programming effects of short prenatal exposure to dexamethasone in sheep
TRANSCRIPT

Programming effects of short prenatal exposureto cortisol
MIODRAG DODIC,1 VICKY HANTZIS, JHODIE DUNCAN,* SANDRA REES,*IRENE KOUKOULAS, KELLI JOHNSON, E. MARELYN WINTOUR, AND KAREN MORITZHoward Florey Institute and *Department of Anatomy and Cell Biology,University of Melbourne, Parkville 3010, Melbourne, Victoria, Australia
ABSTRACT Recent studies have linked fetal expo-sure to a suboptimal intrauterine environment withadult hypertension. The aims of this study were two-fold: 1) to see whether cortisol treatment administeredto the ewe for 2 days at 27 days of gestation (term �150days) resulted in high blood pressure in offspring; 2) tostudy the effect of the same treatment on gene expres-sion in the brain at 130 days of gestation and in lambsat 2 months of age. Mean arterial pressure was signifi-cantly higher in the adult female and male offspring ofsheep treated with cortisol than in the control group(females: 89�2 mmHg vs. 81�2; P<0.05 and males:102�4 mmHg vs. 91�3; P<0.05). Prenatal cortisoltreatment led to up-regulation of angiotensinogen, AT1,MR, and GR mRNA in the hippocampus in fetuses at130 days of gestation but not in the animals at 2 monthsof age. This is the first evidence that short prenatalexposure to cortisol programmed high blood pressurein the adult female and male offspring of sheep.Altered gene expression in the hippocampus couldhave a significant effect on the development of thehippocampus, and on postnatal behavior.—Dodic, M.,Hantzis, V., Duncan, J., Rees, S., Koukoulas, I., John-son, K., Wintour, E. M., Moritz, K. Programming ef-fects of short prenatal exposure to cortisol. FASEB J.16, 1017–1026 (2002)
Key Words: renin-angiotensin system � glucocorticoid � sheep� mineralocorticoid
There is solid epidemiological evidence to suggestthat small size at birth (for gestational age) is associatedwith an increased incidence of adult-onset diseases ordysfunction, including syndrome X (hypertension,non-insulin-dependent diabetes mellitus, and hyperlip-idemia) (1–3). It is thought that an adverse intrauterineenvironment during a critical stage of developmentpermanently alters or ‘programs’ the development offetal tissues, which allows the fetus to survive but withadverse consequences in postnatal life. Two systemsthat could serve as a mechanism for programming arethe hippocampal-hypothalamo-pituitary-adrenal axis(HHPA) and the renin-angiotensin system (RAS).
Low birth weight adults showed greater rates ofurinary glucocorticoid excretion (4), elevated basalplasma cortisol concentrations (5), and greater adreno-
cortical responsiveness to adrenocorticotropic hor-mone (ACTH) (6, 7). These studies proposed that thelink between the size at birth and altered HHPA axisfunction in later life might be a mechanism wherebyprogramming of adult cardiovascular and metabolicdiseases occur. Low birth weight infants have smallkidneys, elevated cord blood renin and angiotensin IIconcentrations, and elevated renin gene expression inthe kidney (8–10).
Animal studies to test the ‘programming’ hypothesishave imposed perturbations such as moderate to severematernal undernutrition, restriction in specific dietarycomponents (iron, protein), or restricting normal pla-cental growth either throughout pregnancy or duringparts of gestation and confirmed that restriction of fetalgrowth leads to elevated blood pressure in the progenyof rats (11, 12). Other models to study the program-ming hypothesis used prenatal glucocorticoid expo-sure, such as elevating endogenous levels of glucocor-ticoids by using carbenoxolone (11�-HSD inhibitor,which blocks placental inactivation of endogenous glu-cocorticoids) (13). Adult rats exposed to a large dose ofcarbenoxolone throughout gestation were of low birthweight and had high blood pressure, increased basalcorticosterone levels, increased corticotropin-releasinghormone levels, and reduced GR mRNA in the hypo-thalamic paraventricular nucleus (14). Other studies totest the programming hypothesis by naturally occurringglucocorticoids used either prenatal exposure to stressor infusions of corticosterone or adrenocorticotrophin(15). Administration of the synthetic glucocorticoiddexamethasone to rats throughout pregnancy led toincreased blood pressure in male and female offspring(16). If dexamethasone was administered only duringthe last week of pregnancy, plasma corticosterone levelswere elevated in 16-wk-old offspring, whereas mineralo-corticoid (MR) and glucocorticoid (GR) mRNA levelsin the hippocampus were decreased (17). Prenatalglucocorticoid exposure also had marked effects onpostnatal behavior (18); in particular, late gestationexposure to dexamethasone impaired coping and
1 Correspondence: Howard Florey Institute, University ofMelbourne, Parkville 3052, Victoria, Australia. E-mail: [email protected]
10170892-6638/02/0016-1017 © FASEB

learning in aversive situations, which was attributed toaltered hippocampal corticosteroid receptor levels.
We were the first to show that exposure of pregnantewes to high levels of dexamethasone for only 2 daysvery early in gestation (at a mean age of 27 days of the150 day gestation period) results in hypertensive femaleoffspring at 3–4 months of age (19). This hypertensionamplifies with age and is associated with increasedcardiac output, left ventricular hypertrophy with re-duced cardiac functional reserve, and increased insulinsensitivity of the inhibition of lipolysis (20–22). Re-cently this finding of increased blood pressure in theprenatally treated offspring has been confirmed in asecond cohort of animals in both females (23) andmales (unpublished data). In late gestation, fetuses ofewes treated with dexamethasone in early pregnancyshowed significant changes in gene expression in thebrain for components of the RAS but no changes in MRand GR mRNAs in the hippocampus (23).
The hypotheses tested in the current study weretwofold: 1) that prenatal treatment with naturally oc-curring glucocorticoid cortisol administered to the ewefor only 2 days at a mean age of 27 days of gestation athigh but still physiological levels would result in highblood pressure in male and female offspring at 1.5 yearsof age; 2) that this cortisol treatment would have effectson gene expression in the brain similar to dexametha-sone treatment. The aim was to show that physiologi-cally relevant concentrations of natural glucocorticoidcould reproduce the programming effects of dexa-methasone in this very early ‘window’ of time.
MATERIALS AND METHODS
Animals
All experiments were approved by the Animal Ethics Com-mittee of the Howard Florey Institute in accordance withNational Health and Medical Research Council guidelines.Pregnant merino ewes (n�55) weighing 45–55 kg were usedin this study. On day 22–23 of gestation, a silastic cannula(inner diameter 0.76 mm, outer diameter 1.65 mm) wasinserted into a maternal jugular vein under local anesthesia(0.5 mL of 0.5% Xylocaine, Astra). Ewes were then infusedwith isotonic saline (0.19 mL/h, n�28) or cortisol (5 mg/h,n�27) for 48 h. In the cortisol-infused ewes carrying singletonfetuses (first cohort, see below), plasma cortisol concentra-tions increased from 51 � 11 to 390 � 23 nmol/L. Plasmacortisol was measured in the maternal plasma using a radio-immunoassay, which has been described in detail and vali-dated (24). Intra-assay and interassay coefficients of variationwere 10.3 and 13%, respectively.
