co2 uptake by the cultivated hemiepiphytic cactus, hylocereus undatus
TRANSCRIPT

1Printed in UK
Corresponding Author E-m
copy 2004 Association of App
CO2 uptake b
Department of Org
The climate of thcultivated in 20 counfactors Under wetCrassulacean acid mreduced 57 by onproduced in the rooH undatus is maximthan those for deserCO
2 uptake become
H undatus whose other cacti and are aits shaded habitat tois maximal at 20 mability which is hig(halftimes of 2-3 mconcentration raisesby even larger perc
Key words Abscisforest water-use ef
Intr
Epiphytic and hemfruits commonly knowMexico the Caribbenorthern South Americamainly in two genera subfamily Cactoideae (Britton amp Rose [18 specfrom H costaricensisRose H polyrhizus (FH triangularis (L) BrH undatus (HaworSelenicereus (Berger) S megalanthus (K Scthe best known edible ofrom S setaceus (APFruits of a few epiphyalso edible such as th(Lemaire) D Don Monative to tropical forethe Greek for forest) clamber on trees Theyfacilitate anchoring alo
ail psnobelbiologyuclaedu
lied Biologists
y the cultivated hemiepiphytic cactus Hylocereus undatus
By PARK S NOBEL and ERICK DE LA BARRERA
anismic Biology Ecology and Evolution University of California Los AngelesCalifornia 90095-1606 USA
(Accepted 11 July 2003 Received 14 April 2003)
Summary
e native tropical forest habitats of Hylocereus undatus a hemiepiphytic cactustries for its fruit can help explain the response of its net CO
2 uptake to environmental
conditions about 85 of the total daily net CO2 uptake occurs at night via
etabolism leading to a high water-use efficiency Total daily net CO2 uptake is
ly 10 days of drought possibly involving stomatal closure induced by abscisic acidts which typically occupy a small substrate volume Total daily net CO
2 uptake for
al at daynight air temperatures of 3020degC optimal temperatures that are highert cacti but representative of ambient temperatures in the tropics its total daily nets zero at daynight air temperatures of 4232degC Stem damage occurs at 45degC forphotosynthetic cells show little acclimation to high temperatures compared withlso sensitive to low temperatures -15degC killing half of these cells Consistent withtal daily net CO
2 uptake is appreciable at a total daily PPF of only 2 mol m-2 day-1 and
ol m-2 day-1 above which photoinhibition reduces net CO2 uptake Net CO
2 uptake
hly correlated with stem nitrogen and chlorophyll contents changes only graduallyonths) as the concentration of applied N is changed Doubling the atmospheric CO
2 the total daily net CO
2 uptake of H undatus by 34 under optimal conditions and
entages under adverse environmental conditions
ic acid nitrogen relations photosynthesis pitahaya temperature tolerance tropicalficiency
oduction
iepiphytic cacti with ediblen as pitahayas are native toan Central America and (Fig 1) These species occurof the tribe Hylocereeae ofAnderson 2001) Hylocereusies edible fruits are obtained (FAC Weber) Britton ampAC Weber) Britton amp Roseitton amp Rose and especiallyth) Britton amp Rose] andBritton amp Rose [28 specieshumann ex Vaupel) Moran isne with fruits also consumed de Candolle) Werdermann]tic cacti in other genera are
ose of Epiphyllum anguligerst of these vine-like cacti arests (ldquohylerdquo in Hylocereus iswhere they tend to climb or produce aerial roots whichng the trunks and have large
flowers and large fruits The climate of the nativehabitats of such epiphytes (plants living on otherplants) and hemiepiphytes (epiphytes that cansecondarily develop roots in the ground) ischaracterised by moderate temperatures with littleannual variation high rainfall leading to high relativehumidity and reduced light levels at the canopylocations where these cacti occur (Croat 1978Benzing 1990 Castillo Martiacutenez et al 1996 Luumlttge1997 Ortiacutez 1999)
This review focuses on how the net CO2 uptake of
pitahaya-bearing cacti responds to climatespecifically to temperature drought and light thelatter represented by wavelengths that can beabsorbed by photosynthetic pigments ndash400 to 700nm the photosynthetic photon flux (PPF) The mostwidely cultivated such vine-like cactus is H undatuswhich currently is grown in 20 countries (Fig 1)and it is also the one most studied in terms of gasexchange Its net CO
2 uptake ability has been
determined with respect to temperature drought andphotosynthetic photon flux (PPF) as well as appliednitrogen concentration and elevated atmosphericCO
2 concentrations
2 PARK S NOBEL amp ERICK DE LA BARRERA
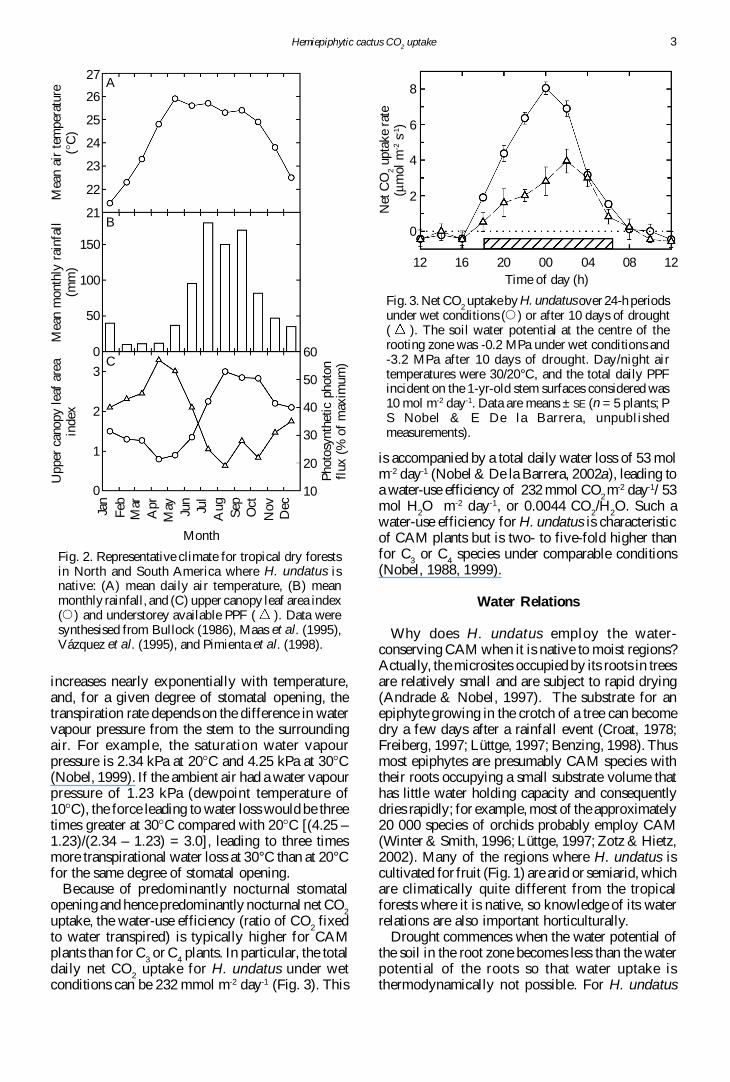
The native distribution of pitahayas (Fig 1) ismostly restricted to tropical dry forests (Barbeau1990) where mean daily air temperature is near24degC ranging from 21degC to 26degC (Fig 2A) Rainfallis usually more than 600 mm year-1 (Rzedowski1983) most of which occurs during the summerwhile the spring is essentially dry (Fig 2B) Suchseasonality for water availability leads to seasonaldifferences in canopy foliage For instance the leafarea index (LAI the ratio of total leaf area to unitground surface area Norman amp Campbell 1989)for the upper canopy is three-fold greater during thewet season than at the end of the dry season (Fig2C) averaging 19 throughout the year A seasonallyvarying PPF occurs under the upper canopyreflecting the changes in water availability and henceLAI instantaneous PPF under the canopy is 50 ofthe maximum during the dry season but only about25 of the maximum during the wet season (Fig2C)
Crassulacean Acid Metabolism
The net CO2 uptake patterns by the stems of
Hylocereus undatus over 24-h periods arecharacteristic of plants employing the Crassulaceanacid metabolism (CAM) pathway of photosynthesisa pathway used by 7 of the nearly 300 000 speciesof vascular plants (Nobel 1991 Winter amp Smith1996) As for other CAM species net CO
2 uptake
by H undatus occurs mainly at night (Fig 3) Inparticular under wet conditions and moderatetemperatures about 85 of its net CO
2 uptake is at
night which is the major period for its stomatalopening (Fig 3 Ortiacutez et al 1996 Ortiacutez 1999) Thisdaynight inversion of the CO
2 uptake pattern
compared with the majority of plants minimiseswater loss by CAM plants About 92 of plantspecies use the C
3 pathway (initial fixation of CO
2catalysed by ribulose-15-bisphosphate carboxylaseoxygenase leading to the formation of a three-carboncompound) and 1 use the C
4 pathway (initial
fixation of CO2 catalysed by phosphoenolpyruvate
carboxylase leading to the formation of a four-carbon compound) such plants take up CO
2 only
during the daytime CO2 uptake at night (Phase I for
CAM plants Osmond 1978 Nobel 1988 1999)involves a binding of CO
2 to a three-carbon
compound catalysed by phosphoenolpyruvatecarboxylase leading to the accumulation of organicacids that are stored in the vacuoles of photosyntheticcells Some net CO
2 uptake generally occurs early
in the morning for H undatus (Fig 3 Phase II) andinvolves the binding of CO
2 by ribulose-15-
bisphosphate carboxylaseoxygenase and also in thelate afternoon (Phase IV) whereas CO
2 tends to be
released during the middle of the daytime (PhaseIII) when the organic acids formed during theprevious night are decarboxylated leading to a highCO
2 concentration within the stem
Air and stem temperatures tend to be lower atnight which is the key to appreciating the water-conserving aspects of CAM Lower temperatureslead to less water loss because air in the intercellularair spaces of the stems is essentially saturated withwater vapour the saturation water vapour pressure
Fig 1 Countries where Hylocereus undatus is currently being cultivated for fruit (back slanted hatching) and whereit as well as other cacti bearing pitahayas are native (forward slanted hatching) Data are from Britton amp Rose (1963)Bravo (1978) Backeberg (1966) Barthlott amp Hunt (1993) Mizrahi amp Nerd (1999) Nerd et al (2002) Ortiacutez (1999)and P S Nobel (unpublished observations)
3Hemiepiphytic cactus CO2 uptake
is accompanied by a total daily water loss of 53 molm-2 day-1 (Nobel amp De la Barrera 2002a) leading toa water-use efficiency of 232 mmol CO
2 m-2 day-1 53
mol H2O m-2 day-1 or 00044 CO
2H
2O Such a
water-use efficiency for H undatus is characteristicof CAM plants but is two- to five-fold higher thanfor C
3 or C
4 species under comparable conditions
(Nobel 1988 1999)
Water Relations
Why does H undatus employ the water-conserving CAM when it is native to moist regionsActually the microsites occupied by its roots in treesare relatively small and are subject to rapid drying(Andrade amp Nobel 1997) The substrate for anepiphyte growing in the crotch of a tree can becomedry a few days after a rainfall event (Croat 1978Freiberg 1997 Luumlttge 1997 Benzing 1998) Thusmost epiphytes are presumably CAM species withtheir roots occupying a small substrate volume thathas little water holding capacity and consequentlydries rapidly for example most of the approximately20 000 species of orchids probably employ CAM(Winter amp Smith 1996 Luumlttge 1997 Zotz amp Hietz2002) Many of the regions where H undatus iscultivated for fruit (Fig 1) are arid or semiarid whichare climatically quite different from the tropicalforests where it is native so knowledge of its waterrelations are also important horticulturally
Drought commences when the water potential ofthe soil in the root zone becomes less than the waterpotential of the roots so that water uptake isthermodynamically not possible For H undatus
increases nearly exponentially with temperatureand for a given degree of stomatal opening thetranspiration rate depends on the difference in watervapour pressure from the stem to the surroundingair For example the saturation water vapourpressure is 234 kPa at 20degC and 425 kPa at 30degC(Nobel 1999) If the ambient air had a water vapourpressure of 123 kPa (dewpoint temperature of10degC) the force leading to water loss would be threetimes greater at 30degC compared with 20degC [(425 ndash123)(234 ndash 123) = 30] leading to three timesmore transpirational water loss at 30degC than at 20degCfor the same degree of stomatal opening
Because of predominantly nocturnal stomatalopening and hence predominantly nocturnal net CO
2uptake the water-use efficiency (ratio of CO
2 fixed
to water transpired) is typically higher for CAMplants than for C
3 or C
4 plants In particular the total
daily net CO2 uptake for H undatus under wet
conditions can be 232 mmol m-2 day-1 (Fig 3) This
Mea
n ai
r te
mpe
ratu
re(deg
C)
27
10
Phot
osyn
thet
ic p
hoto
nfl
ux (
o
f m
axim
um)
Mea
n m
onth
ly r
ainf
all
(mm
)U
pper
can
opy
leaf
are
ain
dex
26
25
2
24
23
22
21
150
100
50
0
3
1
0
A
B
C
20
30
40
50
60
Jan
Feb
Mar
Apr
May Jun
Jul
Aug Se
pO
ctN
ovD
ec
Month
Fig 2 Representative climate for tropical dry forestsin North and South America where H undatus isnative (A) mean daily air temperature (B) meanmonthly rainfall and (C) upper canopy leaf area index(E ) and understorey available PPF ( ) Data weresynthesised from Bullock (1986) Maas et al (1995)Vaacutezquez et al (1995) and Pimienta et al (1998)
Net
CO
2 up
take
rat
e(m
mol
m-2 s
-1)
0
2
4
6
8
12 16 08 120020 04Time of day (h)
Fig 3 Net CO2 uptake by H undatus over 24-h periods
under wet conditions (E ) or after 10 days of drought( ) The soil water potential at the centre of therooting zone was -02 MPa under wet conditions and-32 MPa after 10 days of drought Daynight airtemperatures were 3020degC and the total daily PPFincident on the 1-yr-old stem surfaces considered was10 mol m-2 day-1 Data are means plusmn SE (n = 5 plants PS Nobel amp E De la Barrera unpublishedmeasurements)
4 PARK S NOBEL amp ERICK DE LA BARRERA
rooted in silica sand the total daily net CO2 uptake
does not change appreciably for the first 7 days ofdrought but decreases by 78 at 17 days (Raveh etal 1995) For H undatus rooted in a sandvermiculite mixture the net CO
2 uptake ability
decreases by 57 at 10 days of drought (Fig 3)Also the maximal rate of net CO
2 uptake during
drought tends to occur later in the night (Fig 3)when stem temperatures generally are lower andhence transpiration is lower for a given degree ofstomatal opening
The water vapour conductance for H undatusdecreases during 10 days of drought indicatingstomatal closure but the stem water potential doesnot decrease significantly (Nobel amp De la Barrera2002a) A chemical message probably abscisic acidis apparently sent from the roots in the drying soilto the shoots leading to stomatal closure as a meansfor conserving water Moreover the stomatalresponse to this water-stress hormone prevents theloss of stem water which may be crucial forepiphytic and hemiepiphytic cacti whose rootsexplore only a relatively small soil volume Whenthe roots of H undatus are excised so that no abscisicacid can move from the roots to the shoot duringdrought the decline in net CO
2 uptake is much
slower (50 inhibition is delayed by approximately8 days compared with plants with their roots intact)emphasising the importance of this hormone forconserving stem water during drought (Nobel amp Dela Barrera 2002a)
When H undatus that has been droughted for 10days is rewatered a significant increase in net CO
2uptake occurs in 1 day the CO
2 uptake ability is
half restored in only 2 days and full recovery occursin 7 days (P S Nobel and E De la Barreraunpublished observations) Thus this cactus canrespond rapidly to rainfall events which may becrucial for its growth in tropical regions with frequentrainfalls (Freiberg 1997 Luumlttge 1997) as well asof importance in developing irrigation scheduleswhen it is cultivated as a crop (Mizrahi amp Nerd1999 Nerd et al 2002) Similarly the hydraulicconductivity of the roots (a direct measure of plantwater uptake ability) of two epiphytic cactiEpiphyllum phyllanthus and Rhipsalis bacciferadecreases during drought but increases to the valuesunder wet conditions only 3 days after rewatering(North amp Nobel 1994)
Temperature
Temperature influences nearly every plant processincluding net CO
2 uptake and hence growth The
optimal daynight air temperatures for total daily netCO
2 uptake by Hylocereus undatus are 3020degC (Fig
4) which are relatively high compared withtemperatures optimal for other CAM plants Because
most net CO2 uptake occurs at night the nighttime
temperatures are more crucial for total daily net CO2
uptake than are the daytime ones The optimalnighttime temperatures for total daily net CO
2 uptake
by cacti native to arid or semiarid regions are from10degC to 15degC (Nobel 1988) Apparently the higheroptimal nighttime temperatures for H undatusreflect temperatures characteristic of the primarilytropical regions where it is native (Fig 1) The totaldaily net CO
2 uptake is half of the maximal value at
low daynight temperatures of 188degC and at highdaynight temperatures of 3828degC (Fig 4) Abovedaynight air temperatures of 4232degC a net loss ofCO
2 occurs over a 24-h period (Fig 4) indicating
that this species is not adapted to such hightemperatures
Temperature also affects the survival of cacti andH undatus is more sensitive to temperature extremesthan are most other cacti Based on an assay usingthe uptake of the vital dye neutral red into the centralvacuole of a cell (Onweume 1979 Nobel et al1995) half of the photosynthetic (chlorenchyma)cells are killed by a 1-h low-temperature treatmentat -15degC for H undatus maintained at daynighttemperatures of 2010degC and half are killed at -13degCfor plants maintained at 3020degC (Nobel et al 2002)For 19 other species of cacti half of thephotosynthetic cells are killed at an averagetemperature of -102degC for plants maintained at 2010degC and the low-temperature acclimation orhardening (decrease in lethal temperatures as the daynight temperatures are reduced) averages 07degC asthe daynight temperatures are reduced by 10degC(Nobel 1988) much greater than the 02degC observedfor H undatus Again using the uptake of neutralred into the cell central vacuoles as the assay halfof the photosynthetic cells of H undatus are killedby a 1-h high-temperature treatment at 540degC forplants at 2515degC and at 554degC for plants at 3525degC (Nobel amp De la Barrera 2002b) For 18 otherspecies of cacti half of the photosynthetic cells arekilled at an average temperature of 570degC for plantsmaintained at 3525degC and the high temperatureacclimation (as the daynight temperatures are raisedby 10degC) averages 53degC (Nobel 1988) comparedwith only 14degC for H undatus Thus both the low-temperature and the high-temperature acclimationswhich are the crucial factors for the tolerance ofextreme temperatures (Nobel 1988 Nobel amp De laBarrera 2002b) are less than one-third as great forH undatus as is the average for other cacti
Time of exposure to extreme temperatures is alsocrucial when examining tolerances especially forhigh temperatures For instance H undatus is notexpected to tolerate daynight air temperatures above4232degC on a prolonged basis as the total daily netCO
2 uptake would then be negative (Fig 4) In this
regard the maximal temperatures in the native
5Hemiepiphytic cactus CO2 uptake
habitats of H undatus generally do not exceed 35degC(Croat 1978 Freiberg 1997 Luumlttge 1997 Benzing1998) Maintaining H undatus at 4030degC graduallyleads to visible damage of the stems which firstbecomes evident at 6 wk (Nobel amp De la Barrera2002b) Necrosis increases to about half of the stemsurface area at 15 wk and no new stem growth isobserved at such temperatures Also daytimetemperatures of 45degC in the field greatly reduceflower production by H undatus and brief exposureto 50degC leads to stem necrosis (Mizrahi amp Nerd1999) Thus this species cannot be cultivated wheredaytime maximal temperatures average 40degC forseveral weeks or occasionally exceed 45degC Indeedtotal daily net CO
2 uptake by H undatus is 37
lower after 30 wk at 3525degC compared with at 10wk (Nobel amp De la Barrera 2002b) suggesting thatH undatus does not have phenotypic plasticity totolerate temperatures exceeding those in its nativehabitat for prolonged periods
Photosynthetic Photon Flux
Hylocereus undatus has considerable net CO2
uptake at a total daily PPF of only 2 mol m-2 day-1
(Fig 5) indicating that it is well adapted to the low-light conditions of a tropical forest (total daily netCO
2 uptake is negative for desert CAM species at
this PPF Nobel 1988) [Net CO2 uptake responses
to PPF for CAM plants are not expressed versus theinstantaneous level of such light as is conventionalfor C
3 and C
4 plants but rather versus the total daily
PPF as most CO2 uptake occurs at night in Phase I
(Fig 3) when no light is available] Total daily netCO
2 uptake by H undatus increases as the total daily
PPF increases to about 20 mol m-2 day-1 above whichnet CO
2 uptake decreases with increasing PPF (Fig
5) indicating that high PPF levels damage the
photosynthetic apparatus Total daily net CO2 uptake
for H undatus is about 90 of maximal at only 10mol m-2 day -1 (Fig 5) For CAM plants such as barrelcacti (eg Ferocactus acanthodes) andplatyopuntias (eg Opuntia ficus-indica) a totaldaily PPF of 10 mol m-2 day-1 leads to about 40 ofthe maximal total daily net CO
2 uptake maximal
uptake for such cacti generally occurs from 30ndash40mol m-2 day-1 (Nobel 1988) levels attained bysurfaces facing east or west on clear days in thesummer
Photoinhibition leading to a reduction in total dailynet CO
2 uptake sets in at a total daily PPF of 20 mol
m-2 day-1 for H undatus (Fig 5) compared withabove 40 mol m-2 day-1 for desert CAM species(Nobel 1988) Measurement of chlorophyllfluorescence for H undatus indicates thatphotosynthetic efficiency (total daily net CO
2 uptake
divided by PPF) actually begins to decrease at arelatively low total daily PPF of 5 mol m-2 day-1
(Raveh et al 1995) To avoid exceeding a total dailyPPF of 20 mol m-2 day-1 a vertical climbing orclambering stem of H undatus would need to beshaded by nearly 40 on a clear day for stemsurfaces facing east or west (north- or south-facingsurfaces would need no shading) In this regard Hundatus H polyrhizus and Selenicereusmegalanthus are commercially grown in Israel undershade cloth that attenuates solar irradiation by 30ndash60 (Raveh et al 1993 1998 Mizrahi amp Nerd1999) Such shading is comparable to that in theforests where such species are native (Benzing 1990Barthlott amp Hunt 1993 Luumlttge 1997) Whenexposed to full sun on clear days H undatus in
Fig 4 Total daily net CO2 uptake by H undatus at the
indicated daynight air temperatures Plants weregrowing under wet conditions and the PPF was 10 molm-2 day-1 Data are from Raveh et al (1995) and Nobelamp De la Barrera (2002b)
Tota
l dai
ly n
et C
O2 u
ptak
e(m
mol
m-2 d
ay-1)
-50
0
100
150
200
50
250
155 2515 453535253020 4030Daynight air temperatures
Fig 5 Total daily net CO2 uptake by H undatus at the
indicated total daily PPF incident on the stem surfacePlants were growing under wet conditions and the daynight temperatures were 2515degC Data are from Ravehet al (1995) and P S Nobel amp E De la Barrera(unpublished measurements for 1-yr-old stems withdata indicated by means plusmn SE for n = 5 plants andnormalised to the previous measurements at a totaldaily PPF of 20 mol m-2 day-1)
Tota
l dai
ly n
et C
O2 u
ptak
e(m
mol
m-2 d
ay-1)
0
100
150
200
50
250
0 5 251510 20Total daily PPF (mol m-2 day-1)
30
6 PARK S NOBEL amp ERICK DE LA BARRERA
Mexico (Ortiacutez 1999) and 23 genotypes ofHylocereus in Israel (Nerd et al 1990) developvisible necrosis on the stems However H undatusis cultivated in southeast Asia such as in the MekongDelta of Vietnam without shading (Nerd et al 2002P S Nobel personal observations) Evidently theprevalent cloud cover in such regions reduces thetotal daily PPF to values that do not lead to stembleaching and necrosis
Nutrients
The effects of soil nutrients on net CO2 uptake of
cacti bearing pitahayas have received little attention(Castillo Martiacutenez et al 1996 Ortiacutez 1999) althoughnutrients are known to have substantial effects onother cacti (Nobel 1988 1989) For instanceapplication of nutrients such as nitrogen can alterthe timing of fruiting for Opuntia ficus-indica(Mizrahi amp Nerd 1999) Application of boron andzinc can increase the number of fruits for H undatusand foliar application of magnesium can increasefruit size (Castillo Martiacutenez et al 1996) Variationsin the applied nitrogen concentration for H undatusrooted in a sandvermiculite mixture for 22 wkaffects its net CO
