retinal and choroidal neovascularization in a transgenic mouse model of sickle cell disease
TRANSCRIPT

American Journal ofPathology, Vol. 145, No. 2, August 1994Copyright C) American Societyfor Investigative Pathology
Animal ModelRetinal and Choroidal Neovascularization in aTransgenic Mouse Model of Sickle Cell Disease
Gerard A. Lutty,* D. Scott McLeod,tAgathe Pachnis,- Frank Costantini,AMary E. Fabry,§ and Ronald L. Nagel§From the Wilmer Ophthalmological Institute,* TheJohnsHopkins School ofMedicine, Baltimore, Maryland; TheJohns Hopkins Applied Physics Laboratory,tLaurel, Maryland; Department of Genetics andDevelopment,* College ofPhysicians and Surgeons,Columbia University, New York, New York; and Division ofHematology,5 The Albert Einstein College of Medicine,Bronx, New York
A complication ofsickle ceU disease is prolifera-tive retinopathy. We investigated the eyesfrom atransgenic mouse model of sickle ceU disease(CH.(S[iMDD] type) to determine ifpathologicalchanges occurred in their retinas and choroids.One retina from each animal was processedby flat-embedding adenosine diphosphatase-reacted retinas in glycol methacrylate. The fel-low eyefrom each animal was embedded wholein glycol methacrylate for histopathologicalanalysis of aU ocular structures. Retinal vascu-lar occlusions resulted in nonperfused areas ofretina and arterio-venous anastomoses. Intra-and extraretinal neovascularization was ob-served adjacent to nonperfused areas. Retinalpigmented lesions wereformed by the migrationof retinal pigment epithelial cells into sensoryretina, often ensheathing choroidal neovascu-larization. The incidence ofthis bilateral chorio-retinopathy was 30% in animals older than 15months of age. The ocular histopathologicalchanges we observed in the mouse model mim-icked many aspects of human proliferativesickle ceUl retinopathy. Furthermore, this is thefirst geneticaly derived animal model forchorio-retinal neovascularization. (AmJ Pathol1994, 145:490-497)
Sickle cell anemia results from homozygosity for asingle point mutation in the human 3-globin gene. Theproduct of the mutated gene is a globin subunit (13S)that is incorporated into hemoglobin tetramers, whichpolymerize when deoxygenated, causing the eryth-rocytes to sickle and become less deformable. Theaccelerated destruction of these erythrocytes (hemo-lysis) produces a preponderance of young red cells.Both young deformable red cells and very dense redcells, which include irreversibly sickled cells, arethought to be involved in vaso-occlusions that occurin most organs of the body.1 In the human retina, vaso-occlusion occurs in the far peripheral vasculature,creating ischemic areas; proliferative sickle cell reti-nopathy (PSR) occurs when neovascularization formsadjacent to the ischemic retinal tissue.2 Another le-sion characteristic of sickle cell retinopathy, the blacksunburst lesion, is formed by migration of retinal pig-ment epithelial cells into sensory retina.3
Recently, several transgenic mouse models ofsickle cell disease have been created by injectingconstructs of human 3S-globin or PS variants in com-bination with the human a-globin gene into fertilizedmouse eggs.47 We investigated ocular histopatho-logical changes in the most extensively characterizedtransgenic mouse model for sickle cell anemia, the(aoHs3[I3MDD]) type, which was created by co-
injecting two constructs into fertilized eggs: one con-struct contained the locus control region linked to thehuman sickle P-globin gene and the other, the locuscontrol region plus the human a-globin gene. Thetransgenic line produced was bred into a backgroundhomozygous for a deletion of the mouse 3major globin
Supported by Research to Prevent Blindness and NIH grantsHL45922 (GL), EY01765 (Wilmer Ophthalmological Institute), andHL38655CRLN (MF).Accepted for publication March 18, 1994.Address reprint requests to Dr. Gerard A. Lutty, Wilmer Ophthal-
mological Institute, Johns Hopkins University School of Medicine,600 North Wolfe Street, Baltimore, MD 21287-9115.
