protein turnover in adult drosophila
TRANSCRIPT

3. Insect Physiol., 1970, Vol. 16, pp. 601 to 613. Pergamon Press. Printed in Great Britain
PROTEIN TURNOVER IN ADULT DROSOPHILA
J. MAYNARD SMITH, A. N. BOZCUK, and SUSAN TEBBUTT
School of Biological Sciences, University of Sussex
(Receiwed 10 October 1969)
Abstract-Adult male Drosophila subobscu~u were pulse-labelled with aH leucine, and radioactivity incorporated into protein measured for 20 days after the pulse label, by extracting total protein and by autoradiography. The half- life of these proteins which turn over in the adult is approximately 10 days. The rate of synthesis varies greatly between tissues, being greatest in the wall of the mid-gut and almost zero in thoracic flight muscle.
Larvae were raised axenically in a medium containing labelled amino acids, and the radioactivity in protein followed for 60 days after emergence by extract- ing total protein, and for 25 days by autoradiographs. The activity in total protein declined rapidly during the first 4 days after emergence of adults, and then remained constant. Autoradiographs suggest that the initial drop is largely due to the disappearance of the pupal fat body, which contains a large amount of protein at emergence.
Ignoring storage protein in the fat body, approximately 20 per cent of the total protein in the adult male turns over, with a half-life of approximately 10 days, and the remaining 80 per cent is not replaced. Among the non-replaced proteins are the structural and mitochondrial proteins of the flight muscles. There is active synthesis of ribosomal proteins.
INTRODUCTION
THE IMMEDIATE reason for undertaking the present investigation was to provide information needed for the study of ageing in Drosophila. Changes in proteins whose turnover time is short compared to the lifespan of the fly are unlikely to be important in ageing, since damaged proteins can be replaced by new synthesis. The ‘error catastrophe’ theory of ORGEL (1963) would provide an exception, but there is evidence (DINGLEY and MAYNARD SMITH, 1969) to suggest that such catastrophes do not play an important part in ageing. We have therefore studied the amount and location of the proteins which turn over in adult Droso$hila, and
of those which do not. It was already known (CLARKE and MAYNARD SMITH, 1966; DINGLEY and
MAYNARD SMITH, 1968) that in young adult Drosophila the amount of protein synthesized per hour amounts to approximately 0.1 per cent of the total adult protein. This could mean that all, or almost all, types of protein are turning over with a half-life approximately equal to the adult lifespan after emergence of 60 days at 20°C; or it could mean that some fraction of the protein is turning over more rapidly, and the rest not at all.
601

602 J. MAYNARD SMITH, A. N. BOZCUK, AND SUSAN TEBBU-M
Two methods of distinguishing between these possibilities have been used. First, adult flies have been pulse-labelled with sH amino acids, and the subsequent decline of the label in protein has been followed. Second, larvae have been raised on a medium containing 3H amino acids, and the activity in protein has been followed throughout adult life. Two techniques, total extraction of soluble protein and autoradiography, have been used to measure the activity incorporated. The former technique is less laborious, and in some cases more accurate quanti- tatively; the latter makes it possible to locate synthesis in particular tissues and, in particular, ensures that measurements do not include proteins synthesized by the gut flora. The various methods used have given reasonably consistent results.
Flies
MATERIALS AND METHODS
The adult flies used were male F, hybrids between the B and K inbred lines aged 7 to 14 days unless otherwise stated. All stocks are raised on yeasted maize meal at 20°C. The flies labelled as larvae were B/K F, hybrids.
Labelling
Adults were labelled by hand feeding. Single flies are etherized and as each fly starts to recover, a fine glass pipette containing the labelled amino acid solution (40 PC/ml, 1000 mc/mM sH leucine in 10 per cent sucrose, unless otherwise stated) is held to its proboscis. Flies in this condition readily drink any sweetened solution.
For’ uniform labelling of larvae the following procedure was adopted. F, eggs were collected from F, females 6 to 9 days after emergence. These are induced to lay on a paste of ground killed yeast spread on agar containing acetic acid and absolute alcohol. The eggs are separated from the yeast by a series of filtrations and by suspension in strong sugar solution and finally dechorionated for 10 min in calcium hypochlorite. After washing, the embryos are transferred under a sterile hood to the surface of the culture medium in 15 x 2.5 cm boiling tubes. The technical details of the medium are given by SANG (1956).