A first cohort of ewes carrying single fetuses was allowed tolamb. In these animals blood pressure was measured at �1.5years of age. There were 17 females (saline n�8 and cortisoln�9) and 16 males (saline n�8 and cortisol n�8). Femaleanimals were oophorectomized and carotid artery loops con-structed at 1 year of age (19). In females, tail docking wasperformed at 2 months of age. In males, castration and taildocking was performed at 2 months of age.
A second cohort of ewes (n�5 saline, n�5 cortisol) wasmaintained until fetuses were at 130 days of gestation, at
which time they were killed (100 mg/kg pentobarbitone,Lethabarb, Arnolds, Reading) and fetal organs (kidney,heart, lung, brain, adrenal) weighed and collected. All eweskilled at this stage carried twin fetuses. Thus, there weretissues from 10 fetuses in each treatment group at 130 days(saline group: seven female and three male fetuses; cortisolgroup: six female and four male fetuses). The brain wasfurther dissected and hippocampus, hypothalamus, and me-dulla oblongata were frozen in liquid nitrogen for laterextraction of mRNA. Hippocampus was taken and processedwith paraffin for gene localization studies.
A third cohort of ewes, all carrying twins, was allowed tolamb and suckled their lambs until 2 months of age. From thiscohort, brain tissues were collected from the saline (n�7; 4females and 3 males) and cortisol (n�5; 3 females and 2males) -exposed animals. One lamb in each group diedbetween 1 and 2 months of age.
Placental samples collected from saline-treated ewes (n�3)at the end of the treatment (28 days of gestation) from anearlier study (25) were extracted to assess the presence ofmRNA for 11�-hydroxysteroid dehydrogenase type 2 (11�-HSD2). One whole fetus was sectioned and stained in hema-toxylin and eosin (H&E) to illustrate the stage of braindevelopment at the time of treatment (28 days of gestation)(Fig. 1).
Blood pressure and heart rate measurement protocol
In each group of sheep, mean arterial pressure (MAP) andheart rate (HR) were measured at �1.5 years of age every 10min for 3 days, as described (19). MAP and HR weremeasured via a Tygon cannula (1.0 mm i.d, 1.5 mm o.d)inserted 10 cm proximally into a carotid artery loop andconnected to a pressure transducer (TD XIII, Cobe). Thepressure was corrected to compensate for the height oftransducer above the level of the heart (19). The analogsignal was digitally converted via a DT 301 Board DataTranslation device (Marlboro, MA) and blood pressure andheart rate data were collected at 100 Hz (HEM 3.1; Notocord,Kent Scientific Corp, Litchfield, CT). Heart rate was calcu-lated by software from dP/dt Max and took into account thepick of the dP/dt curve during systole vs. time.
Preparation of RNA
Total RNA was extracted from brain tissues (hippocampus,hypothalamus, and medulla oblongata) using the phenol-chloroform method (26). As pure RNA is required forreal-time PCR, samples were DNase treated to eliminate anyresidual genomic DNA. To 75 �L of RNA in water, thefollowing were added: 10 �L 0.1M DTT, 10 �L 50 mM Mg Cl2,3.3 �L 3M NaOAc, 0.5 �L RNase inhibitor (40 U/�L), and 1�L DNase I (RNase free, 1 U/�l). The mixture was incubatedat 37°C for 15–20 min, then purified using a phenol extrac-tion (twice) and chloroform extraction. Samples were storedat �80°C until use.
Before use in real-time PCR, 1 �g of each sample wasreverse transcribed in a 10 �L reaction containing 1� Taq-Man® RT buffer, 5.5 mM MgCl2, 500 �M each 2�-de-oxynucleoside 5�-triphosphate, 2.5 �M random hexamers, 0.4U/�l RNase inhibitor, and 1.25 U/�l MultiScribe™ reversetranscriptase (PE Applied Biosystems, Foster City, CA). Toensure there was no contaminating genomic DNA, controlreactions without reverse transcriptase were included in aseparate reverse transcription reaction with all total RNAsamples. Reverse transcription was performed in a GeneAmpPCR System 9600 (PE Applied Biosystems) at 25°C for 10 min,
1018 Vol. 16 July 2002 DODIC ET AL.The FASEB Journal

48°C for 30 min, and 95°C for 5 min. Upon completion, allsamples were stored at �80°C until use.
Real-time PCR
A comparative CT (cycle of threshold fluorescence) methodwas used to determine relative mRNA expression levels in thehippocampus, hypothalamus, and medulla oblongata of MRand GR receptor, angiotensin II type 1 and type 2 receptors(AT1 and AT2), angiotensinogen (A’ogen), 11�-hydroxy-steroid dehydrogenase type 2 (11�-HSD2) along with anendogenous reference gene, 18S ribosomal RNA, at 130 days
of gestation. The expression of MR, GR, AT1, and A’ogen wasstudied in hippocampus from lambs killed at 2 months of age.11�-HSD2 was studied in the placenta of saline-treated ewes.This method has been described elsewhere (25, 27). Allprimers and TaqMan® probes were designed using PrimerExpress™ Version 1.0 (PE Biosystems). The primer andTaqMan® probe sequences for all genes are shown in Table1. The TaqMan® probe and primers for 18S were supplied byPE Applied Biosystems in a control reagents kit. PCR reac-tions were carried out in 25 �L volumes consisting of 1�TaqMan® Universal PCR Master Mix (including passive ref-erence), 50 nM TaqMan® 18S probe, 20 nM 18S forward
Figure 1. A sagittal section of the ovineembryo at 28 days of gestation. Originalmagnification: �40.
TABLE 1. Primer and probe concentrations and sequences used in real time PCRa
Probe sequence PositionConcentration
(nM)
AT1 ACCGCTGGCCCTTCGGCAA (596–614) 150AT2 TGGCTTCCCTTCCATGTTCTGACCTTC (596–614) 75A’ogen CCACGGACCCAAATCTCGCTGC (723–744) 75MR TCCTCATTTCTTCAAACGCAGCCTGG (486–511) 100GR AAAGAAGATTTTATCGAACTCTGCACCCCTGG (425–455) 7511�-HSD2 TGCTTGGCTGCCTATGGGACCTCC (681–705) 125
Forward primer sequence
AT1 GGGCTGTCTACACTGCTATGGAA (572–594) 900AT2 TGTTCTGGCGTTCATCATTTG (642–662) 300A’ogen CTCTCCCACGCTCACTAGACTTG (699–721) 300MR TCCAAAGGATGGCCTCAAAA (465–484) 900GR ACTGCCCCAAGTGAAAACAGA (402–421) 90011�-HSD2 AGCAGGAGACATGCCGTTTC (659–679) 300
Reverse primer sequence
AT1 CCGGAAGCGATCTTACATAGGTA (638–616) 900AT2 CCATCCAAGCTAGAGCATCCA (712–692) 300A’ogen CTCTTCTAGTTGTCCAAGTACGTA (769–746) 300MR TTTAATGTAGTTCCTCGACTCTTTCTA (539–513) 900GR TACTTGTCTTTACCGTCTGTAAAATAA (552–527) 90011�-HSD2 GCAATGCCAAGGCTGCTT (725–707) 300
a All primers and probes are from 5� to 3� and all concentrations are in nM (in the final reaction).
