pollination biology of the hemiepiphytic cactus hylocereus undatus in the tehuacán valley, mexico
TRANSCRIPT

ARTICLE IN PRESS
Journal of AridEnvironments
Journal of Arid Environments 68 (2007) 1–8
0140-1963/$ -
doi:10.1016/j
�CorrespoE-mail ad
www.elsevier.com/locate/jnlabr/yjare
Pollination biology of the hemiepiphytic cactusHylocereus undatus in the Tehuacan Valley, Mexico
A. Valiente-Banueta,�, R. Santos Gallya, M.C. Arizmendib, A. Casasc
aDepartamento de Ecologıa de la Biodiversidad, Instituto de Ecologıa, Universidad Nacional Autonoma de Mexico.
Apartado Postal 70-275 Mexico D. F. C.P. 04510, MexicobUnidad de Biologıa, Tecnologıa y Prototipos, FES-Iztacala, Universidad Nacional Autonoma de Mexico. Av. de los
Barrios 1 Tlalnepantla, Estado de Mexico C.P. 54090. Apartado Postal 133, Mexico, MexicocCentro de Investigaciones en Ecosistemas, Universidad Nacional Autonoma de Mexico.
Apartado Postal 27-3 (Xangari). C.P. 58090, Morelia, Michoacan, Mexico
Received 20 October 2003; received in revised form 6 December 2005; accepted 11 April 2006
Available online 30 June 2006
Abstract
Nearly 118 cacti species have been used by Mesoamerican people since pre-Columbian times, and
about 40 of those species show signs of domestication. Among used and domesticated species, the
hemiepiphytic cactus Hylocereus undatus is one of the most appreciated in Mexico because of its
ornamental value and its edible fruits which are consumed at both regional and national scales. This
study analysed the pollination biology of cultivated plants of H. undatus to determine the role of
selfing in fruit production and the relative importance of nocturnal and diurnal flower visitors in
cross-pollination. Flowers were open nearly 17 h. Nocturnal visitors produced significantly higher
fruit production (76.9%) than diurnal visitors (46.1%). During the night, nectar-feeding bats
Leptonycteris curasoae and Choeronycteris mexicana pollinate flowers, whereas during the day the
bee, Apis melifera pollinates flowers. Both self-pollination (100% in unmanipulated treatments and
53.8% in hand self-pollination treatments) and hand cross-pollination treatments (40%) set fruit.
The capability of selfing in this cultivated plant has not been reported elsewhere and may be a result
of intense local cultivation of this plant in Mexico. This self-pollination capability may significantly
reduce labor costs associated with cultivation and allow fruit production to become a viable
economic option for local people.
r 2006 Elsevier Ltd. All rights reserved.
Keywords: Breeding systems; Fruit production; Hemiepiphytic cactus; Hylocereus undatus; Pollination biology;
Tehuacan-Cuicatlan Valley
see front matter r 2006 Elsevier Ltd. All rights reserved.
.jaridenv.2006.04.001
nding author. Tel.: +525556229010; fax: +52 5556229010.
dress: [email protected] (A. Valiente-Banuet).