For the leucine labelling, 1.5 ml of a solution containing 40 PC/ml of 1000 mc/ mM sH leucine was added per 100 ml of defined medium. The standard medium contains 3 mM leucine/lOO ml.
For the phenylalanine labelling, 0.325 ml of a 200 &nl, 1000 mc/mM 3H phenylalanine solution was added per 100 ml of defined medium. The standard medium contains 1.5 mM of phenylalanine per 100 ml.
Protein extraction
Flies are homogenized in 10 per cent trichloracetic acid. The precipitate is washed with TCA at 100°C. The soluble protein is dissolved from the precipitate with 1 ml O-4 N sodium hydroxide which, after centrifugation, is transferred to scintillation vials. The residue is again treated with NaOH and the supematant added to the scintillation vials. The protein is reprecipitated with excess 10%

PROTRIN TURNOVER IN ADULT DROSOPHILA 603
TCA. The precipitate is spun down in the vials and dissolved in 1 ml of hyamine hydroxide (Packard). Ten ml of scintillation mixture (0.3% PPO in AnalaR toluene) is added and the samples counted in a Beckman scintillation counter. Further technical details are given by DINGLEY and MAYNARD SMITH (1968). It is shown in that paper that for periods from 30 to 120 min between feeding and homogenization the activity incorporated is proportional to the time, from which it is concluded that the leucine fed enters rapidly into the pool from which protein is synthesized.
Autoradiography
Flies were fixed in Carnoy’s fluid (6 : 3 : 1) for 2 hr, and dehydrated in 90 per cent and absolute ethanol, being given three changes in each alcohol for at least 1 hr per change. Specimens were not left for more than 6 hr in absolute alcohol. The flies were then cleared in xylene for 45 min and embedded in 56°C paraffin wax by changing three times for at least 1 hr each change, or by keeping overnight in molten wax. Specimens were then blocked, sliced at 6 p and mounted on ‘subbed’ 3 in. x 1 in. Chance microscope slides smeared with glycerine albumen.
After mounting, the sections were cleared in xylene and hydrated through
lOO%, 90%, 70%, 50% and 30% ethanol. The slides were washed in a few changes of CO,-free distilled water. Then in a dark room, under an Ilford 906 iso dark-red safe light, Kodak AR10 stripping film pieces were coated on the sections. The slides were then dried, and stored in light-tight boxes at 4°C for from 1 to 3 weeks. They were then developed with Kodak D19b or D19 for 4 min, washed for 15 set in distilled water, and fixed in 25% Kodafix for 3 min. They were then rinsed in tap water, and washed for 30 min in running tap water, and finally washed in distilled water for 5 min. The slides were stained through the film after development in Ehrlich’s acid haemotoxylin/eosin. Further details are given by P~~c(1956).
The slides were examined under a high-power microscope. Grain counts were made in an area of 36 p2, using an eyepiece grid. For each tissue in each fly, counts were made in four representative squares, and an average count was calcu- lated. The background was estimated for each slide by counting the grain in four representative squares away from the section; in most cases the background was less than one grain/36 p2, but was appreciably higher in flies 20 days after labelling which were exposed for 4 weeks. The background was subtracted from the average count from a tissue, to give an estimate of activity in grains/36 I_L~_ In comparing different flies, this activity was divided by the weeks of exposure, and by the mg of 3H solution fed to that fly. Thus final comparisons, which are propor- tional to the rate of protein synthesis per unit volume of tissue, are given in grains (corrected for background)/36 p2 per mg of solution fed per week of exposure. To check that all free leucine is removed from the sections by the washing procedures described, flies were fed on 3H leucine solution and fixed immediately (i.e. within 5 min of starting to feed). In these experiments, the grain counts in the tissues did not differ from the background level.

604 J. MAYNARD SMITH, A. N. BOZCUK, AND SUSAN TEBBUTT
Comparison of extraction and autoradiographic techniques
To interpret the results of our experiments, it is necessary to know to what extent the same proteins are retained by the techniques of extraction and auto- radiography. Groups of males were fed on 3H-leucine solution (3.3 mc/mM, approximately five times the free leucine pool being fed to each fly), and killed after 4 hr. They were divided into three groups, as follows:
(a) Four groups of 4 males each were extracted in the usual way. (b) Four groups of 2 males each were prepared for autoradiography in the usual
way but, instead of being coated with stripping film, the sections were scraped off the slides, homogenized in TCA with 2 additional flies as carriers, and protein was extracted in the usual way.
