rice ascorbate peroxidase gene family encodes functionally diverse isoforms localized in different...
TRANSCRIPT

ORIGINAL ARTICLE
Felipe Karam Teixeira Æ Larissa Menezes-Benavente
Vinıcius Costa Galvao Æ Rogerio Margis
Marcia Margis-Pinheiro
Rice ascorbate peroxidase gene family encodes functionally diverseisoforms localized in different subcellular compartments
Received: 11 October 2005 / Accepted: 11 December 2005 / Published online: 6 January 2006� Springer-Verlag 2006
Abstract Aerobic organisms evolved a complex anti-oxidant system, which protect the cells against oxida-tive damage caused by partially reduced oxygenintermediates, also known as reactive oxygen species.In plants, ascorbate peroxidases (EC, 1.11.1.11) cata-lyze the conversion of H2O2 to H2O, using ascorbateas the specific electron donor in this enzymatic reac-tion. Previously, eight APx genes were identified in therice (Oryza sativa L.) genome through in silico anal-ysis: two cytosolic isoforms, two putative peroxisomalisoforms, and four putative chloroplastic ones. Usinggene-specific probes, we confirmed the presence of theeight APx genes in the rice genome by Southern blothybridization. Transcript accumulation analysisshowed specific expression patterns for each memberof the APx family according to developmental stageand in response to salt stress, revealing the complexityof the antioxidant system in plants. Finally, the sub-cellular localization of rice APx isoforms was deter-mined using GFP-fusion proteins in BY-2 tobaccocells. In agreement with the initial prediction,OSAPX3 was localized in the peroxisomes. On the
other hand, the OSAPX6-GFP fusion protein wasfound in mitochondria of the BY-2 cells, in contrastto the chloroplastic location predicted by sequenceanalysis. Our findings reveal the functional diversity ofthe rice APx genes and suggest complementation andcoordination of the antioxidant defenses in differentcellular compartments during development and abioticstress.
Keywords Antioxidant metabolism Æ Ascorbateperoxidase Æ BY-2 tobacco cells Æ Green fluorescentprotein Æ Oryza
Abbreviations APx: Ascorbate peroxidase Æ CaMV:Cauliflower mosaic virus Æ CAT: Catalase Æ DHAR:Dehydroascorbate reductase Æ DIC: Differentialinterference contrast Æ EST: Expressed sequencetag Æ GFP: Green fluorescent protein Æ GPx:Glutathione peroxidase Æ GR: Glutathionereductase Æ KO: Knock-out mutants Æ MDHAR:Monodehydroascorbate reductase Æ PCD: Programmedcell death Æ pER: Peroxisomal endoplasmaticreticulum Æ Prx: Peroxidase Æ ROS: Reactive oxygenspecies Æ SOD: Superoxide dismutase Æ SRE: Splicingregulatory cis-element Æ TMRE: Tetramethylrhodamineethyl ester Æ 3-UTR: 3untranslated region
Introduction
Reactive oxygen species (ROS) such as the superoxideradical (O2
�), H2O2, and hydroxyl radical are producedas a consequence of the univalent reduction of molecularoxygen in metabolic reactions, thus, being intrinsic tothe aerobic life style. However, when present in highconcentration in the cells, ROS can diffuse throughmembranes and react with biological compounds suchas proteins, nucleic acids, and lipids, producing extensivedamage and, in severe cases, cellular death (Fridovich1998; Scandalios 2002). In addition, the production and
F. K. TeixeiraL. Menezes-Benavente Æ V. C. GalvaoM. Margis-PinheiroLaboratorio de Genetica Molecular Vegetal,Departamento de Genetica, Universidade Federaldo Rio de Janeiro, 21944-970, Rio de Janeiro, Brasil
R. MargisDepartamento de Bioquımica, Universidade Federal do RioGrande do Sul, 90035-003, Rio Grande do Sul, Brasil
M. Margis-Pinheiro (&)Laboratorio de Genetica Vegetal,Departamento de Genetica, Universidade Federaldo Rio Grande do Sul. Av. Bento Goncalves, 9500,Predio 43323, Porto Alegre, RS, BrasilE-mail: [email protected]: +55-21-25626380
Present address:L. Menezes-BenaventeDepartment of Genetics, North Carolina State University,Raleigh, NC, 27695, USA
Planta (2006) 224: 300–314DOI 10.1007/s00425-005-0214-8

accumulation of ROS increase in response to stressfulenvironmental conditions such as drought, high light,wounding, salinity, or pathogen infection (Karpinskiet al. 1997; Noctor and Foyer 1998; Mittler et al. 2004).Besides that, in plant cells, the intracellular level of ROSmust be tightly regulated since ROS are also signalingmolecules involved in several physiological processessuch as cell cycle, stress perception, gene regulation,programmed cell death (PCD), hypersensitive response,and senescence (Dat et al. 2000; Orozco-Cardenas et al.2001; Mullineaux and Karpinski 2002; Vandenabeeleet al. 2003). Control of intracellular ROS levels is a verycomplex process and involves a large network of genes,whose principal function is to avoid cellular damage thatcould result in oxidative stress and disturbances in cel-lular redox homeostasis (Mittler et al. 2004). The ROS-scavenging system is responsible for removing free rad-icals and keeping the cellular steady-state level of ROSunder tight control. This system consists of low-molec-ular weight compounds including ascorbic acid, gluta-thione, tocoferol, and carotenoids, and several enzymessuch as superoxide dismutases (SOD), ascorbate perox-idases (APx), catalases (CAT), glutathione peroxidases(GPx), and peroxidases (Prx) (Halliwell and Gutteridge1999; Scandalios, 2002; Mittler et al. 2004).
APx (EC, 1.11.1.11) has a key role in scavengingreactive oxygen species and in protecting cells againstthe toxic effects of ROS in higher plants, algae, andEuglena (Shigeoka et al. 1980, 2002)). This class I per-oxidase catalyzes the conversion of H2O2 to H2O and O2
using ascorbate as specific electron donor (Asada 1999).The detoxification of H2O2 by APx is followed by a setof reactions catalyzed by monodehydroascorbatereductase (MDHAR), dehydroascorbate reductase(DHAR), and glutathione reductase (GR). Together,these reactions comprise one of the most importantantioxidant systems in plants—the ascorbate-glutathi-one or Halliwell-Asada cycle. In this cycle, ascorbateand glutathione are used as reducing substrates for thescavenging of H2O2 and are ultimately recycled at theexpense of ATP and NAD(P)H (Alscher et al. 1997;Noctor and Foyer 1998).
The ascorbate-glutathione cycle acts in different cel-lular compartments including chloroplasts, peroxisomes,and cytosol, protecting them against oxidative damage.In particular, the ascorbate-glutathione cycle exerts acritical role in chloroplasts redox protection. Theseorganelles are devoid of catalases and are a major sourceof superoxide and H2O2 as a consequence of the highlyenergetic reactions that take place during photosyntheticactivity. Consequently, ascorbate and glutathione arefound at very high concentrations in these organelles(Alscher et al. 1997; Noctor and Foyer 1998). Recently,a series of reports have convincingly demonstrated theactivity of the ascorbate-glutathione cycle enzymes inthe mitochondria of plant cells, proving that this cyclealso plays an important role in the protection of thisorganelle against the harmful effects of ROS that areregularly produced in the respiratory chain reactions
(Jimenez et al. 1997; Jimenez et al. 1998; De Leonardiset al. 2000; Mittova et al. 2004a).
The distinct APx isoforms are classified according totheir subcellular location: soluble isoforms are found inthe cytosol (Shigeoka et al. 1980; Ishikawa et al. 1995)and in the stroma of plastids (Ishikawa et al. 1996;Madhusudhan et al. 2003); and membrane-bound iso-forms are present in the peroxisomes (Ishikawa et al.1998) and associated with the thylakoid membrane ofchloroplasts (Miyake et al. 1993; Ishikawa et al. 1996).The distinct subcellular location of APx resulted in dif-ferent biochemical properties of the isoforms such asmolecular mass, substrate specificity, ideal pH, andstability in the absence of ascorbate (Ishikawa et al.1998). When compared to the cytosolic and peroxisomalisoforms, the chloroplastic enzymes are more labile andare rapidly inactivated in the absence of ascorbate(Miyake et al. 1993; Ishikawa et al. 1998; Shigeoka et al.2002). Furthermore, the analysis of the phylogeneticsand structural organization of APx genes indicate thatthe different APx isoforms arose by a complex evolu-tionary process involving several gene duplications andsuggest a close relationship among proteins located inthe same subcellular compartment (Teixeira et al. 2004).
APx subcellular location is determined by the pres-ence of targeting peptides and transmembrane domainsin the N- and COOH-terminal regions of the proteins(Shigeoka et al. 2002; Teixeira et al. 2004). The peroxi-somal targeting signal comprises a COOH-terminaltransmembrane domain rich in valine and alanine, fol-lowed by a positively charged domain containing fiveamino acid residues (Mullen and Trelease 2000). Theseisoforms face the cytosol and are indirectly sorted to theperoxisomes via a subdomain of the rough endoplas-matic reticulum, known as pER (Mullen et al. 1999;Lisenbee et al. 2003a). In contrast, all chloroplasticisoforms possess a hydroxylated peptide at the N-ter-minus that is processed in the mature proteins (Mad-husudhan et al. 2003). In addition, thylakoid-boundisoforms present a transmembrane hydrophobic domainat their COOH-terminus (Miyake et al. 1993; Ishikawaet al. 1996). Recently, Chew et al. (2003b) showed thatthe Arabidopsis stromal APx is dual-targeted to plastidsand mitochondria due to the ambiguity of the targetingpeptide at N-terminal of the proteins. Conversely,Mittova et al. (2004b) suggested that distinct genesgenerate the mitochondrial and stromal isoform in to-mato plants, despite the fact that no corresponding gene,cDNA, or protein sequences for the specific mitochon-drial isoform have been described so far.