ARTICLE IN PRESSA. Valiente-Banuet et al. / Journal of Arid Environments 68 (2007) 1–82
1. Introduction
Ethnobotanical studies conducted in the culturally defined region of Mesoamerica insouthern Mexico and northern Central America indicate that cacti have been domesticatedby pre-Columbian cultures (Casas and Barbera, 2002, pp. 143–162). These studies indicatethat from a documented number of 118 species used by Mesoamerican people, about 40species show different degrees of domestication, and are widely cultivated by currentindigenous peoples (Casas and Barbera, 2002, pp. 143–162). Among these species, thehemiepiphytic cactus Hylocereus undatus (Haworth) Britton and Rose is highlyappreciated throughout Mexico because of its ornamental value and its edible fruits thatare consumed at both regional and national scales (Bravo-Hollis, 1978; Ortiz, 1999). Thisspecies is native to tropical deciduous forests of Mexico, the West Indies, Central Americaand northern South America (Bravo-Hollis, 1978; Nobel and de la Barrera, 2002). Thisclimbing, vine-like species is at present cultivated for fruit production in Cambodia,Colombia, Ecuador, Guatemala, Indonesia, Mexico, Nicaragua, Peru, Taiwan andVietnam, with more recent cultivation in Australia, Israel, Japan, New Zealand,Philippines, Spain and the southwestern United States (Nerd et al., 2002a, pp. 185–197;Nobel and de la Barrera, 2002).Because of its importance as crop species, H. undatus has been extensively studied
horticulturally and eco-physiologically (Barcenas and Campbell, 1994; Raveh et al., 1995;Nerd et al., 1999, 2002b; Barcenas-Abogado et al., 2002; Nobel and de la Barrera, 2002;Nobel et al., 2002). Most studies on the reproductive biology of Hylocereus andSelenicereus genera have been conducted under greenhouse conditions (Weiss et al., 1994;Nerd and Mizrahi, 1997; Mizrahi and Nerd, 1999, pp. 358–366). According to thesestudies, flowers are nocturnal and open only for one night, exhibiting bat and large-hawkmoth pollination syndromes (Grant and Grant, 1979; Haber, 1983, pp. 252–253).Breeding systems already described show a wide variety of compatibility systems (Weisset al., 1994). Hylocereus polyrhizus and Hylocereus costaricensis have been reported as self-incompatible, requiring cross-pollination to set fruit. Weiss et al. (1994) reported that H.
undatus cannot produce fruits by automatic self-pollination and produce a reduced numberof seeds by hand self-pollination (50–79.6% fruit set) compared with outcrossed fruits(100% fruit set), while Selenicereus megalanthus is self-compatible, producing fruits bothby automatic self-pollination (60–73% fruit set) and by hand self-pollination (100% fruitset). Apparently, differences between the pollination systems of the two species are due tomorphological differences in the position of anthers and stigma that prevent/allowautomatic self-pollination (Weiss et al., 1994).Fruits of Hylocereus species develop from both the ovary and the receptacule that
surrounds the ovary, can weigh up to approximately 800 g, and exhibit a positivecorrelation between the weight of the fruit and the number of seeds (Weiss et al., 1994;Nerd and Mizrahi, 1997). In all self-fruitful species of Hylocereus analysed by Weiss et al.(1994) fruits obtained by hand cross-pollination, either within a species or by crossingpollen between species, were heavier than those obtained by hand self-pollination.In the Tehuacan Valley in south-central Mexico, H. undatus, called ‘pitahaya’ locally, is
a widely cultivated plant, and according to local people, cultivation of this crop species hasbeen practiced since ancient times. At present, production and commercialization ofpitahaya fruits constitute an important economic activity. Cultivation of pitahaya iscommonly practiced in home gardens by planting stems obtained from other plants at the

ARTICLE IN PRESSA. Valiente-Banuet et al. / Journal of Arid Environments 68 (2007) 1–8 3
base of trees. Preliminary observations by the authors indicate that fruit production inhome gardens is highly productive in the region, but it is unknown if management by localpeople involves the planting of different genotypes or whether this aspect is irrelevantbecause plants are self-compatible. Therefore, in this study we analysed the pollinationbiology of H. undatus in order to determine (1) the role of selfing in fruit production and(2) the relative importance of nocturnal and diurnal pollinators. We then compare thebiological aspects of fruit production in Mexico to data produced by other studies in othercountries.
2. Methods
2.1. Study site
The study was conducted in June 2001 in home gardens of the Tehuacan Valley in thestate of Puebla, Mexico. This is an arid to semiarid region that owes its aridity to the rainshadow produced by the Eastern Sierra Madre (Smith, 1965). It has an average rainfall of380mm and an annual mean temperature of 21 1C with rare frosts (Garcıa, 1973).