490

A Mouse Model of Sickle Cell Retinopathy 491AJP August 1994, Vol. 145, No. 2
gene,8 a strategy that resulted in greater expressionof human P3s and reduced production of mouse
p3-globin.6 In animals from the resultant type (a HfS[f3MDD]) that were used in our study, 72.7 ± 2.4% ofthe 3-globin chains expressed were human P3s and44.7 ± 2.4% of the a-globin chains were human. Thetransgenes seem to be expressed hemizygously,suggesting that homozygosity may be lethal. Themice displayed normal murine mean corpuscular he-moglobin, mean red cell volume, and density distri-bution width with slightly elevated mean corpuscularhemoglobin concentration and total hemoglobin.6 Allof these observations indicated that introduction andexpression of the transgenes on a mouse f-major de-letional background had resulted in a mouse that isnot phenotypically thalassemic. The effects of the hu-man Ps hemoglobin were apparent in that slow de-oxygenation resulted in 95% typical sickled cells.7 Inaddition, magnetic resonance imaging, functional re-
nal tests, and histopathology revealed abnormalitiesin spleens and kidneys of some animals that were
comparable to abnormalities in human sickle cellanemia.7We investigated changes in the retina from one eye
of each animal of the aHfS[IMDD] transgenic mouse
type using a histological technique we developed tostudy the human retinal vasculature in dual perspec-
tive: adenosine diphosphatase-(ADPase) reactedretinas flat-embedded in glycol methacrylate blocks.9With this technique, the pattern of the retinal vascu-
lature (the site of the ADPase reaction product) is vi-sualized en bloc with darkfield illumination. Regionsof interest are then cut in cross section, offering thesecond analytical perspective. The fellow eye fromeach animal was embedded whole in glycol methac-rylate so that changes in the entire globe could alsobe evaluated. These methods demonstrated thatvaso-occlusions occurred in retinal vessels, resultingin nonperfused areas of retina. Adjacent to these pre-
sumably ischemic areas, retinal neovascularizationwas observed. Neovascularization from the choroidalvasculature was also observed in whole eyes, in ad-dition to pigmented chorioretinal lesions.
Materials and Methods
Transgenic mice of the a HIS[1MDD] type,6 ranging inage from 4 to 30 months old, were sacrificed by cer-
vical dislocation. Institutional guidelines regardinganimal experimentation were followed. Mice from thisline not expressing the transgenes, as determined byhigh performance liquid chromatography analysis,6were used as controls. One eye from each animal was
processed by the technique of Lutty and McLeod.9Sensory retina was dissected from the eye at the levelof the photoreceptor outer segments, fixed in 2%paraformaldehyde in 0.1 mol/L cacodylate, pH 7.4,and incubated for enzyme histochemical demonstra-tion of ADPase activity. The retinas were then flattenedand embedded in glycol methacrylate (JB-4 kit, Poly-sciences, Warrington, PA). Lead phosphate ADPasereaction product was viewed and photographed enb/ocwith darkfield illumination, white reaction productin blood vessels was seen against a black back-ground. Pigment in retina was visualized and photo-graphed en bloc using brightfield illumination.Two-p-thick serial sections were then cut from pho-tographically documented areas in the retina usingdry glass knives on a Sorval MT2-B ultramicrotome(Norwalk, CN). ADPase reaction product was devel-oped in some sections with ammonium sulfide, re-sulting in a black reaction product. These sectionswere counterstained with thionin. Other sections werestained with toluidine blue-basic fuchsin (Multiplestain, Polysciences), McManus' periodic acid-Schiffwith hematoxylin counterstain, or hematoxylin andeosin.The fellow eye from each animal was embedded