In Arabidopsis, there are at least six genes encodingAPx isoforms: two cytosolic isoforms (At1g07890 andAt3g09640); two peroxisomal isoforms (At4g35000 andAt4g35970); a thylakoid-bound isoform (At1g77490);and a stromal/mitochondrial isoform (At4g08390)(Chew et al. 2003b; Teixeira et al. 2004; Davletova et al.2005). In contrast to Arabidopsis, stromal and thylakoid-bound isoforms are generated by alternative splicing of asingle gene in spinach, tobacco, pumpkin, and iceplant
301

(Ishikawa et al. 1996; Mano et al. 1997; Yoshimura et al.2002).
The expression of APx encoding genes is modulatedby diverse environmental stimuli such as water and saltstresses, high temperatures, freezing, pathogen attack,H2O2 treatment, and abscisic acid (Zhang et al. 1997;Mittler et al. 1998; Yoshimura et al. 2000; Sato et al.2001; Shigeoka et al. 2002; Agrawal et al. 2003; Mene-zes-Benavente et al. 2004a, b). In Arabidopsis, the twogenes encoding cytosolic isoforms were largely studied:AtAPx1 (At1g07890) is expressed in different tissues andin response to diverse stresses (Pnueli et al. 2003;Fourcroy et al. 2004; Rizhsky et al. 2004); and AtAPx2(At3g09640) is expressed in the photosynthetic bundlesheath cell layer in response to light and wound (Fryeret al. 2003; Chang et al. 2004). Recently, analysis ofArabidopsis knock-out mutants (KO) have pointed toAtAPx1, being a central component of the reactiveoxygen gene network, demonstrating that cytosolic APxisoforms play a key role in chloroplast protection duringlight stress (Pnueli et al. 2003; Davletova et al. 2005).
Despite its obvious importance as a model cerealplant, little is know about the rice APx gene family. In aprevious study, we reported the identification of a mul-tigene family encoding eight ascorbate peroxidases inrice: two cytosolic (OsAPx1 and OsAPx2), two putativeperoxisomal (OsAPx3 and OsAPx4), and four putativechloroplastic isoforms (OsAPx5 to OsAPx8), whereOSAPX8 is a putative thylakoid-bound isoform (Te-ixeira et al. 2004). In the present work, we have ex-panded our analysis toward a comprehensive study ofthe expression profile of each of these genes. Transcriptaccumulation analysis showed a specific expressionpattern for each member of the rice APx family in dif-ferent plant tissues and in response to salt stress, sug-gesting the possible involvement of these genes in ricestress defense and in growth/development. Additionally,we have confirmed the subcellular localization of theperoxisomal isoform OSAPX3. Conversely, in contrastto the initial in silico prediction, we show that OSAPX6was located in the mitochondria of tobacco BY-2 cells.
Materials and methods
Plant material
Rice seeds (O. sativa L. var. indica cv. Taim 7) used inthis study were kindly provided by Centro de PesquisaAgropecuaria de Clima Temperado–CPCAT/EMBRA-PA, Rio Grande do Sul, Brazil. Seeds were germinatedin water at 28�C under dark conditions. Two-day-oldseedlings were transferred to plastic pots containinghalf-strength Furlanis solution (Furlani and Furlani1998) and grown in a greenhouse, at 28+2�C, undernatural sunlight. Nutrients composition of Furlani’ssolution is as follow: 48 mg/l N–NO3
�; 12 mg/l N–NH4�;
200 mg/l Ca; 200 mg/l K; 40.6 mg/l Mg; 8.0 mg/l P;
151 mg/l S; 234 mg/l Cl; 4.85 mg/l Fe; 0.67 mg/l Mn;0.36 mg/l B; 0.20 mg/l Zn; 0.05 mg/l Cu; 0.11 mg/l Mo.Nutrient solution volume and pH were monitoredweekly and changed every 2 weeks. Two-week-oldseedlings were used for salt treatments (250 mM NaCl)as described previously (Menezes-Benavente et al.2004b). Plant materials were sampled at the times indi-cated and immediately frozen at �80�C. For northernblot hybridization analysis, all samples were collected at12 a.m. Two independent repetitions of each experimentwere performed.
Tobacco BY-2 cells (Nicotiana tabacum var. BrightYellow 2) (Kato et al. 1972) were grown in the dark byshaking (120 rpm) in suspension at 28�C using a modi-fied Murashige and Skoog (1962) basal medium sup-plemented with 1.5 mM KH2PO4, 3 mM thiamine,0.55 mM inositol, 87 mM sucrose, and 1 lM 2,4 di-chlorophenoxy acetic acid (2,4-D). Cells were main-tained in the log phase by subculturing weekly into freshmedium at a dilution of 1:50. The tobacco BY-2 calliwere cultured on the above medium solidified with 0.7%(w/v) agar at 28�C in the dark and subcultured every3 weeks.
DNA probes and Southern blot hybridization analysis
A standard reverse transcriptase-polymerase chainreaction (RT-PCR) was used to amplify the OsAPx2, 3,and 8 cDNAs (accession nos. AB053297, AY382617,and AK070842, respectively). For this, the followingprimer pairs and annealing temperatures were used:OsAPx2: forward 5¢-CGCGCCCTCATCGCC-3¢ andreverse 5 ¢-ATCCATAGTTATCTTGACAGCA-3¢(52�C-938 bp); OsAPx3: forward 5¢-ACCTATCACCTCTCCTCGGATCG-3¢ and reverse 5¢-CAACATAGCAGACTTAGCAGCACTCAC-3¢ (56�C-1062 bp); OsAPx8: forward 5¢-TCCGTCTTGGGTGGCACGA-3¢and reverse 5¢-TGGAATCCATTCACATGCCCATC-3¢ (55�C-1246 bp). The cDNA derived from seedlingleaves of 4-week-old rice plants were used. The PCRproducts were cloned onto pGEM�-T Easy (Promega,WI, USA) and sequenced in a MegaBACE 1000(Amersham-Bioscience, Norway) (data not shown).
The probes were prepared from the ESTs (expressedsequence tags) C28227, C29046, AU030079, andAU173022 (NCBI accession number), kindly providedby Dr. Takuji Sasaki from the National Institute ofAgrobiological Resources-Rice Genome Research Pro-gram (NIAR - RGP), Japan. In addition, the EST(NCBI AI978435; Causse et al. 1994) was gently sup-plied by Dr. Susan R. McCouch, from Cornell Univer-sity, USA. The restriction sites used to obtain specificprobes of each APx gene from EST and cDNA amplifiedare described in Table 1.
Rice genomic DNA (20 lg) extracted from leaves wasdigested with HindIII, EcoRI, or XbaI, separated byelectrophoresis on a 1% (w/v) agarose gel, and blottedonto a nylon membrane (Hybond-N, Amersham Phar-
302

macia Biotech, Buckinghamshire, UK). The membraneswere cross-linked for 2 h at 80�C and hybridized with[a-32P]dCTP-labeled specific probes prepared by randompriming (Feinberg and Vogelstein 1983). Hybridizationwas carried out in 0.5 M sodium phosphate (pH 7.2),7% SDS, and 1 mM EDTA at 65�C (Church and Gil-bert 1984). The hybridized membranes were washedunder stringent condition (0.5·SSC and 0.1% (w/v) SDSat 65�C for 1 h) and exposed to X-ray film (BioMax MS-Kodak, Tokyo, Japan) using two intensifying screens at�80�C.
RNA extraction and northern blot hybridizationanalysis
Total RNA was extracted from rice seedling or matureplant leaves and roots as described (Ragueh et al. 1989).Northern blot analyses were carried out as describedpreviously (Menezes-Benavente et al. 2004b). Hybrid-ization, washing, and exposure were done as mentionedin genomic southern analysis. The relative RNA load-ings were indicated by the ethidium bromide-stainedribosomal RNA (rRNA) panels in the relevant figures.In salt stress experiments, quantification of the tran-script levels was performed by measuring the bandintensities by Scion Image program (Scion Corporation,MA, USA). The average measurement from two repli-cate experiments and the calculated error bars wereplotted using Prism 3.02 software (GraphPad SoftwareInc., CA, USA).