2 uptake ability as measured by the
maximal nocturnal net CO2 uptake rate 016 mM N
leads to a maximal nocturnal net CO2 uptake rate of
25 micromol m-2 s-1 16 mM N leads to 56 micromol m-2 s-1and 16 mM N leads to 98 micromol m-2 s-1 (Fig 6) thelatter being the N concentration in full-strengthHoagland solution (Hoagland amp Arnon 1950Johnson et al 1957 Epstein 1972) Thus net CO
2uptake for H undatus is highest under a relativelyhigh applied N concentration (8-16 mM Fig 6) asis the case for intensively managed greenhouse crops
and field-grown vegetables and ornamentals (Huett1996 Langhans amp Tibbitts 1997) A maximalnocturnal net CO
2 uptake rate of 10 micromol m-2 s-1 (Fig
6) places H undatus fourth highest among the 28species of cacti whose maximal net CO
2 uptake rates
have been reported (Nobel amp Bobich 2002) Thusthis species has a high potential growth rate underhigh-nutrient conditions
Shifts in stem properties of H undatus as theapplied nitrogen concentration is changed arerelatively slow For instance the halftime for theshift in net CO
2 uptake ability in response to applied
N is 12ndash13 wk (Nobel amp De la Barrera 2002c) After22 wk at 016 mM applied N the stem N level on adry mass basis decreases to only 09 (Nobel amp Dela Barrera 2002c) After 22 wk at an applied Nconcentration of 16 mM stem N is 25 comparableto values in the shoots of cultivated C
3 and C
4 plants
under high-nutrient conditions (Mengel amp Kirkby1987 Marschner 1995) Also the chlorophyll levelof H undatus at an applied N concentration of 016mM for 23 wk is only 029 g m-2 compared with 063g m-2 at 16 mM N (Nobel amp De la Barrera 2002c)H undatus supplied with 16 mM N for 8 wk has astem chlorophyll level of 049 g m-2 (Raveh et al1998) and H polyrhizus has 045 g chlorophyll m-2
under comparable conditions (Raveh et al 1998)Following 23 wk at low N levels the stem net CO
2uptake ability and chlorophyll level both increaseto maximal values when 16 mM N is applied to Hundatus for 11 wk with a halftime for the increaseof 8 wk (Nobel amp De la Barrera 2002c) Thusresponses of H undatus to changes in the applied Nconcentration are slower than for most C
3 and C
4crops and other cacti for which major responses canoccur in 2 wk (Mengel amp Kirkby 1987 Nerd et al1993 Marschner 1995 Nerd amp Nobel 1995)suggesting lack of genetic plasticity in this recentlydomesticated cactus or relatively slow nutrientuptake by the roots Because the maximal net CO
2uptake rates for H undatus are similar at applied Nconcentrations of 8 and 16 mM (Fig 6) the latterlevel may lead to luxury (excess) N consumptionas the tissue level after 22 wk is 36 higher at 16mM than at 8 mM applied N (Nobel amp De la Barrera2002c)
Elevated Atmospheric CO2
Another environmental variable influencing netCO
2 uptake by cacti is the atmospheric CO
2concentration which currently is increasing bynearly 3 parts per million (09) per year (Schimelet al 1996 Stitt amp Krapp 1999) For all 20 CAMspecies studied biomass productivity increases fora doubled atmospheric CO
2 concentration compared
to the current atmospheric CO2 concentration
(Drennan amp Nobel 2000) The increase averages
Fig 6 Maximal net CO2 uptake rate at night for H
undatus maintained for 22 wk at the indicated nitrogenconcentration Plants were growing under wetconditions the daynight air temperatures were 3020degC and the total daily PPF was 10 mol m-2 day-1The maximal rate occurred between 2200 h and 0200h Data are from Nobel amp De la Barrera (2002c)
Max
imal
net
CO
2 up
take
rat
e(m
mol
m-2 d
ay-1)
4
6
8
2
10
0 2 1464 8Nitrogen concentration (mM)
160
7Hemiepiphytic cactus CO2 uptake
35 at 700 ppm (700 micro litre litre -1 or 70 kPa)compared with 350 ppm which represents anapproximately 1 increase in growth per 10 ppmincrease in atmospheric CO
2 concentration for CAM
plants (Nobel 1991) Data for H undatus areconsistent with the results for other CAM plants asa doubled atmospheric CO
2 concentration increases
the total daily net CO2 uptake by 34 (Raveh et al
1995) Most of the increase occurs during thedaytime in Phases II and IV Moreover increases intotal daily net CO
2 uptake under a doubled
atmospheric CO2 concentration compared with the
current atmospheric CO2 concentration are more
prominent under less favorable or even stressfulclimatic conditions For instance the increase in totaldaily net CO
2 uptake for H undatus due to a doubled
atmospheric CO2 concentration is 63 higher at 35
25degC versus 2515degC 90 higher at 24 days ofdrought versus 14 days and 120 higher at a PPFof 30 mol m-2 day-1 versus 20 mol m-2 day-1 (Ravehet al 1995) Thus the responses to atmospheric CO
2increases will not only lead to greater total daily netCO
2 uptake but also will lead to proportionally
greater net CO2 uptake under adverse environmental
conditions
Conclusions and Future Prospects
A striking feature of H undatus is how rapidlynocturnal stomatal opening and net CO
2 uptake
abili ty decrease during drought Such rapidresponses which undoubtedly are related to the oftensmall substrate volume that is occupied by its rootsmay be mediated by abscisic acid Other rootproperties such as the apparently low nutrient uptakerate and the influence of mycorrhizae also needinvestigation Another striking ecophysiologicalfeature of H undatus compared with barrel andcolumnar cacti which is especially pertinent to itscultivation outside its native regions is its poortolerance of high temperatures such as those above40degC Indeed its photosynthetic cells do not showmuch acclimation as the daynight air temperaturesare gradually raised over a period of weeks contraryto the case for other cacti Comparisons at amolecular level between H undatus and other cactishowing substantial high-temperature acclimationcould help elucidate the mechanisms involved forthe latter group The intolerance of H undatus tohigh PPF presumably relates to its shaded naturalhabitat Levels of stem nitrogen and chlorophyllcorrelate well with the net CO
2 uptake ability of H
undatus providing relatively simple assays for thephysiological status of its stems which changerelatively slowly in response to changes in theconcentration of applied nitrogen Indeedunderstanding how climatic factors in general andhormonal factors in particular affect the gas
exchange of H undatus and related species will beinteresting ecologically and extremely importanthorticulturally for the expanding cultivation ofpitahayas
Acknowledgements
We thank the UCLA-Ben Gurion UniversityProgram of Cooperation through the gift of Dr SolLeshin and the US-Israel Binational AgriculturalResearch and Development Fund grant no IS-3382-01 for financial support
References
Anderson E F 2001 The Cactus Family Portland OregonUSA Timber Press 776 pp
Andrade J L Nobel P S 1997 Microhabitats and waterrelations of epiphytic cacti and ferns in a lowland neotropicalforest Biotropica 29261-270
Backeberg C 1966 Das Kakteenlexikon Jena GermanyGustav Fisher 828 pp
Barbeau G 1990 La pitahaya rouge un nouveau fruit exotiqueFruits 45141-147
Barthlott W Hunt D R 1993 Cactaceae In The Families andGenera of Vascular Plants Vol 2 pp 161-196 Ed KKubitzki Berlin Germany Springer-Verlag
Benzing D H 1990 Vascular Epiphytes General Biology andRelated Biota Cambridge UK Cambridge University Press354 pp
Benzing D H 1998 Vulnerabilities of tropical forests to climatechange The significance of resident epiphytes ClimateChange 39519-540
Bravo H H 1978 Las Cactaacuteceas de Meacutexico Vol I MexicoCity Mexico UNAM 743 pp
Britton N L Rose J N 1963 The Cactaceae Descriptionsand Illustrations of Plants of the Cactus Family Vol II NewYork USA Dover 241 pp
Bullock S H 1986 Climate of Chamela Jalisco and trends inthe south coastal region of Mexico Archives for MeteorologyGeophysics and Bioclimatology Series B 36297-316
Castillo Martiacutenez R Caacutelix De Dios H Rodriacuteguez Canto A1996 Guia Teacutecnica para el Cultivo de Pitahaya ChetumalQuintana Roo Mexico Universidad de Quintana Roo 158pp
Croat T B 1978 Flora of Barro Colorado Island StanfordCalifornia USA Stanford University Press 943 pp
Drennan P M Nobel P S 2000 Responses of CAM species toincreasing atmospheric CO
2 concentrations Plant Cell and
Environment 23767-781Epstein E 1972 Mineral Nutrition of Plants Principles and
Perspectives New York USA Wiley 412 ppFreiberg M 1997 Spatial and temporal pattern of temperature
and humidity of a tropical premontane rain forest tree in CostaRica Selbyana 1877-84
Hoagland D R Arnon D I 1950 The water-culture methodfor growing plants without soil California AgriculturalExperimental Station Circular 3471-32
Huett D O 1996 Prospects for manipulating the vegetative-reproductive balance in horticultural crops through nitrogennutrition A review Australian Journal of AgriculturalResearch 4747-66
Johnson C M Stout P R Broyer T C Carlton A B 1957Comparative chlorine requirements of different plant speciesPlant Soil 8337-353
Langhans R W Tibbitts T W 1997 Plant growth chamberhandbook Ames Iowa USA Iowa State University 240 pp
8 PARK S NOBEL amp ERICK DE LA BARRERA
Luumlttge U 1997 Physiological Ecology of Tropical PlantsBerlin Germany Springer-Verlag 384 pp
Maas J M Vose J M Swank W T Martiacutenez-Yriacutezar A 1995Seasonal changes of leaf area index (LAI) in a tropicaldeciduous forest in west Mexico Forest Ecology andManagement 74171-180
Marschner H 1995 Mineral Nutrition of Higher Plants 2ndEdn London UK Academic Press 889 pp
Mengel K Kirkby A E 1987 Principles of Plant Nutrition4th Edn Bern Switzerland International Potash Institute593 pp
Mizrahi Y Nerd A 1999 Climbing and columnar cacti newarid land fruit crops In Perspectives on New Crops and NewUses pp 358-366 Ed J Janick Alexandria Virginia USAASHS Press
Nerd A Nobel P S 1995 Accumulation partitioning andassimilation of nitrate in Opuntia ficus-indica Journal ofPlant Nutrition 182533-2549
Nerd A Aronson J A Mizrahi Y 1990 Introduction anddomestication of rare and wild fruit and nut trees for desertareas In Advances in New Crops pp 353-363 Eds J Janickand J E Simon Portland Oregon USA Timber Press
Nerd A Mesika R Mizrahi Y 1993 Effect of N fertilizer onautumn floral flush and cladode N in prickly pear (Opuntiaficus-indica (L) Mill) Journal of Horticultural Science68337-342
Nerd A Tel-Zur N Mizrahi Y 2002 Fruits of vine andcolumnar cacti In Cacti Biology and Uses pp 185-197Ed P S Nobel Berkeley California University of CaliforniaPress
Nobel P S 1988 Environmental Biology of Agaves and CactiNew York USA Cambridge University Press 270 pp
Nobel P S 1989 A nutrient index quantifying productivity ofagaves and cacti Applied Ecology 26635-645
Nobel P S 1991 Tansley Review No 32 Achievableproductivities of certain CAM plants basis for high valuescompared with C
3 and C
4 plants New Phytologist 119183-
205Nobel P S 1999 Physicochemical and Environmental Plant
Physiology 2nd Edn San Diego California USA AcademicPress 474 pp
Nobel P S Bobich E G 2002 Environmental biology In CactiBiology and Uses pp 57-74 Ed P S Nobel BerkeleyCalifornia USA University of California Press
Nobel P S De la Barrera E 2002a Stem water relations andnet CO
2 uptake for a hemiepiphytic cactus during short-term
drought Environmental and Experimental Botany 48129-137
Nobel P S De la Barrera E 2002b High temperatures and netCO2 uptake growth and stem damage for the hemiepiphyticcactus Hylocereus undatus Biotropica 34225-231
Nobel P S De la Barrera E 2002c Nitrogen relations for netCO 2 uptake by the cultivated hemiepiphytic cactusHylocereus undatus Scientia Horticulturae 96281-292
Nobel P S De la Barrera E Beilman D W Doherty J HZutta B R 2002 Temperature limitations for cultivation ofedible cacti in California Madrontildeo 49228-236
Nobel P S Wang N Balsamo R A Loik M E Hawke M A1995 Low-temperature tolerance and acclimation of Opuntiaspp after injecting glucose or methylglucose InternationalJournal of Plant Sciences 156496-504
Norman J M Campbell G S 1989 Canopy structure In PlantPhysiological Ecology Field Methods and Instrumentationpp 301-325 Eds R W Pearcy J Ehleringer H A Mooney andP W Rundel New York USA Chapman and Hall
North G B Nobel P S 1994 Changes in root hydraulicconductivity for two tropical epiphytic cacti as soil moisturevaries American Journal of Botany 8146-53
Onweume I C 1979 Rapid plant-conserving estimation of heattolerance in plants Journal of Agricultural ScienceCambridge 92527-536
Ortiacutez H Y D Livera M M Carrillo S J A 1996 Asimilacioacutende CO
2 en tallos joacutevenes de pitahaya (Hylocereus undatus)
Revista Fitotecnia Mexicana 1931-41Ortiacutez H Y D 1999 Pitahaya un Nuevo Cultivo para Meacutexico
Pitahaya a New Crop for Mexico Balderas Mexico CityMexico Editorial Limusa Grupo Noriega Editores 111 pp
Osmond C B 1978 Crassulacean acid metabolism a curiosityin context Annual Review of Plant Physiology 26379-414
Pimienta E Hernandez G Domingues A Nobel P S 1998Growth and development of the arborescent cactusStenocereus queretaroensis in a subtropical semiaridenvironment including effects of gibberellic acid TreePhysiology 1859-64
Raveh E Weiss J Nerd A Mizrahi Y 1993 Pitayas (GenusHylocereus) a new fruit crop for the Negev Desert of IsraelIn New Crops pp 491-495 Eds J Janick and J E SimonNew York USA Wiley
Raveh E Gersani M Nobel P S 1995 CO2 uptake and
fluorescence responses for a shade-tolerant cactus Hylocereusundatus under current and doubled CO
2 concentrations
Physiologia Plantarum 93505-511Raveh E Nerd A Mizrahi Y 1998 Responses of two
hemiepiphytic fruit-crop cacti to different degrees of shadeScientia Horticulturae 73151-164
Rzedowski J 1983 Vegetacioacuten de Meacutexico Mexico CityMexico Editorial Limusa 432 pp
Schimel D Ives D Enting I Heimann M Joos F RaynaudD Wigley T 1996 CO2 and the carbon cycle In ClimateChange 1995 IPCC pp 65-131 Eds J T Houghton L GMeira Pilho B A Callendar N Harris A Kattenberg and KMaskell Cambridge UK Cambridge University Press
Stitt M Krapp A 1999 The interaction between elevatedcarbon dioxide and nitrogen nutrition the physiological andmolecular background Plant Cell and Environment 22583-621
Vaacutezquez J A Cuevas R Cochrane T S Iltis H H Santana FJ Guzmaacuten L 1995 Flora de Manantlaacuten Plantas Vascularesde la Reserva de la Bioacutesfera Sierra de Manantlaacuten Jalisco-Colima Meacutexico SIDA Botanical Miscellany No 13 FortWorth Texas USA Botanical Research Institute of Texas312 pp
Winter K Smith J A C 1996 An introduction to Crassulaceanacid metabolism Biochemical principles and ecologicaldiversity In Crassulacean Acid Metabolism pp 1-3 Eds KWinter and J A C Smith Berlin Germany Springer
Zotz G Hietz P 2002 The physiological ecology of vascularepiphytes current knowledge open questions Journal ofExperimental Botany 522067-2078

2 PARK S NOBEL amp ERICK DE LA BARRERA
The native distribution of pitahayas (Fig 1) ismostly restricted to tropical dry forests (Barbeau1990) where mean daily air temperature is near24degC ranging from 21degC to 26degC (Fig 2A) Rainfallis usually more than 600 mm year-1 (Rzedowski1983) most of which occurs during the summerwhile the spring is essentially dry (Fig 2B) Suchseasonality for water availability leads to seasonaldifferences in canopy foliage For instance the leafarea index (LAI the ratio of total leaf area to unitground surface area Norman amp Campbell 1989)for the upper canopy is three-fold greater during thewet season than at the end of the dry season (Fig2C) averaging 19 throughout the year A seasonallyvarying PPF occurs under the upper canopyreflecting the changes in water availability and henceLAI instantaneous PPF under the canopy is 50 ofthe maximum during the dry season but only about25 of the maximum during the wet season (Fig2C)
Crassulacean Acid Metabolism
The net CO2 uptake patterns by the stems of
Hylocereus undatus over 24-h periods arecharacteristic of plants employing the Crassulaceanacid metabolism (CAM) pathway of photosynthesisa pathway used by 7 of the nearly 300 000 speciesof vascular plants (Nobel 1991 Winter amp Smith1996) As for other CAM species net CO
2 uptake
by H undatus occurs mainly at night (Fig 3) Inparticular under wet conditions and moderatetemperatures about 85 of its net CO
2 uptake is at
night which is the major period for its stomatalopening (Fig 3 Ortiacutez et al 1996 Ortiacutez 1999) Thisdaynight inversion of the CO
2 uptake pattern
compared with the majority of plants minimiseswater loss by CAM plants About 92 of plantspecies use the C
3 pathway (initial fixation of CO
2catalysed by ribulose-15-bisphosphate carboxylaseoxygenase leading to the formation of a three-carboncompound) and 1 use the C
4 pathway (initial
fixation of CO2 catalysed by phosphoenolpyruvate
carboxylase leading to the formation of a four-carbon compound) such plants take up CO
2 only
during the daytime CO2 uptake at night (Phase I for
CAM plants Osmond 1978 Nobel 1988 1999)involves a binding of CO
2 to a three-carbon
compound catalysed by phosphoenolpyruvatecarboxylase leading to the accumulation of organicacids that are stored in the vacuoles of photosyntheticcells Some net CO
2 uptake generally occurs early
in the morning for H undatus (Fig 3 Phase II) andinvolves the binding of CO
2 by ribulose-15-
bisphosphate carboxylaseoxygenase and also in thelate afternoon (Phase IV) whereas CO
2 tends to be
released during the middle of the daytime (PhaseIII) when the organic acids formed during theprevious night are decarboxylated leading to a highCO
2 concentration within the stem
Air and stem temperatures tend to be lower atnight which is the key to appreciating the water-conserving aspects of CAM Lower temperatureslead to less water loss because air in the intercellularair spaces of the stems is essentially saturated withwater vapour the saturation water vapour pressure
Fig 1 Countries where Hylocereus undatus is currently being cultivated for fruit (back slanted hatching) and whereit as well as other cacti bearing pitahayas are native (forward slanted hatching) Data are from Britton amp Rose (1963)Bravo (1978) Backeberg (1966) Barthlott amp Hunt (1993) Mizrahi amp Nerd (1999) Nerd et al (2002) Ortiacutez (1999)and P S Nobel (unpublished observations)
3Hemiepiphytic cactus CO2 uptake
is accompanied by a total daily water loss of 53 molm-2 day-1 (Nobel amp De la Barrera 2002a) leading toa water-use efficiency of 232 mmol CO
2 m-2 day-1 53
mol H2O m-2 day-1 or 00044 CO
2H
2O Such a
water-use efficiency for H undatus is characteristicof CAM plants but is two- to five-fold higher thanfor C
3 or C
4 species under comparable conditions
(Nobel 1988 1999)
Water Relations
Why does H undatus employ the water-conserving CAM when it is native to moist regionsActually the microsites occupied by its roots in treesare relatively small and are subject to rapid drying(Andrade amp Nobel 1997) The substrate for anepiphyte growing in the crotch of a tree can becomedry a few days after a rainfall event (Croat 1978Freiberg 1997 Luumlttge 1997 Benzing 1998) Thusmost epiphytes are presumably CAM species withtheir roots occupying a small substrate volume thathas little water holding capacity and consequentlydries rapidly for example most of the approximately20 000 species of orchids probably employ CAM(Winter amp Smith 1996 Luumlttge 1997 Zotz amp Hietz2002) Many of the regions where H undatus iscultivated for fruit (Fig 1) are arid or semiarid whichare climatically quite different from the tropicalforests where it is native so knowledge of its waterrelations are also important horticulturally
Drought commences when the water potential ofthe soil in the root zone becomes less than the waterpotential of the roots so that water uptake isthermodynamically not possible For H undatus
increases nearly exponentially with temperatureand for a given degree of stomatal opening thetranspiration rate depends on the difference in watervapour pressure from the stem to the surroundingair For example the saturation water vapourpressure is 234 kPa at 20degC and 425 kPa at 30degC(Nobel 1999) If the ambient air had a water vapourpressure of 123 kPa (dewpoint temperature of10degC) the force leading to water loss would be threetimes greater at 30degC compared with 20degC [(425 ndash123)(234 ndash 123) = 30] leading to three timesmore transpirational water loss at 30degC than at 20degCfor the same degree of stomatal opening
Because of predominantly nocturnal stomatalopening and hence predominantly nocturnal net CO
2uptake the water-use efficiency (ratio of CO
2 fixed
to water transpired) is typically higher for CAMplants than for C
3 or C
4 plants In particular the total
daily net CO2 uptake for H undatus under wet
conditions can be 232 mmol m-2 day-1 (Fig 3) This
Mea
n ai
r te
mpe
ratu
re(deg
C)
27
10
Phot
osyn
thet
ic p
hoto
nfl
ux (
o
f m
axim
um)
Mea
n m
onth
ly r
ainf
all
(mm
)U
pper
can
opy
leaf
are
ain
dex
26
25
2
24
23
22
21
150
100
50
0
3
1
0
A
B
C
20
30
40
50
60
Jan
Feb
Mar
Apr
May Jun
Jul
Aug Se
pO
ctN
ovD
ec
Month
Fig 2 Representative climate for tropical dry forestsin North and South America where H undatus isnative (A) mean daily air temperature (B) meanmonthly rainfall and (C) upper canopy leaf area index(E ) and understorey available PPF ( ) Data weresynthesised from Bullock (1986) Maas et al (1995)Vaacutezquez et al (1995) and Pimienta et al (1998)
Net
CO
2 up
take
rat
e(m
mol
m-2 s
-1)
0
2
4
6
8
12 16 08 120020 04Time of day (h)
Fig 3 Net CO2 uptake by H undatus over 24-h periods
under wet conditions (E ) or after 10 days of drought( ) The soil water potential at the centre of therooting zone was -02 MPa under wet conditions and-32 MPa after 10 days of drought Daynight airtemperatures were 3020degC and the total daily PPFincident on the 1-yr-old stem surfaces considered was10 mol m-2 day-1 Data are means plusmn SE (n = 5 plants PS Nobel amp E De la Barrera unpublishedmeasurements)
4 PARK S NOBEL amp ERICK DE LA BARRERA
rooted in silica sand the total daily net CO2 uptake