1019PROGRAMMING OF HYPERTENSION IN SHEEP

primer, 80 nM 18S reverse primer, and the appropriateconcentration of primers and TaqMan® probe for the MR,GR, AT1, AT2, A’ogen, and 11�-HSD2. These concentrationshad been determined in preliminary experiments and areshown in Table 1. Additional preliminary experiments hadshown that there was no difference in CT values when wecompared any of these genes in a nonmultiplex reaction to amultiplex reaction (containing 18S). The amplification effi-ciency of these genes was equal to that of 18S over a range oftemplate concentrations (50 ng to 0.5 pg). cDNA (50 ng) andno reverse transcriptase preparations were amplified at 50°Cfor 2 min and 95°C for 10 min, followed by 40 cycles of 95°Cfor 15 s and 60°C for 1 min.
Calculations for real-time analysis
The CT value (obtained by subtracting the CT value for 18Sfrom the CT value of the gene of interest) of the calibrator wassubtracted from the CT value of each sample to give a CTvalue. The equation of 2�CT was used to obtain a final valuefor each sample relative to the calibrator. Coefficients ofvariation for one sample run five times in one assay were 23%,15%, 40%, 18%, and 23%, respectively, for MR, GR, AT1,A’ogen, and 11�-HSD2. We have used the mean CT value ofthe saline group for any particular gene as the calibrator. Fivealiquots of adult kidney cDNA were also run in each assay todetermine the relative fetal/adult levels of expression of eachgene.
Hybridization histochemistry
A plasmid containing 574 bp of partial ovine MR cDNAsequence was kindly donated by Dr. Anthony Albiston. Afterthe recombinant plasmid was linearized, antisense (T3 Pro-motor) and sense (T7 negative control) riboprobes wereprepared by in vitro transcription using the Promega ribo-probe kit (Promega, Madison, WI), where [-35S] UTP(100CI mmol�1) was incorporated (Bresatec, Thebarton,Australia). The riboprobes were hydrolyzed, precipitated, andresuspended in 10 mM DTT before hybridization histochem-istry. A plasmid containing a 900 bp partial ovine GR cDNAsequence was kindly donated by Dr. G. L. Hammond (28).The same procedure as above was carried out for preparationof the riboprobe for hybridization histochemistry.
The riboprobe was used at a final concentration of 0.02ng/�l in hybridization buffer consisting of 50% deionizedformamide, 20% dextran sulfate, 10% 10� salts (3M Na Cl,100 mM Na2HPO4, pH 6.8, 100 mM Tris-HCl pH 7.5, 50 mMEDTA pH 8.0, 0.2% BSA, 0.2% Ficoll 400, 0.2% polyvinylpyrrolidone), 3.5% tRNA (20 mg/mL), and 1% DTT. Allslides were hybridized in duplicate and sense probes wereused as negative controls. In brief, 5 �m paraffin sectionswere cut and mounted on 2% silanized slides, dried overnightat 37°C, dewaxed, and rehydrated. Sections were prehybrid-ized with Pronase E (125 �g/mL, Sigma, St. Louis, MO) at37°C for 10 min, postfixed in 4% paraformaldehyde for 10min, dehydrated, and air dried. Sections were treated with 80�L of riboprobe, covered with a coverslip, and left overnightat 55°C in a humidified chamber. RNase A digestion (150�g/mL, Sigma-Aldrich) was performed the next day for 2 h at37°C. Slides were dehydrated, air dried and placed on a FujiPhosphor Imaging plate (BASIII) overnight before scanningon a Fujix BAS 2000 scanner to determine possible sites oflabeling. Autoradiography was achieved by dipping slides inliquid emulsion (Ilford, Essex, UK) and leaving the slides atroom temperature for 10 days. Slides were developed for 2min in filtered Kodak D19 developer before fixing in IlfordHypam fixer (1:5 dilution) and staining in H&E.
Statistical analysis
The analysis of differences in gene expression and bloodpressure of the two treatment groups was made by anunpaired t test. Where data were not normally distributed, theMann-Whitney test was used and data presented as median,25% and 75%; otherwise data are presented as mean � se.Statistical analysis was performed using Sigmastat softwarepackage. Statistical significance was set at P � 0.05.
RESULTS
A morphological appearance of the ovine fetus at 28days of gestation
As shown in Fig. 1, at the time of treatment (26–28 daysof gestation) the ovine embryo has only the transientform of the kidney (mesonephros). The neural tube isclosed; by 28 days of gestation the telencephalic vesiclefrom which the hippocampus will form, is present (Fig.1). The metencephalon, which gives rise to the cere-bellum and pons, together with myelencephalon,which gives rise to the medulla oblongata, have formedfrom the rhombencephalon vesicle (Fig. 1).
Effects of prenatal cortisol treatment on bloodpressure in adult female and male lambs
Prenatal cortisol treatment had no effect on growthpatterns in female and male group of animals. Thebirth weights were similar in the two groups of animals(males: 4.5�0.2 kg in saline group vs. 4.8�0.2 kg incortisol group; females: 4.6�0.1 kg in saline group vs.4.3�0.2 kg in cortisol group).
Blood pressure was examined in female lambs at16 � 1 (saline; n�8) and 18 � 1 (cortisol group; n�9)months of age. Body weights were similar between thetwo groups of animals (38�1 kg in saline group vs.37�1 in cortisol group). As shown in Fig. 2, basal meanarterial pressure was significantly higher in the femaleoffspring of sheep treated with prenatal cortisol than infemales in the control group (89�2 mmHg vs. 81�2;P�0.05).
In males, blood pressure was examined when lambswere 17 � 1 (saline; n�8) and 18 � 1 (cortisol group;n�8) months old. Body weights were similar betweenthe two groups of animals (39�1 kg in saline group vs.40�1 in cortisol group). Basal mean arterial pressurewas significantly higher in the offspring of sheeptreated with prenatal cortisol than in the control group(102�4 mmHg vs. 91�3; P�0.05) (Fig. 2). In femaleand male lambs, heart rate was similar between the twogroups of animals (Fig. 2). The basal mean arterialpressure in males from the control group was higherthan basal mean arterial pressure in the respectivegroup of females (91�3 mmHg vs. 81�2; P�0.01).
