prenatal development of rat primary afferent fibers: i. peripheral projection
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 355589-600 (1995)
Prenatal Development of Rat Primary Afferent Fibers: I. Peripheral Projections
KAROLY MIRNICS AND H. RICHARD KOERBER Department of Neurobiology, University of Pittsburgh, School of Medicine,
Pittsburgh, Pennsylvania 15261
ABSTRACT Development of the peripheral innervation patterns of the Ll-S1 lumbosacral ganglia and
motor segments in embryonic day 12-17 (E12-17) rat embryos was examined using carbocya- nine dyes. Individual dorsal root ganglia (DRGs) and/or isolated ventral horn (VH) segments, or individual peripheral nerves, were isolated in rat embryos fixed at different stages and filled with one of three carbocyanine dyes; DiI, DiA, and DiO. Individual experimental preparations included labeling of 1) single DRGs; 2) multiple DRGs with alternating dyes, DiO, DiI, and DiA; 3) single isolated VH segments; 4) multiple VH segments with alternating dyes; 5) single VH segments and the corresponding segmental DRGs with different dyes; and 6) two or more individual peripheral nerves labeled with different dyes.
Results from these preparations have shown that the first fibers exited the lumbar ventral horn and DRGs at E12. At E l 3 major nerve trunks (e.g., femoral and sciatic) were visible as they exited the plexus region. By E l 4 afferent fibers were present in the epidermis of the proximal hindlimb, and the major nerve trunks extended into the leg. Fibers originating from L3 to L5 (DRG and VH) reached the paw by E14.5-El5, and the epidermis of the most distal toes was innervated by E16-E16.5. While afferent fibers and motor axons of the same segmental origin mixed extensively in the spinal nerve, fibers of different segmental origin combined in the plexus and major nerve trunks with little or no interfascicular mixing.
Dermatomes observed at E l 4 were in general spotty and non-contiguous. However, by E l 6 the dermatomes resembled mature forms with substantial overlap only between adjacent ones. Thus the adult pattern of spatial relationships between cutaneous afferent fibers in the periphery is established early in development. D 1995 Wiley-Liss, Inc.
Indexing terms: dorsal root ganglion, spinal cord, somatotopy, lumbosacral plexus, dermatome
In the adult central nervous system, primary afferent fibers map a precise representation of the body onto second-order neurons in the spinal cord (reviewed in Brown et al., 1992). In addition, primary afferent fibers that innervate different peripheral tissue types and those that innervate different types of peripheral receptors project to characteristic laminar locations within the spinal cord. The developmental processes responsible for this precise map- ping of location and modality have been the subject of numerous investigations. These investigations have exam- ined aspects of both the development of primary afferent fiber projections into the periphery (e.g., Scott, 1982, 1987; Reynolds et al., 1991) as well as their central projections (Smith, 1983; Davis et al., 1989; Woodbury and Scott, 1991; Mendelson et al., 1992). In these studies one consistent finding has been that development of afferent projections in the spinal gray is delayed with respect to the innervation of the hindlimb (Honig, 1982; Scott, 1982; Smith, 1983; Fitzgerald et al., 1991). That is, while the central and peripheral processes of primary afferents begin their growth
simultaneously, peripheral innervation of the hindlimb proceeds while the growth of the central processes is confined within the oval bundle of His (e.g., Honig, 1982; Scott, 1982; Fitzgerald, 1991).
The delay in central ingrowth into the spinal gray has resulted in many hypotheses suggesting that peripheral innervation may be a prerequisite for appropriate central growth. For example, Smith and Frank (1988) suggested that peripheral innervation may be the stimulus for the onset of growth into the gray matter. Subsequently, Fitzger- ald et al. (1991) combined this hypothesis with other findings, such as the fact that proximal hindlimb skin is innervated before distal skin, to suggest that those fibers innervating the proximal hindlimb enter the spinal gray first and occupy the lateral part of the developing dorsal horn. Then, given an orderly progression of the innervation
Accepted October 20, 1994. Address reprint requests to H. Richard Koerber, Department of Neurobiol-
ogy, University of Pittsburgh, School of Medicine, Pittsburgh, PA 15261.
O 1995 WILEY-LISS, INC.

590 K. MIRNICS AND H.R. KOERBER
phosphate buffer (pH = 7.4). Younger embryos (E12-El4) were submerged in 8% paraformaldehyde without perfu- sion. Individual embryos were staged according to crown- rump lengths and external morphological criteria (Hebel and Stromberg, 1986). The perfused embryos were stored for up to 12 months in phosphate-buffered 4% paraformal- dehyde (pH = 7.4) at 4°C.
Individual embryos were placed on a small piece of Sylgard, and a lateral laminectomy was performed exposing the spinal cord and the dorsal root ganglia (DRG) from the lower thoracic to the mid-caudal level. Individual DRGs were separated from the underlying spinal cord and connec- tive tissue by cutting the dorsal roots, elevating the ganglia, and inserting a small piece of aluminum foil beneath the ganglia. The entire preparation was then covered with 19% gelatin (Fisher Scientific) in 0.1 M phosphate buffer
of more distal skin and the corresponding delays of dorsal horn penetration, fibers innervating more distal skin would occupy sequentially more medial locations in the dorsal horn. Such an organizational scheme could obviously result in an initial topographic organization in the presynaptic neuropil of the spinal dorsal horn.
The experiments in the present and companion study (Mirnics and Koerber, 1995) were designed to investigate whether such mechanisms play a role in the formation of the somatotopically organized presynaptic neuropil in the dorsal horn. The initial phase of these studies was to determine the timing and segmental organization of the peripheral innervation of the rat hindlimb. Specifically, what peripheral spatial information is available to primary afferent axons before the central projections penetrate the gray matter?
The development of the segmental sensory and motor innervation of the hindlimb has been studied extensively in the chick (e.g., Landmesser, 1978, 1984; Lance-Jones and Landmesser, 1981; Honig, 1982; Scott, 1982). Although these studies examined many different questions concern- ing the development of hindlimb innervation (e.g., axonal guidance), when taken together they provide an overview which suggests that the adult patterns of segmental inner- vation are established early in development (reviewed in Scott, 1992). The foundation of these early segmental patterns is believed to be established in the region of the lumbar plexus (i.e., crural and sciatic plexuses in chick) as sensory and motor axons from different segmental origins cross over and mix to form major nerve trunks that extend into the developing limb bud (Lance-Jones and Land- messer, 1981; Honig, 1982; Scott, 1982).
The basic organization of developing avian and mamma- lian limbs and their innervation are considered to be similar (reviewed in Scott, 1992). However, due to difficulties inherent in studying prenatal development, relatively little is known about the timing (Reynolds et al., 1991) and the segmental contributions (Wessels et al., 1990) to this innervation in the rat. The recent development of the lipophilic carbocyanine dyes (Honig and Hume, 1986) has afforded us the opportunity to address this question in a unique and comprehensive manner. For example, the use of combinations of different fluorescent dyes (i.e., DiI, DiO, and DiA) allows the visualization of segmental contribu- tions to the innervation of the skin and their interactions at specific times during development. In addition, the segmen- tal contributions to the formation of peripheral nerves can be followed as they course through the region of the lumbar plexus and extend into the hindlimb. Therefore, the experi- ments in the present study were designed to take advantage of these new techniques to determine the timing and organization of segmental innervation of the rat hindlimb with emphasis on the cutaneous innervation. Preliminary results have been reported elsewhere (Mirnics et al., 1992).
