograph. This is the most difficult task to perform in the research process as the individual marks must be quantified in each of the micrographs obtained. The final aim is to obtain feature quantification. The combined use of image processing and quantitative techniques [85] can be used to distinguish different shapes. Using digitalized micrographs of the enamel surface, some authors apply a power spectra analysis and obtain the corresponding Fourier transformate [86-87]. The Fourier series are formed by sins and cosines from multiples of a variable that allows a convergent series to be developed for some types of discontinuous variables.
Science, Technology and Education of Microscopy: an Overview
426
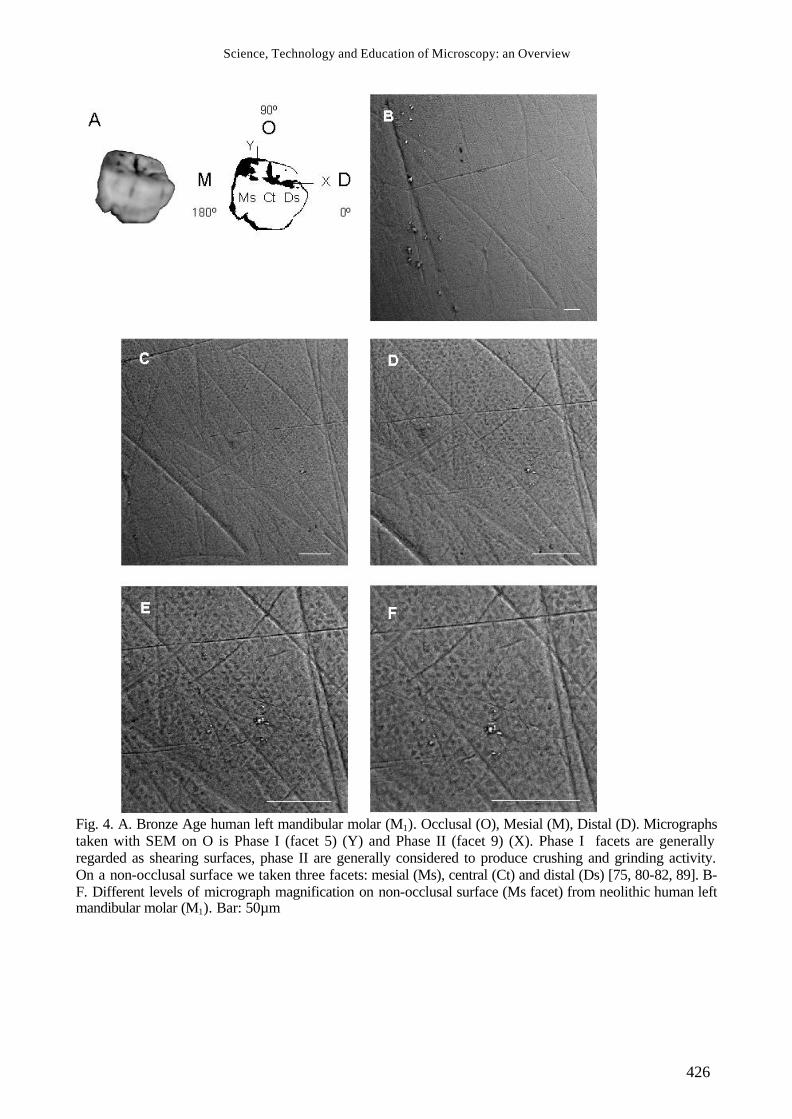
Fig. 4. A. Bronze Age human left mandibular molar (M1). Occlusal (O), Mesial (M), Distal (D). Micrographs taken with SEM on O is Phase I (facet 5) (Y) and Phase II (facet 9) (X). Phase I facets are generally regarded as shearing surfaces, phase II are generally considered to produce crushing and grinding activity. On a non-occlusal surface we taken three facets: mesial (Ms), central (Ct) and distal (Ds) [75, 80-82, 89]. B-F. Different levels of micrograph magnification on non-occlusal surface (Ms facet) from neolithic human left mandibular molar (M1). Bar: 50µm

Science, Technology and Education of Microscopy: an Overview
427
Fg
M A nt AxL O nst SL SB O nsc
B X1
00 0.455 32 330.1±16
6.1 119.9±34 32 0
C X200
0.114 41 162.1±83 112.4±42.8
40 266.6 12.04 134.24 1
D X300
0.051 33 117.6±61.4
129.7±48.2
26 152.8±64.1
5.09±2.011
123.8±30.2
7
E X400
0.028 30 91.3±42.8
149.9±54.7
20 120.1±52.4
4.13±1.761
118.3±32.9
10
F
X500
0.018 23 74.5±38.6
117.1±55.3
16 112±34.2
3.2±1.065
99.8±49.3
7
Table 1. Figures 4B-F; M, micrograph magnification; A, area (mm2); AxL, mean axis length (in µm); O, preferred orientation (in degrees) ; SL, mean scratch length (in µm); SB, scratch breadth (in µm); nt, total number of features; nst, total number of striations; nsc, total number of scratches. All micrographs were analyzed using a semi-automated image analysis software package, Microwear 3.0 [84]
Fig. 5. Micrographs of different facets of molar surfaces (see Fig. 4A). Areas on occlusal surface approx.0.04 mm2, Y facet (A) and X facet (B). In nonocclusal (buccal) approx.0.05 mm2, Ms facet (C), Ct (D), Ds (E). Bar: 50 µm. In occlusal facets it is possible to see different features on both phases [80-81]. Features on buccal surface are different since they do not come into regular contact with opposing teeth.

