plastids are major regulators of light signaling in arabidopsis
TRANSCRIPT

Available online at www.sciencedirect.com
Integration of light and plastid signalsRobert M Larkin1,2 and Michael E Ruckle1,2
Light and plastid signals promote chloroplast biogenesis and
are among the most potent inducers and repressors of
photosynthesis-related gene expression, respectively. These
signals can be likened to a ‘gas and brake system’ that
promotes efficient chloroplast biogenesis and function. Recent
findings indicate that a particular plastid signal can ‘rewire’ a
light signaling network, converting it from an inducer into a
repressor of particular photosynthesis-related genes.
Therefore, a plastid signal appears to be an endogenous
regulator of light signaling rather than a signal acting
independently from light. This integration of light and plastid
signals may allow plants to proactively manage chloroplast
dysfunction when performing chloroplast biogenesis and
maintenance in adverse light conditions.
Addresses1 Michigan State University, Department of Energy Plant Research
Laboratory, United States2 Department of Biochemistry and Molecular Biology, Michigan State
University, East Lansing, MI 48824, United States
Corresponding author: Larkin, Robert M ([email protected]) and Ruckle,
Michael E ([email protected])
Current Opinion in Plant Biology 2008, 11:593–599
This review comes from a themed issue on
Cell Biology
Edited by David Ehrhardt and Federica Brandizzi
Available online 22nd October 2008
1369-5266/$ – see front matter
# 2008 Elsevier Ltd. All rights reserved.
DOI 10.1016/j.pbi.2008.10.004
IntroductionProteins that perform functions related to photosynthesis
are encoded by both nuclear and chloroplast genomes.
The chloroplast contains a small genome that encodes
less than 100 proteins, most of which contribute to
photosynthesis and the expression of the chloroplast
genome. By contrast, approximately 3000 nuclear genes
are predicted to encode chloroplast proteins in Arabidop-
sis and rice [1]. Coordinating the expression of photosyn-
thesis-associated nuclear genes (PhANGs) with the
expression of photosynthesis-associated plastidic genes
is essential for proper chloroplast biogenesis and main-
tenance, because photosynthesis depends on large multi-
subunit protein complexes that are composed of both
chloroplast-encoded and nuclear-encoded proteins.
Environmental and endogenous cues such as light, the
circadian clock, tissue-specific signals, carbohydrates, and
www.sciencedirect.com
hormones control the expression of nuclear genes that
encode chloroplast proteins [2–4], including those that
control the expression of the plastid genome [5,6]. There-
fore, the regulation of nuclear gene expression by these
signals has a major role in coordinating the expression of
both genomes and controlling chloroplast biogenesis and
function.
Although far from autonomous, chloroplasts are not com-
pletely subordinate to the nucleus. In fact, signals that are
triggered by plastid dysfunction are major regulators of
nuclear gene expression, especially of a number of genes
that encode proteins active in photosynthesis. Plastids
were first shown to affect the extraplastidic synthesis of
plastidic enzymes thirty years ago [7]. Subsequent studies
showed that plastid factors can repress the transcription of
nuclear genes and that this plastid-mediated repression
depends on particular cis-acting elements in the promo-
ters of nuclear genes. Up to several thousand genes can be
regulated by plastid dysfunction in Arabidopsis. Several
plastid signals have been identified, such as buildup of
the chlorophyll precursor Mg-protoporphyrin IX, signals
that are transduced when the expression of the plastid
genome is inhibited, signals derived from the photosyn-
thetic electron transport chain, and reactive oxygen
species such as singlet oxygen and hydrogen peroxide.
The molecular nature of signals that are derived from the
expression of the plastid genome and the photosynthetic
electron transport chain have not been defined. Many of
the proteins that are required for plastid signal biosyn-
thesis and transduction remain unknown. Also, although
plastid signals affect growth and development, their full
impact of plastid signals on growth and development has
not been completely established. These aspects of plas-
tid-to-nucleus signaling have been reviewed recently [8–12]. Here, we will review recent findings on the integ-
ration of light and plastid signals, topics not covered by
these other reviews.
Plastid signals and plastid-to-nucleussignaling mechanismsBlocking chloroplast biogenesis was one of the first
approaches for studying plastid-to-nucleus signaling.
