role of gpib in phagocytosis of platelets by macrophages
TRANSCRIPT

2090 TRANSFUSION
Volume 46, December 2006
Blackwell Publishing IncMalden, USATRFTransfusion0041-11322006 American Association of Blood BanksDecember 2006461220902099Original Article
PLATELET GPIb
a
AS TRIGGER FOR PHAGOCYTOSISBADLOU ET AL.
ABBREVIATIONS:
AA
=
amino acids; GlcNAc
=
N
-
acetylglucosamine; GP
=
glycoprotein; PGI
2
=
prostacyclin; PMA
=
phorbol 12-myristate 13-acetate; TAPI
=
tumor necrosis factor-
α
protease inhibitor.
From the Thrombosis and Haemostasis Laboratory, DCCH, the
Department of Immunology, UMC-Utrecht, the Institute for
Biomembranes Utrecht University, and the Sanquin Blood Bank
Region North-West, Utrecht, the Netherlands; and the Laboratory
for Thrombosis Research, IRC, KU Leuven Campus, Kortrijk
Belgium.
Address reprint requests to:
Prof. Dr. J.W.N. Akkerman,
Thrombosis and Haemostasis Laboratory (G.03.647), DCCH,
University Medical Center Utrecht, Heidelberglaan 100, 3584 CX
Utrecht, the Netherlands; e-mail: [email protected].
This work was supported by Sanquin Blood Supply
Foundation (Grant PPO 01.019). JWNA is supported by the
Netherlands Thrombosis Foundation.
Received for publication March 8, 2006; revision received
May 24, 2006, and accepted May 30, 2006.
doi: 10.1111/j.1537-2995.2006.01034.x
TRANSFUSION
2006;46:2090-2099.
C E L L P H Y S I O L O G Y
Role of glycoprotein Ib
a
in phagocytosis of platelets by macrophages
Bahram A. Badlou, Gerrit Spierenburg, Hans Ulrichts, Hans Deckmyn, W. Martin Smid, and
Jan-Willem N. Akkerman
BACKGROUND:
Platelet (PLT) storage at 0 to 4
°
C suppresses bacterial multiplication, but induces clusters of glycoprotein (GP) Ib
α
that trigger their phagocytosis by macrophages and reduce their survival after transfusion. A method was sought that detects cold-induced changes in GPIb
α
involved in phagocytosis.
STUDY DESIGN AND METHODS:
Human PLTs were isolated and stored for up to 48 hours at 0
°
C. Binding of a phycoerythrin (PE)-labeled antibody directed against amino acids (AA) 1-35 on GPIb
α
(AN51-PE) was compared with phagocytosis of PLTs by matured monocytic THP-1 cells, analyzed by fluorescence-activated cell sorting.
RESULTS:
Freshly isolated PLTs were detected as a single population of AN51-PE–positive particles and showed less than 5 percent phagocytosis. Cold storage led to a decrease in AN51-PE binding and an increase in phagocytosis.
N
-Acetylglucosamine, known to interfere with macrophage recognition of GPIb
α
clusters, restored normal AN51-PE binding to cold-stored PLTs and suppressed phagocytosis.
CONCLUSIONS:
It is concluded that binding of an antibody against AA 1-35 on GPIb
α
reflects changes in GPIb
α
that make PLTs targets for phagocytosis by macrophages.
he storage of platelet (PLT) concentrates at roomtemperature facilitates bacterial multiplicationand introduces changes in PLTs indicative foractivation and apoptosis.
1-3
Improvements havebeen sought in lowering the storage temperature therebyreducing bacterial growth and possibly prolonging thestorage period.
1,4-6
A disadvantage of cold storage is that itinduces the so called cold-induced PLT lesion with shapechange and actin reassembly,
7
myosin activation, andan increase in cytosolic Ca
2
+
.
8
In addition, cold storageinduces clusters of the von Willebrand factor (VWF) recep-tor glycoprotein (GP) Ib
α
, which are recognized by inte-grin
α
M
β
2
(Mac-1, CR3) receptors on liver macrophagestriggering rapid PLT destruction in vitro and their removalfrom the circulation in vivo.
6
GPIb
α
clusters expose
N
-acetylglucosamine (GlcNAc) to which
α
M
β
2
binds throughthe integrin’s affinity for N-linked glycans. Enzymaticgalactosylation of chilled PLTs blocks
α
M
β
2
recognitionand prolongs the circulation of cold-stored PLTs.
9
T

PLATELET GPIb
a
AS TRIGGER FOR PHAGOCYTOSIS
Volume 46, December 2006
TRANSFUSION 2091
GPIb
α
is a member of the GPIb-V-IX complex and amajor sialoglycoprotein with multiple N- and O-linkedoligosaccharides.
10-12
The complex is formed in the endo-plasmic reticulum and transported to the Golgi for furthermodification.
13
GPIb
α
serves in the transient binding ofPLTs to activated VWF at sites of vascular damage slowingdown their velocity in flowing blood and enabling otherreceptors to firmly attach PLTs to the wound.
14
Inter-estingly, the cold-induced rearrangement of GPIb
α
leavesthe hemostatic functions of the receptor unchanged.The VWF-binding site is located within amino acids (AA)36-200 at the N-terminal part of GPIb
α
, which containsseven leucine-rich repeats. It is part of glycocalicin, aregion of GPIb
α
that is rapidly released by proteolyticdegradation.
12,15,16
The expression of GPIb
α
on the PLT surface is readilydetected by fluorescence-activated cell sorting (FACS)analysis after incubation with a phycoerythrin (PE)- orfluorescein isothiocyanate (FITC)-labeled monoclonalantibody (MoAb). Storage at room temperature is knownto inflict changes on the PLT surface that decrease anti-body binding to GPIb
α
leading to a shift to lower fluores-cence intensity.
17
This is the result of proteolytic cleavageby the metallopeptidase tumor necrosis factor (TNF)-
α
–converting enzyme or ADAM-17.
18,19
ADAM-17 is a plasmaconstituent and present in PLT concentrates prepared inmixtures of additive solution and plasma. GPIb
α
sharesthe sensitivity to proteolytic damage with other PLT sur-face receptors such as GPV
20
and GPVI.