Science, Technology and Education of Microscopy: an Overview
428
Fg
F Features Scratches Pits
MaxL Po nt xL xB po n xL xW n 8A
Y 99.6±50.6
179.4±36.2
24 102.8±53.3
2.5±0.9 178.7±37.0
18
8B
X 37.9±35.3
31.2±4
6.6
62 62.5±30.
3
2.4±0.9 13.5±26.
3
34
8.1±4.2
5.9±3.1
28
8C
Ms
95.3±52.7
83.9±3
1.2
56 164.1±60.1
3.26±1.7 79.6±15.
9
7
8D
Ct
94.7±39.9
71.3±3
1.7
74 141.3±41.1
2.59±0.7 62.3±23.
1
10
8E
Ds
93.0±51.2
65.7±3
1.1
76 218.6±23.8
4.71±2.1 72.4±5.6 4
Table 2. Dental microwear analysis of the figures in this text (Fg), facet examined (F) (see Fig. 4A). General pattern of features (nt): mean major axis length (MaxL), preferred orientation (po). Scratches and pits examined: mean axis length (xL), mean scratch breadth (xB), mean pit width (xW). Its application on Australopithecus and Paranthropus [88] revealed, in the former, bands concentrated at right angles to the predominant longer axes of the wear scratches. In the latter, the spectrum is more diffuse with high frequencies of occlusal pits. Paranthropus are classified as having a more abrasive diet. Although this data was previously known [74,89], thresholding techniques have been used to isolate features from the image [90]. Others studies have used a substraction of smoothed versions of digital images in which the larger features of surface relief are removed [91]. References [1] S.L. Washburn: Transact. New York Acad. Sci. Vol. 13 (1951), p. 298 [2] S.M. Garn: Am. J. Phys. Anthropol. Vol. 21 (1963), p. 225 [3] S. Jarcho: Human Paleopathology (New Haven, Yale UP 1966). [4] R.F. Macadam, A.T. Sandison: Med. Hist. Vol. 13 (1969), p. 81 [5] A. Ascenzi: in Science in Archaeology, D. Brothwell, E. Higgs (eds.) (Thames & Hudson, London 1969), p. 526 [6] D. Brothwell : in Science in Archaeology, D. Brothwell, E. Higgs (eds.) (Thames & Hudson, London 1969), p. 564 [7] S. Olsen (ed.): Scanning Electron Microscopy in Archaeology (B.A.R. Int. Series. Vol. 452, Oxford 1988). [8] J.J. Rose: Am. J. Phys. Anthropol. Vol. 62 (1983), p. 255 [9] T.G. Bromage: Scanning Microsc. Vol. 1 (1987), p. 607 [10] A.D. Beynon: Scanning Microsc. Vol. 1 (1987), p. 663 [11] M.F. Teaford, O.J. Oyen: Am. J. Phys. Anthropol. Vol. 80 (1989), p. 73 [12] P.D. Taylor: Paleontology. Vol. 29 (1986), p. 685 [13] S.M. McCormack, F.J. Tormo, J.D.B. Featherstone: Scanning Microsc. Vol. 5 (1991), p. 269

Science, Technology and Education of Microscopy: an Overview
429
[14] T. King, P.Andrews, B. Boz: Am. J. Phys.Anthropol. Vol. 108 (1999), p.359 [15] J.B. Lambert, S.V. Simpson, J.E. Buikstra, D. Hanson: Am. J. Phys. Anthropol. Vol. 62 (1983), p. 409 [16] R.L. Ciochon, D.R. Piperno, R.G. Thompson: Proc. Natl. Acad. Sci., USA, Vol. 87 (1990), p. 8120 [17] L.S. Bell, M.F. Skinner, S.J. Jones: Forens. Sci. Int. Vol. 82 (1996), p. 129 [18] A.C. Walker: in A.K. Behrensmeyer, A.P. Hill (eds.) Fossils in the Making (Univ. Chicago Press, Chicago 1980), p. 182 [19] R. Dart: Am. J. Phys. Anthropol. Vol. 7 (1949), p. 1 [20] R. Ardrey: