podoplanin requires sialylated o-glycans for stable expression on lymphatic endothelial cells and...
TRANSCRIPT

Regular Article
VASCULAR BIOLOGY
Podoplanin requires sialylated O-glycans for stable expression onlymphatic endothelial cells and for interaction with plateletsYanfang Pan,1-3 Tadayuki Yago,1 Jianxin Fu,1,4 Brett Herzog,1,2 J. Michael McDaniel,1 Padmaja Mehta-D’Souza,1
Xiaofeng Cai,1 Changgeng Ruan,4 Rodger P. McEver,1,2 Christopher West,2 Kesheng Dai,4 Hong Chen,1,2 and Lijun Xia1,2,4
1Cardiovascular Biology Research Program, Oklahoma Medical Research Foundation, Oklahoma City, OK; 2Department of Biochemistry and Molecular
Biology, University of Oklahoma Health Sciences Center, Oklahoma City, OK; 3Zhejiang Provincial Center for Disease Control and Prevention, Hangzhou,
Zhejiang, China; 4Jiangsu Institute of Hematology, Collaborative Innovation Center of Hematology, Key Laboratory of Thrombosis and Hemostasis of
Ministry of Health, The First Affiliated Hospital of Soochow University, Suzhou, Jiangsu, China
Key Points
• Sialylated O-glycans protectsPDPN from proteolyticdegradation.
• Sialylated core 1 O-glycansof PDPN on lymphaticendothelial cells (LECs) arerequired for interacting withplatelets.
O-glycosylation of podoplanin (PDPN) on lymphatic endothelial cells is critical for the
separation of blood and lymphatic systems by interacting with platelet C-type lectin-like
receptor 2 during development. However, how O-glycosylation controls endothelial
PDPN function and expression remains unclear. In this study, we report that core 1
O-glycan–deficient or desialylated PDPN was highly susceptible to proteolytic degrada-
tion by various proteases, including metalloproteinases (MMP)-2/9. We found that the
lymphcontainedactivatedMMP-2/9 and incubationof the lymphreducedsurface levelsof
PDPNoncore 1O-glycan–deficient endothelial cells, but not onwild-typeECs. The lymph
frommice with sepsis induced by cecal ligation and puncture, which contained bacteria-
derived sialidase, reduced PDPN levels on wild-type ECs. The MMP inhibitor, GM6001,
rescued these reductions.Additionally,GM6001 treatment rescued the reductionofPDPN
level on lymphatic endothelial cells in mice lacking endothelial core 1 O-glycan or cecal
ligation and puncture-treatedmice. Furthermore, core 1O-glycan–deficient or desialylated PDPN impaired platelet interaction under
physiological flow. These data indicate that sialylatedO-glycans of PDPN are essential for platelet adhesion and prevent PDPN from
proteolytic degradation primarily mediated by MMPs in the lymph. (Blood. 2014;124(24):3656-3665)
Introduction
Podoplanin (PDPN), a type 1 transmembrane mucin-type O-glycoprotein, was initially discovered as a platelet aggregation-inducing glycoprotein expressed on tumor cells.1 Subsequentstudies have shown that it is expressed on several cell types, in-cluding lymphatic endothelial cells (LECs).2-5 PDPN on LECsis critical for the initiation and maintenance of an independentlymphatic vascular system.
PDPN consists of 172 amino acids in mice and 163 amino acidsin humans. It has an extracellular domain, a transmembrane do-main, and a short cytoplasmic domain. A striking feature of theextracellular domain of PDPN is a high content of serine andthreonine residues that can potentially be attached by mucin-typeO-glycans (also known as O-N-acetylgalactosamine [GalNAc] gly-cans or simply O-glycans for this study).5-7 O-glycosylation isa common form of posttranslational modification of membrane andsecreted proteins.8-10 It occurs in the Golgi apparatus via sequentialreactions catalyzed by specific glycosyltransferases. The core of allmucin-type O-glycans is serine/threonine-linked GalNAc, alsoknown as the Tn antigen, which is normally not exposed due tofurther modification in forming distinct subtypes of O-glycans.Among them, core 1 O-glycans are a predominant form. Core 1O-glycans are synthesized by adding galactose in b3 linkage to Tn
antigen, which is catalyzed solely by the T-synthase (core 1synthase, C1galt1).8-11 Core 1 structure can be further branched toform extended core 1 or core 2 structures, or can be modified byadding sialic acids. These glycans are known as core 1-derivedO-glycans.10,11
The molecular weight of the core PDPN polypeptide is about17 kDa; however, PDPN isolated from different cell types hasan apparent molecular weight ranging from 37 kDa to 41 kDa,suggesting extensive, heterogeneous O-glycosylation. In our pre-vious study, mice lacking endothelial core 1 O-glycans display im-paired PDPN expression that is required for the development andmaintenance of an independent lymphatic vascular system.8,12,13
PDPN binding to the platelet C-type lectin-like receptor 2 (CLEC-2)induces platelet aggregation that seals initial blood-lymphaticvascular connections during embryonic development.12 Sialyl core 1O-glycans on PDPN are required for interacting with CLEC-2 usingChinese hamster ovary cells (CHO) or synthetic glycopeptides.6,7,14
However, CHO cells lack core 2 and extended core 1 O-glycans.15
Therefore, whether PDPN on endothelial cells (ECs) requires similarO-glycosylation to interact with CLEC-2 is unclear. Additionally,howO-glycosylation regulates function and expression of PDPN onLECs remains to be addressed.
Submitted April 24, 2014; accepted October 2, 2014. Prepublished online as
Blood First Edition paper, October 21, 2014; DOI 10.1182/blood-2014-04-
572107.
The online version of this article contains a data supplement.
The publication costs of this article were defrayed in part by page charge
payment. Therefore, and solely to indicate this fact, this article is hereby
marked “advertisement” in accordance with 18 USC section 1734.
© 2014 by The American Society of Hematology
3656 BLOOD, 4 DECEMBER 2014 x VOLUME 124, NUMBER 24
For personal use only.on December 8, 2014. by guest www.bloodjournal.orgFrom

In this study, we show that core 1 O-glycan–deficient ordesialylated PDPN on LECs is highly susceptible to proteolyticdegradation, suggesting the importance of O-glycosylation for con-trolling stability on EC surfaces. Notably, matrix metalloproteinases(MMPs) in the lymph are essential for the proteolytic degradation ofcore 1 O-glycan–deficient or desialylated PDPN. Furthermore, core1 O-glycan–deficient or desialylated PDPN on LECs exhibits im-paired interactions with platelets under flow. Our results providestrong evidence that sialylatedO-glycans control the expression andfunction of endothelial PDPN.
Materials and methods
Mice and cells
Mice lacking EC and hematopoietic core (EHC) 1O-glycans (C1galt12/2 orC1galt1f/f; Tie2Cre) and platelet-specific Clec22/2 mice were describedpreviously.8,13 Mice with doxycycline-inducible global deficiency of core 1O-glycans (inducibleC1galt12/2)were generated by crossingC1galt1f/fmicewith Rosa26-rtTA, tetO-Cre Tg mice (The Jackson Laboratory). PrimaryLECs were isolated from different tissues as described in “Results.” Some ofthe LECs were used for culture. Stable PDPN-eGFP–expressed ECs areestablished by transfection of immortalized wild-type (WT) or C1galt12/2
ECs8 with DNA construct of murine PDPN tagged with a C-terminal eGFP(PDPN-eGFP). CHO cells were cotransfected with complementary DNAconstructs encoding PDPN-eGFP and glycosyltransferases. All mouseexperiments were performed in compliance with protocols approved by theInstitutional Animal Care and Use Committee of the Oklahoma MedicalResearch Foundation.
Platelet adhesion under flow
WT or Clec22/2 platelet adhesion on ECs or CHO cells expressing PDPN-eGFP were performed under flow.16
Lymph or protease treatment
PDPN-eGFP transfectedWTorC1galt12/2ECswere incubatedwith 200mLof mouse lymph or buffer containing proteases.