Quantitative reverse transcriptase PCR
Approximately one microgram of total RNA was ad-ded to each cDNA synthesis reaction using theSuperscript-II RT pre-amplification system (Invitrogen,CA, USA). Reactions were performed at 40�C for 1 husing the primer T23V (5¢ TTT TTT TTT TTT TTTTTT TTT TTV). Real-time PCR amplification wascarried out using specific primer pairs designed withOligo Calculator version 3.02 (http://www.ba-sic.nwu.edu/biotools/oligocalc.html) and synthesized byRW-Genes. Quantitative reverse transcriptase PCR
(QRT-PCR) was carried out in an Applied-Biosystem7500 real-time cycler. Reaction settings were composedof an initial denaturation step of 5 min at 94�C fol-lowed by 45 cycles of 10 s at 94�C, 15 s at 63�C, and15 s at 72�C; samples were kept for 2 min at 40�C forreannealing and then heated from 55 to 99�C with aramp of 1�C/s to acquire data to produce the dena-turing curve of the amplified products. QRT-PCRswere made in 10 ll final volume composed of 5 ll ofeach reverse transcription sample diluted 50 times in1 ll Platinum Taq 10· PCR buffer, 0.6 ll MgCl250 mM, 0.2 ll dNTPs 5 mM, 0.2 ll primer pairs10 lM, 2.45 ll H2O2, 0.5 ll SYBR (1:100,000 -Molecular Probes, OR, USA), and 0.05 ll PlatinumTaq DNA polymerase (5 U/ll) (Invitrogen, CA, USA).The melting temperatures of all primers were adjustedto 63�C. Primer pairs OsAPx1 to OsAPx8 were asfollow: OsAPx1: OsAPX1-F 5¢-CTCGAGCTACAAGGAGGCCCACCTCA-3¢ and OsAPX1-R 5¢-GGTACCTCAGCCGCATTTCATACCAACACA-3¢ (185 bp);OsAPX2: OsAPX2-F 5¢-CTCGAGCCAAGTGACAAAGCCCTCAT-3¢ and OsAPX2-R 5¢-GGTACCTCGAGAAGGCGCAAAATACAAATCG-3¢ (198 bp); OsAPX3: OsAPX3-F 5¢-GGTACCTCGAGCGAAAGATCAGGATTTGATGGT-3¢ and OsAPX3-R 5¢-CTCGAGCCCAGCTCAGATAGCTTCTTGT-3¢ (228 bp);OsAPX4: OsAPX4-F 5¢-CTCGAGTGACAAGGCATTGTTGGAAG-3¢ and OsAPX4-R 5¢-GGTACCTCGAGCAGCTGCAGCAACAGCTACC-3¢ (180 bp);OsAPX5: OsAPX5-F 5¢-CTCGAGAGGGCAATCTTGGACATCTG-3¢ and OsAPX5-R 5¢-GGTACCTCGAGGATCAAACTTTGCCCCAAGA -3 ¢ (215 bp);OsAPX6: OsAPX6-F 5¢-CTCGAGAGGGCAATCTTGGACATCAC-3¢ and OsAPX6-R 5¢-GGTACCTCGAGGATCAAACTTTGCTCCGAGA-3¢ (227 bp); OsAPX7: OsAPX7-F 5¢-GAGCAATCTGGGTGCAAAAT-3¢ and OsAPX7-R 5¢-GGTACCTCGAGGACTCGTGGTCAGGAAAAGC-3¢ (193 bp); OsAPX8: OsAPX8-F 5¢-CTCGAGGCTGCGAAATACTCCTACGG-3¢and OsAPX8-R 5¢-GGTACCTCGAGAGGAGGTCATCAGACCATCG-3¢ (197 bp). The product lengthsamplified were in the range of 180–228 bp. The adh3gene (NCBI accession number U77637) was used as aninternal control to normalize the amount of mRNApresent in each sample (Margis-Pinheiro et al. 2005).
Table 1 Probes used for Southern blot and northern blot analyses
Gene cDNA (size in bp)a Probe (cDNA or EST)a Fragment (size in bp) Source
OsAPx1 D45423 (983) C28227 PstI/SstI (266) NIAS/RGPOsAPx2 AB053297 (1160) – BglII/SstI (543) –OsAPx3 AY382617 (1060) AY382617 NsiI/SstI (386) –OsAPx4 AK070842 (1244) C29046 HindIII/SstI (426) NIAS/RGPOsAPx5 AK073910 (1129) AU030079 XhoI/SstI (508) NIAS/RGPOsAPx6 AK061107 (1288) AU173022 BglII/NotI (424) NIAS/RGPOsAPx7 AK103344 (1392) AI978435 SstI (403) Dra. McCouchOsAPx8 AB114856 (1698) – BamHI/EcoRI (496) –
aGenebank accession numbers
303

All results were expressed as a relative quantification ofthe specific APx gene and the adh3 internal controlgene using the 2�AACT method described by Livak andSchmittgen (2001). Two independent biological repeti-tions of each experiment were performed, with fourinternal replications per experiment.
Plasmid constructs
The cDNAs encoding OsAPx3 and OsAPx6 (NCBIaccession number AY382617 and AK061107, respec-tively) were cloned in translational fusion with greenfluorescent protein (GFP) by using the Gateway�cloning technology (Invitrogen, CA, USA). Two roundsof PCR were performed in order to add the flankingattB-sites to APx cDNA clones. In the first round, theprimer pairs and annealing temperature were: OsAPx3:forward 5¢-AAAAAGCAGGCTTCACAATGTCGGCGGCGCCG-3¢ and reverse 5¢-AGAAAGCTGGGTCTTAGCCAAGCCTCTT-3¢ (48�C); OsAPx6: forward 5¢-AAAAAGCAGGCTTCACAATGGCCGTCGTCCACCGCCTC-3¢ and reverse 5¢-AGAAAGCTGGGTCTGATTCCAGTGAAATACCCTTT GG-3¢ (61�C). In thesecond round of amplification, the primer pair con-taining the entire recombination site used and theannealing temperature was: forward primer AttB1 5¢-GGGGACAAGTTTGTACAAAAAAGCAGGCT-3¢;and reverse primer AttB2 5¢-GGGGACCACTTTGTACAAGAAAGCTGGGT-3¢ (47�C).
The first recombination reaction (BP recombination)involved cloning of the attB-flanked PCR products ontothe pDONR�201 vector (Invitrogen, CA, USA) to ob-tain the pENTER-OsAPx plasmids. In the secondrecombination reaction (LR recombination) the pEN-TER-OsAPx6 plasmid was recombined with pK7FWG2to generate a COOH-terminal APx::GFP fusion underthe control of the CaMV35S promoter. The pENTR-OsAPx3 was recombined with pK7WGF2, generatingan N-terminal GFP::APx fusion driven by theCaMV35S promoter. The gateway destination vectorspK7FWG2 and pK7WGF2 (Karimi et al. 2002) werekindly provided by Dr. Mansour Karimi (Ghent Uni-versity, Belgium).
Transformation of BY-2 tobacco cells
Agrobacterium tumefaciens strain LBA4404.pBBR1MCS-5.virGN54D (van der Fits et al. 2000) was cultivatedin yeast extract Bacto tryptone medium and transformedaccording to Hofgen and Willmitzer (1988). For a stableBY-2 transformation, 100 ll of 2-day-old culture of thetransformed A. tumefaciens containing a binary vectorconstruct was cocultivated with 4 ml of a 4-day-oldculture of tobacco BY-2 cells for 48 h at 28�C, and thenthe transformed cells were selected by plating ontosolidified BY-2 medium supplemented with 500 mg l�1
vancomycin and 50 mg l�1 kanamycin and incubating at
28�C in the dark. The transformed BY-2 cells weremaintained in solidified and liquid medium containing50 mg l�1 kanamycin.
Immunofluorescent labeling
BY-2 cells, 4–5 days after subculture, were harvested bycentrifugation (100·g, 5 min) and fixed for 1 h in twovolumes of fixative solution (4% formaldehyde, 0.25 Mmannitol, 50 mM sodium phosphate pH 7.2). Fixed cellswere washed three times in PBS 1X (2.7 mM KCl,137 mM NaCl, 1.5 mM KH2PO4, 7.5 mM Na2HPO4)and incubated in 0.05% cellulase RS (Yakult, Tokyo,Japan), 0.05% macerozyme R10 (Yakult, Tokyo, Ja-pan), 0.5% Triton X-100, 0.25 M mannitol, and prote-ase inhibitors for 30 min at room temperature.Permeabilized cells were washed twice in PBS 1X andincubated at 4�C overnight in a 1:500 dilution of theprimary antibody (rabbit anti-cottonseed catalase affin-ity-purified IgGs, kindly provided by Richard Trelease)(Kunce et al. 1988) in PBS. After washing in PBS (threetimes, 5 min each), cells were incubated again at roomtemperature for 90 min with goat anti-rabbit Cy3 (Sig-ma, MO, USA)and diluted 1:5000 in PBS. Cells werewashed again for three times before observation. Con-trol experiments in which primary antibodies wereomitted were performed to verify the specificity of thelabeling.
Fluorescence microscopy analysis
Fixed and living transformed BY-2 cells were observedin a confocal laser scanning microscope LSM model 510META Zeiss Corporation equipped with a lens alphaPlan-Fluar 100X. For the mitochondrial-specific stain-ing, prior observation cells were incubated in 250 nMTMRE (Molecular Probes, OR, USA) for 15 min atroom temperature in the dark. Excitation and emissionwavelengths were 488 and 505–530 nm (BP 505–530filter) for GFP, 543 and 560–615 nm (BP 560–615 filter)for TMRE, and 543 and 560–615 nm (BP 560–615 filter)for goat anti-rabbit Cy3. All images were processed onZeiss LSM Image Browser 3.2 program (Zeiss Corpo-ration, Germany).