21
In the course of our studies on PLT storage, we foundthat the binding of a PE-labeled MoAb directed against AA1-35 in the N-terminal flank of the leucine-rich region ofGPIb
α
named AN51 decreases during storage at 0
°
C. Thisis a condition in which PLTs express properties that makethem targets for phagocytosis by macrophages. In thisstudy we investigated the nature of the affinity loss forAN51 and its possible impact on the mechanisms thatcontrol the binding of PLTs to macrophages and theirdestruction.
MATERIALS AND METHODS
We obtained phorbol 12-myristate 13-acetate (PMA),mepacrine (Quinacrine), mannose, and GlcNAc fromSigma Chemicals (Mannheim, Germany). The anti-human CD42b (GPIb
α
)-PE (R7014 clone AN51 directedagainst AA 1-35 of the N-terminal flank of GPIb
α
), anti-human CD14-FITC, and FITC-labeled immunoglobulin Gas a negative control were from Dako A/S (Glusdorp, Den-mark). Serum-free cell culture medium RPMI 1640 wasfrom Corning Inc. (Corning, NY), fetal calf serum (FCS)from Cambrex (Viers, Belgium), penicillin and streptomy-cin sulfate from Gibco Invitrogen Corp. (Grand Island,NY), and prostacyclin (PGI
2
) from Cayman ChemicalCompany (Ann Arbor, MI).
N
-(
R
)-[2-(Hydroxyaminocar-
bonyl)methyl]-4-methylpentanoyl-
L
-alanine amine TNF-
α
protease inhibitor (TAPI) was from Calbiochem (Louis-ville, KY).
D
-Glucose was from BDH analaR (Poole, Dorset,UK). The MoAb 6D1 against AA 104-128 of the VWF-binding domain was a gift of Dr Ruggeri from the Depart-ment of Molecular and Experimental Medicine, TheScripps Research Institute (La Jolla, CA). MoAb 6B4directed against AA 201-268 partially overlapping thebinding sites for thrombin and the VWF-ristocetin com-plex and MoAb 10H9 against AA 276-282 adjacent to thesebinding sites have been described.
22,23
PLT isolation and storage
Freshly drawn venous blood from healthy volunteers(40 mL) was collected with informed consent into 1:10 (v/v) 130 mmol per L trisodium citrate. The donors claimednot to have taken any medication during 2 weeks beforeblood collection. PLT-rich plasma was prepared by centrif-ugation (200
×
g
, 15 min, 20
°
C). ACD (0.1 vol of 2.5 g triso-dium citrate, 1.5 g citric acid and 2.0 g
D
-glucose in 100 mLof distilled water) was added to decrease the pH value to6.0 and prevent PLT activation during further isolation.The suspensions were centrifuged (330
×
g
, 15 min, 22
°
C)and resuspended in Hepes-Tyrode (137 mmol/L NaCl,2.68 mmol/L KCl, 0.42 mmol/L NaH
2
PO
4
, 1.7 mmol/LMgCl
2
, and 11.9 mmol/L NaHCO
3
, pH 7.2) with 5 mmol/Lglucose to a final concentration of 2
×
10
8
PLTs per mL. PLTcount was measured on a cell counter (Model AL871,Molab, Hilden, Germany). PLTs were stored on ice (termedC0 suspension) for up to 48 hours, a condition known toinduce clustering of GPIb
α
and binding and phagocytosisby macrophages.
6,9
The suspensions were stored in closedtubes impermeable for gas exchange without agitation.Before each measurement, the C0 suspensions wererewarmed for 15 minutes at 37
°
C, as described.
1
Analysis of GPIb
a
binding to AN51-PE antibody
The expression of GPIb
α
was measured by incubating0.1 mL of PLT suspension (2
×
10
8
PLTs/mL) with PE-labeled AN51 antibody at the indicated concentrations for15 minutes at 37
°
C. Subsequently, 300
µ
l of phosphate-buffered saline (PBS) was added and 10,000 events wereanalyzed on a flow cytometer (FACSCalibur, BD Bio-sciences, San Jose, CA). The distribution of AN51-PEfluorescence on fresh PLTs was gated and defined as M1-fraction. Loss of AN51-PE binding during storage resultedin a shift to lower fluorescence, defined as M2-fraction.Quantitative measurements of changes in GPIb
α
fluores-cence were evaluated by computer software (WinMDI,The Scripps Research Institute, La Jolla, CA). Some exper-iments with stored PLTs were performed after prior incu-bation for 30 minutes at 37
°
C, with either 5 mmol per Lethylenediaminetetraacetate (EDTA), 110 nmol per L

BADLOU ET AL.
2092 TRANSFUSION
Volume 46, December 2006
TAPI, 100 mmol per L GlcNAc, 100 mmol per L glucose, or100 mmol per L mannose. To investigate which regions ofGPIb
α
contributed to affinity changes for AN51-PE bind-ing, PLTs were incubated with 10
µ
g per mL MoAb 6D1,6B4, and 10H9 for 60 minutes at 37
°
C, before incubationwith 0.5 to 2
µ
g per mL AN51-PE (depending on PLTpreparations).
Binding and phagocytosis assay
The binding and phagocytosis assay was performed asdescribed.