does not change appreciably for the first 7 days ofdrought but decreases by 78 at 17 days (Raveh etal 1995) For H undatus rooted in a sandvermiculite mixture the net CO
2 uptake ability
decreases by 57 at 10 days of drought (Fig 3)Also the maximal rate of net CO
2 uptake during
drought tends to occur later in the night (Fig 3)when stem temperatures generally are lower andhence transpiration is lower for a given degree ofstomatal opening
The water vapour conductance for H undatusdecreases during 10 days of drought indicatingstomatal closure but the stem water potential doesnot decrease significantly (Nobel amp De la Barrera2002a) A chemical message probably abscisic acidis apparently sent from the roots in the drying soilto the shoots leading to stomatal closure as a meansfor conserving water Moreover the stomatalresponse to this water-stress hormone prevents theloss of stem water which may be crucial forepiphytic and hemiepiphytic cacti whose rootsexplore only a relatively small soil volume Whenthe roots of H undatus are excised so that no abscisicacid can move from the roots to the shoot duringdrought the decline in net CO
2 uptake is much
slower (50 inhibition is delayed by approximately8 days compared with plants with their roots intact)emphasising the importance of this hormone forconserving stem water during drought (Nobel amp Dela Barrera 2002a)
When H undatus that has been droughted for 10days is rewatered a significant increase in net CO
2uptake occurs in 1 day the CO
2 uptake ability is
half restored in only 2 days and full recovery occursin 7 days (P S Nobel and E De la Barreraunpublished observations) Thus this cactus canrespond rapidly to rainfall events which may becrucial for its growth in tropical regions with frequentrainfalls (Freiberg 1997 Luumlttge 1997) as well asof importance in developing irrigation scheduleswhen it is cultivated as a crop (Mizrahi amp Nerd1999 Nerd et al 2002) Similarly the hydraulicconductivity of the roots (a direct measure of plantwater uptake ability) of two epiphytic cactiEpiphyllum phyllanthus and Rhipsalis bacciferadecreases during drought but increases to the valuesunder wet conditions only 3 days after rewatering(North amp Nobel 1994)
Temperature
Temperature influences nearly every plant processincluding net CO
2 uptake and hence growth The
optimal daynight air temperatures for total daily netCO
2 uptake by Hylocereus undatus are 3020degC (Fig
4) which are relatively high compared withtemperatures optimal for other CAM plants Because
most net CO2 uptake occurs at night the nighttime
temperatures are more crucial for total daily net CO2
uptake than are the daytime ones The optimalnighttime temperatures for total daily net CO
2 uptake
by cacti native to arid or semiarid regions are from10degC to 15degC (Nobel 1988) Apparently the higheroptimal nighttime temperatures for H undatusreflect temperatures characteristic of the primarilytropical regions where it is native (Fig 1) The totaldaily net CO
2 uptake is half of the maximal value at
low daynight temperatures of 188degC and at highdaynight temperatures of 3828degC (Fig 4) Abovedaynight air temperatures of 4232degC a net loss ofCO
2 occurs over a 24-h period (Fig 4) indicating
that this species is not adapted to such hightemperatures
Temperature also affects the survival of cacti andH undatus is more sensitive to temperature extremesthan are most other cacti Based on an assay usingthe uptake of the vital dye neutral red into the centralvacuole of a cell (Onweume 1979 Nobel et al1995) half of the photosynthetic (chlorenchyma)cells are killed by a 1-h low-temperature treatmentat -15degC for H undatus maintained at daynighttemperatures of 2010degC and half are killed at -13degCfor plants maintained at 3020degC (Nobel et al 2002)For 19 other species of cacti half of thephotosynthetic cells are killed at an averagetemperature of -102degC for plants maintained at 2010degC and the low-temperature acclimation orhardening (decrease in lethal temperatures as the daynight temperatures are reduced) averages 07degC asthe daynight temperatures are reduced by 10degC(Nobel 1988) much greater than the 02degC observedfor H undatus Again using the uptake of neutralred into the cell central vacuoles as the assay halfof the photosynthetic cells of H undatus are killedby a 1-h high-temperature treatment at 540degC forplants at 2515degC and at 554degC for plants at 3525degC (Nobel amp De la Barrera 2002b) For 18 otherspecies of cacti half of the photosynthetic cells arekilled at an average temperature of 570degC for plantsmaintained at 3525degC and the high temperatureacclimation (as the daynight temperatures are raisedby 10degC) averages 53degC (Nobel 1988) comparedwith only 14degC for H undatus Thus both the low-temperature and the high-temperature acclimationswhich are the crucial factors for the tolerance ofextreme temperatures (Nobel 1988 Nobel amp De laBarrera 2002b) are less than one-third as great forH undatus as is the average for other cacti
Time of exposure to extreme temperatures is alsocrucial when examining tolerances especially forhigh temperatures For instance H undatus is notexpected to tolerate daynight air temperatures above4232degC on a prolonged basis as the total daily netCO
2 uptake would then be negative (Fig 4) In this
regard the maximal temperatures in the native
5Hemiepiphytic cactus CO2 uptake
habitats of H undatus generally do not exceed 35degC(Croat 1978 Freiberg 1997 Luumlttge 1997 Benzing1998) Maintaining H undatus at 4030degC graduallyleads to visible damage of the stems which firstbecomes evident at 6 wk (Nobel amp De la Barrera2002b) Necrosis increases to about half of the stemsurface area at 15 wk and no new stem growth isobserved at such temperatures Also daytimetemperatures of 45degC in the field greatly reduceflower production by H undatus and brief exposureto 50degC leads to stem necrosis (Mizrahi amp Nerd1999) Thus this species cannot be cultivated wheredaytime maximal temperatures average 40degC forseveral weeks or occasionally exceed 45degC Indeedtotal daily net CO
2 uptake by H undatus is 37
lower after 30 wk at 3525degC compared with at 10wk (Nobel amp De la Barrera 2002b) suggesting thatH undatus does not have phenotypic plasticity totolerate temperatures exceeding those in its nativehabitat for prolonged periods
Photosynthetic Photon Flux
Hylocereus undatus has considerable net CO2
uptake at a total daily PPF of only 2 mol m-2 day-1
(Fig 5) indicating that it is well adapted to the low-light conditions of a tropical forest (total daily netCO
2 uptake is negative for desert CAM species at
this PPF Nobel 1988) [Net CO2 uptake responses
to PPF for CAM plants are not expressed versus theinstantaneous level of such light as is conventionalfor C
3 and C
4 plants but rather versus the total daily
PPF as most CO2 uptake occurs at night in Phase I
(Fig 3) when no light is available] Total daily netCO
2 uptake by H undatus increases as the total daily
PPF increases to about 20 mol m-2 day-1 above whichnet CO
2 uptake decreases with increasing PPF (Fig
5) indicating that high PPF levels damage the
photosynthetic apparatus Total daily net CO2 uptake
for H undatus is about 90 of maximal at only 10mol m-2 day -1 (Fig 5) For CAM plants such as barrelcacti (eg Ferocactus acanthodes) andplatyopuntias (eg Opuntia ficus-indica) a totaldaily PPF of 10 mol m-2 day-1 leads to about 40 ofthe maximal total daily net CO
2 uptake maximal
uptake for such cacti generally occurs from 30ndash40mol m-2 day-1 (Nobel 1988) levels attained bysurfaces facing east or west on clear days in thesummer
Photoinhibition leading to a reduction in total dailynet CO
2 uptake sets in at a total daily PPF of 20 mol
m-2 day-1 for H undatus (Fig 5) compared withabove 40 mol m-2 day-1 for desert CAM species(Nobel 1988) Measurement of chlorophyllfluorescence for H undatus indicates thatphotosynthetic efficiency (total daily net CO
2 uptake
divided by PPF) actually begins to decrease at arelatively low total daily PPF of 5 mol m-2 day-1
(Raveh et al 1995) To avoid exceeding a total dailyPPF of 20 mol m-2 day-1 a vertical climbing orclambering stem of H undatus would need to beshaded by nearly 40 on a clear day for stemsurfaces facing east or west (north- or south-facingsurfaces would need no shading) In this regard Hundatus H polyrhizus and Selenicereusmegalanthus are commercially grown in Israel undershade cloth that attenuates solar irradiation by 30ndash60 (Raveh et al 1993 1998 Mizrahi amp Nerd1999) Such shading is comparable to that in theforests where such species are native (Benzing 1990Barthlott amp Hunt 1993 Luumlttge 1997) Whenexposed to full sun on clear days H undatus in
Fig 4 Total daily net CO2 uptake by H undatus at the
indicated daynight air temperatures Plants weregrowing under wet conditions and the PPF was 10 molm-2 day-1 Data are from Raveh et al (1995) and Nobelamp De la Barrera (2002b)
Tota
l dai
ly n
et C
O2 u
ptak
e(m
mol
m-2 d
ay-1)
-50
0
100
150
200
50
250
155 2515 453535253020 4030Daynight air temperatures
Fig 5 Total daily net CO2 uptake by H undatus at the
indicated total daily PPF incident on the stem surfacePlants were growing under wet conditions and the daynight temperatures were 2515degC Data are from Ravehet al (1995) and P S Nobel amp E De la Barrera(unpublished measurements for 1-yr-old stems withdata indicated by means plusmn SE for n = 5 plants andnormalised to the previous measurements at a totaldaily PPF of 20 mol m-2 day-1)
Tota
l dai
ly n
et C
O2 u
ptak
e(m
mol
m-2 d
ay-1)
0
100
150
200
50
250
0 5 251510 20Total daily PPF (mol m-2 day-1)
30
6 PARK S NOBEL amp ERICK DE LA BARRERA
Mexico (Ortiacutez 1999) and 23 genotypes ofHylocereus in Israel (Nerd et al 1990) developvisible necrosis on the stems However H undatusis cultivated in southeast Asia such as in the MekongDelta of Vietnam without shading (Nerd et al 2002P S Nobel personal observations) Evidently theprevalent cloud cover in such regions reduces thetotal daily PPF to values that do not lead to stembleaching and necrosis
Nutrients
The effects of soil nutrients on net CO2 uptake of
cacti bearing pitahayas have received little attention(Castillo Martiacutenez et al 1996 Ortiacutez 1999) althoughnutrients are known to have substantial effects onother cacti (Nobel 1988 1989) For instanceapplication of nutrients such as nitrogen can alterthe timing of fruiting for Opuntia ficus-indica(Mizrahi amp Nerd 1999) Application of boron andzinc can increase the number of fruits for H undatusand foliar application of magnesium can increasefruit size (Castillo Martiacutenez et al 1996) Variationsin the applied nitrogen concentration for H undatusrooted in a sandvermiculite mixture for 22 wkaffects its net CO
2 uptake ability as measured by the
maximal nocturnal net CO2 uptake rate 016 mM N
leads to a maximal nocturnal net CO2 uptake rate of
25 micromol m-2 s-1 16 mM N leads to 56 micromol m-2 s-1and 16 mM N leads to 98 micromol m-2 s-1 (Fig 6) thelatter being the N concentration in full-strengthHoagland solution (Hoagland amp Arnon 1950Johnson et al 1957 Epstein 1972) Thus net CO
2uptake for H undatus is highest under a relativelyhigh applied N concentration (8-16 mM Fig 6) asis the case for intensively managed greenhouse crops
and field-grown vegetables and ornamentals (Huett1996 Langhans amp Tibbitts 1997) A maximalnocturnal net CO
2 uptake rate of 10 micromol m-2 s-1 (Fig
6) places H undatus fourth highest among the 28species of cacti whose maximal net CO
2 uptake rates
have been reported (Nobel amp Bobich 2002) Thusthis species has a high potential growth rate underhigh-nutrient conditions
Shifts in stem properties of H undatus as theapplied nitrogen concentration is changed arerelatively slow For instance the halftime for theshift in net CO
2 uptake ability in response to applied
N is 12ndash13 wk (Nobel amp De la Barrera 2002c) After22 wk at 016 mM applied N the stem N level on adry mass basis decreases to only 09 (Nobel amp Dela Barrera 2002c) After 22 wk at an applied Nconcentration of 16 mM stem N is 25 comparableto values in the shoots of cultivated C
3 and C
4 plants
under high-nutrient conditions (Mengel amp Kirkby1987 Marschner 1995) Also the chlorophyll levelof H undatus at an applied N concentration of 016mM for 23 wk is only 029 g m-2 compared with 063g m-2 at 16 mM N (Nobel amp De la Barrera 2002c)H undatus supplied with 16 mM N for 8 wk has astem chlorophyll level of 049 g m-2 (Raveh et al1998) and H polyrhizus has 045 g chlorophyll m-2
under comparable conditions (Raveh et al 1998)Following 23 wk at low N levels the stem net CO
2uptake ability and chlorophyll level both increaseto maximal values when 16 mM N is applied to Hundatus for 11 wk with a halftime for the increaseof 8 wk (Nobel amp De la Barrera 2002c) Thusresponses of H undatus to changes in the applied Nconcentration are slower than for most C
3 and C
4crops and other cacti for which major responses canoccur in 2 wk (Mengel amp Kirkby 1987 Nerd et al1993 Marschner 1995 Nerd amp Nobel 1995)suggesting lack of genetic plasticity in this recentlydomesticated cactus or relatively slow nutrientuptake by the roots Because the maximal net CO
2uptake rates for H undatus are similar at applied Nconcentrations of 8 and 16 mM (Fig 6) the latterlevel may lead to luxury (excess) N consumptionas the tissue level after 22 wk is 36 higher at 16mM than at 8 mM applied N (Nobel amp De la Barrera2002c)
Elevated Atmospheric CO2
Another environmental variable influencing netCO
2 uptake by cacti is the atmospheric CO
2concentration which currently is increasing bynearly 3 parts per million (09) per year (Schimelet al 1996 Stitt amp Krapp 1999) For all 20 CAMspecies studied biomass productivity increases fora doubled atmospheric CO
2 concentration compared
to the current atmospheric CO2 concentration
(Drennan amp Nobel 2000) The increase averages
Fig 6 Maximal net CO2 uptake rate at night for H
undatus maintained for 22 wk at the indicated nitrogenconcentration Plants were growing under wetconditions the daynight air temperatures were 3020degC and the total daily PPF was 10 mol m-2 day-1The maximal rate occurred between 2200 h and 0200h Data are from Nobel amp De la Barrera (2002c)
Max
imal
net
CO
2 up
take
rat
e(m
mol
m-2 d
ay-1)
4
6
8
2
10
0 2 1464 8Nitrogen concentration (mM)
160
7Hemiepiphytic cactus CO2 uptake
35 at 700 ppm (700 micro litre litre -1 or 70 kPa)compared with 350 ppm which represents anapproximately 1 increase in growth per 10 ppmincrease in atmospheric CO
2 concentration for CAM
plants (Nobel 1991) Data for H undatus areconsistent with the results for other CAM plants asa doubled atmospheric CO
2 concentration increases
the total daily net CO2 uptake by 34 (Raveh et al
1995) Most of the increase occurs during thedaytime in Phases II and IV Moreover increases intotal daily net CO
2 uptake under a doubled
atmospheric CO2 concentration compared with the
current atmospheric CO2 concentration are more
prominent under less favorable or even stressfulclimatic conditions For instance the increase in totaldaily net CO
2 uptake for H undatus due to a doubled
atmospheric CO2 concentration is 63 higher at 35
25degC versus 2515degC 90 higher at 24 days ofdrought versus 14 days and 120 higher at a PPFof 30 mol m-2 day-1 versus 20 mol m-2 day-1 (Ravehet al 1995) Thus the responses to atmospheric CO
2increases will not only lead to greater total daily netCO
2 uptake but also will lead to proportionally
greater net CO2 uptake under adverse environmental
conditions
Conclusions and Future Prospects
A striking feature of H undatus is how rapidlynocturnal stomatal opening and net CO
2 uptake
abili ty decrease during drought Such rapidresponses which undoubtedly are related to the oftensmall substrate volume that is occupied by its rootsmay be mediated by abscisic acid Other rootproperties such as the apparently low nutrient uptakerate and the influence of mycorrhizae also needinvestigation Another striking ecophysiologicalfeature of H undatus compared with barrel andcolumnar cacti which is especially pertinent to itscultivation outside its native regions is its poortolerance of high temperatures such as those above40degC Indeed its photosynthetic cells do not showmuch acclimation as the daynight air temperaturesare gradually raised over a period of weeks contraryto the case for other cacti Comparisons at amolecular level between H undatus and other cactishowing substantial high-temperature acclimationcould help elucidate the mechanisms involved forthe latter group The intolerance of H undatus tohigh PPF presumably relates to its shaded naturalhabitat Levels of stem nitrogen and chlorophyllcorrelate well with the net CO
2 uptake ability of H
undatus providing relatively simple assays for thephysiological status of its stems which changerelatively slowly in response to changes in theconcentration of applied nitrogen Indeedunderstanding how climatic factors in general andhormonal factors in particular affect the gas
exchange of H undatus and related species will beinteresting ecologically and extremely importanthorticulturally for the expanding cultivation ofpitahayas
Acknowledgements
We thank the UCLA-Ben Gurion UniversityProgram of Cooperation through the gift of Dr SolLeshin and the US-Israel Binational AgriculturalResearch and Development Fund grant no IS-3382-01 for financial support
References
Anderson E F 2001 The Cactus Family Portland OregonUSA Timber Press 776 pp
Andrade J L Nobel P S 1997 Microhabitats and waterrelations of epiphytic cacti and ferns in a lowland neotropicalforest Biotropica 29261-270
Backeberg C 1966 Das Kakteenlexikon Jena GermanyGustav Fisher 828 pp
Barbeau G 1990 La pitahaya rouge un nouveau fruit exotiqueFruits 45141-147
Barthlott W Hunt D R 1993 Cactaceae In The Families andGenera of Vascular Plants Vol 2 pp 161-196 Ed KKubitzki Berlin Germany Springer-Verlag
Benzing D H 1990 Vascular Epiphytes General Biology andRelated Biota Cambridge UK Cambridge University Press354 pp
Benzing D H 1998 Vulnerabilities of tropical forests to climatechange The significance of resident epiphytes ClimateChange 39519-540
Bravo H H 1978 Las Cactaacuteceas de Meacutexico Vol I MexicoCity Mexico UNAM 743 pp
Britton N L Rose J N 1963 The Cactaceae Descriptionsand Illustrations of Plants of the Cactus Family Vol II NewYork USA Dover 241 pp
Bullock S H 1986 Climate of Chamela Jalisco and trends inthe south coastal region of Mexico Archives for MeteorologyGeophysics and Bioclimatology Series B 36297-316
Castillo Martiacutenez R Caacutelix De Dios H Rodriacuteguez Canto A1996 Guia Teacutecnica para el Cultivo de Pitahaya ChetumalQuintana Roo Mexico Universidad de Quintana Roo 158pp
Croat T B 1978 Flora of Barro Colorado Island StanfordCalifornia USA Stanford University Press 943 pp
Drennan P M Nobel P S 2000 Responses of CAM species toincreasing atmospheric CO
2 concentrations Plant Cell and
Environment 23767-781Epstein E 1972 Mineral Nutrition of Plants Principles and
Perspectives New York USA Wiley 412 ppFreiberg M 1997 Spatial and temporal pattern of temperature
and humidity of a tropical premontane rain forest tree in CostaRica Selbyana 1877-84
Hoagland D R Arnon D I 1950 The water-culture methodfor growing plants without soil California AgriculturalExperimental Station Circular 3471-32
Huett D O 1996 Prospects for manipulating the vegetative-reproductive balance in horticultural crops through nitrogennutrition A review Australian Journal of AgriculturalResearch 4747-66
Johnson C M Stout P R Broyer T C Carlton A B 1957Comparative chlorine requirements of different plant speciesPlant Soil 8337-353
Langhans R W Tibbitts T W 1997 Plant growth chamberhandbook Ames Iowa USA Iowa State University 240 pp
8 PARK S NOBEL amp ERICK DE LA BARRERA
Luumlttge U 1997 Physiological Ecology of Tropical PlantsBerlin Germany Springer-Verlag 384 pp
Maas J M Vose J M Swank W T Martiacutenez-Yriacutezar A 1995Seasonal changes of leaf area index (LAI) in a tropicaldeciduous forest in west Mexico Forest Ecology andManagement 74171-180
Marschner H 1995 Mineral Nutrition of Higher Plants 2ndEdn London UK Academic Press 889 pp
Mengel K Kirkby A E 1987 Principles of Plant Nutrition4th Edn Bern Switzerland International Potash Institute593 pp
Mizrahi Y Nerd A 1999 Climbing and columnar cacti newarid land fruit crops In Perspectives on New Crops and NewUses pp 358-366 Ed J Janick Alexandria Virginia USAASHS Press
Nerd A Nobel P S 1995 Accumulation partitioning andassimilation of nitrate in Opuntia ficus-indica Journal ofPlant Nutrition 182533-2549
Nerd A Aronson J A Mizrahi Y 1990 Introduction anddomestication of rare and wild fruit and nut trees for desertareas In Advances in New Crops pp 353-363 Eds J Janickand J E Simon Portland Oregon USA Timber Press
Nerd A Mesika R Mizrahi Y 1993 Effect of N fertilizer onautumn floral flush and cladode N in prickly pear (Opuntiaficus-indica (L) Mill) Journal of Horticultural Science68337-342
Nerd A Tel-Zur N Mizrahi Y 2002 Fruits of vine andcolumnar cacti In Cacti Biology and Uses pp 185-197Ed P S Nobel Berkeley California University of CaliforniaPress
Nobel P S 1988 Environmental Biology of Agaves and CactiNew York USA Cambridge University Press 270 pp
Nobel P S 1989 A nutrient index quantifying productivity ofagaves and cacti Applied Ecology 26635-645
Nobel P S 1991 Tansley Review No 32 Achievableproductivities of certain CAM plants basis for high valuescompared with C
3 and C
4 plants New Phytologist 119183-
205Nobel P S 1999 Physicochemical and Environmental Plant
Physiology 2nd Edn San Diego California USA AcademicPress 474 pp
Nobel P S Bobich E G 2002 Environmental biology In CactiBiology and Uses pp 57-74 Ed P S Nobel BerkeleyCalifornia USA University of California Press
Nobel P S De la Barrera E 2002a Stem water relations andnet CO
2 uptake for a hemiepiphytic cactus during short-term
drought Environmental and Experimental Botany 48129-137
Nobel P S De la Barrera E 2002b High temperatures and netCO2 uptake growth and stem damage for the hemiepiphyticcactus Hylocereus undatus Biotropica 34225-231
