regulation of hemolymph osmolytes and gill na+/k+-atpase activities during acclimation to saline...
TRANSCRIPT

LJournal of Experimental Marine Biology and Ecology,215 (1997) 81–91
1 1Regulation of hemolymph osmolytes and gill Na /K -ATPase activities during acclimation to saline media in the
freshwater shrimp Macrobrachium olfersii (Wiegmann, 1836)(Decapoda, Palaemonidae)
b a , c*Alice Gon¢calves Lima , John C. McNamara , Walter Ribeiro Terraa ˜ ˜ ˜Departamento de Biologia, FFCLRP, Universidade de Sao Paulo, Ribeirao Preto, 14040-901 Sao Paulo,
Brazilb ˆ ˜Departamento de Fisiologia, Instituto de Biociencias, USP, Sao Paulo, Brazil
c ´ ´ ˜Departamento de Bioquımica, Instituto de Quımica, USP, Sao Paulo, Brazil
Received 6 May 1996; revised 1 November 1996; accepted 16 December 1996
Abstract
To evaluate the mechanisms of long-term osmotic adaptation to saline media, total hemolymph os-1 1molytes, hemolymph sodium and chloride concentrations, and gill Na /K -ATPase activities were
measured in the freshwater shrimp Macrobrachium olfersii (Wiegman) after acclimation for 10 or 20days to media of , 0.5, 21 and 28‰ salinity. Total hemolymph osmolytes are maintained strongly hy-perosmotic to the external medium in low and moderate salinities, becoming slightly hyperosmotic at
1 2high salinity. In contrast, however, hemolymph [Na ] and [Cl ] are distinctly hyporegulated at thehigher salinities. This difference in response pattern to acclimation apparently results from the pres-ence of free amino acids in the hemolymph as a consequence of the synthesis of intracellular organicosmolytes. The resulting hyperosmotic regulation both avoids water loss from the animal in salinemedia and provides an osmotic gradient, allowing the uptake of water from the medium for excretion
1 1of the salt load. Gill Na /K -ATPase activities decrease by ¯ 35% after acclimation to saline media,suggesting that the rates of the cellular mechanisms responsible for salt uptake in freshwater become
1reduced, avoiding excessive salt loading. A Na -ATPase activity is salinity independent. These dataare examined with regard to the intrinsic coupling between the physiological alterations taking placein hemolymph osmolytes during acclimation to saline media and the concomitant ultrastructural re-arrangements in the salt transporting tissues of the gills, and their possible neurosecretory controlmechanisms. 1997 Elsevier Science B.V.
1 1Keywords: Salinity acclimation; Gill Na /K -ATPase; Osmoregulation; Hemolymph osmolytes;Macrobrachium
*Corresponding author.
0022-0981/97/$17.00 1997 Elsevier Science B.V. All rights reservedPII S0022-0981( 97 )00016-6

82 A.G. Lima et al. / J. Exp. Mar. Biol. Ecol. 215 (1997) 81 –91
1. Introduction
Freshwater decapods, particularly those of the palaemonid genus Macrobrachium,display varying degrees of physiological adaption to their limnetic habitat. Some species,like M. olfersii and M. acanthurus, are entirely dependent on saline water forreproduction and metamorphosis, exhibiting a long larval developmental sequence;others, like M. potiuna and M. iheringi, are hololimnetic and exhibit salinity-in-dependent reproductive strategies with a very reduced number of zoeal stages (seeMoreira et al., 1983, for a discussion). These life histories reflect the varying capacitiesfor osmotic adaptation that have evolved during the differential penetration of thissuccessful palaemonid group into the freshwater habitat, and disclose differing degreesof independence from the marine biotope.
Osmoregulatory ability, the primary physiological means by which these hyper-regulators maintain the osmolality and ionic concentrations of their intra- and extracellu-lar fluids fairly constant also differs widely among the different species of Macrobrach-ium although all are strong hyperosmotic regulators in freshwater, their usual medium(Moreira et al., 1983; Read, 1984). Several species do show some differentiation interms of their hyporegulatory osmotic and ionic capability in saline media; however, theshort-term response to strong saline gradients is at best time-dependent, moderatehyporegulation (e.g., Denne, 1968; Armstrong et al., 1981; Castille and Lawrence, 1981;Read, 1984; McNamara, 1987).
Macrobrachium olfersii (Wiegman) is a euryhaline species common to the freshwaterstreams of the Atlantic coast of southern Brazil and is physiologically representative ofthe genus. In this strong hyper-regulator (McNamara, 1987), many of the processesinvolved in anisosmotic, extracellular osmo- and ion regulation in response to short-termsalinity exposure are under the control of neurosecretory factors released fromneurosecretory cells located in the principal ganglia of the central nervous system(McNamara et al., 1990, 1991; McNamara, 1993; Santos and McNamara, 1996). Theregulation of hemolymph free amino acid concentrations that occurs during intracellularisosmotic regulation also appears to be similarly modulated (Freire et al., 1995).
The principal effector of ion uptake in M. olfersii, and in various species of21 1 1Macrobrachium, is the Mg -dependent, Na /K -ATPase (see Stern et al., 1984, for
M. rosenbergii and Proverbio et al., 1990 for M. amazonicum), an ion transportingenzyme apparently located in the numerous membrane invaginations of the mito-chondria-rich, intralamellar, septal cells in the gill lamellae (Freire and McNamara,
11995; McNamara and Lima, 1997). This enzyme actively transports Na , exchanged1 1across the apical membranes of the gill pillar cells by a Na /NH counter transporter4
´(see Armstrong et al., 1981; Pequeux, 1995) from the external medium, up aconcentration gradient to the hemolymph, and is responsible for the strong hyper-regulation of the hemolymph in freshwater.