(c) Four groups of 2 males each were prepared for autoradiography, and then scraped off the slides into 1 ml of hyamine.
The results are shown in Table 1. Only the difference between groups (a) and (b) is significant (O-01 <P< O-05). Approximately 70 per cent of the NaOH- soluble protein is retained in the autoradiographs. There is a suggestion that some labelled protein is retained in the autoradiographs which is insoluble and so not
TABLE ~-COMPARISON OF PROTEIN FRACTIONS RETAINED BY DIFFERENT PROCEDURES
Dis/min per fly per 4 hr
(a) Total NaOH-soluble counts 1289f 72 (b) NaOH-soluble counts retained in autoradiographs 887 + 133 (c) Total counts in autoradiographs 1138 f 144
measured in the extraction procedure ; measurements of radioactivity in the NaOH-insoluble residue during the extraction procedure has confirmed that a small fraction (approximately 10 per cent) of the turning over protein remains in this fraction.
The technique of pulse labelring
Individual males were fed by hand on 3H leucine (40 &ml, 100 mc/mM in 10% sucrose), and weighed before and after feeding to determine the amount taken. After 2 hr, they were fed a solution of cold O-1 M leucine; on this occasion the flies were not weighed, but each fly was fed until swollen, which requires 0.3 to O-4 mg of solution per fly. Unfortunately this is insufficient to reduce the specific activity of the leucine pool to a level below that of the leucine incorporated into protein. It is therefore necessary to wait for 24 hr, when most of the excess leucine has been excreted, and then to give a third feed, again of cold O-1 M leucine. After this second feed, the specific activity of leucine in the pool is lower than in turning over protein; the fly has therefore been pulse-labelled. The evidence for these statements, and for the belief that any radioactivity incorporated into protein is incorporated as leucine, will now be given.

PROTEIN TURNOVER IN ADULT DROSOPHILA 605
Thirty-eight males 10 days after emergence were fed a solution of 14C leucine, 165 mc/mM, 5 PC/ml, in 10% sucrose. These flies took a total of 4.33 mg of solution, containing 43,300 disjmin. After 2 hr, each fly was fed by hand on cold O-1 M leucine, and transferred to yeasted maize meal. After a further 24 hr, they were homogenized in 1 ml of 10% TCA containing O-4 PM of n-leucine as a standard. The homogenate was separated into a supernatant, containing any free leucine remaining in the pool, and a precipitate containing any leucine incor- porated into protein. The precipitate was then hydrolysed in 5.7 N HCl.
Aliquots of 10 per cent of the supernatant and one-sixth of the hydrolysate were counted in a scintillation counter with an efficiency of 75 per cent. The remainder of the supernatant and the hydrolysate were run through an amino acid analyser, and the amount of leucine in each, and the radioactivity associated with each peak, were measured. Running a separate standard containing 20 rnt+ of i4C leucine gave a counting efficiency of 32 per cent.
Considering first the supernatant, measurement of the n-leucine peak showed that 64 per cent of the original sample remained (instead of 90 per cent if there had been no losses). No radioactivity above background was found except in the leucine peak, which contained 528 counts/min above background. Hence
528 1 100 v64’ -x 0.32 43,300 - = 5.95 per cent of the 14C leucine
which was originally fed remained in the pool. Measurement of the leucine peak gave 2.49 mpM of free leucine per fly. This is approximately three times the normal level, so even after 24 hr the excess leucine fed has not been fully excreted. Since 30 to 40 mpM of cold leucine was fed per fly, the value of 2.49 mpM remaining agrees well with the value of 5-95 per cent obtained by measuring radioactivity.
Turning to the hydrolysate, there was again no activity except in the leucine peak, which gave 2587 counts/min above background. Assuming that the hydro- lysate was 64 per cent of the original total, this corresponds to 29.2 per cent of the i4C leucine originally fed. Measurement of the leucine peak gave 91 mpM/fly. Earlier measurements (IMLAH, 1967) g ave 108 wM/fly of leucine in protein, so the figure of 64 per cent recovery is not badly wrong.