Nobel P S De la Barrera E 2002c Nitrogen relations for netCO 2 uptake by the cultivated hemiepiphytic cactusHylocereus undatus Scientia Horticulturae 96281-292
Nobel P S De la Barrera E Beilman D W Doherty J HZutta B R 2002 Temperature limitations for cultivation ofedible cacti in California Madrontildeo 49228-236
Nobel P S Wang N Balsamo R A Loik M E Hawke M A1995 Low-temperature tolerance and acclimation of Opuntiaspp after injecting glucose or methylglucose InternationalJournal of Plant Sciences 156496-504
Norman J M Campbell G S 1989 Canopy structure In PlantPhysiological Ecology Field Methods and Instrumentationpp 301-325 Eds R W Pearcy J Ehleringer H A Mooney andP W Rundel New York USA Chapman and Hall
North G B Nobel P S 1994 Changes in root hydraulicconductivity for two tropical epiphytic cacti as soil moisturevaries American Journal of Botany 8146-53
Onweume I C 1979 Rapid plant-conserving estimation of heattolerance in plants Journal of Agricultural ScienceCambridge 92527-536
Ortiacutez H Y D Livera M M Carrillo S J A 1996 Asimilacioacutende CO
2 en tallos joacutevenes de pitahaya (Hylocereus undatus)
Revista Fitotecnia Mexicana 1931-41Ortiacutez H Y D 1999 Pitahaya un Nuevo Cultivo para Meacutexico
Pitahaya a New Crop for Mexico Balderas Mexico CityMexico Editorial Limusa Grupo Noriega Editores 111 pp
Osmond C B 1978 Crassulacean acid metabolism a curiosityin context Annual Review of Plant Physiology 26379-414
Pimienta E Hernandez G Domingues A Nobel P S 1998Growth and development of the arborescent cactusStenocereus queretaroensis in a subtropical semiaridenvironment including effects of gibberellic acid TreePhysiology 1859-64
Raveh E Weiss J Nerd A Mizrahi Y 1993 Pitayas (GenusHylocereus) a new fruit crop for the Negev Desert of IsraelIn New Crops pp 491-495 Eds J Janick and J E SimonNew York USA Wiley
Raveh E Gersani M Nobel P S 1995 CO2 uptake and
fluorescence responses for a shade-tolerant cactus Hylocereusundatus under current and doubled CO
2 concentrations
Physiologia Plantarum 93505-511Raveh E Nerd A Mizrahi Y 1998 Responses of two
hemiepiphytic fruit-crop cacti to different degrees of shadeScientia Horticulturae 73151-164
Rzedowski J 1983 Vegetacioacuten de Meacutexico Mexico CityMexico Editorial Limusa 432 pp
Schimel D Ives D Enting I Heimann M Joos F RaynaudD Wigley T 1996 CO2 and the carbon cycle In ClimateChange 1995 IPCC pp 65-131 Eds J T Houghton L GMeira Pilho B A Callendar N Harris A Kattenberg and KMaskell Cambridge UK Cambridge University Press
Stitt M Krapp A 1999 The interaction between elevatedcarbon dioxide and nitrogen nutrition the physiological andmolecular background Plant Cell and Environment 22583-621
Vaacutezquez J A Cuevas R Cochrane T S Iltis H H Santana FJ Guzmaacuten L 1995 Flora de Manantlaacuten Plantas Vascularesde la Reserva de la Bioacutesfera Sierra de Manantlaacuten Jalisco-Colima Meacutexico SIDA Botanical Miscellany No 13 FortWorth Texas USA Botanical Research Institute of Texas312 pp
Winter K Smith J A C 1996 An introduction to Crassulaceanacid metabolism Biochemical principles and ecologicaldiversity In Crassulacean Acid Metabolism pp 1-3 Eds KWinter and J A C Smith Berlin Germany Springer
Zotz G Hietz P 2002 The physiological ecology of vascularepiphytes current knowledge open questions Journal ofExperimental Botany 522067-2078
3Hemiepiphytic cactus CO2 uptake
is accompanied by a total daily water loss of 53 molm-2 day-1 (Nobel amp De la Barrera 2002a) leading toa water-use efficiency of 232 mmol CO
2 m-2 day-1 53
mol H2O m-2 day-1 or 00044 CO
2H
2O Such a
water-use efficiency for H undatus is characteristicof CAM plants but is two- to five-fold higher thanfor C
3 or C
4 species under comparable conditions
(Nobel 1988 1999)
Water Relations
Why does H undatus employ the water-conserving CAM when it is native to moist regionsActually the microsites occupied by its roots in treesare relatively small and are subject to rapid drying(Andrade amp Nobel 1997) The substrate for anepiphyte growing in the crotch of a tree can becomedry a few days after a rainfall event (Croat 1978Freiberg 1997 Luumlttge 1997 Benzing 1998) Thusmost epiphytes are presumably CAM species withtheir roots occupying a small substrate volume thathas little water holding capacity and consequentlydries rapidly for example most of the approximately20 000 species of orchids probably employ CAM(Winter amp Smith 1996 Luumlttge 1997 Zotz amp Hietz2002) Many of the regions where H undatus iscultivated for fruit (Fig 1) are arid or semiarid whichare climatically quite different from the tropicalforests where it is native so knowledge of its waterrelations are also important horticulturally
Drought commences when the water potential ofthe soil in the root zone becomes less than the waterpotential of the roots so that water uptake isthermodynamically not possible For H undatus
increases nearly exponentially with temperatureand for a given degree of stomatal opening thetranspiration rate depends on the difference in watervapour pressure from the stem to the surroundingair For example the saturation water vapourpressure is 234 kPa at 20degC and 425 kPa at 30degC(Nobel 1999) If the ambient air had a water vapourpressure of 123 kPa (dewpoint temperature of10degC) the force leading to water loss would be threetimes greater at 30degC compared with 20degC [(425 ndash123)(234 ndash 123) = 30] leading to three timesmore transpirational water loss at 30degC than at 20degCfor the same degree of stomatal opening
Because of predominantly nocturnal stomatalopening and hence predominantly nocturnal net CO
2uptake the water-use efficiency (ratio of CO
2 fixed
to water transpired) is typically higher for CAMplants than for C
3 or C
4 plants In particular the total
daily net CO2 uptake for H undatus under wet
conditions can be 232 mmol m-2 day-1 (Fig 3) This
Mea
n ai
r te
mpe
ratu
re(deg
C)
27
10
Phot
osyn
thet
ic p
hoto
nfl
ux (
o
f m
axim
um)
Mea
n m
onth
ly r
ainf
all
(mm
)U
pper
can
opy
leaf
are
ain
dex
26
25
2
24
23
22
21
150
100
50
0
3
1
0
A
B
C
20
30
40
50
60
Jan
Feb
Mar
Apr
May Jun
Jul
Aug Se
pO
ctN
ovD
ec
Month
Fig 2 Representative climate for tropical dry forestsin North and South America where H undatus isnative (A) mean daily air temperature (B) meanmonthly rainfall and (C) upper canopy leaf area index(E ) and understorey available PPF ( ) Data weresynthesised from Bullock (1986) Maas et al (1995)Vaacutezquez et al (1995) and Pimienta et al (1998)
Net
CO
2 up
take
rat
e(m
mol
m-2 s
-1)
0
2
4
6
8
12 16 08 120020 04Time of day (h)
Fig 3 Net CO2 uptake by H undatus over 24-h periods
under wet conditions (E ) or after 10 days of drought( ) The soil water potential at the centre of therooting zone was -02 MPa under wet conditions and-32 MPa after 10 days of drought Daynight airtemperatures were 3020degC and the total daily PPFincident on the 1-yr-old stem surfaces considered was10 mol m-2 day-1 Data are means plusmn SE (n = 5 plants PS Nobel amp E De la Barrera unpublishedmeasurements)
4 PARK S NOBEL amp ERICK DE LA BARRERA
rooted in silica sand the total daily net CO2 uptake
does not change appreciably for the first 7 days ofdrought but decreases by 78 at 17 days (Raveh etal 1995) For H undatus rooted in a sandvermiculite mixture the net CO
2 uptake ability
decreases by 57 at 10 days of drought (Fig 3)Also the maximal rate of net CO
2 uptake during
drought tends to occur later in the night (Fig 3)when stem temperatures generally are lower andhence transpiration is lower for a given degree ofstomatal opening
The water vapour conductance for H undatusdecreases during 10 days of drought indicatingstomatal closure but the stem water potential doesnot decrease significantly (Nobel amp De la Barrera2002a) A chemical message probably abscisic acidis apparently sent from the roots in the drying soilto the shoots leading to stomatal closure as a meansfor conserving water Moreover the stomatalresponse to this water-stress hormone prevents theloss of stem water which may be crucial forepiphytic and hemiepiphytic cacti whose rootsexplore only a relatively small soil volume Whenthe roots of H undatus are excised so that no abscisicacid can move from the roots to the shoot duringdrought the decline in net CO
2 uptake is much
slower (50 inhibition is delayed by approximately8 days compared with plants with their roots intact)emphasising the importance of this hormone forconserving stem water during drought (Nobel amp Dela Barrera 2002a)
When H undatus that has been droughted for 10days is rewatered a significant increase in net CO
2uptake occurs in 1 day the CO
2 uptake ability is
half restored in only 2 days and full recovery occursin 7 days (P S Nobel and E De la Barreraunpublished observations) Thus this cactus canrespond rapidly to rainfall events which may becrucial for its growth in tropical regions with frequentrainfalls (Freiberg 1997 Luumlttge 1997) as well asof importance in developing irrigation scheduleswhen it is cultivated as a crop (Mizrahi amp Nerd1999 Nerd et al 2002) Similarly the hydraulicconductivity of the roots (a direct measure of plantwater uptake ability) of two epiphytic cactiEpiphyllum phyllanthus and Rhipsalis bacciferadecreases during drought but increases to the valuesunder wet conditions only 3 days after rewatering(North amp Nobel 1994)
Temperature
Temperature influences nearly every plant processincluding net CO
2 uptake and hence growth The
optimal daynight air temperatures for total daily netCO
2 uptake by Hylocereus undatus are 3020degC (Fig
4) which are relatively high compared withtemperatures optimal for other CAM plants Because
most net CO2 uptake occurs at night the nighttime
temperatures are more crucial for total daily net CO2
uptake than are the daytime ones The optimalnighttime temperatures for total daily net CO
2 uptake
by cacti native to arid or semiarid regions are from10degC to 15degC (Nobel 1988) Apparently the higheroptimal nighttime temperatures for H undatusreflect temperatures characteristic of the primarilytropical regions where it is native (Fig 1) The totaldaily net CO
2 uptake is half of the maximal value at
low daynight temperatures of 188degC and at highdaynight temperatures of 3828degC (Fig 4) Abovedaynight air temperatures of 4232degC a net loss ofCO
2 occurs over a 24-h period (Fig 4) indicating
that this species is not adapted to such hightemperatures
Temperature also affects the survival of cacti andH undatus is more sensitive to temperature extremesthan are most other cacti Based on an assay usingthe uptake of the vital dye neutral red into the centralvacuole of a cell (Onweume 1979 Nobel et al1995) half of the photosynthetic (chlorenchyma)cells are killed by a 1-h low-temperature treatmentat -15degC for H undatus maintained at daynighttemperatures of 2010degC and half are killed at -13degCfor plants maintained at 3020degC (Nobel et al 2002)For 19 other species of cacti half of thephotosynthetic cells are killed at an averagetemperature of -102degC for plants maintained at 2010degC and the low-temperature acclimation orhardening (decrease in lethal temperatures as the daynight temperatures are reduced) averages 07degC asthe daynight temperatures are reduced by 10degC(Nobel 1988) much greater than the 02degC observedfor H undatus Again using the uptake of neutralred into the cell central vacuoles as the assay halfof the photosynthetic cells of H undatus are killedby a 1-h high-temperature treatment at 540degC forplants at 2515degC and at 554degC for plants at 3525degC (Nobel amp De la Barrera 2002b) For 18 otherspecies of cacti half of the photosynthetic cells arekilled at an average temperature of 570degC for plantsmaintained at 3525degC and the high temperatureacclimation (as the daynight temperatures are raisedby 10degC) averages 53degC (Nobel 1988) comparedwith only 14degC for H undatus Thus both the low-temperature and the high-temperature acclimationswhich are the crucial factors for the tolerance ofextreme temperatures (Nobel 1988 Nobel amp De laBarrera 2002b) are less than one-third as great forH undatus as is the average for other cacti
Time of exposure to extreme temperatures is alsocrucial when examining tolerances especially forhigh temperatures For instance H undatus is notexpected to tolerate daynight air temperatures above4232degC on a prolonged basis as the total daily netCO
2 uptake would then be negative (Fig 4) In this
regard the maximal temperatures in the native
5Hemiepiphytic cactus CO2 uptake
habitats of H undatus generally do not exceed 35degC(Croat 1978 Freiberg 1997 Luumlttge 1997 Benzing1998) Maintaining H undatus at 4030degC graduallyleads to visible damage of the stems which firstbecomes evident at 6 wk (Nobel amp De la Barrera2002b) Necrosis increases to about half of the stemsurface area at 15 wk and no new stem growth isobserved at such temperatures Also daytimetemperatures of 45degC in the field greatly reduceflower production by H undatus and brief exposureto 50degC leads to stem necrosis (Mizrahi amp Nerd1999) Thus this species cannot be cultivated wheredaytime maximal temperatures average 40degC forseveral weeks or occasionally exceed 45degC Indeedtotal daily net CO
2 uptake by H undatus is 37
lower after 30 wk at 3525degC compared with at 10wk (Nobel amp De la Barrera 2002b) suggesting thatH undatus does not have phenotypic plasticity totolerate temperatures exceeding those in its nativehabitat for prolonged periods
Photosynthetic Photon Flux
Hylocereus undatus has considerable net CO2
uptake at a total daily PPF of only 2 mol m-2 day-1
(Fig 5) indicating that it is well adapted to the low-light conditions of a tropical forest (total daily netCO
2 uptake is negative for desert CAM species at
this PPF Nobel 1988) [Net CO2 uptake responses
to PPF for CAM plants are not expressed versus theinstantaneous level of such light as is conventionalfor C
3 and C
4 plants but rather versus the total daily
PPF as most CO2 uptake occurs at night in Phase I
(Fig 3) when no light is available] Total daily netCO
2 uptake by H undatus increases as the total daily
PPF increases to about 20 mol m-2 day-1 above whichnet CO
2 uptake decreases with increasing PPF (Fig
5) indicating that high PPF levels damage the
photosynthetic apparatus Total daily net CO2 uptake
for H undatus is about 90 of maximal at only 10mol m-2 day -1 (Fig 5) For CAM plants such as barrelcacti (eg Ferocactus acanthodes) andplatyopuntias (eg Opuntia ficus-indica) a totaldaily PPF of 10 mol m-2 day-1 leads to about 40 ofthe maximal total daily net CO
2 uptake maximal
uptake for such cacti generally occurs from 30ndash40mol m-2 day-1 (Nobel 1988) levels attained bysurfaces facing east or west on clear days in thesummer
Photoinhibition leading to a reduction in total dailynet CO
2 uptake sets in at a total daily PPF of 20 mol
m-2 day-1 for H undatus (Fig 5) compared withabove 40 mol m-2 day-1 for desert CAM species(Nobel 1988) Measurement of chlorophyllfluorescence for H undatus indicates thatphotosynthetic efficiency (total daily net CO
2 uptake
divided by PPF) actually begins to decrease at arelatively low total daily PPF of 5 mol m-2 day-1
(Raveh et al 1995) To avoid exceeding a total dailyPPF of 20 mol m-2 day-1 a vertical climbing orclambering stem of H undatus would need to beshaded by nearly 40 on a clear day for stemsurfaces facing east or west (north- or south-facingsurfaces would need no shading) In this regard Hundatus H polyrhizus and Selenicereusmegalanthus are commercially grown in Israel undershade cloth that attenuates solar irradiation by 30ndash60 (Raveh et al 1993 1998 Mizrahi amp Nerd1999) Such shading is comparable to that in theforests where such species are native (Benzing 1990Barthlott amp Hunt 1993 Luumlttge 1997) Whenexposed to full sun on clear days H undatus in
Fig 4 Total daily net CO2 uptake by H undatus at the
indicated daynight air temperatures Plants weregrowing under wet conditions and the PPF was 10 molm-2 day-1 Data are from Raveh et al (1995) and Nobelamp De la Barrera (2002b)
Tota
l dai
ly n
et C
O2 u
ptak
e(m
mol
m-2 d
ay-1)
-50
0
100
150
200
50
250
155 2515 453535253020 4030Daynight air temperatures
Fig 5 Total daily net CO2 uptake by H undatus at the
indicated total daily PPF incident on the stem surfacePlants were growing under wet conditions and the daynight temperatures were 2515degC Data are from Ravehet al (1995) and P S Nobel amp E De la Barrera(unpublished measurements for 1-yr-old stems withdata indicated by means plusmn SE for n = 5 plants andnormalised to the previous measurements at a totaldaily PPF of 20 mol m-2 day-1)
Tota
l dai
ly n
et C
O2 u
ptak
e(m
mol
m-2 d
ay-1)
0
100
150
200
50
250
0 5 251510 20Total daily PPF (mol m-2 day-1)
30
6 PARK S NOBEL amp ERICK DE LA BARRERA
Mexico (Ortiacutez 1999) and 23 genotypes ofHylocereus in Israel (Nerd et al 1990) developvisible necrosis on the stems However H undatusis cultivated in southeast Asia such as in the MekongDelta of Vietnam without shading (Nerd et al 2002P S Nobel personal observations) Evidently theprevalent cloud cover in such regions reduces thetotal daily PPF to values that do not lead to stembleaching and necrosis
Nutrients
The effects of soil nutrients on net CO2 uptake of
cacti bearing pitahayas have received little attention(Castillo Martiacutenez et al 1996 Ortiacutez 1999) althoughnutrients are known to have substantial effects onother cacti (Nobel 1988 1989) For instanceapplication of nutrients such as nitrogen can alterthe timing of fruiting for Opuntia ficus-indica(Mizrahi amp Nerd 1999) Application of boron andzinc can increase the number of fruits for H undatusand foliar application of magnesium can increasefruit size (Castillo Martiacutenez et al 1996) Variationsin the applied nitrogen concentration for H undatusrooted in a sandvermiculite mixture for 22 wkaffects its net CO
2 uptake ability as measured by the
maximal nocturnal net CO2 uptake rate 016 mM N
leads to a maximal nocturnal net CO2 uptake rate of
25 micromol m-2 s-1 16 mM N leads to 56 micromol m-2 s-1and 16 mM N leads to 98 micromol m-2 s-1 (Fig 6) thelatter being the N concentration in full-strengthHoagland solution (Hoagland amp Arnon 1950Johnson et al 1957 Epstein 1972) Thus net CO
2uptake for H undatus is highest under a relativelyhigh applied N concentration (8-16 mM Fig 6) asis the case for intensively managed greenhouse crops
and field-grown vegetables and ornamentals (Huett1996 Langhans amp Tibbitts 1997) A maximalnocturnal net CO
2 uptake rate of 10 micromol m-2 s-1 (Fig
6) places H undatus fourth highest among the 28species of cacti whose maximal net CO
2 uptake rates
have been reported (Nobel amp Bobich 2002) Thusthis species has a high potential growth rate underhigh-nutrient conditions
Shifts in stem properties of H undatus as theapplied nitrogen concentration is changed arerelatively slow For instance the halftime for theshift in net CO
2 uptake ability in response to applied
N is 12ndash13 wk (Nobel amp De la Barrera 2002c) After22 wk at 016 mM applied N the stem N level on adry mass basis decreases to only 09 (Nobel amp Dela Barrera 2002c) After 22 wk at an applied Nconcentration of 16 mM stem N is 25 comparableto values in the shoots of cultivated C
3 and C
4 plants
under high-nutrient conditions (Mengel amp Kirkby1987 Marschner 1995) Also the chlorophyll levelof H undatus at an applied N concentration of 016mM for 23 wk is only 029 g m-2 compared with 063g m-2 at 16 mM N (Nobel amp De la Barrera 2002c)H undatus supplied with 16 mM N for 8 wk has astem chlorophyll level of 049 g m-2 (Raveh et al1998) and H polyrhizus has 045 g chlorophyll m-2
under comparable conditions (Raveh et al 1998)Following 23 wk at low N levels the stem net CO
2uptake ability and chlorophyll level both increaseto maximal values when 16 mM N is applied to Hundatus for 11 wk with a halftime for the increaseof 8 wk (Nobel amp De la Barrera 2002c) Thusresponses of H undatus to changes in the applied Nconcentration are slower than for most C
3 and C
4crops and other cacti for which major responses canoccur in 2 wk (Mengel amp Kirkby 1987 Nerd et al1993 Marschner 1995 Nerd amp Nobel 1995)suggesting lack of genetic plasticity in this recentlydomesticated cactus or relatively slow nutrientuptake by the roots Because the maximal net CO
2uptake rates for H undatus are similar at applied Nconcentrations of 8 and 16 mM (Fig 6) the latterlevel may lead to luxury (excess) N consumptionas the tissue level after 22 wk is 36 higher at 16mM than at 8 mM applied N (Nobel amp De la Barrera2002c)
Elevated Atmospheric CO2
Another environmental variable influencing netCO
2 uptake by cacti is the atmospheric CO
2concentration which currently is increasing bynearly 3 parts per million (09) per year (Schimelet al 1996 Stitt amp Krapp 1999) For all 20 CAMspecies studied biomass productivity increases fora doubled atmospheric CO
2 concentration compared
to the current atmospheric CO2 concentration
(Drennan amp Nobel 2000) The increase averages
Fig 6 Maximal net CO2 uptake rate at night for H
undatus maintained for 22 wk at the indicated nitrogenconcentration Plants were growing under wetconditions the daynight air temperatures were 3020degC and the total daily PPF was 10 mol m-2 day-1The maximal rate occurred between 2200 h and 0200h Data are from Nobel amp De la Barrera (2002c)
Max
imal
net
CO
2 up
take
rat
e(m
mol
m-2 d
ay-1)
4
6
8
2
10
0 2 1464 8Nitrogen concentration (mM)
160
7Hemiepiphytic cactus CO2 uptake
35 at 700 ppm (700 micro litre litre -1 or 70 kPa)compared with 350 ppm which represents anapproximately 1 increase in growth per 10 ppmincrease in atmospheric CO
2 concentration for CAM
plants (Nobel 1991) Data for H undatus areconsistent with the results for other CAM plants asa doubled atmospheric CO
2 concentration increases
the total daily net CO2 uptake by 34 (Raveh et al
1995) Most of the increase occurs during thedaytime in Phases II and IV Moreover increases intotal daily net CO
2 uptake under a doubled
atmospheric CO2 concentration compared with the
current atmospheric CO2 concentration are more
prominent under less favorable or even stressfulclimatic conditions For instance the increase in totaldaily net CO
2 uptake for H undatus due to a doubled