MATERIALS AND METHODS Timed-pregnant Sprague-Dawley rats were anesthetized
by a mixture of ketamine (90 mgikg) and xylazine (2 mgikg) i.m. The abdominal cavity was opened and the embryos (E12-El7) were rapidly removed and anesthetized by cooling to +PC. The dam was killed by anesthetic overdose. Cold anesthetized embryos aged E15-El7 were perfused through the heart with 1-2 ml of saline (4°C) followed by 2-5 ml cold (4°C) 4% paraformaldehyde in
Fig. 1. A: Photomicrograph of a whole mount containing multiple dorsal root ganglia (DRGs) (Ll-S1) loaded with carbocyanine dyes DiI, DiA, and DiO in an E l 4 embryo (Ll, L4, S1-DiI; L2, L5-DA; L3, L6-DiO). Color separation was achieved by taking double-exposed photographs under fluorescein isothiocyanate (FITC) and rhodamine fluorescence. The inset indicates the orientation of the embryo (D, dorsal; V, ventral; R, rostral; C, caudal). Note the separate fascicles exiting the DRGs (arrowhead) and the separation of ventral (asterisk) and dorsal (circle) rami. B: Micrograph of a whole mount containing multiple ventral horn (VH) segments loaded alternately with different dyes in an E l 4 embryo (Ll, L4, S1-DiI; L2, L5-DiO; L3, L6-DiA). The exposure and body position are the same as in figure 1A. C: Low-power photomicrograph of a parasagittal section though an E l4 embryo in which multiple VH segments were filled alternately with one of the three dyes (L4-DiO; L2, L5-DiA; L1, L3, L6-DiI). The contributions of the L3-L6 ventral roots to the lumbosacral plexus and peripheral nerves are labeled. The orientation of the embryo is similar to that shown in A (double-exposure under violet and rhodamine excitation). Note the combining of motor fascicles of different segmen- tal origin into the iliohypogastric (ih), ilioinguinal (ii), femoral (0, and sciatic (s) nerve trunks. D: Micrographs of a cross section through an E l 4 spinal nerve (L3) in which the isolated VH segment was loaded with DiI and the corresponding DRG with DiA. This composite figure shows the same field of view under rhodamine (I), violet (II), and rhodamine + violet excitation (111). Note that most fascicles contain both motor and sensory fibers. However, distinct fascicles containing only sensory fibers (arrowheads) are also present. E: Micrograph of a parasagittal section through the lumbosacral plexus of an E 13 embryo in which the L2-L5 DRGs were filled with one of the three dyes. The contributions of the individual dorsal roots are labeled (FITCI rhodamine double exposure). Fascicles of different segmental origin remain separate as they grow through the plexus and form the femoral (D and sciatic (sl nerve trunks. F: Micrograph of a parasagittal section through the lumbosacral plexus region and proximal hindlimb in an E l 4 embryo in which the L2-L5 DRGs were labeled with different dyes. The contributions of the individual dorsal roots are labeled. (FITCI rhodamine double exposure). Note the merging of fascicles into the sciatic (s) and femoral (0 nerve trunks. G Micrograph of a parasagittal section through the lumbosacral plexus region of an E l 4 embryo in which the L2-L6 VH segments were labeled. The contribution of each dorsal root is labeled separately. f, femoral; s, sciatic; g, genitofemoral; arrowhead identifies a small bundle of fibers connecting the genitofemo- ral and femoral nerves (violetirhodamine double exposure; orientation as shown in A). H: Micrograph of a more distal section through the limb of the same E l 4 embryo shown in G. Note the small concise L2 contribution (yellow) in the femoral nerve trunk (arrowhead). I: Micrograph of longitudinal section through an E 14 hindlimb displaying a peripheral branch point of the femoral nerve. The L3 DRG was labeled with DiA and the L4 DRG with DiI (FITCirhodamine double exposure). Note that fascicle separation is maintained as the nerve extends into the limb. J Photomicrograph of a section through an E l 4 hindlimb showing the femoral nerve in cross section at a similar proximodistal location as in I. The L2-L4 VH segments were labeled with different dyes (violetirhodamine double exposure). Scale bars = 250 pm in A-C, 75 pm in D, 100 pm in E, I, and J, 200 km in F-H.

PRENATAL HINDLIMB INNERVATION 591
Figure 7

592 K. MIRNICS AND H.R. KOERBER
(pH = 7.4) at 37°C. The gelatin was quickly hardened by cooling to 4"C, and individual ganglia were exposed by removing the overlying gelatin. Crystals of the dyes or in some cases dyes dissolved in grease (see below) were then applied to the surface of the ganglia and, when possible, inserted into the ganglia (Fig. 1A); a small drop of 70% ethanol was applied to the ganglia to help dissolve the dyes. The entire preparation was then covered with gelatin and stored in 4% paraformaldehyde a t 37°C.
The dyes were occasionally dissolved in grease by taking a small amount ( - 10 mg) of DiI C18 or DiA C18 (Molecular Probes) which was crushed to dust and mixed with 25 mg of silicon grease (Thomas Scientific-"Lubriseal"). Three ali- quots (25 pl) of absolute ethanol were applied separately to the mixture during vigorous stirring. The dye grease prod- uct was stored at -25°C. It was found that this mixture is very easy to manipulate and that this technique increased the diffusion rate by approximately 50% over application of crystals or the dyes dissolved in alcohol.
Individual spinal segments were isolated by first exposing the dorsal root ganglia, cutting the dorsal roots, and reflecting the DRGs. This allowed the visualization of the ventral roots. Small strips of aluminum foil were then inserted into the spinal cord at the segmental boundaries. The dorsal one-third to one-half of the spinal cord was removed, and the preparation was covered with gelatin. Once the gelatin was cooled, the isolated ventral horn segments were exposed, and the different dyes were applied directly to the cut surface (dorsal) of the individual seg- ments (Fig. 1B). The entire preparation was then covered with gelatin and stored in 4% paraformaldehyde at 37°C. A third preparation involved labeling the primary afferents of a given spinal nerve with one dye and the motor axons of the same segment with a different dye. This was accom- plished by first labeling an isolated ventral horn (VH) segment exactly as described above and then freeing the appropriate ganglia from the gelatin and loading it, also as described above.
In another series of experiments individual peripheral nerves were labeled with different dyes in the periphery to determine the distribution of cell somata and fibers in different DRGs. This labeling was accomplished by isolating the individual nerves from surrounding tissue, cutting the nerves, and placing them on a small piece of foil. The rest of the preparation was then covered with gelatin to avoid leakage into surrounding tissue. The individual nerves were labeled by applying dyes directly to the proximal stump of the severed nerves. In these preparations it was found that applying the dyes dissolved in grease resulted in better labeling than applying crystals alone.
Working with fixed tissue allowed us to use several different experimental preparations which were useful in addressing different aspects of the developmental process. These preparations included labeling of 1) single DRGs; 2) multiple DRGs (Ll-Sl) with alternating DiO, DiI, and DiA, 3) single ventral horn segments; 4) multiple isolated VH segments (Ll-S1) with alternating dyes; 5) single isolated VH segments and the corresponding segmental DRGs with different dyes (DiI and DiA); 6) individual peripheral nerves with two different dyes (DiI and DiA).
The stored preparations were examined frequently under fluorescent illumination ( 4 ~ ) to record the progress of the dyes as they diffused through the axonal membranes (Mendelson et al., 1992). Examples of the resolution seen in these whole mounts are provided in Figure 2A-C. After
diffusion was judged to be complete, individual prepara- tions were cooled to +4"C. The filled ganglia and/or VH segments were removed, and the limb with the attached pelvic region was dissected and mounted for sagittal section- ing in 19% gelatin. The embedded preparation was placed in phosphate-buffered (pH = 7.4) 8% paraformaldehyde and left at 4°C for 18-24 hours before sectioning. This fixation stiffened the gelatin, which improved sectioning on the Vibratome.