Intraperitoneal (IP) injection of MMP inhibitor
IP injections of GM6001 were performed. Primary LECs isolated frommesentery and cryosections of ileum were used for staining to examine Tn,Lyve-1, and PDPN expression.
Cecal ligation and puncture (CLP) sepsis model
CLP was performed with or without IP injection of sialidase inhibitor orGM6001. The lymph and mesentery were collected for EC treatment andflow cytometry.
More details of materials and methods used are described in thesupplemental Methods, available on the BloodWeb site.
Statistics
The unpaired Student t test was used to determine P values as indicated in thefigures.
Results
Lack of core 1 O-glycosylation reduces PDPN on LECs in vivo
Wereport here thatmice lacking core 1O-glycans (EHCC1galt12/2)exhibited reduced levels of PDPN in lymphatic microvessels.8 To
determine whether the reduction of PDPN occurred on largercollecting lymphatic vessels, we performed immunostaining on lym-phatic vessels in embryonic day (E17)mesentery.Wefirst probed theProx1-positive LECs of the collecting lymphatic vessels of EHCC1galt12/2 mesenteries with mAb to Tn antigen (Figure 1A).8,9
Anti-Tn mAb stained C1galt12/2 but not WT LECs, indicatingefficient excision of the C1galt1 gene. Desialylation did not ap-preciably affect the intensity of anti-Tn staining (data not shown),indicating that most exposed Tn is not sialylated. Immunofluores-cent images revealed that the collecting lymphatic vessels of EHCC1galt12/2 mesenteries had reduced PDPN levels relative to WTcollecting lymphatic vessels. Using primary LECs, which were de-fined as CD311/Lyve-11 cells (supplemental Figure 1A), flowcytometric analyses showed that the surface level of PDPN onmesenteric LECs from EHC C1galt12/2 mice was reduced com-paredwith that fromWTmice (Figure 1B). Reduced surface levels ofPDPN were also found on isolated primary LECs from collectinglymphatic vessels in 3-week-old and 8-week-old EHC C1galt12/2
mesentery (supplemental Figure 1B), on isolated primary LECsfrom P1 (supplemental Figure 1C), or in cryosections of lymphaticmicrovessels of 8-week-old EHC C1galt12/2 small intestine (sup-plementalFigure1D).These results indicate that loss of core1O-glycanscauses defective expression of PDPN in different types of C1galt12/2
lymphatic vessels.Loss of core 1 O-glycans occurs during embryonic development
in EHC C1galt12/2 mice. To determine whether postnatal loss ofcore 1O-glycosylation impairs PDPN expression, we used inducibleC1galt12/2mice. Sixweeks after a doxycycline diet, Tn antigenwasdetected in the intestine of inducibleC1galt12/2mice but not inWTlittermates (supplemental Figure 1E). Lyve-1–positive submucosallymphatic vessels of the WT intestine expressed a high level ofPDPN (Figure 1C, left). In contrast, the level of PDPN was di-minished in those of the inducible C1galt12/2 intestine. Flowcytometric analyses showed reduced PDPN levels on primary LECsfrom the inducible C1galt12/2 mesentery compared with that fromWT littermates (Figure 1C, right). These data indicate that reducedPDPN levels in lymphatic vessels of EHC C1galt12/2 mice occurduring both developmental and postnatal stages.
EHC C1galt12/2 mice exhibit disorganized and blood-filledlymphatic vessels.8 To determine whether these lymphatic vascular ab-normalities contribute to defective PDPN expression, we examinedPDPN levels on LECs isolated frommice lacking CLEC-2 (Clec22/2),which exhibit the same lymphatic vascular abnormalities.12,13 PDPNexpression on LECs fromClec22/2micewas equivalent to that fromWT mice, indicating that lymphatic abnormalities do not cause thereduced PDPN expression (Figure 1D). WT and C1galt12/2 LECsexpressed a similar amount of PDPN messenger RNA, suggest-ing that lack of core 1 O-glycans does not affect the transcription(supplemental Figure 2). Collectively, these data indicate that lackof core 1 O-glycans causes a reduction of PDPN in C1galt12/2
lymphatic vessels.
Lack of core 1 O-glycosylation does not affect intracellular
trafficking or protein turnover of PDPN
O-glycan deficiency may result in defective synthesis, impaired in-tracellular trafficking, or increased degradation of glycoproteins.8,17-24
To determine how the lack of core 1 O-glycan resulted in reducedexpression of PDPN, we isolated primary skin ECs enriched in LECsfrom WT or EHC C1galt12/2 mice,8 and then cultured the isolatedcells for 5 days to increase cell numbers for analysis (referring tocultured primary LECs). Most of these cells were CD31 and Lyve-1
BLOOD, 4 DECEMBER 2014 x VOLUME 124, NUMBER 24 SIALYLATED O-GLYCANS CONTROL PODOPLANIN EXPRESSION 3657
For personal use only.on December 8, 2014. by guest www.bloodjournal.orgFrom

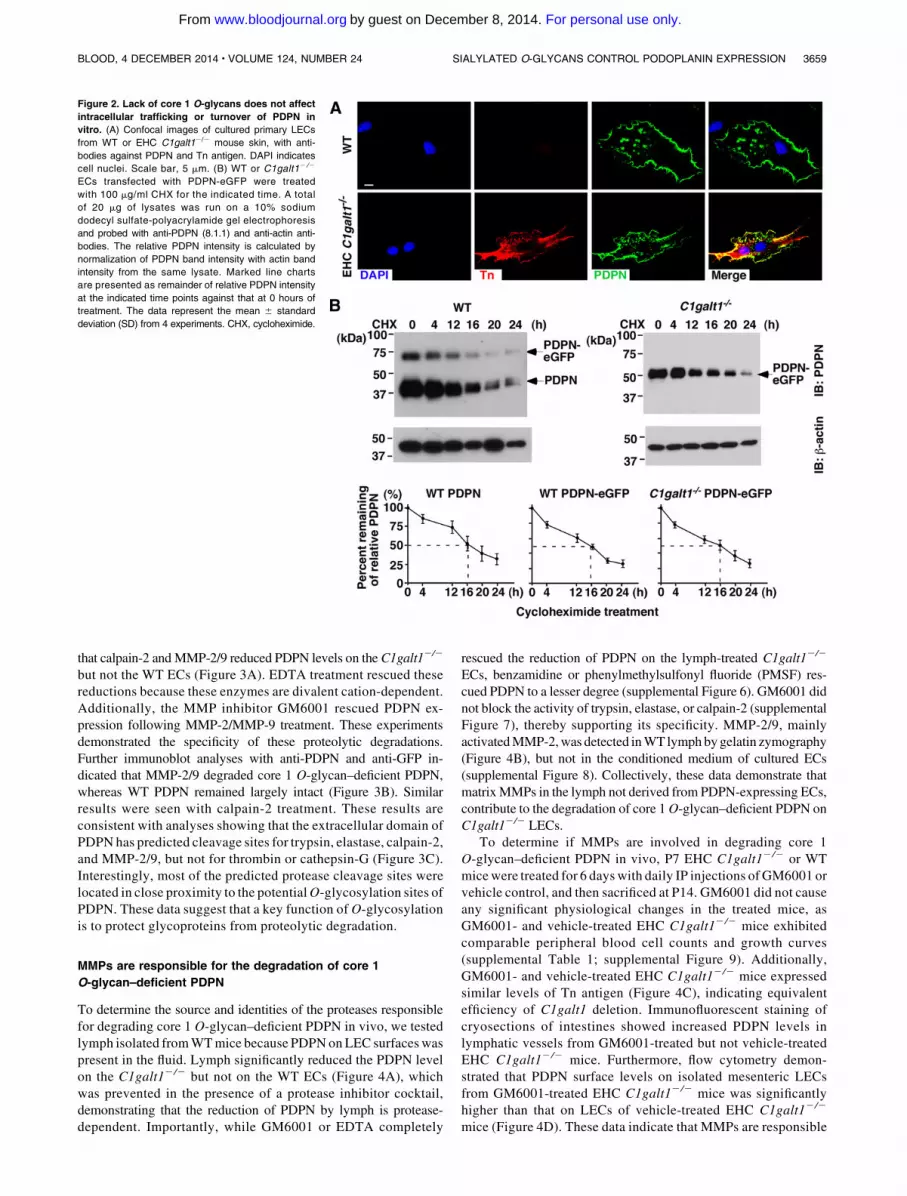
positive, consistent with their LEC phenotype (supplementalFigure 3). Anti-Tn antibody stained C1galt12/2 but not WTcultured primary LECs (Figure 2A). Immunofluorescent stainingof permeabilized cells revealed similar levels of PDPN inside theWT and C1galt12/2 cultured primary LECs, indicating that thelack of O-glycans does not affect the intracellular localization ofPDPN.