The underlying physiological responses of the above osmoregulatory systems tolong-term high salinity acclimation are not well known in Macrobrachium or in anyother freshwater palaemonid. Essentially, during acclimation, hemolymph proteinconcentration decreases while intracellular (muscle) free amino acid concentrationsincrease in both a time- (Armstrong et al., 1981) and salinity-dependent manner (Tan

A.G. Lima et al. / J. Exp. Mar. Biol. Ecol. 215 (1997) 81 –91 83
and Choong, 1981). However, little information is available concerning alterations in theinorganic and organic osmolytes of the hemolymph after salinity acclimation. Consider-ing that this fluid constitutes one of the primary barriers against alterations inintracellular osmolality and changes in cell volume, modifications in the concentrationsof the principal hemolymph osmolytes during salinity acclimation warrant investigation.
1 1Increases in the activity of the Na /K -ATPase transport system have been reportedin response to short-term exposure to dilute media in several brachyuran species(Kamemoto, 1991), apparently regulated by neurohormonal mediation. However,virtually nothing is known of the long-term changes in ATPase activity that may occurin response to high salinity acclimation in freshwater hyper-regulators where the rate offunction of inwardly directed systems of salt capture must necessarily decrease.
To address this lack of data on the consequences of long-term salinity acclimation infreshwater hyperegulators, the present study examines the alterations induced in the
1 1concentrations of the principal hemolymph osmolytes, and in gill Na /K -ATPaseactivity, in the freshwater shrimp Macrobrachium olfersii.
2. Material and methods
Intermolt, female specimens of Macrobrachium olfersii, of 4–6 cm total length, were´collected from the marginal vegetation of the Pauba River (238 479 S; 458 339 W) in the
˜State of Sao Paulo, Brazil. The shrimps were maintained in tanks containing 100 l ofwater from the collection site ( , 0.5‰) and were fed fish, meat or carrot three times aweek.
Three groups of 24 shrimps each were acclimated for a 10 day period in 100 l ofmedium of either 0 ( , 0.5‰), 21 or 28‰ salinity. After this exposure period, individualhemolymph samples were obtained by puncturing the pericardial sinus with a [25-8needle. Approximately 70 ml of hemolymph was drawn into a 1 ml plastic syringe,transferred to a 1.5 ml Eppendorf tube and frozen at 2 208C.
Hemolymph osmolality was measured using a Wescor model 5500 vapour pressureosmometer in 10 ml aliquots. Sodium concentration was estimated by emissionspectrophotometry with a Zeiss PMQII flame spectrophotometer, and chloride con-centration by titration with mercuric nitrate using a Beckman/Spinco microtitrator,employing s-diphenylcarbazone as the indicator.
1 1 1Gill Na /K -ATPase and Na -ATPase activities were measured in 3 groups ofshrimps (17 , N , 30) acclimated for 20 days to salinities of 0 ( , 0.5‰), 21 or 28‰.Each group was subdivided to provide at least 5 replicate experiments.
After being killed by cooling, the 5 large, posterior gill pairs were removed from eachshrimp and were homogenized in a solution containing (in mM) imidazole–HCl 20,sucrose 250, EDTA 6 and sodium deoxycholate, 2.4, at pH 6.8. The crude homogenatewas centrifuged at 9500 rpm (Sorvall, rotor SS34) for 35 min, the supernatantultracentrifuged at 35 000 rpm (Hitachi Ultracentrifuge, rotor 50.2) for a further 60 min,and the precipitate was suspended in 1 ml of homogenate solution.
Three, replicate, 20 ml aliquots of the mitochondria-free, vesicle preparations from

84 A.G. Lima et al. / J. Exp. Mar. Biol. Ecol. 215 (1997) 81 –91
each of the homogenates for each acclimation group were incubated for 30 min at 308Cin 1 ml of a medium containing (in mM) imidazole–HCl 20, MgCl 50, tris-ATP 2 and2
1 1the requisite combinations of Na 100 and K 30, or choline 100 (as chlorides) andouabain 2, according to the particular medium required. The reaction was stopped byimmersion in boiling water, and inorganic phosphate (P ) was measured in 100 mli
aliquots of reaction medium, using Baginski and Zak’s (Baginski and Zak, 1960)1 1technique. The Na /K -ATPase activity was calculated as the difference in P liberatedi
in the complete incubation medium and that in this medium plus ouabain, a specific1 1 1inhibitor of Na /K -ATPase activity. The Na -ATPase activity was calculated as the
1 1 1difference in P liberated in K -free medium and that in K - and Na -free medium.i
Protein content was measured using Bradford’s (Bradford, 1976) Coomassie BrilliantBlue technique.
1 2All data (osmolality, [Na ], [Cl ], ATPase activities) were verified for normality ofdistribution (Kolmogorov-Smirnov test) and a one-way analysis of variance was thenperformed to detect the effect of acclimation salinity. When present, differences betweenmeans were located using the Student-Newman-Keuls test. All tests were performedusing a minimum significance level of P 5 0.05. Data are presented in the text as themean6SEM (n).
The data for hemolymph osmolytes were adjusted to second-degree polynomialequations using the curve-adjusting function of SlideWrite 5 1 (Advanced GraphicsSoftware).
3. Results
3.1. Hemolymph osmolytes
After a 10 day acclimation period, Macrobrachium olfersii maintains the osmolalityof its hemolymph well above that of the medium in low and moderate salinities,becoming slightly hyperosmotic in a high salinity medium (Fig. 1). Osmolality increasessignificantly in animals exposed to 21 (7376222.2 mOsm H O/kg, n 5 9) and 28‰2
salinity (8686195.3, n 5 14) compared to those kept in freshwater (FW) (340680.7,n 5 6).
