predicted climate-driven bird distribution changes and forecasted conservation conflicts in a...
TRANSCRIPT

Contributed Paper
Predicted Climate-Driven Bird Distribution Changesand Forecasted Conservation Conflicts in aNeotropical SavannaMIGUEL ANGELO MARINI,∗ MORGANE BARBET-MASSIN,† LEONARDO ESTEVES LOPES,‡AND FREDERIC JIGUET†∗Departamento de Zoologia, IB, Universidade de Brasılia, 70910-900, Brasılia, DF, Brazil, email [email protected]†CRBPO, UMR 5173 MNHN-CNRS-UPMC Conservation des Especes, Restauration et Suivi des Populations, 55 rue Buffon, CP 51,75005 Paris, France‡Laboratorio de Ornitologia, Departamento de Zoologia, ICB, Universidade Federal de Minas Gerais, 31270-910, Belo Horizonte,MG, Brazil
Abstract: Climate-change scenarios project significant temperature changes for most of South America. We
studied the potential impacts of predicted climate-driven change on the distribution and conservation of 26
broad-range birds from South America Cerrado biome (a savanna that also encompass tracts of grasslands
and forests). We used 12 temperature or precipitation-related bioclimatic variables, nine niche modeling
techniques, three general circulation models, and two climate scenarios (for 2030, 2065, 2099) for each species
to model distribution ranges. To reach a consensus scenario, we used an ensemble-forecasting approach to
obtain an average distribution for each species at each time interval. We estimated the range extent and
shift of each species. Changes in range size varied across species and according to habitat dependency;
future predicted range extent was negatively correlated with current predicted range extent in all scenarios.
Evolution of range size under full or null dispersal scenarios varied among species from a 5% increase to an
80% decrease. The mean expected range shifts under null and full-dispersal scenarios were 175 and 200 km,
respectively (range 15–399 km), and the shift was usually toward southeastern Brazil. We predicted larger
range contractions and longer range shifts for forest- and grassland-dependent species than for savanna-
dependent birds. A negative correlation between current range extent and predicted range loss revealed that
geographically restricted species may face stronger threat and become even rarer. The predicted southeasterly
direction of range changes is cause for concern because ranges are predicted to shift to the most developed and
populated region of Brazil. Also, southeastern Brazil is the least likely region to contain significant dispersal
corridors, to allow expansion of Cerrado vegetation types, or to accommodate creation of new reserves.
Keywords: BIOMOD, Cerrado, climate change, conservation planning, ecological-niche modeling, ensembleforecasting, maximum entropy
Cambios Previstos en la Distribucion de Aves Debido al Cambio Climatico y Pronostico de Conflictos de Conser-vacion en una Sabana Neotropical
Resumen: Los escenarios del cambio climatico proyectan cambios significativos en la temperatura de casi
toda America del Sur. Estudiamos los impactos potenciales de cambios –debidos al cambio climatico – previstos
en la distribucion y conservacion de 26 especies de aves de amplia distribucion en el bioma Cerrado de America
del Sur (una sabana que tambien comprende extensiones de campos y bosques). Utilizamos 12 variables
bioclimaticas relacionadas con la temperatura y la precipitacion, nueve tecnicas de modelado de nicho, tres
modelos generales de circulacion y dos escenarios climaticos (para 2030, 2065 y 2099) para cada especie
para modelar sus rangos de distribucion. Para alcanzar un consenso de escenario, utilizamos un metodo
Paper submitted January 20, 2009; revised manuscript accepted February 5, 2009.
1558Conservation Biology, Volume 23, No. 6, 1558–1567C©2009 Society for Conservation BiologyDOI: 10.1111/j.1523-1739.2009.01258.x

Marini et al. 1559
de pronostico de ensamble para obtener una distribucion promedio para cada especie en cada intervalo de
tiempo. Estimamos la extension y el cambio de rango en cada especie. Los cambios en el area de distribucion
variaron en las especies en funcion de la dependencia de habitat; el rango de distribucion futuro pronosticado
se correlaciono negativamente con el rango actual en todos los escenarios. La evolucion del tamano del rango
bajo escenarios de dispersion total o nula vario entre especies desde un 5% de incremento hasta 80% de
decremento. Los cambios promedio esperados bajos escenarios de dispersion nula y total fueron 175 y 200
km, respectivamente (rango 15 a 399 km), y los cambios generalmente fueron hacia el sureste de Brasil.
Pronosticamos mayores contracciones de rango y cambios de rango mas largos para especies dependientes
de bosque y de campos que para las aves dependientes de la sabana. Una correlacion negativa entre el rango
actual y la perdida pronosticada revelo que una especie geograficamente restringida puede enfrentar mayor
amenaza y volverse aun mas rara. La direccion sureste pronosticada en los cambios de rango es motivo de
preocupacion porque se pronostico que los rangos cambiaran hacia la region mas desarrollada y poblada
de Brasil. Tambien, el sureste de Brasil es la region que contiene menos corredores de dispersion, como para
permitir la expansion de los tipos de vegetacion del Cerrado, o acomodar la creacion de nuevas reservas.
Palabras Clave: BioMod, cambio climatico, Cerrado, entropıa maxima, modelado de nicho ecologico, planifi-cacion de la conservacion, pronostico de ensamble
Introduction
Climate-change scenarios project significant temperaturechanges for most of South America. Mean predictions ofthree general circulation models (GCMs) from the In-tergovernmental Panel on Climate Change (IPCC 2007)project an increase in temperature of as much as 3.5 ◦C(16.4%) and a small increase in precipitation of 2% acrossSouth America between the 1960–1990 period and the2080–2099 period. Despite high diversity and endemism,few researchers have considered the potential impacts ofclimate change on Neotropical (e.g., Pounds et al. 1999;Peterson et al. 2001) or Brazilian (Siqueira & Peterson2003; Anciaes & Peterson 2006; Nunes et al. 2007) biodi-versity. Major declines in potential distributional areas arepredicted for 162 tree species from the Cerrado (savanna-like vegetation) region (Siqueira & Peterson 2003). Abird study predicts that present-day distributions of abouthalf the manakins (Pipridae) will decrease over 80% andthat Cerrado bird diversity may be severely threatened(Anciaes & Peterson 2006).