Next, we compared the stability of PDPN using PDPN-eGFPtransfected WT or C1galt12/2 EC lines.8 Immortalized WT ECsexpressed endogenous PDPN, but C1galt12/2 ECs had lost theirendogenous PDPN at the messenger RNA level (supplementalFigure 4A). After transfection with PDPN-eGFP, PDPN expressedon theWT and C1galt12/2 EC surfaces was detected similarly bySyrian hamster anti-mouse PDPN monoclonal antibody (mAb,clone 8.1.1), indicating that the antibody binds to PDPN in anO-glycosylation–independent manner (supplemental Figure 4B). BothECs expressed endothelial (CD31) and lymphatic (Lyve-1) markers.Tn antigen was expressed in C1galt12/2 ECs, indicating the loss ofcore 1O-glycans (supplemental Figure 4C). TheWT ECs expressedendogenous PDPN (38 kDa) and PDPN-eGFP (64 kDa), whereas theC1galt12/2 ECs only expressed an underglycosylated form of thetransfected PDPN-eGFP (52 kDa) due to lack of the core 1O-glycan(Figure 2B). PDPN protein synthesis was inhibited by cyclohex-imide, and the decay of PDPN over time was determined byimmunoblot analysis. WT and core 1 O-glycan–deficient PDPN-eGFP, as well as endogenous WT PDPN, were similarly reduced inlevel by 50% at ;16 hours. The internal control b-actin remainedstable for about 20 hours, consistent with published results.25 Takentogether, these results support that lack of core 1 O-glycosylationdoes not affect intracellular synthesis, trafficking, or protein turnoverof PDPN.
Core 1 O-glycans protect PDPN from proteolytic degradation
To test whether core 1 O-glycan–deficient PDPN is sensitiveto proteolytic degradation, we first treated the PDPN-transfectedECs described above with serine protease trypsin. Flow cytometry
demonstrated that trypsin treatment significantly reducedthe PDPN level on the surfaces of C1galt12/2 but not WT ECs.This reduction was rescued by trypsin-like serine protease in-hibitor benzamidine, illustrating its specificity (Figure 3A).The WT or C1galt12 /2 ECs with buffer control or trypsintreatment were also lysed for immunoblot analyses (Figure 3B).Control WT EC lysates exhibited two major bands when probedwith anti-PDPN (8.1.1). Although 8.1.1 detected no obviousdegraded PDPN, anti-GFP antibody recognized a 30-kDa bandfrom lysates of trypsin-treated WT ECs, suggesting that trypsingenerated a minor cleaved product of WT PDPN under theconditions tested in this study. Control core 1 O-glycan–deficientPDPN from the C1galt12/2 ECs was blotted around 52 kDa with8.1.1, whereas 8.1.1 did not detect PDPN from the trypsin-treatedC1galt12/2 ECs. These results suggest that trypsin degradedthe core 1 O-glycan–deficient PDPN and eliminated epitopesrecognized by the 8.1.1 antibody. Consistent with this, anti-GFPantibody detected several fragments of core 1 O-glycan–deficientPDPN after trypsin treatment. To confirm these results, we used acomplementary cell surface biotinylation method (supplementalFigure 5A). Surface-biotinylated nontransfectedWTECs or transfectedstable C1galt12/2 ECs were lysed, and then PDPN in cell lysates wasimmunoprecipitated with anti-PDPN 8.1.1-conjugated agarose beads.The beads were treated with trypsin or buffer control. Immunoblotsusing the eluted proteins demonstrated that the control nontransfectedWT ECs expressed only endogenous 38 kDa PDPN, whereas thecontrol transfected C1galt12/2 ECs expressed 52 kDa PDPN-eGFP. Trypsin treatment resulted in a partial degradation of WTPDPN, whereas it degraded all core 1 O-glycan–deficient PDPN,as no intact PDPN was detected after treatment. Further analysesindicated that core 1 O-glycan–deficient PDPN was also highlysusceptible to degradation by another serine protease, elastase(Figure 3A-B), but not by thrombin, plasmin, or cathepsin-G(supplemental Figure 5B).
To determine whether core 1 O-glycan–deficient PDPN issusceptible to other types of proteases, we examined cysteineprotease calpain-2 and matrix MMP-2/9. Our results indicated
Figure 1. Lack of core 1 O-glycans causes reduced
PDPN levels on LECs. (A) Whole mount images of
WT or EHC C1galt12/2 mouse mesentery at E17 with
antibodies against Prox-1, Tn, and PDPN. Scale bar,
10 mm. (B) Flow cytometric analyses of cell surface
PDPN levels on freshly isolated mesenteric LECs from
WT or EHC C1galt12/2 mice at P1. Dot plots are
shown after gating on CD311/Lyve-11 LEC cells.
Percentage and mean fluorescence intensity (MFI) (in
parenthesis) are indicated for PDPN in each quadrant.
(C, left) C1galt1f/f;tetO-Cre1 mice and littermate WT
mice were fed a doxycycline diet for 6 weeks, starting
at 6-weeks old. Confocal images of ileal cryosections
from doxycycline-inducible or littermate control mice
with antibodies against PDPN and Lyve-1 are shown.
Scale bar, 20 mm. (C, right) Flow cytometric analyses
of PDPN on CD311/Lyve-11 isolated mesenteric LECs
from doxycycline-inducible or littermate control mice,
and (D) CD311/Lyve-11 isolated mesenteric LECs from
12-week-old WT or Clec22/2 mice. The images and
data are representative of at least 3 experiments. E17,
embryonic day 17; P1, postnatal day. SSC, side scatter.
3658 PAN et al BLOOD, 4 DECEMBER 2014 x VOLUME 124, NUMBER 24
For personal use only.on December 8, 2014. by guest www.bloodjournal.orgFrom
that calpain-2 and MMP-2/9 reduced PDPN levels on the C1galt12/2
but not the WT ECs (Figure 3A). EDTA treatment rescued thesereductions because these enzymes are divalent cation-dependent.Additionally, the MMP inhibitor GM6001 rescued PDPN ex-pression following MMP-2/MMP-9 treatment. These experimentsdemonstrated the specificity of these proteolytic degradations.Further immunoblot analyses with anti-PDPN and anti-GFP in-dicated that MMP-2/9 degraded core 1 O-glycan–deficient PDPN,whereas WT PDPN remained largely intact (Figure 3B). Similarresults were seen with calpain-2 treatment. These results areconsistent with analyses showing that the extracellular domain ofPDPN has predicted cleavage sites for trypsin, elastase, calpain-2,and MMP-2/9, but not for thrombin or cathepsin-G (Figure 3C).Interestingly, most of the predicted protease cleavage sites werelocated in close proximity to the potentialO-glycosylation sites ofPDPN. These data suggest that a key function of O-glycosylationis to protect glycoproteins from proteolytic degradation.