Chloroplast biogenesis can be blocked using particular
genetic backgrounds (e.g., variegated mutants, plastid
ribosome-deficient, or carotenoid-deficient mutants).
Alternatively, chloroplast biogenesis can be blocked using
photobleaching herbicides or antibiotics that inhibit the
expression of the plastid genome. Seedlings treated with
these inhibitors of chloroplast biogenesis are viable when
they are provided sucrose, and become green when
transferred to media that does not contain one of these
Current Opinion in Plant Biology 2008, 11:593–599
594 Cell Biology
Figure 1
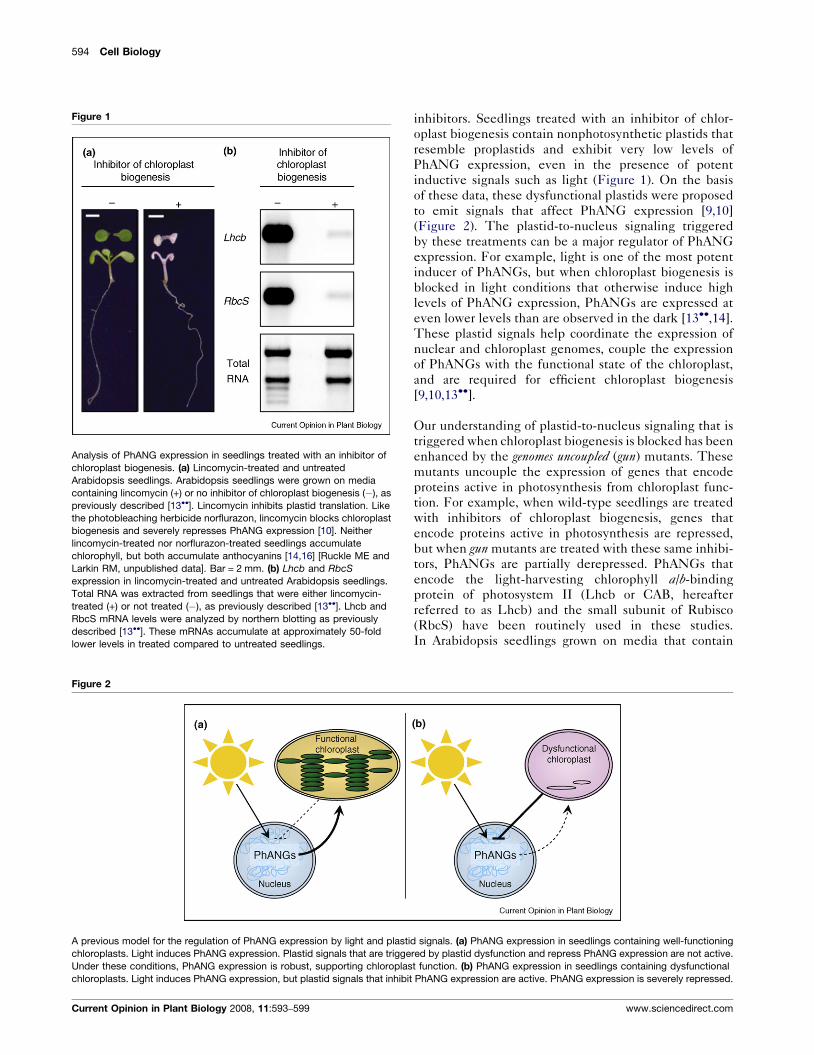
Analysis of PhANG expression in seedlings treated with an inhibitor of
chloroplast biogenesis. (a) Lincomycin-treated and untreated
Arabidopsis seedlings. Arabidopsis seedlings were grown on media
containing lincomycin (+) or no inhibitor of chloroplast biogenesis (�), as
previously described [13��]. Lincomycin inhibits plastid translation. Like
the photobleaching herbicide norflurazon, lincomycin blocks chloroplast
biogenesis and severely represses PhANG expression [10]. Neither
lincomycin-treated nor norflurazon-treated seedlings accumulate
chlorophyll, but both accumulate anthocyanins [14,16] [Ruckle ME and
Larkin RM, unpublished data]. Bar = 2 mm. (b) Lhcb and RbcS
expression in lincomycin-treated and untreated Arabidopsis seedlings.