The Cerrado is the largest tropical savanna in the worldand the second-largest vegetation formation in SouthAmerica, and it occurs primarily in Brazil (Ratter et al.1997). It is a biodiversity hotspot and is already classifiedas one of the most threatened biomes on Earth (Mitter-meier et al. 1999; Myers et al. 2000; Silva & Bates 2002).Its vegetation varies considerably in physiognomy, fromtreeless grasslands to tall savanna woodland and galleryforests along watercourses (Eiten 1972, 1978). Biologicaldiversity is high, with plant endemism reaching 44% of10,000 species. Among birds, 30 of the 856 species listedfor the biome are endemic (Silva 1995, 1997; Silva &Bates 2002; Silva & Santos 2005). Currently, the Cerradois one of the regions of the world suffering the most fromanthropogenic impacts (Dias 1990; Ratter et al. 1997),with only 2.2% of its area under legal protection (Klink
& Machado 2005). Because recent estimates indicate thatonly 40–50% of its original vegetation remains (Machadoet al. 2004; BRASIL 2007; Brannstrom et al. 2008) andwhat is left is highly fragmented, there are limited op-portunities for large-scale species range shifts, as in otherhotspots such as Madagascar (Hannah et al. 2008).
The ecological-niche modeling approach is based onthe principle that one can model species’ ecologicalniches given the assumption that distribution of knownlocalities reflects species’ environmental preferences.This principle is supported by the results of studies thatshow the evolutionary conservatism of ecological niches(Peterson et al. 1999). There is one conceptual frame-work of niche modeling but several mathematical ap-proaches for modeling distributions (Elith et al. 2006;Heikkinen et al. 2006) that may yield very different pre-dictions (e.g., Thuiller 2004; Araujo et al. 2005b; Pearsonet al. 2006). The use of multiple models within an en-semble forecasting framework is one solution to accountfor model variability, and this approach has been used inclimatic change studies (e.g., Thuiller 2004; Araujo et al.2005a, 2005b).
Models of the effects of climate change on tropical or-ganisms are few but necessary, especially in biodiversityhotspots (Peterson et al. 2005). Bioclimatic niche modelsare useful for predicting the impacts of climate changeon species distribution (Pearson & Dawson 2003). Niche-based modeling has been used in reserve planning(Wilson et al. 2005), prediction of biological invasions(Broennimann & Guisan 2008), and identification of un-known distributional areas of rare species (Guisan et al.2006). Such assessments are critical in rapidly develop-ing landscapes (Pressey et al. 2007), where conservationopportunities become limited as natural lands are con-verted to agriculture or urban development. Proper man-agement of the matrix is necessary in such areas (Fon-seca et al. 2005). In addition, models can increase the
Conservation Biology
Volume 23, No. 6, 2009

1560 Climate-Driven Changes in Cerrado Birds
cost-effectiveness of conservation plans, which is criticalin regions with limited resources, such as in the Cerradoof South America.
In conservation planning it is important to evaluateinteractions between climate and other factors, such asland-use change (Sala et al. 2000; Laurance & Williamson2001) and to avoid conservation inferences based solelyon bioclimatic models (Hampe 2004; Thuiller 2004). Fu-ture distributions may differ from the predictions of bio-climatic models for several reasons. Species’ inherenttraits, such as dispersal ability, may limit changes in theirgeographical occurrence. The vegetation on which thespecies depend may not disperse to the new geographicareas or may disperse at a different rate. Biotic interac-tions, such as competition, predation, and parasitism,may also change and help or preclude the realizationof range shift (Araujo & Luoto 2007; Preston et al. 2008).Furthermore, the predicted future geographical range,suitable on the basis of climatic conditions, may alreadybe occupied and converted by anthropogenic activities,such as plantations or urban development.
We studied the potential impacts of predicted climate-driven changes on the distribution and conservation of26 endemic Cerrado bird species with broad ranges. Weexamined predicted future geographical distributions inlight of current and projected land use, developmentrates, and distribution of reserves so as to provide sup-port for conservation planning. We worked under the as-sumption of temporal constancy in the bioclimatic nicheof each species, even though some species may adapt tonew climatic conditions.
Methods
We modeled the bioclimatic niches of 26 bird species en-demic to the Cerrado. To decrease the impact of modelvariability with a consensus method, we used nine dif-ferent niche-models, three climate models, and two cli-mate scenarios for each species. With this approach wemaximized the predictive accuracy of species distribu-tions and the reliability of future potential distributions.We used Maxent (Phillips et al. 2006) and eight mod-eling techniques in the BIOMOD computational frame-work under R application (Thuiller 2003) in an ensemble-forecasting approach (use of multiple models) to reach aconsensus scenario (Thuiller 2003; Marmion et al. 2009;see Supporting Information). Models were evaluated witha relative operating characteristic (ROC) curve and areaunder the curve (AUC) (see Supporting Information).
To explore the distributional response to climatechange, we characterized every modeled distribution(current and future) by its range and position (latitudeand longitude of the range centroid). We estimated theratio of range extents (full dispersal hypothesis) and range
overlaps (null dispersal hypothesis) for predicted presentand future ranges for each species and the distance anddirection of range shift of present and future range cen-troids. Under the full dispersal hypothesis, we assumedspecies would be able to move through the landscapewithout physiological or environmental impediments,which is a reasonable hypothesis for birds. Under thenull dispersal hypothesis, we assumed species would notdisperse at all. We modeled both dispersal hypotheses forall 26 species for three reasons. First, Neotropical birdshave different dispersal abilities (e.g., Andrade & Marini2001; van Houtan et al. 2007). Second, the ability of these26 species to disperse through natural and altered land-scapes is still poorly studied and understood. Third, byconsidering both hypotheses we provide a range of best-and worst-case scenarios within which species responsesmust lie. The knowledge of the likelihood of movementof organisms is needed to improve the use of models(Peterson et al. 2005).
Species Occurrence Localities
To determine occurrence points for the 26 species, weused three data sources as detailed in Lopes (2008). First,we collated unpublished field records of L.E.L. and sev-eral other experienced observers. Second, we revieweda large literature, scanning papers listed in bibliographiccompilations primarily for the names of the species fromBrazil, Paraguay, and Bolivia. Third, we collated data fromornithological specimens deposited in seven Brazilianmuseums and the Museum National d’Histoire Naturelle(Paris) and from on-line databases of Brazilian and NorthAmerican museums. We obtained geographical coordi-nates of specimen collection locations from the originalsources, Paynter and Traylor (1991), or Vanzolini (1992).To reduce modeling errors associated with small sam-ple sizes, we excluded from analyses species with fewerthan 20 spatially independent localities (1◦ × 1◦ latitude× longitude grid cells). We plotted species data in 100× 100 km2 (1◦ × 1◦) grid cells, removing duplicate pres-ence records within a cell. The final list contained 26species with 20–131 spatially independent localities. Allthe species had broad geographic distributions, weremostly endemic or primarily restricted to the Cerrado(Silva 1995, 1997; Silva & Bates 2002), and included sixspecies currently classified as vulnerable to extinction(IBAMA 2003; IUCN 2008). We classified species accord-ing to their main habitat dependency (grassland, savanna,or forest) on the basis of field experience and the lit-erature. We followed the checklist of Comite Brasileirode Registros Ornitologicos (http://www.cbro.org.br) forbird nomenclature.