whole in glycol methacrylate for analysis of all ocularstructures. Eyes were fixed whole in 1/4 strength Kar-novsky's paraformaldehyde/glutaraldehyde fixativeand dehydrated, infiltrated, and embedded in JB-4solution as recommended by the manufacturer.Blocks were cut and sections stained as describedabove.
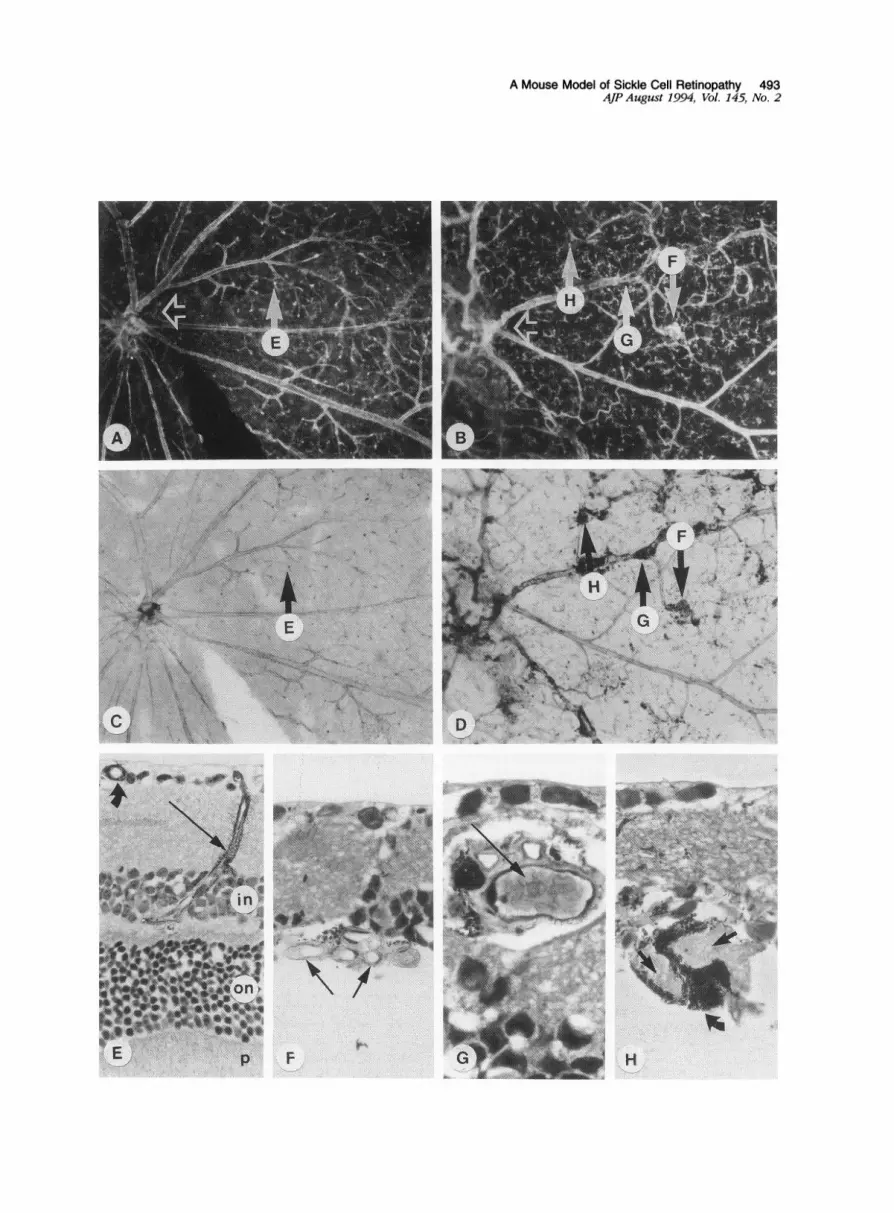
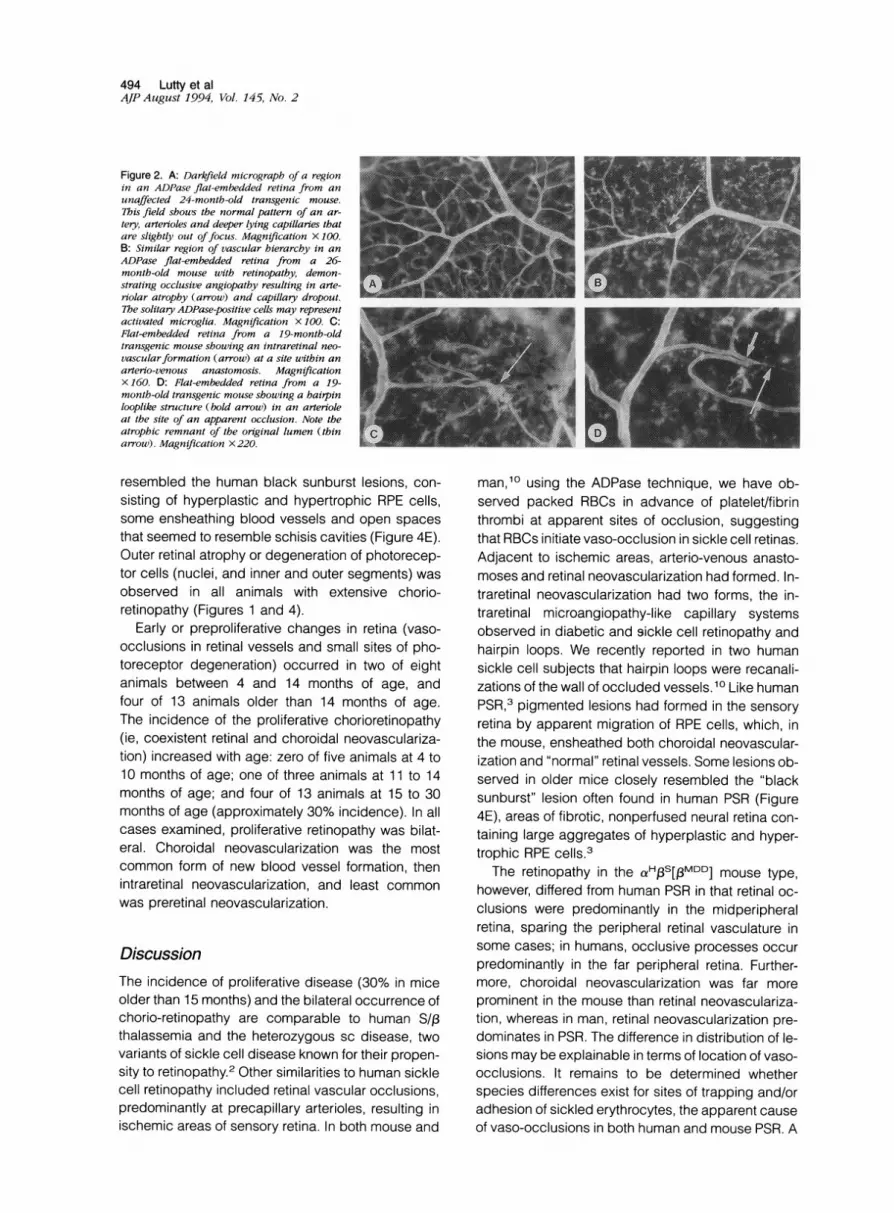
ResultsIn four of five mice younger than 10 months old andall control animals (not expressing transgenes), theretinal vasculature appeared normal. The primaryvascular system, viewed en bloc in the ADPase flat-embedded retinas, had pairs of arteries and veinsextending radially from the optic nerve head (Figure1A). The precapillary arterioles traversed the innerretina to supply the capillaries in the deep vascularnetwork within the inner nuclear layer (Figures 1 E and2A) and venules returned to the nerve fiber layer. Be-cause our dissections were at the level of the pho-toreceptor outer segments, there were no retinal pig-ment epithelial cells in the normal sensory retina,when the retinas were viewed with transmitted orbright field illumination (Figure 1C).The initial vascular changes observed in the
ADPase-reacted retinas of mice expressing the trans-genes were occlusions (indicated by loss of ADPase

492 Lutty et alAJP August 1994, Vol. 145, No. 2
activity), most often in retinal precapillary arterioles(Figure 2B) but also, in more advanced stages of dis-ease, in veins (Figure 1, B and G). At some sites ofocclusion, packed red blood cells (RBCs) were ob-served in advance of platelet/fibrin thrombi (Figure1G). The occlusive processes resulted in nonper-
fused areas of retina (loss of capillaries in the innernuclear layer) (Figure 2B) and in formation of arterio-venous anastomoses (Figures 1B and 2C). SolitaryADPase-positive cells with a ramified morphologywere observed in nonperfused areas of retina (Figure2, B to D). We have previously observed similar cellsin nonperfused areas of diabetic retinas and assume
they represent activated microglia.9 In whole eyes,
other early changes were observed in both retina andchoroid. Small retinal detachments were observed inthree mice. Small sites of photoreceptor degenerationwere observed in areas that appeared to lack patentchoriocapillaris lumens or the lumens were engorgedwith RBCs (Figure 3, B to D). The earliest form of thischange was reduced numbers of photoreceptor cellnuclei in the outer nuclear layer (Figure 3, B to C).
Proliferative changes were observed in more
advanced stages of retinopathy and consisted ofintraretinal and, in some cases, preretinal neovas-
cularization (Figure 4B). The intraretinal neovas-