Total RNA was extracted from seedlings that were either lincomycin-
treated (+) or not treated (�), as previously described [13��]. Lhcb and
RbcS mRNA levels were analyzed by northern blotting as previously
described [13��]. These mRNAs accumulate at approximately 50-fold
lower levels in treated compared to untreated seedlings.
Figure 2
A previous model for the regulation of PhANG expression by light and plasti
chloroplasts. Light induces PhANG expression. Plastid signals that are trigge
Under these conditions, PhANG expression is robust, supporting chloroplas
chloroplasts. Light induces PhANG expression, but plastid signals that inhibit
Current Opinion in Plant Biology 2008, 11:593–599
inhibitors. Seedlings treated with an inhibitor of chlor-
oplast biogenesis contain nonphotosynthetic plastids that
resemble proplastids and exhibit very low levels of
PhANG expression, even in the presence of potent
inductive signals such as light (Figure 1). On the basis
of these data, these dysfunctional plastids were proposed
to emit signals that affect PhANG expression [9,10]
(Figure 2). The plastid-to-nucleus signaling triggered
by these treatments can be a major regulator of PhANG
expression. For example, light is one of the most potent
inducer of PhANGs, but when chloroplast biogenesis is
blocked in light conditions that otherwise induce high
levels of PhANG expression, PhANGs are expressed at
even lower levels than are observed in the dark [13��,14].
These plastid signals help coordinate the expression of
nuclear and chloroplast genomes, couple the expression
of PhANGs with the functional state of the chloroplast,
and are required for efficient chloroplast biogenesis
[9,10,13��].
Our understanding of plastid-to-nucleus signaling that is
triggered when chloroplast biogenesis is blocked has been
enhanced by the genomes uncoupled (gun) mutants. These
mutants uncouple the expression of genes that encode
proteins active in photosynthesis from chloroplast func-
tion. For example, when wild-type seedlings are treated
with inhibitors of chloroplast biogenesis, genes that
encode proteins active in photosynthesis are repressed,
but when gun mutants are treated with these same inhibi-
tors, PhANGs are partially derepressed. PhANGs that
encode the light-harvesting chlorophyll a/b-binding
protein of photosystem II (Lhcb or CAB, hereafter
referred to as Lhcb) and the small subunit of Rubisco
(RbcS) have been routinely used in these studies.
In Arabidopsis seedlings grown on media that contain
d signals. (a) PhANG expression in seedlings containing well-functioning
red by plastid dysfunction and repress PhANG expression are not active.
t function. (b) PhANG expression in seedlings containing dysfunctional
PhANG expression are active. PhANG expression is severely repressed.
www.sciencedirect.com

Plastid-light interactions Larkin and Ruckle 595
Figure 3
The GUN1-dependent plastid signaling. When Arabidopsis seedlings are
treated with inhibitors of chloroplast biogenesis, the buildup of Mg-
protoporphyrin IX (Mg-Proto) or inhibition of plastid gene expression
(PGE) triggers the production of a second messenger (X) that requires
GUN1 for its biosynthesis or transduction (dashed arrows). High-
intensity light and glucose treatments can also induce the biosynthesis
or transduction of X. Light induces PhANG expression by inducing the
activity of G-box binding factors (GBFs) that induce PhANG
transcription, as described in the text. X induces the ABI4-dependent
repression of PhANGs. ABI4 has been proposed to repress PhANGs by
binding to CCAC and displacing GBFs, which bind the light-responsive
G-box (CACGTG). This model was adapted from Koussevitzky et al.
[18��].
various inhibitors of chloroplast biogenesis, Lhcb and
RbcS mRNAs accumulate at 2–3% of the levels observed
in untreated seedlings. By contrast, when strong gunalleles are treated with these same inhibitors, mRNAs
encoded by these PhANGs accumulate to 10–20% of the
levels observed in untreated seedlings [13��,15]. The
derepression of PhANG expression in gun mutants is
thought to result from defects in plastid-to-nucleus sig-
naling that repress PhANGs [9,10]. The coordination of
the nuclear and chloroplastic genomes is also impaired in
gun1 mutants [16].