Serial sagittal vibratome sections (50 pm) of the limb and body were collected and mounted on slides in cold 0.1 M phosphate buffer (pH = 7.4). An example of the orientation of these sections can be seen in the insert in Figure 1A and for labeled motor fibers in Figure 1C. In those preparations in which peripheral nerves were labeled the individual ganglia were first photographed and videotaped as whole mounts then removed and sectioned (50-150 pm) using a vibratome. The fluorescent images of all serial sections (4 X, l ox , 20x1 were recorded on videotape and/or photo- graphed. In order to get a better color separation in the color slide films the multiple dye-loaded preparations were double exposed under different excitation wavelengths and using different barrier filters (see Table 1). The stored videotape images of the serial sections were used later to reconstruct the dermatomal map and the branching pat- terns of peripheral nerves.
Separation of fluorescent dyes Separation of these fluorescent dyes was not straightfor-
ward. Excitation and emission wavelengths originally re- ported for the dyes dissolved in methanol varied consider- ably from that observed when they were dissolved in axonal membranes. However, the appropriate excitation wave- lengths and barrier filters were identified, and adequate separation of the individual dyes was achieved. Settings and equipment (Olympus) are summarized in Table 1. It should be noted that the emission wavelengths did vary slightly depending on the batch of dye, the solvent employed, the transport media, and the age of the embryo.
RESULTS A total of 80 embryos was used to label different neuronal
elements with different combinations of dyes. In order to be included in the analysis individual preparations had to meet the following criteria: 1) all elements of the structure were labeled; 2) there were no signs of leakage between adjacent roots; 3) diffusion was judged to be complete. Of the original 80 preparations, 48 met these criteria. Of these, 18 were in preparations in which DRGs were labeled, 14 in which ventral horn segments were labeled, and 16 in which both the dorsal roots and ventral horn segments were labeled. In an additional 8 embryos individual peripheral nerves were adequately labeled.
The amount of time necessary for acceptable diffusion of the dyes varied considerably (12 hours to >60 days) depending upon several variables. The most consistent variable was the age of the embryos, as all three dyes diffused much more rapidly in younger embryos. Other variables included 1) size of the embryo; 2) quality of perfusion; 3) differences in individual batches of dyes; 4) concentration of dye achieved in the loading process. Al- though these parameters were identified and controlled for, it was still very difficult to predict the amount of time
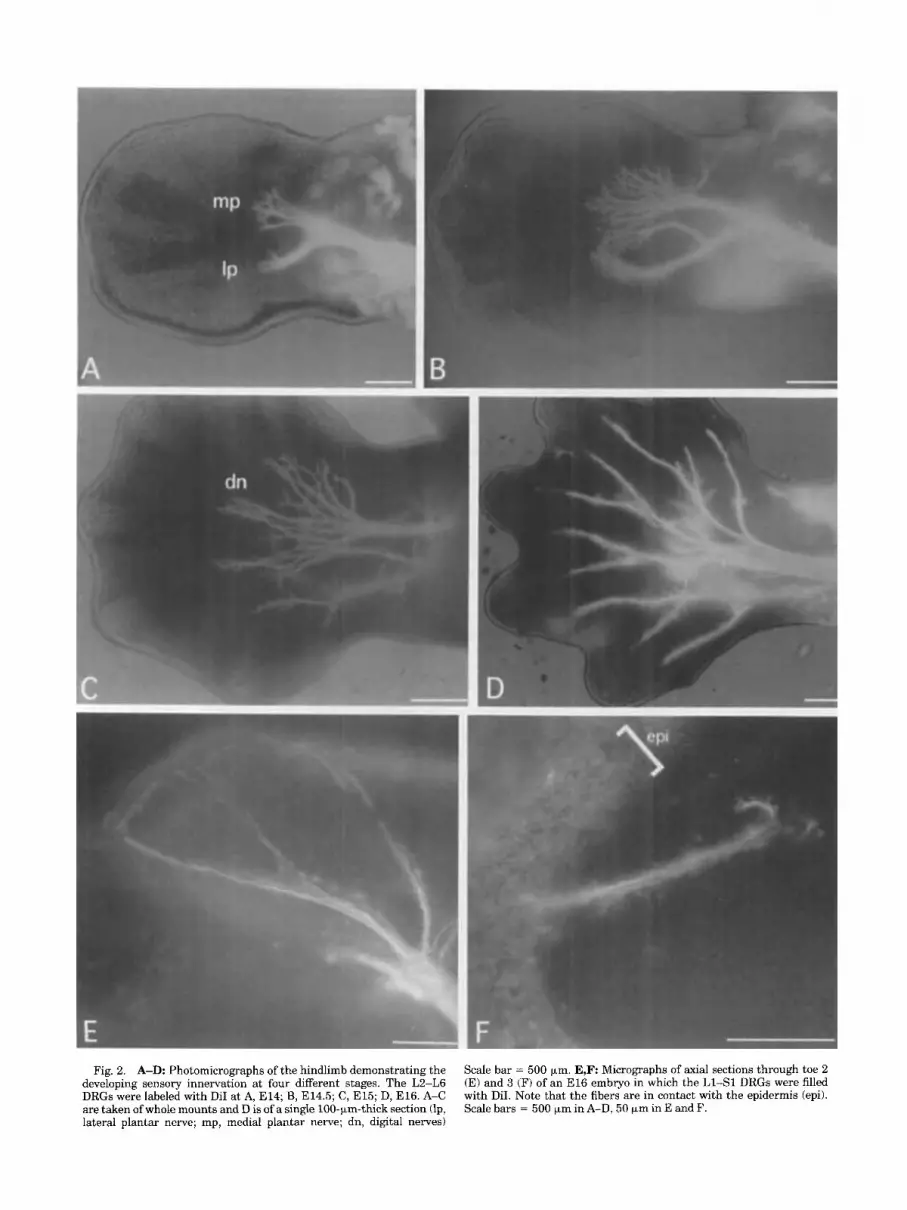
Fig. 2. A-D: Photomicrographs of the hindlimb demonstrating the developing sensory innervation at four different stages. The L2-L6 DRGs were labeled with DiI at A, E14; B, E14.5; C, E15; D, E16. A-C are taken of whole mounts and D is of a single lOO-)*rn-thick section (lp, lateral plantar nerve; mp, medial plantar nerve; dn, digital nerves)
Scale bar = 500 km. E,F Micrographs of axial sections through toe 2 (E) and 3 (F) of an El6 embryo in which the L1-S1 DRGs were filled with DiI. Note that the fibers are in contact with the epidermis (epi). Scale bars = 500 +m in A-D, 50 km in E and F.

594 K. MIRNICS AND H.R. KOERBER
TABLE 1.
Dichroic Excitation Barrier Visible dye Blank dye mirror filter (nm) filter (nm)
DiI (dark red) DiA and DiO DM545 BP545 & 546DF10 R610 (rhodamine) Passes 545-550 Passes 610 & t
DiA (dark green) DiI DM490 BP490 530DF15 (FIT0 Passes 410490 Passes 530 ? 7.5
DiO (dark green) DiI and DiA DM455 BP405 0510 (violet) Passes 390-410 Passes510 61 1
DiI (light red) None DM490 RP490 0515 DiA (yellow) (FIT0 Passes 410-490 Passes 515 & T DiO (green)
necessary for complete diffusion. Therefore, frequent moni- toring of the diffusion of the dyes was always necessary.