Environmental Data Sets
Range boundaries of birds correlate with climatic factors,such as temperature and precipitation (reviews in Gaston
Conservation Biology
Volume 23, No. 6, 2009

Marini et al. 1561
2003; Newton 2003). Thus, we chose these variables tocapture both the annual and the strong seasonal charac-teristics of the Cerrado bioclimate and to describe thegeneral climatic environment of pixels. Precipitation ishighly seasonal in the Cerrado, with rains mostly concen-trated during 7 months and as much as 5 months withno or very little precipitation (see Supporting Informa-tion). In the Cerrado region, mean annual temperature isexpected to increase by as much as 3.3 ◦C (13.9%) andprecipitation is expected to remain nearly the same over100 years, according to mean predictions of three GCMs(IPCC 2007) (Supporting Information).
We used 12 bioclimatic variables relating to tempera-ture and precipitation to run the models. We extractedmean, maximum, and minimum temperatures and to-tal precipitation for each month for 1961–1990 fromthe 2007 International Panel on Climate Change (IPCC)database (http://www.ipcc-data.org/). All downloadedenvironmental variables had resolutions broader than 1◦
× 1◦ latitude × longitude. Thus, we interpolated thesevalues to a 1◦ × 1◦ latitude × longitude grid over SouthAmerica (1468 pixels total) with MATLAB. Mean valueswere averages for the period 1961–1990. From these fourextracted variables, we created the following 12 variablesfor the niche modeling: mean annual temperature, tem-perature seasonality (SD of mean monthly temperatures),minimum temperature of the coldest month, maximumtemperature of the hottest month, mean temperature ofthe coldest and hottest quarter, annual precipitation, pre-cipitation seasonality (coefficient of variation of monthlyprecipitation), precipitation of the driest month, precip-itation of the wettest month, and precipitation of thewettest and driest quarter. Their heuristic estimates ofrelative contributions to the models were calculated au-tomatically by the programs. To determine the estimates,in each iteration of the training algorithm the increasein gain is added or subtracted to the contribution of thecorresponding variable.
Climate-Change Scenarios
We realized projections over three time intervals: 2011–2030, 2046–2065, and 2080–2099. Future projectionsperformed were derived from three GCMs (BCM2,MIROHIC3_2-HI, and MK3). These models are consideredthe most advanced tools currently available for simulatingthe response of the global climate system to increasinggreenhouse gas concentrations. For each one of these cli-mate models, we used climate parameters derived fromtwo IPCC (2007) Special Report Emissions Scenarios(SRES) (A1B and B1), which reflect the potential impactsof different assumptions about demographic changes, so-cioeconomic, and technological developments on the re-lease of greenhouse gases. Thus, for each of the 9 niche-modeling tools, we had 18 projections (three GCMs ×
two SRES scenarios, for three time intervals) for eachspecies (details in Supporting Information).
Statistical Analyses
To test the hypothesis of differences in range shifts orrange contractions among the three habitat types, weused Kruskal–Wallis tests because variances were usuallyunequal.
Results
Five of the nine models (random forest, generalizedboosting model, artificial neural networks, generalizedadditive model, and multivariate adaptive regressionsplines) were selected as the best models in more than80% of cases. The mixture discriminant analysis was se-lected 33% of the time, and the other three models (Max-ent, generalized additive model, and classification treeanalysis) were selected <8% of the time (see Supplemen-tary Material for model details.) Temperature was moreimportant in the predictive models than precipitation.The six temperature-related variables contributed 70% tothe models averaged over the five best models for the26 species, whereas the six precipitation variables av-eraged over the five best models contributed only 30%.The maximum temperature of the hottest month (20.5%)and temperature seasonality (18.9%) were the two biocli-matic variables with highest relative importance for mostspecies. The other 10 variables had relative importancesof 2.5–8.5%. The mean AUC of the five models selectedfor every species was high and ranged from 0.919 to0.994 (mean = 0.974).
Range Extents
Predicted changes in range size varied across species ac-cording to their main habitat and were negatively corre-lated with their current range size in all scenarios. Theexpected range contractions under null dispersal acrossall species were 7–38% by 2030, 11–64% by 2065, and14–80% by 2099 (Table 2; Fig. 1a). When species wereassumed to be able to fully disperse from current to fu-ture ranges, the expected changes in range size were3% increase to 37% decrease by 2030, 1% increase to57% decrease by 2065, and 5% increase to 74% decreaseby 2099 (Table 2; Fig. 1b; Supporting Information). Forthe full dispersal scenario, only one species increased itsrange through time.