However, the concentrations of the two principal inorganic hemolymph osmolytes,1 2 1Na and Cl , do not follow this response pattern. While Na concentrations increase
significantly both in 21 (258622.3 mEq/ l, n 5 19) and 28‰ salinity (330620, n 5 14)compared to FW-acclimated shrimps (154622.8, n 5 12) (Fig. 2), they are moderately
1hyporegulated below the respective Na concentrations of these two media. The1 1 2iso-ionic point for Na was calculated to be 216 mEq Na / l. Hemolymph Cl
concentrations also increase significantly in both 21 (266620 mEq/ l, n 5 15) and 28‰salinity (358622.2, n 5 13) compared to FW-exposed shrimps (170642.1, n 5 13) (Fig.
23) but are strongly hyporegulated below the respective Cl concentrations in these2 2media. The iso-ionic point for Cl was 192 mEq Cl / l.
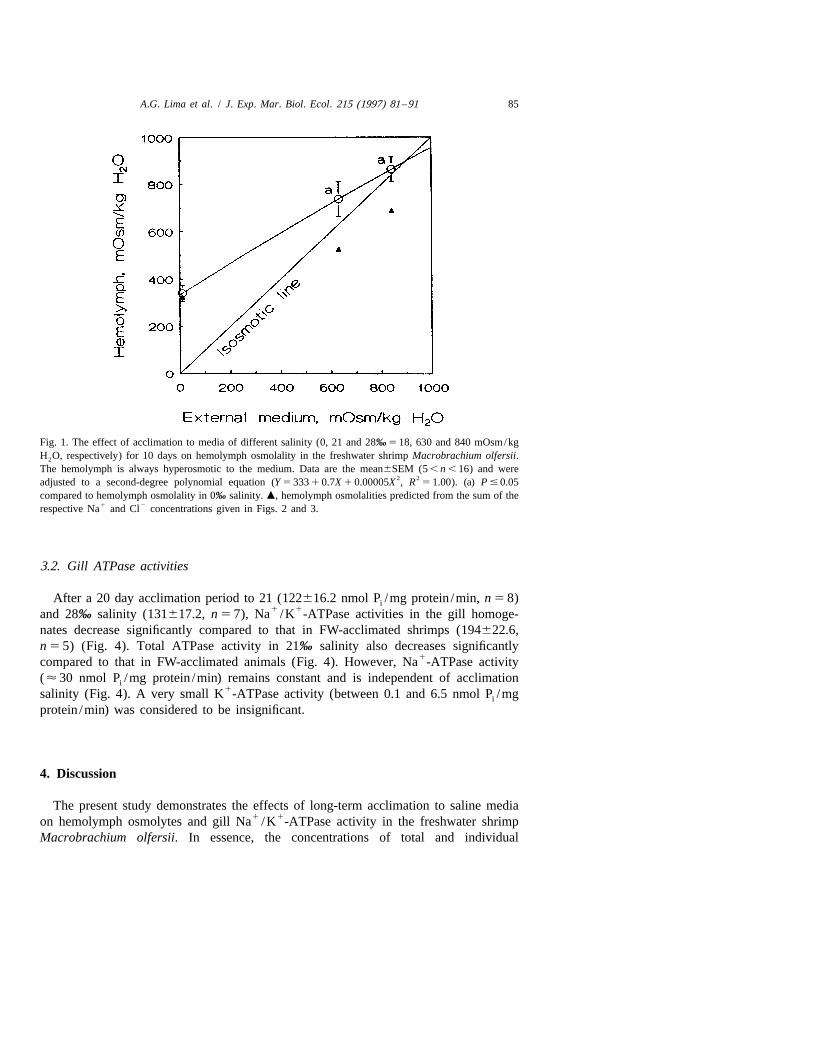
A.G. Lima et al. / J. Exp. Mar. Biol. Ecol. 215 (1997) 81 –91 85
Fig. 1. The effect of acclimation to media of different salinity (0, 21 and 28‰ 5 18, 630 and 840 mOsm/kgH O, respectively) for 10 days on hemolymph osmolality in the freshwater shrimp Macrobrachium olfersii.2
The hemolymph is always hyperosmotic to the medium. Data are the mean6SEM (5 , n , 16) and were2 2adjusted to a second-degree polynomial equation (Y 5 333 1 0.7X 1 0.00005X , R 5 1.00). (a) P # 0.05
compared to hemolymph osmolality in 0‰ salinity. m, hemolymph osmolalities predicted from the sum of the1 2respective Na and Cl concentrations given in Figs. 2 and 3.
3.2. Gill ATPase activities
After a 20 day acclimation period to 21 (122616.2 nmol P /mg protein /min, n 5 8)i1 1and 28‰ salinity (131617.2, n 5 7), Na /K -ATPase activities in the gill homoge-
nates decrease significantly compared to that in FW-acclimated shrimps (194622.6,n 5 5) (Fig. 4). Total ATPase activity in 21‰ salinity also decreases significantly
1compared to that in FW-acclimated animals (Fig. 4). However, Na -ATPase activity( ¯ 30 nmol P /mg protein /min) remains constant and is independent of acclimationi
1salinity (Fig. 4). A very small K -ATPase activity (between 0.1 and 6.5 nmol P /mgi
protein /min) was considered to be insignificant.