atmospheric CO2 concentration is 63 higher at 35
25degC versus 2515degC 90 higher at 24 days ofdrought versus 14 days and 120 higher at a PPFof 30 mol m-2 day-1 versus 20 mol m-2 day-1 (Ravehet al 1995) Thus the responses to atmospheric CO
2increases will not only lead to greater total daily netCO
2 uptake but also will lead to proportionally
greater net CO2 uptake under adverse environmental
conditions
Conclusions and Future Prospects
A striking feature of H undatus is how rapidlynocturnal stomatal opening and net CO
2 uptake
abili ty decrease during drought Such rapidresponses which undoubtedly are related to the oftensmall substrate volume that is occupied by its rootsmay be mediated by abscisic acid Other rootproperties such as the apparently low nutrient uptakerate and the influence of mycorrhizae also needinvestigation Another striking ecophysiologicalfeature of H undatus compared with barrel andcolumnar cacti which is especially pertinent to itscultivation outside its native regions is its poortolerance of high temperatures such as those above40degC Indeed its photosynthetic cells do not showmuch acclimation as the daynight air temperaturesare gradually raised over a period of weeks contraryto the case for other cacti Comparisons at amolecular level between H undatus and other cactishowing substantial high-temperature acclimationcould help elucidate the mechanisms involved forthe latter group The intolerance of H undatus tohigh PPF presumably relates to its shaded naturalhabitat Levels of stem nitrogen and chlorophyllcorrelate well with the net CO
2 uptake ability of H
undatus providing relatively simple assays for thephysiological status of its stems which changerelatively slowly in response to changes in theconcentration of applied nitrogen Indeedunderstanding how climatic factors in general andhormonal factors in particular affect the gas
exchange of H undatus and related species will beinteresting ecologically and extremely importanthorticulturally for the expanding cultivation ofpitahayas
Acknowledgements
We thank the UCLA-Ben Gurion UniversityProgram of Cooperation through the gift of Dr SolLeshin and the US-Israel Binational AgriculturalResearch and Development Fund grant no IS-3382-01 for financial support
References
Anderson E F 2001 The Cactus Family Portland OregonUSA Timber Press 776 pp
Andrade J L Nobel P S 1997 Microhabitats and waterrelations of epiphytic cacti and ferns in a lowland neotropicalforest Biotropica 29261-270
Backeberg C 1966 Das Kakteenlexikon Jena GermanyGustav Fisher 828 pp
Barbeau G 1990 La pitahaya rouge un nouveau fruit exotiqueFruits 45141-147
Barthlott W Hunt D R 1993 Cactaceae In The Families andGenera of Vascular Plants Vol 2 pp 161-196 Ed KKubitzki Berlin Germany Springer-Verlag
Benzing D H 1990 Vascular Epiphytes General Biology andRelated Biota Cambridge UK Cambridge University Press354 pp
Benzing D H 1998 Vulnerabilities of tropical forests to climatechange The significance of resident epiphytes ClimateChange 39519-540
Bravo H H 1978 Las Cactaacuteceas de Meacutexico Vol I MexicoCity Mexico UNAM 743 pp
Britton N L Rose J N 1963 The Cactaceae Descriptionsand Illustrations of Plants of the Cactus Family Vol II NewYork USA Dover 241 pp
Bullock S H 1986 Climate of Chamela Jalisco and trends inthe south coastal region of Mexico Archives for MeteorologyGeophysics and Bioclimatology Series B 36297-316
Castillo Martiacutenez R Caacutelix De Dios H Rodriacuteguez Canto A1996 Guia Teacutecnica para el Cultivo de Pitahaya ChetumalQuintana Roo Mexico Universidad de Quintana Roo 158pp
Croat T B 1978 Flora of Barro Colorado Island StanfordCalifornia USA Stanford University Press 943 pp
Drennan P M Nobel P S 2000 Responses of CAM species toincreasing atmospheric CO
2 concentrations Plant Cell and
Environment 23767-781Epstein E 1972 Mineral Nutrition of Plants Principles and
Perspectives New York USA Wiley 412 ppFreiberg M 1997 Spatial and temporal pattern of temperature
and humidity of a tropical premontane rain forest tree in CostaRica Selbyana 1877-84
Hoagland D R Arnon D I 1950 The water-culture methodfor growing plants without soil California AgriculturalExperimental Station Circular 3471-32
Huett D O 1996 Prospects for manipulating the vegetative-reproductive balance in horticultural crops through nitrogennutrition A review Australian Journal of AgriculturalResearch 4747-66
Johnson C M Stout P R Broyer T C Carlton A B 1957Comparative chlorine requirements of different plant speciesPlant Soil 8337-353
Langhans R W Tibbitts T W 1997 Plant growth chamberhandbook Ames Iowa USA Iowa State University 240 pp
8 PARK S NOBEL amp ERICK DE LA BARRERA
Luumlttge U 1997 Physiological Ecology of Tropical PlantsBerlin Germany Springer-Verlag 384 pp
Maas J M Vose J M Swank W T Martiacutenez-Yriacutezar A 1995Seasonal changes of leaf area index (LAI) in a tropicaldeciduous forest in west Mexico Forest Ecology andManagement 74171-180
Marschner H 1995 Mineral Nutrition of Higher Plants 2ndEdn London UK Academic Press 889 pp
Mengel K Kirkby A E 1987 Principles of Plant Nutrition4th Edn Bern Switzerland International Potash Institute593 pp
Mizrahi Y Nerd A 1999 Climbing and columnar cacti newarid land fruit crops In Perspectives on New Crops and NewUses pp 358-366 Ed J Janick Alexandria Virginia USAASHS Press
Nerd A Nobel P S 1995 Accumulation partitioning andassimilation of nitrate in Opuntia ficus-indica Journal ofPlant Nutrition 182533-2549
Nerd A Aronson J A Mizrahi Y 1990 Introduction anddomestication of rare and wild fruit and nut trees for desertareas In Advances in New Crops pp 353-363 Eds J Janickand J E Simon Portland Oregon USA Timber Press
Nerd A Mesika R Mizrahi Y 1993 Effect of N fertilizer onautumn floral flush and cladode N in prickly pear (Opuntiaficus-indica (L) Mill) Journal of Horticultural Science68337-342
Nerd A Tel-Zur N Mizrahi Y 2002 Fruits of vine andcolumnar cacti In Cacti Biology and Uses pp 185-197Ed P S Nobel Berkeley California University of CaliforniaPress
Nobel P S 1988 Environmental Biology of Agaves and CactiNew York USA Cambridge University Press 270 pp
Nobel P S 1989 A nutrient index quantifying productivity ofagaves and cacti Applied Ecology 26635-645
Nobel P S 1991 Tansley Review No 32 Achievableproductivities of certain CAM plants basis for high valuescompared with C
3 and C
4 plants New Phytologist 119183-
205Nobel P S 1999 Physicochemical and Environmental Plant
Physiology 2nd Edn San Diego California USA AcademicPress 474 pp
Nobel P S Bobich E G 2002 Environmental biology In CactiBiology and Uses pp 57-74 Ed P S Nobel BerkeleyCalifornia USA University of California Press
Nobel P S De la Barrera E 2002a Stem water relations andnet CO
2 uptake for a hemiepiphytic cactus during short-term
drought Environmental and Experimental Botany 48129-137
Nobel P S De la Barrera E 2002b High temperatures and netCO2 uptake growth and stem damage for the hemiepiphyticcactus Hylocereus undatus Biotropica 34225-231
Nobel P S De la Barrera E 2002c Nitrogen relations for netCO 2 uptake by the cultivated hemiepiphytic cactusHylocereus undatus Scientia Horticulturae 96281-292
Nobel P S De la Barrera E Beilman D W Doherty J HZutta B R 2002 Temperature limitations for cultivation ofedible cacti in California Madrontildeo 49228-236
Nobel P S Wang N Balsamo R A Loik M E Hawke M A1995 Low-temperature tolerance and acclimation of Opuntiaspp after injecting glucose or methylglucose InternationalJournal of Plant Sciences 156496-504
Norman J M Campbell G S 1989 Canopy structure In PlantPhysiological Ecology Field Methods and Instrumentationpp 301-325 Eds R W Pearcy J Ehleringer H A Mooney andP W Rundel New York USA Chapman and Hall
North G B Nobel P S 1994 Changes in root hydraulicconductivity for two tropical epiphytic cacti as soil moisturevaries American Journal of Botany 8146-53
Onweume I C 1979 Rapid plant-conserving estimation of heattolerance in plants Journal of Agricultural ScienceCambridge 92527-536
Ortiacutez H Y D Livera M M Carrillo S J A 1996 Asimilacioacutende CO
2 en tallos joacutevenes de pitahaya (Hylocereus undatus)
Revista Fitotecnia Mexicana 1931-41Ortiacutez H Y D 1999 Pitahaya un Nuevo Cultivo para Meacutexico
Pitahaya a New Crop for Mexico Balderas Mexico CityMexico Editorial Limusa Grupo Noriega Editores 111 pp
Osmond C B 1978 Crassulacean acid metabolism a curiosityin context Annual Review of Plant Physiology 26379-414
Pimienta E Hernandez G Domingues A Nobel P S 1998Growth and development of the arborescent cactusStenocereus queretaroensis in a subtropical semiaridenvironment including effects of gibberellic acid TreePhysiology 1859-64
Raveh E Weiss J Nerd A Mizrahi Y 1993 Pitayas (GenusHylocereus) a new fruit crop for the Negev Desert of IsraelIn New Crops pp 491-495 Eds J Janick and J E SimonNew York USA Wiley
Raveh E Gersani M Nobel P S 1995 CO2 uptake and
fluorescence responses for a shade-tolerant cactus Hylocereusundatus under current and doubled CO
2 concentrations
Physiologia Plantarum 93505-511Raveh E Nerd A Mizrahi Y 1998 Responses of two
hemiepiphytic fruit-crop cacti to different degrees of shadeScientia Horticulturae 73151-164
Rzedowski J 1983 Vegetacioacuten de Meacutexico Mexico CityMexico Editorial Limusa 432 pp
Schimel D Ives D Enting I Heimann M Joos F RaynaudD Wigley T 1996 CO2 and the carbon cycle In ClimateChange 1995 IPCC pp 65-131 Eds J T Houghton L GMeira Pilho B A Callendar N Harris A Kattenberg and KMaskell Cambridge UK Cambridge University Press
Stitt M Krapp A 1999 The interaction between elevatedcarbon dioxide and nitrogen nutrition the physiological andmolecular background Plant Cell and Environment 22583-621
Vaacutezquez J A Cuevas R Cochrane T S Iltis H H Santana FJ Guzmaacuten L 1995 Flora de Manantlaacuten Plantas Vascularesde la Reserva de la Bioacutesfera Sierra de Manantlaacuten Jalisco-Colima Meacutexico SIDA Botanical Miscellany No 13 FortWorth Texas USA Botanical Research Institute of Texas312 pp
Winter K Smith J A C 1996 An introduction to Crassulaceanacid metabolism Biochemical principles and ecologicaldiversity In Crassulacean Acid Metabolism pp 1-3 Eds KWinter and J A C Smith Berlin Germany Springer
Zotz G Hietz P 2002 The physiological ecology of vascularepiphytes current knowledge open questions Journal ofExperimental Botany 522067-2078

4 PARK S NOBEL amp ERICK DE LA BARRERA
rooted in silica sand the total daily net CO2 uptake
does not change appreciably for the first 7 days ofdrought but decreases by 78 at 17 days (Raveh etal 1995) For H undatus rooted in a sandvermiculite mixture the net CO
2 uptake ability
decreases by 57 at 10 days of drought (Fig 3)Also the maximal rate of net CO
2 uptake during
drought tends to occur later in the night (Fig 3)when stem temperatures generally are lower andhence transpiration is lower for a given degree ofstomatal opening
The water vapour conductance for H undatusdecreases during 10 days of drought indicatingstomatal closure but the stem water potential doesnot decrease significantly (Nobel amp De la Barrera2002a) A chemical message probably abscisic acidis apparently sent from the roots in the drying soilto the shoots leading to stomatal closure as a meansfor conserving water Moreover the stomatalresponse to this water-stress hormone prevents theloss of stem water which may be crucial forepiphytic and hemiepiphytic cacti whose rootsexplore only a relatively small soil volume Whenthe roots of H undatus are excised so that no abscisicacid can move from the roots to the shoot duringdrought the decline in net CO
2 uptake is much
slower (50 inhibition is delayed by approximately8 days compared with plants with their roots intact)emphasising the importance of this hormone forconserving stem water during drought (Nobel amp Dela Barrera 2002a)
When H undatus that has been droughted for 10days is rewatered a significant increase in net CO
2uptake occurs in 1 day the CO
2 uptake ability is
half restored in only 2 days and full recovery occursin 7 days (P S Nobel and E De la Barreraunpublished observations) Thus this cactus canrespond rapidly to rainfall events which may becrucial for its growth in tropical regions with frequentrainfalls (Freiberg 1997 Luumlttge 1997) as well asof importance in developing irrigation scheduleswhen it is cultivated as a crop (Mizrahi amp Nerd1999 Nerd et al 2002) Similarly the hydraulicconductivity of the roots (a direct measure of plantwater uptake ability) of two epiphytic cactiEpiphyllum phyllanthus and Rhipsalis bacciferadecreases during drought but increases to the valuesunder wet conditions only 3 days after rewatering(North amp Nobel 1994)
Temperature
Temperature influences nearly every plant processincluding net CO
2 uptake and hence growth The
optimal daynight air temperatures for total daily netCO
2 uptake by Hylocereus undatus are 3020degC (Fig
4) which are relatively high compared withtemperatures optimal for other CAM plants Because
most net CO2 uptake occurs at night the nighttime
temperatures are more crucial for total daily net CO2
uptake than are the daytime ones The optimalnighttime temperatures for total daily net CO
2 uptake
by cacti native to arid or semiarid regions are from10degC to 15degC (Nobel 1988) Apparently the higheroptimal nighttime temperatures for H undatusreflect temperatures characteristic of the primarilytropical regions where it is native (Fig 1) The totaldaily net CO
2 uptake is half of the maximal value at
low daynight temperatures of 188degC and at highdaynight temperatures of 3828degC (Fig 4) Abovedaynight air temperatures of 4232degC a net loss ofCO
2 occurs over a 24-h period (Fig 4) indicating
that this species is not adapted to such hightemperatures
Temperature also affects the survival of cacti andH undatus is more sensitive to temperature extremesthan are most other cacti Based on an assay usingthe uptake of the vital dye neutral red into the centralvacuole of a cell (Onweume 1979 Nobel et al1995) half of the photosynthetic (chlorenchyma)cells are killed by a 1-h low-temperature treatmentat -15degC for H undatus maintained at daynighttemperatures of 2010degC and half are killed at -13degCfor plants maintained at 3020degC (Nobel et al 2002)For 19 other species of cacti half of thephotosynthetic cells are killed at an averagetemperature of -102degC for plants maintained at 2010degC and the low-temperature acclimation orhardening (decrease in lethal temperatures as the daynight temperatures are reduced) averages 07degC asthe daynight temperatures are reduced by 10degC(Nobel 1988) much greater than the 02degC observedfor H undatus Again using the uptake of neutralred into the cell central vacuoles as the assay halfof the photosynthetic cells of H undatus are killedby a 1-h high-temperature treatment at 540degC forplants at 2515degC and at 554degC for plants at 3525degC (Nobel amp De la Barrera 2002b) For 18 otherspecies of cacti half of the photosynthetic cells arekilled at an average temperature of 570degC for plantsmaintained at 3525degC and the high temperatureacclimation (as the daynight temperatures are raisedby 10degC) averages 53degC (Nobel 1988) comparedwith only 14degC for H undatus Thus both the low-temperature and the high-temperature acclimationswhich are the crucial factors for the tolerance ofextreme temperatures (Nobel 1988 Nobel amp De laBarrera 2002b) are less than one-third as great forH undatus as is the average for other cacti
Time of exposure to extreme temperatures is alsocrucial when examining tolerances especially forhigh temperatures For instance H undatus is notexpected to tolerate daynight air temperatures above4232degC on a prolonged basis as the total daily netCO
2 uptake would then be negative (Fig 4) In this
regard the maximal temperatures in the native
5Hemiepiphytic cactus CO2 uptake
habitats of H undatus generally do not exceed 35degC(Croat 1978 Freiberg 1997 Luumlttge 1997 Benzing1998) Maintaining H undatus at 4030degC graduallyleads to visible damage of the stems which firstbecomes evident at 6 wk (Nobel amp De la Barrera2002b) Necrosis increases to about half of the stemsurface area at 15 wk and no new stem growth isobserved at such temperatures Also daytimetemperatures of 45degC in the field greatly reduceflower production by H undatus and brief exposureto 50degC leads to stem necrosis (Mizrahi amp Nerd1999) Thus this species cannot be cultivated wheredaytime maximal temperatures average 40degC forseveral weeks or occasionally exceed 45degC Indeedtotal daily net CO
2 uptake by H undatus is 37
lower after 30 wk at 3525degC compared with at 10wk (Nobel amp De la Barrera 2002b) suggesting thatH undatus does not have phenotypic plasticity totolerate temperatures exceeding those in its nativehabitat for prolonged periods
Photosynthetic Photon Flux
Hylocereus undatus has considerable net CO2
uptake at a total daily PPF of only 2 mol m-2 day-1
(Fig 5) indicating that it is well adapted to the low-light conditions of a tropical forest (total daily netCO
2 uptake is negative for desert CAM species at
this PPF Nobel 1988) [Net CO2 uptake responses
to PPF for CAM plants are not expressed versus theinstantaneous level of such light as is conventionalfor C
3 and C
4 plants but rather versus the total daily
PPF as most CO2 uptake occurs at night in Phase I
(Fig 3) when no light is available] Total daily netCO
2 uptake by H undatus increases as the total daily
PPF increases to about 20 mol m-2 day-1 above whichnet CO
2 uptake decreases with increasing PPF (Fig
5) indicating that high PPF levels damage the
photosynthetic apparatus Total daily net CO2 uptake
for H undatus is about 90 of maximal at only 10mol m-2 day -1 (Fig 5) For CAM plants such as barrelcacti (eg Ferocactus acanthodes) andplatyopuntias (eg Opuntia ficus-indica) a totaldaily PPF of 10 mol m-2 day-1 leads to about 40 ofthe maximal total daily net CO
2 uptake maximal
uptake for such cacti generally occurs from 30ndash40mol m-2 day-1 (Nobel 1988) levels attained bysurfaces facing east or west on clear days in thesummer
Photoinhibition leading to a reduction in total dailynet CO
2 uptake sets in at a total daily PPF of 20 mol
m-2 day-1 for H undatus (Fig 5) compared withabove 40 mol m-2 day-1 for desert CAM species(Nobel 1988) Measurement of chlorophyllfluorescence for H undatus indicates thatphotosynthetic efficiency (total daily net CO
2 uptake
divided by PPF) actually begins to decrease at arelatively low total daily PPF of 5 mol m-2 day-1
(Raveh et al 1995) To avoid exceeding a total dailyPPF of 20 mol m-2 day-1 a vertical climbing orclambering stem of H undatus would need to beshaded by nearly 40 on a clear day for stemsurfaces facing east or west (north- or south-facingsurfaces would need no shading) In this regard Hundatus H polyrhizus and Selenicereusmegalanthus are commercially grown in Israel undershade cloth that attenuates solar irradiation by 30ndash60 (Raveh et al 1993 1998 Mizrahi amp Nerd1999) Such shading is comparable to that in theforests where such species are native (Benzing 1990Barthlott amp Hunt 1993 Luumlttge 1997) Whenexposed to full sun on clear days H undatus in
Fig 4 Total daily net CO2 uptake by H undatus at the
indicated daynight air temperatures Plants weregrowing under wet conditions and the PPF was 10 molm-2 day-1 Data are from Raveh et al (1995) and Nobelamp De la Barrera (2002b)
Tota
l dai
ly n
et C
O2 u
ptak
e(m
mol
m-2 d
ay-1)
-50
0
100
150
200
50
250
155 2515 453535253020 4030Daynight air temperatures
Fig 5 Total daily net CO2 uptake by H undatus at the
indicated total daily PPF incident on the stem surfacePlants were growing under wet conditions and the daynight temperatures were 2515degC Data are from Ravehet al (1995) and P S Nobel amp E De la Barrera(unpublished measurements for 1-yr-old stems withdata indicated by means plusmn SE for n = 5 plants andnormalised to the previous measurements at a totaldaily PPF of 20 mol m-2 day-1)
Tota
l dai
ly n
et C
O2 u
ptak
e(m
mol
m-2 d
ay-1)
0
100
150
200
50
250
0 5 251510 20Total daily PPF (mol m-2 day-1)
30
6 PARK S NOBEL amp ERICK DE LA BARRERA
Mexico (Ortiacutez 1999) and 23 genotypes ofHylocereus in Israel (Nerd et al 1990) developvisible necrosis on the stems However H undatusis cultivated in southeast Asia such as in the MekongDelta of Vietnam without shading (Nerd et al 2002P S Nobel personal observations) Evidently theprevalent cloud cover in such regions reduces thetotal daily PPF to values that do not lead to stembleaching and necrosis
Nutrients
The effects of soil nutrients on net CO2 uptake of
cacti bearing pitahayas have received little attention(Castillo Martiacutenez et al 1996 Ortiacutez 1999) althoughnutrients are known to have substantial effects onother cacti (Nobel 1988 1989) For instanceapplication of nutrients such as nitrogen can alterthe timing of fruiting for Opuntia ficus-indica(Mizrahi amp Nerd 1999) Application of boron andzinc can increase the number of fruits for H undatusand foliar application of magnesium can increasefruit size (Castillo Martiacutenez et al 1996) Variationsin the applied nitrogen concentration for H undatusrooted in a sandvermiculite mixture for 22 wkaffects its net CO
2 uptake ability as measured by the
maximal nocturnal net CO2 uptake rate 016 mM N
leads to a maximal nocturnal net CO2 uptake rate of
25 micromol m-2 s-1 16 mM N leads to 56 micromol m-2 s-1and 16 mM N leads to 98 micromol m-2 s-1 (Fig 6) thelatter being the N concentration in full-strengthHoagland solution (Hoagland amp Arnon 1950Johnson et al 1957 Epstein 1972) Thus net CO
2uptake for H undatus is highest under a relativelyhigh applied N concentration (8-16 mM Fig 6) asis the case for intensively managed greenhouse crops
and field-grown vegetables and ornamentals (Huett1996 Langhans amp Tibbitts 1997) A maximalnocturnal net CO
2 uptake rate of 10 micromol m-2 s-1 (Fig
6) places H undatus fourth highest among the 28species of cacti whose maximal net CO
2 uptake rates
have been reported (Nobel amp Bobich 2002) Thusthis species has a high potential growth rate underhigh-nutrient conditions
Shifts in stem properties of H undatus as theapplied nitrogen concentration is changed arerelatively slow For instance the halftime for theshift in net CO