MMPs are responsible for the degradation of core 1
O-glycan–deficient PDPN
To determine the source and identities of the proteases responsiblefor degrading core 1 O-glycan–deficient PDPN in vivo, we testedlymph isolated fromWTmice because PDPN onLEC surfaces waspresent in the fluid. Lymph significantly reduced the PDPN levelon the C1galt12/2 but not on the WT ECs (Figure 4A), whichwas prevented in the presence of a protease inhibitor cocktail,demonstrating that the reduction of PDPN by lymph is protease-dependent. Importantly, while GM6001 or EDTA completely
rescued the reduction of PDPN on the lymph-treated C1galt12/2
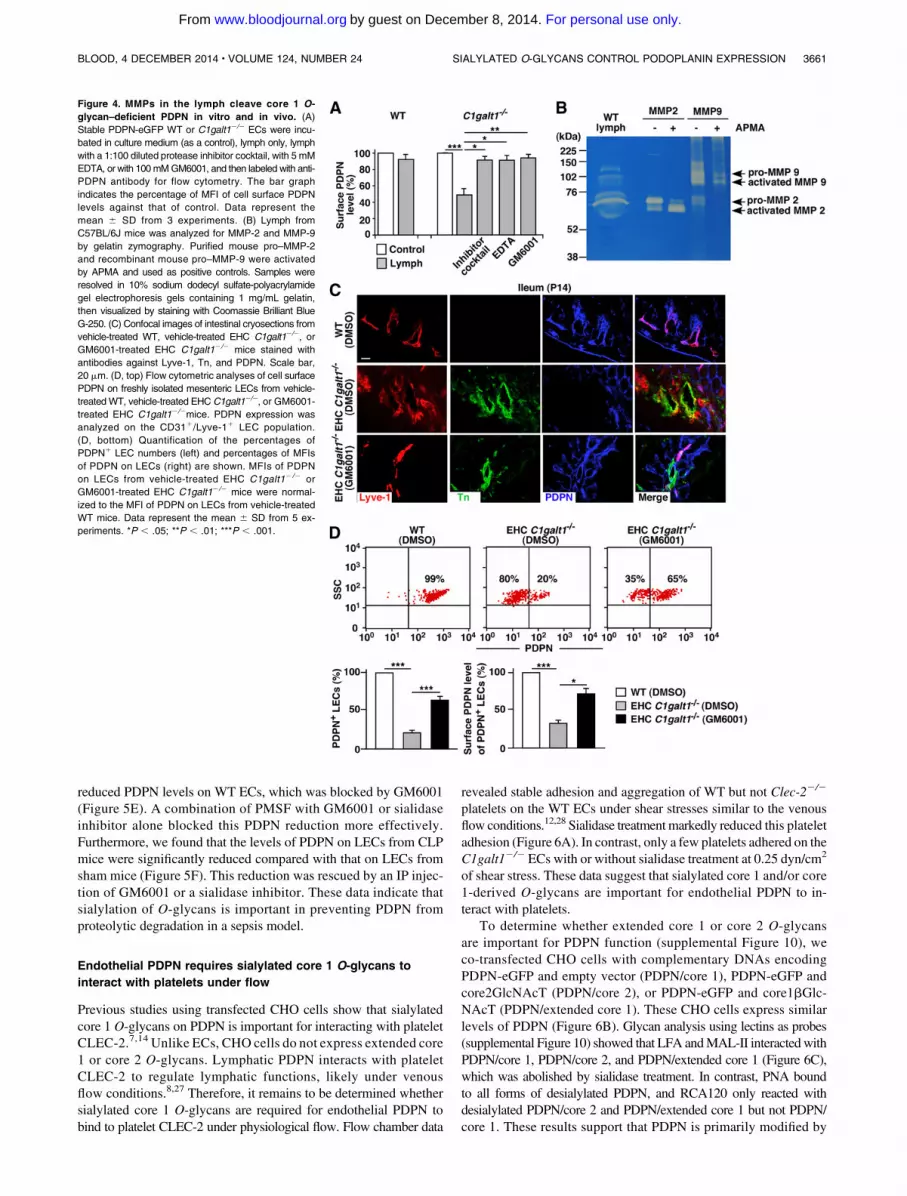
ECs, benzamidine or phenylmethylsulfonyl fluoride (PMSF) res-cued PDPN to a lesser degree (supplemental Figure 6). GM6001 didnot block the activity of trypsin, elastase, or calpain-2 (supplementalFigure 7), thereby supporting its specificity. MMP-2/9, mainlyactivatedMMP-2,was detected inWT lymphbygelatin zymography(Figure 4B), but not in the conditioned medium of cultured ECs(supplemental Figure 8). Collectively, these data demonstrate thatmatrixMMPs in the lymph not derived from PDPN-expressing ECs,contribute to the degradation of core 1O-glycan–deficient PDPN onC1galt12/2 LECs.
To determine if MMPs are involved in degrading core 1O-glycan–deficient PDPN in vivo, P7 EHC C1galt12/2 or WTmice were treated for 6 dayswith daily IP injections of GM6001 orvehicle control, and then sacrificed at P14. GM6001 did not causeany significant physiological changes in the treated mice, asGM6001- and vehicle-treated EHC C1galt12/2 mice exhibitedcomparable peripheral blood cell counts and growth curves(supplemental Table 1; supplemental Figure 9). Additionally,GM6001- and vehicle-treated EHC C1galt12/2 mice expressedsimilar levels of Tn antigen (Figure 4C), indicating equivalentefficiency of C1galt1 deletion. Immunofluorescent staining ofcryosections of intestines showed increased PDPN levels inlymphatic vessels from GM6001-treated but not vehicle-treatedEHC C1galt12/2 mice. Furthermore, flow cytometry demon-strated that PDPN surface levels on isolated mesenteric LECsfrom GM6001-treated EHC C1galt12/2 mice was significantlyhigher than that on LECs of vehicle-treated EHC C1galt12/2
mice (Figure 4D). These data indicate that MMPs are responsible
Figure 2. Lack of core 1 O-glycans does not affect
intracellular trafficking or turnover of PDPN in
vitro. (A) Confocal images of cultured primary LECs
from WT or EHC C1galt12/2 mouse skin, with anti-
bodies against PDPN and Tn antigen. DAPI indicates
cell nuclei. Scale bar, 5 mm. (B) WT or C1galt12 /2
ECs transfected with PDPN-eGFP were treated
with 100 mg/ml CHX for the indicated time. A total
of 20 mg of lysates was run on a 10% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis
and probed with anti-PDPN (8.1.1) and anti-actin anti-
bodies. The relative PDPN intensity is calculated by
normalization of PDPN band intensity with actin band
intensity from the same lysate. Marked line charts
are presented as remainder of relative PDPN intensity
at the indicated time points against that at 0 hours of
treatment. The data represent the mean 6 standard
deviation (SD) from 4 experiments. CHX, cycloheximide.
BLOOD, 4 DECEMBER 2014 x VOLUME 124, NUMBER 24 SIALYLATED O-GLYCANS CONTROL PODOPLANIN EXPRESSION 3659
For personal use only.on December 8, 2014. by guest www.bloodjournal.orgFrom

for proteolytic degradation of PDPN on LECs in EHC C1galt12/2
mice.