4. Discussion
The present study demonstrates the effects of long-term acclimation to saline media1 1on hemolymph osmolytes and gill Na /K -ATPase activity in the freshwater shrimp
Macrobrachium olfersii. In essence, the concentrations of total and individual
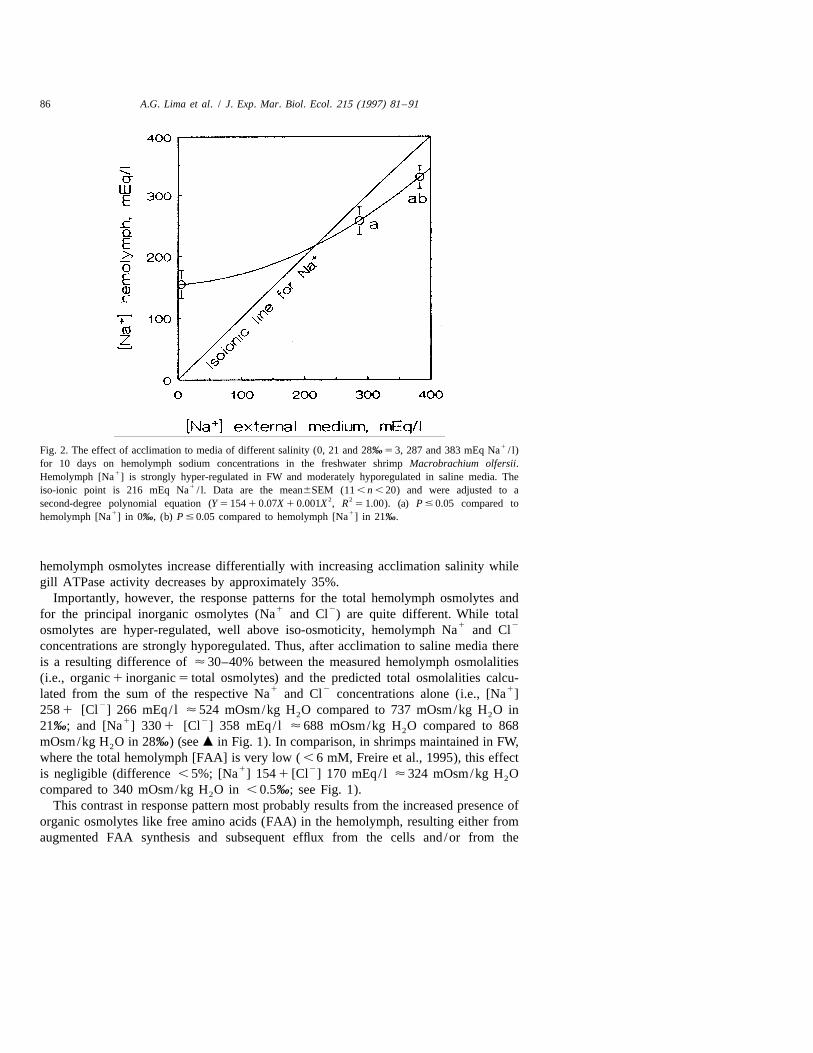
86 A.G. Lima et al. / J. Exp. Mar. Biol. Ecol. 215 (1997) 81 –91
1Fig. 2. The effect of acclimation to media of different salinity (0, 21 and 28‰ 5 3, 287 and 383 mEq Na / l)for 10 days on hemolymph sodium concentrations in the freshwater shrimp Macrobrachium olfersii.
1Hemolymph [Na ] is strongly hyper-regulated in FW and moderately hyporegulated in saline media. The1iso-ionic point is 216 mEq Na / l. Data are the mean6SEM (11 , n , 20) and were adjusted to a
2 2second-degree polynomial equation (Y 5 154 1 0.07X 1 0.001X , R 5 1.00). (a) P # 0.05 compared to1 1hemolymph [Na ] in 0‰, (b) P # 0.05 compared to hemolymph [Na ] in 21‰.
hemolymph osmolytes increase differentially with increasing acclimation salinity whilegill ATPase activity decreases by approximately 35%.
Importantly, however, the response patterns for the total hemolymph osmolytes and1 2for the principal inorganic osmolytes (Na and Cl ) are quite different. While total
1 2osmolytes are hyper-regulated, well above iso-osmoticity, hemolymph Na and Clconcentrations are strongly hyporegulated. Thus, after acclimation to saline media thereis a resulting difference of ¯ 30–40% between the measured hemolymph osmolalities(i.e., organic 1 inorganic 5 total osmolytes) and the predicted total osmolalities calcu-
1 2 1lated from the sum of the respective Na and Cl concentrations alone (i.e., [Na ]2258 1 [Cl ] 266 mEq/ l ¯ 524 mOsm/kg H O compared to 737 mOsm/kg H O in2 2
1 221‰; and [Na ] 330 1 [Cl ] 358 mEq/ l ¯ 688 mOsm/kg H O compared to 8682
mOsm/kg H O in 28‰) (see m in Fig. 1). In comparison, in shrimps maintained in FW,2
where the total hemolymph [FAA] is very low ( , 6 mM, Freire et al., 1995), this effect1 2is negligible (difference , 5%; [Na ] 154 1 [Cl ] 170 mEq/ l ¯ 324 mOsm/kg H O2
compared to 340 mOsm/kg H O in , 0.5‰; see Fig. 1).2