Results
Characterization of the rice APx gene familyby Southern blot hybridization analyses
The presence of the eight APx genes in the rice genomewas confirmed by Southern blot hybridizations. To ob-tain gene-specific DNA probes for the eight members ofthe rice ascorbate peroxidase gene family, rice EST se-quences were selected according to BLAST searches
304

against the NCBI database using the gene sequences asqueries. Homologous ESTs for each rice APx gene werethus obtained. When ESTs were not available, full-length or partial cDNAs were cloned by RT-PCR(OsAPx2, OsAPx3, and OsAPx8). After sequence vali-dation, restriction fragments corresponding to the 3-UTR of the rice APx genes were used as probes ingenomic Southern blot analyses (O. sativa var. indica cv.Taim 7). The results confirmed the presence of the eightAPx genes in the rice genome and revealed that theprobes were gene-specific, since different banding pat-terns were observed for each gene (Fig. 1). Probes dataare summarized in Table 1.
Steady-state transcript levels of the membersof the rice APx gene family
The steady-state transcript levels of the rice APx geneswere characterized by Quantitative reverse transcriptasePCR (QRT-PCR) and northern blot hybridizationanalyses. Initially, we investigated the transcript accu-mulation pattern of the APx genes in rice seeds collectedat 0.5, 12, 24, 48, 96, and 192 h after imbibition (Fig. 2).The results demonstrated that most of the APx tran-scripts were detectable from 48 h after imbibition on-wards, suggesting a coordinated regulation of theantioxidant machinery. In contrast, OsAPx8 transcript
Fig. 1 Southern blot analysis of the rice APx gene family. Ricegenomic DNA (20 lg) (O. sativa var. indica cv. Taim 7) extractedfrom seedling leaves was digested with HindIII, EcoRI, and XbaI,separated electrophoretically on a 1% (w/v) agarose gel, and
blotted onto nylon membranes (Hybond-N, Amersham). The blotswere hybridized with [a-32P]dCTP-labeled probes covering the 3¢-UTR of the rice APx genes (Table 1). DNA marker sizes areindicated on the left (in kb)
305

abundance was positively correlated with the appear-ance of green shoots in the latest intervals of the kinetics(96 and 192 h).
Using the 3-UTR gene-specific probes, northern blotanalyses were conducted with shoots and roots of two-,four-, and six-week-old rice plants (Fig. 3). Each riceAPx gene exhibited a particular pattern of transcript
accumulation, suggesting that these genes are not func-tionally redundant, but complementary. OsAPx1,OsAPx6, and OsAPx7 transcripts were preferentiallyaccumulated in roots, while increased transcript accu-mulation for OsAPx2, OsAPx3, OsAPx4, and OsAPx5were found in rice shoots. Furthermore, the analysisshowed a reduction in the transcript accumulation of the
Fig. 2 Quantitative reverse transcriptase PCR (QRT-PCR) analy-sis of the rice APx family transcript abundance in germinatingseedlings. Total RNA from rice seeds was reverse-transcribed, andeach cDNA was assayed using real-time quantitative PCR.
Seedlings were harvested at 0.5, 12, 24, 48, 96, and 192 h afterimbibition. Transcript levels were normalized relative to the adh3expression level as internal standard and time 0.5 as the unit ofexpression
306

cytosolic isoforms OsAPx1 and OsAPx2 in six-week-oldroot tissues. Transcripts of the putative thylakoid-boundisoform OsAPx8 were specifically accumulated inshoots, particularly at the 2-week-old stage, in agree-ment to its predicted subcellular localization.
We also investigated the steady-state transcript levelof the APx genes in leaves, flowers, and stems of 4-month-old rice plants by northern blot analyses (Fig. 4).
Similar transcript levels were found in the different tis-sues for OsAPx2, OsAPx4, OsAPx5, OsAPx6, OsAPx7,and OsAPx8. On the other hand, OsAPx1 transcriptswere preferentially accumulated in stems in comparisonto flowers and leaves, while OsAPx3 was specificallyexpressed in stem tissues.
Fig. 4 Northern blot analysis of the APx transcripts from leaves,stems, and flowers of 4-month-old rice plants. Total RNA (20 lg)was separated electrophoretically, blotted onto nylon membranes(Hybond-N, Amersham), and hybridized with [a-32P]dCTP-labeledprobes covering the 3¢-UTR of the rice APx genes (Table 1). Lowerpanel corresponds to ethidium bromide stained gels showing RNAloading for each sample
Fig. 3 Northern blot analysis of the APx transcripts from shootsand roots of two-, four-, and six-week-old rice plants. Total RNA(20 lg) was separated electrophoretically, blotted onto nylonmembranes (Hybond-N, Amersham), and hybridized with[a-32P]dCTP-labeled probes covering the 3¢-UTR of the rice APxgenes (Table 1). Lower panel corresponds to ethidium bromidestained gels showing RNA loading for each sample
Fig. 5 Steady-state transcript level of the rice APx genes inresponse to salt stress (OsAPx1, OsAPx3, OsAPx4, OsAPx5, andOsAPx6). a Northern blot analysis of the APx transcripts from saltstressed plants (0.25 M NaCl). Total RNA (20 lg) was separatedelectrophoretically, blotted onto nylon membranes (Hybond-N,Amersham), and hybridized with [a-32P]dCTP-labeled probes
consisting of the 3¢-UTR of the rice APx genes (Table 1). Lowerpanel corresponds to ethidium bromide stained gels showing RNAloading for each sample. b Relative transcript accumulation levelswere quantified by densitometry and normalized to the respectiveRNA loading. The values shown represent the mean value anderror bars of two independent experiments
307
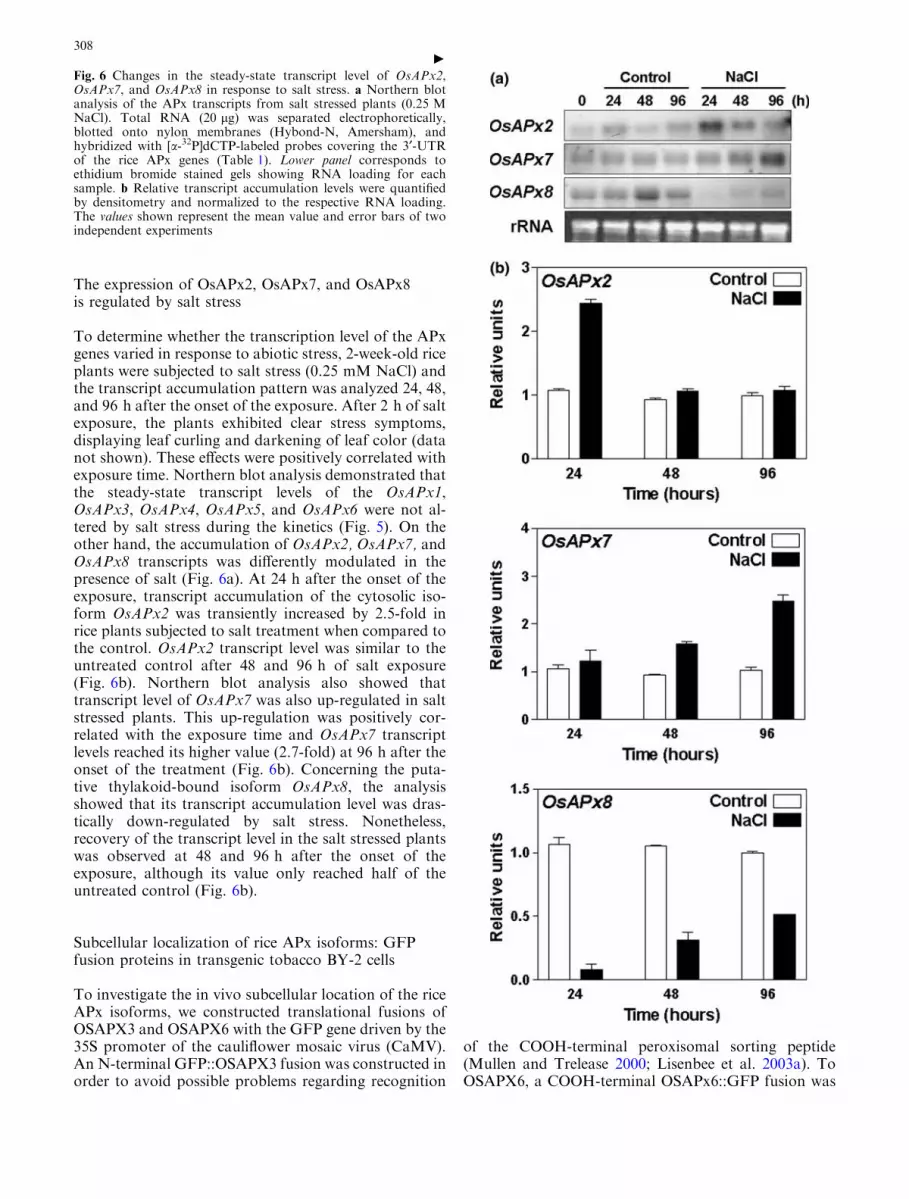
The expression of OsAPx2, OsAPx7, and OsAPx8is regulated by salt stress
To determine whether the transcription level of the APxgenes varied in response to abiotic stress, 2-week-old riceplants were subjected to salt stress (0.25 mM NaCl) andthe transcript accumulation pattern was analyzed 24, 48,and 96 h after the onset of the exposure. After 2 h of saltexposure, the plants exhibited clear stress symptoms,displaying leaf curling and darkening of leaf color (datanot shown). These effects were positively correlated withexposure time. Northern blot analysis demonstrated thatthe steady-state transcript levels of the OsAPx1,OsAPx3, OsAPx4, OsAPx5, and OsAPx6 were not al-tered by salt stress during the kinetics (Fig. 5). On theother hand, the accumulation of OsAPx2, OsAPx7, andOsAPx8 transcripts was differently modulated in thepresence of salt (Fig. 6a). At 24 h after the onset of theexposure, transcript accumulation of the cytosolic iso-form OsAPx2 was transiently increased by 2.5-fold inrice plants subjected to salt treatment when compared tothe control. OsAPx2 transcript level was similar to theuntreated control after 48 and 96 h of salt exposure(Fig. 6b). Northern blot analysis also showed thattranscript level of OsAPx7 was also up-regulated in saltstressed plants. This up-regulation was positively cor-related with the exposure time and OsAPx7 transcriptlevels reached its higher value (2.7-fold) at 96 h after theonset of the treatment (Fig. 6b). Concerning the puta-tive thylakoid-bound isoform OsAPx8, the analysisshowed that its transcript accumulation level was dras-tically down-regulated by salt stress. Nonetheless,recovery of the transcript level in the salt stressed plantswas observed at 48 and 96 h after the onset of theexposure, although its value only reached half of theuntreated control (Fig. 6b).