The Hunting Hypothesis: A personal Conclusion Concerning the Evolutionary Nature of Man (New York, Atheneum 1976). [21] L.C. Aiello, P. Wheeler: Curr. Anthropol. Vol. 36 (1995), p. 199 [22] H.T. Bunn: Nature Vol. 291 (1981), p. 574 [23] R. Potts, P. Shipman: Nature Vol. 291 (1981), p. 577 [24] A.K. Behrensmeyer, K.D. Gordon, G.T. Yanagi: Nature Vol. 319 (1986), p.768 [25] R.J. Blumenschine, M.M. Selvaggio: Nature Vol. 333 (1988), p.763 [26] R.J. Blumenschine, C.W. Marean, S.D. Capaldo: J. Archaeol. Sci. Vol. 23 (1996), p. 493 [27] Y. Fernández-Jalvo, J.C. Díez, J.Mª. Bermúdez de Castro, E. Carbonell, J.L. Arsuaga: Science Vol. 271 (1996), p. 277 [28] Y. Fernández-Jalvo, J.C. Díez, I. Cáceres, J. Rosell: J. Hum. Evol. Vol. 37 (1999), p. 591 [29] P.Villa, C. Bouville, J. Courtin, D. Helmer, E. Mahieu, P. Shipman, G. Belluomini, M. Branca: Science Vol. 233 (1986), p. 431 [30] P. Shipman, J. Rose: J. Anthropol. Archaeol. Vol. 2 (1983), p. 57 [31] T.G. Bromage, A. Boyde: Am. J. Phys. Anthropol. Vol. 65 (1984), p. 359 [32] T. Kaiser, H. Katterwe: Int. J. Osteoarchaeol. Vol. 11 (2001), p. 350 [33] K.W. Alt, S.L. Pichler: in K.W. Alt, F.W. Rösing, M. Teschler-Nicola (eds.), Dental Anthropology: Fundamentals, limits, prospects (Gustav-Fischer, Stuttgart 1998), p. 387 [34] G.R. Milner, C.S. Larsen: in M.A. Kelley, C.S. Larsen (eds.), Advances in Dental Anthropology (Alan R. Liss, New York 1991), p. 357 [35] P.S. Ungar, F.E. Grine, M.F. Teaford, A. Pérez-Pérez: Arch. Oral Biol. Vol. 46 (2001), p. 285 [36] D.W. Frayer, M.D. Russel: Am. J. Phys. Anthropol. Vol. 74 (1987), p. 393 [37] C. Lalueza, A. Pérez-Pérez, D. Turbón: J. Hum. Evol. Vol. 24 (1993), p. 281 [38] J.Mª. Bermúdez de Castro, J.L. Arsuaga, P.J. Pérez: Am. J. Phys. Anthropol. Vol.102 (1997), p. 369 [39] D.H. Ubelaker, T.W. Phenice, W.M. Bass: Am. J. Phys. Anthropol. Vol. 30 (1969), p. 145 [40] J.A. Wallace: Am. J. Phys. Anthropol. Vol. 40 (1974), p. 385 [41] J.Mª. Bermúdez de Castro, J.L. Arsuaga: L´Anthropologie Vol. 87 (1983), p. 521 [42] J.Mª. Be rmúdez de Castro, P.J. Pérez: Paleopathol. Newsl. Vol. 54 (1986), p. 5 [43] J.R. Luckas, R.F. Pastor: Am. J. Phys. Anthropol. Vol. 76 (1988), p. 377 [44] T. Brown, S. Molnar: Am. J. Phys. Anthropol. Vol. 81 (1991), p. 545 [45] C.L. Brace: Curr. Anthropol. Vol. 16 (1975), p. 396 [46] J.Mª. Bermúdez de Castro, T. Bromage, Y. Fernández -Jalvo: J. Hum. Evol. Vol. 17 (1988), p. 403 [47] C. Lalueza Fox, D.W. Frayer: Int. J. Osteoarchaeol. Vol. 7 (1997), p. 133 [48] K.D. Gordon: Am. J. Phys. Anthropol. Vol. 63 (1984), p. 315 [49] J.S. Bax, P.S. Ungar: Int. J. Osteoarchaeol. Vol. 9 (1999), p. 189 [50] P.S. Ungar, M.A. Spencer: Am. J. Phys. Anthropol. Vol 109 (1999), p. 387 [51] L.W. Konigsberg, S.R. Frankenberg: Am. J. Phys. Anthropol. Vol. 89 (1992), p. 235 [52] D. Lucy, A.M. Pollard, C.A. Roberts: J. Archaeol. Sci. Vol. 22 (1995), p. 417 [53] M.C. Dean: J. Hum. Evol. Vol. 16 (1987), p. 157 [54] T.G. Bromage, M.C. Dean: Nature Vol. 317 (1985), p. 525 [55] M.C. Dean, C.B. Stringer, T.G. Bromage: Am. J. Phys. Anthropol. 70 (1986), p. 301