Effects of cortisol treatment at 130 days of gestationand 2 months of age
Body and placental weights and crown rump lengthsmeasured at 130 days of gestation and in animals 2
1020 Vol. 16 July 2002 DODIC ET AL.The FASEB Journal
months of age were not different between the twotreatment groups. The twin fetuses of ewes treated withsaline weighed 2.8 � 0.1 kg, whereas those from ewestreated with cortisol were 3.1 � 0.1 kg. As shown inTable 2, there was no difference in organ weightsbetween the saline and cortisol fetuses. Volumes andcomposition of fetal fluids (amniotic and allantoic)were similar in all treatment groups. The compositionof fetal urine and plasma (sodium, potassium, chloride,osmolality, urea, creatinine) was similar in the twotreatment groups (data not shown).
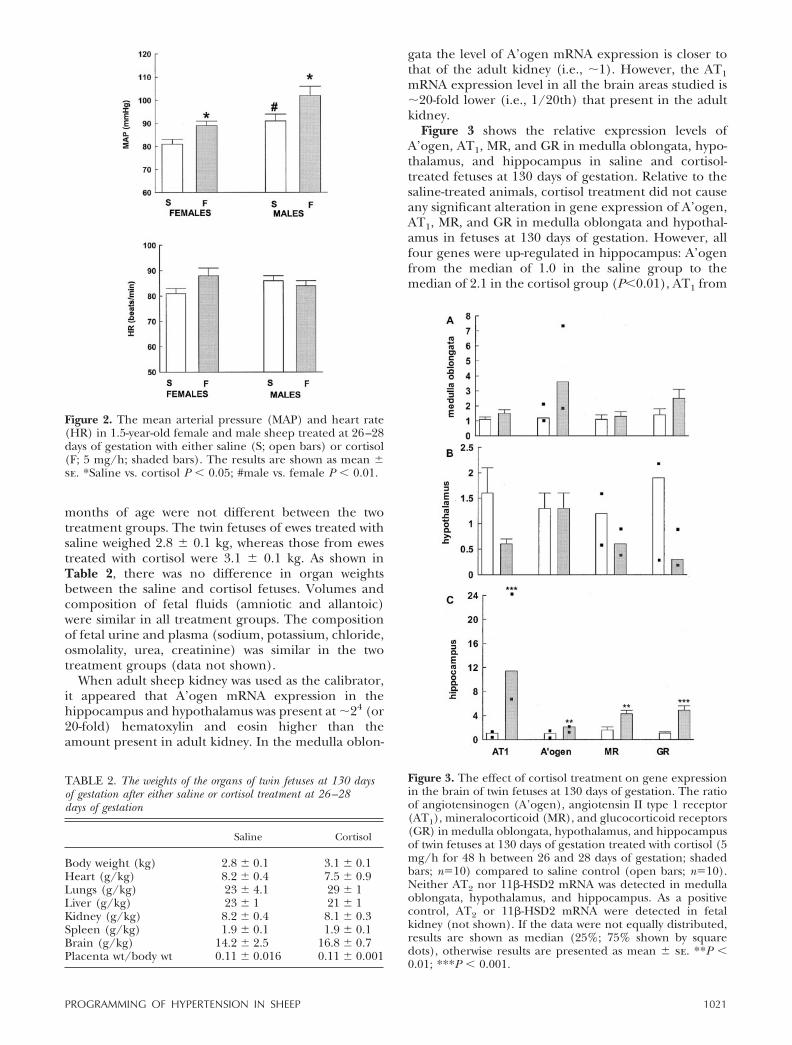
When adult sheep kidney was used as the calibrator,it appeared that A’ogen mRNA expression in thehippocampus and hypothalamus was present at �24 (or20-fold) hematoxylin and eosin higher than theamount present in adult kidney. In the medulla oblon-
gata the level of A’ogen mRNA expression is closer tothat of the adult kidney (i.e., �1). However, the AT1mRNA expression level in all the brain areas studied is�20-fold lower (i.e., 1/20th) that present in the adultkidney.
Figure 3 shows the relative expression levels ofA’ogen, AT1, MR, and GR in medulla oblongata, hypo-thalamus, and hippocampus in saline and cortisol-treated fetuses at 130 days of gestation. Relative to thesaline-treated animals, cortisol treatment did not causeany significant alteration in gene expression of A’ogen,AT1, MR, and GR in medulla oblongata and hypothal-amus in fetuses at 130 days of gestation. However, allfour genes were up-regulated in hippocampus: A’ogenfrom the median of 1.0 in the saline group to themedian of 2.1 in the cortisol group (P�0.01), AT1 from
Figure 3. The effect of cortisol treatment on gene expressionin the brain of twin fetuses at 130 days of gestation. The ratioof angiotensinogen (A’ogen), angiotensin II type 1 receptor(AT1), mineralocorticoid (MR), and glucocorticoid receptors(GR) in medulla oblongata, hypothalamus, and hippocampusof twin fetuses at 130 days of gestation treated with cortisol (5mg/h for 48 h between 26 and 28 days of gestation; shadedbars; n�10) compared to saline control (open bars; n�10).Neither AT2 nor 11�-HSD2 mRNA was detected in medullaoblongata, hypothalamus, and hippocampus. As a positivecontrol, AT2 or 11�-HSD2 mRNA were detected in fetalkidney (not shown). If the data were not equally distributed,results are shown as median (25%; 75% shown by squaredots), otherwise results are presented as mean � se. **P �0.01; ***P � 0.001.
Figure 2. The mean arterial pressure (MAP) and heart rate(HR) in 1.5-year-old female and male sheep treated at 26–28days of gestation with either saline (S; open bars) or cortisol(F; 5 mg/h; shaded bars). The results are shown as mean �se. *Saline vs. cortisol P � 0.05; #male vs. female P � 0.01.
TABLE 2. The weights of the organs of twin fetuses at 130 daysof gestation after either saline or cortisol treatment at 26–28days of gestation
Saline Cortisol
Body weight (kg) 2.8 � 0.1 3.1 � 0.1Heart (g/kg) 8.2 � 0.4 7.5 � 0.9Lungs (g/kg) 23 � 4.1 29 � 1Liver (g/kg) 23 � 1 21 � 1Kidney (g/kg) 8.2 � 0.4 8.1 � 0.3Spleen (g/kg) 1.9 � 0.1 1.9 � 0.1Brain (g/kg) 14.2 � 2.5 16.8 � 0.7Placenta wt/body wt 0.11 � 0.016 0.11 � 0.001
1021PROGRAMMING OF HYPERTENSION IN SHEEP

the median of 1.1 in the saline group to the median of11.4 in the cortisol group (P�0.001), MR from 1.6 �0.5 in the saline group to 4.3 � 0.6 in the cortisol group(P�0.01), and GR from 1.1 � 0.2 in the saline group to4.9 � 0.7 in the cortisol group (P�0.001). As shown inTable 3, these changes in hippocampal gene expres-sion were not seen in animals exposed to cortisol for48 h at 26–28 days of gestation and killed at 2 monthsof age. AT2 was not detected in medulla oblongata,hypothalamus, and hippocampus. Although 11�-HSD2mRNA was not detected in any brain region studied inthree placentas at 28 days, the levels were 39%, 42%,and 100% of that in the adult kidney.