Subcellular localization of rice APx isoforms: GFPfusion proteins in transgenic tobacco BY-2 cells
To investigate the in vivo subcellular location of the riceAPx isoforms, we constructed translational fusions ofOSAPX3 and OSAPX6 with the GFP gene driven by the35S promoter of the cauliflower mosaic virus (CaMV).An N-terminal GFP::OSAPX3 fusion was constructed inorder to avoid possible problems regarding recognition
of the COOH-terminal peroxisomal sorting peptide(Mullen and Trelease 2000; Lisenbee et al. 2003a). ToOSAPX6, a COOH-terminal OSAPx6::GFP fusion was
Fig. 6 Changes in the steady-state transcript level of OsAPx2,OsAPx7, and OsAPx8 in response to salt stress. a Northern blotanalysis of the APx transcripts from salt stressed plants (0.25 MNaCl). Total RNA (20 lg) was separated electrophoretically,blotted onto nylon membranes (Hybond-N, Amersham), andhybridized with [a-32P]dCTP-labeled probes covering the 3¢-UTRof the rice APx genes (Table 1). Lower panel corresponds toethidium bromide stained gels showing RNA loading for eachsample. b Relative transcript accumulation levels were quantifiedby densitometry and normalized to the respective RNA loading.The values shown represent the mean value and error bars of twoindependent experiments
c308

generated since the N-terminal targeting signals tomitochondria and plastid are usually processed afterimport into the organelle (Madhusudhan et al. 2003;Chew and Whelan 2004) (Fig. 7a). Stable transgenic to-bacco BY-2 cells expressing the fusion proteins wereestablished by Agrobacterium-mediated transformationand analyzed in a confocal laser-scanning microscope(Fig. 7b–i). The fluorescence of GFP::OSAPX3 was de-tected in a reticular/circular structure around the nucleusand in spherical spots distributed diffusely throughoutthe periphery of the cytosol of BY-2 cells (Fig. 7c).Immunofluorescent labeling using the anti-cottonseedperoxisomal catalase antibody (Kunce et al. 1988) wasperformed as a peroxisome marker (Fig. 7d). Themerged image indicated that OSAPX3::GFP fusion wastargeted to peroxisomes and to a reticular compartmentcirculating the nucleus, possibly the pER (Fig. 7e).Confocal analysis of BY-2 cells stably expressing theOSAPX6::GFP fusion (Fig. 7f–i) revealed that GFPlabeling was highly specific to a small organelle, prefer-entially distributed around the cytosol periphery(Fig. 7g). The same pattern was observed when BY-2cells were stained with a mitochondria-specific fluores-cent dye, TMRE (Fig. 7h). The composite image ob-tained after merging DIC, GFP fluorescence, and TMRE
labeling images confirmed that the OSAPX6::GFP pro-tein fusion was specifically targeted to the mitochondriaof the BY-2 cells (Fig. 7i).
Discussion
In a previous study, we demonstrated the presence ofeight APx encoding genes in the nuclear genome of riceby in silico analysis. The products of these genes werepredicted to be located in different subcellular com-partments: two cytosolic isoforms (OsAPx1 andOsAPx2); two putative peroxisomal isoforms (OsAPx3and OsApx4); and four putative chloroplastic isoforms(OsAPx5 to OsAPx8) (Teixeira et al. 2004). In order tounderstand the function of each rice APx gene, we per-formed a systematic and comprehensive analysis of theexpression profile of the rice APx encoding genes and thecharacterization of the subcellular location of theirprotein products.
Firstly, we confirmed the presence of these eight APxloci in the rice genome (O. sativa var. indica cv. Taim 7)by Southern blot hybridization analysis using gene-spe-cific probes (Table 1 and Fig. 1). The results demon-strated the specificity of the probes (Fig. 1). With the
Fig. 7 Subcellular localization of APx-GFP fusion proteins intransgenic tobacco BY-2 cells. (a) Schematic representation of theT-DNA construction used in Tobacco BY-2 cells transformation.(b–i) Confocal analysis of BY-2 cells expressing GFP::OSAPX3 (b–e) and OSAPX6::GFP (f–i) fusions. (b,f) Differential interference
contrast (DIC) images. (c,g) GFP fluorescence of stable transfor-mant BY-2 cells. (d) Imumunofluorescence labeling with anti-cottonseed catalase. (e) Merged image from (b–d). (h) Mitochon-drial-specific staining (TMRE) fluorescence. (i) Merged image from(f–h). Bars in (b–i)=10 lm
309

exception of the HindIII digestion on the OsAPx1 blot,the restriction fragment banding profiles obtained in thegenomic Southern blot analysis matched perfectly withvirtual restriction analysis based on the rice genomicsequence (data not shown). The difference presented inthe OsAPx1 analysis may be due to the existence ofpolymorphisms between the cultivars used in the ricegenome project and in our experiments. In conclusion,the southern experiments confirmed our analysis indi-cating the presence of eight APx loci in the rice genome.
Subcellular localization of rice APx isoforms
The putative subcellular location of the rice APx pro-teins was previously assigned based on the presence oftargeting signal and transmembrane domains (Teixeiraet al. 2004). In this work, we investigated the in vivosubcellular location of two rice APx proteins (OSAPX3and OSAPX6) by the stable expression of GFP trans-lational fusions in BY-2 tobacco cells (Fig. 7).
The GFP fluorescence of BY-2 cells expressing theGFP::OSAPX3 fusion was observed in sphericalorganelles dispersed in the cytosol, but was also associ-ated with a reticular network around BY-2 nucleus(Fig. 7c). The spherical organelles were characterized asperoxisomes by the BY-2 cell staining with a peroxi-somal protein marker (anti-catalase) (Fig. 7d). Themerged image confirmed that the GFP::OSPAX3 fusionwas targeted to the peroxisomes (Fig. 7e). Nevertheless,it was previously shown that APx isoforms are indirectlysorted to BY-2 cell peroxisomes via an endoplasmicreticulum subdomain, also known as pER (peroxisomalendoplasmatic reticulum) (Mullen and Trealease 2000;Lisenbee et al. 2003a). In addition, Lisenbee et al.(2003b) have shown that the pER can be observed as areticular structure only when a peroxisomal APx-GFPfusion is overexpressed. Since our GFP::OSAPX3 fusionprotein was driven by the CaMV 35S promoter, weconcluded that OSAPX3 is a peroxisomal isoformindirectly sorted via pER, as shown for Arabidopsis andpumpkin peroxisomal APx isoforms.
The GFP fluorescence of BY-2 cells expressing theOSAPX6::GFP fusion was observed in isolated smallorganelles concentrated around the periphery of the cell(Fig. 7g). The same cells stained with the mitochondria-specific dye TMRE (Zottini et al. 2002) (Fig. 7h) re-vealed that the OSAPX6::GFP fusion was specificallytargeted to the mitochondria (Fig. 7i). Based on in silicoanalysis, OSAPX6 was predicted to be targeted to thechloroplast (Teixeira et al. 2004). The majority ofmitochondrial and plastid proteins are encoded by nu-clear genes and are targeted to their final organelle aftertranslation. In general, these proteins are preceded by anN-terminal targeting peptide containing hydroxilated-amino acid residues, which is processed after import intothe organelle. Plastid and mitochondrial targeting sig-nals are very similar to each other, making it difficult topredict the exact subcellular location of these proteins
(Chew and Whelan 2004). Furthermore, there are a largenumber of proteins that are dual-targeted to mitochon-dria and plastids (Silva-Filho 2003). For instance, inArabidopsis, the stromal isoform (At4g08390) wassimultaneously targeted to chloroplast and mitochon-dria (Chew et al. 2003b). On the other hand, theN. tabacum stromal isoform is exclusively targeted to theplastids of BY-2 cells, suggesting that a distinct genemay encode the mitochondrial isoform (Madhusudhanet al. 2003). In tomato, western blot analyses suggestthat the mitochondrial and choroplastic APx isoformsmay have originated from different genes (Mittova et al.2004b). To our knowledge, OSAPX6 is the first APxisoform known to be exclusively located in mitochon-dria. It is not possible to rule out, however, that in ricethis protein may also be found in chloroplasts, in addi-tion to a mitochondrial location.