Recent analysis of gun mutants indicates that accumu-
lation of the chlorophyll precursor Mg-protoporphyrin IX,
inhibition of chloroplast genome expression, high levels
of glucose, and exposure to high-intensity light all pro-
duce a second messenger that triggers plastid-to-nucleus
signaling that represses PhANG expression. This second
messenger remains undefined. GUN1 appears to be cru-
cial for the production or transduction of this second
messenger. GUN1 is a chloroplastic pentatricopeptide
repeat protein that colocalizes with nucleoids. Because
these proteins contribute to post-transcriptional processes
such as splicing, editing, processing, and translation of
RNA in chloroplasts and mitochondria [17], these findings
are consistent with the expression of the plastid genome
affecting nuclear gene expression when chloroplast bio-
genesis is blocked. ABSCISIC ACID-INSENSITIVE 4
(ABI4), an Apetala 2-type transcription factor, acts down-
stream of GUN1 and appears to repress PhANG tran-
scription by binding CCAC in PhANG promoters, a
sequence that is adjacent to or overlapping with the G-
box [18��]. Because G-boxes can contribute to the light
induction of PhANGs [2,19], these data suggest that ABI4
and a G-box binding factor(s) (GBF) might compete for a
binding site on PhANG promoters. However, not all
GUN1-regulated promoters have a CCAC motif overlap-
ping or in close proximity to a G-box [18��]. Moreover,
ABI4 has also been reported to have no effect on the
plastid regulation of reporter genes driven by particular
RbcS promoters [20]. These findings suggest that GUN1-
dependent plastid signals may regulate transcription fac-
tors other than ABI4. Nonetheless, GUN1 and ABI4
acting downstream of multiple plastid signals is consistent
with a model proposed several years ago in which a master
switch integrates diverse plastid signals [21]. These find-
ings also suggest that GUN1 and ABI4 function as part of
this master switch [18��] (Figure 3).
Regulation of PhANG expression by lightLight regulates at least 20% of the transcriptome in
Arabidopsis and rice. A number of photoreceptors and
downstream signaling components have been shown to
function in light-regulated transcriptional networks [2].
Photoreceptor signaling mechanisms are complex, invol-
ving the regulation of activity, subcellular localization,
and concentrations of particular photoreceptors and
www.sciencedirect.com
downstream signaling components [2]. The expression
of Lhcb, RbcS, and other PhANGs is induced by phyto-
chromes and cryptochromes, which sense far-red, red, and
blue light [19,22,23]. Much of this induction results from
increased rates of transcription. A combination of at least
two promoter elements such as G-boxes, GT-elements, I-
boxes or GATA motifs, AT-rich motifs, and others have
been shown to function as light-responsive promoter
elements in PhANGs [2,19].
Interaction between plastid signaling and lightsignalingNo light signaling mutants were obtained from the
original gun screen, with the exceptions of gun2 and
gun3, which are allelic to hy1 and hy2 [15]. These genes
encode heme oxygenase and phytochromobilin synthase
Current Opinion in Plant Biology 2008, 11:593–599
596 Cell Biology
and are required for the biosynthesis of the chromophore
for phytochromes [24]. The gun phenotypes of these
mutants are thought to arise from reduced levels of
Mg-protoporphyrin IX rather than as a result of defective
phytochrome signaling [15,25,26]. However, although
photoreceptors such as cry1 induce Lhcb expression in
greening and green seedlings [19,23], cry1 alleles were
recently isolated from a new gun mutant screen. These
data indicate that cry1 represses Lhcb when chloroplast
biogenesis is blocked. cry1 mutants that are treated with
inhibitors of chloroplast biogenesis accumulate approxi-
mately 5–10% of the Lhcb mRNA that accumulates in
wild-type seedlings that are not treated with inhibitors of
chloroplast biogenesis, compared to the 2–3% that
accumulates in treated wild type. This subtle gun phe-
notype understates the significance of cry1 in this context.