cularization demonstrated elevated ADPase activityand was most often located at sites within arterio-venous anastomoses (Figure 2C). Some formationswere corkscrewlike systems of capillaries (Figures 1 Fand 2C), similar to the intraretinal microangiopathyobserved in both human diabetic and sickle cell reti-nopathy, whereas other formations had the appear-ance of hairpin loops (Figure 2D), which are common
in human sickle cell retinas.10 Preretinal neovascu-
larization, new blood vessels that extended fromretina into the normally avascular vitreous cavity (Fig-ure 4B), consisted of one or more large lumenal di-ameter vessels and was far less prevalent than in-traretinal neovascularization. In animals with severe
proliferative retinopathy, the entire outer nuclear layer
(photoreceptor nuclei) was degenerative, resulting inouter retinal atrophy (Figure 4, C to E). This was ac-companied by a paucity of choriocapillaris lumens,and the lumens observed were often engorged withRBCs. The choroids from these eyes also exhibited anincrease in number and size of large round pig-mented cells that were similar to the clump cells ofKoganei.11 In humans, these cells are prominent in irisand ciliary body and are believed to be resident mac-rophages (histiocytes), which scavenge shed pig-ment from melanocytes.11 In mice expressing thetransgenes, the number and size of clump cells wasincreased throughout the uveal tract, which includesiris, ciliary body, and choroid (Figure 4, C and E).When diseased flat-embedded retinas were
viewed en bloc with brightfield illumination, pig-mented areas were observed within sensory retina(Figure 1 D), which is normally devoid of pigmentedcells (Figure 1C). In addition to discrete pigmentedlesions, the pigmented cells were often intimately as-
sociated with blood vessels (Figure 1D). It was ap-
parent in 2-p sections through these vessels that theperivascular pigment consisted of hyperplastic andhypertrophic retinal pigment epithelial (RPE) cells thathad migrated into the sensory retina and ensheathednearby blood vessels (Figure 1G). Many of the RPE-ensheathed vessels had unusually dilated lumensand coursed through the outer retina (Figure 1 H). Nor-mally, only the secondary vasculature or deep cap-
illary network and postcapillary venules are observedin outer retina (Figures 1 E and 4A). In sections takenfrom the whole fellow eye of each animal, some RPE-ensheathed vessels within the retina were apparentlyderived from the choroidal vasculature, ie, choroidalneovascularization (Figure 4D). In a few instances,the choroidal neovascularization seemed to anasto-mose with retinal vessels (chorio-retinal anastomosis)(Figure 4C). Many of the choroidal neovascularizationformations appeared infarcted because their lumenscontained amorphous material and a paucity of endo-thelial cells. The most extensive pigmented lesions
Figure 1. A: En bloc darkfield micrograph ofan ADPaseflat-embedded retina from a 12-month-old transgenic mouse without retinopathy, dem-onstrates a normal spoke-wheel pattern with arteries and veins extending radiallyfrom the optic nerve head (open arrow). Magnification X58. B:Similar region in an ADPasefiat-embedded retinafrom a 19-month-old transgenic mouse with retinopathy (open arrow = optic nerve head). Reti-nopatby is characterized by occlusions resulting in nonperfused regions of retina, atterio-venous anastomoses and neovascularization. Magnifi-cation X58. C: Brighfield