24
THP-1 monocytic cell lines were cultured to adensity of (1-2)
×
10
6
cells per mL in RPMI 1640 contain-ing 10 percent FCS, 2 mmol per L glutamine, penicillin(100 U/mL), and streptomycin (100
µ
g/mL) at 37
°
C. THP-1 cells were counted in a counting chamber (Burker-Turk,Marienfeld, Emergo, Landsmeer, the Netherlands), and1 mL of suspension containing 1
×
10
6
cells was added toa well of a 48-well plate (Corning Inc.). Maturation ofmonocytes to macrophages was induced by incubationwith 500 nmol per L PMA for 24 hours at 37
°
C. Before thebinding and phagocytosis experiments, 100
µ
L of PLTsuspension was labeled with 1
µ
mol per L mepacrine inHepes-Tyrode (pH 7.2, 5 min, 22
°
C). Mepacrine is a fluo-rescent polyphenolic compound that emits at 519 nm,which is in the range of the emission of FITC(530
±
15 nm). Free mepacrine was removed by a washingstep (5 min, 350
×
g
, 22
°
C, with soft mode) while prevent-ing PLT activation with 10 ng per mL PGI
2
. The pelletswere then resuspended in 25
µ
L of Hepes-Tyrode buffer(pH 7.2) and 2
×
10
6
PLTs per mL were added to 1
×
10
6
macrophages in 1 mL per well. After addition of 2 mmolper L CaCl
2
and 2 mmol per L MgCl
2
(final concentra-tions), suspensions were incubated for 30 minutes at37
°
C. Subsequently, wells were gently washed three timeswith Hanks’ buffered salt solution (HBSS) buffer(0.3 mmol/L KH
2
PO
4
, 13.7 mmol/L NaCl, 417 mmol/LNaHCO
3
, 31 mmol/L Na
2
HPO
4
, and 0.5 mmol/L KCl indistilled water), and removed PLTs were stored. Two-hun-dred microliters of HBSS was added to each well followedby addition of EDTA (5 mmol/L final concentration) andincubated for 15 minutes on ice. Stored PLTs were washedunder protection of PGI
2
to remove RPMI and added tothe main suspension. Samples (100
µ
L) were incubatedwith 2
µ
g per mL AN51-PE and 2
µ
g per mL anti-humanCD14-FITC antibody for 15 minutes at 37
°
C. Then, 300
µ
Lof PBS was added and 20,000 events were measured on aFACSCalibur. Quantitative analysis was performed withWinMDI software. Binding of PLTs to macrophages wasexpressed as the percentage of CD42b/CD14-positiveparticles of the total number of CD42b- and/or CD14-positive particles. Phagocytosis of PLTs by PMA-maturedTHP-1 cells was measured by FACS analysis ofmepacrine-positive CD14 cells that were inaccessible tothe CD42b-PE antibody and expressed as percentage of
total number of CD14-positive, CD42b-negative particles,as described.24
Statistical analysis
Data are expressed as means ± SEM with number ofobservations. Statistical analysis was based on a pairedt test or one-way analysis of variance (with post t test) forcomparison between two and more groups. Differenceswere considered significant at a p value of less than 0.05.
RESULTS
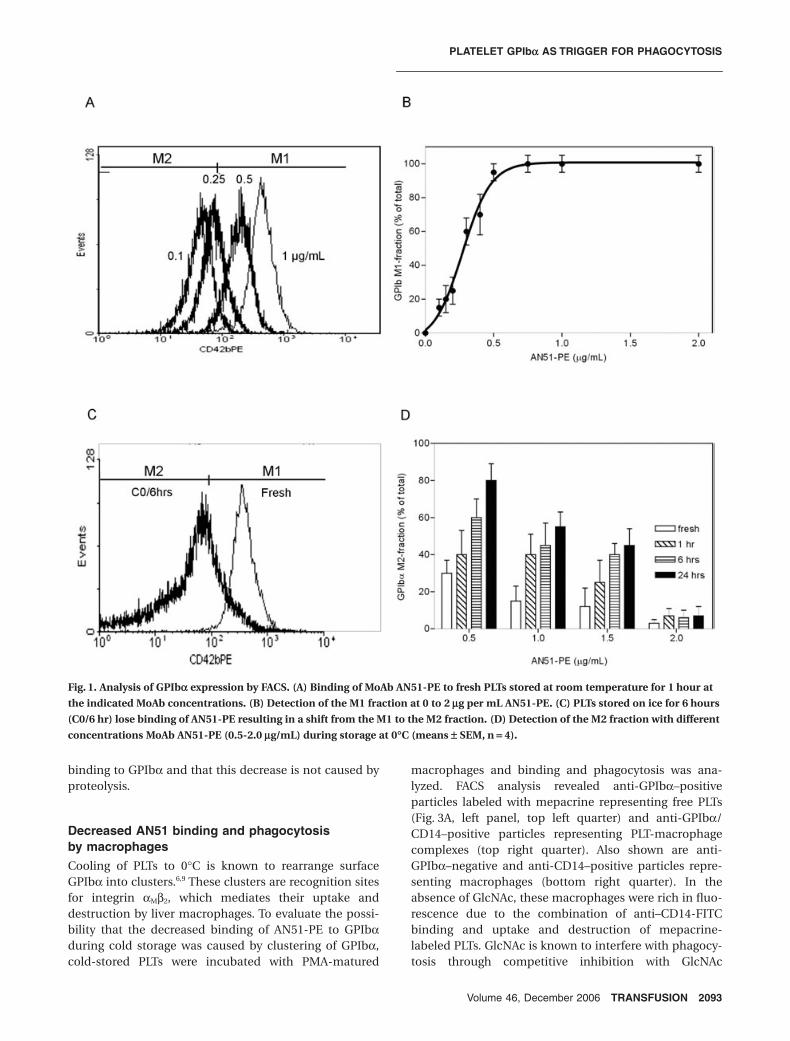
AN51-PE binding after prolonged PLT storage at 0∞CWhen freshly collected PLTs were washed and incubatedwith 1 µg per mL GPIbα antibody AN51-PE, FACS analysisshowed a single population of PLTs, which was gated anddefined as M1 fraction (Fig. 1A). When the concentrationof AN51-PE was lowered while keeping PLT count con-stant, fluorescence decreased to a range below M1, whichwas defined as M2 fraction. Titrations revealed that at2 × 108 PLTs per mL, a concentration of 0.5 to 1.5 µg permL AN51-PE was just sufficient to saturate GPIbα recep-tors (Fig. 1B). When PLTs were cooled and thereafterstored at 0°C for 6 hours, subsequent incubation with 1 µgper mL AN51-PE showed a shift from M1 to M2 fraction,reflecting changes in GPIbα that decreased the binding ofAN51-PE (Fig. 1C). Indeed, within a critical range of anti-body concentrations (0.5-1.5 µg/mL, depending on PLTpreparations), AN51-PE binding was a sensitive means todetect changes in GPIbα induced by storage at 0°C(Fig. 1D).