This contrast in response pattern most probably results from the increased presence oforganic osmolytes like free amino acids (FAA) in the hemolymph, resulting either fromaugmented FAA synthesis and subsequent efflux from the cells and/or from the
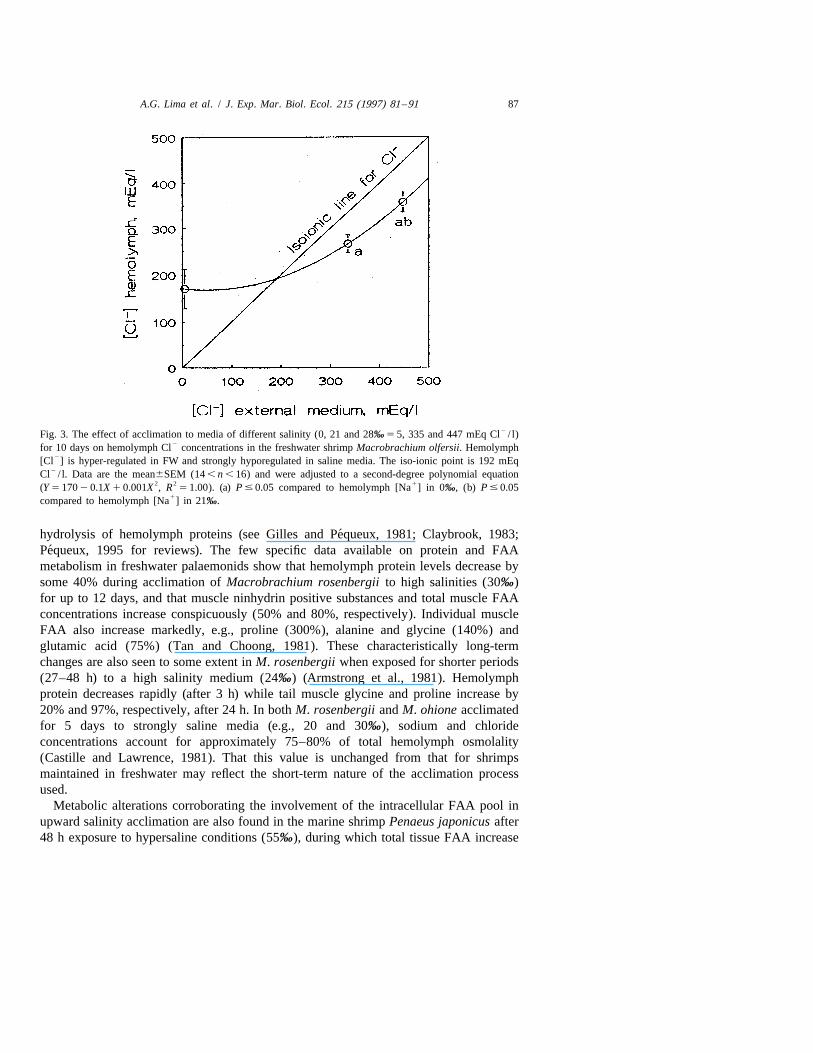
A.G. Lima et al. / J. Exp. Mar. Biol. Ecol. 215 (1997) 81 –91 87
2Fig. 3. The effect of acclimation to media of different salinity (0, 21 and 28‰ 5 5, 335 and 447 mEq Cl / l)2for 10 days on hemolymph Cl concentrations in the freshwater shrimp Macrobrachium olfersii. Hemolymph
2[Cl ] is hyper-regulated in FW and strongly hyporegulated in saline media. The iso-ionic point is 192 mEq2Cl / l. Data are the mean6SEM (14 , n , 16) and were adjusted to a second-degree polynomial equation
2 2 1(Y 5 170 2 0.1X 1 0.001X , R 5 1.00). (a) P # 0.05 compared to hemolymph [Na ] in 0‰, (b) P # 0.051compared to hemolymph [Na ] in 21‰.
´hydrolysis of hemolymph proteins (see Gilles and Pequeux, 1981; Claybrook, 1983;´Pequeux, 1995 for reviews). The few specific data available on protein and FAA
metabolism in freshwater palaemonids show that hemolymph protein levels decrease bysome 40% during acclimation of Macrobrachium rosenbergii to high salinities (30‰)for up to 12 days, and that muscle ninhydrin positive substances and total muscle FAAconcentrations increase conspicuously (50% and 80%, respectively). Individual muscleFAA also increase markedly, e.g., proline (300%), alanine and glycine (140%) andglutamic acid (75%) (Tan and Choong, 1981). These characteristically long-termchanges are also seen to some extent in M. rosenbergii when exposed for shorter periods(27–48 h) to a high salinity medium (24‰) (Armstrong et al., 1981). Hemolymphprotein decreases rapidly (after 3 h) while tail muscle glycine and proline increase by20% and 97%, respectively, after 24 h. In both M. rosenbergii and M. ohione acclimatedfor 5 days to strongly saline media (e.g., 20 and 30‰), sodium and chlorideconcentrations account for approximately 75–80% of total hemolymph osmolality(Castille and Lawrence, 1981). That this value is unchanged from that for shrimpsmaintained in freshwater may reflect the short-term nature of the acclimation processused.