Double mutant studies show that Lhcb is synergistically
derepressed in cry1 gun1 double mutants and that cry1 and
GUN1 are responsible for most if not all of the repression
of Lhcb in blue light. These data indicate that a plastid
signal can convert cry1 into a negative regulator of Lhcband suggest that the fluence rate should control the
amount of Lhcb repression. Indeed, when seedlings are
treated with inhibitors of chloroplast biogenesis, the
repression of Lhcb is increased 10-fold in bright light
compared to dim light [13��].
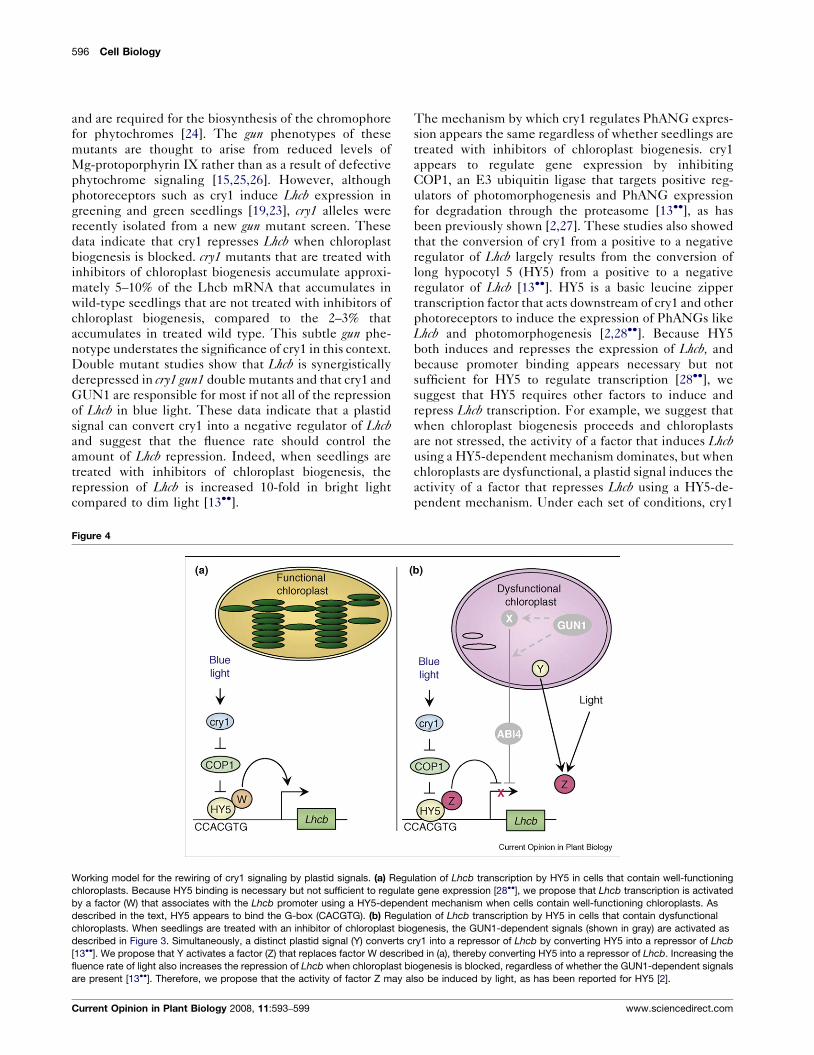
Figure 4
Working model for the rewiring of cry1 signaling by plastid signals. (a) Regu
chloroplasts. Because HY5 binding is necessary but not sufficient to regulate
by a factor (W) that associates with the Lhcb promoter using a HY5-depend
described in the text, HY5 appears to bind the G-box (CACGTG). (b) Regul
chloroplasts. When seedlings are treated with an inhibitor of chloroplast bio
described in Figure 3. Simultaneously, a distinct plastid signal (Y) converts c
[13��]. We propose that Y activates a factor (Z) that replaces factor W describ
fluence rate of light also increases the repression of Lhcb when chloroplast b
are present [13��]. Therefore, we propose that the activity of factor Z may a
Current Opinion in Plant Biology 2008, 11:593–599
The mechanism by which cry1 regulates PhANG expres-
sion appears the same regardless of whether seedlings are
treated with inhibitors of chloroplast biogenesis. cry1
appears to regulate gene expression by inhibiting
COP1, an E3 ubiquitin ligase that targets positive reg-
ulators of photomorphogenesis and PhANG expression
for degradation through the proteasome [13��], as has
been previously shown [2,27]. These studies also showed
that the conversion of cry1 from a positive to a negative
regulator of Lhcb largely results from the conversion of
long hypocotyl 5 (HY5) from a positive to a negative
regulator of Lhcb [13��]. HY5 is a basic leucine zipper
transcription factor that acts downstream of cry1 and other
photoreceptors to induce the expression of PhANGs like
Lhcb and photomorphogenesis [2,28��]. Because HY5
both induces and represses the expression of Lhcb, and
because promoter binding appears necessary but not
sufficient for HY5 to regulate transcription [28��], we
suggest that HY5 requires other factors to induce and
repress Lhcb transcription. For example, we suggest that
when chloroplast biogenesis proceeds and chloroplasts
are not stressed, the activity of a factor that induces Lhcbusing a HY5-dependent mechanism dominates, but when
chloroplasts are dysfunctional, a plastid signal induces the
activity of a factor that represses Lhcb using a HY5-de-
pendent mechanism. Under each set of conditions, cry1
lation of Lhcb transcription by HY5 in cells that contain well-functioning
gene expression [28��], we propose that Lhcb transcription is activated
ent mechanism when cells contain well-functioning chloroplasts. As