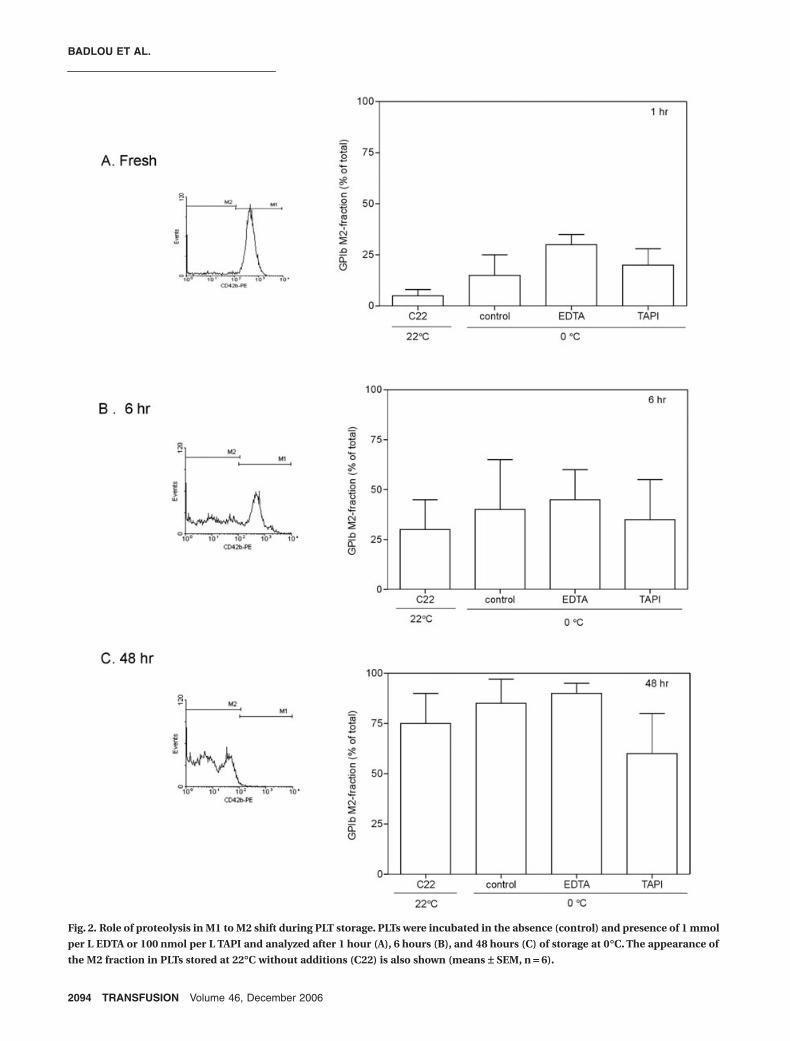
To investigate whether the decrease in antibodybinding observed during cold storage was the resultof proteolytic damage of GPIbα, incubations wererepeated in the presence of EDTA, which inhibitsmetalloproteinases18,21 and fibrinolytic proteinases,25 andTAPI,19 an inhibitor of ADAM-17. A concurrently run con-trol suspension without additions was stored at roomtemperature, defined as C22 (Fig. 2). A first analysis after1 hour of incubation at 0°C already showed 15 percent ofthe PLTs in the low-affinity M2 fraction. Neither EDTA norTAPI reduced this expression, suggesting that the decreasein AN51-PE binding was not caused by proteolysis. Thefresh control suspension kept at room temperatureshowed little GPIbα affinity loss, indicating that the cool-ing step was the cause of the decrease in GPIbα affinityfor AN51-PE. Subsequent storage at 0°C led to a furtherincrease in low-affinity GPIbα. The increase in M2 fractionwas also seen in the presence of proteolytic inhibitors,indicating again that it was not caused by proteolysis. AlsoPLTs stored at room temperature without additionsshowed a decrease in MoAb binding, but here the causewas damage of GPIbα, as reported earlier.16 These datashow that cold storage leads to a decrease in AN51-PE
PLATELET GPIba AS TRIGGER FOR PHAGOCYTOSIS
Volume 46, December 2006 TRANSFUSION 2093
binding to GPIbα and that this decrease is not caused byproteolysis.
Decreased AN51 binding and phagocytosis by macrophages
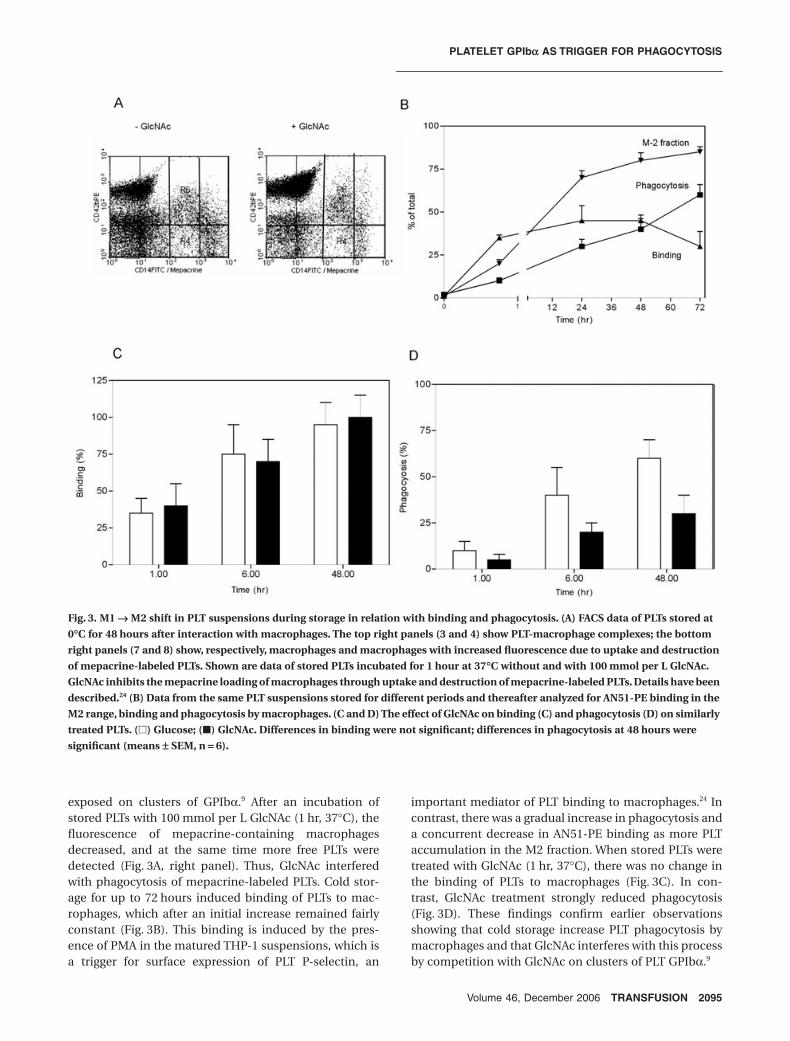
Cooling of PLTs to 0°C is known to rearrange surfaceGPIbα into clusters.6,9 These clusters are recognition sitesfor integrin αMβ2, which mediates their uptake anddestruction by liver macrophages. To evaluate the possi-bility that the decreased binding of AN51-PE to GPIbαduring cold storage was caused by clustering of GPIbα,cold-stored PLTs were incubated with PMA-matured
macrophages and binding and phagocytosis was ana-lyzed. FACS analysis revealed anti-GPIbα–positiveparticles labeled with mepacrine representing free PLTs(Fig. 3A, left panel, top left quarter) and anti-GPIbα/CD14–positive particles representing PLT-macrophagecomplexes (top right quarter). Also shown are anti-GPIbα–negative and anti-CD14–positive particles repre-senting macrophages (bottom right quarter). In theabsence of GlcNAc, these macrophages were rich in fluo-rescence due to the combination of anti–CD14-FITCbinding and uptake and destruction of mepacrine-labeled PLTs. GlcNAc is known to interfere with phagocy-tosis through competitive inhibition with GlcNAc
Fig. 1. Analysis of GPIba expression by FACS. (A) Binding of MoAb AN51-PE to fresh PLTs stored at room temperature for 1 hour at
the indicated MoAb concentrations. (B) Detection of the M1 fraction at 0 to 2 mg per mL AN51-PE. (C) PLTs stored on ice for 6 hours
(C0/6 hr) lose binding of AN51-PE resulting in a shift from the M1 to the M2 fraction. (D) Detection of the M2 fraction with different
concentrations MoAb AN51-PE (0.5-2.0 mg/mL) during storage at 0∞C (means ± SEM, n = 4).