Figure 4 shows the hybridization histochemistry ofhippocampus (CA3 and CA1 regions) for GR and CA3for MR in 130 day ovine fetuses. The CA3 region of thesaline-treated, 130 days of gestation ovine fetus (Fig. 4A,B) shows weak labeling with the GR probe. The sameregion in the cortisol-treated, 130 days of gestationovine fetus (Fig. 4C, D) shows much stronger labeling.The inserts in Fig. 4A, C show no detectible specificlabeling with the sense probe. The CA1 region of thecortisol-treated, 130 days of gestation ovine fetus (Fig.4E, F) shows weaker labeling compared with the CA3region in the same fetus. The MR riboprobe showsstronger labeling in the CA3 region of the cortisol-treated, 130 days of gestation fetus (Fig. 4H) than in thesaline-treated, 130 days of gestation ovine fetus (Fig.4G). The expression of MR and GR was highest in thedentate gyrus � CA3 � CA1 (not shown).
DISCUSSION
The most important finding of this study was thatcortisol, administered to the ewe for only 48 h, at theend of the first month of gestation led to high bloodpressure in the male and female offspring of sheep at1.5 years of age. The significance of this finding lies inthe fact that increased blood pressure was programmed
with high but still physiological levels of the naturallyoccurring glucocorticoid cortisol. The cortisol treat-ment used in this study increased maternal plasmacortisol concentration to levels similar to those shownpreviously (25). Also, this treatment resulted in asignificant increase in maternal glucose concentrationsand suppressed plasma ACTH concentrations to unde-tectable levels (all P�0.05).
Timing of the programming event
We were the first to show that exposure of pregnantewes to pharmacological levels of dexamethasone for
Figure 4. Bright- and dark-field photomicrographs of differ-ent regions of 130 days fetal hippocampus hybridized with theGR (A–F) (magnification �40) and MR (G, H) riboprobes(magnification �100). The CA3 region of the saline-treated,130 days of gestation ovine fetus (A, B) shows weak labelingwith the GR probe. The same region in the cortisol-treated,130 days of gestation ovine fetus (C, D) shows much strongerlabeling. The inserts (A, C) show no detectible specificlabeling with the sense probe. The CA1 region of the cortisol-treated, 130 days of gestation ovine fetus (E, F) shows weakerlabeling than the CA3 region in the same fetus. The MRriboprobe shows stronger labeling in the CA3 region of thecortisol-treated, 130 days of gestation fetus (H) vs. the saline-treated, 130 days of gestation ovine fetus (G).
TABLE 3. The effect of cortisol treatment on gene expressionin the hippocampus in lambs at 2 months of age
Gene Treatment Hippocampus
A’ogen S 1.5 � 0.4F 1.4 � 0.3
AT1 S 1.4 � 0.4F 1.2 � 0.6
MR S 1.1 � 0.2F 0.8 � 0.1
GR S 1.25 (0.7; 1.5)F 0.87 (0.7; 1.5)
a The ratio of angiotensinogen (A’ogen), angiotensin II type 1receptor (AT1), mineralocorticoid (MR), and glucocorticoid recep-tors (GR), relative to calibrator (mean of saline-treated animals) inhippocampus of 2-month-old lambs treated with cortisol (5 mg/h for48 h between 26 and 28 days of gestation; n�5) compared to salinecontrol (n�7). If the data were not equally distributed, the resultsare shown as median (25%; 75%); otherwise results are presented asmean � se.
1022 Vol. 16 July 2002 DODIC ET AL.The FASEB Journal

only 2 days very early in gestation (at a mean age of 27days of the 150 day gestation period) results in hyper-tensive female offspring at 3–4 months of age (19).Subsequently, we demonstrated that this type of hyper-tension amplifies with age and is associated with in-creased cardiac output, left ventricular hypertrophywith reduced cardiac functional reserve, and increasedinsulin sensitivity of the inhibition of lipolysis (20–22).Taken together, it seems that in sheep there is a criticalstage of development, a window, at the end of the firstmonth of gestation when short exposure to eithersynthetic or natural glucocorticoid for only 48 h canprogram high blood pressure in offspring. There issome evidence that disturbance of the intrauterineenvironment, particularly early in pregnancy, can haveprofound effects on the health of the adult. Exposureto malnutrition during the Dutch famine, especiallyduring the first gestational trimester, led to a greateroccurrence of coronary heart disease, an atherogeniclipid profile, and obesity in 50-year-old adults (29–31).An early origin for programming of cardiovasculardisease was also documented in rats (32). Undernutri-tion of the pregnant rat confined to the preimplanta-tion period (0–4.5 days) produced hypertension, butonly in male offspring (32).
Other evidence for programming effects of naturalsteroids on blood pressure
Regardless of the experimental perturbation applied tothe mother causing long-term programming of thefetus, there is a good evidence that one common factorthat may mediate the effect is exposure of the fetus toexcess glucocorticoids (33). Adult rats exposed to alarge dose of carbenoxolone (11�-HSD inhibitor,which blocks placental inactivation of endogenous glu-cocorticoids) throughout gestation were of low birthweight and had high blood pressure, increased basalcorticosterone levels, increased corticotropin-releasinghormone levels, and reduced GR mRNA in the hypo-thalamic paraventricular nucleus (14). However, theseeffects were not reproduced when smaller doses ofcarbenoxolone (still sufficient to elevate maternalblood pressure by �20 mmHg) were administered inrats throughout gestation (34). It is known that themechanism by which carbenoxolone, particularly whenadministered at high doses, increases blood pressure ismore complex than simple inhibition of 11�-HSD(35–37). These studies were based on a proposal thatplacental 11�-HSD2 serves as a mechanism that pro-tects the developing fetus from the excess of maternalglucocorticoids (38). In this study, we found that eventhough placental 11�-HSD2 was present at the levels40%–100% of adult kidney, it was not sufficient to blockthe long-term effects after high but still physiologicallevels of maternal cortisol. The absence of 11�-HSD2from the brain of the late gestation fetus supports thefindings of others (39). Undernutrition during earlygestation in sheep (15% reduction in maternal foodintake) led to a reduced pituitary and adrenal respon-
siveness in late gestation (40), but switched to en-hanced responsiveness postnatally (41). In these ani-mals, exaggerated responsiveness of the HPA axis wasassociated with higher blood pressure (41). However,in our study the effect of cortisol appeared to be timedependent since no change in MR or GR expressionlevels was seen in 2-month-old lambs. We have recentlyreported that a brief exposure to dexamethasone at26–28 days of gestation resulted in hypertensive off-spring, which were allowed to survive for 7 years butshowed no change in MR or GR mRNA in the hip-pocampus or hypothalamus (42). Taken together,these findings suggest that in sheep the HPA may notbe the mechanism whereby prenatal exposure to dexa-methasone programmed hypertension in adult off-spring. More studies are required to prove conclusivelythat such is the case in the adult hypertension pro-grammed by cortisol.