Metabolic alterations corroborating the involvement of the intracellular FAA pool inupward salinity acclimation are also found in the marine shrimp Penaeus japonicus after48 h exposure to hypersaline conditions (55‰), during which total tissue FAA increase
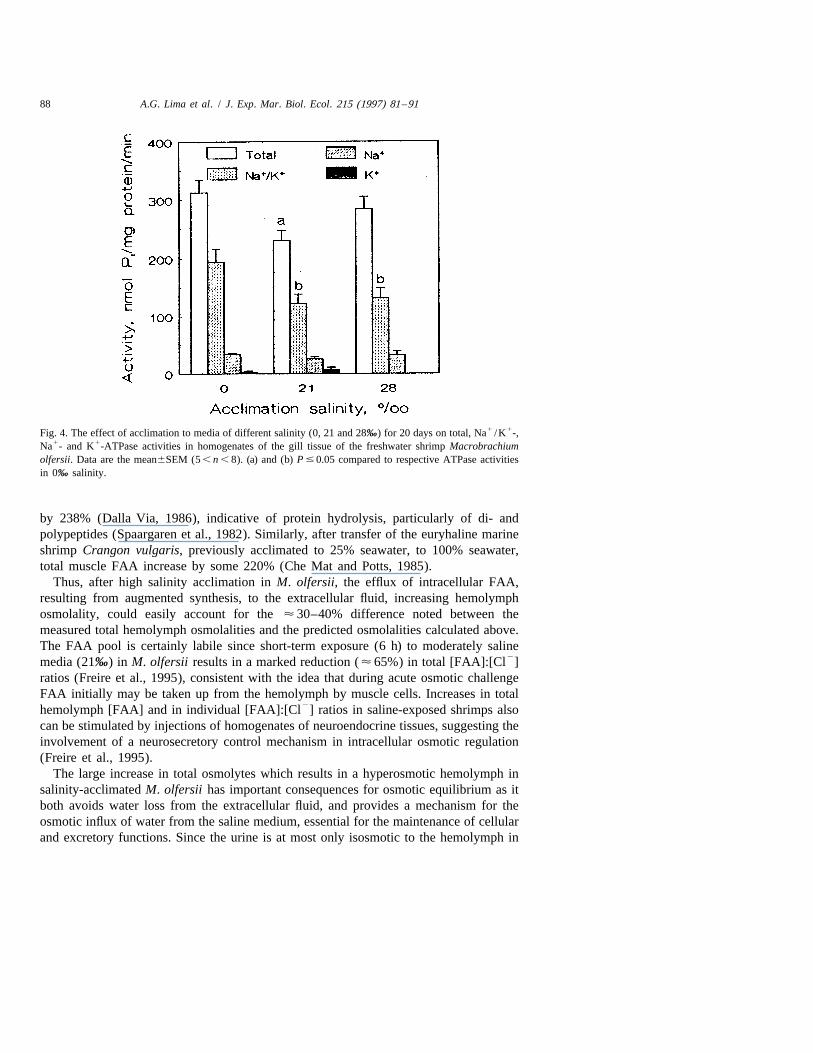
88 A.G. Lima et al. / J. Exp. Mar. Biol. Ecol. 215 (1997) 81 –91
1 1Fig. 4. The effect of acclimation to media of different salinity (0, 21 and 28‰) for 20 days on total, Na /K -,1 1Na - and K -ATPase activities in homogenates of the gill tissue of the freshwater shrimp Macrobrachium
olfersii. Data are the mean6SEM (5 , n , 8). (a) and (b) P # 0.05 compared to respective ATPase activitiesin 0‰ salinity.
by 238% (Dalla Via, 1986), indicative of protein hydrolysis, particularly of di- andpolypeptides (Spaargaren et al., 1982). Similarly, after transfer of the euryhaline marineshrimp Crangon vulgaris, previously acclimated to 25% seawater, to 100% seawater,total muscle FAA increase by some 220% (Che Mat and Potts, 1985).
Thus, after high salinity acclimation in M. olfersii, the efflux of intracellular FAA,resulting from augmented synthesis, to the extracellular fluid, increasing hemolymphosmolality, could easily account for the ¯ 30–40% difference noted between themeasured total hemolymph osmolalities and the predicted osmolalities calculated above.The FAA pool is certainly labile since short-term exposure (6 h) to moderately saline
2media (21‰) in M. olfersii results in a marked reduction ( ¯ 65%) in total [FAA]:[Cl ]ratios (Freire et al., 1995), consistent with the idea that during acute osmotic challengeFAA initially may be taken up from the hemolymph by muscle cells. Increases in total
2hemolymph [FAA] and in individual [FAA]:[Cl ] ratios in saline-exposed shrimps alsocan be stimulated by injections of homogenates of neuroendocrine tissues, suggesting theinvolvement of a neurosecretory control mechanism in intracellular osmotic regulation(Freire et al., 1995).
The large increase in total osmolytes which results in a hyperosmotic hemolymph insalinity-acclimated M. olfersii has important consequences for osmotic equilibrium as itboth avoids water loss from the extracellular fluid, and provides a mechanism for theosmotic influx of water from the saline medium, essential for the maintenance of cellularand excretory functions. Since the urine is at most only isosmotic to the hemolymph in

A.G. Lima et al. / J. Exp. Mar. Biol. Ecol. 215 (1997) 81 –91 89
freshwater palaemonids exposed to saline media (see Denne, 1968; Born, 1968),sufficient extracellular water would then be available for the excretion of the increased
1 2Na and Cl load. The necessary osmotic influx of water apparently could not begenerated by the principal, inorganic, hemolymph osmolytes alone as these are distinctlyhyporegulated at higher salinities.
1 1Gill Na /K -ATPase activity in M. olfersii is maximal in FW. This active transport´system (see Pequeux, 1995), located in the extensive membrane invaginations of the
intralamellar septal cells (McNamara and Torres, unpubl. data; McNamara and Lima,1 11997), apparently functions in concert with a Na /NH counter transporter (Armstrong4
et al., 1981) present in the apical membranes of the pillar cells, and is responsible for1 1most of the Na uptake to the hemolymph, thus maintaining the strong Na gradient
1 1seen in this medium. After acclimation to saline media for 20 days, gill Na /K -ATPase activity decreases by ¯ 35%, concomitant with hyporegulation of hemolymph
1 1Na concentrations. During the initial exposure to saline media, the external [Na ] (2871and 383 mEq/ l) is considerably higher than the hemolymph [Na ] (154 mEq/ l) in FW,
1resulting in a passive influx of Na to the hemolymph, in addition to the actively1 1transported Na component. This influx results in a rapid increase in hemolymph [Na ]
which stabilizes after 6 h (McNamara et al., 1991) but continues to increase after 12 hexposure (McNamara et al., 1990). In M. rosenbergii exposed to saline media,
1hemolymph [Na ] is finally iso-ionically regulated only after 48 h exposure (Armstronget al., 1981).