Control for leakage of dyes The preparations were closely monitored to determine if
there was any leakage of the dyes into adjacent ventral horn segments, DRGs, or peripheral nerves. These determina- tions were very straightforward, and any preparations in which leakage occurred were excluded from analysis. How- ever, there were also concerns about other possible sources of leakage. For example, axons growing together through the plexus may come in contact, suggesting the possibility that the dyes would transfer from one to the other. In order to assure that such leakage did not occur, the unlabeled component was always left intact. For example, when the DRGs were loaded the ventral roots were left intact and vice versa. The unlabeled tissue was also embedded, sectioned, and viewed. There were never any signs of such leakage even after 60 days at 37°C.
Initial outgrowth The first motor axons were observed leaving lumbar
ventral horn segments at E12. The growth cones of these fibers displayed a very complicated morphology with numer- ous swellings and complex filopodia (Tosney and Land- messer, 1985b). Although the onset of this initial out- growth was delayed relative to the onset of such growth in cervical and thoracic segments, it began simultaneously at all lumbar levels. Within 12 hours (E12.5) large numbers of fibers were visible leaving the ventral spinal cord. Out- growth from the lumbar ganglia also began at E12, and by E12.5-El3 the dorsal and ventral rami of the spinal nerves were formed.
Formation of spinal nerves and lumbar plexus The formation of individual spinal nerves was visualized
at these early stages (E12-El3) in those preparations in which the DRGs and isolated ventral horns of the same segmental level were labeled with different dyes. Overall there was a large amount of mixing of sensory and motor fibers, as numerous mixed fascicles (sensory/motor) were observed proximal to the plexus region (Fig. 1D). However, several fascicles containing only sensory fibers were also visible in the spinal nerves at this proximodistal location (Fig. 1D). In addition, fascicles devoid of motor axons were observed at all successive proximodistal levels through the plexus and into the limb.
In the rostral lumbar levels the ventral rami of the spinal nerves quickly split and joined with fascicles from adjacent spinal nerves to form axial and proximal peripheral nerves such as the ilioinguinal and iliohypogastric (Fig. 1C). Both the sensory and motor contributions to the lumbar plexus from all levels were in place by E13. There was a very
characteristic organization of the plexus with L2, L3, and L4 spinal nerves contributing to the femoral nerve trunk and elements from L4, L5, and L6 joining to form the sciatic nerve trunk (Fig. 1E-G). However, occasionally a small branch from L3 spinal nerve also contributed to the sciatic (Fig. 1F). The relative contributions from the different spinal nerves to the major nerve trunks varied as expected due to pre- and post-fixation of the plexus (Romanes, 1951). For example, in some preparations most of the ventral ramus of the L4 spinal nerve was included in the femoral nerve trunk, whereas in others a majority of L4 was included in the sciatic nerve trunk. In these cases the contributions from other segments to these major nerve trunks were shifted accordingly.
Although there was merging of fiber fascicles from several segmental levels in the formation of these major nerve trunks, this process was quite different in appearance from that seen in the formation of individual spinal nerves. In contrast to the impressive mixing of individual fibers of different modality (motor vs. sensory) into fascicles seen in the formation of spinal nerves (Fig. lD), contributions from the different segmental levels apparently remained in sepa- rate fascicles which joined and formed the major nerve trunks (Fig. 1E-G). The result of this joining of fascicles was visible in sections taken through the nerve trunks in the proximal hindlimb (Fig. 1H). In addition, the fascicles which joined to form the individual nerve trunks appar- ently did so on a nearest-neighbor basis. For example, fascicles from the caudal part of the L4 ventral rami did not cross over more rostral fascicles to join the femoral nerve trunk. This finding was consistent for both sensory and motor fibers in all the animals and ages studied. However, when these fibers were viewed in sections cut in the same plane as the plexus (Fig. 1F) a few fascicles of different segmental origin were observed which did cross. It should be noted that examination of serial sections consistently revealed that these fascicles had separated in the dorsoven- tral axis, before they crossed paths.
Segregation of segmental components persisted as the nerves grew further into the hindlimb. At peripheral branch points in the limb, fascicles from different segmental levels joined with others to form individual nerves much as they had in the plexus (Fig. 11,J). In Figure 11 it appears that there was some mixing as a small strip of yellow can be seen between the bundles of green and red fluorescing fibers. However, this was due to the fact that fibers from one segmental origin were running slightly behind the other, and this overlap of red and green fluorescing fibers pro- duced the yellow color in the photomicrographs. Separation of fascicles in the peripheral nerves was also obvious in cross sections taken a t a similar location in the limb (Fig. 1 J). Although these fascicles apparently contained fibers from one segmental level, given our level of resolution, it was difficult to determine whether individual fibers joined different fascicles at these peripheral branch points (Tosney and Landmesser, 1985a,c). However, if present, they were few in number.
In general the early pattern of growth of these fibers was similar to that described previously in the chick (reviewed in Scott, 1992) and for GAP 43 expression in the rat (Reynolds et al., 1991). However, there were some differ- ences in the timing of certain events. Unlike the chick (Landmesser, 1978), motor and sensory fibers simulta- neously exited the ventral horn and the DRGs. In addition, fibers labeled with these dyes in the rat did not pause in the

PRENATAL HINDLIMB INNERVATION 595
plexus region as reported in the chick (Landmesser, 1978) and for GAP 43 expression in the rat (Reynolds et al., 1991), at least when viewed at 12-hour intervals. Although this may reflect differences in organizational strategies between the different species, it could also simply reflect a more rapid process in mammals.
By E l 4 the growing fibers extended well into the leg (Fig. 2A) and formed medial and lateral plantar nerves. Half a day later these nerve trunks penetrated well into the foot (Fig. 2B). At E l 5 fibers were in the skin over the ankle and the proximal toes. In addition, individual digital nerves were also visible at this stage (Fig. 2C). By E l 6 the medial and lateral branches of the individual digital nerves were clearly visible (Fig. 2D), and the first fibers were in the epidermis of the tips of the most distal toes (Fig. 2E,F).
Overall the formation of the various nerve trunks in the plexus appeared to be very precise. However, there were apparently some adjustments made in the periphery, as small fascicles of fibers were seen which changed their course after the plexus to join other peripheral nerves. An example of this apparent course correction can be seen in Figure lG, where a small fascicle of motor axons leaves the genitofemoral nerve trunk and returns to the femoral nerve.
Development of dermatomes Individual cutaneous nerves were visualized distal to the
lumbosacral plexus by E14. These nerves were readily visible when dorsal and ventral roots of the same segment were labeled with different dyes. By E14.5-El5 the cutane- ous fibers were in axial and proximal hindlimb skin, and many of these fibers were visible in the epidermis (Fig. 3A-D). Once the fibers penetrated the epidermis the fibers branched out rapidly into the surrounding skin. As these fibers penetrated the epidermis they often encircled particu- lar epidermal cells. These cells were often labeled with dye, suggesting intimate contact between the growing cutane- ous afferent fibers and these cells. Innervation of the rest of the hindlimb skin proceeded rapidly as fibers reached the tips of the most distal toes by E16-16.5. Although innerva- tion of the tips of the toes was initiated at this stage, the entire epidermis of the distal toes was not innervated until a day later (E17).