Home gardens in the study area are characterized by the presence of a spiny tropicalevergreen forest called ‘mezquital’ dominated by the tree Prosopis laevigata (Humb. andBonpl. Ex Willd.) M. C. Johnston. Other species growing within the study site areStenocereus pruinosus (Otto) F. Buxb., Celtis pallida Torrey, Pachycereus hollianus (F.A.C.Weber) F. Buxb., and Morkillia mexicana (Mocino and Sesse) Rose and Painter (Valiente-Banuet et al., 2000). A total of 10 Prosopis laevigata trees reaching ca. 6m tall hadH. undatus plants rooted at the base of each tree.
2.2. Floral biology
One flower from each of 10 plants was used for measuring flower dimensions.Measurements included flower length (i.e. from petal tip to flower base), distance tonectarious chamber (i.e. from petal tip to the nectarious chamber) and external diameter atthe corolla apex at the point of anther dehiscence. We also used this sample to verifywhether flowers were hermaphroditic (i.e. we checked ovaries for presence of ovules andanthers for pollen). In order to describe flower opening and closure, we measured thedistance between opposite tepal tips in a sample of 10 flowers from 10 plants every tenminutes starting at bud opening. Flower receptivity was recorded by monitoring antherdehiscence and stigma turgidity in 10 flowers from 10 plants every 2 h. No nectarproduction was detected in a sample of 10 bagged flowers from nine plants that werebagged before anthesis with bridal veil netting and followed until closure.
2.3. Breeding system
We conducted pollination experiments in order to determine whether flowers are self-compatible and the effectiveness of nocturnal and diurnal visitors. We used a total of 72tagged flowers in six pollination treatments across 10 plants. Different numbers of flowerswere assigned to each of the following treatments: (1) hand self-pollination (n ¼ 13 on 10plants), flower buds were bagged and 2 h after the flowers opened they were hand-pollinated using pollen obtained from the same flower; (2) nocturnal pollination (n ¼ 13 on

ARTICLE IN PRESSA. Valiente-Banuet et al. / Journal of Arid Environments 68 (2007) 1–84
10 plants), flower buds were tagged before opening and were exposed to nocturnal visitorsduring the night and excluded from diurnal visitors by bagging flowers at sunrise; (3)diurnal pollination (n ¼ 13 on 9 plants), flower buds were bagged and remainedunavailable to nocturnal visitors, then, flowers were exposed to diurnal visitors byremoving the bag at sunrise until the flowers closed; (4) hand cross-pollination treatment(n ¼ 10 on 10 plants), flower buds were bagged, and when flowers opened they were hand-pollinated by saturating the stigma with fresh pollen obtained from another plant; (5)open-pollinated control (n ¼ 20 on 10 plants), flowers that opened during 2 consecutivedays were tagged and these flowers were available to nocturnal and diurnal visitors. Also, atotal of three flowers were bagged in order to determine if the plants are able to producefruits through unmanipulated (automatic) self-pollination (6). The fate (aborted ordeveloping fruit) of the tagged flowers from pollination treatments was scored. Fruits weremonitored every month until they matured. Mature fruits were collected in mid-July.Number of seeds per fruit in each treatment was obtained by collecting one nearly maturefruit from three plants. We obtained the mean weight per seed by weighting 10 groups of50 mature seeds (to the nearest 0.1mg). Then we weighted total seeds per fruit and dividedby mean seed weight to obtain the number of seeds per fruit. Number of seeds amongtreatments was analysed by a one-way ANOVA. All statistical analyses used JMP 3.1software (JMP Statistics and Graphics 1995).
2.4. Flower visitors
Nocturnal and diurnal visitors to H. undatus flowers were captured by placing three mistnets (12m long� 2m tall; separated by E20m) across the home garden area for 2 days. Inaddition, a black-light trap was used to capture insects during the night. For each animalcaught, pollen preparations were made by rubbing a cube of fuchsin-stained jelly (Beattie,1971) over the animal’s body. The cube was placed on a microslide, melted, and coveredwith a coverslip for later examination under the microscope. Pollen presence was regardedas proof of flower visitation. Pollen grains from animal samples were later compared withthose obtained directly from flowers of H. undatus.