Sialylated O-glycans protect PDPN from proteolysis in the CLP
sepsis model
Sialic acids are the common capping structure of O-glycans.Using the WT and C1galt12/2 ECs with similar levels of PDPN(Figure 5A) that was not altered by sialidase treatment, the glycanprofile showed that limax flavus agglutinin (LFA) (to sialic acids)andmaackia amurensis lectin II (MAL-II) (toa2,3 sialylated glycans),but not peanut agglutinin (PNA) (to desialylated core 1 O-glycans),sambucus nigra agglutinin (toa2,6 sialylated GalNAc-Ser/Thr), helixpomatia agglutinin (HPA) (to Tn antigen), and ricinus communisagglutinin (RCA120) (to LacNAc), interacted strongly with WTPDPN (Figure 5B). Sialidase treatment decreased reactivity of WTPDPN with LFA and MAL-II, and increased that with PNA andRCA120. These results indicate that WT PDPN is modified bya2,3 sialylated core 1 and core 1-derived O-glycans (supplemental
Figure 10). In contrast, core 1 O-glycan–deficient PDPN with orwithout sialidase treatment reacted only with HPA, indicating thatcore 1 O-glycan–deficient PDPN presents terminal GalNAc withoutdetectable sialylation. These data suggest that lack of sialylation orcore 1O-glycosylation, or both in combination,may contribute to theincreased PDPN degradation on C1galt12/2 LECs. To test this, wepretreated theWT ECs with sialidases and then incubated them withMMP-2/9 or trypsin (Figure 5C and supplemental Figure 11).Surface level of PDPN on sialidase-treated WT ECs was reduced byMMP-2/9 or trypsin, but not as much as on the C1galt12/2 ECs.These reductions were rescued with GM6001 or benzamidine. Thisresult supports that lack of sialylation is sufficient to cause significantdegradation of PDPN.
To determine the pathological relevance of this finding, we useda CLP sepsis model, which has increased sialidase activity inserum.26 Firstly, lymph was collected from CLP or sham mice.Consistent with the published data, increased sialidase activity wasdetected in the CLP lymph, which was blocked by the sialidaseinhibitor but not in the sham lymph (Figure 5D). The CLP lymph
Figure 3. Core 1 O-glycans protect PDPN from pro-
teolytic degradation. (A) Stable PDPN-eGFP trans-
fected WT or C1galt12/2 ECs were incubated in buffer
(control), 0.05% trypsin (with or without 5 mM
benzamidine), 100 mg/ml elastase (with or without
2 mMPMSF), 10 mg/ml calpain-2 (with 2 mMCa11, with
or without 5 mM EDTA), or 2 mg/ml activated MMP-2/
MMP-9 (with or without 100 mM GM6001), and then
stained with anti-PDPN antibody for flow cytometry.
The bar graph indicates the percentage of MFI of cell
surface PDPN levels against that of control. The data
represent the mean 6 SD from 3 to 5 experiments.
***P , .001. (B) Stable WT or C1galt12/2 ECs were
treated with or without trypsin, elastase, calpain-2, or
activated MMP-2/MMP-9, and then lysed. All lysates
were analyzed by immunoblot using anti-PDPN
mAb (8.1.1) or rabbit anti-GFP Ab. Glyceraldehyde-3-
phosphate dehydrogenase was used as a loading
control. (C) Potential O-glycosylation sites (black dots)
and predicted cleavage sites of PDPN by trypsin
(yellow), elastase (green), calpain-2 (red), or MMP-2/
MMP-9 (purple) in the extracellular domain of PDPN.
3660 PAN et al BLOOD, 4 DECEMBER 2014 x VOLUME 124, NUMBER 24
For personal use only.on December 8, 2014. by guest www.bloodjournal.orgFrom
reduced PDPN levels on WT ECs, which was blocked by GM6001(Figure 5E). A combination of PMSF with GM6001 or sialidaseinhibitor alone blocked this PDPN reduction more effectively.Furthermore, we found that the levels of PDPN on LECs from CLPmice were significantly reduced compared with that on LECs fromsham mice (Figure 5F). This reduction was rescued by an IP injec-tion of GM6001 or a sialidase inhibitor. These data indicate thatsialylation of O-glycans is important in preventing PDPN fromproteolytic degradation in a sepsis model.
Endothelial PDPN requires sialylated core 1 O-glycans to
interact with platelets under flow
Previous studies using transfected CHO cells show that sialylatedcore 1 O-glycans on PDPN is important for interacting with plateletCLEC-2.7,14 Unlike ECs, CHO cells do not express extended core1 or core 2 O-glycans. Lymphatic PDPN interacts with plateletCLEC-2 to regulate lymphatic functions, likely under venousflow conditions.8,27 Therefore, it remains to be determined whethersialylated core 1 O-glycans are required for endothelial PDPN tobind to platelet CLEC-2 under physiological flow. Flow chamber data
revealed stable adhesion and aggregation of WT but not Clec-22/2
platelets on the WT ECs under shear stresses similar to the venousflow conditions.12,28 Sialidase treatmentmarkedly reduced this plateletadhesion (Figure 6A). In contrast, only a few platelets adhered on theC1galt12/2 ECs with or without sialidase treatment at 0.25 dyn/cm2
of shear stress. These data suggest that sialylated core 1 and/or core1-derived O-glycans are important for endothelial PDPN to in-teract with platelets.
To determine whether extended core 1 or core 2 O-glycansare important for PDPN function (supplemental Figure 10), weco-transfected CHO cells with complementary DNAs encodingPDPN-eGFP and empty vector (PDPN/core 1), PDPN-eGFP andcore2GlcNAcT (PDPN/core 2), or PDPN-eGFP and core1bGlc-NAcT (PDPN/extended core 1). These CHO cells express similarlevels of PDPN (Figure 6B). Glycan analysis using lectins as probes(supplemental Figure 10) showed that LFA andMAL-II interactedwithPDPN/core 1, PDPN/core 2, and PDPN/extended core 1 (Figure 6C),which was abolished by sialidase treatment. In contrast, PNA boundto all forms of desialylated PDPN, and RCA120 only reacted withdesialylated PDPN/core 2 and PDPN/extended core 1 but not PDPN/core 1. These results support that PDPN is primarily modified by
Figure 4. MMPs in the lymph cleave core 1 O-
glycan–deficient PDPN in vitro and in vivo. (A)
Stable PDPN-eGFP WT or C1galt12/2 ECs were incu-
bated in culture medium (as a control), lymph only, lymph
with a 1:100 diluted protease inhibitor cocktail, with 5 mM
EDTA, or with 100mMGM6001, and then labeled with anti-
PDPN antibody for flow cytometry. The bar graph
indicates the percentage of MFI of cell surface PDPN
levels against that of control. Data represent the
mean 6 SD from 3 experiments. (B) Lymph from
C57BL/6J mice was analyzed for MMP-2 and MMP-9
by gelatin zymography. Purified mouse pro–MMP-2
and recombinant mouse pro–MMP-9 were activated
by APMA and used as positive controls. Samples were
resolved in 10% sodium dodecyl sulfate-polyacrylamide
gel electrophoresis gels containing 1 mg/mL gelatin,
then visualized by staining with Coomassie Brilliant Blue
G-250. (C) Confocal images of intestinal cryosections from
vehicle-treated WT, vehicle-treated EHC C1galt12/2, or
GM6001-treated EHC C1galt12/2 mice stained with
antibodies against Lyve-1, Tn, and PDPN. Scale bar,
20 mm. (D, top) Flow cytometric analyses of cell surface
PDPN on freshly isolated mesenteric LECs from vehicle-
treatedWT, vehicle-treated EHC C1galt12/2, or GM6001-
treated EHC C1galt12/2mice. PDPN expression was
analyzed on the CD311/Lyve-11 LEC population.
(D, bottom) Quantification of the percentages of
PDPN1 LEC numbers (left) and percentages of MFIs
of PDPN on LECs (right) are shown. MFIs of PDPN
on LECs from vehicle-treated EHC C1galt12/2 or
GM6001-treated EHC C1galt12/2 mice were normal-
ized to the MFI of PDPN on LECs from vehicle-treated
WT mice. Data represent the mean 6 SD from 5 ex-
periments. *P , .05; **P , .01; ***P , .001.