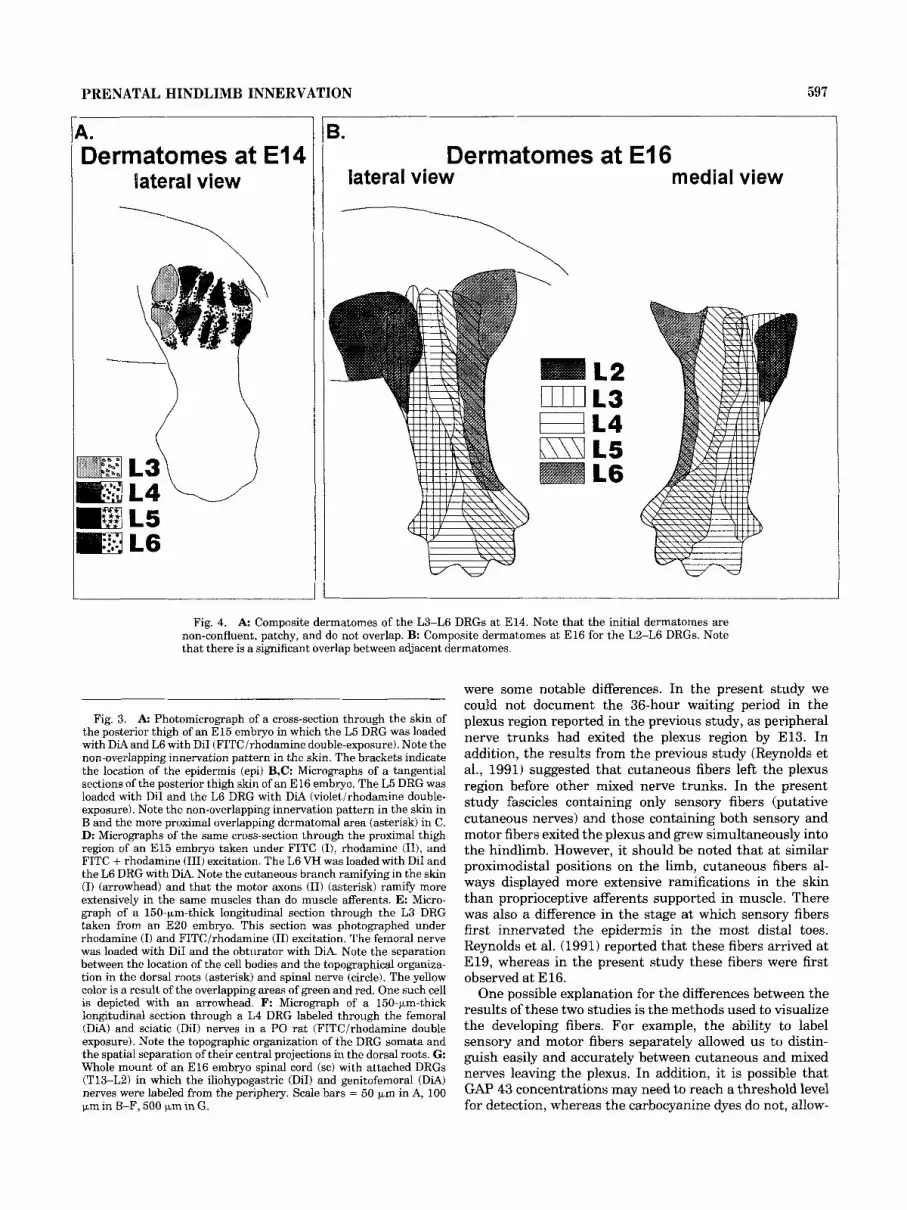
The initial dermatomes (E14-15) in the proximal limb were patchy and mostly non-contiguous (Fig. 4A). By E l 6 there was an increase in the density of innervation of axial and proximal limb skin, and the ramifications of the fibers were more complex. Gaps between dermatomes were filled in, and areas of overlap between adjacent dermatomes were observed (Fig. 4B). At later stages (>E17), the peripheral patterns were not easily reconstructed for technical rea- sons, including l) the increased diffusion distance; 2) the inability to visualize the innervation patterns in whole mounts; and 3) difficulties in obtaining good serial vibra- tome sections of the older limbs. Although individual dermatomes were difficult to visualize in older embryos, the proximal parts of individual dermatomes were easily viewed in E l 6 embryos. Then by using segmental contributions to the digital nerves as templates the distal parts of the dermatomes were estimated and composite representations of the dermatomes were constructed (Fig. 4B).
As mentioned above, the labeling of the dorsal and ventral roots of the same spinal segment allowed the identification of individual cutaneous or muscle nerve fascicles very early in development (E14). Once these
fascicles were formed they apparently grew as a unit through the plexus. Observed at the level of light micros- copy, fascicles containing only sensory fibers (cutaneous nerve trunks) and those containing both sensory and motor fibers exited the plexus simultaneously and proceeded into the hindlimb in unison. Within individual mixed nerve trunks sensory and motor axons also penetrated the hind- limb in unison. However, once reaching their final target, the motor axons clearly developed more extensive and complex ramifications within the muscles than the corre- sponding afferent fibers (Fig. 3D).
Topographic organization of dorsal root ganglia
The early fasciculation of fibers during the formation of spinal nerves and the conservation of these fascicles through the plexus and into the distal parts of the hindlimb suggested a high degree of segregation of fibers at this level. Evidence for topographic organization at the level of the DRGs in adults has been difficult to obtain (e.g., McLachlan and Janig, 1983; Ygge, 1984). However, other investigators have reported some degree of topology (e.g., Burton and McFarlane, 1973; Kausz and Rethelyi, 1985). Likewise, developmental studies have also yielded different results, with Wessels et al. (1990) reporting evidence in support of a topographic organization in lumbar ganglia and Smith (1983) reporting a lack of organization in thoracic ganglia.
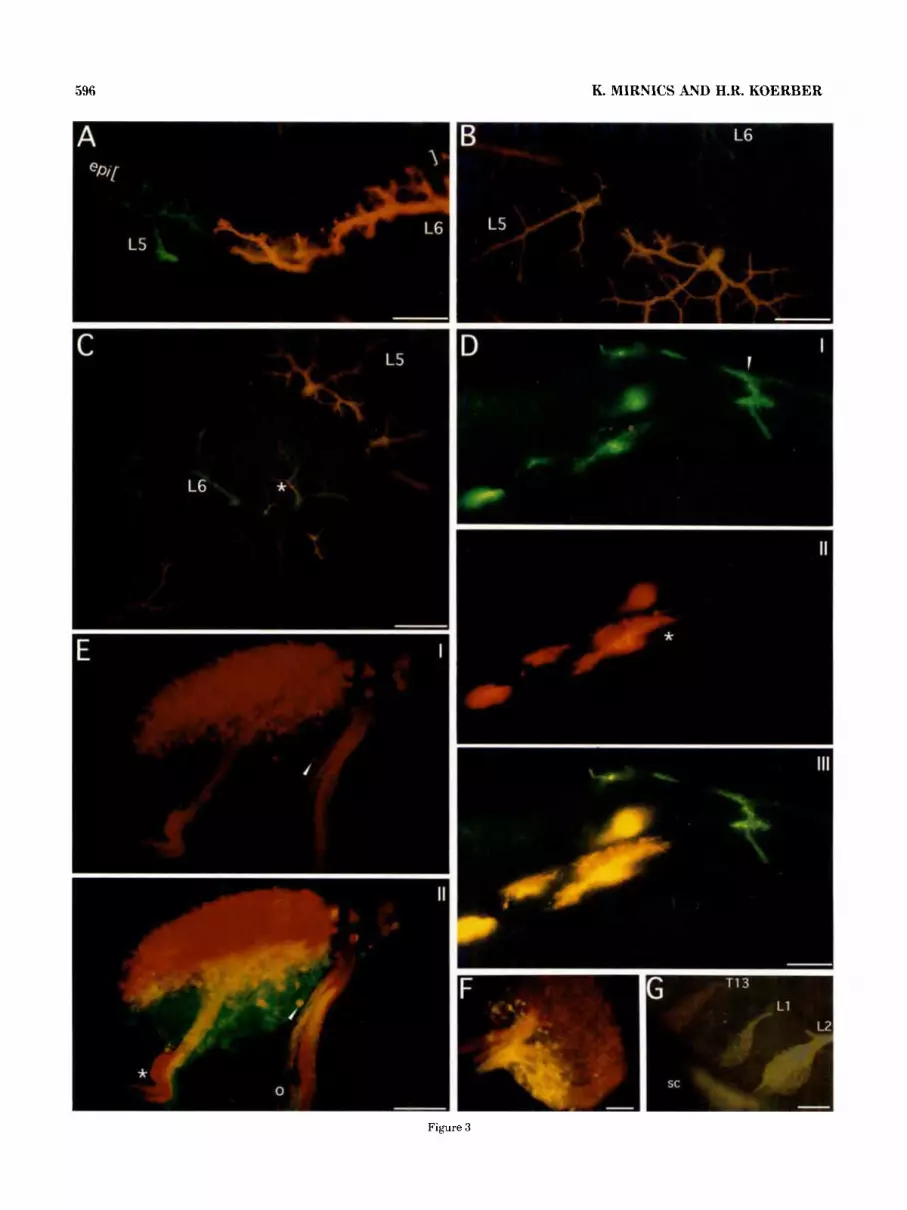
The ability to use different dyes in the same preparations has given us the opportunity to address this question with a high degree of precision. Examples of the results obtained with such preparations can be seen in Figure 3E-G. There was a clear separation of the cell bodies of primary afferents whose fibers grouped together to form different peripheral nerves. This segregation was maintained in both the central and peripheral projections of the afferent fibers (Fig. 3E,F). I t is important to note that although this separation of bundles of fibers was difficult to photograph, under micro- scopic examination the separation was quite distinct. These findings suggest that the destination and possibly the specificity (e.g., muscle or cutaneous) of primary afferents may be decided very early in development during the formation of spinal nerves.