Enzymatic activity of the additional ascorbate-glu-tathione cycle enzymes (MDHAR, DHAR, and GR)was also found in the mitochondria of tomato, potato,and peas (Jimenez et al. 1997; De Leonardis et al. 2000;Mittova et al. 2004a). Interestingly, Obara et al. (2002)demonstrated that the dual-targeting of the ArabidopsisMDHAR (At1g63940) is related to multiple transcrip-tion initiation sites of a single gene. In addition, analysisof in vitro import revealed the dual targeting of APx,MDHAR, and GR in Arabidopsis to both chloroplastand mitochondria, while DHAR was only targeted to alater one (Chew et al. 2003b). Similarly, the dual tar-geting of a pea GR isoform has been reported (Chewet al. 2003a). The mitochondrial ascorbate-glutathionecycle was suggested to be very important in the controlof the photosynthetic stress and during senescence inpeas (Jimenez et al. 1997; Jimenez et al. 1998), and alsoin protecting tomato leaves and roots against salt stress(Mittova et al. 2004b). Together, these data demonstratethe importance of the ascorbate-glutathione cycle to theROS scavenging in the mitochondria.
Gene expression analyses reveal the functional diversityof the eight members of the rice APx gene family
Although the expression profile of individual APxencoding genes is largely reported in the literature, therehas been no systematic study of the regulation of theexpression patterns of the APx gene family in a singleplant species. Furthermore, there are few reportsdescribing the developmental regulation of the APxgenes. To evaluate the importance of the APx genesduring seed germination, we collected rice seedlings at0.5, 12, 24, 48, 96, and 192 h after imbibition andinvestigated the transcript accumulation by QRT-PCRanalyses (Fig. 2). The results demonstrated that almostall rice APx transcripts were maximally accumulatedafter 48 h of imbibition, coinciding with the emergenceof the radicle tip from the seed coat. This synchroniza-tion of expression patterns revealed that the APx genesare coordinately regulated in the initial stages of plant
310

development, possibly related to the scavenging of ROSproduced due to the continuous mechanical stressedtissues of the radicle tip and to the mobilization of theendosperm carbohydrate/lipid reserves. However, theexpression pattern of the APx isoforms might also berelated to protecting the cells against the harmful effectsof high ROS concentrations that are produced in highlyproliferating tissues such as radicle tip. Accordingly, theexpression of the cytosolic APx isoform was previouslyreported in tomato seeds, but also in the root and shootapical meristems (Gadea et al. 1999). On the other hand,OsAPx8 transcript abundance levels increased in olderseedlings, correlated with the appearance of etiolatedtissues. Curiously, the spinach thylakoid-bound APxwas also specifically expressed in the etiolated tissues(Mano et al. 1997; Yoshimura et al. 2002).
The availability of specific probes for each member ofthe rice APx family (Table 1) allowed us to assess thespecific expression profiles of these genes by northernblot. The analyses of the transcript accumulation of APxin seedlings showed that the cytosolic isoforms havedistinct expression patterns, with OsAPx1 accumulatingpreferentially in roots, while OsAPx2 is expressed inshoots and roots (Fig. 3). In roots, the expression ofboth cytosolic genes was reduced in older plants, sug-gesting their involvement in the scavenging of ROS inthe initial stages of the plant development. In tomatoand Arabidopsis plants, LeAPx1 and AtAPx1 genesshow similar expression profile to OsAPx1 (Gadea et al.1999; Fourcroy et al. 2004). In addition, expressionanalysis of promoter-GUS fusions in transgenic plantsshowed that LeAPx1 is differentially expressed duringplant development (Gadea et al. 1999). Furthermore,Arabidopsis KO-plants for the cytosolic AtAPx1 isoformshow delayed development and flowering (Pnueli et al.2003).
In higher plants, peroxisomes are classified accord-ing to their function depending on the cell type: (i)glyoxysomes, present in storage organs, such as endo-sperm and cotyledons, are specialized for the b-oxida-tion of fatty acids and the glyoxylate cycle; (ii) leafperoxisomes, present in the photosynthetic organs andimplicated in photorespiratory metabolism; and (iii)unspecialized peroxisomes, present in roots and stems(Nishimura et al. 1996). In addition, microarray anal-yses performed with 263 Arabidopsis peroxisomal genesrevealed the existence of specific groups of genes foreach peroxisome class (Kamada et al. 2003). The riceperoxisomal isoforms, OsAPx3 and OsAPx4, werepreferentially expressed in seedling shoots (Fig. 3).Furthermore, OsAPx3 transcript was exclusively accu-mulated in the stems of 4-month-old plants (Fig. 4),suggesting that this isoform may be closely related tothe unspecialized peroxisomes present in these tissues.The majority of studies on cDNA isolation and anal-ysis of expression of the APx peroxisomal isoformswere performed using leaf and cotyledon tissues, sug-gesting that these isoforms play important roles in thedetoxification of H2O2 in these tissues (Zhang et al.
1997; Ishikawa et al. 1998; Yoshimura et al. 2000). Inaddition, the expression of the Arabidopsis peroxisso-mal AtAPx3 gene (At4g35000) was twofold lower inroots when compared to the shoots, and threefoldlower when compared to the cotyledons (Kamada et al.2003). Taken together, the results suggest that peroxi-some APx isoforms are largely related to the ROSdetoxification in the shoots of the plants, acting pri-marily in leaf peroxisomes, unspecialized stem peroxi-somes, and glyoxysomes.
The transcript accumulation profiles of OsAPx5 andOsAPx6 in shoots and roots revealed differential reg-ulation of these genes (Fig. 3). Although these geneshave been likely originated by a very recent duplicationevent as indicated by the high identity of their nucle-otide sequence and by their in tandem organization inthe rice genome (Teixeira et al. 2004), the results sug-gest divergences in their promoter regions. On theother hand, the putative chloroplastic isoform OsAPx7accumulated preferentially in root tissues where no truechloroplasts were present. However, OsAPx7 couldencode a root-expressed plastid stromal isoform, suchas the stromal spinach isoform, which is also highlyexpressed in roots (Yoshimura et al. 2002). In agree-ment, the tobacco stromal isoforms were specificallytargeted to the BY-2 cell plastids (Madhusudhan et al.2003).
In spinach, tobacco, and pumpkin, the stromal andthe thylakoid-bound isoforms are generated by alterna-tive splicing of a single gene (Ishikawa et al. 1996; Manoet al. 1997; Yoshimura et al. 2000). In spinach, a splicingregulatory cis-element (SRE) identified in the chloro-plastic APx encoding gene has been shown to beresponsible for the tissue-specific alternative splicing.Although the transcription of the choroplastic spinachAPx gene is almost equal in shoots and roots, the rela-tive level of the transcripts encoding the stromal andthylakoid-bound isoforms were different. In leaves, bothtranscripts are present in the same proportion. In con-trast, 95% of the root transcripts correspond to thestromal isoform, suggesting that the thylakoid-boundisoform is not expressed in non-etiolated tissues due tothe absence of chloroplasts in these tissues (Yoshimuraet al. 2002). In agreement, transcripts of the OsAPx8gene accumulated only in rice shoots (Fig. 3). Associ-ated with the presence of a putative N-terminal chloro-plastic sorting peptide and a COOH-terminaltransmembrane domain, these results strongly suggestthat OsAPx8 encodes a thylakoid-bound isoform.Therefore, in contrast to spinach, tobacco, and pump-kin, two different genes encode the stromal and thethylakoid-bound isoforms in rice, and that the tran-scription of these genes is differentially regulated in atissue-specific manner.
We have also investigated the regulation of the riceAPx genes in response to salt stress. Salinity causesdisruption of ionic, osmotic, and cell-water homeostasis,resulting in perturbation of plant growth, development,and reproduction (Zhu 2001). Our results showed that
311

transcript accumulation of the majority of the rice APxgenes (OsAPx1, OsAPx3, OsAPx4, OsAPx5, andOsAPx6) remained fairly constant when plants weretreated with 0.25 M NaCl (Fig. 5). Accordingly, tran-scripts levels of the Arabidopsis and spinach peroxisomalisoforms were unaffected by salt stress (Zhang et al.1997; Yoshimura et al. 2000).