BLOOD, 4 DECEMBER 2014 x VOLUME 124, NUMBER 24 SIALYLATED O-GLYCANS CONTROL PODOPLANIN EXPRESSION 3661
For personal use only.on December 8, 2014. by guest www.bloodjournal.orgFrom

a sialylated core 1 structure.Consistentwith this,WTplatelets interactedsimilarly with CHO cells expressing PDPN/core 1 and with PDPN/core 2 (Figure 6D). Interestingly, extended core 1 O-glycosylationappeared to reduce PDPN interaction with WT platelets, as there wasa modest reduction of platelet binding to CHO cells with PDPN/extended core 1. Taken together, sialylated core 1 O-glycosylation ofPDPN on ECs is sufficient to interact with platelet CLEC-2 under flow.
Discussion
We previously reported that core 1 O-glycosylation is critical for theexpression of functional PDPN during the development of lymphaticvessels.8 However, the mechanism of how O-glycosylation regulatesPDPN function and expression was unclear. In this study, we dem-onstrate that sialylated core 1O-glycosylation is essential for the stabilityof PDPN, by protecting it from proteolytic degradation byMMPs in thelymph, and for platelet adhesion and aggregation on LECs under flowconditions.
The protective role of O-glycosylation against proteolysis ofmodified proteins has been previously demonstrated in vitro.19,21
However, the nature of the proteases and whether this functionis important in vivo have not been elucidated. Our results showthat core 1 O-glycan–deficient PDPN is susceptible to differ-ent types of proteases that have predicted cleavage sites in theextracellular domain of PDPN, such as serine protease, trypsinand elastase, cysteine protease calpain-2, and MMP2/9. In-terestingly, most of the predicted protease cleavage sites arelocated in close proximity to the potential O-glycosylation sitesof PDPN (Figure 3C).Mucin-typeO-glycoproteins typically havea rod conformation.29 The first a-linked GalNAc is essential forthe extended conformation.30 Because the a-GalNAc remainsattached to the peptide backbone of PDPN in the absence of core 1O-glycosylation, it is unlikely that the increased proteolyticdegradation of core 1-deficient PDPN is due to its conformationalchanges that favor the accessibility of proteases to their cleavagesites.
Negatively charged sialic acids are common capping struc-tures in O-glycans and are reported to be important for protecting
Figure 5. Sialic acids play an important role in the
protection of PDPN from proteolytic degradation.
(A) Flow cytometric analyses of cell surface PDPN
on WT or C1galt12/2 ECs with or without sialidase
treatment. Cells were incubated with hamster anti-
PDPN antibody 8.1.1, followed by PE-conjugated anti-
hamster IgG. (B) Glycan profile of WT PDPN-eGFP or
core 1 O-glycan–deficient PDPN-GFP with or with-
out sialidase treatment. WT PDPN-eGFP or core 1
O-glycan–deficient PDPN-eGFP was purified by mouse
anti-GFP monoclonal antibody agarose beads from WT
ECs or C1galt12/2 ECs. The fluorescence intensity of
anti-PDPN antibody or lectin binding is indicated with
bars. Lectin specificities are as follows: LFA (sialic acid);
MAL-II (Siaa2-3Galb1-3 6 [Siaa2-6]GalNAc); sambucus
nigra agglutinin (Siaa2-6Gal or Siaa2-6GalNAc); PNA
(desialylated Galb1-3GalNAc [core 1]); HPA (terminal
GalNAc [Tn]); and RCA120 (desialylated Galb1-4GlcNAc
[LacNAc]). The data represent the mean 6 SD from
3 experiments. (C) WT ECs were pretreated with
sialidase and then incubated in buffer (as a control), or
2 mg/ml activated MMP-2/MMP-9 with or without
100 mMGM6001. The cells were stained with anti-PDPN
antibody for flow cytometry. The bar graph indicates
the percentage of MFI of cell surface PDPN levels against
that of control. The data represent the mean6 SD from 3
experiments. (D) Sialidase activity was measured
using RCA120 reactivity with fetuin treated by culture
medium, the lymph from sham, the lymph from CLP
mice, or the lymph from CLP mice with 300 mg/ml of
sialidase inhibitor at 37°C for 2 hours. The fluores-
cence intensity of RCA120 binding is indicated with
bars. The data represent the mean 6 SD from 3
experiments. (E) WT ECs were incubated in culture
medium (as a control), lymph from sham, or lymph from
CLP mice with or without 100 mM GM6001, 100 mM
GM6001 and 2 mM PMSF, or 300 mg/ml sialidase
inhibitor. The bar graph indicates the percentage of
MFI of cell surface PDPN levels against that of control.
The data represent the mean 6 SD from 3 experi-
ments. (F, top) Flow cytometric analyses of cell sur-
face PDPN on isolated mesenteric LECs from mice
treated by sham with vehicle, CLP with vehicle, CLP
with GM6001, or CLP with sialidase inhibitor. PDPN
expression was analyzed on CD311/Lyve-11 LEC
population. (F, bottom) Quantification of the percent-
ages of PDPN1 LEC numbers (left) and percentages
of MFIs of PDPN on LECs (right) are shown. MFIs
of PDPN on LECs from CLP mice were normalized
to the MFI of PDPN on LECs from sham mice. Data
represent the mean 6 SD from 4 experiments.
*P , .05; **P , .01; ***P , .001. ND, not detected.
3662 PAN et al BLOOD, 4 DECEMBER 2014 x VOLUME 124, NUMBER 24
For personal use only.on December 8, 2014. by guest www.bloodjournal.orgFrom

glycoproteins from proteolysis.31-33 Adding exogenous GalNActo ldl-DCHO cells, which cannot synthesize uridine diphosphate-Gal/uridine diphosphate-GalNAc, generates sialylated and more stableproteins against proteolysis comparedwith theproteinswithoutGalNAc,suggesting a protective mechanism of sialylated GalNAc.19,34 Inter-estingly, core 1 O-glycan–deficient PDPN is not sialylated. Thelack of sialylation may contribute to increased proteolysis ofcore 1 O-glycan–deficient PDPN. Supporting this idea, our datashowed that desialylation significantly increased susceptibility ofPDPN to the proteases, but not as much as lack of core 1O-glycan(supplemental Figure 11). Core 1 O-glycans can be extended toform extended core 1 or core 2 O-glycans. Mice lacking theseglycan structures35,36 or lacking core 3-derived O-glycans (in-cluding core 4),37 do not exhibit lymphatic vascular defects thatresemble mice without core 1 O-glycans. Together, these dataindicate that sialylation and basic core 1 structure cooperate to-gether to protect PDPN from proteolytic degradation. Desialyla-tion of glycans are known to contribute to increased clearanceof platelets and vonWillebrand factor in bacteria-mediated sepsismodels.38 Consistently, increased sialidase activity in serum isreported in sepsis patients39 and in rodent CLP sepsis models.26
Our data showed the increased sialidase in the lymph from CLPmice, which caused desialylation and promoted degradationof PDPN on LECs primarily by MMPs in the lymph. Althoughit remains to be further determined, these results imply thatdesialylation of O-glycans leads to abnormal lymphatic functionin pathological conditions, such as sepsis.
The lymph of humans and rodents contains proteases, includingMMP-2/9, and serine proteases thrombin, plasmin, and factor X.40,41
Therefore, although our in vitro and in vivo analyses showedthat MMPs are a major form of proteases in degrading core 1O-glycan–deficient or desialylated PDPN, other proteases such asserine proteases, may also be involved in the regulation of PDPNlevels.42-44
PDPN induces platelet aggregation by interacting with CLEC-2on platelets.12,45 Previous studies using transfected CHO cells andthe glioblastoma cell line6 demonstrated that sialylated core 1 O-glycans on PDPN is required for platelet aggregation.7,14 Our results,using Lyve-1–positive ECs show that sialylated core 1 structure ispresent on WT PDPN, consistent with published data.6 Plateletsadhered and aggregated onWT, but not on core 1O-glycan–deficientPDPN, under shears. Our studies further demonstrate that extendedcore 1 or core 2O-glycans are not required for PDPN to interact withplatelet CLEC-2. These results support that sialylated core 1 O-glycan on endothelial PDPN is sufficient to mediate stable plateletadhesion and aggregation under shears.