The largest potential loss in range size under fulldispersal was predicted to occur among grassland andforest-dependent species in all time frames (Table 2).Savanna-dependent species showed significantly lowerrange reduction than species from the other two habitatsfor all three time frames (all H > 9.58, p < 0.008). Wide-ranging species showed smaller range contractions than
Conservation Biology
Volume 23, No. 6, 2009
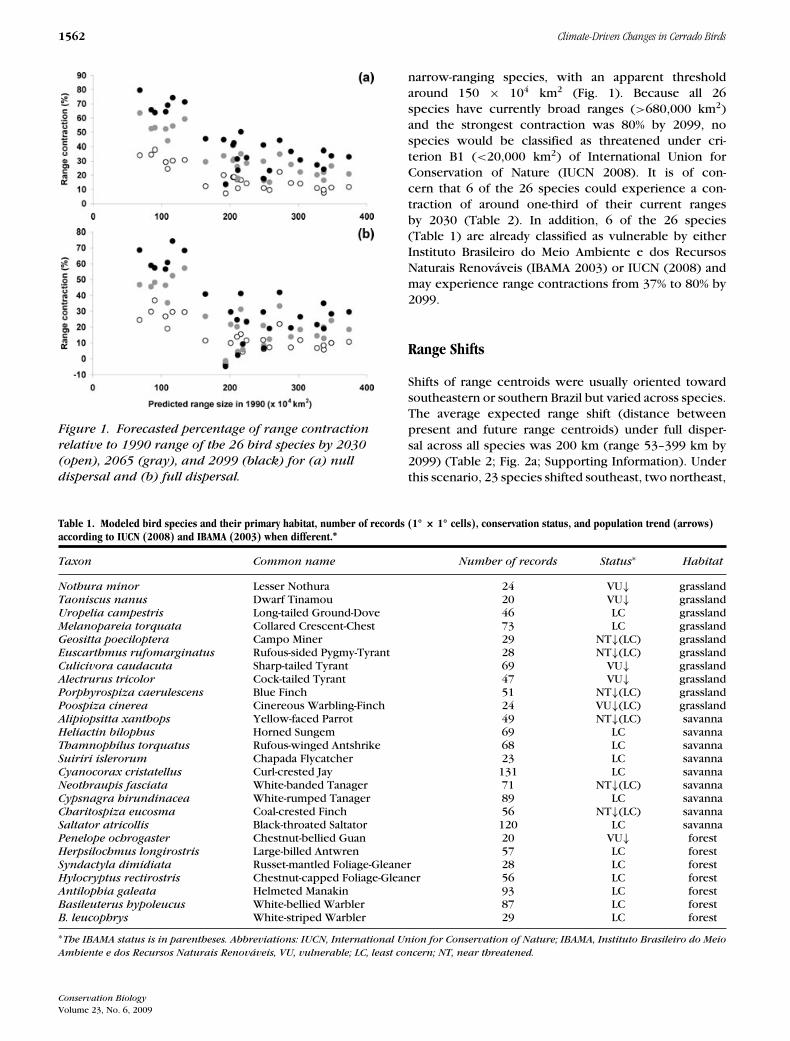
1562 Climate-Driven Changes in Cerrado Birds
Figure 1. Forecasted percentage of range contraction
relative to 1990 range of the 26 bird species by 2030
(open), 2065 (gray), and 2099 (black) for (a) null
dispersal and (b) full dispersal.
Table 1. Modeled bird species and their primary habitat, number of records (1◦ × 1◦ cells), conservation status, and population trend (arrows)according to IUCN (2008) and IBAMA (2003) when different.∗
Taxon Common name Number of records Status∗ Habitat
Nothura minor Lesser Nothura 24 VU↓ grasslandTaoniscus nanus Dwarf Tinamou 20 VU↓ grasslandUropelia campestris Long-tailed Ground-Dove 46 LC grasslandMelanopareia torquata Collared Crescent-Chest 73 LC grasslandGeositta poeciloptera Campo Miner 29 NT↓(LC) grasslandEuscarthmus rufomarginatus Rufous-sided Pygmy-Tyrant 28 NT↓(LC) grasslandCulicivora caudacuta Sharp-tailed Tyrant 69 VU↓ grasslandAlectrurus tricolor Cock-tailed Tyrant 47 VU↓ grasslandPorphyrospiza caerulescens Blue Finch 51 NT↓(LC) grasslandPoospiza cinerea Cinereous Warbling-Finch 24 VU↓(LC) grasslandAlipiopsitta xanthops Yellow-faced Parrot 49 NT↓(LC) savannaHeliactin bilophus Horned Sungem 69 LC savannaThamnophilus torquatus Rufous-winged Antshrike 68 LC savannaSuiriri islerorum Chapada Flycatcher 23 LC savannaCyanocorax cristatellus Curl-crested Jay 131 LC savannaNeothraupis fasciata White-banded Tanager 71 NT↓(LC) savannaCypsnagra hirundinacea White-rumped Tanager 89 LC savannaCharitospiza eucosma Coal-crested Finch 56 NT↓(LC) savannaSaltator atricollis Black-throated Saltator 120 LC savannaPenelope ochrogaster Chestnut-bellied Guan 20 VU↓ forestHerpsilochmus longirostris Large-billed Antwren 57 LC forestSyndactyla dimidiata Russet-mantled Foliage-Gleaner 28 LC forestHylocryptus rectirostris Chestnut-capped Foliage-Gleaner 56 LC forestAntilophia galeata Helmeted Manakin 93 LC forestBasileuterus hypoleucus White-bellied Warbler 87 LC forestB. leucophrys White-striped Warbler 29 LC forest
∗The IBAMA status is in parentheses. Abbreviations: IUCN, International Union for Conservation of Nature; IBAMA, Instituto Brasileiro do Meio
Ambiente e dos Recursos Naturais Renovaveis, VU, vulnerable; LC, least concern; NT, near threatened.
narrow-ranging species, with an apparent thresholdaround 150 × 104 km2 (Fig. 1). Because all 26species have currently broad ranges (>680,000 km2)and the strongest contraction was 80% by 2099, nospecies would be classified as threatened under cri-terion B1 (<20,000 km2) of International Union forConservation of Nature (IUCN 2008). It is of con-cern that 6 of the 26 species could experience a con-traction of around one-third of their current rangesby 2030 (Table 2). In addition, 6 of the 26 species(Table 1) are already classified as vulnerable by eitherInstituto Brasileiro do Meio Ambiente e dos RecursosNaturais Renovaveis (IBAMA 2003) or IUCN (2008) andmay experience range contractions from 37% to 80% by2099.
Range Shifts
Shifts of range centroids were usually oriented towardsoutheastern or southern Brazil but varied across species.The average expected range shift (distance betweenpresent and future range centroids) under full disper-sal across all species was 200 km (range 53–399 km by2099) (Table 2; Fig. 2a; Supporting Information). Underthis scenario, 23 species shifted southeast, two northeast,
Conservation Biology
Volume 23, No. 6, 2009
Marini et al. 1563
Table 2. Estimated range size in 1990, percentage of range loss relative to 1990 (negative in case of range extension), and extent and direction ofrange shift by 2099 under full and null dispersal scenarios for 26 bird species classified by their habitat dependency.