2 uptake ability in response to applied
N is 12ndash13 wk (Nobel amp De la Barrera 2002c) After22 wk at 016 mM applied N the stem N level on adry mass basis decreases to only 09 (Nobel amp Dela Barrera 2002c) After 22 wk at an applied Nconcentration of 16 mM stem N is 25 comparableto values in the shoots of cultivated C
3 and C
4 plants
under high-nutrient conditions (Mengel amp Kirkby1987 Marschner 1995) Also the chlorophyll levelof H undatus at an applied N concentration of 016mM for 23 wk is only 029 g m-2 compared with 063g m-2 at 16 mM N (Nobel amp De la Barrera 2002c)H undatus supplied with 16 mM N for 8 wk has astem chlorophyll level of 049 g m-2 (Raveh et al1998) and H polyrhizus has 045 g chlorophyll m-2
under comparable conditions (Raveh et al 1998)Following 23 wk at low N levels the stem net CO
2uptake ability and chlorophyll level both increaseto maximal values when 16 mM N is applied to Hundatus for 11 wk with a halftime for the increaseof 8 wk (Nobel amp De la Barrera 2002c) Thusresponses of H undatus to changes in the applied Nconcentration are slower than for most C
3 and C
4crops and other cacti for which major responses canoccur in 2 wk (Mengel amp Kirkby 1987 Nerd et al1993 Marschner 1995 Nerd amp Nobel 1995)suggesting lack of genetic plasticity in this recentlydomesticated cactus or relatively slow nutrientuptake by the roots Because the maximal net CO
2uptake rates for H undatus are similar at applied Nconcentrations of 8 and 16 mM (Fig 6) the latterlevel may lead to luxury (excess) N consumptionas the tissue level after 22 wk is 36 higher at 16mM than at 8 mM applied N (Nobel amp De la Barrera2002c)
Elevated Atmospheric CO2
Another environmental variable influencing netCO
2 uptake by cacti is the atmospheric CO
2concentration which currently is increasing bynearly 3 parts per million (09) per year (Schimelet al 1996 Stitt amp Krapp 1999) For all 20 CAMspecies studied biomass productivity increases fora doubled atmospheric CO
2 concentration compared
to the current atmospheric CO2 concentration
(Drennan amp Nobel 2000) The increase averages
Fig 6 Maximal net CO2 uptake rate at night for H
undatus maintained for 22 wk at the indicated nitrogenconcentration Plants were growing under wetconditions the daynight air temperatures were 3020degC and the total daily PPF was 10 mol m-2 day-1The maximal rate occurred between 2200 h and 0200h Data are from Nobel amp De la Barrera (2002c)
Max
imal
net
CO
2 up
take
rat
e(m
mol
m-2 d
ay-1)
4
6
8
2
10
0 2 1464 8Nitrogen concentration (mM)
160
7Hemiepiphytic cactus CO2 uptake
35 at 700 ppm (700 micro litre litre -1 or 70 kPa)compared with 350 ppm which represents anapproximately 1 increase in growth per 10 ppmincrease in atmospheric CO
2 concentration for CAM
plants (Nobel 1991) Data for H undatus areconsistent with the results for other CAM plants asa doubled atmospheric CO
2 concentration increases
the total daily net CO2 uptake by 34 (Raveh et al
1995) Most of the increase occurs during thedaytime in Phases II and IV Moreover increases intotal daily net CO
2 uptake under a doubled
atmospheric CO2 concentration compared with the
current atmospheric CO2 concentration are more
prominent under less favorable or even stressfulclimatic conditions For instance the increase in totaldaily net CO
2 uptake for H undatus due to a doubled
atmospheric CO2 concentration is 63 higher at 35
25degC versus 2515degC 90 higher at 24 days ofdrought versus 14 days and 120 higher at a PPFof 30 mol m-2 day-1 versus 20 mol m-2 day-1 (Ravehet al 1995) Thus the responses to atmospheric CO
2increases will not only lead to greater total daily netCO
2 uptake but also will lead to proportionally
greater net CO2 uptake under adverse environmental
conditions
Conclusions and Future Prospects
A striking feature of H undatus is how rapidlynocturnal stomatal opening and net CO
2 uptake
abili ty decrease during drought Such rapidresponses which undoubtedly are related to the oftensmall substrate volume that is occupied by its rootsmay be mediated by abscisic acid Other rootproperties such as the apparently low nutrient uptakerate and the influence of mycorrhizae also needinvestigation Another striking ecophysiologicalfeature of H undatus compared with barrel andcolumnar cacti which is especially pertinent to itscultivation outside its native regions is its poortolerance of high temperatures such as those above40degC Indeed its photosynthetic cells do not showmuch acclimation as the daynight air temperaturesare gradually raised over a period of weeks contraryto the case for other cacti Comparisons at amolecular level between H undatus and other cactishowing substantial high-temperature acclimationcould help elucidate the mechanisms involved forthe latter group The intolerance of H undatus tohigh PPF presumably relates to its shaded naturalhabitat Levels of stem nitrogen and chlorophyllcorrelate well with the net CO
2 uptake ability of H
undatus providing relatively simple assays for thephysiological status of its stems which changerelatively slowly in response to changes in theconcentration of applied nitrogen Indeedunderstanding how climatic factors in general andhormonal factors in particular affect the gas
exchange of H undatus and related species will beinteresting ecologically and extremely importanthorticulturally for the expanding cultivation ofpitahayas
Acknowledgements
We thank the UCLA-Ben Gurion UniversityProgram of Cooperation through the gift of Dr SolLeshin and the US-Israel Binational AgriculturalResearch and Development Fund grant no IS-3382-01 for financial support
References
Anderson E F 2001 The Cactus Family Portland OregonUSA Timber Press 776 pp
Andrade J L Nobel P S 1997 Microhabitats and waterrelations of epiphytic cacti and ferns in a lowland neotropicalforest Biotropica 29261-270
Backeberg C 1966 Das Kakteenlexikon Jena GermanyGustav Fisher 828 pp
Barbeau G 1990 La pitahaya rouge un nouveau fruit exotiqueFruits 45141-147
Barthlott W Hunt D R 1993 Cactaceae In The Families andGenera of Vascular Plants Vol 2 pp 161-196 Ed KKubitzki Berlin Germany Springer-Verlag
Benzing D H 1990 Vascular Epiphytes General Biology andRelated Biota Cambridge UK Cambridge University Press354 pp
Benzing D H 1998 Vulnerabilities of tropical forests to climatechange The significance of resident epiphytes ClimateChange 39519-540
Bravo H H 1978 Las Cactaacuteceas de Meacutexico Vol I MexicoCity Mexico UNAM 743 pp
Britton N L Rose J N 1963 The Cactaceae Descriptionsand Illustrations of Plants of the Cactus Family Vol II NewYork USA Dover 241 pp
Bullock S H 1986 Climate of Chamela Jalisco and trends inthe south coastal region of Mexico Archives for MeteorologyGeophysics and Bioclimatology Series B 36297-316
Castillo Martiacutenez R Caacutelix De Dios H Rodriacuteguez Canto A1996 Guia Teacutecnica para el Cultivo de Pitahaya ChetumalQuintana Roo Mexico Universidad de Quintana Roo 158pp
Croat T B 1978 Flora of Barro Colorado Island StanfordCalifornia USA Stanford University Press 943 pp
Drennan P M Nobel P S 2000 Responses of CAM species toincreasing atmospheric CO
2 concentrations Plant Cell and
Environment 23767-781Epstein E 1972 Mineral Nutrition of Plants Principles and
Perspectives New York USA Wiley 412 ppFreiberg M 1997 Spatial and temporal pattern of temperature
and humidity of a tropical premontane rain forest tree in CostaRica Selbyana 1877-84
Hoagland D R Arnon D I 1950 The water-culture methodfor growing plants without soil California AgriculturalExperimental Station Circular 3471-32
Huett D O 1996 Prospects for manipulating the vegetative-reproductive balance in horticultural crops through nitrogennutrition A review Australian Journal of AgriculturalResearch 4747-66
Johnson C M Stout P R Broyer T C Carlton A B 1957Comparative chlorine requirements of different plant speciesPlant Soil 8337-353
Langhans R W Tibbitts T W 1997 Plant growth chamberhandbook Ames Iowa USA Iowa State University 240 pp
8 PARK S NOBEL amp ERICK DE LA BARRERA
Luumlttge U 1997 Physiological Ecology of Tropical PlantsBerlin Germany Springer-Verlag 384 pp
Maas J M Vose J M Swank W T Martiacutenez-Yriacutezar A 1995Seasonal changes of leaf area index (LAI) in a tropicaldeciduous forest in west Mexico Forest Ecology andManagement 74171-180
Marschner H 1995 Mineral Nutrition of Higher Plants 2ndEdn London UK Academic Press 889 pp
Mengel K Kirkby A E 1987 Principles of Plant Nutrition4th Edn Bern Switzerland International Potash Institute593 pp
Mizrahi Y Nerd A 1999 Climbing and columnar cacti newarid land fruit crops In Perspectives on New Crops and NewUses pp 358-366 Ed J Janick Alexandria Virginia USAASHS Press
Nerd A Nobel P S 1995 Accumulation partitioning andassimilation of nitrate in Opuntia ficus-indica Journal ofPlant Nutrition 182533-2549
Nerd A Aronson J A Mizrahi Y 1990 Introduction anddomestication of rare and wild fruit and nut trees for desertareas In Advances in New Crops pp 353-363 Eds J Janickand J E Simon Portland Oregon USA Timber Press
Nerd A Mesika R Mizrahi Y 1993 Effect of N fertilizer onautumn floral flush and cladode N in prickly pear (Opuntiaficus-indica (L) Mill) Journal of Horticultural Science68337-342
Nerd A Tel-Zur N Mizrahi Y 2002 Fruits of vine andcolumnar cacti In Cacti Biology and Uses pp 185-197Ed P S Nobel Berkeley California University of CaliforniaPress
Nobel P S 1988 Environmental Biology of Agaves and CactiNew York USA Cambridge University Press 270 pp
Nobel P S 1989 A nutrient index quantifying productivity ofagaves and cacti Applied Ecology 26635-645
Nobel P S 1991 Tansley Review No 32 Achievableproductivities of certain CAM plants basis for high valuescompared with C
3 and C
4 plants New Phytologist 119183-
205Nobel P S 1999 Physicochemical and Environmental Plant
Physiology 2nd Edn San Diego California USA AcademicPress 474 pp
Nobel P S Bobich E G 2002 Environmental biology In CactiBiology and Uses pp 57-74 Ed P S Nobel BerkeleyCalifornia USA University of California Press
Nobel P S De la Barrera E 2002a Stem water relations andnet CO
2 uptake for a hemiepiphytic cactus during short-term
drought Environmental and Experimental Botany 48129-137
Nobel P S De la Barrera E 2002b High temperatures and netCO2 uptake growth and stem damage for the hemiepiphyticcactus Hylocereus undatus Biotropica 34225-231
Nobel P S De la Barrera E 2002c Nitrogen relations for netCO 2 uptake by the cultivated hemiepiphytic cactusHylocereus undatus Scientia Horticulturae 96281-292
Nobel P S De la Barrera E Beilman D W Doherty J HZutta B R 2002 Temperature limitations for cultivation ofedible cacti in California Madrontildeo 49228-236
Nobel P S Wang N Balsamo R A Loik M E Hawke M A1995 Low-temperature tolerance and acclimation of Opuntiaspp after injecting glucose or methylglucose InternationalJournal of Plant Sciences 156496-504
Norman J M Campbell G S 1989 Canopy structure In PlantPhysiological Ecology Field Methods and Instrumentationpp 301-325 Eds R W Pearcy J Ehleringer H A Mooney andP W Rundel New York USA Chapman and Hall
North G B Nobel P S 1994 Changes in root hydraulicconductivity for two tropical epiphytic cacti as soil moisturevaries American Journal of Botany 8146-53
Onweume I C 1979 Rapid plant-conserving estimation of heattolerance in plants Journal of Agricultural ScienceCambridge 92527-536
Ortiacutez H Y D Livera M M Carrillo S J A 1996 Asimilacioacutende CO
2 en tallos joacutevenes de pitahaya (Hylocereus undatus)
Revista Fitotecnia Mexicana 1931-41Ortiacutez H Y D 1999 Pitahaya un Nuevo Cultivo para Meacutexico
Pitahaya a New Crop for Mexico Balderas Mexico CityMexico Editorial Limusa Grupo Noriega Editores 111 pp
Osmond C B 1978 Crassulacean acid metabolism a curiosityin context Annual Review of Plant Physiology 26379-414
Pimienta E Hernandez G Domingues A Nobel P S 1998Growth and development of the arborescent cactusStenocereus queretaroensis in a subtropical semiaridenvironment including effects of gibberellic acid TreePhysiology 1859-64
Raveh E Weiss J Nerd A Mizrahi Y 1993 Pitayas (GenusHylocereus) a new fruit crop for the Negev Desert of IsraelIn New Crops pp 491-495 Eds J Janick and J E SimonNew York USA Wiley
Raveh E Gersani M Nobel P S 1995 CO2 uptake and
fluorescence responses for a shade-tolerant cactus Hylocereusundatus under current and doubled CO
2 concentrations
Physiologia Plantarum 93505-511Raveh E Nerd A Mizrahi Y 1998 Responses of two
hemiepiphytic fruit-crop cacti to different degrees of shadeScientia Horticulturae 73151-164
Rzedowski J 1983 Vegetacioacuten de Meacutexico Mexico CityMexico Editorial Limusa 432 pp
Schimel D Ives D Enting I Heimann M Joos F RaynaudD Wigley T 1996 CO2 and the carbon cycle In ClimateChange 1995 IPCC pp 65-131 Eds J T Houghton L GMeira Pilho B A Callendar N Harris A Kattenberg and KMaskell Cambridge UK Cambridge University Press
Stitt M Krapp A 1999 The interaction between elevatedcarbon dioxide and nitrogen nutrition the physiological andmolecular background Plant Cell and Environment 22583-621
Vaacutezquez J A Cuevas R Cochrane T S Iltis H H Santana FJ Guzmaacuten L 1995 Flora de Manantlaacuten Plantas Vascularesde la Reserva de la Bioacutesfera Sierra de Manantlaacuten Jalisco-Colima Meacutexico SIDA Botanical Miscellany No 13 FortWorth Texas USA Botanical Research Institute of Texas312 pp
Winter K Smith J A C 1996 An introduction to Crassulaceanacid metabolism Biochemical principles and ecologicaldiversity In Crassulacean Acid Metabolism pp 1-3 Eds KWinter and J A C Smith Berlin Germany Springer
Zotz G Hietz P 2002 The physiological ecology of vascularepiphytes current knowledge open questions Journal ofExperimental Botany 522067-2078
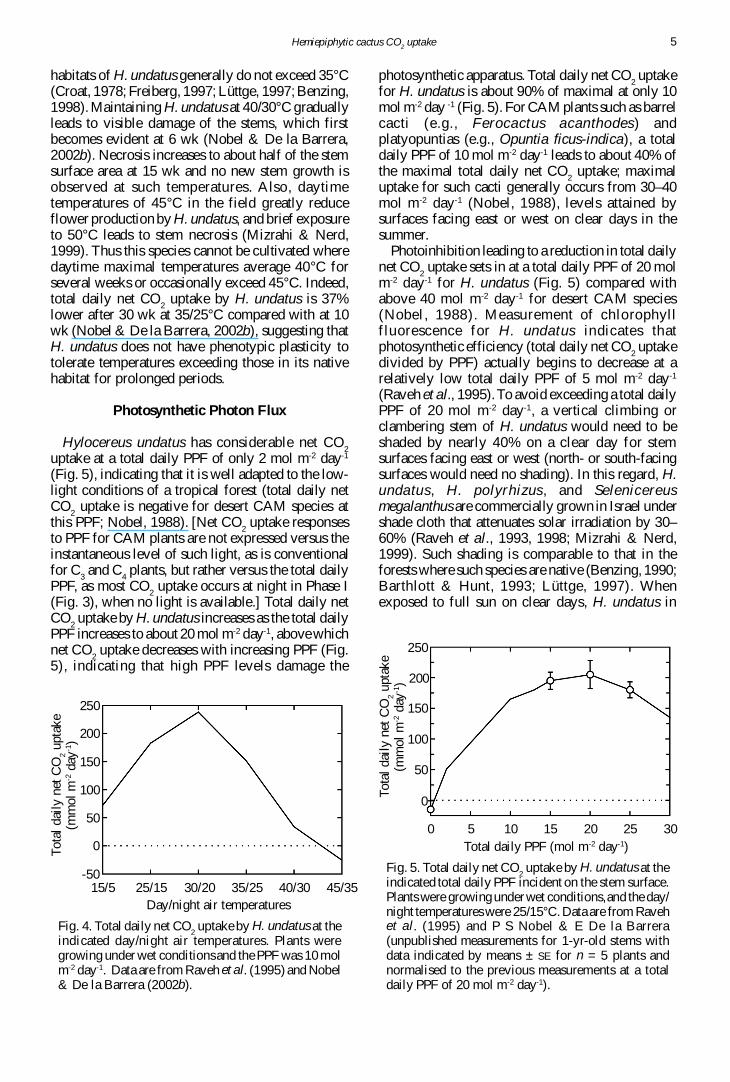
5Hemiepiphytic cactus CO2 uptake
habitats of H undatus generally do not exceed 35degC(Croat 1978 Freiberg 1997 Luumlttge 1997 Benzing1998) Maintaining H undatus at 4030degC graduallyleads to visible damage of the stems which firstbecomes evident at 6 wk (Nobel amp De la Barrera2002b) Necrosis increases to about half of the stemsurface area at 15 wk and no new stem growth isobserved at such temperatures Also daytimetemperatures of 45degC in the field greatly reduceflower production by H undatus and brief exposureto 50degC leads to stem necrosis (Mizrahi amp Nerd1999) Thus this species cannot be cultivated wheredaytime maximal temperatures average 40degC forseveral weeks or occasionally exceed 45degC Indeedtotal daily net CO
2 uptake by H undatus is 37
lower after 30 wk at 3525degC compared with at 10wk (Nobel amp De la Barrera 2002b) suggesting thatH undatus does not have phenotypic plasticity totolerate temperatures exceeding those in its nativehabitat for prolonged periods
Photosynthetic Photon Flux
Hylocereus undatus has considerable net CO2
uptake at a total daily PPF of only 2 mol m-2 day-1
(Fig 5) indicating that it is well adapted to the low-light conditions of a tropical forest (total daily netCO
2 uptake is negative for desert CAM species at
this PPF Nobel 1988) [Net CO2 uptake responses
to PPF for CAM plants are not expressed versus theinstantaneous level of such light as is conventionalfor C
3 and C
4 plants but rather versus the total daily
PPF as most CO2 uptake occurs at night in Phase I
(Fig 3) when no light is available] Total daily netCO
2 uptake by H undatus increases as the total daily
PPF increases to about 20 mol m-2 day-1 above whichnet CO
2 uptake decreases with increasing PPF (Fig
5) indicating that high PPF levels damage the
photosynthetic apparatus Total daily net CO2 uptake
for H undatus is about 90 of maximal at only 10mol m-2 day -1 (Fig 5) For CAM plants such as barrelcacti (eg Ferocactus acanthodes) andplatyopuntias (eg Opuntia ficus-indica) a totaldaily PPF of 10 mol m-2 day-1 leads to about 40 ofthe maximal total daily net CO
2 uptake maximal
uptake for such cacti generally occurs from 30ndash40mol m-2 day-1 (Nobel 1988) levels attained bysurfaces facing east or west on clear days in thesummer
Photoinhibition leading to a reduction in total dailynet CO
2 uptake sets in at a total daily PPF of 20 mol
m-2 day-1 for H undatus (Fig 5) compared withabove 40 mol m-2 day-1 for desert CAM species(Nobel 1988) Measurement of chlorophyllfluorescence for H undatus indicates thatphotosynthetic efficiency (total daily net CO
2 uptake
divided by PPF) actually begins to decrease at arelatively low total daily PPF of 5 mol m-2 day-1
(Raveh et al 1995) To avoid exceeding a total dailyPPF of 20 mol m-2 day-1 a vertical climbing orclambering stem of H undatus would need to beshaded by nearly 40 on a clear day for stemsurfaces facing east or west (north- or south-facingsurfaces would need no shading) In this regard Hundatus H polyrhizus and Selenicereusmegalanthus are commercially grown in Israel undershade cloth that attenuates solar irradiation by 30ndash60 (Raveh et al 1993 1998 Mizrahi amp Nerd1999) Such shading is comparable to that in theforests where such species are native (Benzing 1990Barthlott amp Hunt 1993 Luumlttge 1997) Whenexposed to full sun on clear days H undatus in
Fig 4 Total daily net CO2 uptake by H undatus at the
indicated daynight air temperatures Plants weregrowing under wet conditions and the PPF was 10 molm-2 day-1 Data are from Raveh et al (1995) and Nobelamp De la Barrera (2002b)
Tota
l dai
ly n
et C
O2 u
ptak
e(m
mol
m-2 d
ay-1)
-50
0
100
150
200
50
250
155 2515 453535253020 4030Daynight air temperatures
Fig 5 Total daily net CO2 uptake by H undatus at the
indicated total daily PPF incident on the stem surfacePlants were growing under wet conditions and the daynight temperatures were 2515degC Data are from Ravehet al (1995) and P S Nobel amp E De la Barrera(unpublished measurements for 1-yr-old stems withdata indicated by means plusmn SE for n = 5 plants andnormalised to the previous measurements at a totaldaily PPF of 20 mol m-2 day-1)
Tota
l dai
ly n
et C
O2 u
ptak
e(m
mol
m-2 d
ay-1)
0
100
150
200
50
250
0 5 251510 20Total daily PPF (mol m-2 day-1)
30
6 PARK S NOBEL amp ERICK DE LA BARRERA
Mexico (Ortiacutez 1999) and 23 genotypes ofHylocereus in Israel (Nerd et al 1990) developvisible necrosis on the stems However H undatusis cultivated in southeast Asia such as in the MekongDelta of Vietnam without shading (Nerd et al 2002P S Nobel personal observations) Evidently theprevalent cloud cover in such regions reduces thetotal daily PPF to values that do not lead to stembleaching and necrosis
Nutrients
The effects of soil nutrients on net CO2 uptake of
cacti bearing pitahayas have received little attention(Castillo Martiacutenez et al 1996 Ortiacutez 1999) althoughnutrients are known to have substantial effects onother cacti (Nobel 1988 1989) For instanceapplication of nutrients such as nitrogen can alterthe timing of fruiting for Opuntia ficus-indica(Mizrahi amp Nerd 1999) Application of boron andzinc can increase the number of fruits for H undatusand foliar application of magnesium can increasefruit size (Castillo Martiacutenez et al 1996) Variationsin the applied nitrogen concentration for H undatusrooted in a sandvermiculite mixture for 22 wkaffects its net CO
2 uptake ability as measured by the
maximal nocturnal net CO2 uptake rate 016 mM N
leads to a maximal nocturnal net CO2 uptake rate of
25 micromol m-2 s-1 16 mM N leads to 56 micromol m-2 s-1and 16 mM N leads to 98 micromol m-2 s-1 (Fig 6) thelatter being the N concentration in full-strengthHoagland solution (Hoagland amp Arnon 1950Johnson et al 1957 Epstein 1972) Thus net CO
2uptake for H undatus is highest under a relativelyhigh applied N concentration (8-16 mM Fig 6) asis the case for intensively managed greenhouse crops
and field-grown vegetables and ornamentals (Huett1996 Langhans amp Tibbitts 1997) A maximalnocturnal net CO
2 uptake rate of 10 micromol m-2 s-1 (Fig
6) places H undatus fourth highest among the 28species of cacti whose maximal net CO
2 uptake rates
have been reported (Nobel amp Bobich 2002) Thusthis species has a high potential growth rate underhigh-nutrient conditions
Shifts in stem properties of H undatus as theapplied nitrogen concentration is changed arerelatively slow For instance the halftime for theshift in net CO
2 uptake ability in response to applied
N is 12ndash13 wk (Nobel amp De la Barrera 2002c) After22 wk at 016 mM applied N the stem N level on adry mass basis decreases to only 09 (Nobel amp Dela Barrera 2002c) After 22 wk at an applied Nconcentration of 16 mM stem N is 25 comparableto values in the shoots of cultivated C
3 and C
4 plants
under high-nutrient conditions (Mengel amp Kirkby1987 Marschner 1995) Also the chlorophyll levelof H undatus at an applied N concentration of 016mM for 23 wk is only 029 g m-2 compared with 063g m-2 at 16 mM N (Nobel amp De la Barrera 2002c)H undatus supplied with 16 mM N for 8 wk has astem chlorophyll level of 049 g m-2 (Raveh et al1998) and H polyrhizus has 045 g chlorophyll m-2
under comparable conditions (Raveh et al 1998)Following 23 wk at low N levels the stem net CO
2uptake ability and chlorophyll level both increaseto maximal values when 16 mM N is applied to Hundatus for 11 wk with a halftime for the increaseof 8 wk (Nobel amp De la Barrera 2002c) Thusresponses of H undatus to changes in the applied Nconcentration are slower than for most C
3 and C
4crops and other cacti for which major responses canoccur in 2 wk (Mengel amp Kirkby 1987 Nerd et al1993 Marschner 1995 Nerd amp Nobel 1995)suggesting lack of genetic plasticity in this recentlydomesticated cactus or relatively slow nutrientuptake by the roots Because the maximal net CO
2uptake rates for H undatus are similar at applied Nconcentrations of 8 and 16 mM (Fig 6) the latterlevel may lead to luxury (excess) N consumptionas the tissue level after 22 wk is 36 higher at 16mM than at 8 mM applied N (Nobel amp De la Barrera2002c)
Elevated Atmospheric CO2
Another environmental variable influencing netCO
2 uptake by cacti is the atmospheric CO
2concentration which currently is increasing bynearly 3 parts per million (09) per year (Schimelet al 1996 Stitt amp Krapp 1999) For all 20 CAMspecies studied biomass productivity increases fora doubled atmospheric CO
2 concentration compared