BADLOU ET AL.
2094 TRANSFUSION Volume 46, December 2006
Fig. 2. Role of proteolysis in M1 to M2 shift during PLT storage. PLTs were incubated in the absence (control) and presence of 1 mmol
per L EDTA or 100 nmol per L TAPI and analyzed after 1 hour (A), 6 hours (B), and 48 hours (C) of storage at 0∞C. The appearance of
the M2 fraction in PLTs stored at 22∞C without additions (C22) is also shown (means ± SEM, n = 6).
PLATELET GPIba AS TRIGGER FOR PHAGOCYTOSIS
Volume 46, December 2006 TRANSFUSION 2095
exposed on clusters of GPIbα.9 After an incubation ofstored PLTs with 100 mmol per L GlcNAc (1 hr, 37°C), thefluorescence of mepacrine-containing macrophagesdecreased, and at the same time more free PLTs weredetected (Fig. 3A, right panel). Thus, GlcNAc interferedwith phagocytosis of mepacrine-labeled PLTs. Cold stor-age for up to 72 hours induced binding of PLTs to mac-rophages, which after an initial increase remained fairlyconstant (Fig. 3B). This binding is induced by the pres-ence of PMA in the matured THP-1 suspensions, which isa trigger for surface expression of PLT P-selectin, an
important mediator of PLT binding to macrophages.24 Incontrast, there was a gradual increase in phagocytosis anda concurrent decrease in AN51-PE binding as more PLTaccumulation in the M2 fraction. When stored PLTs weretreated with GlcNAc (1 hr, 37°C), there was no change inthe binding of PLTs to macrophages (Fig. 3C). In con-trast, GlcNAc treatment strongly reduced phagocytosis(Fig. 3D). These findings confirm earlier observationsshowing that cold storage increase PLT phagocytosis bymacrophages and that GlcNAc interferes with this processby competition with GlcNAc on clusters of PLT GPIbα.9
Fig. 3. M1 Æ M2 shift in PLT suspensions during storage in relation with binding and phagocytosis. (A) FACS data of PLTs stored at
0∞C for 48 hours after interaction with macrophages. The top right panels (3 and 4) show PLT-macrophage complexes; the bottom
right panels (7 and 8) show, respectively, macrophages and macrophages with increased fluorescence due to uptake and destruction
of mepacrine-labeled PLTs. Shown are data of stored PLTs incubated for 1 hour at 37∞C without and with 100 mmol per L GlcNAc.
GlcNAc inhibits the mepacrine loading of macrophages through uptake and destruction of mepacrine-labeled PLTs. Details have been
described.24 (B) Data from the same PLT suspensions stored for different periods and thereafter analyzed for AN51-PE binding in the
M2 range, binding and phagocytosis by macrophages. (C and D) The effect of GlcNAc on binding (C) and phagocytosis (D) on similarly
treated PLTs. (�) Glucose; (�) GlcNAc. Differences in binding were not significant; differences in phagocytosis at 48 hours were
significant (means ± SEM, n = 6).
BADLOU ET AL.