Full dispersal Null dispersal
Range range direction range directionarea in range loss (%) shift by of range range loss (%) shift by of range1990 2099 change 2099 change
Habitat, species (104 km2) 2030 2065 2099 (km) (◦to North) 2030 2065 2099 (km) (◦to North)
GrasslandTaoniscus nanus 85 34 53 66 199 151 30 45 59 147 112Nothura minor 106 29 52 64 229 167 27 46 57 236 135Uropelia campestris 211 11 18 24 135 54 4 5 2 117 99Melanopareia torquata 337 8 16 24 109 165 6 13 19 95 130Geositta poeciloptera 139 31 59 71 399 143 30 57 68 390 134Euscarthmus rufomarginatus 224 14 29 32 76 62 12 23 23 72 110Culicivora caudacuta 289 15 28 37 365 192 7 14 20 209 173Alectrurus tricolor 201 15 32 46 257 189 10 22 30 178 145Porphyrospiza caerulescens 250 11 16 17 69 173 6 8 7 188 124Poospiza cinerea 90 38 53 64 221 153 37 48 57 54 154
SavannaAlipiopsitta xanthops 219 7 11 15 64 137 4 6 9 58 124Heliactin bilophus 211 17 26 31 175 125 14 20 25 175 138Thamnophilus torquatus 337 10 22 37 194 159 8 20 35 167 154Suiriri islerorum 194 7 13 14 118 307 −3 −1 −5 15 142Cyanocorax cristatellus 348 11 26 34 188 157 10 24 28 170 131Neothraupis fasciata 302 14 22 31 173 114 12 18 27 200 106Cypsnagra hirundinacea 327 11 20 27 143 122 7 15 22 155 115Charitospiza eucosma 258 10 15 23 53 138 7 12 19 56 130Saltator atricollis 374 12 21 33 178 144 11 19 30 186 127
ForestPenelope ochrogaster 68 34 64 80 262 141 25 47 69 170 127Herpsilochmus longirostris 249 18 30 41 289 144 12 21 30 263 141Syndactyla dimidiata 109 24 44 69 303 127 19 35 61 247 127Hylocryptus rectirostris 164 12 29 45 203 140 11 27 41 209 124Antilophia galeata 273 22 35 44 284 152 22 33 42 270 146Basileuterus leucophrys 116 30 54 74 270 124 30 52 74 270 124B. hypoleucus 216 17 35 50 239 166 16 31 41 246 135
and one northwest. The mean expected range shift undernull dispersal across all species was 175 km (range 15–390 km by 2099) (Table 2; Fig. 2b). Under this scenarioall species shifted southeast. The largest mean potentialrange shift under full dispersal occurred among forest-dependent (mean = 264 km) and grassland (mean = 206km) species. Range shift of savanna-dependent specieswas lower (mean = 143 km) (Table 2). Only forest-dependent species differed from the other two groupsof species either for the full (Kruskal–Wallis, H = 10.40,p = 0.006; post hoc: savanna and forest species, p =0.001, grassland and forest species, p = 0.07) or null(Kruskal–Wallis, H = 9.85, p = 0.007; post hoc: savannaand forest, p = 0.002, grassland and forest, p = 0.035)dispersal scenarios.
Discussion
Model Performance
The ensemble forecasting allowed us to make consen-sus projections for each species on the basis of the five
best performing models from the nine different niche-based modeling techniques. By averaging projections,model-specific uncertainty was reduced, and by includ-ing linear and nonlinear techniques, the robustness ofmodel predictions increased (Araujo et al. 2005b). Fiveof the nine modeling techniques performed consistentlybetter in predicting current species distributions. Eventhough precipitation seasonality was predicted to havemuch greater effects than temperature seasonality (Sup-porting Information), extreme temperatures seemed tobe the most important factor limiting bird distributions,which reveals their physiological tolerances (Jiguet et al.2006). In the Cerrado the maximum temperature of thehottest month may increase by 4 ◦C by 2100 to approxi-mately 40 ◦C (Supporting Information).
A model of the distributional changes of the HelmetedManakin (Antilophia galeata) under climate changeyielded a directional shift in range similar to ours, butshowed a much larger contraction in suitable habitat(Anciaes & Peterson 2006). This model predicts a rangereduction of 67% and a range shift by 317 km to the south-east by 2070 (2040–2069) under a null dispersal scenario.
Conservation Biology
Volume 23, No. 6, 2009

1564 Climate-Driven Changes in Cerrado Birds
Figure 2. Predicted range shifts for 26 bird species of
the Cerrado between modeled current and future
ranges for (a) null dispersal and (b) full dispersal.
Base of arrows represents centroid of the modeled
current distribution (1990), and tip of arrow
represents the projected centroid of distribution by
2099.
Our predictions for that species were a 33% loss in distri-bution and a 231-km shift by 2065, also in a southeasterlydirection. Results from Anciaes and Peterson (2006) andour study were somewhat similar despite the fact thatAnciaes and Peterson modeled the distribution with asingle modeling tool (GARP), another general circulationmodel, and both topographic and climatic (precipitationand temperature) layers.
Range Extents
Range sizes are expected to decrease over time, and sig-nificantly so as soon as 2030. Our projections, however,did not predict species extinctions due to range contrac-tions under full or null dispersal scenarios. These two
extreme scenarios assumed species were either able todisperse between current and future distribution ranges,so that they would completely occupy the future pre-dicted range, or were unable to disperse at all and wouldoccupy only the overlap area between predicted currentand future distribution ranges. In contrast, Anciaes andPeterson (2006) predicted that 20% of Cerrado manakinspecies could become extinct in the Cerrado by 2055,although their study used a nonconsensus approach.We predicted, however, the full dispersal scenarios mightbe expected for at least some species, such as Cock-tailed Tyrant (Alectrurus tricolor), which is migratory;the Curl-crested Jay (Cyanocorax cristatellus), whichhas expanded with the expansion of Cerrado vege-tation; Helmeted Manakin and the White-bellied War-bler (Basileuterus hypoleucus), which are able to movethrough fragmented landscapes: and Black-throated Salta-tor (Saltator atricollis), which is adapted to altered veg-etation.
Different conservation measures should be undertakendepending on bird habitat dependencies because theresponse to climate change differed between grasslandand forest birds relative to savanna birds. Grassland- andforest-dependent birds will probably have the worst fu-ture conservation scenarios because these habitat typesare the least common. Among the three broadly definedvegetation types, savanna vegetation is the only type thatcurrently can easily and quickly expand into disturbed ar-eas. Forests usually represent <10% of landscapes in theCerrado region (Eiten 1978), and natural grasslands arebecoming increasingly rare (Ratter et al. 1997). Dispersalabilities of forest birds are expected to be lower for forestthan for open-vegetation birds. Several forest-dependentbirds from the Cerrado region have decreased dispersalrates with increasing forest distance (Andrade & Marini2001) and are already disappearing from small forest frag-ments (Christiansen & Pitter 1997; Marini 2001). On theother hand, other forest-dependent birds endemic to theCerrado survive well in small forest patches (Andrade &Marini 2001; Marini 2001). The scenario for some grass-land birds may be even worse than for most other speciesbecause some of them are very sensitive to habitat distur-bance and are already disappearing at a fast rate (Willis &Oniki 1992).