Recently, PDPN-mediated platelet activation has been found tobe critical in maintaining the integrity of high endothelial venules inthe lymph node and for lymphovenous hemostasis in the adultmouse.13,27 Therefore, O-glycosylation in regulating the functionand expression of PDPN is important not only for the developmentof an independent lymphatic vascular system, but also for themaintenance of the established blood-lymphatic vascular systemsthroughout life. Abnormal glycosylation of PDPNmay contribute to
Figure 6. Sialylated core 1 O-glycans of PDPN
plays an important role in platelet adhesion on ECs
under flow. (A) PHK26 labeled platelets from WT or
platelet-specific Clec-22/2 mice were perfused and
allowed to accumulate on WT or C1galt12/2 ECs with
or without sialidase treatment under shear stress of
0.25 dyn/cm2 for 10 minutes. After changing to platelet-
free buffer, fluid shear stress was increased every 30
seconds. Dual bright field and fluorescence images of
platelet adhesion and aggregation on ECs under shear
stress of 0.25 dyn/cm2 are shown. Scale bar, 20 mm. The
images are representative of 3 independent experi-
ments. ECs in bright fields are shown as blue color,
and fluorescent PHK-labeled platelets are shown as
red color. The data quantify the covered platelet area
on each EC under the indicated shear stresses and
represent the mean 6 SD from 3 experiments. (B)
Flow cytometric analyses of cell surface PDPN on
CHO cells transfected with PDPN-eGFP and control
vector (PDPN/core1), PDPN-eGFP and core2GlcNAcT
(PDPN/core2), or PDPN-eGFP and core1bGlcNAcT
(PDPN/extended core1). Cells were incubated with
hamster anti-PDPN antibody 8.1.1, followed by PE-
conjugated anti-hamster IgG. (C) Glycan profile of
PDPN/core1, PDPN/core2, or PDPN/extended core 1
with or without sialidase treatment is shown. PDPN-
eGFP from different CHO cell lines was captured on
agarose beads conjugated with mouse anti-GFP
monoclonal antibody. Glycan profile of PDPN was
analyzed based on lectin binding. See Figure 5 for
lectin specificities. The fluorescence intensity of anti-
PDPN antibody or lectin binding is indicated in the bar
graphs. The data represent the mean 6 SD from 3
experiments. (D) PDPN/core1, PDPN/core2, or PDPN/
extended core1 CHO cells were cultured in 35 mm
dishes, and PHK26-labeled platelets from WT were
perfused and allowed to accumulate on CHO cells
under shear stress of 0.25 dyn/cm2 for 10 minutes. The
data quantify the covered platelet area on CHO
cells under the indicated shear stress, and repre-
sent the mean 6 SD from 3 experiments. **P , .01.
Plt, platelets.
BLOOD, 4 DECEMBER 2014 x VOLUME 124, NUMBER 24 SIALYLATED O-GLYCANS CONTROL PODOPLANIN EXPRESSION 3663
For personal use only.on December 8, 2014. by guest www.bloodjournal.orgFrom

pathogenesis of diseases such as sepsis, as demonstrated in the CLPmodel.
Acknowledgments
This study was supported by grants from the National Institutes ofHealth (National Institute of General Medical Sciences: GM103441;and National Heart, Lung, and Blood Institute: HL085607, HL093242,and HL118676), Department of Defense (W81XWH-11-1-0226),Oklahoma Center for the Advancement of Science (HR13-160 andHR13-020), National Natural Science Foundation of China (30928010and 31400692), Jiangsu Provincial Special Program ofMedical Science(BL2012005), Jiangsu Province’s Key Medical Center (ZX201102),and the American Heart Association (SDG7410022).
Authorship
Contribution: Y.P., T.Y., B.H., and J.M.M. performed research;Y.P., T.Y., J.F., P.M-D., X.C., C.R., R.P.M., C.W., K.D., H.C., andL.X. analyzed data; and T.Y. and L.X. designed research and wrotethe paper.
Conflict-of-interest disclosure: The authors declare no competingfinancial interests.
Correspondence: Lijun Xia, Cardiovascular Biology ResearchProgram, Oklahoma Medical Research Foundation, MS #45, 825N.E. 13th St, Oklahoma City, OK 73104; e-mail: [email protected];and Tadayuki Yago, Cardiovascular Biology Research Program,Oklahoma Medical Research Foundation, 825 NE 13th St, OklahomaCity, OK 73104; e-mail: [email protected].
References
1. Watanabe M, Okochi E, Sugimoto Y, Tsuruo T.Identification of a platelet-aggregating factorof murine colon adenocarcinoma 26: Mr44,000 membrane protein as determined bymonoclonal antibodies. Cancer Res. 1988;48(22):6411-6416.
2. Breiteneder-Geleff S, Matsui K, Soleiman A, et al.Podoplanin, novel 43-kd membrane protein ofglomerular epithelial cells, is down-regulated inpuromycin nephrosis. Am J Pathol. 1997;151(4):1141-1152.
3. Ramirez MI, Millien G, Hinds A, Cao Y, Seldin DC,Williams MC. T1alpha, a lung type I celldifferentiation gene, is required for normal lungcell proliferation and alveolus formation at birth.Dev Biol. 2003;256(1):61-72.
4. Schacht V, Ramirez MI, Hong YK, et al. T1alpha/podoplanin deficiency disrupts normal lymphaticvasculature formation and causes lymphedema.EMBO J. 2003;22(14):3546-3556.
5. Schietinger A, Philip M, Yoshida BA, et al.A mutant chaperone converts a wild-type proteininto a tumor-specific antigen. Science. 2006;314(5797):304-308.
6. Kaneko MK, Kato Y, Kameyama A, et al.Functional glycosylation of human podoplanin:glycan structure of platelet aggregation-inducingfactor. FEBS Lett. 2007;581(2):331-336.
7. Kaneko M, Kato Y, Kunita A, Fujita N, Tsuruo T,Osawa M. Functional sialylated O-glycan toplatelet aggregation on Aggrus (T1alpha/Podoplanin) molecules expressed in Chinesehamster ovary cells. J Biol Chem. 2004;279(37):38838-38843.
8. Fu J, Gerhardt H, McDaniel JM, et al. Endothelialcell O-glycan deficiency causes blood/lymphaticmisconnections and consequent fatty liverdisease in mice. J Clin Invest. 2008;118(11):3725-3737.
9. Ju T, Brewer K, D’Souza A, Cummings RD,Canfield WM. Cloning and expression of humancore 1 beta1,3-galactosyltransferase. J BiolChem. 2002;277(1):178-186.
10. Xia L, Ju T, Westmuckett A, et al. Defectiveangiogenesis and fatal embryonic hemorrhage inmice lacking core 1-derived O-glycans. J Cell Biol.2004;164(3):451-459.
11. Ju T, Cummings RD, Canfield WM. Purification,characterization, and subunit structure of rat core1 Beta1,3-galactosyltransferase. J Biol Chem.2002;277(1):169-177.
12. Bertozzi CC, Schmaier AA, Mericko P,et al. Platelets regulate lymphatic vasculardevelopment through CLEC-2-SLP-76 signaling.Blood. 2010;116(4):661-670.
13. Herzog BH, Fu J, Wilson SJ, et al. Podoplaninmaintains high endothelial venule integrity byinteracting with platelet CLEC-2. Nature. 2013;502(7469):105-109.
14. Kato Y, Kaneko MK, Kunita A, et al. Molecularanalysis of the pathophysiological binding of theplatelet aggregation-inducing factor podoplaninto the C-type lectin-like receptor CLEC-2. CancerSci. 2008;99(1):54-61.