view of the same region shown in (A) illustrates lack ofpigment in the retinalpreparationfrom unaffected mice. Mag-nification X58. D: Brightfield view of the flat-embedded retina shown in (B) demonstrates abnormal perivascular pigment accumulation inanimals with retinopathy. Magnification X58. E: Section collected through the region shown by arrow "E" in the flat views of the nondiseasedretina (A and C) illustrates the primary vasculature of the nerve fiber layer (curved arrow) and a precapillary arteiole (arrow) that traverses theinner retinal layers to supply the secondary capillary network at the outer margin of the inner nuclear layer (in). Th-e photoreceptor cell nuclei arelocated in the outer nuclear layer (on). Photoreceptor inner and outer segments (p) are also shown. ADPase reaction product developed with am-monium sulfide and thionin counterstain. Magnification X 430. F: Section collectedfrom the region indicated in theflat view (arrow "F" in B andD), shows intraretinal neovascularization consisting of a corkscrewlike formation of capillaries. Toluidine blue-basic fuchsin, magnificationx 680. G: Section collected through the site ofan apparent occlusion in a vein ensheathed with pigment (arrow "G" in theflat views). Packed sick-led erythrocytes within the lumen are shown (arrow). Toluidine blue-basic fuchsin, magnification x 1,100. H: Section collected through a pig-mented lesion (arrow "H" in the flat views) reveals cores offibrillar material (arrows) surrounded by hypertrophic retinalpigment epithelial cells(curved arrow). Toluidine blue-basic fuchsin, magnification X 680.
A Mouse Model of Sickle Cell Retinopathy 493AJP August 1994, Vol. 145, No. 2
ssJ'
* er 8 * >..
Ve...: * ,>. ^*.
*: 411¢' "* * !WrO^
, .,...; E< * L. tQ J '#5 g.''
$'Ntit*- #? .; * ,>
Wr s . >* ss .
.. s.
.s.. jwf .. , .is.*S ^N
.. :N *, s
r:.'
9ss g ,.
_g* *
¾ .
N,\
v~~~F-HW-: H
494 Lutty et alAJP August 1994, Vol. 145, No. 2
Figure 2. A: Darkfield micrograph of a regionin an ADPase flat-embedded retina from anunaffected 24-month-old transgenic mouse.7his field shouws the normal pattern of an ar-tery, arterioles and deeper lying capillaries thatare slightly out offocus. Magnification X 100.B: Similar region of vascular hierarchy in anADPase flat-embedded retina from a 26-month-old mouse with retinopathy, demon-strating occlusive angiopathy resulting in arte-riolar atrophy (arrow) and capillary dropout.The solitary ADPase-positive cells may representactivated microglia. Magnification X 100. C:Flat-embedded retina from a 19-month-oldtransgenic mouse showing an intraretinal neo-vascularformation (arrow) at a site within anarterio-venous anastomosis. MagnificationX 160. D: Flat-embedded retina from a 19-month-old transgenic mouse showing a hairpinlooplike structure (bold arrow) in an arterioleat the site of an apparent occlusion. Note theatrophic remnant of the original lumen (thinarrow). Magnification X220.
resembled the human black sunburst lesions, con-
sisting of hyperplastic and hypertrophic RPE cells,some ensheathing blood vessels and open spacesthat seemed to resemble schisis cavities (Figure 4E).Outer retinal atrophy or degeneration of photorecep-tor cells (nuclei, and inner and outer segments) was
observed in all animals with extensive chorio-retinopathy (Figures 1 and 4).