DISCUSSION The present studies were designed to establish the devel-
opmental timing and organization of the segmental innerva- tion in the rat hindlimb. In a study of this nature the accurate staging of the embryos was of obvious importance. As stated in Materials and Methods we employed the staging criteria proposed by Hebel and Stromberg (19861, and in both this study and the companion paper the staging agreed with previous reports for the timing of the initial outgrowth (e.g., Reynolds et al., 1991) and spinal cord development (Altman and Bayer, 1984). However, due to the fact that individual embryos were usually staged to the nearest half day and that small variations within litters were observed, all dates reported here should be viewed as having an error of f % day.
The only previous report on the development of periph- eral innervation of the hindlimb in the rat described the timing and distribution of GAP 43 expression (Reynolds et al., 1991). Although the results of the present study generally agreed with the GAP 43 study (e.g., initial outgrowth from lumbar segments begins a t E12), there
596 K. MIRNICS AND H.R. KOERBER
Figure 3
PRENATAL HINDLIMB INNERVATION 597
L Dermatomes at El4
lateral view
3. Dermatomes at El6
lateral view medial view
L2 rn L3
Fig. 4. A: Composite dermatomes of the L3-L6 DRGs at E14. Note that the initial dermatornes are non-confluent, patchy, and do not overlap. B: Composite dermatomes at E l6 for the L2-L6 DRGs. Note that there is a significant overlap between adjacent dermatomes.
Fig. 3. A: Photomicrograph of a cross-section through the slun of the posterior thigh of an E l 5 embryo in which the L5 DRG was loaded with DiA and L6 with DiI (FITCirhodamine double-exposure). Note the non-overlapping innervation pattern in the skin. The brackets indicate the location of the epidermis (epi) B,C: Micrographs of a tangential sections of the posterior thigh skin of an E l 6 embryo. The L5 DRG was loaded with DiI and the L6 DRG with DiA (violetirhodamine douhle- exposure). Note the non-overlapping innervation pattern in the skin in B and the more proximal overlapping dermatomal area (asterisk) in C. D: Micrographs of the same cross-section through the proximal thigh region of an E l 5 embryo taken under FITC (I), rhodamine (II), and FITC + rhodamine (111) excitation. The L6 VH was loaded with DiI and the L6 DRG with DiA. Note the cutaneous branch ramifying in the skin (I) (arrowhead) and that the motor axons (11) (asterisk) ramify more extensively in the same muscles than do muscle afferents. E: Micro- graph of a 150-pm-thick longitudinal section through the L3 DRG taken from an E20 embryo. This section was photographed under rhodamine (I) and FITC/rhodamine (11) excitation. The femoral nerve was loaded with DiI and the obturator with DiA. Note the separation between the location of the cell bodies and the topographical organiza- tion in the dorsal roots (asterisk) and spinal nerve (circle). The yellow color is a result of the overlapping areas of green and red. One such cell is depicted with an arrowhead. F: Micrograph of a 150-pm-thick longitudinal section through a L4 DRG labeled through the femoral (DiA) and sciatic (DiI) nerves in a PO rat (FITCirhodamine double exposure). Note the topographic organization of the DRG somata and the spatial separation of their central projections in the dorsal roots. G: Whole mount of an E l6 embryo spinal cord (sc) with attached DRGs (T13-LZ) in which the iliohypogastric (DiI) and genitofemoral (DiA) nerves were labeled from the periphery. Scale bars = 50 Fm in A, 100 Vrn in B-F, 500 pm in G.
were some notable differences. In the present study we could not document the 36-hour waiting period in the plexus region reported in the previous study, as peripheral nerve trunks had exited the plexus region by E13. In addition, the results from the previous study (Reynolds et al., 1991) suggested that cutaneous fibers left the plexus region before other mixed nerve trunks. In the present study fascicles containing only sensory fibers (putative cutaneous nerves) and those containing both sensory and motor fibers exited the plexus and grew simultaneously into the hindlimb. However, it should be noted that at similar proximodistal positions on the limb, cutaneous fibers al- ways displayed more extensive ramifications in the skin than proprioceptive afferents supported in muscle. There was also a difference in the stage at which sensory fibers first innervated the epidermis in the most distal toes. Reynolds et al. (1991) reported that these fibers arrived at E19, whereas in the present study these fibers were first observed at E16.
One possible explanation for the differences between the results of these two studies is the methods used to visualize the developing fibers. For example, the ability to label sensory and motor fibers separately allowed us to distin- guish easily and accurately between cutaneous and mixed nerves leaving the plexus. In addition, it is possible that GAP 43 concentrations may need to reach a threshold level for detection, whereas the carbocyanine dyes do not, allow-

598
ing for the visualization of small numbers of fibers and their ramifications in the epidermis.
Segregation of segmental components One of the most striking results of this study was the
clear separation of fiber fascicles from different segmental origins in the plexus region and the maintenance of this separation within peripheral nerves in the more distal hindlimb. The orderly patterns formed by adjacent fascicles from different segmental levels are consistent with an organizational scheme based on general guidance cues and the spatiotemporal relationships of growing fibers. For example, fascicular (segmental) components may be select- ing which peripheral nerve trunk to enter based entirely on general guidance cues, i.e., areas permissive or inhibitory to growth (Tosney and Oakley, 1990; Oakley and Tosney, 19911, or mechanical limitations, i.e., how many fascicles can the pathway handle and/or competition between fibers vying for the same permissive paths.
A similar organizational scheme was described by Horder (1978). He theorized that a combination of contact guid- ance, differential growth, and limb rotation could account for the observed patterns of motor innervation. In fact Horder’s theoretical model has accurately predicted many of the results of the present study. For example, his model predicted that fibers (motor or sensory) would retain their segmental (topographic) relationships within peripheral nerve trunks.