Programming effects of natural steroids on behavior
Only a few studies have demonstrated programming ofthe HPA in rat offspring after prenatal infusions ofeither corticosterone or adrenocorticotrophin (15).However, none of these studies report on the effect ofsuch prenatal stress/steroid exposure on adult bloodpressure. Offspring exposed in utero to repeated loudunanticipated noise (such as experienced by peopleliving under flight paths of busy airports) or living in acountry preparing and ultimately going to war (Arab-Israeli war) showed growth retardation and delays inattaining motor, verbal, and social skills (43). Otherstudies to test the programming hypothesis by naturallyoccurring glucocorticoids studied the effects of prena-tal exposure of adult rats to either restraint stress oruncontrollable electric shocks mainly on HPA axis andbehavioral changes of the offspring (44). Such prena-tally stressed adult rats showed elevated plasma ACTHand corticosterone levels and lower MR and GR densi-ties in the hippocampus (45, 46). Adult rats exposed tocarbenoxolone throughout gestation had increasedbasal corticosterone levels, increased corticotropin-re-leasing hormone levels and reduced GR mRNA in thehypothalamic paraventricular nucleus (14). Dexameth-asone administration only during the last week ofpregnancy led to elevated plasma corticosterone levelsas well as decreasing significantly the MR and GRmRNA in the hippocampus in 16-wk-old rat (17). Inthis study, we showed that brief cortisol exposure at theend of the first month of gestation resulted in highgene expression for MR and GR in hippocampus butnot in the hypothalamus or medulla oblongata of thelate gestational ovine fetus (�4 months after the treat-ment ceased). An increase in glucocorticoid sensitivityto negative feedback and a subsequently decreasedstress reactivity in adulthood are seen in rats handledneonatally (18, 47). Such an effect has been linked toincreased GR and MR in the hippocampus, as well as toincreased GABAA and benzodiazepine receptor levelsin the locus ceruleus and the nucleus tractus solitarius
1023PROGRAMMING OF HYPERTENSION IN SHEEP

of the medulla oblongata. If decreased levels of GR andMR are associated with impaired coping and learning(18), one might propose that the cortisol-pretreatedsheep would have better skills. This remains to betested. However, the transient increase in MR in thehippocampus may have had a significant effect on thedevelopment of the hippocampus, since genetic disrup-tion of the MR but not GR in mice results in hippocam-pal granule cell degeneration (48). At the time oftreatment (26–28 days) in the current experiments, thecentral nervous system of the fetus is relatively under-developed, as illustrated in Fig. 1. The neural tubecloses over in the period from 23 to 26 days, in thesheep, which is comparable to E9.5-E10 in the mouse,and 27–29 days in the human fetus (49). By 28 days, theprimordial plexiform layer contains two cells of differ-ent sizes and occupies about one-third of the wall of thetelencephalon (50). In the posterior lateral wall, thesubventricular zone is beginning to differentiate butthe medial wall remains two-layered. Thus, the hip-pocampal cells that show the changed gene expressionin late gestation are not yet present as such.
The RAS and programming
Another system shown to be programmed by prenatalundernutrition and potentially glucocorticoid expo-sure is the renin-angiotensin system (51). Studies in therat model have shown that early administration of ACEinhibitor captopril 2–4 wk postpartum prevents thedevelopment of hypertension, programmed by intra-uterine exposure to a maternal low-protein diet (52),suggesting these effects may have been mediated bylocal RASs. The renal AT1 receptor expression has beenshown to be increased, at term, in the offspring ofsheep after exposure to undernutrition from day 28 today 77 of gestation (51). There is no information in theliterature concerning the effects that various maternalperturbations might have on the expression of thecomponents of the RAS in the brain. We found thatbrief cortisol exposure at the end of the first month ofgestation resulted in high gene expression for AT1 andA’ogen in hippocampus, but not in hypothalamus ormedulla oblongata of the late gestational ovine fetus(�4 months after the treatment ceased). As it is widelyknown in the literature, Ang II exerts a variety ofactions on the brain including central control of BP,modulation of drinking behavior, salt appetite andsensory functions, effects on memory and learning, andstimulation of pituitary hormone release (53). It isunlikely that high gene expression for AT1 and A’ogenin hippocampus after brief prenatal exposure to corti-sol is related to high blood pressure seen in theseanimals at 1.5 years of age. It is possible the develop-ment of the hippocampus may have been accelerated;when A’ogen is not present in KO mice, there is adecreased density in the granular cell layer (54). In theadult rat, expression of AT1 binding is relatively low inthe hippocampus compared to immature rat brain(55), suggesting a role of Ang II in the maturation of
hippocampal formation (54). In the rat, pharmacolog-ical and anatomical evidence suggests that Ang II playsa role in hippocampal function, memory, and recogni-tion (56, 57). Ang II in the hippocampus is alsospeculated to play a major role in long-term potentia-tion. Studies have shown that Ang II injection above thehippocampus inhibits long-term potentiation in den-tate granule cells in rats (56, 58), which would affectlearning and memory. For MR and GR, this effect ofcortisol appeared to be time dependent since nochange in AT1 and A’ogen in hippocampus was seen in2-month-old lambs. It is possible that even a transientincrease in AT1 and A’ogen in hippocampus couldresult in morphological and functional changes to thehippocampus.
CONCLUSION
This is the first evidence in the literature where shortexposure to the naturally occurring glucocorticoid cor-tisol at the end of the first month of gestation in levelsseen at times of stress programmed high blood pressurein the adult offspring of sheep. Although the effects ongene expression in the hippocampus are probably notrelated to programmed high blood pressure, the effectscould have a significant effect on the development ofthe hippocampus and behavior of the offspring.
This work was supported by a Block Grant from NH&MRC(983001) and a grant in aid from BHP. The Applied Biosys-tems PRISM sequence detector system was purchased withdonations from the Philip Bushell Foundation, the Haroldand Cora Brennen Benevolent Trust, the Viertel Foundation,and the Ramaciotti Foundation. We thank Jehan Jeyaseelanfor help in designing the probes and primers used in real-time PCR.