Finally, measurements on the aliquots gave 165 counts/mm in the super- natant and 1246 counts/min in the hydrolysate. Allowing for losses and for counting efficiency, this suggests that of the total i4C leucine fed, 7.2 per cent remained in the free pool and 32.3 per cent was incorporated into protein. These figures agree well with those obtained from the analyser, and show that the activity in leucine, free and in protein, can adequately account for the activity of the fly after 24 hr.
To conclude, 24 hr after feeding on cold leucine, 6 per cent of the original i4C leucine is in the free leucine pool, 29 per cent is in leucine incorporated into protein, and there is no detectable label elsewhere. The free pool, at 2.5 mpM/fly, is approximately three times its normal level.

606 j. MAYNARD SMITH, A. N. BOZCUK, AND SUSAN TEBBUTT
In the pulse-labelling experiments described below, the flies were then given a third feed, of approximately 0.3 mg of cold O-1 M leucine. After this feed, there would be, approximately: 6 per cent of the original label in 30 mpM of pool leucine, and 29 per cent of the original label in 100 WM of protein leucine.
Hence after the second cold feed the specific activity of the protein leucine is still only slightly higher than that of the pool leucine. However, it is shown below that only one-third or less of the total protein is turning over. Only this protein will be labelled, and its specific activity will be approximately five times that of the pool. Flies were subsequently kept on yeasted maize meal, so that the pool continued to be diluted with cold leucine.
Protein turnwer measured by pulse label&g
Three experiments have been performed to measure the activity incorporated into protein after pulse labelling. The results are summarized in Table 2. Each
TABLE 2--PBRCENTAGE OF 8H LEUCINR ORIGINALLY FED WHICH IS INCORPORATED INTO
PROTEIN
Time in days Experiment Mean for three after experiments
pulse label 1 2 3
0 26.2 * 2.9 30.4 f 4.6 37.1 * 7.8 31.2 3 21.5 f 1.6 23.5 f 3.6 27.0 zk 4.7 24.0 9 12.4 + 1.0 19.5 f 8.3 17.7 + 2.9 16.5
19 - 12.9* 13.3 f 2-l 13.1
* Three flies only.
percentage value (with one exception which is indicated in the table) is the average of between four and six values each based on a single fly. The initial values, varying from 26.2 to 37.1 per cent, agree well with the value of 29.2 per cent, obtained in the experiment described in the previous section.
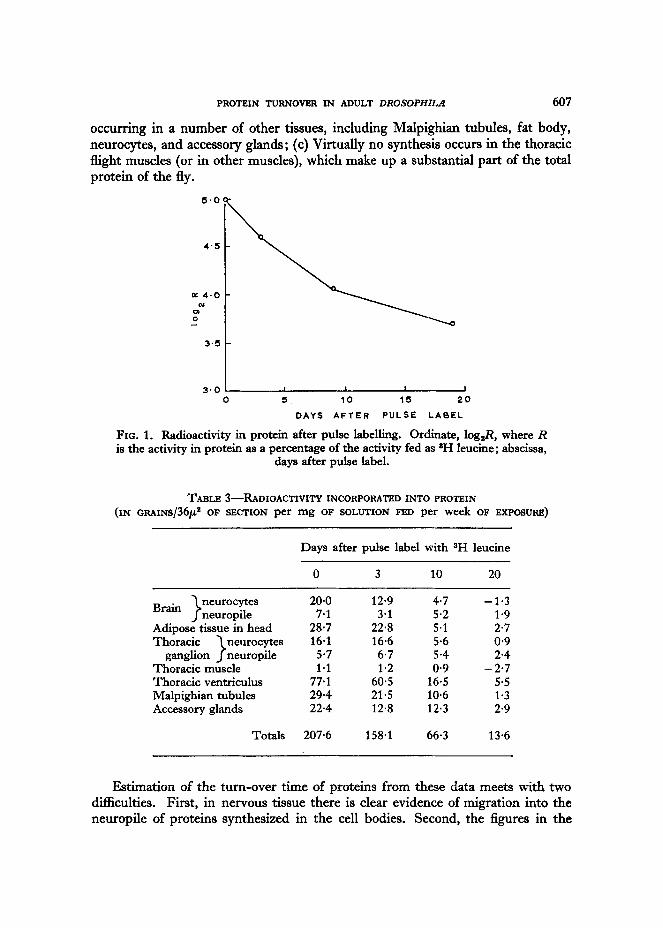
After 9 days on maize meal, the activity fell to approximately half its initial value. In Fig. 1, the unweighted means for the three experiments are plotted on a logarithmic scale. The departure from linearity shows that there is some hetero- geneity of turn-over time for different proteins.