3. Results
3.1. Floral biology
During the flowering peak of H. undatus, plants produced a large number of flowers everynight, averaging 6576.89 flowers in the home garden studied. All dissected flowers werehermaphroditic, started to open at dusk (ca. 1900h), began to close at 1100h and had ananthesis duration of ca. 17h. No nectar production was detected in any of the flowers analysed.Flower dimensions in centimeters were (mean71 se): 34.570.58 (flower length), 29.9570.61(distance from the tip to nectarious chamber), and 13.670.15 for external diameter.
3.2. Breeding system
Flowers of H. undatus that were hand self-pollinated had a 53.8% fruit set whereas thethree flowers excluded from any visitor (automatic self-pollination) had 100% fruit set(Table 1). Nocturnal visitors produced a high fruit set rate (76.9%), whereas diurnal
ARTICLE IN PRESSA. Valiente-Banuet et al. / Journal of Arid Environments 68 (2007) 1–8 5
visitation produced 46% fruit set. Flowers visited by both nocturnal and diurnal visitors(open pollinated control) had 100% fruit set (Table 1).
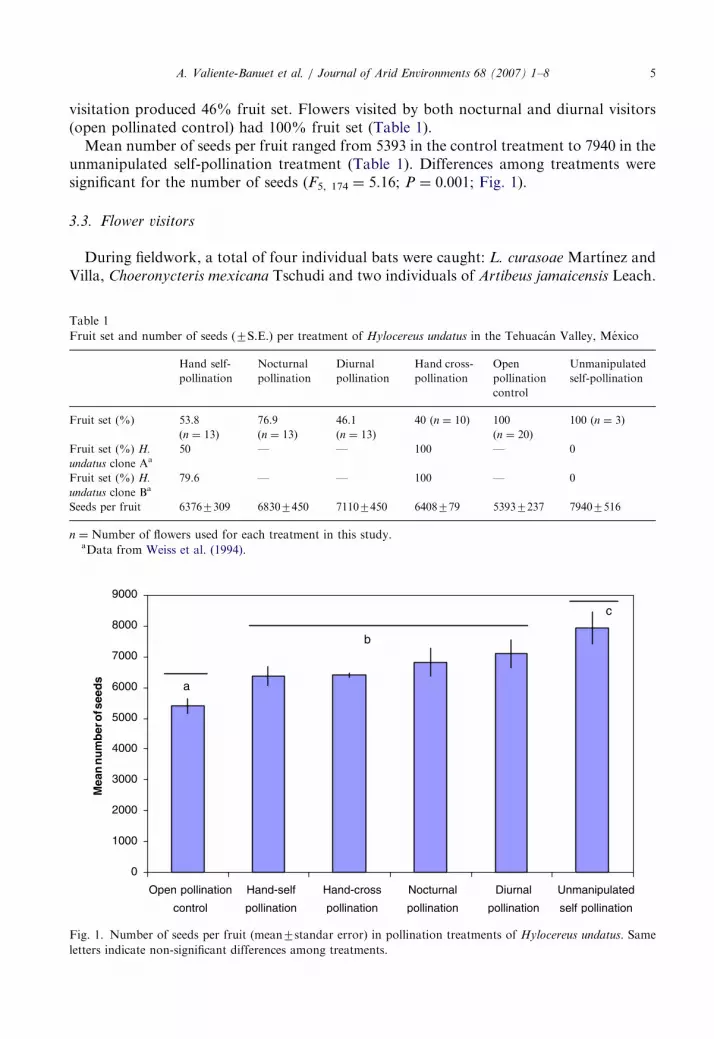
Mean number of seeds per fruit ranged from 5393 in the control treatment to 7940 in theunmanipulated self-pollination treatment (Table 1). Differences among treatments weresignificant for the number of seeds (F5, 174 ¼ 5.16; P ¼ 0:001; Fig. 1).