REFERENCES
1. Barker, D. J. P. (1998) In utero programming of chronicdisease. Clin. Sci. 95, 115–128
2. Huxley, R. R., Shiell, A. W., and Law, C. M. (2000) The role ofsize at birth and postnatal catch-up growth in determiningsystolic blood pressure: a systemic review of the literature.J. Hypertens. 18, 815–831
3. Law, C. M., and Shiell, A. W. (1996) Is blood pressure inverselyrelated to birth weight? The strength of evidence from asystemic review of the literature. J. Hypertens. 14, 935–941
4. Clark, P. M., Hindmarch, P. C., Shiel, A. W., Honour, J. W., andBarker, D. J. P. (1996) Size at birth and adrenocortical functionin childhood. Clin. Endocrinol. 45, 721–726
5. Phillips, D. I. W., Walker, B. R., Reynolds, R. M., Flanagan,D. E. H., Wood, P. J., Osmond, C., Barker, D. J. P., andWhorwood, C. B. (2000) Low birth weight predicts elevatedplasma cortisol concentrations in adults from 3 populations.Hypertension 35, 1301–1306
6. Levitt, N. S., Lambert, E. V., Woods, D. L., Hales, C. N., Andrew,R., and Seckl, J. R. (2000) Impaired glucose tolerance andelevated blood pressure in low birth weight, non obese, youngSouth African adults: early programming of cortisol axis. J. Clin.Endocrinol. Metab. 85, 4611–4618
7. Reynolds, R. M., Walker, B. R., Syddall, H. E., Andrew, R., Wood,P. J., and Whorwood, C. B., Phillips, D. I. W. (2001) Alteredcontrol of cortisol secretion in adult men with low birth weight
1024 Vol. 16 July 2002 DODIC ET AL.The FASEB Journal

and cardiovascular risk factors. J. Clin. Endocrinol. Metab. 86,245–250
8. Konje, J. C., Bell, S. C., Morton, J. J., de Chazal, R., Taylor, D. J(1996) Human fetal kidney morphometry during gestation andthe relationship between weight, kidney morphometry andplasma active renin concentration at birth. Clin. Sci. 91, 169–175
9. Kingdom, J. C., McQueen, J., Connell, J. M., Whittle, M. J.(1993) Fetal angiotensin II levels and vascular (type I) angio-tensin receptors in pregnancies complicated by intrauterinegrowth retardation. Br. J. Obstet. Gynaecol. 100, 476–482
10. Kingdom, J. C., Hayes, M., McQueen, J., Howatson, A. G., andLindop, G. B. (1999) Intrauterine growth restriction is associ-ated with persistent juxtamedullary expression of renin in thefetal kidney. Kidney Int. 55, 424–429
11. Langley-Evans, S. C., Welham, S. J. M., Sherman, R. C., andJackson, A. A. (1996) Weanling rats exposed to maternallow-protein diets during discrete periods of gestation exhibitdiffering severity of hypertension. Clin. Sci. 91, 607–615
12. Woodall, S. M., Johnston, B. M., Breier, B. H., and Gluckman,P. D. (1996) Chronic maternal undernutrition in the rat leads todelayed postnatal growth and elevated blood pressure of off-spring. Pediatric Res. 40, 438–443
13. Lindsay, R. S., Lindsay, R. M., Edwards, C. R. W., and Seckl, J. R.(1996) Inhibition of 11�-hydroxysteroid dehydrogenase inpregnant rats and the programming of blood pressure in theoffspring. Hypertension 27, 1200–1204
14. Welberg, L. A. M., Seckl, J. R., and Holmes, M. C. (2000)Inhibition of 11�-hydroxysteroid dehydrogenase, the fetal pla-cental barrier to maternal glucocorticoids, permanently pro-grams amygdala GR mRNA expression and anxiety-like behav-iour in the offspring. Eur. J. Neurosci. 12, 1047–1054
15. Welberg, L. A. M., and Seckl, J. R. (2001) Prenatal stress,glucocorticoids and the programming of the brain. J. Neuroen-docrinol. 13, 113–128
16. Benediktsson, R., Lindsay, R. S., Noble, J., Seckl, J. R., andEdwards, C. R. W. (1993) Glucocorticoid exposure in utero: newmodel for adult hypertension. Lancet 341, 339–341
17. Levitt, N. S., Lindsay, R. S., Holmes, M. C., and Seckl, J. R.(1996) Dexamethasone in the last week of pregnancy attenuateshippocampal glucocorticoid receptor gene expression and ele-vates blood pressure in the adult offspring in the rat. Neuroen-docrinology 64, 412–418
18. Welberg, L. A. M., Seckl, J. R., and Holmes, M. C. (2001)Prenatal glucocorticoid programming of brain receptors andcorticotrophin-releasing hormone: possible implications for be-haviour. Neuroscience 104, 71–79
19. Dodic, M., May, C. N., Wintour, E. M., and Coghlan, J. P. (1998)An early prenatal exposure to excess glucocorticoid leads tohypertensive offspring in sheep. Clin. Sci. 94, 149–155
20. Dodic, M., Peers, A., Coghlan, J. P., May, C. N., Lumbers, E. R.,Yu Z,.-Y., and Wintour, E. M. (1999) Altered cardiovascularhemodynamics and baroreceptor-heart rate reflex in adultsheep after prenatal exposure to dexamethasone. Clin. Sci. 97,103–109
21. Gatford, K. L., Wintour, E. M., De Blasio, J. J., Owens, J. A., andDodic, M. (2000) Differential timing for programming of glu-cose homeostasis, sensitivity to insulin and blood pressure by inutero exposure to dexamethasone in sheep. Clin. Sci. 98, 553–560
22. Dodic, M., Samuel, C., Moritz, K., Wintour, E. M., Morgan, J.,Grigg, L., and Wong, J. (2001) Impaired cardiac functionalreserve and left ventricular hypertrophy in adult sheep afterprenatal dexamethasone exposure. Circ. Res. 89, 623–629
23. Dodic, M., Baird, R., Hantzis, V., Koukoulas, I., Moritz, K., Peers,A., and Wintour, E. M. (2001) Organs/Systems potentiallyinvolved in one model of programmed hypertension in sheep.Clin. Exp. Pharmacol. Physiol. 28 (11), 952–956
24. Tangalakis, K., Roberts, F. E., and Wintour, E. M. (1992) Thetime-course of ACTH stimulation of cortisol synthesis by theimmature ovine fetal adrenal gland. J. Steroid Biochem. Mol. Biol.42, 527–532
25. Peers, A., Hantzis, V., Dodic, M., Koukoulas, I., Gibson, A.,Baird, R., Salemi, R., and Wintour, E. M. (2001) Functionalglucocorticoid receptors in the mesonephros of the ovine fetus.Kidney Int. 59, 425–433
26. Chomezynski, P., and Saachi, N. (1987) Single step extraction ofRNA using guanidium thiocynate phenol chloroform extrac-tion. Anal. Chem. 162, 156–159
27. Hantzis, V., Albiston, A., Matsacos, D., Wintour, E. M., Peers, A.,Koukoulas, I., Myles, K., Moritz, K., and Dodic, M. (2002)Ontogeny of mineralocorticoid and glucocorticoid receptors inthe developing ovine kidney and the effect of early glucocorti-coid treatment on their expression in the late gestation kidney.Kidney Int. 61, 405–413
28. Young, K., Hammond, G. L., and Challis, J. R. G. (1992) Charac-terization of an ovine glucocorticoid receptor cDNA and develop-mental changes in its mRNA levels in the fetal sheep hypothala-mus, pituitary gland and adrenal. J. Mol. Endocrinol. 8, 173–180