Science, Technology and Education of Microscopy: an Overview
430
[56] A.D. Beynon, B.A. Wood: Nature Vol. 326 (1987), p. 493 [57] A.E. Mann, J.M. Monge, M. Lampl: Am. J. Phys. Anthropol. Vol. 86 (1991), p. 175 [58] A. Boyde: Proc. Roy. Soc. Med. Vol. 60 (1967), p. 13 [59] D.G. Gantt, D. Pilbean, G.P. Steward: Science Vol. 198 (1977), p. 1155 [60] S. Molnar, D.G. Gantt: Am. J. Phys. Anthropol. Vol. 46 (1977), p. 447 [61] E.S. Vrba, F.E. Grine: Science Vol. 202 (1978), p. 890 [62] L. Martin: Nature Vol. 314 (1985), p. 260 [63] D.R. Brothwell: Digging Up Bones. (British Museum (Natural History), London 1963). [64] E.C. Scott: Am. J. Phys. Anthropol. Vol. 51 (1979), p. 203 [65] B.H. Smith: Am. J. Phys. Anthropol. Vol. 63 (1984), p. 39 [66] L.F. Zuccotti, M.D. Williamson, F.W. Limp, P.S. Ungar: Am. J. Phys. Anthropol. Vol. 107 (1998), p. 137 [67] P. Ungar, M. Williamson: Palaeontologia Electronica. Vol. 3 (2000), 18 p. [68] G.V. Shkurkin, A.J. Almqvist, A.A. Pfeihofer, E.L. Stoddard: J. Dent. Res. Vol. 54 (1975), p. 401 [69] P.F. Puech: in H. de Lumley (ed.), La Prehistoire Francaise (C.N.R.S., Paris 1976), p. 709 [70] P.L. Walker: Am. J. Phys. Anthropol. Vol. 45 (1976), p. 299 [71] J.M. Rensberger: in P.M. Butler, K.A. Joysey (eds.), Development, Function and Evolution of Teeth (Academic Press, London 1978), p. 415 [72] A. Walker, H.N. Hoeck, L. Perez: Science Vol. 201 (1978), p. 908 [73] A.C. Walker: Philosoph. Trans. Roy. Soc. London Vol. 292 (1981), p. 57 [74] F.E. Grine: S. Afr. J. Sci. Vol. 77 (1981), p. 203 [75] K.D. Gordon: Am. J. Phys. Anthropol. Vol. 59 (1982), p. 195 [76] K.D. Gordon: Scanning Microsc. Vol. 2 (1988), p. 1139 [77] M.F. Teaford: Scanning Microsc. Vol. 2 (1988), p. 1149 [78] M.F. Teaford: Evol. Anthropol. Vol. 3 (1994), p. 17 [79] J.C. Rose, P.S Ungar: in K.W. Alt, F.W. Rösing, M. Teschler-Nicola (eds.), Dental Anthropology: Fundamentals, limits, prospects (Gustav-Fischer, Stuttgart 1998), p. 349 [80] R.F. Kay, K.M. Hiiemae: Am. J. Phys. Anthropol. Vol. 40 (1974), p. 227 [81] M.F. Teaford: in M.A. Kelley, C.S. Larsen (eds.), Advances in Dental Anthropology (Alan R. Liss, New York 1991), p. 341 [82] A. Romero, J. De Juan: in Proceedings of the Microscopy, Barcelona 2001 (Universitat de Barcelona, Barcelona 2001), p. 239 [83] P.S. Ungar, J.C. Simons, J.W. Cooper: Scanning Vol. 13 (1991), p. 31 [84] P.S. Ungar: Scanning Vol. 17 (1995), p. 57 [85] J. Frank, B. Shimkin, H. Dowse: Ultramicroscopy Vol. 6 (1981), p. 343 [86] K.R. Castleman: Digital Image Processing (Prentice Hall Inc., Englewood NJ 1979). [87] R.F. Kay: Scanning Microsc. Vol. 1 (1987), p. 657 [88] F.E. Grine, R.F. Kay: Nature Vol. 333 (1988), p. 756 [89] F.E. Grine, J. Hum. Evol. Vol. 15 (1986), p. 783 [90] P.L. Walker, S.A. Bernstein, K.D. Gordon: Am. J. Phys. Anthropol. Vol. 72 (1987), p. 267. [91] A. Boyde, M. Fortelius: Scanning, Vol. 13 (1991), p. 429