The fact that the segmental segregation of fibers extends into the distal hindlimb would suggest that the initial formation of fascicles in the spinal nerves may greatly impact the resulting topographic organization. This initial formation of fascicles could also be accomplished by a relatively simple mechanical process based on nearest neighbors and numbers of fibers which will fasciculate together. This possibility is also supported by the striking topographic organization within the DRG. Interanimal variation of topographic patterns observed in individual ganglia and dorsal roots would then depend o n variations in the spatial relationship of the growing fibers and the developing limb bud (e.g., pre- and post-fixation in the plexus region).
The maintenance of this topography in the dorsal roots suggests that these spatial relationships are maintained in the central projections (e.g. Fig. 3F,G). This implies that fibers which travel together in the periphery also project together centrally. This arrangement could obviously influ- ence the topographic organization of growth in the spinal cord (see Mirnics and Koerber, 1995, this issue).
Taken together, the present results, and their correspon- dence with Horder’s theoretical model and recent studies describing the peripheral distributions of molecules shown to be either inhibitory to growth (e.g., Oakley and Tosney, 1991) or growth promoting (e.g. Davies, 1994) suggest a compatible picture of the plexus region. That is, this region could be viewed as a three-dimensional framework of permissive openings which lead into specific permissive pathways or conduits into the developing limb. Fibers from different segmental levels may simply enter the first appro- priate permissive path they encounter.
This reliance on general peripheral guidance cues would seem to contradict the earlier studies in which it was reported that fibers from one segment cross over other fibers to mix and form different nerve trunks (e.g., Honig, 1982). However, as shown above, fascicles that crossed in our preparations (e.g., Fig. 1F) traveled in different dorso-
K. MIRNICS AND H.R. KOERBER
ventral planes within the limb. Therefore, it is possible that the fascicles of fibers cross because they are already follow- ing permissive paths into the periphery which eventually cross, owing to continued differential growth and subse- quent limb rotation (see Horder, 1978).
Although our results are in agreement with such a model, it should be noted that other studies involving small neural tube and neural crest reversals (Honig et al., 1986; Scott, 1986) have produced results which are not always consis- tent with this hypothesis. For example, in some cases displaced sensory neurons are capable of innervating periph- eral targets appropriate for their original locations (Honig et al., 1986; Scott, 1986). Thus, although general guidance cues are sufficient to describe the patterns observed in normal development, we cannot rule out the possibility that additional specific cues may also have a significant role.
Although the relatively simple model described above can account for many aspects of the spatial organization of hindlimb innervation it does not address modality segrega- tion (e.g., muscle vs. cutaneous). For example, purely cutaneous nerves exit the plexus region (e.g., lateral femo- ral cutaneous) suggesting that they have chosen a specific pathway to the skin. Possible explanations for this level of modality sorting have been suggested. For example, cutane- ous afferent fibers may have the ability to select “cutane- ous” pathways based on trophic factors produced by the skin (Dell and Munger, 1986; Tosney and Hageman, 1989). Other evidence strongly suggests that motor axons play an instrumental role in selecting the appropriate pathways for the formation of muscle nerves (Honig et al., 1986; Land- messer and Honig, 1986; Scott, 1988). Interestingly, the latter explanation also receives support from studies in adults where regenerating motor fibers apparently retain the ability to select appropriate paths back to muscle (Brushart, 1988).
The results of the present study provide evidence consis- tent with both of these assertions. For example, cutaneous nerves (sensory only) can be observed early in development. In addition, although large numbers of afferent fibers mix intimately with motor axons in the spinal nerve, pure sensory fascicles (Fig. 1D) are visible as early as E l 3 proximal to the plexus region. The fact that some afferent fibers remain separate from motor axons at these locations suggests the possibility that these fascicles of sensory fibers are destined to enter cutaneous pathways in the plexus region. On the other hand, those in intimate contact with motor axons remain with them to form muscle nerves.
Initial dermatomes The embryonic dermatomes presented in this study are,
in general, similar to those described in the adult rat and in various other species (e.g., Kuhn, 1953; Kow and Pfaf, 1975; Brown and Koerber, 1978; Scott, 1982). This result is in agreement with the finding that the adult patterns of innervation are established early in chick development (reviewed in Scott, 1992). These embryonic dermatomes are also in register with the central projections patterns for individual dorsal roots presented in the companion study (Mirnics and Koerber, 1995). For example, whereas the central projections of adjacent dorsal roots show substan- tial overlap in the dorsal horn, there is little or no overlap in the projections of non-adjacent dorsal roots (Mirnics and Koerber, 1995). Likewise, in this study only the derma- tomes of adjacent dorsal roots show substantial overlap. This pattern of overlap is also consistent with that seen in

PRENATAL HINDLIMB INNERVATION
the adult rat (Kow and Pfaff, 1975) and cat (Brown and Koerber, 1978).
The fact that fibers from adjacent segmental levels do not overlap during the initial innervation of the skin is in agreement with classic Golgi studies (e.g., Ramon y Cajal, 1911) which describes a lack of competition between cutane- ous afferents. This lack of direct interaction of cutaneous fibers could be due to the abundance of non-innervated skin available. Then once all available skin has been innervated the existing fibers, or new ones just arriving in the skin, begin to compete in earnest for available sites resulting in dermatomal overlap. However, one restriction to this com- petition appears to be between fibers contained within different cutaneous nerves as their innervation fields do not overlap in the periphery (Koerber and Brown, 1980 and 1982; Woolf and Fitzgerald, 1986).
Another possibility is that significant competition does not occur and that cutaneous fibers innervate skin solely on the basis of which ones arrive first. This would predict that the initial dermatomes with spotty innervation patterns are actually maintained in the adult. For example, the overlap- ping areas depicted in composite dermatomal drawings may be incorrect in that the areas of overlap between derma- tomes are actually interdigitations of cutaneous fibers from different segmental origins and possibly islands of fibers from one dorsal root surrounded by those from another. However, overlap should be present to some extent at least at the border regions given the fact that some cutaneous receptors can be multiply innervated (i.e., hair follicles) by fibers projecting over more than one dorsal root (Munger, 1983).
In summary, the results of these experiments using multiple dyes present a unique view of the normal segmen- tal innervation of the developing hindlimb. In addition these results give us a comprehensive description of the timing of this innervation. This information will be instru- mental in determining the level of peripheral spatial infor- mation available to primary afferent fibers at different developmental stages as they locate their appropriate cen- tral targets.
599
ACKNOWLEDGMENTS We thank Drs. Cynthia Lance-Jones and Paul B. Brown
for helpful comments and suggestions on this manuscript and Sara Glickstein and Randy Adolph for excellent techni- cal assistance. This work was supported by NIH grant R01 NS23725 (H.R.K.).
LITERATURE CITED Altman, J., and S.A. Bayer (1984) The development of the rat spinal cord.
Adv. Anat. Embryol. Cell Biol. 85:l-164. Brown, P.B., and H.R. Koerber (1978) Cat hindlimb tactile dermatomes
determined with single unit recordings. J. Neurophysiol. 41.260-267. Brown, P.B., H.R. Koerber, and L.A. Ritz (1992) Somatotopic organization of
primary afferent projections to the spinal cord. In S.A. Scott (ed): Sensory Neurons: Diversity, Development and Plasticity. London: Ox- ford University Press, pp. 116130.
Brushart, T.M.E. (1988) Preferential reinnervation of motor nerves by regenerating motor axons. J. Neurosci. 8r1026-1031.
Burton, H., and J.J. McFarlane (1973) The organization of the seventh lumbar spinal ganglion of the cat. J. Comp. Neurol. 149r215-232.