It is concluded that those proteins which are turning over, and which therefore incorporate sH leucine, have a half-life of approximately 10 days.
Protein turnwer measured by autoradiography
Table 3 shows the results of pulse labelling with SH leucine in various tissues. The 0 and 10 day values are the averages for 4 flies, and the 3 and 20 day values for 3 flies.
The first column of the table shows: (a) The most rapid protein synthesis occurs in the wall of the mid-gut, i.e. the thoracic ventriculus; (b) Synthesis is also
PROTEIN TURNOVER IN ADULT DROSOPHILA 607
occurring in a number of other tissues, including Malpighian tubules, fat body, neurocytes, and accessory glands; (c) Virtually no synthesis occurs in the thoracic flight muscles (or in other muscles), which make up a substantial part of the total protein of the fly.
I I I I 0 5 10 15 20
DAYS AFTER PULSE LABEL
FIG. 1. Radioactivity in protein after pulse labelling. Ordinate, log& where R is the activity in protein as a percentage of the activity fed as *H leucine; abscissa,
days after pulse label.
TABLE 3--RADIOACTIVITY INCORPORATED INTO PROTEIN
(IN GRAINS/~~$ OF SECTION per mg OF SOLUTION FED per week OF EXPOSURE)
Days after pulse label with 8H leucine
0 3 10 20
Brain neurocytes neuropile
Adipose tissue in head Thoracic
>
neurocytes ganglion neuropile
Thoracic muscle Thoracic ventriculus Malpighian tubules Accessory glands
20.0 12.9 4.7 -1.3 7.1 3.1 5.2 1.9
28.7 22.8 5.1 2.7 16.1 16.6 5.6 0.9
5.7 6.7 5.4 2.4 1.1 1.2 0.9 - 2.7
77-l 60.5 16.5 5.5 29.4 21.5 10.6 1.3 22.4 12.8 12.3 2.9
Totals 207.6 158.1 66.3 13.6
Estimation of the turn-over time of proteins from these data meets with two difhculties. First, in nervous tissue there is clear evidence of migration into the neuropile of proteins synthesized in the cell bodies. Second, the figures in the
608 J. MAYNARD SMITH, A. N. BOZCUK, AND SUSAN TEBBUTT
final column, for flies 20 days after labelling, are very inaccurate, since in most tissues the activity is only slightly above background, and in a few cases is actually lower than background.
To obtain an estimate corresponding to that obtained by total extraction of protein, it would be necessary to weigh the value for each tissue by the volume of that tissue. However, an approximate idea can be obtained from the totals for each column. These totals suggest that the activity has fallen to one-third of its initial value after 10 days, agreeing reasonably well with the data from total extraction. There is also an indication that the turn-over time for the most rapidly synthesizing tissue, the thoracic ventriculus, is shorter than in other tissues; this would confirm the finding that there is some heterogeneity of turn-over time.
Total labelling of larvae
The data of the previous section suggest that the half-life of turning-over proteins in the adult is approximately 10 days. Hence approximately 1/35Oth of the turning-over protein is replaced per hr. Data on the rate of synthesis show that the protein synthesized per hr is approximately 0.05 per cent of the total protein. Hence 20 per cent, or somewhat less, of the total protein in the adult is turning over, and 80 per cent is not.
A direct check on this conclusion can be obtained by raising larvae on a medium containing 3H amino acids, and following incorporated radioactivity throughout adult life. Fig. 2 shows the results for two groups of Aies, raised as larvae on SH
20000 r
I , 10 20 30 46 50 60
AGE IN DAYS
FIG. 2. Radioactivity in flies labelled as larvae. Full line, labelled with *H phenyl- alanine; broken line, labelled with *H leucine. Background <200 dis/min.
leucine (60 ~43 mM Leu per 100 ml medium) and SH phenylalanine (65 41.5 mM Phe per 100 ml medium) respectively. In both cases there was a rapid fall of

PROTEIN TURNOVER IN ADULT DROSOPHILA 609
incorporated label during the first 4 days of adult life. Subsequently there was no significant change for the remaining 60 days of adult life. This is as expected from the pulse-labelling experiments, and confirms the presence in young adults of a substantial component of the total protein which lasts unreplaced throughout adult life.