The negative correlation between current range ex-tent and predicted range loss revealed that geographi-cally restricted birds could become even rarer. The lackof predicted change in IUCN threat classification for thesespecies is not surprising because they are all broad-rangespecies. Nevertheless, the negative correlation we foundreveals that even more attention should be paid to thepredictions for range-restricted species, as suggested byJetz et al. (2007), especially those under 150 × 104 km2.This special attention should occur shortly because someof these more restricted-range species might lose up toone-third of their ranges by 2030.
Conservation Biology
Volume 23, No. 6, 2009

Marini et al. 1565
Range Shifts
Range shifts were predicted to differ among birdsfrom different habitats. Forest- and grassland-dependentspecies should undergo stronger range shifts thansavanna-dependent species and should require specificconservation actions. In addition, there were major dif-ferences in range shifts among grassland species, whichrevealed that efficient conservation measures should bemore species oriented than habitat oriented. Conversely,predicted range shifts of forest species were more ho-mogeneous, which revealed that conservation measuresfor these species should be more habitat oriented thanspecies oriented.
The predicted direction of change is of obvious con-servation concern because of potential conflicts with hu-man development, lack of potential new reserves, andsynergy with other factors, such as habitat fragmentationand dispersal constraints. Range shifts, even though lim-ited on average (approximately 200 km), were predictedto occur mostly toward the southeast, which is the mostdeveloped and populated region of Brazil. Similarly, dra-matic range contractions of a high number of birds arepredicted in southeastern Brazil under several socioeco-nomic scenarios (Jetz et al. 2007). Thus, potential rangeshifts may not occur or may be constrained because landuse in most of southeastern Brazil is highly shaped by hu-mans (Ratter et al. 1997) and because land-use changesand natural habitat loss are predicted to increase by atleast 45% in savannas and by 78% in tropical forests (vanVuuren et al. 2006). Furthermore, land use is predictedto be a more important driver of biodiversity change ingrasslands and savannas than climate (Sala et al. 2000).In addition, the creation of new reserves to account forthese projected changes may not be feasible because ofintense land use and lack of undisturbed areas, a similarproblem to be solved in several regions of the world (vanRensburg et al. 2004).
Moreover, the high level of habitat fragmentation incentral-southeastern Brazil further limits the availabilityof large habitat patches and dispersal corridors. For ex-ample, only 0.95% of the original Cerrado vegetation stillexists in the state of Sao Paulo (Kronka et al. 1998), wherelosses of grassland birds have long been documented(Willis & Oniki 1992). This shortage of dispersal corri-dors will be much more limiting to forest species than toopen-vegetation species. Current evidence suggests thatmost Cerrado birds will not change their ranges withan expansion of savanna (Cerrado) into degraded areasto the southeast (Lopes 2008) or will disappear from ordecrease in abundance in disturbed open areas (Tubelis& Cavalcanti 2000). It is estimated that loss of Cerradohabitat will be from 12% to 60%, depending on the sce-nario of global warming (Malcolm et al. 2006). Also, itis predicted that endemic Cerrado trees will loose >50%of their potential distributional area (Siqueira & Peterson
2003). The loss of vascular-plant diversity is projected tobe worst in species from savannas, shrub, tropical for-est, and tropical woodlands (van Vuuren et al. 2006).Climate change interacts synergistically with habitat frag-mentation and increases extinction risk of populationsof butterflies (McLaughlin et al. 2002). These problemsresemble closely the climate-change land-use conserva-tion challenges occurring in South Africa (Williams et al.2005).
The Atlantic Forest mountain ridge may be another ob-stacle to range shifts of birds because they might not beable to disperse over it or because they are used to in-habiting the flatter Cerrado terrains. It was predicted thatclimate may become suitable up to the Atlantic coast forseveral species even though today the natural dominantvegetation is Atlantic Forest. Suitable habitat, however,may decrease with climate change (Preston et al. 2008),and it is difficult to predict whether species will adapt ornot to these new conditions. Nevertheless, natural grass-lands in the mountains of southeastern Brazil (Ratter et al.1997) may become a habitat to grassland birds that dis-perse south.
Our results add to the consensus that several similarconservation problems are occurring around the globe.First, the southern range shifts predicted here mirror thenorthern range shifts predicted for European (Thomas& Lennon 1999) and North American (Hitch & Leberg2007) birds (shift toward the poles; reviewed in Parmesan& Yohe 2003), which supports the existence of a globalpattern. Second, synergistic effects of climate change andhabitat fragmentation or other factors, such as biotic in-teractions, may hasten conservation even more. Lastly,conflicts between predicted conservation needs underclimatic change and current or future land use tend toincrease, especially in tropical high-diversity regions. Mit-igation of the predicted effects on biodiversity, however,may be constrained by local or regional characteristicsand will require an integrated set of ecological, economic,and sociological approaches (Hannah et al. 2002).
Acknowledgments
M.A.M. was supported by a postdoctoral fellowship fromCAPES and L.E.L. by a doctoral fellowship from FAPEMIG.We thank W. Thuiller for providing the updated versionof BIOMOD.
Supporting Information
Detailed explanations of species-climate modeling andensemble forecasting methods (Appendix S1); tempera-ture and precipitation values for the Cerrado region for1960–1990, 2011–2030, 2046–2065, and 2080–2099 for
Conservation Biology
Volume 23, No. 6, 2009

1566 Climate-Driven Changes in Cerrado Birds
scenarios A1 and B1 of three general circulation models(BCM2, MIROHIC3_2-HI and MK3 (Appendix S2); andexamples of predicted distribution ranges of four birdspecies for 1990, 2030, 2065, and 2099 (obtained with anensemble-forecasting approach) (Appendix S3) are avail-able as part of the on-line article. The author is responsi-ble for the content and functionality of these materials.Queries (other than absence of the material) should bedirected to the corresponding author.
Literature Cited
Anciaes, M., and A. T. Peterson. 2006. Climate change effects onNeotropical manakin diversity based on ecological niche modeling.Condor 108:778–791.
Andrade, R. D., and M. A. Marini. 2001. Bird movement between nat-ural forest patches in southeast Brazil. Pages 125–136 in J. L. B.Albuquerque, J. F. Candido Jr., F. C. Straube, and A. L. Ross, editors.Ornitologia e conservacao: da ciencia as estrategias. Editora UNISUL(Universidade do Sul de Santa Catarina), Tubarao.
Araujo, M. B., and M. Luoto. 2007. The importance of biotic interactionsfor modelling species distributions under climate change. GlobalEcology and Biogeography 16:743–753.
Araujo, M. B., R. G. Pearson, W. Thuiller, and M. Erhard. 2005a. Valida-tion of species-climate impact models under climate change. GlobalChange Biology 11:1504–1513.