2096 TRANSFUSION Volume 46, December 2006
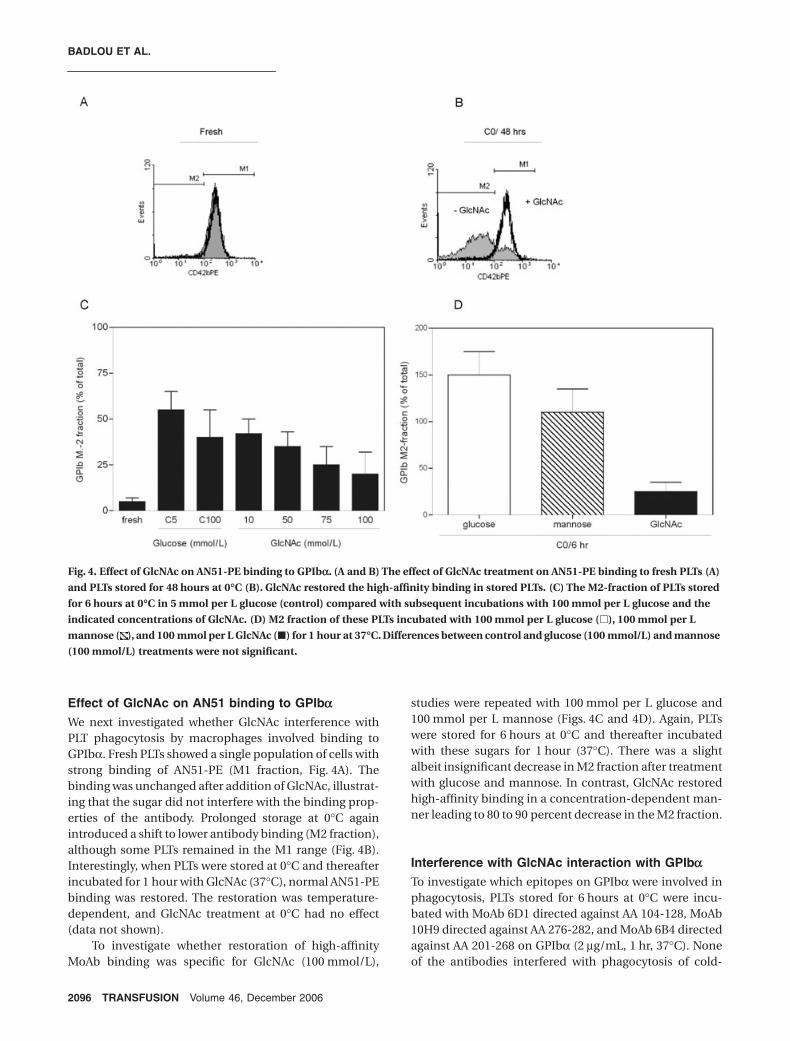
Effect of GlcNAc on AN51 binding to GPIbaWe next investigated whether GlcNAc interference withPLT phagocytosis by macrophages involved binding toGPIbα. Fresh PLTs showed a single population of cells withstrong binding of AN51-PE (M1 fraction, Fig. 4A). Thebinding was unchanged after addition of GlcNAc, illustrat-ing that the sugar did not interfere with the binding prop-erties of the antibody. Prolonged storage at 0°C againintroduced a shift to lower antibody binding (M2 fraction),although some PLTs remained in the M1 range (Fig. 4B).Interestingly, when PLTs were stored at 0°C and thereafterincubated for 1 hour with GlcNAc (37°C), normal AN51-PEbinding was restored. The restoration was temperature-dependent, and GlcNAc treatment at 0°C had no effect(data not shown).
To investigate whether restoration of high-affinityMoAb binding was specific for GlcNAc (100 mmol/L),
studies were repeated with 100 mmol per L glucose and100 mmol per L mannose (Figs. 4C and 4D). Again, PLTswere stored for 6 hours at 0°C and thereafter incubatedwith these sugars for 1 hour (37°C). There was a slightalbeit insignificant decrease in M2 fraction after treatmentwith glucose and mannose. In contrast, GlcNAc restoredhigh-affinity binding in a concentration-dependent man-ner leading to 80 to 90 percent decrease in the M2 fraction.
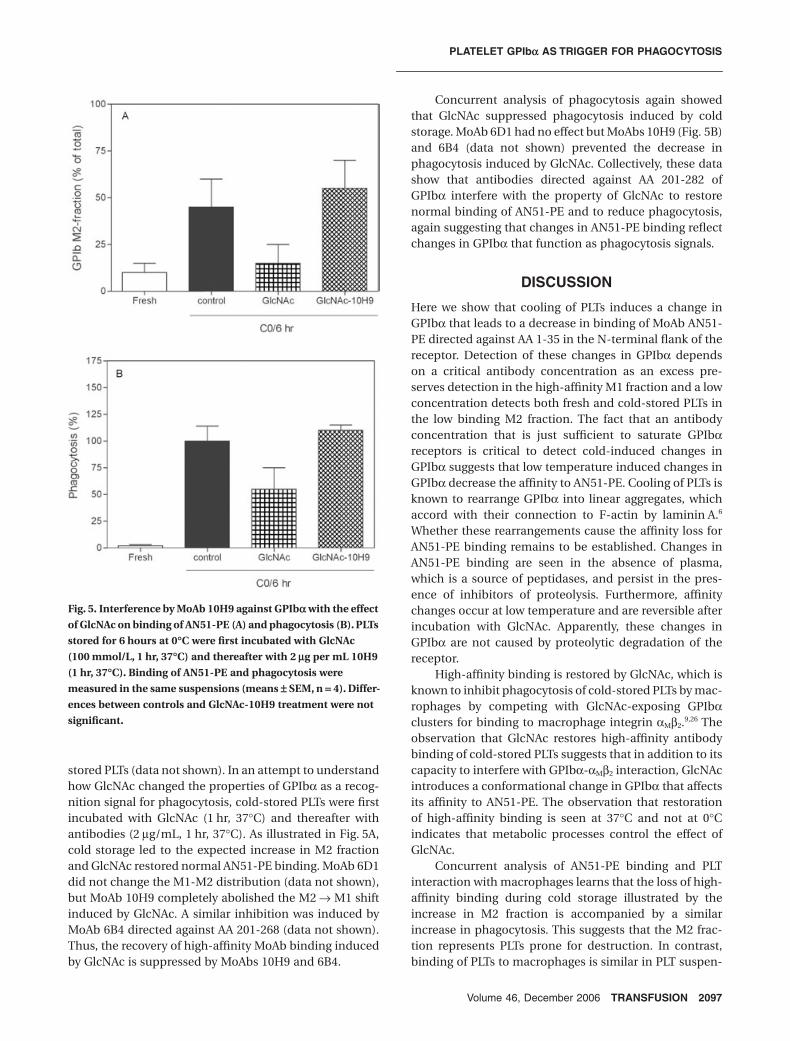
Interference with GlcNAc interaction with GPIbaTo investigate which epitopes on GPIbα were involved inphagocytosis, PLTs stored for 6 hours at 0°C were incu-bated with MoAb 6D1 directed against AA 104-128, MoAb10H9 directed against AA 276-282, and MoAb 6B4 directedagainst AA 201-268 on GPIbα (2 µg/mL, 1 hr, 37°C). Noneof the antibodies interfered with phagocytosis of cold-
Fig. 4. Effect of GlcNAc on AN51-PE binding to GPIba. (A and B) The effect of GlcNAc treatment on AN51-PE binding to fresh PLTs (A)
and PLTs stored for 48 hours at 0∞C (B). GlcNAc restored the high-affinity binding in stored PLTs. (C) The M2-fraction of PLTs stored
for 6 hours at 0∞C in 5 mmol per L glucose (control) compared with subsequent incubations with 100 mmol per L glucose and the
indicated concentrations of GlcNAc. (D) M2 fraction of these PLTs incubated with 100 mmol per L glucose (�), 100 mmol per L
mannose ( ), and 100 mmol per L GlcNAc (�) for 1 hour at 37∞C. Differences between control and glucose (100 mmol/L) and mannose
(100 mmol/L) treatments were not significant.