3.3. Flower visitors
During fieldwork, a total of four individual bats were caught: L. curasoae Martınez andVilla, Choeronycteris mexicana Tschudi and two individuals of Artibeus jamaicensis Leach.
Table 1
Fruit set and number of seeds (7S.E.) per treatment of Hylocereus undatus in the Tehuacan Valley, Mexico
Hand self-
pollination
Nocturnal
pollination
Diurnal
pollination
Hand cross-
pollination
Open
pollination
control
Unmanipulated
self-pollination
Fruit set (%) 53.8
(n ¼ 13)
76.9
(n ¼ 13)
46.1
(n ¼ 13)
40 (n ¼ 10) 100
(n ¼ 20)
100 (n ¼ 3)
Fruit set (%) H.
undatus clone Aa
50 — — 100 — 0
Fruit set (%) H.
undatus clone Ba
79.6 — — 100 — 0
Seeds per fruit 63767309 68307450 71107450 6408779 53937237 79407516
n ¼ Number of flowers used for each treatment in this study.aData from Weiss et al. (1994).
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
Open pollination
control
Hand-self
pollination
Hand-cross
pollination
Nocturnal
pollination
Diurnal
pollination
Unmanipulated
self pollination
Mea
n n
um
ber
of s
eed
s a
b
c
Fig. 1. Number of seeds per fruit (mean7standar error) in pollination treatments of Hylocereus undatus. Same
letters indicate non-significant differences among treatments.

ARTICLE IN PRESSA. Valiente-Banuet et al. / Journal of Arid Environments 68 (2007) 1–86
The bodies of all bats were carrying pollen of H. undatus. Pollen loads in L. curasoae andChoeronycteris mexicana were qualitatively more abundant than in the two individuals ofArtibeus jamaicensis captured. One of these frugivorous bats was carrying a fruit of‘chicozapote’ Manilkara zapota (L.) Royen. During the day, individuals of the bee Apis
mellifera L. were observed visiting flowers, but no insects were captured at night.
4. Discussion
H. undatus in the Tehuacan-Cuicatlan Valley has high fruit production, and in this studynocturnal pollinators were the most important group of pollinators. Nocturnal pollinationwas effected by L. curasoae and Choeronycteris mexicana, which were carrying abundantpollen loads when captured. It is possible that these bats visit flowers looking for pollen,considering that flowers do not produce nectar. Pollen of H. undatus was also recorded inArtibeus jamaicensis, but pollen loads consisted of no more of 10 grains per individual, andso it is doubtful that this species is an effective pollinator.Both control and unmanipulated self-pollination treatments had 100% fruit set, thus
flowers are capable of setting fruit in the absence of any visitor. These fruit set valuescontrast with those obtained by Weiss et al. (1994) who found that automatic self-pollination produced 0 fruit set, while hand cross-pollination and hand self-pollinationwere 100% and 50–79.6%, respectively. Although our result of 100% fruit set in theautomatic self-pollination treatment was obtained from a reduced sample compared to ourother treatments, the existence of automatic self-pollination in H. undatus growing undernative conditions is a remarkable trait for commercial exploitation that has not beenreported elsewhere. In terms of commercial production, Mizrahi and Nerd (1999,pp. 358–366) and Lichtenzveig et al. (2000) report that in Israel, hand pollination (eithercross-pollination or non-automatic self-pollination) is required for fruit production, andthis requirement results in a tremendous increase in labor costs for producers. It is possiblethat the ability to self-pollinate is genetically fixed in clones and H. undatus clones analysedby Mizrahi and Nerd (1999, pp. 358–366) differed in pollination syndrome from thoseexamined in this study. However, differences in our results from previous studies may alsobe attributable to environmental differences between Israel and Tehuacan, and bothexplanations merit investigation. We suggest further study to (1) address morphologicalbarriers reported by Weiss et al. (1994), (2) obtain a more extensive sample and (3) make acomparison between wild and cultivated plants.Many studies have shown that during the course of domestication, plants commonly
evolve the capability of selfing. This transformation has not been common amongdomesticated cacti (Casas et al., 1999; Cruz and Casas, 2002; Oaxaca, 2003). However, incolumnar cacti, Otero-Arnaiz et al. (2003) documented an increase in selfing individualsand flowers in silviculturally managed and cultivated populations of Polaskia chichipe
(Rol.-Goss) Backeberg relative to wild individuals. H. undatus have been used since ancienttimes by local people and it is possible that selfing evolved during domestication. Thiswould explain high fruit production rates and a loss of the ability to produce nectar.However, this hypothesis should be tested by comparing the pollination biology andbreeding systems of wild populations of H. undatus to those of cultivated populations.As has been reported in other studies, Hylocereus species flower in waves, each wave
lasting about one week and hence ripening also occurs in waves (Weiss et al., 1994). In theTehuacan Valley the H. undatus flowering period falls between May and October, and

ARTICLE IN PRESSA. Valiente-Banuet et al. / Journal of Arid Environments 68 (2007) 1–8 7
flowering cycles occur during 10–15 days periods with a large number of flowers perindividual.