1 1In M. olfersii, the significant reduction in gill Na /K -ATPase activity after 20 daysevidently constitutes an important component of the long-term regulatory response since
1total Na influx to the hemolymph would consequently decrease. After acclimation tosaline media for 10 days, quantifiable, ultrastructural modifications take place in theapical flanges of the gill pillar cells, i.e., marked reductions in the height and density ofthe apical microvilli and in pillar flange thickness (McNamara and Lima, 1997), and inthe transporting regions of the gill intralamellar septal cells (Freire and McNamara,1995). These structural alterations apparently reduce the apical permeability of the pillarcells to ions in the medium, through the removal of membrane ion counter transporters,
1 1further reducing Na influx to the point where urinary Na excretion appears to equal1 1Na influx, resulting in a dynamic equilibrium in which hemolymph Na is hyporegu-
lated.1 1It is not clear precisely what the signal or mechanism for reduction in gill Na /K -
1ATPase activity might be in salinity-acclimated shrimps. Increased intracellular [Na ],and increases in cellular second messengers and modulators like cyclic AMP, diacyl
1 1glycerol and protein kinase C, cause a reduction in Na /K -ATPase activity in renaltissues over the short term (Bertorello and Katz, 1993). However, long-term responses
1 1may involve the degradation and/or removal of Na /K -ATPase molecules from theseptal cell membranes by membrane internalization, or a reduction in the rate of
1 1Na /K -ATPase subunit synthesis on cytoplasmic ribosomes, leading to reducedrenewal and turnover. Neuroendocrine control of these processes seems likely sinceseveral studies have shown that the dopamine /cyclic AMP second messenger cascade isinvolved in the increase in gill ATPase activity seen in crabs exposed to dilute media(see Kamemoto, 1991): bilateral eyestalk ligation decreases enzyme activity while the

90 A.G. Lima et al. / J. Exp. Mar. Biol. Ecol. 215 (1997) 81 –91
injection of brain homogenates restores gill ATPase activity (Kamemoto and Tullis,1972). Similarly, bilateral eyestalk ablation, and the injection of cerebral ganglion
1 1extracts, reduce kidney tubule Na /K -ATPase activity in the freshwater crayfishProcambarus clarkii which can be restored by the injection of eyestalk homogenates(Kamemoto and Tullis, 1972).
1The relevance of a significant Na -ATPase activity ( ¯ 30 nmol P /mg protein /min)i1in the gill homogenates of salinity-acclimated M. olfersii is not clear. Intense Na -
ATPase activity ( ¯ 185 nmol P /mg protein /min) has been reported in the gill tissue ofi
M. amazonicum (Proverbio et al., 1990) where the enzyme is suggested to be located inthe basolateral plasmalemmae of the gill cells ( 5 membrane invaginations of theintralamellar septal cells, Freire and McNamara, 1995). In M. olfersii, the activity of thissecond active component of total sodium influx is unaffected by acclimation to saline
1 1media. Both this activity and the remaining Na /K -ATPase activity would provide a1means of Na uptake for metabolic processes despite the ultrastructural modifications
1leading to reduced ionic (Na ?) permeability across the apical surface of the gillepithelium.
1 1These various data demonstrate the intrinsic coupling between gill Na /K -ATPase1 2activity and long-term regulation of hemolymph Na and Cl in M. olfersii; they also
reveal a notable difference in the response patterns of total and individual osmolytes, andsuggest that alterations in these important components of osmotic acclimation may beregulated by neurohormonal mechanisms.
Acknowledgments
This study represents part of an M.Sc. thesis submitted by AGL to the Departamentode Fisiologia / IBUSP, and was financed by a research grant to JCM (FAPESP 91/2467-2) and a postgraduate scholarship (FAPESP 90/3807-9). The authors thank Dr. L.
˜Carlos Salomao (DF/IBUSP) and Dr. J. Luneta (CEBIMAR) for access to necessaryfacilities.
References
Armstrong, D.A., K. Strange, J. Crowe, A. Knight and M. Simmons, 1981. High salinity acclimation by theprawn Macrobrachium rosenbergii: Uptake of exogenous ammonia and changes in endogenous nitrogencompounds. Biol. Bull. Mar. Biol. Lab., Woods Hole, Vol. 160, pp. 349–365.
Baginski, E. and B. Zak, 1960. Micro-determination of serum phosphate and phospholipids. Clin. Chim. Acta,Vol. 5, pp. 834–838.
Bertorello, A.M. and A.I. Katz, 1993. Short-term regulation of renal Na-K-ATPase activity: Physiologicalrelevance and cellular mechanisms. Am. J. Physiol., Vol. 265, pp. F743–F755.
Born, J.W., 1968. Osmoregulatory capacities of two caridean shrimps, Syncaris pacifica (Atyidae) andPalaemon macrodactylus (Palaemonidae). Biol. Bull. Mar. Biol. Lab., Woods Hole, Vol. 134, pp. 235–244.
Bradford, M.M., 1976. Rapid and sensitive method for the quantitation of microgram quantities of proteinutilizing the principle of protein-dye binding. Anal. Biochem., Vol. 72, pp. 248–254.
Castille, F.L. Jr. and A.L. Lawrence, 1981. The effect of salinity on the osmotic, sodium and chlorideconcentrations in the hemolymph of the freshwater shrimps, Macrobrachium ohione Smith and Macrob-rachium rosenbergii (de Man). Comp. Biochem. Physiol., Vol. 70A, pp. 47–52.

A.G. Lima et al. / J. Exp. Mar. Biol. Ecol. 215 (1997) 81 –91 91
Che Mat, C.R. and W.T. Potts, 1985. Intracellular osmotic regulation in Crangon vulgaris. Comp. Biochem.Physiol., Vol. 82A, pp. 719–724.