ation of Lhcb transcription by HY5 in cells that contain dysfunctional
genesis, the GUN1-dependent signals (shown in gray) are activated as
ry1 into a repressor of Lhcb by converting HY5 into a repressor of Lhcb
ed in (a), thereby converting HY5 into a repressor of Lhcb. Increasing the
iogenesis is blocked, regardless of whether the GUN1-dependent signals
lso be induced by light, as has been reported for HY5 [2].
www.sciencedirect.com

Plastid-light interactions Larkin and Ruckle 597
Figure 5
Interaction between plastid signaling and light signaling that affects Lhcb
and RbcS expression. (a) Regulation of Lhcb and RbcS expression in
seedlings containing chloroplasts that are not stressed. During de-
etiolation or in green seedlings photoreceptors that perceive red and
blue light induce the expression of Lhcb and RbcS, as described in the
text. Chloroplasts that are not stressed have not been shown to
influence the regulation of these PhANGs by affecting light-regulated
transcriptional networks. (b) The rewiring of light signaling networks by
plastid signals. When chloroplast biogenesis is blocked, a plastid signal
(X) triggers the GUN1-dependent signaling, as described in Figure 3
(shown in gray). Under these same conditions, a distinct plastid signal(s)
(Y) converts the blue and/or red light signaling network, which have
otherwise been reported to induce both Lhcb and RbcS, into repressors
of Lhcb and RbcS. Plastid–phytochrome interactions are not as well
understood as plastid–cry1 interactions. The repression of RbcS by Y
may result from a light-dependent or a light-independent signal (dotted
arrow and dotted T-bar, respectively). Y may be more than one signal.
Subpart (b) was adapted from Ruckle et al. [13��].
promotes the activity of the regulatory complex by bind-
ing and inhibiting COP1, thereby protecting HY5 from
degradation (Figure 4). The conversion of particular
transcription factors from positive to negative regulators
and vice versa has been reported in other systems. For
example, nuclear receptors and Myc are known to either
positively or negatively regulate transcription, depending
on the coregulators or transcription factors they bind [29–31]. Other mechanisms are possible.
The findings of Koussevitzky et al. [18��] and Ruckle et al.[13��] indicate that ABI4 and HY5 are simultaneously
required for plastid-dependent repression of Lhcb. There-
fore, a model that is more complex than ABI4 displacing
GBFs such as HY5 [32,33] from Lhcb promoters seems
likely. These findings lead us to suggest that either both
factors simultaneously bind Lhcb promoters when chlor-
oplast biogenesis is blocked or that at least one of these
factors represses Lhcb by an indirect mechanism.
Although HY5 has been shown to bind Lhcb promoters
in light-grown and etiolated tissue by chromatin immu-
noprecipitation [28��], neither ABI4 nor HY5 has been
tested for binding Lhcb or other PhANG promoters in
seedlings treated with inhibitors of chloroplast bio-
genesis.