Three rice APx genes (OsAPx2, OsAPx7, andOsAPx8) showed altered transcript levels in response toNaCl treatment (Fig. 6). The cytosolic OsAPx2 tran-script level of the increased 2.5-fold after 24 h of saltstress returned to the control level after 48 h of treat-ment, suggesting a role of OsAPx2 in response to thestress and in controlling the H2O2 concentration in thecytosol. Elevation of APx activity was previously re-ported in rice plants subjected to salt stress (Lee et al.2001). The rapid increase in OsAPx2 mRNA levelsmay be related to the maintenance of high APx activityin the cytosol to protect the cellular componentsagainst the effects of ROS produced as consequence ofthe oxidative stress. Analysis of KO-plants demon-strated that the absence of the cytosolic isoform APX1(KO-AtAPx1) causes suppression of the photosyntheticactivity, increase of H2O2, and protein oxidation, sug-gesting that the cytosolic APX protects the chloroplastduring light stress (Davletova et al. 2005). In addition,the increase of the transcript level of the cytosolic APxisoforms in response to salt stress was also reported inNicotiana plumbaginifolia (Savoure et al. 1999), rice(Menezes-Benavente et al. 2004b), and maize (Menezes-Benavente et al. 2004a). Conversely, the cytosolic APxisoform was down-regulated when spinach plants weresubmitted to salt treatment (Yoshimura et al. 2000).Nevertheless, Mittler et al. (1998) showed that theArabidopsis cytosolic isoform AtAPx1 is post-transla-tionally regulated during PCD mediated by pathogenattack, indicating that APx transcript accumulationmay not always correlate with protein concentration orenzymatic activity.
The OsAPx7 transcripts accumulated progressivelyin the salt-stressed plants, reaching a maximum (2.7-fold) level of 96 h after the onset of salt treatment(Fig. 6). This result suggests that OsAPx7 might play arole in the defense mechanism against oxidative stresscaused by salinity. In contrast, spinach plants subjectedto oxidative stress did not show alterations in chloro-plastic APx neither in transcript or protein levels(Yoshimura et al. 2000). An opposite result was ob-served for OsAPx8 transcript accumulation, which wasstrongly suppressed in plants subjected to salt treat-ment (Fig. 6). Similar results were obtained by Yabutaet al. (2002) with the tobacco thylakoid-bound isoformwhen plants were subjected to light stress, suggestingthat these isoforms are prematurely suppressed duringoxidative stress. The thylakoid-bound isoform wasfurther suggested to be a limiting factor in the anti-oxidant system during the photo-oxidative stress inArabidopsis and wheat (Yabuta et al. 2002; Dannaet al. 2003).
Conclusions
Taken together, these results reveal the importance ofdifferent isoforms of ascorbate peroxidase in the elimi-nation of ROS, both in plant defense during an oxida-tively stressful situation and during plant development,suggesting coordinated regulation of the antioxidantenzymes. In addition, the large number of APx encodinggenes in rice, the differential regulation of each memberof this gene family, and the presence of their proteins indifferent subcellular compartments also highlight thevalue of the specificity of the response and the com-plexity of the antioxidant system in plants.
Acknowledgements We greatly acknowledge Dr. Peter Waterhouseand Dr. John Watson for critically reading the manuscript. We aregrateful to Bernardo Pascarelli and Dr. Radovan Borojevic forhelping with the confocal analysis. We thank Dr. Marcio AlvesFerreira, Dr. Gilberto Saccheto Martins, and Dr. Maite Vaslin forhelpful discussions and suggestions. We greatly acknowledge thefollowing organizations and researchers for providing us with ESTclones, vectors, and antibodies: Dr. Takuji Sasaki, National Insti-tute of Agrobiological Resources-Rice Genome Research Program(NIAS-RGP), Japan; Dr. Susan McCouch, Cornell University,USA; Dr. Mansour Karimi, Ghent University, Belgium; and Dr.Richard Trelease, Arizona State University, USA. This work wassupported by FAPERJ, CNPq, and CAPES.
References
Agrawal GK, Jwa N, Iwahashi H, Rakwal R (2003) Importance ofascorbate peroxidases OsAPx1 and OsAPx2 in the rice patho-gen response pathway and growth and reproduction revealed bytheir transcriptional profiling. Gene 322:93–103
Alscher RG, Donahue JL, Cramer CL (1997) Reactive oxygenspecies and antioxidants: relationships in green cells. PhysiolPlant 100:224–233
Asada K (1999) The water-water cycle in chloroplasts: scavengingof active oxygen and dissipation of excess photons. Annu RevPlant Physiol Plant Mol Biol 50:601–639
Causse MA et al (1994) Saturated molecular map of the rice gen-ome based on an interspecific backcross population. Genetics138:1251–1274
Chang CC, Ball L, Fryer MJ, Baker N, Karpinski S, MullineauxPM (2004) Induction of ASCORBATE PEROXIDASE 2expression in wounded Arabidopsis leaves does not involveknown wound-signalling pathways but is associated withchanges in photosynthesis. Plant J 38:499–511
Chew O, Whelan J (2004) Just read the message: a model forsorting of proteins between mitochondria and chloroplasts.Trends Plant Sci 9:318–319
Chew O, Rudhe C, Glaser E, Whelan J (2003a) Characterization ofthe targeting signal of dual-targeted pea glutathione reductase.Plant Mol Biol 53:341–356
Chew O, Whelan J, Millar H (2003b) Molecular definition of theascorbate-glutathione cycle in Arabidopsis mitochondria revealsdual targeting of antioxidant defenses in plants. J Biol Chem278:46869–46877
Church GM, Gilbert W (1984) Genomic sequencing. Proc NatlAcad Sci 81:1991–1995
Danna CH, Bartoli CG, Sacco F, Ingala LR, Santa-Marıa GE,Guiamet JJ, Ugalde RA (2003) Thylakoid-bound ascorbateperoxidase mutant exhibits impaired electron transport andphotosynthetic activity. Plant Physiol 132:2116–2125
Dat J, Vandenabeele S, Vranova E, Van Montagu M, Inze D, VanBreusegem F (2000) Dual action of the active oxygen speciesduring plant stress response. Cell Mol Life Sci 57:779–795
312

Davletova S, Rizhsky L, Liang H, Shengqiang Z, Oliver DJ, CoutuJ, Shulaev V, Schlauch K, Mittler R (2005) Cytosolic ascorbateperoxidase 1 is a central component of the reactive oxygen genenetwork of arabidopsis. Plant Cell 17:268–281
De Leonardis S, Dipierro N, Dipierro S (2000) Purification andcharacterization of an ascorbate peroxidase from potato tubermitochondria. Plant Physiol Biochem 38:773–779
Feinberg AP, Vogelstein B (1983). A technique for radiolabellingDNA restriction endonuclease fragments to high specific affin-ity. Anal Biochem 132:6–13
van der Fits L, Deakin E, Hoge H, Memelink J (2000) The ternarytransformation system: constitutive virG on a compatible plas-mid dramatically increases Agrobacterium-mediated planttransformation. Plant Mol Biol 43:495–502
Fourcroy P, Vansuyt G, Kushnir S, Inze D, Briat J (2004) Iron-regulated expression of a cytosolic ascorbate peroxidase en-coded by the APX1 gene in arabidopsis seedlings. Plant Physiol134:605–613
Fridovich I (1998) Oxygen toxicity: a radical explanation. J ExpBot 201:1203–1209
Fryer MJ, Ball L, Oxborough K, Karpinski S, Mullineaux PM,Baker NR (2003) Control of Ascorbate Peroxidase 2 expressionby hydrogen peroxide and leaf water status during excess lightstress reveals a functional organization of Arabidopsis leaves.Plant J 33:691–705
Furlani AMC, Furlani PR (1998) Composicao e pH de solucoesnutritivas para estudos fisiologicos e de selecao de plantas emcondicoes nutricionais adversas. Technical bulletin 121, Insti-tuto Agronomico de Campinas pp 1–34
Gadea J, Conejero V, Vera P (1999) Developmental regulation of acytosolic ascorbate peroxidase gene from tomato plants. MolGen Genet 262:212–219
Halliwell B, Gutteridge JM (1999) Free radicals in biology andmedicine. Oxford University Press, New York p 936
Hofgen R, Willmitzer L (1988) Storage of competent cells forAgrobacterium transformation. Nucleic Acids Res 16:9877
Ishikawa T, Sakai K, Takeda T, Shigeoka S (1995) Cloning andexpression of cDNA encoding a new type of ascorbate peroxi-dase in spinach. FEBS Lett 367:28–32
Ishikawa T, Sakai K, Yoshimura K, Takeda T, Shigeoka S (1996)cDNAs encoding spinach stromal and thylakoid-bound ascor-bate peroxidase, differing in the presence or absence of their 3¢-coding regions. FEBS Lett 384:289–293
Ishikawa T, Yoshimura K, Sakai K, Tamoi M, Takeda T, Shi-geoka S (1998) Molecular characterization and physiologicalrole of a glyoxysome-bound ascorbate peroxidase from spinach.Plant Cell Physiol 39:23–34