Claybrook, D.L., 1983. Nitrogen metabolism. In, The Biology of Crustacea, edited by D.E. Bliss, Vol. 5,Internal anatomy and physiological regulation, edited by L.H. Mantel, Academic Press, New York, pp.163–213.
Dalla Via, G.J., 1986. Salinity responses of the juvenile penaeid shrimp Penaeus japonicus. Part II. Free aminoacids. Aquaculture, Vol. 55, pp. 307–316.
Denne, L.H., 1968. Some aspects of osmotic and ionic regulation in the prawns Macrobrachium australiense(Holthuis) and M. equidens (Dana). Comp. Biochem. Physiol., Vol. 26, pp. 17–30.
Freire, C.A. and J.C. McNamara, 1995. Fine structure of the gills of the fresh-water shrimp Macrobrachiumolfersii (Decapoda): Effect of acclimation to high salinity medium and evidence for involvement of theintralamellar septum in ion uptake. J. Crust. Biol., Vol. 15, pp. 103–116.
Freire, C.A., J.C. McNamara, J.C. Rosa and L.J. Greene, 1995. Neuroendocrine control of osmotic regulationin the freshwater shrimp Macrobrachium olfersii (Wiegmann) (Crustacea, Decapoda): Free amino acidconcentrations in the hemolymph. Gen. Comp. Endocrinol., Vol. 100, pp. 83–91.
´Gilles, R. and A.J. Pequeux, 1981. Cell volume regulation in crustaceans: Relationship between mechanismsfor controlling the osmolality of extracellular and intracellular fluids. J. Exp. Zool., Vol. 215, pp. 351–362.
Kamemoto, F.I., 1991. Neuroendocrinology of osmoregulation in crabs. Zool. Sci., Vol. 8, pp. 827–833.Kamemoto, F.I. and R.E. Tullis, 1972. Hydromineral regulation in decapod Crustacea. Gen. Comp.
Endocrinol., Suppl. 3, pp. 299–307.McNamara, J.C., 1987. The time course of osmotic regulation in the freshwater shrimp Macrobrachium
olfersii (Wiegmann) (Decapoda, Palaemonidae). J. Exp. Mar. Biol. Ecol., Vol. 107, pp. 245–251.McNamara, J.C., 1993. Exposure to high salinity medium and neurosecretion in the anteromedial cells of the
supraesophageal ganglion of the fresh-water shrimp Macrobrachium olfersii (Decapoda). J. Crust. Biol.,Vol. 13, pp. 409–422.
McNamara, J.C. and A.G. Lima, 1997. The route of ion and water movements across the gill epithelium of thefreshwater shrimp Macrobrachium olfersii (Decapoda, Palaemonidae): Evidence from ultrastructuralchanges induced by acclimation to saline media. Biol. Bull. Mar. Biol. Lab., Woods Hole. Vol. 192.
˜McNamara, J.C., L.C. Salomao and E.A. Ribeiro, 1990. The effect of eyestalk ablation on hemolymph osmoticand ionic concentrations during acute salinity exposure in the freshwater shrimp Macrobrachium olfersii(Wiegmann) (Crustacea, Decapoda). Hydrobiologia, Vol. 199, pp. 193–200.
˜McNamara, J.C., L.C. Salomao and E.A. Ribeiro, 1991. Neuroendocrine regulation of osmotic and ionicconcentrations in the hemolymph of the freshwater shrimp Macrobrachium olfersii (Wiegmann) (Crustacea,Decapoda). Gen. Comp. Endocrinol., Vol. 84, pp. 16–26.
Moreira, G.S., J.C. McNamara, S.E. Shumway and P.S. Moreira, 1983. Osmoregulation and respiratorymetabolism in Brazilian Macrobrachium (Decapoda, Palaemonidae). Comp. Biochem. Physiol., Vol. 74A,pp. 57–62.
´Pequeux, A., 1995. Osmotic regulation in crustaceans. J. Crust. Biol., Vol. 15, pp. 1–60.1 1Proverbio, T., I.P. Zanders, R. Marin, J.M. Rodrigues and F. Proverbio, 1990. Effects of Na and/or K on the
21Mg -dependent ATPase activities in shrimp (Macrobrachium amazonicum) gill homogenates. Comp.Biochem. Physiol., Vol. 97B, pp. 383–390.
Read, G.H., 1984. Intraspecific variation in the osmoregulatory capacity of larval, postlarval, juvenile and adultMacrobrachium petersii (Hilgendorf). Comp. Biochem. Physiol., Vol. 78A, pp. 501–506.
Santos, F.H. and J.C. McNamara, 1996. Neuroendocrine modulation of osmoregulatory parameters in thefreshwater shrimp Macrobrachium olfersii (Wiegmann) (Crustacea, Decapoda). J. Exp. Mar. Biol. Ecol.,Vol. 206, pp. 109–120.
Spaargaren, D.H., P. Richard and H.J. Ceccaldi, 1982. Excretion of nitrogenous products by Penaeus japonicus(Bate) in relation to environmental osmotic conditions. Comp. Biochem. Physiol., Vol. 72A, pp. 673–678.
Stern, S., A. Borut and D. Cohen, 1984. Characterization of Na-K ATPase from the gills of the freshwaterprawn Macrobrachium rosenbergii (de Man). Comp. Biochem. Physiol., Vol. 79B, pp. 47–50.
Tan, C.H. and K.Y. Choong, 1981. Effect of hyperosmotic stress on hemolymph protein, muscle ninhydrin-positive substances and free amino acids in Macrobrachium rosenbergii (de Man). Comp. Biochem.Physiol., Vol. 70A, pp. 485–489.