Early or preproliferative changes in retina (vaso-occlusions in retinal vessels and small sites of pho-toreceptor degeneration) occurred in two of eightanimals between 4 and 14 months of age, andfour of 13 animals older than 14 months of age.The incidence of the proliferative chorioretinopathy(ie, coexistent retinal and choroidal neovasculariza-tion) increased with age: zero of five animals at 4 to10 months of age; one of three animals at 11 to 14months of age; and four of 13 animals at 15 to 30months of age (approximately 30% incidence). In allcases examined, proliferative retinopathy was bilat-eral. Choroidal neovascularization was the mostcommon form of new blood vessel formation, thenintraretinal neovascularization, and least commonwas preretinal neovascularization.
DiscussionThe incidence of proliferative disease (30% in miceolder than 15 months) and the bilateral occurrence ofchorio-retinopathy are comparable to human S/Ifthalassemia and the heterozygous sc disease, twovariants of sickle cell disease known for their propen-sity to retinopathy.2 Other similarities to human sicklecell retinopathy included retinal vascular occlusions,predominantly at precapillary arterioles, resulting inischemic areas of sensory retina. In both mouse and
man,10 using the ADPase technique, we have ob-served packed RBCs in advance of platelet/fibrinthrombi at apparent sites of occlusion, suggestingthat RBCs initiate vaso-occlusion in sickle cell retinas.Adjacent to ischemic areas, arterio-venous anasto-moses and retinal neovascularization had formed. In-traretinal neovascularization had two forms, the in-traretinal microangiopathy-like capillary systemsobserved in diabetic and sickle cell retinopathy andhairpin loops. We recently reported in two humansickle cell subjects that hairpin loops were recanali-zations of the wall of occluded vessels. 10 Like humanPSR,3 pigmented lesions had formed in the sensoryretina by apparent migration of RPE cells, which, inthe mouse, ensheathed both choroidal neovascular-ization and "normal" retinal vessels. Some lesions ob-served in older mice closely resembled the "blacksunburst" lesion often found in human PSR (Figure4E), areas of fibrotic, nonperfused neural retina con-
taining large aggregates of hyperplastic and hyper-trophic RPE cells.3
The retinopathy in the a HfS[fMDD] mouse type,however, differed from human PSR in that retinal oc-
clusions were predominantly in the midperipheralretina, sparing the peripheral retinal vasculature insome cases; in humans, occlusive processes occur
predominantly in the far peripheral retina. Further-more, choroidal neovascularization was far more
prominent in the mouse than retinal neovasculariza-tion, whereas in man, retinal neovascularization pre-
dominates in PSR. The difference in distribution of le-sions may be explainable in terms of location of vaso-occlusions. It remains to be determined whetherspecies differences exist for sites of trapping and/oradhesion of sickled erythrocytes, the apparent causeof vaso-occlusions in both human and mouse PSR. A
I
I

A Mouse Model of Sickle Cell Retinopathy 495AJP August 1994, Vol. 145, No. 2
....... . ,,. *, . * t~~~~~~~~~~~~W/
*i* f .* :*%v
Figure 3. Comparison ofsections through outer retina and choroidfrom whole eyes ofa 6-month-old control mouse (A) andfrom transgenic miceexpressing 72% Ps (B, a 34-montb-old; C and D a 24-month-old). A: In control mice, 10 to 12 rows ofphotoreceptor nuclei were typically observedin the otuter nuclear layer (n). Posterior to the outer nuclear layer are the photoreceptor inner and outer segments (S) and the pigment epithelium(curved arrow), the outermost cell type in retina that is eliminatedfrom ADPaseflat-embedded preparations. The normal choroid consists of cho-riocapillaris (arrows), large choroidal vessels, and stromal melanocytes (pigmented cells). Toluidine blue-basicfuchsin, magnification X 400. B: Ina 34-month-old transgenic mice that did not demonstrate proliferative retinopathy, diffuse photoreceptor degeneration was observed. The outernuclear (n) layer is reduced to several layerm of nuclei, the outer segments are short (S), and some retinalpigment epithelial cells appear hypertro-phic (curved arrowhead). The choroid underlying this region demonstrates packed RBCs uwithin engorged choriocapillaris (arrow) and an in-crease in pigmented cells. Other areas of this retina had the normal number ofphotoreceptor nuclei and length of outer segments. Toluidine blue-basic fuchsin, magnification X 400. C: Similar field from a preproliferative transgenic mouse demonstrates a focal area of photoreceptordegeneration (loss of nuclei in the outer nuclear layer) (open arrow) associated with vacuolated RPE cells (curved arrow). The red cell aggregatewithin the underlying choriocapillaris (arrow) may represent a choroidal occlusion. Hematoxylin and eosin, magnification X 400. D: Higher mag-nification micrograph of an adjacent section to the one shown in (C) demonstrates sickled erytbrocytes within the choriocapillaris lumen (thinarrow) and normal erytbrocytes in large choroidal vessels (bold arrouw). Hematoxylin and eosin, magnification X 1,800.