to the current atmospheric CO2 concentration
(Drennan amp Nobel 2000) The increase averages
Fig 6 Maximal net CO2 uptake rate at night for H
undatus maintained for 22 wk at the indicated nitrogenconcentration Plants were growing under wetconditions the daynight air temperatures were 3020degC and the total daily PPF was 10 mol m-2 day-1The maximal rate occurred between 2200 h and 0200h Data are from Nobel amp De la Barrera (2002c)
Max
imal
net
CO
2 up
take
rat
e(m
mol
m-2 d
ay-1)
4
6
8
2
10
0 2 1464 8Nitrogen concentration (mM)
160
7Hemiepiphytic cactus CO2 uptake
35 at 700 ppm (700 micro litre litre -1 or 70 kPa)compared with 350 ppm which represents anapproximately 1 increase in growth per 10 ppmincrease in atmospheric CO
2 concentration for CAM
plants (Nobel 1991) Data for H undatus areconsistent with the results for other CAM plants asa doubled atmospheric CO
2 concentration increases
the total daily net CO2 uptake by 34 (Raveh et al
1995) Most of the increase occurs during thedaytime in Phases II and IV Moreover increases intotal daily net CO
2 uptake under a doubled
atmospheric CO2 concentration compared with the
current atmospheric CO2 concentration are more
prominent under less favorable or even stressfulclimatic conditions For instance the increase in totaldaily net CO
2 uptake for H undatus due to a doubled
atmospheric CO2 concentration is 63 higher at 35
25degC versus 2515degC 90 higher at 24 days ofdrought versus 14 days and 120 higher at a PPFof 30 mol m-2 day-1 versus 20 mol m-2 day-1 (Ravehet al 1995) Thus the responses to atmospheric CO
2increases will not only lead to greater total daily netCO
2 uptake but also will lead to proportionally
greater net CO2 uptake under adverse environmental
conditions
Conclusions and Future Prospects
A striking feature of H undatus is how rapidlynocturnal stomatal opening and net CO
2 uptake
abili ty decrease during drought Such rapidresponses which undoubtedly are related to the oftensmall substrate volume that is occupied by its rootsmay be mediated by abscisic acid Other rootproperties such as the apparently low nutrient uptakerate and the influence of mycorrhizae also needinvestigation Another striking ecophysiologicalfeature of H undatus compared with barrel andcolumnar cacti which is especially pertinent to itscultivation outside its native regions is its poortolerance of high temperatures such as those above40degC Indeed its photosynthetic cells do not showmuch acclimation as the daynight air temperaturesare gradually raised over a period of weeks contraryto the case for other cacti Comparisons at amolecular level between H undatus and other cactishowing substantial high-temperature acclimationcould help elucidate the mechanisms involved forthe latter group The intolerance of H undatus tohigh PPF presumably relates to its shaded naturalhabitat Levels of stem nitrogen and chlorophyllcorrelate well with the net CO
2 uptake ability of H
undatus providing relatively simple assays for thephysiological status of its stems which changerelatively slowly in response to changes in theconcentration of applied nitrogen Indeedunderstanding how climatic factors in general andhormonal factors in particular affect the gas
exchange of H undatus and related species will beinteresting ecologically and extremely importanthorticulturally for the expanding cultivation ofpitahayas
Acknowledgements
We thank the UCLA-Ben Gurion UniversityProgram of Cooperation through the gift of Dr SolLeshin and the US-Israel Binational AgriculturalResearch and Development Fund grant no IS-3382-01 for financial support
References
Anderson E F 2001 The Cactus Family Portland OregonUSA Timber Press 776 pp
Andrade J L Nobel P S 1997 Microhabitats and waterrelations of epiphytic cacti and ferns in a lowland neotropicalforest Biotropica 29261-270
Backeberg C 1966 Das Kakteenlexikon Jena GermanyGustav Fisher 828 pp
Barbeau G 1990 La pitahaya rouge un nouveau fruit exotiqueFruits 45141-147
Barthlott W Hunt D R 1993 Cactaceae In The Families andGenera of Vascular Plants Vol 2 pp 161-196 Ed KKubitzki Berlin Germany Springer-Verlag
Benzing D H 1990 Vascular Epiphytes General Biology andRelated Biota Cambridge UK Cambridge University Press354 pp
Benzing D H 1998 Vulnerabilities of tropical forests to climatechange The significance of resident epiphytes ClimateChange 39519-540
Bravo H H 1978 Las Cactaacuteceas de Meacutexico Vol I MexicoCity Mexico UNAM 743 pp
Britton N L Rose J N 1963 The Cactaceae Descriptionsand Illustrations of Plants of the Cactus Family Vol II NewYork USA Dover 241 pp
Bullock S H 1986 Climate of Chamela Jalisco and trends inthe south coastal region of Mexico Archives for MeteorologyGeophysics and Bioclimatology Series B 36297-316
Castillo Martiacutenez R Caacutelix De Dios H Rodriacuteguez Canto A1996 Guia Teacutecnica para el Cultivo de Pitahaya ChetumalQuintana Roo Mexico Universidad de Quintana Roo 158pp
Croat T B 1978 Flora of Barro Colorado Island StanfordCalifornia USA Stanford University Press 943 pp
Drennan P M Nobel P S 2000 Responses of CAM species toincreasing atmospheric CO
2 concentrations Plant Cell and
Environment 23767-781Epstein E 1972 Mineral Nutrition of Plants Principles and
Perspectives New York USA Wiley 412 ppFreiberg M 1997 Spatial and temporal pattern of temperature
and humidity of a tropical premontane rain forest tree in CostaRica Selbyana 1877-84
Hoagland D R Arnon D I 1950 The water-culture methodfor growing plants without soil California AgriculturalExperimental Station Circular 3471-32
Huett D O 1996 Prospects for manipulating the vegetative-reproductive balance in horticultural crops through nitrogennutrition A review Australian Journal of AgriculturalResearch 4747-66
Johnson C M Stout P R Broyer T C Carlton A B 1957Comparative chlorine requirements of different plant speciesPlant Soil 8337-353
Langhans R W Tibbitts T W 1997 Plant growth chamberhandbook Ames Iowa USA Iowa State University 240 pp
8 PARK S NOBEL amp ERICK DE LA BARRERA
Luumlttge U 1997 Physiological Ecology of Tropical PlantsBerlin Germany Springer-Verlag 384 pp
Maas J M Vose J M Swank W T Martiacutenez-Yriacutezar A 1995Seasonal changes of leaf area index (LAI) in a tropicaldeciduous forest in west Mexico Forest Ecology andManagement 74171-180
Marschner H 1995 Mineral Nutrition of Higher Plants 2ndEdn London UK Academic Press 889 pp
Mengel K Kirkby A E 1987 Principles of Plant Nutrition4th Edn Bern Switzerland International Potash Institute593 pp
Mizrahi Y Nerd A 1999 Climbing and columnar cacti newarid land fruit crops In Perspectives on New Crops and NewUses pp 358-366 Ed J Janick Alexandria Virginia USAASHS Press
Nerd A Nobel P S 1995 Accumulation partitioning andassimilation of nitrate in Opuntia ficus-indica Journal ofPlant Nutrition 182533-2549
Nerd A Aronson J A Mizrahi Y 1990 Introduction anddomestication of rare and wild fruit and nut trees for desertareas In Advances in New Crops pp 353-363 Eds J Janickand J E Simon Portland Oregon USA Timber Press
Nerd A Mesika R Mizrahi Y 1993 Effect of N fertilizer onautumn floral flush and cladode N in prickly pear (Opuntiaficus-indica (L) Mill) Journal of Horticultural Science68337-342
Nerd A Tel-Zur N Mizrahi Y 2002 Fruits of vine andcolumnar cacti In Cacti Biology and Uses pp 185-197Ed P S Nobel Berkeley California University of CaliforniaPress
Nobel P S 1988 Environmental Biology of Agaves and CactiNew York USA Cambridge University Press 270 pp
Nobel P S 1989 A nutrient index quantifying productivity ofagaves and cacti Applied Ecology 26635-645
Nobel P S 1991 Tansley Review No 32 Achievableproductivities of certain CAM plants basis for high valuescompared with C
3 and C
4 plants New Phytologist 119183-
205Nobel P S 1999 Physicochemical and Environmental Plant
Physiology 2nd Edn San Diego California USA AcademicPress 474 pp
Nobel P S Bobich E G 2002 Environmental biology In CactiBiology and Uses pp 57-74 Ed P S Nobel BerkeleyCalifornia USA University of California Press
Nobel P S De la Barrera E 2002a Stem water relations andnet CO
2 uptake for a hemiepiphytic cactus during short-term
drought Environmental and Experimental Botany 48129-137
Nobel P S De la Barrera E 2002b High temperatures and netCO2 uptake growth and stem damage for the hemiepiphyticcactus Hylocereus undatus Biotropica 34225-231
Nobel P S De la Barrera E 2002c Nitrogen relations for netCO 2 uptake by the cultivated hemiepiphytic cactusHylocereus undatus Scientia Horticulturae 96281-292
Nobel P S De la Barrera E Beilman D W Doherty J HZutta B R 2002 Temperature limitations for cultivation ofedible cacti in California Madrontildeo 49228-236
Nobel P S Wang N Balsamo R A Loik M E Hawke M A1995 Low-temperature tolerance and acclimation of Opuntiaspp after injecting glucose or methylglucose InternationalJournal of Plant Sciences 156496-504
Norman J M Campbell G S 1989 Canopy structure In PlantPhysiological Ecology Field Methods and Instrumentationpp 301-325 Eds R W Pearcy J Ehleringer H A Mooney andP W Rundel New York USA Chapman and Hall
North G B Nobel P S 1994 Changes in root hydraulicconductivity for two tropical epiphytic cacti as soil moisturevaries American Journal of Botany 8146-53
Onweume I C 1979 Rapid plant-conserving estimation of heattolerance in plants Journal of Agricultural ScienceCambridge 92527-536
Ortiacutez H Y D Livera M M Carrillo S J A 1996 Asimilacioacutende CO
2 en tallos joacutevenes de pitahaya (Hylocereus undatus)
Revista Fitotecnia Mexicana 1931-41Ortiacutez H Y D 1999 Pitahaya un Nuevo Cultivo para Meacutexico
Pitahaya a New Crop for Mexico Balderas Mexico CityMexico Editorial Limusa Grupo Noriega Editores 111 pp
Osmond C B 1978 Crassulacean acid metabolism a curiosityin context Annual Review of Plant Physiology 26379-414
Pimienta E Hernandez G Domingues A Nobel P S 1998Growth and development of the arborescent cactusStenocereus queretaroensis in a subtropical semiaridenvironment including effects of gibberellic acid TreePhysiology 1859-64
Raveh E Weiss J Nerd A Mizrahi Y 1993 Pitayas (GenusHylocereus) a new fruit crop for the Negev Desert of IsraelIn New Crops pp 491-495 Eds J Janick and J E SimonNew York USA Wiley
Raveh E Gersani M Nobel P S 1995 CO2 uptake and
fluorescence responses for a shade-tolerant cactus Hylocereusundatus under current and doubled CO
2 concentrations
Physiologia Plantarum 93505-511Raveh E Nerd A Mizrahi Y 1998 Responses of two
hemiepiphytic fruit-crop cacti to different degrees of shadeScientia Horticulturae 73151-164
Rzedowski J 1983 Vegetacioacuten de Meacutexico Mexico CityMexico Editorial Limusa 432 pp
Schimel D Ives D Enting I Heimann M Joos F RaynaudD Wigley T 1996 CO2 and the carbon cycle In ClimateChange 1995 IPCC pp 65-131 Eds J T Houghton L GMeira Pilho B A Callendar N Harris A Kattenberg and KMaskell Cambridge UK Cambridge University Press
Stitt M Krapp A 1999 The interaction between elevatedcarbon dioxide and nitrogen nutrition the physiological andmolecular background Plant Cell and Environment 22583-621
Vaacutezquez J A Cuevas R Cochrane T S Iltis H H Santana FJ Guzmaacuten L 1995 Flora de Manantlaacuten Plantas Vascularesde la Reserva de la Bioacutesfera Sierra de Manantlaacuten Jalisco-Colima Meacutexico SIDA Botanical Miscellany No 13 FortWorth Texas USA Botanical Research Institute of Texas312 pp
Winter K Smith J A C 1996 An introduction to Crassulaceanacid metabolism Biochemical principles and ecologicaldiversity In Crassulacean Acid Metabolism pp 1-3 Eds KWinter and J A C Smith Berlin Germany Springer
Zotz G Hietz P 2002 The physiological ecology of vascularepiphytes current knowledge open questions Journal ofExperimental Botany 522067-2078
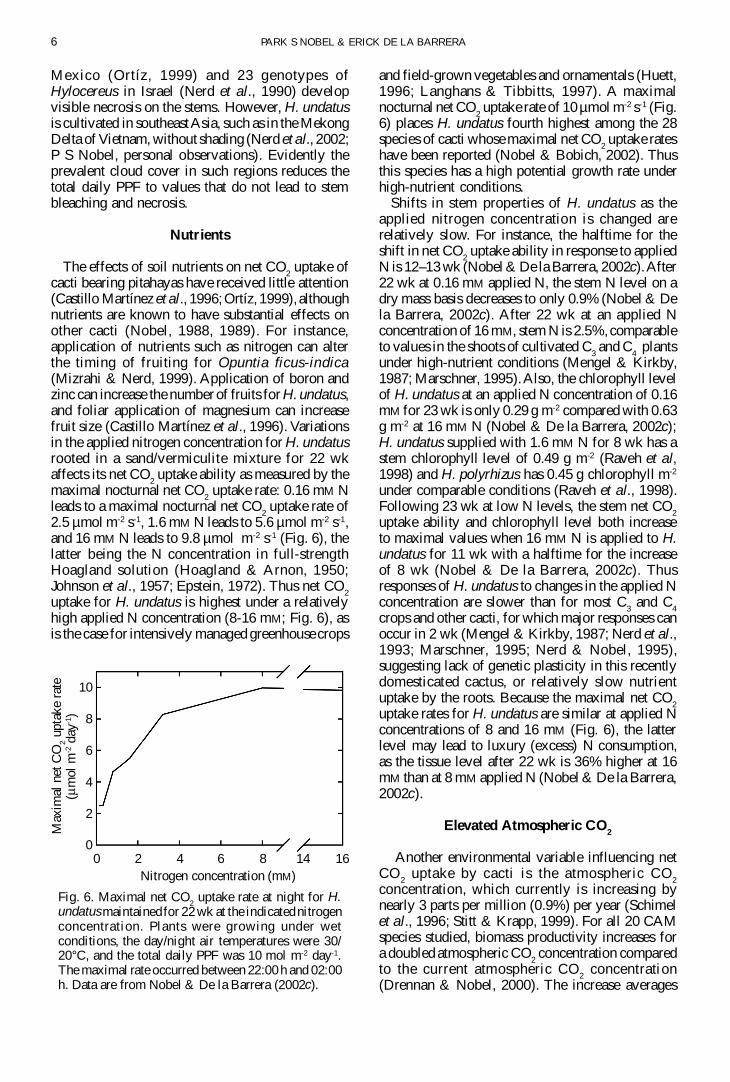
6 PARK S NOBEL amp ERICK DE LA BARRERA
Mexico (Ortiacutez 1999) and 23 genotypes ofHylocereus in Israel (Nerd et al 1990) developvisible necrosis on the stems However H undatusis cultivated in southeast Asia such as in the MekongDelta of Vietnam without shading (Nerd et al 2002P S Nobel personal observations) Evidently theprevalent cloud cover in such regions reduces thetotal daily PPF to values that do not lead to stembleaching and necrosis
Nutrients
The effects of soil nutrients on net CO2 uptake of
cacti bearing pitahayas have received little attention(Castillo Martiacutenez et al 1996 Ortiacutez 1999) althoughnutrients are known to have substantial effects onother cacti (Nobel 1988 1989) For instanceapplication of nutrients such as nitrogen can alterthe timing of fruiting for Opuntia ficus-indica(Mizrahi amp Nerd 1999) Application of boron andzinc can increase the number of fruits for H undatusand foliar application of magnesium can increasefruit size (Castillo Martiacutenez et al 1996) Variationsin the applied nitrogen concentration for H undatusrooted in a sandvermiculite mixture for 22 wkaffects its net CO
2 uptake ability as measured by the
maximal nocturnal net CO2 uptake rate 016 mM N
leads to a maximal nocturnal net CO2 uptake rate of
25 micromol m-2 s-1 16 mM N leads to 56 micromol m-2 s-1and 16 mM N leads to 98 micromol m-2 s-1 (Fig 6) thelatter being the N concentration in full-strengthHoagland solution (Hoagland amp Arnon 1950Johnson et al 1957 Epstein 1972) Thus net CO
2uptake for H undatus is highest under a relativelyhigh applied N concentration (8-16 mM Fig 6) asis the case for intensively managed greenhouse crops
and field-grown vegetables and ornamentals (Huett1996 Langhans amp Tibbitts 1997) A maximalnocturnal net CO
2 uptake rate of 10 micromol m-2 s-1 (Fig
6) places H undatus fourth highest among the 28species of cacti whose maximal net CO
2 uptake rates
have been reported (Nobel amp Bobich 2002) Thusthis species has a high potential growth rate underhigh-nutrient conditions
Shifts in stem properties of H undatus as theapplied nitrogen concentration is changed arerelatively slow For instance the halftime for theshift in net CO
2 uptake ability in response to applied
N is 12ndash13 wk (Nobel amp De la Barrera 2002c) After22 wk at 016 mM applied N the stem N level on adry mass basis decreases to only 09 (Nobel amp Dela Barrera 2002c) After 22 wk at an applied Nconcentration of 16 mM stem N is 25 comparableto values in the shoots of cultivated C
3 and C
4 plants
under high-nutrient conditions (Mengel amp Kirkby1987 Marschner 1995) Also the chlorophyll levelof H undatus at an applied N concentration of 016mM for 23 wk is only 029 g m-2 compared with 063g m-2 at 16 mM N (Nobel amp De la Barrera 2002c)H undatus supplied with 16 mM N for 8 wk has astem chlorophyll level of 049 g m-2 (Raveh et al1998) and H polyrhizus has 045 g chlorophyll m-2
under comparable conditions (Raveh et al 1998)Following 23 wk at low N levels the stem net CO
2uptake ability and chlorophyll level both increaseto maximal values when 16 mM N is applied to Hundatus for 11 wk with a halftime for the increaseof 8 wk (Nobel amp De la Barrera 2002c) Thusresponses of H undatus to changes in the applied Nconcentration are slower than for most C
3 and C
4crops and other cacti for which major responses canoccur in 2 wk (Mengel amp Kirkby 1987 Nerd et al1993 Marschner 1995 Nerd amp Nobel 1995)suggesting lack of genetic plasticity in this recentlydomesticated cactus or relatively slow nutrientuptake by the roots Because the maximal net CO
2uptake rates for H undatus are similar at applied Nconcentrations of 8 and 16 mM (Fig 6) the latterlevel may lead to luxury (excess) N consumptionas the tissue level after 22 wk is 36 higher at 16mM than at 8 mM applied N (Nobel amp De la Barrera2002c)
Elevated Atmospheric CO2
Another environmental variable influencing netCO
2 uptake by cacti is the atmospheric CO
2concentration which currently is increasing bynearly 3 parts per million (09) per year (Schimelet al 1996 Stitt amp Krapp 1999) For all 20 CAMspecies studied biomass productivity increases fora doubled atmospheric CO
2 concentration compared
to the current atmospheric CO2 concentration
(Drennan amp Nobel 2000) The increase averages
Fig 6 Maximal net CO2 uptake rate at night for H
undatus maintained for 22 wk at the indicated nitrogenconcentration Plants were growing under wetconditions the daynight air temperatures were 3020degC and the total daily PPF was 10 mol m-2 day-1The maximal rate occurred between 2200 h and 0200h Data are from Nobel amp De la Barrera (2002c)
Max
imal
net
CO
2 up
take
rat
e(m
mol
m-2 d
ay-1)
4
6
8
2
10
0 2 1464 8Nitrogen concentration (mM)
160
7Hemiepiphytic cactus CO2 uptake
35 at 700 ppm (700 micro litre litre -1 or 70 kPa)compared with 350 ppm which represents anapproximately 1 increase in growth per 10 ppmincrease in atmospheric CO
2 concentration for CAM
plants (Nobel 1991) Data for H undatus areconsistent with the results for other CAM plants asa doubled atmospheric CO
2 concentration increases
the total daily net CO2 uptake by 34 (Raveh et al
1995) Most of the increase occurs during thedaytime in Phases II and IV Moreover increases intotal daily net CO
2 uptake under a doubled
atmospheric CO2 concentration compared with the
current atmospheric CO2 concentration are more
prominent under less favorable or even stressfulclimatic conditions For instance the increase in totaldaily net CO
2 uptake for H undatus due to a doubled
atmospheric CO2 concentration is 63 higher at 35
25degC versus 2515degC 90 higher at 24 days ofdrought versus 14 days and 120 higher at a PPFof 30 mol m-2 day-1 versus 20 mol m-2 day-1 (Ravehet al 1995) Thus the responses to atmospheric CO
2increases will not only lead to greater total daily netCO
2 uptake but also will lead to proportionally
greater net CO2 uptake under adverse environmental
conditions
Conclusions and Future Prospects
A striking feature of H undatus is how rapidlynocturnal stomatal opening and net CO
2 uptake
abili ty decrease during drought Such rapidresponses which undoubtedly are related to the oftensmall substrate volume that is occupied by its rootsmay be mediated by abscisic acid Other rootproperties such as the apparently low nutrient uptakerate and the influence of mycorrhizae also needinvestigation Another striking ecophysiologicalfeature of H undatus compared with barrel andcolumnar cacti which is especially pertinent to itscultivation outside its native regions is its poortolerance of high temperatures such as those above40degC Indeed its photosynthetic cells do not showmuch acclimation as the daynight air temperaturesare gradually raised over a period of weeks contraryto the case for other cacti Comparisons at amolecular level between H undatus and other cactishowing substantial high-temperature acclimationcould help elucidate the mechanisms involved forthe latter group The intolerance of H undatus tohigh PPF presumably relates to its shaded naturalhabitat Levels of stem nitrogen and chlorophyllcorrelate well with the net CO
2 uptake ability of H
undatus providing relatively simple assays for thephysiological status of its stems which changerelatively slowly in response to changes in theconcentration of applied nitrogen Indeedunderstanding how climatic factors in general andhormonal factors in particular affect the gas
exchange of H undatus and related species will beinteresting ecologically and extremely importanthorticulturally for the expanding cultivation ofpitahayas
Acknowledgements
We thank the UCLA-Ben Gurion UniversityProgram of Cooperation through the gift of Dr SolLeshin and the US-Israel Binational AgriculturalResearch and Development Fund grant no IS-3382-01 for financial support
References
Anderson E F 2001 The Cactus Family Portland OregonUSA Timber Press 776 pp
Andrade J L Nobel P S 1997 Microhabitats and waterrelations of epiphytic cacti and ferns in a lowland neotropicalforest Biotropica 29261-270
Backeberg C 1966 Das Kakteenlexikon Jena GermanyGustav Fisher 828 pp
Barbeau G 1990 La pitahaya rouge un nouveau fruit exotiqueFruits 45141-147
Barthlott W Hunt D R 1993 Cactaceae In The Families andGenera of Vascular Plants Vol 2 pp 161-196 Ed KKubitzki Berlin Germany Springer-Verlag
Benzing D H 1990 Vascular Epiphytes General Biology andRelated Biota Cambridge UK Cambridge University Press354 pp
Benzing D H 1998 Vulnerabilities of tropical forests to climatechange The significance of resident epiphytes ClimateChange 39519-540
Bravo H H 1978 Las Cactaacuteceas de Meacutexico Vol I MexicoCity Mexico UNAM 743 pp
Britton N L Rose J N 1963 The Cactaceae Descriptionsand Illustrations of Plants of the Cactus Family Vol II NewYork USA Dover 241 pp
Bullock S H 1986 Climate of Chamela Jalisco and trends inthe south coastal region of Mexico Archives for MeteorologyGeophysics and Bioclimatology Series B 36297-316
Castillo Martiacutenez R Caacutelix De Dios H Rodriacuteguez Canto A1996 Guia Teacutecnica para el Cultivo de Pitahaya ChetumalQuintana Roo Mexico Universidad de Quintana Roo 158pp
Croat T B 1978 Flora of Barro Colorado Island StanfordCalifornia USA Stanford University Press 943 pp
Drennan P M Nobel P S 2000 Responses of CAM species toincreasing atmospheric CO
2 concentrations Plant Cell and
Environment 23767-781Epstein E 1972 Mineral Nutrition of Plants Principles and
Perspectives New York USA Wiley 412 ppFreiberg M 1997 Spatial and temporal pattern of temperature
and humidity of a tropical premontane rain forest tree in CostaRica Selbyana 1877-84
Hoagland D R Arnon D I 1950 The water-culture methodfor growing plants without soil California AgriculturalExperimental Station Circular 3471-32
Huett D O 1996 Prospects for manipulating the vegetative-reproductive balance in horticultural crops through nitrogennutrition A review Australian Journal of AgriculturalResearch 4747-66
Johnson C M Stout P R Broyer T C Carlton A B 1957Comparative chlorine requirements of different plant speciesPlant Soil 8337-353
Langhans R W Tibbitts T W 1997 Plant growth chamberhandbook Ames Iowa USA Iowa State University 240 pp
8 PARK S NOBEL amp ERICK DE LA BARRERA
Luumlttge U 1997 Physiological Ecology of Tropical PlantsBerlin Germany Springer-Verlag 384 pp
Maas J M Vose J M Swank W T Martiacutenez-Yriacutezar A 1995Seasonal changes of leaf area index (LAI) in a tropicaldeciduous forest in west Mexico Forest Ecology andManagement 74171-180