Araujo, M. B., R. J. Whittaker, R. J. Ladle, and M. Erhard. 2005b. Reducinguncertainty in projections of extinction risk from climate change.Global Ecology and Biogeography 14:529–538.
Brannstrom, C., W. Jepson, A. M. Filippi, D. Redo, Z. Xu, and S. Ganesh.2008. Land change in the Brazilian savanna (Cerrado), 1986–2002:comparative analysis and implications for land-use policy. Land UsePolicy 25:579–595.
BRASIL. 2007. Mapas de cobertura vegetal dos biomas brasileiros. Avail-able from http:www.mma.gov.br (accessed December 2008).
Broennimann, O., and A. Guisan. 2008. Predicting current and futurebiological invasions: both native and invaded ranges matter. BiologyLetters 4:585–589.
Christiansen, M. B., and E. Pitter. 1997. Species loss in a forest birdcommunity near Lagoa Santa in southeastern Brazil. Biological Con-servation 80:23–32.
Dias, B. F. S. 1990. Conservacao da natureza no Cerrado brasileiro. Pages583–640 in M. N. Pinto, editor. Cerrado: caracterizacao, ocupacaoe perspectivas. Editora Universidade de Brasılia, Brasılia.
Eiten, G. 1972. The Cerrado vegetation of Brazil. Botanical Reviews38:201–341.
Eiten, G. 1978. Delimitation of the Cerrado concept. Vegetatio 36:169–178.
Elith, J., et al. 2006. Novel methods improve prediction of species’distributions from occurrence data. Ecography 29:129–151.
Fonseca, G. A. B., W. Sechrest, and J. Oglethorpe. 2005. Managingthe matrix. Pages 346–358 in T. E. Lovejoy and L. Hannah, editors.Climate change and biodiversity. Yale University Press, New Haven,Connecticut.
Gaston, K. J. 2003. The structure and dynamics of geographic ranges.Oxford University Press, Oxford, United Kingdom.
Guisan, A., O. Broennimann, R. Engler, M. Vust, N. G. Yoccoz, A.Lehmann, and N. E. Zimmermann. 2006. Using niche-based mod-els to improve the sampling of rare species. Conservation Biology20:501–511.
Hampe, A. 2004. Bioclimate envelope models: what they detect andwhat they hide. Global Ecology and Biogeography 13:469–476.
Hannah, L., et al. 2008. Climate change adaptation for conservation inMadagascar. Biology Letters 4:590–594.
Hannah, L., G. F. Midgley, and D. Millar. 2002. Climate change-
integrated conservation strategies. Global Ecology and Biogeogra-phy 11:485–495.
Heikkinen, R. K., M. Luoto, M. B. Araujo, R. Virkkala, W. Thuiller, and M.T. Sykes. 2006. Methods and uncertainties in bioclimatic envelopemodelling under climate change. Progress in Physical Geography30:751–777.
Hickling, R., D. B. Roy, J. K. Hill, R. Fox, and C. D. Thomas. 2006. Thedistributions of a wide range of taxonomic groups are expandingpolewards. Global Change Biology 12:450–455.
Hitch, A. T., and P. L. Leberg. 2007. Breeding distributions of NorthAmerican bird species moving north as a result of climate change.Conservation Biology 21:534–539.
IBAMA (Instituto Brasileiro do Meio Ambiente e dos Recursos Natu-rais Renovaveis). 2003. Lista das especies da fauna ameacada deextincao. Instrucao Normativa n◦ 3, de 27 de maio de 2003. IBAMA,Ministerio do Meio Ambiente, Brasılia (in Portuguese).
IPCC (Intergovenmental Panel on Climate Change). 2007. Climatechange 2007: impacts, adaptation, and vulnerability. Cambridge Uni-versity Press, Cambridge, United Kingdom.
IUCN (International Union for Conservation of Nature). 2008. 2008IUCN Red List of threatened species. IUCN, Gland, Switzerland.Available from http:www.iucnredlist.org (accessed October 2008).
Jetz, W., D. S. Wilcove, and A. P. Dobson. 2007. Projected impacts ofclimate and land-use change on the global diversity of birds. PublicLibrary of Science Biology DOI: 10.1371/journal.pbio.0050157.
Jiguet, F., R. Julliard, C. D. Thomas, O. Dehorter, S. E. Newson, and D.Couvet. 2006. Thermal range predicts bird population resilience toextreme high temperatures. Ecology Letters 9:1321–1330.
Klink, C. A., and R. B. Machado. 2005. Conservation of the BrazilianCerrado. Conservation Biology 19:707–713.
Kronka, F. J. N., et al. 1998. Areas de domınio de Cerrado no estado deSao Paulo. Secretaria de Estado do Meio Ambiente, Instituto Flore-stal, Sao Paulo.
Laurance, W. F., and G. B. Williamson. 2001. Positive feedbacks amongforest fragmentation, drought, and climate change in the Amazon.Conservation Biology 15:1529–1535.
Lopes, L. E. 2008. The range of the curl-crested jay: lessons for evalu-ating bird endemism in the South American Cerrado. Diversity andDistributions 14:561–568.
Machado, R. B., M. B. Ramos Neto, P. G. P. Pereira, E. F. Caldas, D. A.Goncalves, N. S. Santos, K. Tabor, and M. Steininger. 2004. Estima-tivas de perda da area do Cerrado brasileiro. Conservation Interna-tional do Brasil, Brasılia.
McLaughlin, J. F., J. J. Hellmann, C. L. Boggs, and P. R. Ehrlich. 2002. Cli-mate change hastens population extinctions. Proceedings of the Na-tional Academy of Sciences of the United States of America 99:6070–6074.
Malcolm, J. R., C. Liu, R. P. Neilson, L. Hansen, and L. Hannah. 2006.Global warming and extinctions of endemic species from biodiver-sity hotspots. Conservation Biology 20:538–548.
Marini, M. A. 2001. Effects of forest fragmentation on birds of the Cer-rado region, Brazil. Bird Conservation International 11:11–23.
Marmion, M., M. Parviainen, M. Luoto, R. K. Heikkinen, and W. Thuiller.2009. Evaluation of consensus methods in predictive species distri-bution modelling. Diversity and Distributions 15:56–69.
Mittermeier, R. A., N. Myers, P. R. Gil, and C. G. Mittermeier. 1999.Hotspots: earth’s biologically richest and most endangered terres-trial ecoregions. CEMEX/Agrupacion Sierra Madre, Mexico City.
Myers, N., R. A. Mittermeier, C. G. Mittermeier, G. A. Fonseca, and J.Kent. 2000. Biodiversity hotspots for conservation priorities. Nature403:853–858.