PLATELET GPIba AS TRIGGER FOR PHAGOCYTOSIS
Volume 46, December 2006 TRANSFUSION 2097
stored PLTs (data not shown). In an attempt to understandhow GlcNAc changed the properties of GPIbα as a recog-nition signal for phagocytosis, cold-stored PLTs were firstincubated with GlcNAc (1 hr, 37°C) and thereafter withantibodies (2 µg/mL, 1 hr, 37°C). As illustrated in Fig. 5A,cold storage led to the expected increase in M2 fractionand GlcNAc restored normal AN51-PE binding. MoAb 6D1did not change the M1-M2 distribution (data not shown),but MoAb 10H9 completely abolished the M2 → M1 shiftinduced by GlcNAc. A similar inhibition was induced byMoAb 6B4 directed against AA 201-268 (data not shown).Thus, the recovery of high-affinity MoAb binding inducedby GlcNAc is suppressed by MoAbs 10H9 and 6B4.
Concurrent analysis of phagocytosis again showedthat GlcNAc suppressed phagocytosis induced by coldstorage. MoAb 6D1 had no effect but MoAbs 10H9 (Fig. 5B)and 6B4 (data not shown) prevented the decrease inphagocytosis induced by GlcNAc. Collectively, these datashow that antibodies directed against AA 201-282 ofGPIbα interfere with the property of GlcNAc to restorenormal binding of AN51-PE and to reduce phagocytosis,again suggesting that changes in AN51-PE binding reflectchanges in GPIbα that function as phagocytosis signals.
DISCUSSION
Here we show that cooling of PLTs induces a change inGPIbα that leads to a decrease in binding of MoAb AN51-PE directed against AA 1-35 in the N-terminal flank of thereceptor. Detection of these changes in GPIbα dependson a critical antibody concentration as an excess pre-serves detection in the high-affinity M1 fraction and a lowconcentration detects both fresh and cold-stored PLTs inthe low binding M2 fraction. The fact that an antibodyconcentration that is just sufficient to saturate GPIbαreceptors is critical to detect cold-induced changes inGPIbα suggests that low temperature induced changes inGPIbα decrease the affinity to AN51-PE. Cooling of PLTs isknown to rearrange GPIbα into linear aggregates, whichaccord with their connection to F-actin by laminin A.6
Whether these rearrangements cause the affinity loss forAN51-PE binding remains to be established. Changes inAN51-PE binding are seen in the absence of plasma,which is a source of peptidases, and persist in the pres-ence of inhibitors of proteolysis. Furthermore, affinitychanges occur at low temperature and are reversible afterincubation with GlcNAc. Apparently, these changes inGPIbα are not caused by proteolytic degradation of thereceptor.
High-affinity binding is restored by GlcNAc, which isknown to inhibit phagocytosis of cold-stored PLTs by mac-rophages by competing with GlcNAc-exposing GPIbαclusters for binding to macrophage integrin αMβ2.9,26 Theobservation that GlcNAc restores high-affinity antibodybinding of cold-stored PLTs suggests that in addition to itscapacity to interfere with GPIbα-αMβ2 interaction, GlcNAcintroduces a conformational change in GPIbα that affectsits affinity to AN51-PE. The observation that restorationof high-affinity binding is seen at 37°C and not at 0°Cindicates that metabolic processes control the effect ofGlcNAc.
Concurrent analysis of AN51-PE binding and PLTinteraction with macrophages learns that the loss of high-affinity binding during cold storage illustrated by theincrease in M2 fraction is accompanied by a similarincrease in phagocytosis. This suggests that the M2 frac-tion represents PLTs prone for destruction. In contrast,binding of PLTs to macrophages is similar in PLT suspen-
Fig. 5. Interference by MoAb 10H9 against GPIba with the effect
of GlcNAc on binding of AN51-PE (A) and phagocytosis (B). PLTs
stored for 6 hours at 0∞C were first incubated with GlcNAc
(100 mmol/L, 1 hr, 37∞C) and thereafter with 2 mg per mL 10H9
(1 hr, 37∞C). Binding of AN51-PE and phagocytosis were
measured in the same suspensions (means ± SEM, n = 4). Differ-
ences between controls and GlcNAc-10H9 treatment were not
significant.

BADLOU ET AL.
2098 TRANSFUSION Volume 46, December 2006
sions with approximately 20 percent M2 PLTs and approx-imately 80 percent M2 PLTs suggesting a different controlmechanism that is independent of changes in GPIbα. Thisaccords with previous findings showing that binding ofPLTs to macrophage is mediated through surface expres-sion of P-selectin and independent of exposed phosphati-dylinositol and changes in GPIbα, which serve exclusivelyin phagocytosis.24
Both the increase in M2 fraction and the increase inphagocytosis during cold storage are reversed by GlcNAc.This is another illustration of the tight relation betweenM1-M2 shifts and control of phagocytosis signals onGPIbα and indicates that the changes inflicted duringcooling and storage at low temperature are reversible innature. How GlcNAc restores the normal GPIbα confor-mation awaits further studies. MoAb 6B4 directed againstAA 201-268 partially overlapping the binding sites forthrombin and the VWF-ristocetin complex and 10H9against AA 276-282 in the anionic-sulfated sequence adja-cent to these binding sites interfered with restoration ofhigh-affinity binding by GlcNAc, suggesting that theregion between AA 201 and AA 282 is sensitive to GlcNAcaddition. The MoAb 6D1 against AA 104-128 had no effect.This suggests that GlcNAc binds to a specific part of thereceptor that controls the generation of phagocytosissignals.