Marschner H 1995 Mineral Nutrition of Higher Plants 2ndEdn London UK Academic Press 889 pp
Mengel K Kirkby A E 1987 Principles of Plant Nutrition4th Edn Bern Switzerland International Potash Institute593 pp
Mizrahi Y Nerd A 1999 Climbing and columnar cacti newarid land fruit crops In Perspectives on New Crops and NewUses pp 358-366 Ed J Janick Alexandria Virginia USAASHS Press
Nerd A Nobel P S 1995 Accumulation partitioning andassimilation of nitrate in Opuntia ficus-indica Journal ofPlant Nutrition 182533-2549
Nerd A Aronson J A Mizrahi Y 1990 Introduction anddomestication of rare and wild fruit and nut trees for desertareas In Advances in New Crops pp 353-363 Eds J Janickand J E Simon Portland Oregon USA Timber Press
Nerd A Mesika R Mizrahi Y 1993 Effect of N fertilizer onautumn floral flush and cladode N in prickly pear (Opuntiaficus-indica (L) Mill) Journal of Horticultural Science68337-342
Nerd A Tel-Zur N Mizrahi Y 2002 Fruits of vine andcolumnar cacti In Cacti Biology and Uses pp 185-197Ed P S Nobel Berkeley California University of CaliforniaPress
Nobel P S 1988 Environmental Biology of Agaves and CactiNew York USA Cambridge University Press 270 pp
Nobel P S 1989 A nutrient index quantifying productivity ofagaves and cacti Applied Ecology 26635-645
Nobel P S 1991 Tansley Review No 32 Achievableproductivities of certain CAM plants basis for high valuescompared with C
3 and C
4 plants New Phytologist 119183-
205Nobel P S 1999 Physicochemical and Environmental Plant
Physiology 2nd Edn San Diego California USA AcademicPress 474 pp
Nobel P S Bobich E G 2002 Environmental biology In CactiBiology and Uses pp 57-74 Ed P S Nobel BerkeleyCalifornia USA University of California Press
Nobel P S De la Barrera E 2002a Stem water relations andnet CO
2 uptake for a hemiepiphytic cactus during short-term
drought Environmental and Experimental Botany 48129-137
Nobel P S De la Barrera E 2002b High temperatures and netCO2 uptake growth and stem damage for the hemiepiphyticcactus Hylocereus undatus Biotropica 34225-231
Nobel P S De la Barrera E 2002c Nitrogen relations for netCO 2 uptake by the cultivated hemiepiphytic cactusHylocereus undatus Scientia Horticulturae 96281-292
Nobel P S De la Barrera E Beilman D W Doherty J HZutta B R 2002 Temperature limitations for cultivation ofedible cacti in California Madrontildeo 49228-236
Nobel P S Wang N Balsamo R A Loik M E Hawke M A1995 Low-temperature tolerance and acclimation of Opuntiaspp after injecting glucose or methylglucose InternationalJournal of Plant Sciences 156496-504
Norman J M Campbell G S 1989 Canopy structure In PlantPhysiological Ecology Field Methods and Instrumentationpp 301-325 Eds R W Pearcy J Ehleringer H A Mooney andP W Rundel New York USA Chapman and Hall
North G B Nobel P S 1994 Changes in root hydraulicconductivity for two tropical epiphytic cacti as soil moisturevaries American Journal of Botany 8146-53
Onweume I C 1979 Rapid plant-conserving estimation of heattolerance in plants Journal of Agricultural ScienceCambridge 92527-536
Ortiacutez H Y D Livera M M Carrillo S J A 1996 Asimilacioacutende CO
2 en tallos joacutevenes de pitahaya (Hylocereus undatus)
Revista Fitotecnia Mexicana 1931-41Ortiacutez H Y D 1999 Pitahaya un Nuevo Cultivo para Meacutexico
Pitahaya a New Crop for Mexico Balderas Mexico CityMexico Editorial Limusa Grupo Noriega Editores 111 pp
Osmond C B 1978 Crassulacean acid metabolism a curiosityin context Annual Review of Plant Physiology 26379-414
Pimienta E Hernandez G Domingues A Nobel P S 1998Growth and development of the arborescent cactusStenocereus queretaroensis in a subtropical semiaridenvironment including effects of gibberellic acid TreePhysiology 1859-64
Raveh E Weiss J Nerd A Mizrahi Y 1993 Pitayas (GenusHylocereus) a new fruit crop for the Negev Desert of IsraelIn New Crops pp 491-495 Eds J Janick and J E SimonNew York USA Wiley
Raveh E Gersani M Nobel P S 1995 CO2 uptake and
fluorescence responses for a shade-tolerant cactus Hylocereusundatus under current and doubled CO
2 concentrations
Physiologia Plantarum 93505-511Raveh E Nerd A Mizrahi Y 1998 Responses of two
hemiepiphytic fruit-crop cacti to different degrees of shadeScientia Horticulturae 73151-164
Rzedowski J 1983 Vegetacioacuten de Meacutexico Mexico CityMexico Editorial Limusa 432 pp
Schimel D Ives D Enting I Heimann M Joos F RaynaudD Wigley T 1996 CO2 and the carbon cycle In ClimateChange 1995 IPCC pp 65-131 Eds J T Houghton L GMeira Pilho B A Callendar N Harris A Kattenberg and KMaskell Cambridge UK Cambridge University Press
Stitt M Krapp A 1999 The interaction between elevatedcarbon dioxide and nitrogen nutrition the physiological andmolecular background Plant Cell and Environment 22583-621
Vaacutezquez J A Cuevas R Cochrane T S Iltis H H Santana FJ Guzmaacuten L 1995 Flora de Manantlaacuten Plantas Vascularesde la Reserva de la Bioacutesfera Sierra de Manantlaacuten Jalisco-Colima Meacutexico SIDA Botanical Miscellany No 13 FortWorth Texas USA Botanical Research Institute of Texas312 pp
Winter K Smith J A C 1996 An introduction to Crassulaceanacid metabolism Biochemical principles and ecologicaldiversity In Crassulacean Acid Metabolism pp 1-3 Eds KWinter and J A C Smith Berlin Germany Springer
Zotz G Hietz P 2002 The physiological ecology of vascularepiphytes current knowledge open questions Journal ofExperimental Botany 522067-2078

7Hemiepiphytic cactus CO2 uptake
35 at 700 ppm (700 micro litre litre -1 or 70 kPa)compared with 350 ppm which represents anapproximately 1 increase in growth per 10 ppmincrease in atmospheric CO
2 concentration for CAM
plants (Nobel 1991) Data for H undatus areconsistent with the results for other CAM plants asa doubled atmospheric CO
2 concentration increases
the total daily net CO2 uptake by 34 (Raveh et al
1995) Most of the increase occurs during thedaytime in Phases II and IV Moreover increases intotal daily net CO
2 uptake under a doubled
atmospheric CO2 concentration compared with the
current atmospheric CO2 concentration are more
prominent under less favorable or even stressfulclimatic conditions For instance the increase in totaldaily net CO
2 uptake for H undatus due to a doubled
atmospheric CO2 concentration is 63 higher at 35
25degC versus 2515degC 90 higher at 24 days ofdrought versus 14 days and 120 higher at a PPFof 30 mol m-2 day-1 versus 20 mol m-2 day-1 (Ravehet al 1995) Thus the responses to atmospheric CO
2increases will not only lead to greater total daily netCO
2 uptake but also will lead to proportionally
greater net CO2 uptake under adverse environmental
conditions
Conclusions and Future Prospects
A striking feature of H undatus is how rapidlynocturnal stomatal opening and net CO
2 uptake
abili ty decrease during drought Such rapidresponses which undoubtedly are related to the oftensmall substrate volume that is occupied by its rootsmay be mediated by abscisic acid Other rootproperties such as the apparently low nutrient uptakerate and the influence of mycorrhizae also needinvestigation Another striking ecophysiologicalfeature of H undatus compared with barrel andcolumnar cacti which is especially pertinent to itscultivation outside its native regions is its poortolerance of high temperatures such as those above40degC Indeed its photosynthetic cells do not showmuch acclimation as the daynight air temperaturesare gradually raised over a period of weeks contraryto the case for other cacti Comparisons at amolecular level between H undatus and other cactishowing substantial high-temperature acclimationcould help elucidate the mechanisms involved forthe latter group The intolerance of H undatus tohigh PPF presumably relates to its shaded naturalhabitat Levels of stem nitrogen and chlorophyllcorrelate well with the net CO
2 uptake ability of H
undatus providing relatively simple assays for thephysiological status of its stems which changerelatively slowly in response to changes in theconcentration of applied nitrogen Indeedunderstanding how climatic factors in general andhormonal factors in particular affect the gas
exchange of H undatus and related species will beinteresting ecologically and extremely importanthorticulturally for the expanding cultivation ofpitahayas
Acknowledgements
We thank the UCLA-Ben Gurion UniversityProgram of Cooperation through the gift of Dr SolLeshin and the US-Israel Binational AgriculturalResearch and Development Fund grant no IS-3382-01 for financial support
References
Anderson E F 2001 The Cactus Family Portland OregonUSA Timber Press 776 pp
Andrade J L Nobel P S 1997 Microhabitats and waterrelations of epiphytic cacti and ferns in a lowland neotropicalforest Biotropica 29261-270
Backeberg C 1966 Das Kakteenlexikon Jena GermanyGustav Fisher 828 pp
Barbeau G 1990 La pitahaya rouge un nouveau fruit exotiqueFruits 45141-147
Barthlott W Hunt D R 1993 Cactaceae In The Families andGenera of Vascular Plants Vol 2 pp 161-196 Ed KKubitzki Berlin Germany Springer-Verlag
Benzing D H 1990 Vascular Epiphytes General Biology andRelated Biota Cambridge UK Cambridge University Press354 pp
Benzing D H 1998 Vulnerabilities of tropical forests to climatechange The significance of resident epiphytes ClimateChange 39519-540
Bravo H H 1978 Las Cactaacuteceas de Meacutexico Vol I MexicoCity Mexico UNAM 743 pp
Britton N L Rose J N 1963 The Cactaceae Descriptionsand Illustrations of Plants of the Cactus Family Vol II NewYork USA Dover 241 pp
Bullock S H 1986 Climate of Chamela Jalisco and trends inthe south coastal region of Mexico Archives for MeteorologyGeophysics and Bioclimatology Series B 36297-316
Castillo Martiacutenez R Caacutelix De Dios H Rodriacuteguez Canto A1996 Guia Teacutecnica para el Cultivo de Pitahaya ChetumalQuintana Roo Mexico Universidad de Quintana Roo 158pp
Croat T B 1978 Flora of Barro Colorado Island StanfordCalifornia USA Stanford University Press 943 pp
Drennan P M Nobel P S 2000 Responses of CAM species toincreasing atmospheric CO
2 concentrations Plant Cell and
Environment 23767-781Epstein E 1972 Mineral Nutrition of Plants Principles and
Perspectives New York USA Wiley 412 ppFreiberg M 1997 Spatial and temporal pattern of temperature
and humidity of a tropical premontane rain forest tree in CostaRica Selbyana 1877-84
Hoagland D R Arnon D I 1950 The water-culture methodfor growing plants without soil California AgriculturalExperimental Station Circular 3471-32
Huett D O 1996 Prospects for manipulating the vegetative-reproductive balance in horticultural crops through nitrogennutrition A review Australian Journal of AgriculturalResearch 4747-66
Johnson C M Stout P R Broyer T C Carlton A B 1957Comparative chlorine requirements of different plant speciesPlant Soil 8337-353
Langhans R W Tibbitts T W 1997 Plant growth chamberhandbook Ames Iowa USA Iowa State University 240 pp
8 PARK S NOBEL amp ERICK DE LA BARRERA
Luumlttge U 1997 Physiological Ecology of Tropical PlantsBerlin Germany Springer-Verlag 384 pp
Maas J M Vose J M Swank W T Martiacutenez-Yriacutezar A 1995Seasonal changes of leaf area index (LAI) in a tropicaldeciduous forest in west Mexico Forest Ecology andManagement 74171-180
Marschner H 1995 Mineral Nutrition of Higher Plants 2ndEdn London UK Academic Press 889 pp
Mengel K Kirkby A E 1987 Principles of Plant Nutrition4th Edn Bern Switzerland International Potash Institute593 pp
Mizrahi Y Nerd A 1999 Climbing and columnar cacti newarid land fruit crops In Perspectives on New Crops and NewUses pp 358-366 Ed J Janick Alexandria Virginia USAASHS Press
Nerd A Nobel P S 1995 Accumulation partitioning andassimilation of nitrate in Opuntia ficus-indica Journal ofPlant Nutrition 182533-2549
Nerd A Aronson J A Mizrahi Y 1990 Introduction anddomestication of rare and wild fruit and nut trees for desertareas In Advances in New Crops pp 353-363 Eds J Janickand J E Simon Portland Oregon USA Timber Press
Nerd A Mesika R Mizrahi Y 1993 Effect of N fertilizer onautumn floral flush and cladode N in prickly pear (Opuntiaficus-indica (L) Mill) Journal of Horticultural Science68337-342
Nerd A Tel-Zur N Mizrahi Y 2002 Fruits of vine andcolumnar cacti In Cacti Biology and Uses pp 185-197Ed P S Nobel Berkeley California University of CaliforniaPress
Nobel P S 1988 Environmental Biology of Agaves and CactiNew York USA Cambridge University Press 270 pp
Nobel P S 1989 A nutrient index quantifying productivity ofagaves and cacti Applied Ecology 26635-645
Nobel P S 1991 Tansley Review No 32 Achievableproductivities of certain CAM plants basis for high valuescompared with C
3 and C
4 plants New Phytologist 119183-
205Nobel P S 1999 Physicochemical and Environmental Plant
Physiology 2nd Edn San Diego California USA AcademicPress 474 pp
Nobel P S Bobich E G 2002 Environmental biology In CactiBiology and Uses pp 57-74 Ed P S Nobel BerkeleyCalifornia USA University of California Press
Nobel P S De la Barrera E 2002a Stem water relations andnet CO
2 uptake for a hemiepiphytic cactus during short-term
drought Environmental and Experimental Botany 48129-137
Nobel P S De la Barrera E 2002b High temperatures and netCO2 uptake growth and stem damage for the hemiepiphyticcactus Hylocereus undatus Biotropica 34225-231
Nobel P S De la Barrera E 2002c Nitrogen relations for netCO 2 uptake by the cultivated hemiepiphytic cactusHylocereus undatus Scientia Horticulturae 96281-292
Nobel P S De la Barrera E Beilman D W Doherty J HZutta B R 2002 Temperature limitations for cultivation ofedible cacti in California Madrontildeo 49228-236
Nobel P S Wang N Balsamo R A Loik M E Hawke M A1995 Low-temperature tolerance and acclimation of Opuntiaspp after injecting glucose or methylglucose InternationalJournal of Plant Sciences 156496-504
Norman J M Campbell G S 1989 Canopy structure In PlantPhysiological Ecology Field Methods and Instrumentationpp 301-325 Eds R W Pearcy J Ehleringer H A Mooney andP W Rundel New York USA Chapman and Hall
North G B Nobel P S 1994 Changes in root hydraulicconductivity for two tropical epiphytic cacti as soil moisturevaries American Journal of Botany 8146-53
Onweume I C 1979 Rapid plant-conserving estimation of heattolerance in plants Journal of Agricultural ScienceCambridge 92527-536
Ortiacutez H Y D Livera M M Carrillo S J A 1996 Asimilacioacutende CO
2 en tallos joacutevenes de pitahaya (Hylocereus undatus)
Revista Fitotecnia Mexicana 1931-41Ortiacutez H Y D 1999 Pitahaya un Nuevo Cultivo para Meacutexico
Pitahaya a New Crop for Mexico Balderas Mexico CityMexico Editorial Limusa Grupo Noriega Editores 111 pp
Osmond C B 1978 Crassulacean acid metabolism a curiosityin context Annual Review of Plant Physiology 26379-414
Pimienta E Hernandez G Domingues A Nobel P S 1998Growth and development of the arborescent cactusStenocereus queretaroensis in a subtropical semiaridenvironment including effects of gibberellic acid TreePhysiology 1859-64
Raveh E Weiss J Nerd A Mizrahi Y 1993 Pitayas (GenusHylocereus) a new fruit crop for the Negev Desert of IsraelIn New Crops pp 491-495 Eds J Janick and J E SimonNew York USA Wiley
Raveh E Gersani M Nobel P S 1995 CO2 uptake and
fluorescence responses for a shade-tolerant cactus Hylocereusundatus under current and doubled CO
2 concentrations
Physiologia Plantarum 93505-511Raveh E Nerd A Mizrahi Y 1998 Responses of two
hemiepiphytic fruit-crop cacti to different degrees of shadeScientia Horticulturae 73151-164
Rzedowski J 1983 Vegetacioacuten de Meacutexico Mexico CityMexico Editorial Limusa 432 pp
Schimel D Ives D Enting I Heimann M Joos F RaynaudD Wigley T 1996 CO2 and the carbon cycle In ClimateChange 1995 IPCC pp 65-131 Eds J T Houghton L GMeira Pilho B A Callendar N Harris A Kattenberg and KMaskell Cambridge UK Cambridge University Press
Stitt M Krapp A 1999 The interaction between elevatedcarbon dioxide and nitrogen nutrition the physiological andmolecular background Plant Cell and Environment 22583-621
Vaacutezquez J A Cuevas R Cochrane T S Iltis H H Santana FJ Guzmaacuten L 1995 Flora de Manantlaacuten Plantas Vascularesde la Reserva de la Bioacutesfera Sierra de Manantlaacuten Jalisco-Colima Meacutexico SIDA Botanical Miscellany No 13 FortWorth Texas USA Botanical Research Institute of Texas312 pp
Winter K Smith J A C 1996 An introduction to Crassulaceanacid metabolism Biochemical principles and ecologicaldiversity In Crassulacean Acid Metabolism pp 1-3 Eds KWinter and J A C Smith Berlin Germany Springer
Zotz G Hietz P 2002 The physiological ecology of vascularepiphytes current knowledge open questions Journal ofExperimental Botany 522067-2078

8 PARK S NOBEL amp ERICK DE LA BARRERA
Luumlttge U 1997 Physiological Ecology of Tropical PlantsBerlin Germany Springer-Verlag 384 pp
Maas J M Vose J M Swank W T Martiacutenez-Yriacutezar A 1995Seasonal changes of leaf area index (LAI) in a tropicaldeciduous forest in west Mexico Forest Ecology andManagement 74171-180
Marschner H 1995 Mineral Nutrition of Higher Plants 2ndEdn London UK Academic Press 889 pp
Mengel K Kirkby A E 1987 Principles of Plant Nutrition4th Edn Bern Switzerland International Potash Institute593 pp
Mizrahi Y Nerd A 1999 Climbing and columnar cacti newarid land fruit crops In Perspectives on New Crops and NewUses pp 358-366 Ed J Janick Alexandria Virginia USAASHS Press
Nerd A Nobel P S 1995 Accumulation partitioning andassimilation of nitrate in Opuntia ficus-indica Journal ofPlant Nutrition 182533-2549
Nerd A Aronson J A Mizrahi Y 1990 Introduction anddomestication of rare and wild fruit and nut trees for desertareas In Advances in New Crops pp 353-363 Eds J Janickand J E Simon Portland Oregon USA Timber Press
Nerd A Mesika R Mizrahi Y 1993 Effect of N fertilizer onautumn floral flush and cladode N in prickly pear (Opuntiaficus-indica (L) Mill) Journal of Horticultural Science68337-342
Nerd A Tel-Zur N Mizrahi Y 2002 Fruits of vine andcolumnar cacti In Cacti Biology and Uses pp 185-197Ed P S Nobel Berkeley California University of CaliforniaPress
Nobel P S 1988 Environmental Biology of Agaves and CactiNew York USA Cambridge University Press 270 pp
Nobel P S 1989 A nutrient index quantifying productivity ofagaves and cacti Applied Ecology 26635-645
Nobel P S 1991 Tansley Review No 32 Achievableproductivities of certain CAM plants basis for high valuescompared with C
3 and C
4 plants New Phytologist 119183-
205Nobel P S 1999 Physicochemical and Environmental Plant
Physiology 2nd Edn San Diego California USA AcademicPress 474 pp
Nobel P S Bobich E G 2002 Environmental biology In CactiBiology and Uses pp 57-74 Ed P S Nobel BerkeleyCalifornia USA University of California Press
Nobel P S De la Barrera E 2002a Stem water relations andnet CO
2 uptake for a hemiepiphytic cactus during short-term
drought Environmental and Experimental Botany 48129-137
Nobel P S De la Barrera E 2002b High temperatures and netCO2 uptake growth and stem damage for the hemiepiphyticcactus Hylocereus undatus Biotropica 34225-231
Nobel P S De la Barrera E 2002c Nitrogen relations for netCO 2 uptake by the cultivated hemiepiphytic cactusHylocereus undatus Scientia Horticulturae 96281-292
Nobel P S De la Barrera E Beilman D W Doherty J HZutta B R 2002 Temperature limitations for cultivation ofedible cacti in California Madrontildeo 49228-236
Nobel P S Wang N Balsamo R A Loik M E Hawke M A1995 Low-temperature tolerance and acclimation of Opuntiaspp after injecting glucose or methylglucose InternationalJournal of Plant Sciences 156496-504
Norman J M Campbell G S 1989 Canopy structure In PlantPhysiological Ecology Field Methods and Instrumentationpp 301-325 Eds R W Pearcy J Ehleringer H A Mooney andP W Rundel New York USA Chapman and Hall
North G B Nobel P S 1994 Changes in root hydraulicconductivity for two tropical epiphytic cacti as soil moisturevaries American Journal of Botany 8146-53
Onweume I C 1979 Rapid plant-conserving estimation of heattolerance in plants Journal of Agricultural ScienceCambridge 92527-536
Ortiacutez H Y D Livera M M Carrillo S J A 1996 Asimilacioacutende CO
2 en tallos joacutevenes de pitahaya (Hylocereus undatus)
Revista Fitotecnia Mexicana 1931-41Ortiacutez H Y D 1999 Pitahaya un Nuevo Cultivo para Meacutexico
Pitahaya a New Crop for Mexico Balderas Mexico CityMexico Editorial Limusa Grupo Noriega Editores 111 pp
Osmond C B 1978 Crassulacean acid metabolism a curiosityin context Annual Review of Plant Physiology 26379-414
Pimienta E Hernandez G Domingues A Nobel P S 1998Growth and development of the arborescent cactusStenocereus queretaroensis in a subtropical semiaridenvironment including effects of gibberellic acid TreePhysiology 1859-64
Raveh E Weiss J Nerd A Mizrahi Y 1993 Pitayas (GenusHylocereus) a new fruit crop for the Negev Desert of IsraelIn New Crops pp 491-495 Eds J Janick and J E SimonNew York USA Wiley
Raveh E Gersani M Nobel P S 1995 CO2 uptake and
fluorescence responses for a shade-tolerant cactus Hylocereusundatus under current and doubled CO
2 concentrations
Physiologia Plantarum 93505-511Raveh E Nerd A Mizrahi Y 1998 Responses of two
hemiepiphytic fruit-crop cacti to different degrees of shadeScientia Horticulturae 73151-164
Rzedowski J 1983 Vegetacioacuten de Meacutexico Mexico CityMexico Editorial Limusa 432 pp
Schimel D Ives D Enting I Heimann M Joos F RaynaudD Wigley T 1996 CO2 and the carbon cycle In ClimateChange 1995 IPCC pp 65-131 Eds J T Houghton L GMeira Pilho B A Callendar N Harris A Kattenberg and KMaskell Cambridge UK Cambridge University Press
Stitt M Krapp A 1999 The interaction between elevatedcarbon dioxide and nitrogen nutrition the physiological andmolecular background Plant Cell and Environment 22583-621
Vaacutezquez J A Cuevas R Cochrane T S Iltis H H Santana FJ Guzmaacuten L 1995 Flora de Manantlaacuten Plantas Vascularesde la Reserva de la Bioacutesfera Sierra de Manantlaacuten Jalisco-Colima Meacutexico SIDA Botanical Miscellany No 13 FortWorth Texas USA Botanical Research Institute of Texas312 pp
Winter K Smith J A C 1996 An introduction to Crassulaceanacid metabolism Biochemical principles and ecologicaldiversity In Crassulacean Acid Metabolism pp 1-3 Eds KWinter and J A C Smith Berlin Germany Springer
Zotz G Hietz P 2002 The physiological ecology of vascularepiphytes current knowledge open questions Journal ofExperimental Botany 522067-2078