third difference between the mouse model and hu-man sickle cell retinopathy is that the preretinal neo-vascular formations observed in human PSR areelaborate fronds of blood vessels called "sea fans",2,3whereas mouse preretinal neovascular formationsconsisted of only a few large vessels. Finally, retinalhemorrhages were rarely observed in the mice,whereas in human PSR, retinal hemorrhages are con-sidered a major component.2'3
Although there was degeneration of outer retina inanimals with severe chorioretinopathy, we do not be-lieve that the complete loss of photoreceptors is dueto a retinal degeneration process invoked perhaps byinsertion of the transgenes. Loss of photoreceptorshas been observed during murine retinal degenera-tions, but, even in the slow onset retinal degenerationmodels (rds, pcd, and nr), the degenerative processis complete (loss of photoreceptor nuclei, and innerand outer segments) by age 6 to 12 months.12 Theproliferative retinopathy with total loss of photorecep-tors we report was primarily observed at 14 months orolder. In rodent retinal degenerations, all animalscompletely lose their photoreceptors in a very definitechronological progression. The retinopathy we ob-served occurred in only 30% of the animals older than
14 months of age and older transgenic mice withoutretinopathy had histologically viable photoreceptors.Furthermore, focal areas of photoreceptor degenera-tion were observed in eyes at the preproliferativestage, where the remainder of their photoreceptorlayer appeared normal (Figure 3).We propose that photoreceptor degeneration in
our mouse line is caused by choriocapillaris occlu-sion, either permanent or transient, and subsequentischemia that occurred in the course of the disease.This vasculature is the source of oxygen and nutrientsfor the high metabolic demands of photoreceptors,and loss of patency in choriocapillaris might causesuch a wholesale death of the photoreceptors. Fouldsand Lee13 have demonstrated that choroidal ische-mia in human subjects results in degeneration of pho-toreceptor elements and outer retinal atrophy. Thepredominance of choroidal neovascularization in themouse model suggests that major vaso-occlusiveevents have occurred in choroids of affected animals.In a preliminary study using horse radish peroxidaseas a vascular tracer, we have observed small areas ofphotoreceptor degeneration associated with appar-ently nonperfused areas of choriocapillaris, ie, areaslacking horse radish peroxidase in choriocapillaris lu-
ik -

496 Lutty et alAJP Augtust 1994, Vol. 145, No. 2
P
w}~~~ ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.........- t sLlilt!;.....tE>jv
w>''X'<i>!S~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~......
Figure 4. A: Cross-section ofa whole eyefrom a 12-month-old control mouse, illustrating the appearance ofnormal retina and choroid. There aretwo nuclear layers in sensory retina, inner (in) and outer (on), and photoreceptor inner (is) and outer segments (os). Toluidine blue-basic fuch-sin, magnification X340. B: Preretinal neovascularization in an 18-month-old transgenic mouse near the optic nerve. 7Te lumens (arrows) of thenew vessels are relatively large and are partially surrounded by RPE-like cells (curved arrows). Note the absence of nuclear layers andphotorecep-tors. Toluidine blue-basic fuchsin, magnification X510. C: In an 18-month-old transgenic mouse, a choroidal vessel traverses retina and anasto-moses with a retinal vein (a chorio-retinal anastomosis [arrows!). Only the nervefiber and inner nuclear layers of retina are present, ie, the pho-toreceptor cells are absent. Toluidine blue-basic fuchsin, magnification X510. D: Choroidal neovascularization (arrow) traverses a discontinuousRPE cell layer (curved arrow) and invades sensory retina in a 20-month-old transgenic mouse. 7Te slew vascular lumens in sensory retina areensheathed with RPE cells (arrowheads). There are no photoreceptors present. Toluidine blue-basic fuchsin, magnification X 610. E: The apparentmigration of RPE cells into sensory retina in this 24-month-old transgenic mouse has created a chorio-retinal lesion that resembles the "black sun-burst" lesion observed in human PSR. The lesion contains schisislike cavities (curved arrow) and several large vascular lumens (arrow) that areensheathed with RPE cells and resemble the channels shown in Figure 1 H. The choroid in mice with retinopathy was quite thick and heavily pig-mented compared to unaffected transgenic and normal animals (Figure 4A). The increase in pigmentation was at least partially due to an in-crease in ntumber and size ofpigmented clump cells (open white arrow). Hematoxylin and eosin, magnification X330.
mens (unpublished data). The increase in size andnumber of clump cells suggests that some pathologi-cal process has occurred throughout the uveal tract,which includes the choriocapillaris and large choroi-dal vessels. In human iris and ciliary body, the num-ber of clump cells increases during pathologicalprocesses.11 In human specimens examined re-cently,10'14 we have observed outer retinal atrophy,vaso-occlusions of the choriocapillaris, and choroidalneovascularization. Furthermore, using our recentlydeveloped alkaline phosphatase flat-embedded cho-roid technique15 we have observed areas of nonper-fused choriocapillaris in 20- and 28-year-old sicklecell subjects (unpublished data).