To our knowledge, the flowers of H. undatus are among the largest recorded withinCactaceae and the fruits in the Tehuacan Valley contain a large number of seeds.Considering the positive correlation between fruit weight and number of seeds (Weisset al., 1994; Nerd and Mizrahi, 1997), fruits obtained during our pollination experimentsshould be among the largest fruits recorded. Likewise, Weiss et al. (1994) report 71847613ovules per fruit, which when compared with our number of seeds per fruit indicates a seedset rate of 90% or more in most of our pollination treatments.
H. undatus is a relatively new crop in many countries around the world and an old cropin Mexico. Our pollination study indicates that nectar-feeding bats are the mainpollinators but that plants also have a high capability of selfing. These results stronglysuggest that this vine-like cactus in the Tehuacan Valley deserves considerably moreattention in order to offer effective economic options for local habitants.
Acknowledgments
We thank ‘Dona Panchita’ for facilities provided during our study. Financial supportwas provided by DGAPA IN-208301, IN-227605 to A.V-B and DGAPA IN-207305 toMCA. We thank Kristina Smucker of the University of Montana for the revision of themanuscript.
References
Barcenas, P., Campbell, R.J., 1994. Effect of three substrates on rooting and development of pitahaya (Hylocereus
undatus). Proceedings of the InterAmerican Society of Tropical Horticulture 38, 120–121.
Barcenas-Abogado, A., Tijerina-Chavez, P., Martınez-Garza, L., Becerril-Roman, A., Larque-Saavedra, A.E.,
Colinas de Leon, A.M.T., 2002. Respuesta de tres materiales del genero Hylocereus a la salinidad sulfatico-
clorhıdrica. Terra 20, 123–127.
Beattie, A.J., 1971. A technique for the study of insect-borne pollen. Pan Pacific Entomologist 47, 82.
Bravo-Hollis, H., 1978. Las cactaceas de Mexico. Universidad Nacional Autonoma de Mexico, Mexico D.F.,
743pp.
Casas, A., Barbera, G., 2002. Mesoamerican domestication and diffusion. In: Nobel, P.S. (Ed.), Cacti Biology and
Uses. University of California Press, pp. 143–162, 280pp.
Casas, A., Valiente-Banuet, A., Rojas-Martınez, A., Davila, P., 1999. Reproductive biology and the process of
domestication of the columnar cactus Stenocereus stellatus in central Mexico. American Journal of Botany 86,
534–542.
Cruz, M., Casas, A., 2002. Morphological variation and reproductive biology of Polaskia chende (Cactaceae)
under domestication in central Mexico. Journal of Arid Environments 51, 561–576.
Garcıa, E., 1973. Modificaciones al sistema de clasificacion climatica de Koppen. Instituto de Geografıa,
Universidad Nacional Autonoma de Mexico, Mexico, 217pp.