Jimenez A, Hernandez JA, del Rıo LA, Sevilla F (1997) Evidencefor the presence of the ascorbate-glutathione cycle in mito-chondria and peroxisomes of pea leaves. Plant Physiol 114:275–284
Jimenez A, Hernandez JA, Pastori G, del Rıo LA, Sevilla F (1998)Role of the ascorbate-glutathione cycle of mitochondria andperoxisomes in the senescence of pea leaves. Plant Physiol118:1327–1335
Kamada T, Nito K, Hayashi H, Mano S, Hayashi M, NishimuraM (2003) Functional differentiation of peroxisomes revealed byexpression profiles of peroxisomal genes in Arabidopsis thaliana.Plant Cell Physiol 44:1275–1289
Karimi M, Inze D, Depicker A (2002) GATEWAY� vectors forAgrobacterium-mediated plant transformation. Trends Plant Sci7:193–195
Karpinski S, Escobar C, Karpinska B, Creissen G, Mullineaux PM(1997) Photosynthetic electron transport regulates the expres-sion of cytosolic ascorbate peroxidase genes in arabidopsisduring excess light stress. Plant Cell 9:627–640
Kato K, Matsumoto T, Koiwai S, Mizusaki S, Nishida K, NogushiM, Tamaki E (1972) Liquid suspension culture of tobacco cells.In: Terui G (ed) Ferment technology today. Society of Fer-mentation Technology, Osaka, pp 689–695
Kunce CM, Trelease RN, Turley RB (1988) Purification and bio-synthesis of cottonseed (Gossypium hirsutum L). Biochem J251:147–155
Lee HD, Kim YS, Lee CB (2001) The inductive responses of theantioxidant enzymes by salt stress in the rice (Oryza sativa L.). JPlant Physiol 158:737–745
Lisenbee CS, Heinze M, Trelease RN (2003a) Peroxisomal ascor-bate peroxidase resides within a subdomain of rough endo-plasmic reticulum in wild-type arabidopsis cells. Plant Physiol132:870–882
Lisenbee CS, Karnik SK, Trelease RN (2003b) Overexpression andmislocalization of a tail-anchored GFP redefines the identity ofperoxisomal ER. Traffic 4:491–501
Livak KJ, Schmittgen TD (2001) Analysis of relative expressiondata using real-time quantitative PCR and the 2�DDCt method.Methods 25:402–408
Madhusudhan R, Ishikawa T, Sawa Y, Shigeoka S, Shibata H(2003) Characterization of an ascorbate peroxidase in plastidsof tobacco BY-2 cells. Physiol Plant 117:550–557
Mano S, Yamaguchi K, Hayashi M, Nishimura M (1997) Stromaland thylakoid-bound ascorbate peroxidases are produced byalternative splicing in pumpkin. FEBS Lett 413:21–26
Margis-Pinheiro M, Zhou XR, Zhu QH, Dennis ES, UpadhyayaNM (2005) Isolation and characterization of a Ds-tagged rice(Oryza sativa L.) GA-responsive dwarf mutant defective in anearly step of the gibberellin biosynthesis pathway. Plant CellRep 23:819–33
Menezes-Benavente L, Kernodle SP, Margis-Pinheiro M, Scanda-lios JG (2004a) Salt-induced antioxidant metabolism defenses inmaize (Zea mays L.) seedlings. Redox Rep 9:29–36
Menezes-Benavente L, Teixeira FK, Kamei CL, Margis-PinheiroM (2004b) Salt stress induces expression of genes encodingantioxidant enzymes in seedlings of a Brazilian indica rice(Oryza sativa L.). Plant Sci 166:323–331
Mittler R, Feng X, Cohen M (1998) Post-transcriptional suppres-sion of cytosolic ascorbate peroxidase expression during path-ogen-induced programmed cell death in tobacco. Plant Cell10:461–473
Mittler R, Vanderauwera S, Gallery M, Van Breusegem F (2004)Reactive oxygen gene network of plants. Trends Plant Sci9:490–498
Mittova V, Guy M, Tal M, Volokita M (2004a) Salinity up-regu-lates the antioxidative system in root mitochondria and per-oxisomes of the wild salt-tolerant tomato species Lycopersiconpennellii. J Exp Bot 55:1105–1113
Mittova V, Theodoulou FL, Kiddle G, Volokita M, Tal M, FoyerCH, Guy M (2004b) Comparison of mitochondrial ascorbateperoxidase in the cultivated tomato, Lycopersicon esculentum,and its wild, salt-tolerant relative, L. pennellii—a role for matrixisoforms in protection against oxidative damage. Plant CellEnviron 27:237–250
Miyake C, Cao WH, Asada K (1993) Purification and molecularproperties of thylakoid-bound acorbate peroxidase in spinachchloroplasts. Plant Cell Physiol 34:881–889
Mullen RT, Trelease RN (2000) The sorting signals for peroxi-somal membrane-bound ascorbate peroxidase are within its C-terminal tail. J Biol Chem 275:16337–16344
Mullen RT, Lisenbee CS, Miernyk JA, Trelease RN (1999) Per-oxisomal membrane ascorbate peroxidase is sorted to a mem-branous network that resembles a subdomain of theendoplasmic reticulum. Plant Cell 11:2167–2185
Mullineaux P, Karpinski S (2002) Signal transduction in responseto excess light: getting out of the chloroplast. Curr Opin PlantBiol 5:43–48
Murashige T, Skoog F (1962) A revised medium for rapid growthand bioassays with tobacco tissue cultures. Plant Physiol15:473–497
Nishimura M, Hayashi M, Kato A, Yamaguchi H, Mano S (1996)Functional transformation of microbodies in higher plant cells.Cell Struct Funct 21:387–393
313

Noctor G, Foyer CH (1998) Ascorbate and glutathione: keepingactive oxygen under control. Annu Rev Plant Physiol Plant MolBiol 49:249–79
Obara K, Sumi K, Fukuda H (2002) The use of multiple tran-scription starts causes the dual targeting of Arabidopsis putativemondehydroascorbate reductase to both mitochondria andchloroplasts. Plant Cell Physiol 43:697–705
Orozco-Cardenas ML, Narvaez-Vasquez J, Ryan CA (2001)Hydrogen peroxide acts as a second messenger for the inductionof defense genes in tomato plants in response to wounding,systemin, and methyl-jasmonate. Plant Cell 13:179–191
Pnueli L, Liang H, Rozenberg M, Mittler R (2003) Growth sup-pression, altered stomatal responses, and augmented inductionof heat shock protein in cytosolic ascobate peroxidase (Apx1)-deficient Arabidopsis plants. Plant J 34:187–203
Ragueh F, Fescure N, Roby D, Marco Y (1989) Gene expression inNicotiana tabacum in response to compatible and incompatibleisolates of Pseudomonas solonaciarum. Physiol Mol Plant Pa-thol 35:23–33
Rizhsky L, Davletova S, Liang H, Mittler R (2004) The zing fingerprotein Zat12 is required for cytosolic ascorbate peroxidase 1expression during oxidative stress in Arabidopsis. J Biol Chem279:11736–11743
Sato Y, Murakami T, Funatsuki H, Matsuba S, Saruyama H,Tanida M (2001) Heat shock-mediated APX gene expressionand protection against chilling injury in rice seedlings. J ExpBot 52:145–151
Savoure A, Thorin D, Xeu-Jun H, Van Montagu M, Inze D,Verbruggen N (1999) NaCl and CuSO4 treatments trigger dis-tinct oxidative defense mechanisms in Nicotiana plumbaginifoliaL. Plant Cell Environ 22:387–396
Scandalios JG (2002) The rise of ROS. Trends Biochem Sci 27:483–486
Shigeoka S, Nakano Y, Kitaoka S (1980) Metabolism of hydrogenperoxide in Euglena gracilis Z by L-ascorbic acid peroxidase.Biochem J 186:377–380
Shigeoka S, Ishikawa T, Tamoi M, Miyagawa Y, Takeda T,Yabuta Y, Yoshimura K (2002) Regulation and function ofascorbate peroxidase isoenzymes. J Exp Bot 53:1305–1319
Silva-Filho MC (2003) One ticket for multiple destinations: dualtargeting of proteins to distinct subcellular locations. Curr OpinPlant Biol 6:589–595
Teixeira FK, Menezes-Benavente L, Margis R, Margis-Pinheiro M(2004) Analysis of the molecular evolutionary history of theascorbate peroxidase gene family: inferences from the ricegenome. J Mol Evol 59:761–770
Vandenabeele S, Van Der Kelen K, Dat J, Gadjev I, Boonefaes T,Morsa S, Rottiers P, Slooten L, Van Montagu M, Zabeau M,Inze D, Van Breusegem F (2003) A comprehensive analysis ofhydrogen peroxide-induced gene expression in tobacco. ProcNatl Acad Sci USA 100:16113–16118
Yabuta Y, Motoki T, Yoshimura K, Takeda T, Ishikawa, Shi-geoka S (2002) Thylakoid membrane-bound ascorbate peroxi-dase is a limiting factor of antioxidative systems under photo-oxidative stress. Plant J 32:915–925
Yoshimura K, Yabuta Y, Ishikawa T, Shigeoka S (2000) Expres-sion of spinach ascorbate peroxidase isoenzymes in response tooxidative stresses. Plant Physiol 123:223–233
Yoshimura K, Yabuta Y, Ishikawa T, Shigeoka S (2002) Identifi-cation of a cis element for tissue-specific alternative splicing ofchloroplast ascorbate peroxidase pre-mRNA in higher plants. JBiol Chem 277:40623–40632
Zhang H, Wang J, Nickel U, Allen RD, Goodman HM (1997)Cloning and expression of an Arabidopsis gene encoding aputative peroxisomal ascorbate peroxidase. Plant Mol Biol34:967–971
Zhu JK (2001) Plant salt tolerance. Trends Plant Sci 6:66–67Zottini M, Formentin E, Scattolin M, Carimi F, Lo Schiavo F,
Terzi M (2002) Nitric oxide affects plant mitochondrial func-tionality in vivo. FEBS Lett 515:75–78
314