The histopathological results presented clearlydemonstrate similarities in the ocular effects of sicklecell disease in the a HfS[EMDD] transgenic mousetype and human. Anatomical variations in the vascu-latures of mammalian species may account for thepropensity for choroidal involvement in the mouse.The few differences between human PSR and our
mouse PSR model do not detract from its suitability asa model for human sickle cell retinopathy and, per-haps, as an animal model for evaluating the effects ofantisickling therapies on the incidence and severity ofsickle cell retinopathy. Furthermore, this is the first ge-netically derived animal model for choroidal and reti-nal neovascularization. As such, it should be a usefulmodel for the evaluation of antiangiogenic sub-stances for their potential use in controlling both cho-roidal and retinal neovascularization, which occurs inage-related macular degeneration and diabetic andsickle cell retinopathy respectively.
Acknowledgments
The authors acknowledge the excellent technical as-sistance of Carol Merges, Sandra Suzuka, SylviaCrone, and Mellisa Katz. We also acknowledge Drs.D. Zack and S. C. Selden for their critical evaluationof this manuscript.
-7WAMAMIIW --:

A Mouse Model of Sickle Cell Retinopathy 497AJP August 1994, Vol. 145, No. 2
References
1. Kaul DK, Fabry ME, Nagel RL: Erythrocytic and vascu-lar factors influencing the microcirculatory behavior ofblood in sickle cell anemia. Ann NY Acad Sci 1989,565:316-326
2. Goldberg MF: Retinal vaso-occlusion in sickling hemo-globinopathies. Birth Defects 1976, 12:475-515
3. Romayananda N, Goldberg MF, Green WR: Histopa-thology of sickle cell retinopathy. Trans Am Acad Oph-thalmol Otolaryngol 1973, 77:652-676
4. Trudel M, Saadane N, Garel M-C, Bardakdjian-MichauJ, Blouquit Y, Guerquin-Kern J-L, Rouyer-Fessard P,Vivaud D, Pachnis A, Romeo P-H, Beuzard Y, Costan-tini F: Towards a transgenic mouse model of sickle celldisease: hemoglobin SAD. EMBO J 1991, 10:3157-3165
5. Ryan E, Townes TM, Reilly MP, Asakura T, Palmiter RD,Brinster RL, Behringer RR: Human sickle hemoglobinin transgenic mice. Science 1990, 247:566-568
6. Fabry ME, Nagel RL, Pachnis A, Suzuka SM, Constan-tini F: High expression of human Ps and a-globins intransgenic mice: hemoglobin composition and hema-tological consequences. Proc Natl Acad Sci USA1992, 89:12150-12154
7. Fabry ME, Costantini F, Pachnis A, Suzuki SM, BankN, Aynedjian HS, Factor SM, Nagel RL.: High expres-sion of human fs- and a-genes in transgenic mice:
erythrocyte abnormalities, organ damage, and the ef-fect of hypoxia. Proc Natl Acad Sci USA 1992, 89:12155-12159
8. Skow LC, Burkhart BA, Johnson FM, Popp RA, PoppDM, Goldberg SZ, Anderson WF, Barnett LB, LewisSE: A mouse model for ,B-thalassemia. Cell 1983, 34:1043-1052
9. Lutty GA, McLeod DS: A new technique for visualiza-tion of the human retinal vasculature. Arch Ophthalmol1992, 110:267-276
10. McLeod D, Goldberg M, Lutty G: Dual perspectiveanalysis of vascular formations in sickle cell retinopa-thy. Arch Ophthalmol 1993, 111:1234-1245
11. Wobmann P, Fine B: The clump cells of Koganei: alight and electron microscopic study. Am J Ophthal-mol 1972, 73:90-101
12. LaVail MM: Analysis of neurological mutants with in-herited retinal degeneration. Invest Ophthalmol Vis Sci1981, 21:638-657
13. Foulds W, Lee W: Clinical and pathological aspects ofchoroidal ischaemia. Trans Ophthalmol Soc UK 1971,91:323-341
14. Lutty G, Merges C, Crone S, McLeod D: Immunohisto-chemical insights into sickle cell retinopathy. Curr EyeRes 1994, 13:125-138
15. McLeod DS, Lutty GL: High resolution histologicalanalysis of the human choroidal vasculature. InvestOphthalmol Vis Sci 1994 (in press)