Davies, A.M. (1994) Tracking neurotrophin function. Nature 368r193-194. Davis, B.M., E. Frank, F.A. Johnson, and S.A. Scott (1989) Development of
central projections of lumbosacral sensory neurons in the chick. J. Comp. Neurol. 279:556-566.
Dell, D.A., and B.L. Munger (1986) The early embryogenesis of papillary (sweat duct) ridges in primate glabrous skin: the derrnatotopic map of cutaneous mechanoreceptors and dermatoglyphics. J. Comp. Neurol. 244t511-532.
Fitzgerald, M. (1991) A physiological study of the prenatal development of cutaneous sensory inputs to dorsal horn cells in the rat. J. Physiol. (Lond.) 432473482.
Fitzgerald, M., M.L. Reynolds, and L.I. Benowitz (1991) GAP-43 expression in the developing rat lumbar spinal cord. Neuroscience 41r187-199.
Hebel, R., and M.W. Stromberg (1986) Anatomy and Embryology of the Laboratory Rat. Worthsee: BioMed Verlag Worthsee.
Honig, M.G. (1982) The development of sensory projection patterns in embryonic chick hind limb. J. Physiol. (Lond.) 33Or175-202.
Honig, M.G., and R. Hume (1986) Fluorescent carbocyanine dyesallow living neurons of identified origin to be studied in long-term cultures. J. Cell. Biol. 103: 171-187.
Honig, M.G., C. Lance-Jones, and L.T. Landmesser (1986) The development of sensory projection patterns in embryonic chick hindlimbs under experimental conditions. Dev. Biol. 118:532-548.
Horder, T.J. (1978) Functional adaptability and morphogenetic opportun- ism, the only rules for limb development? Zoon 6r181-192.
Kausz M., and M. Rethelyi (1985) Lamellar arrangement of neuronal somata in the dorsal root ganglion in the cat. Somatosens. Res. 2193-204.
Koerber, H.R., and P.B. Brown (1980) Projections of two hindlimb nerves to cat dorsal horn. J. Neurophysiol. 44:259-269.
Koerber, H.R., and P.B. Brown (1982) Somatotopic organization of hindlimb cutaneous nerve projections to cat dorsal horn. J. Neurophysiol. 48:481- 489.
Kow, L.M., and D.W. Pfaff (1975) Dorsal root recording relevant for mating reflexes in female rats: identification of receptive fields and effects of peripheral denervation. J. Neurobiol. 6:23-37.
Kuhn, R.A. (1953) Organization of tactile dermatomes in cat and monkey. J. Neurophysiol. 16: 169-182.
Lance-Jones, C., and L.T. Landmesser (1981) Pathway selection by chick lumbosacral motoneurons during normal development. Proc. R. SOC. Lond. [Biol.] 214r19-52.
Landmesser, L.T. (1978) The development of motor projection pattern in the chick hind limb. J. Physiol. (Lond.) 284:391-414.
Landmesser, L.T. (1984) The development of specific motor pathways in the chick embryo. TINS 7:336-339.
Landmesser, L.T., and M.G. Honig (1986) Altered sensory projections in the chick hind limb following early removal of motoneurons. Dev. Biol. 11 8r5 11-531.
McLachlan, F.M., and W. Janig (1983) The cell bodies of origin of sympa- thetic and sensory axons in some skin and muscle nerve of the cat hindlimb. J. Comp. Neurol. 214r115-130.
Mendelson, B., H.R. Koerber, and E. Frank (1992) Development of cutane- ous and proprioceptive afferent projections in the chick spinal cord. Neurosci. Lett. 138:72-76.
Mirnics, K., and H.R. Koerher (1995) Prenatal development of rat primary afferent fibers: 11. Central projections. J. Comp. Neurol. 335:601-614.
Mirnics, K., R.H. Adolph, and H.R. Koerber (1992) Earlydevelopment of the segmental innervation of the rat hindlimb. SOC. Neurosci. Abstr. 18:609.
Munger, B.L. (1983) Multiple segmental innervation of single primate hairs and rete ridges. Anat. Rec. 205r137.
Oakley, R.A., and K.W. Tosney (1991) Peanut agglutinin and chondroitin-6- sulfate are molecular markers for tissues that act as barriers to axon advance in the avian embryo. Dev. Biol. 147:187-206.
Ramon y Cajal, S. (1911) Histologie du Systkme Nerveux de l'Homme et des Vertebbres, Vol 1. Paris: A. Maloine.
Reynolds, M.L., M. Fitzgerald, and L.I. Benowitz (1991) GAP-43 expression in developing cutaneous and muscle nerves in the rat hindlimb. Neurosci- ence41:201-211.
Romanes, G.J. (1951) The motor cell columns of the lumbosacral cord of the cat. J. Comp. Neurol. 94:313-364.
Scott, S.A. (1982) The development of the segmental pattern of skin sensory innervation in embryonic chick hindlimb. J. Physiol. (Lond.) 330r203- 220.
Scott, S.A. (1986) Skin sensory innervation patterns in embryonic chick hindlimb following dorsal root ganglion reversals. J. Neurobiol. 173349- 668.
Scott, S.A. (1987) The development of skin sensory innervation patterns. TINS lOr468-473.

600 K. MIRNICS AND H.R. KOERBER
Scott, S.A. (1988) Skin sensory innervation patterns in embryonic chick hindlimbs deprived of motoneurons. Dev. Biol. 126:362--374.
Scott, S.A. (1992) The development of peripheral sensory innervation patterns. In S.A. Scott (ed): Sensory Neurons: Diversity, Development and Plasticity. London: Oxford University Press, pp. 242-263.
Smith, C.L. (1983) Development and postnatal organization of primary afferent projections to the rat thoracic spinal cord. J. Comp. Neurol. 220:29-43.
Smith, C.L., and E. Frank (1988) Specific sensory projections to the spinal cord during development in bullfrogs. J. Comp. Neurol. 269;96-108.
Tosney, K.W., and M.S. Hageman (1989) Different subsets of axonal guidance cues are essential for sensory neurite outgrowth to cutaneous and muscle targets in the dorsal ramus of the embryonic chick. J. Exp. 2001.251:232-244.
Tosney, K.W., and L.T. Landmesser (1985a) Growth cone morphology and trajectory in the lumbosacral region of the chick embryo. J. Neurosci. 5.2345-2358.
Tosney, K.W., and L.T. Landmesser (198513) Specificity of early motoneuron growth cone outgrowth in the chick embryo. J. Neurosci. 5.2336-2344.
Tosney, K.W., and L.T. Landmesser (1985~) Development of the major pathways for neurite outgrowth in the chick hindlimb. Dev. Biol. 109: 193-2 14.
Tosney, K.W., and R.A. Oakley 11990) The perinotochordal mesenchyme acts as a barrier to axon advance in the chick embryo: implications for a general mechanism of axonal guidance. Exp. Neurol. 109:7549.
Wessels, W.J.T., H.K.P. Feirabend, and E. Marani (1990) Evidence for a rostrocaudal organization in dorsal root ganglia during development as demonstrated by intra-uterine WGA-HRP injections into the hindlimb of rat fetuses. Dev. Brain Res. 54.273-281.
Woodbury, C.J., and S.A. Scott (1991) Somatotopic organization of hindlimb skin sensory inputs to the dorsal horn of hatchling chicks (GulEus g. domesticus). J. Comp. Neurol. 314,237-256.
Woolf, C.J., and M. Fitzgerald (1986) Somatotopic organization of cutaneous afferent terminals and dorsal horn neuronal receptive fields in the superficial and deep laminae of the rat lumbar spinal cord. J. Comp. Neurol. 2515 17-53 1.
Ygge, J. (1984) On the organization of the thoracic spinal ganglion and nerve in the rat. Dev. Brain Res. 55:395-401.