29. Roseboom, T. J., van der Meulen, J. H. P., Osmond, C., Barker,D. J. P., Ravelli, A. C. J., Schroder-Tanaka, J. M., van Montfrans,G. A., Michels, R. P. J., and Bleker, O. P. (2000) Coronary heartdisease after prenatal exposure to the Dutch famine, 1944–45.Heart 84, 595–598
30. Roseboom, T. J., van der Meulen, J. H. P., Osmond, C., Barker,D. J. P., Ravelli, A. C. J., Bleker, O. P. (2000) Plasma lipidprofiles in adults after prenatal exposure to the Dutch famine.Am. J. Clin. Nutr. 72, 1101–1106
31. Ravelli, A. C. J., van der Meulen, J. H. P., Osmond, C., Barker,D. J. P., and Bleker, O. P. (1999) Obesity at the age of 50 y inmen and women exposed to famine prenatally. Am. J. Clin. Nutr.70, 811–816
32. Kwong, W. Y., Wild, A. E., Roberts, P., Willis, A. C., and Fleming,T. P. (2000) Maternal undernutrition during the preimplanta-tion period of rat development causes blastocyst abnormalitiesand programming of postnatal hypertension. Development 127,4195–4202
33. Langley-Evans, S. C. (1997) Hypertension induced by fetalexposure to a maternal low-protein diet, in the rat, is preventedby pharmacological blockade of maternal glucocorticoid synthe-sis. J. Hypertens. 15, 537–544
34. Gomez-Sanchez, E. P., and Gomez-Sanchez, C. E. (1999) Mater-nal hypertension and progeny blood pressure. Hypertension 33,1369–1373
35. Armanini, D., Karbowiak, I., and Funder, J. W. (1983) Affinity ofliquorice derivatives for mineralocorticoid and glucocorticoidreceptors. Clin. Endocrinol. 19, 609–612
36. Walker, B. R., Sang, K. S., Williams, B. C., and Edwards, C. R. W.(1994) Direct and indirect effect of carbenoxolone on re-sponses to glucocorticoids and noradrenaline in rat aorta.J. Hypertens. 12, 33–39
37. Ullian, M. E., Hazen-Martin, D. J., Walsh, L. G., Davda, R. K.,and Egan, B. M. (1996) Carbenoxolone damages endotheliumand enhances vasoconstrictor action in aortic rings. Hypertension27, 1346–1352
38. Seckl, J. R., Cleasby, M., and Nyirenda, M. J. (2000) Glucocor-ticoids, 11�-hydroxysteroid dehydrogenase, and fetal program-ming. Kidney Int. 57, 1412–1417
39. Seckl, J. R. (1997) 11�-hydroxysteroid dehydrogenase in thebrain: a novel regulator of glucocorticoid action? Front. Neuroen-docrinol. 18, 49–59
40. Hawkins, P., Steyn, C., McGarrigle, H. H. G., Saito, T., Ozaki, T.,Stratford, L. L., Noakes, D. E., and Hanson, M. A. (1999) Effectof maternal nutrient restriction in early gestation on develop-ment of the hypothalamic-pituitary-adrenal axis in fetal sheep at0.8–0.9 gestation. J. Endocrinol. 163, 553–561
41. Hoet, J. J., and Hanson, M. A. (1999) Intrauterine nutrition: itsimportance during critical periods for cardiovascular an endo-crine development. J. Physiol. (London) 514, 617–627
42. Dodic, M., Peers, A., Moritz, K., Hantzis, V., and Wintour, E. M.(2002) No evidence for HPA reset in adult sheep with highblood pressure due to short prenatal exposure to dexametha-sone. Am. J. Physiol. 282, R343–R350
43. Weinstock, M., Fride, E., and Hertzberg, R. (1988) Prenatalstress effects on functional development of the offspring. Prog.Brain Res. 73, 319–330
44. Takahashi, L. K. (1998) Prenatal stress: consequences of glu-cocorticoids on hippocampal development and function. Int. J.Dev. Neurosci. 16, 199–207
45. Takahashi, L. K., Baker, E. W., and Kalin, N. H. (1990)Ontogeny of behavioural and hormonal responses to stress inprenatally stressed male rat pups. Physiol. Behav. 47, 357–364
46. Henry, C., Kabbaj, M., Simon, H., Le Moal, M., and Maccari, S.(1994) Prenatal stress increases the hypothalmo-pituitary-adre-nal axis response in young and adult rats. J. Neuroendocrinol. 6,341–345
1025PROGRAMMING OF HYPERTENSION IN SHEEP

47. Meaney, M. J. (2001) Maternal care, gene expression, and thetransmission of individual differences in stress reactivity acrossgenerations. Annu. Rev. Neurosci. 24, 1161–1192
48. Gass, P., Kretz, O., Wolfer, D. P., Berger, S., Tronche, F.,Reichardt, H. M., Kellendonk, C., Lipp, H. P., Schmid, W., andSchutz, G. (2000) Genetic disruption of mineralocorticoidreceptor leads to impaired neurogenesis and granule cell de-generation in the hippocampus of adult mice. EMBO J. 11,447–451
49. Tillet, Y., and Thibault, J. (1987) Early ontogeny of cat-echolaminergic structures in the sheep brain. Anat. Embryol.177, 173–181
50. Reynolds, M. L., and Mollgard, K. (1985) The distribution ofplasma proteins in the cortex and early allocortex of thedeveloping sheep brain. Anat. Embryol. 171, 41–60
51. Whorwood, C. B., Firth, K. M., Budge, H., Symonds, M. E.(2001) Maternal undernutrition during early to mid-gestationprograms tissue specific alterations in the expression of theglucocorticoid receptor, 11�-hydroxysteroid dehydrogenase iso-forms and type-1 angiotensin II receptor in neonatal sheep.Endocrinology 142, 2854–2864
52. Sherman, R. C., Langley-Evans, S. C. (1999) Antihypertensivetreatment in early postnatal life modulates prenatal dietaryinfluences upon blood pressure in the rat. Clin. Sci. 98, 269–275
53. Allen, A. M., McKinley, M. J., Lee, J. H., and Mendelsohn,F. A. O. (2000) Angiotensin II and the central nervoussystem. In Angiotensin II Receptor Antagonists (Epstein, M., andBrunner, H., eds) Hanley and Belfus, pp 129 –148, Philadel-phia
54. Kakinuma, Y., Hama, H., Sugiyama, F., Goto, K., Murakami, K.,and Fukamizu, A. (1997) Anti-apoptotic action of angiotensinfragments to neuronal cells from angiotensinogen knock-outmice. Neurosci. Lett. 232, 167–170
55. Johren, O., and Saavedra, J. M. (1996) Expression of AT1A andAT1B angiotensin II receptor messenger RNA in forebrain of2-wk-old rats. Am. J. Physiol. 271, E104–E112
56. Denny, J. B., Polan-Curtain, J., Wayner, M. J., and Armstrong,D. L. (1991) Angiotensin II blocks hippocampal long-termpotentiation. Brain Res. 567, 321–324
57. Wright, J. W., and Harding, J. W. (1994) Brain angiotensinreceptors subtypes in the control of physiological and behav-ioural responses. Review. Neurosci. Bihav. Rev. 18, 21–53
58. Wayener, M. J., Polan-Curtain, J., and Armstrong, D. L. (1995)Dose and time dependency of angiotensin II inhibition ofhippocampal long-term potentiation. Peptides 16, 1079–1082
Received for publication January 2, 2002.Revised for publication March 27, 2002.
1026 Vol. 16 July 2002 DODIC ET AL.The FASEB Journal