The initial fall in activity is, however, greater than would be predicted, amount- ing to from one-half to two-thirds of the total protein of the freshly emerged fly. This implies a fall in the total protein of the fly during the first few days, since the rate of synthesis during this period is no higher than the typical adult figure of 0.05 per cent per hr, which is insufficient to replace the loss of one-half of the total protein in 4 days.
An explanation for this discrepancy was obtained by autoradiography, carried out on flies from the same culture tubes as those on which the phenylalanine data in Fig. 2 were obtained. The results are given in Table 4. Fig. 3 shows the values in the pupal fat body, and the average value for the remaining tissues, for the
TABLE ~-RADIOACTIVITY INCORPORATED INTO PROTEIN IN FLIES LABELLED WITH SH PHENYLALANINEAS LARVAE(IN GRAINS/36$ OF SECTION per 15 days OFRXPOSURE)
Days after emergence
0 1 2 5 15 25
Number of flies 2 2 4 3 5 5 Brain (neuropile) 3.12 2.37 5.28 5.58 6.25 4.15 Thoracic ganglion 3.00 4.87 5.81 5.25 6.25 6.90 Flight muscle 2.62 3.87 4.56 3.83 5.40 5.55 Cardia 3.25 2.87 2.03 2.25 1.55 1.15 Thoracic ventriculus 2.00 2.12 3.34 2.50 1.80 1.00 Testis 3.37 3.75 4,81 3.75 5.45 4.90 Accessory glands 2.62 1.37 2.00 2.25 2.58 2.35 Fat body 6.75 6.37 3.34 1.25 l-55 1.00
first 25 days after adult emergence. The loss of radioactivity after emergence is confined to the pupal fat body in the abdomen. This activity is in the large pupal cells; the low residual level of activity after 5 days is in the small cells which compose the adult fat body. The rise in other tissues is not statistically significant ; it may however indicate net synthesis of protein in these tissues, using amino acids derived from the breakdown of the fat body. The main conclusions to emerge are :
(i) The loss of activity during the first five days is mainly from the pupal fat body, which is a massive structure in freshly emerged flies, with a high protein content.
(ii) Even in those tissues in which appreciable protein synthesis occurs in adults, there is an appreciable protein component which does not turn over.
In other experiments using total protein extraction to follow label incorporated during larval life, the fall in activity during the first 2 days was less than that shown

610 J. hlAyNARD SMITH, A. N. BOZCUK, AND SUSAN TEBBUTT
in Fig. 2; this variability probably reflects variations in the size of the fat body in different batches of flies.
I I I 1
5 10 15 20 25
AGE IN DAYS
FIG. 3. 8H phenylalanine in protein in flies labelled as larvae. Full line, fat body; broken line, average for seven other tissues.
CONCLUSIONS
It is clear from Fig. 2 that an appreciable proportion of the total protein of an adult fly is not replaced during adult life. This conclusion is important in inter- preting the causes of ageing in Drosophila.
The magnitude of the initial drop shown in Fig. 2 suggests that only between one-half and one-third of the total protein of a fly falls into the non-replaceable category. This is an underestimate, however, because much of the drop is accoun- ted for by the disappearance of the pupal fat body. A better estimate of the proportion of protein which is turning over can be obtained from estimates of the rate of synthesis of protein and of the half-life of turning-over protein. Thus the incorporation of 1289 dis/min per fly per 4 hr (Table 1) corresponds to the syn- thesis of protein containing 0.053 mpM of leucine/fly per hr. The total protein of an adult male does not change significantly during adult life, and contains 108 mpM of leucine. Hence approximately 1 part in 2000 of the total protein is replaced per hr. The half-life of the turning-over protein is approximately 10 days. Com- bining these estimates, slightly less than 20 per cent of the total protein is turning over, and 80 per cent is not replaced.