Newton, I. 2003. The speciation and biogeography of birds. AcademicPress, London.
Nunes, M. F. C., M. Galetti, S. Marsden, R. S. Pereira, and A. T. Peter-son. 2007. Are large-scale distributional shifts of the blue-wingedmacaw (Primolius maracana) related to climate change? Journalof Biogeography 34:816–827.
Conservation Biology
Volume 23, No. 6, 2009

Marini et al. 1567
Parmesan, C., and G. Yohe. 2003. A globally coherent fingerprint ofclimate change impacts across natural systems. Nature 421:37–42.
Paynter, R. A., Jr., and M. A. Traylor Jr. 1991. Ornithological gazetteerof Brazil. 2 volumes. Museum of Comparative Zoology, Cambridge,Massachusetts.
Pearson, R. G., and T. P. Dawson. 2003. Predicting the impacts of cli-mate change on the distribution of species: are bioclimate envelopemodels useful? Global Ecology and Biogeography 12:361–371.
Pearson, R. G., W. Thuiller, M. B. Araujo, E. Martınez-Meyer, L. Brotons,C. McClean, L. Miles, P. Segurado, T. P. Dawson, and D. C. Lees.2006. Model-based uncertainty in species range prediction. Journalof Biogeography 33:1704–1711.
Peterson, A. T., J. Soberon, and V. Sanchez-Cordero. 1999. Conservatismof ecological niches in evolutionary time. Science 285:1265–1267.
Peterson, A. T., V. Sanchez-Cordero, J. Soberon, J. Bartley, R. W. Bud-demeier, and A. G. Navarro-Siguenza. 2001. Effects of global climatechange on geographic distribution of Mexican Cracidae. EcologicalModeling 144:21–30.
Peterson, A. T., H. Tian, E. Martınez-Meyer, J. Soberon, V. Sanchez-Cordero, and B. Huntley. 2005. Modeling distributional shifts ofindividual species and biomes. Pages 211–228 in T. E. Lovejoy andL. Hannah. Climate change and biodiversity. Yale University Press,New Haven, Connecticut.
Phillips, S. J., R. P. Anderson, and R. E. Schapire. 2006. Maximumentropy modelling of species geographic distributions. EcologicalModelling 190:231–259.
Pounds, J. A., M. P. L. Fogden, and J. H. Campbell. 1999. Biologicalresponse to climate change on a tropical mountain. Nature 398:611–615.
Pressey, R. L., M. Cabeza, M. E. Watts, R. M. Cowling, and K. A. Wilson.2007. Conservation planning in a changing world. Trends in Ecology& Evolution 22:583–592.
Preston, K. L., J. T. Rotenberry, R. A. Redak, and M. F. Allen. 2008.Habitat shifts of endangered species altered climate conditions: im-portance of biotic interactions. Global Change Biology 14:1–15.
Ratter, J. A., J. F. Ribeiro, and S. Bridgewater. 1997. The Brazilian Cer-rado vegetation and threats to its biodiversity. Annals of Botany80:223–230.
Sala, O. E., et al. 2000. Global biodiversity scenarios for the year 2100.Science 287:1770–1774.
Silva, J. M. C. 1995. Birds of the Cerrado region, South America. Steen-strupia 21:69–92.
Silva, J. M. C. 1997. Endemic bird species and conservation in the Cer-rado region, South America. Biodiversity and Conservation 6:435–450.
Silva, J. M. C., and J. M. Bates. 2002. Biogeographic patterns and conser-vation in the South American Cerrado: a tropical savanna hotspot.BioScience 52:225–233.
Silva, J. M. C., and M. P. D. Santos. 2005. A importancia relativa dosprocessos biogeograficos na formacao da avifauna do Cerrado e deoutros biomas brasileiros. Pages 220–233 in A. Scariot, J. C. Souza-Silva, and J. M. Felfili, editors. Cerrado: ecologia, biodiversidade econservacao. Ministerio do Meio Ambiente, Brasılia.
Siqueira, M. F. de, and A. T. Peterson. 2003. Consequences ofglobal climate change for geographic distributions of Cerrado treespecies. Biota Neotropica 3:http://www.biotaneotropica.org.br/v3n2/pt/abstract?article+BN00803022003.
Thomas, C. D., and J. L. Lennon. 1999. Birds extend their ranges north-wards. Nature 399:213.
Thuiller, W. 2003. BIOMOD—optimizing predictions of species distri-butions and projecting potential future shifts under global change.Global Change Biology 9:1353–1362.
Thuiller, W. 2004. Patterns and uncertainties of species’ range shiftsunder climate change. Global Change Biology 10:2020–2027.
Tubelis, D. P., and R. B. Cavalcanti. 2000. A comparison of bird com-munities in natural and disturbed non-wetland open habitats inthe Cerrado’s central region, Brazil. Bird Conservation International10:331–350.
van Houtan, K. S., S. L. Pimm, J. M. Halley, R. O. Bierregaard Jr., and T.E. Lovejoy. 2007. Dispersal of Amazonian birds in continuous andfragmented forest. Ecology Letters 10:219–229.
van Rensburg, B. J., B. F. N. Erasmus, A. S. van Jaarsveld, K. J. Gaston,and S. L. Chown. 2004. Conservation during times of change: cor-relations between birds, climate and people in South Africa. SouthAfrican Journal of Science 100:266–272.
van Vuuren, D. P., O. E. Sala, and H. M. Pereira. 2006. The future ofvascular plant diversity under four global scenarios. Ecology andSociety 11: http://www.ecologyandsociety.org/vol11/iss2/art25/.
Vanzolini, P. E. 1992. A supplement to the ornithological gazetteer ofBrazil. Museu de Zoologia, Universidade de Sao Paulo, Sao Paulo.
Williams, P., L. Hannah, S. Andelman, G. Midgley, M. Araujo, G. Hughes,L. Manne, E. Martinez-Meyer, and R. Pearson. 2005. Planning for cli-mate change: identifying minimum-dispersal corridors for the CapeProteaceae. Conservation Biology 19:1063–1074.
Willis, E. O., and Y. Oniki. 1992. The losses of Sao Paulo birds are worsein the interior than in Atlantic forests. Ciencia e Cultura 44:326–328.
Wilson, K. A., M. I. Westphal, H. P. Possingham, and J. Elith. 2005. Sensi-tivity of conservation planning to different approaches to using pre-dicted species distribution data. Biological Conservation 122:99–112.
Conservation Biology
Volume 23, No. 6, 2009