15. Mitoma J, Petryniak B, Hiraoka N, Yeh JC,Lowe JB, Fukuda M. Extended core 1 andcore 2 branched O-glycans differentiallymodulate sialyl Lewis X-type L-selectinligand activity. J Biol Chem. 2003;278(11):9953-9961.
16. Wang Y, Zhao L, Suzuki A, et al. Plateletslacking PIP5KIg have normal integrin activationbut impaired cytoskeletal-membrane integrityand adhesion. Blood. 2013;121(14):2743-2752.
17. Alfalah M, Jacob R, Preuss U, Zimmer KP, Naim H,Naim HY. O-linked glycans mediate apical sortingof human intestinal sucrase-isomaltase throughassociation with lipid rafts. Curr Biol. 1999;9(11):593-596.
18. Altschuler Y, Kinlough CL, Poland PA, et al.Clathrin-mediated endocytosis of MUC1 ismodulated by its glycosylation state. Mol Biol Cell.2000;11(3):819-831.
19. Kozarsky K, Kingsley D, Krieger M. Use ofa mutant cell line to study the kinetics and functionof O-linked glycosylation of low density lipoproteinreceptors. Proc Natl Acad Sci USA. 1988;85(12):4335-4339.
20. Potter BA, Hughey RP, Weisz OA. Role of N- andO-glycans in polarized biosynthetic sorting. Am JPhysiol Cell Physiol. 2006;290(1):C1-C10.
21. Reddy P, Caras I, Krieger M. Effects ofO-linked glycosylation on the cell surfaceexpression and stability of decay-acceleratingfactor, a glycophospholipid-anchoredmembrane protein. J Biol Chem. 1989;264(29):17329-17336.
22. Tian E, Ten Hagen KG. Recent insights intothe biological roles of mucin-type O-glycosylation.Glycoconj J. 2009;26(3):325-334.
23. Valencia JC, Rouzaud F, Julien S, et al. Sialylatedcore 1 O-glycans influence the sorting of Pmel17/gp100 and determine its capacity to form fibrils.J Biol Chem. 2007;282(15):11266-11280.
24. van der Post S, Subramani DB, Backstrom M,et al. Site-specific O-glycosylation on theMUC2 mucin protein inhibits cleavage by thePorphyromonas gingivalis secreted cysteineprotease (RgpB). J Biol Chem. 2013;288(20):14636-14646.
25. Antecol MH, Darveau A, Sonenberg N,Mukherjee BB. Altered biochemical propertiesof actin in normal skin fibroblasts fromindividuals predisposed to dominantly inheritedcancers. Cancer Res. 1986;46(4, pt 1):1867-1873.
26. Hof L, Loegering DJ. Increase of plasmaneuraminidase activity in experimental peritonitis.Proc Soc Exp Biol Med. 1982;169(4):501-505.
27. Hess PR, Rawnsley DR, Jakus Z, et al. Plateletsmediate lymphovenous hemostasis to maintainblood-lymphatic separation throughout life. J ClinInvest. 2014;124(1):273-284.
28. Malek AM, Alper SL, Izumo S. Hemodynamicshear stress and its role in atherosclerosis. JAMA.1999;282(21):2035-2042.
29. Jentoft N. Why are proteins O-glycosylated?Trends Biochem Sci. 1990;15(8):291-294.
30. Coltart DM, Royyuru AK, Williams LJ, et al.Principles of mucin architecture: structuralstudies on synthetic glycopeptides bearingclustered mono-, di-, tri-, and hexasaccharideglycodomains. J Am Chem Soc. 2002;124(33):9833-9844.
31. Varki A, Gagneux P. Multifarious roles of sialicacids in immunity. Ann N Y Acad Sci. 2012;1253:16-36.
32. McGrath RT, McKinnon TA, Byrne B, et al.Expression of terminal alpha2-6-linked sialic acidon von Willebrand factor specifically enhancesproteolysis by ADAMTS13. Blood. 2010;115(13):2666-2673.
33. Varki A, Schauer R. Sialic Acids. In: Varki A,Cummings RD, Esko JD, et al., eds. Essentials ofGlycobiology. 2nd ed. Cold Spring Harbor, NY:Cold Spring Harbor Laboratory Press; 2009.
34. Kozarsky KF, Call SM, Dower SK, Krieger M.Abnormal intracellular sorting of O-linkedcarbohydrate-deficient interleukin-2 receptors.Mol Cell Biol. 1988;8(8):3357-3363.
35. Mitoma J, Bao X, Petryanik B, et al. Criticalfunctions of N-glycans in L-selectin-mediated lymphocyte homing and recruitment.
Nat Immunol. 2007;8(4):409-418.
36. Stone EL, Ismail MN, Lee SH, et al.Glycosyltransferase function in core 2-typeprotein O glycosylation. Mol Cell Biol. 2009;29(13):3770-3782.
37. An G, Wei B, Xia B, et al. Increased susceptibilityto colitis and colorectal tumors in mice lackingcore 3-derived O-glycans. J Exp Med. 2007;204(6):1417-1429.
38. Grewal PK, Uchiyama S, Ditto D, et al. TheAshwell receptor mitigates the lethal
3664 PAN et al BLOOD, 4 DECEMBER 2014 x VOLUME 124, NUMBER 24
For personal use only.on December 8, 2014. by guest www.bloodjournal.orgFrom

coagulopathy of sepsis. Nat Med. 2008;14(6):648-655.
39. Piagnerelli M, Boudjeltia KZ, Rapotec A, et al.Neuraminidase alters red blood cells in sepsis.Crit Care Med. 2009;37(4):1244-1250.
40. Clement CC, Aphkhazava D, Nieves E, et al.Protein expression profiles of human lymph andplasma mapped by 2D-DIGE and 1D SDS-PAGEcoupled with nanoLC-ESI-MS/MS bottom-upproteomics. J Proteomics. 2013;78:172-187.
41. Mittal A, Middleditch M, Ruggiero K, et al. Theproteome of rodent mesenteric lymph. Am JPhysiol Gastrointest Liver Physiol. 2008;295(5):G895-G903.
42. Dollery CM, Libby P. Atherosclerosis andproteinase activation. Cardiovasc Res. 2006;69(3):625-635.
43. Koo BH, Park MY, Jeon OH, Kim DS.Regulatory mechanism of matrixmetalloprotease-2 enzymatic activity by factor
Xa and thrombin. J Biol Chem. 2009;284(35):23375-23385.
44. Liu Z, Li N, Diaz LA, Shipley M, Senior RM, WerbZ. Synergy between a plasminogen cascade andMMP-9 in autoimmune disease. J Clin Invest.2005;115(4):879-887.
45. Suzuki-Inoue K, Kato Y, Inoue O, et al. Involvementof the snake toxin receptor CLEC-2, in podoplanin-mediated platelet activation, by cancer cells. J BiolChem. 2007;282(36):25993-26001.
BLOOD, 4 DECEMBER 2014 x VOLUME 124, NUMBER 24 SIALYLATED O-GLYCANS CONTROL PODOPLANIN EXPRESSION 3665
For personal use only.on December 8, 2014. by guest www.bloodjournal.orgFrom

online October 21, 2014 originally publisheddoi:10.1182/blood-2014-04-572107
2014 124: 3656-3665
Dai, Hong Chen and Lijun XiaMehta-D'Souza, Xiaofeng Cai, Changgeng Ruan, Rodger P. McEver, Christopher West, Kesheng Yanfang Pan, Tadayuki Yago, Jianxin Fu, Brett Herzog, J. Michael McDaniel, Padmaja lymphatic endothelial cells and for interaction with platelets
-glycans for stable expression onOPodoplanin requires sialylated
http://www.bloodjournal.org/content/124/24/3656.full.htmlUpdated information and services can be found at:
(462 articles)Vascular Biology (507 articles)Platelets and Thrombopoiesis
Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society
For personal use only.on December 8, 2014. by guest www.bloodjournal.orgFrom