Grant, V., Grant, K.A., 1979. The pollination spectrum in the southwestern American cactus flora. Plant
Sytematics and Evolution 133, 29–37.
Haber, W.A., 1983. Hylocereus costarricensis (Pitahaya silvestre, wild pitahaya). In: Janzen, D.H. (Ed.), Costa
Rican Natural History. The University of Chicago Press, Chicago, 789pp.
Lichtenzveig, J., Abbo, S., Nerd, A., Tel-Zur, N., Mizrahi, Y., 2000. Cytology and mating systems in the climbing
cacti Hylocereus and Selenicereus. American Journal of Botany 87, 1058–1065.
Mizrahi, Y., Nerd, A., 1999. Climbing and columnar cacti: new arid land fruit crops. In: Janick, J. (Ed.),
Perspectives on New Crops and New Uses. ASHS Press, Alexandria, VA, 528pp.
Nerd, A., Mizrahi, Y., 1997. Reproductive biology of fruit cacti. Horticultural Review 18, 322–346.

ARTICLE IN PRESSA. Valiente-Banuet et al. / Journal of Arid Environments 68 (2007) 1–88
Nerd, A., Gutman, F., Mizrahi, Y., 1999. Ripening and postharvest behaviour of the fruits of two Hylocereus
species (Cactaceae). Postharvest Biology and Technology 17, 39–45.
Nerd, A., Tel-Zur, N., Mizrahi, J., 2002a. Fruits of vine and columnar cacti. In: Nobel, P.S. (Ed.), Cacti Biology
and Uses. University of California Press, Berkeley, CA, 280pp.
Nerd, A., Sitrit, Y., Kaushik, R.A., Mizrahi, Y., 2002b. High summer temperatures inhibit flowering in vine
pitaya crops (Hylocereus spp.). Scientia Horticulturae 96, 343–350.
Nobel, P.S., de la Barrera, E., 2002. High temperatures and Net CO2 uptake, growth, and stem damage for the
hemiepiphytic cactus Hylocereus undatus. Biotropica 34, 225–231.
Nobel, P.S., de la Barrera, E., Beilman, D.W., Doherty, J.H., Zutta, B.R., 2002. Temperature limitations of edible
cacti in California. Madrono 49, 228–236.
Oaxaca, B., 2003. Biologıa reproductiva de Escontria chiotilla (Weber) Rose (Cactaceae) en el Valle de Tehuacan,
Puebla, Mexico. B. Sc. Dissertation. Universidad Michoacana de San Nicolas de Hidalgo, Morelia,
Michoacan, Mexico. 128pp.
Ortiz, H.Y.J., 1999. Pitahaya: Un Nuevo Cultivo para Mexico. Coleccion de Textos Politecnicos. Serie
Biotecnologıas. Limusa/Noriega Editores, Mexico, 111pp.
Otero-Arnaiz, A., Casas, A., Bartolo, C., Perez-Negron, E., Valiente-Banuet, A., 2003. Evolution of Polaskia
chichipe (Cactaceae) under domestication in the Tehuacan-Valley, central Mexico: reproductive biology.
American Journal of Botany 90, 593–602.
Raveh, E., Gersani, M., Nobel, P.S., 1995. CO2 uptake and fluorescence response for a shade-tolerant cactus
Hylocereus undatus under current and double CO2 concentration. Physilogia Plantarum 93, 505–511.
Smith, C.E., 1965. Flora Tehuacan Valley. Fieldiana Botany 31, 107–143.
Valiente-Banuet, A., Casas, A., Alcantara, A., Davila, P., Flores, N., Arizmendi, M.C., Villasenor, J.L., Ortega-
Ramırez, J., 2000. La vegetacion del Valle de Tehuacan-Cuicatlan. Boletın de la Sociedad Botanica de Mexico
67, 25–74.
Weiss, J., Nerd, A., Mizrahi, Y., 1994. Flowering behavior and pollination requirements in climbing cacti with
fruit crop potential. HortScience 29, 1487–1492.