More important than the precise proportion of turning-over protein is the question of which particular proteins belong to the non-replaced category. From Fig. 3 it is clear that there are non-replaced proteins in all tissues, including those in which synthesis is most rapid. The almost total absence of synthesis in the thoracic flight muscles shows not only that the structural proteins of the muscles are
611
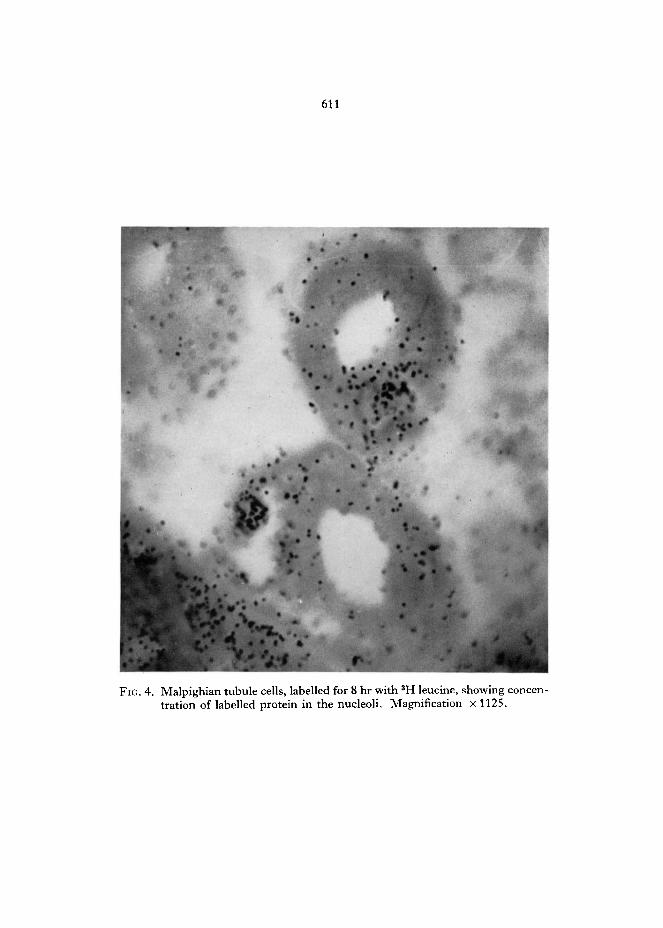
FIG. 4. Malpighian tubule cells, labelled for 8 hr with 3H leucine, showing concen- tration of labelled protein in the nucleoli. Magnification x 1125.


PROTRIN TURNOVRR IN ADULT DROSOPHILA 613
permanent, but also that the proteins of the flight muscle mitochondrialast through- out adult lie. This is important in view of TRIBE’S demonstration (1967) that the coupling between oxidation and phosphorylation in the flight muscle mitochondria of blowflies deteriorates with age.
The autoradiographs also give some evidence on the types of protein which are turning over. Fig. 4 shows an autoradiograph in which flies were kept for 8 hr after feeding with 3H leucine. In both mid-gut and Malpighian tubule cells there is a concentration of grains in the nucleoli; this concentration is not seen in flies kept only 4 hr after labelling. This demonstrates a migration of recently syn- thesized proteins to the nucleolus; these are presumably ribosomal proteins. If so, a substantial part of the protein synthesis in adults is of ribosomal proteins.
Acknowledgement.-This work was supported by the Medical Research Council.
REFERENCES
CLARKE J. M. and MAYNARD SMITH J. (1966) Increase in the rate of protein synthesis with age in Drosophila subobscura. Nature, Lond. 209, 627-629.
DINGLEY F. and MAYNARD SMITH J. (1968) Temperature acclimatization in the absence of protein synthesis in Drosophila subobscura. J. Insect Physiol. 14, 1185-1194.
DINGLEY F. and MAYNARD SMITH J. (1969) Ab sence of a life-shortening effect of amino acid analogues on adult Drosophila. Exp. Geront. 4, 145-149.
IMLAH J. G. (1967) Studies on the amino acid metabolism of adult Drosophila subobscura. Ph.D. Thesis, University of London.
ORGEL L. E. (1963) The maintenance of the accuracy of protein synthesis and its relevance to ageing. Proc. U.S. nat. Acad. Sci. 49, 517-521.
PELC S. R. (1956) The stripping-film technique of autoradiography. Int.J. appl. Radiat. Isotopes 1, 172-177.
SANG J. H. (1956) The quantitative nutritional requirements of Drosophila melanogaster. J. exp. Biol. 33, 45-72.
TRIBE M. A. (1967) Changes taking place in the respiratory efficiency of isolated flight muscle sarcosomes, associated with the age of the blowfly, Calliphora erythrocephala. Comp. Biochem. Physiol. 23, 607-620.