How GlcNAc controls GPIbα affinity for AN51-PE andthe recognition by macrophages is at present unknown.The observation that exposed GlcNAc residues on GPIbαcan be masked by galactosylation in the absence of addedenzymes9 suggests that the PLT surface contains a galac-tosyltransferase that couples galactose to GPIbα. Thepresent results suggest that similar transferases mightcontrol the coupling between GlcNAc and GPIbα but theprecise characterization awaits further investigations.Although these interactions greatly affect PLT destructionby matured THP-1 cells, their importance for the in vivodestruction of PLTs by macrophages remains to be estab-lished. If the same relation between PLT binding of AN51-PE and phagocytosis remains valid, a simple FACS analysismight predict the fate of transfused PLTs in patients witha shortage in PLT number and/or function.
REFERENCES
1. Badlou BA, Ijseldijk MJ, Smid WM, Akkerman JW. Prolonged
platelet preservation by transient metabolic suppression.
Transfusion 2005;45:214-22.
2. Perrotta PL, Perrotta CL, Snyder EL. Apoptotic activity in
stored human platelets. Transfusion 2003;43:526-35.
3. Wadhawan V, Karim ZA, Mukhopadhyay S, et al. Platelet
storage under in vitro condition is associated with calcium-
dependent apoptosis-like lesions and novel reorganization
in platelet cytoskeleton. Arch Biochem Biophys 2004;422:
183-90.
4. Xiao HY, Matsubayashi H, Bonderman DP, et al. Generation
of annexin V-positive platelets and shedding of
microparticles with stimulus-dependent procoagulant
activity during storage of platelets at 4 degrees C.
Transfusion 2000;40:420-7.
5. Snyder EL, Rinder HM. Platelet storage—time to come in
from the cold? N Engl J Med 2003;348:2032-3.
6. Hoffmeister KM, Felbinger TW, Falet H, et al. The clearance
mechanism of chilled blood platelets. Cell 2003;112:87-97.
7. Hartwig JH. Mechanisms of actin rearrangements mediating
platelet activation. J Cell Biol 1992;118:1421-42.
8. Kawakami H, Higashihara M, Ohsaka M, et al. Myosin light
chain phosphorylation is correlated with cold-induced
changes in platelet shape. J Smooth Muscle Res 2001;37:113-
22.
9. Hoffmeister KM, Josefsson EC, Isaac NA, et al. Glycosylation
restores survival of chilled blood platelets. Science 2003;301:
1531-4.
10. Andrews RK, Gardiner EE, Shen Y, Whisstock JC, Berndt MC.
Glycoprotein Ib-IX-V. Int J Biochem Cell Biol 2003;35:
1170-4.
11. Canobbio I, Balduini C, Torti M. Signalling through the
platelet glycoprotein Ib-V-IX complex. Cell Signal 2004;16:
1329-44.
12. Huizinga EG, Tsuji S, Romijn RA, et al. Structures of
glycoprotein Ibalpha and its complex with von Willebrand
factor A1 domain. Science 2002;297:1176-9.
13. Chou CF, Omary MB. Mitotic arrest with anti-microtubule
agents or okadaic acid is associated with increased
glycoprotein terminal GlcNAc’s. J Cell Sci 1994;107:1833-43.
14. Wu YP, Vink T, Schiphorst M, et al. Platelet thrombus
formation on collagen at high shear rates is mediated by von
Willebrand factor-glycoprotein Ib interaction and inhibited
by von Willebrand factor-glycoprotein IIb/IIIa interaction.
Arterioscler Thromb Vasc Biol 2000;20:1661-7.
15. Kawasaki T, Fujimura Y, Usami Y, et al. Complete amino acid
sequence and identification of the platelet glycoprotein Ib-
binding site of jararaca GPIb-BP, a snake venom protein
isolated from Bothrops jararaca. J Biol Chem 1996;271:
10635-9.
16. Bessos H, Murphy WG, Seghatchian MJ. Monitoring the
release of glycocalicin in platelet concentrates by ELISA.
Blood Coagul Fibrinolysis 1991;2:373-6.
17. Adelman B, Michelson AD, Greenberg J, Handin RI.
Proteolysis of platelet glycoprotein Ib by plasmin is
facilitated by plasmin lysine-binding regions. Blood
1986;68:1280-4.
18. Bergmeier W, Burger PC, Piffath CL, et al. Metalloproteinase
inhibitors improve the recovery and hemostatic function of
in vitro-aged or -injured mouse platelets. Blood 2003;102:
4229-35.
19. Bergmeier W, Piffath CL, Cheng G, et al. Tumor necrosis
factor-alpha-converting enzyme (ADAM17) mediates
GPIbalpha shedding from platelets in vitro and in vivo. Circ
Res 2004;95:677-83.

PLATELET GPIba AS TRIGGER FOR PHAGOCYTOSIS
Volume 46, December 2006 TRANSFUSION 2099
20. Rabie T, Strehl A, Ludwig A, et al. Evidence for a role of
ADAM17 (TACE) in the regulation of platelet glycoprotein V.
J Biol Chem 2005;280:14462-8.
21. Bergmeier W, Rabie T, Strehl A, et al. GPVI down-regulation
in murine platelets through metalloproteinase-dependent
shedding. Thromb Haemost 2004;91:951-8.
22. Cauwenberghs N, Vanhoorelbeke K, Vauterin S, et al.
Epitope mapping of inhibitory antibodies against platelet
glycoprotein Ibalpha reveals interaction between the
leucine-rich repeat N-terminal and C-terminal flanking
domains of glycoprotein Ibalpha. Blood 2001;98:652-60.
23. Deckmyn H, Cauwenberghs N, Wu D, Depraetere H,
Vanhoorelbeke K. Development of antibodies that interfere
with the collagen-VWF-GPIb axis as new antithrombotics.
Verh K Acad Geneeskd Belg 2005;67:55-65.
24. Badlou BA, Wu YP, Smid WM, Akkerman JW. Regulation
of platelet phagocytosis by macrophages. Transfusion
2006;46:1432-43.
25. Bello CA, Hermogenes AL, Magalhaes A, et al. Isolation and
biochemical characterization of a fibrinolytic proteinase
from Bothrops leucurus (white-tailed jararaca) snake
venom. Biochimie 2006;88:189-200.
26. Josefsson EC, Gebhard HH, Stossel TP, Hartwig JH,
Hoffmeister KM. The macrophage alphaMbeta2 integrin
alphaM lectin domain mediates the phagocytosis of chilled
platelets. J Biol Chem 2005;280:18025-32.