Several observations suggest that plastid–light inter-
actions are probably more complex than described above.
For example, although cry1 and GUN1 are responsible for
most if not all repression of Lhcb when chloroplast bio-
genesis is blocked in blue light, repression in white light
appears more complex. This additional complexity may
be contributed by other photoreceptors. Indeed, phyB
contributes to the repression of Lhcb when chloroplast
biogenesis is blocked in white light, but whether all
repression of Lhcb in white light depends on GUN1,
cry1, and phyB remains to be tested. The complexity
of plastid–light interactions is also apparent from an
analysis of RbcS expression. Although Lhcb and RbcSare similarly repressed when Arabidopsis seedlings are
treated with inhibitors of chloroplast biogenesis in white
light, cry1 remains a positive regulator of RbcS regardless
of the functional and developmental state of the chlor-
oplast. Analysis of RbcS mRNA levels in seedlings trea-
ted with inhibitors of chloroplast biogenesis and
untreated seedlings in different light qualities are con-
sistent with (1) a light quality besides blue or (2) a light-
independent signal that is distinct from GUN1-depend-
ent signals repressing RbcS when chloroplast biogenesis is
blocked [13��] (Figure 5).
Interactions between plastid and light signals are not
surprising. Because light promotes chloroplast biogenesis
and photosynthesis, light also induces chloroplast stress
[34]. Although plastid signals such as the light-indepen-
dent signals that require GUN1 allow plants to react to
chloroplast stress, plants may need to do more than simply
www.sciencedirect.com
react. By ‘rewiring’ light signaling, plastid signals could
enable plants to be proactive rather than simply reactive,
and to respond not only to chloroplast dysfunction but
also to the potential for continued photooxidative damage
when regulating nuclear gene expression. Consistent with
these plastid–light interactions contributing to photooxi-
dative stress tolerance, cry1 and hy5 single mutants are more
sensitive to high-intensity light than wild type [13��,35].
Additionally, a striking increase in the sensitivity to
Current Opinion in Plant Biology 2008, 11:593–599

598 Cell Biology
high-intensity light was observed in the cry1 gun1 and hy5gun1 double mutants compared to the corresponding single
mutants and wild type [13��]. Another example of plastid–light interactions that is consistent with this model was
recently reported by Danon et al. [36��]. These authors
showed that singlet oxygen stress within the chloroplast
requires cry1 to regulate nuclear gene expression and
trigger a programmed cell death response.
ConclusionsAt least two genetically distinct plastid signals have been
defined that are triggered by treatments with inhibitors of
chloroplast biogenesis: the signal that remodels a light
signaling network and the light-independent plastid sig-
nals that depend on GUN1. Lhcb mRNA has been
reported to accumulate at similar levels in particular
gun mutants treated with inhibitors of chloroplast bio-
genesis and untreated wild-type seedlings only in blue or
far-red light and not when seedlings are grown in dark-
ness, white light, red light, or in any light condition that
induces RbcS mRNA [13��]. These findings suggest that
we are unable to completely knock out plastid-to-nucleus
signaling and may not know the full impact of plastid-to-
nucleus signaling during growth and development of
plants at this time. We do know that the GUN1-depend-
ent signals and plastid signals that remodel light signaling
networks regulate nuclear gene expression and promote
efficient chloroplast biogenesis [9,10,13��]. Recent
analyses of gun1 cry1 double mutants indicate that plastid
signals can also affect the accumulation of anthocyanins
and the photomorphogenesis of cotyledons and hypoco-
tyls (Ruckle ME and Larkin RM, unpublished data).
Additionally, plastid signals were recently shown to affect
the circadian clock [37�]. These recent findings suggest
that the significance of signals emanating from the plastid
may be underappreciated, not only in regulating chlor-
oplast biogenesis and function but also in other processes.
AcknowledgementsThis work was supported by DOE grant no. DE–FG02–91ER20021 andNSF grant no. IOB 0517841 to RML.
References and recommended readingPapers of particular interest, published within the period of review,have been highlighted as:
� of special interest
�� of outstanding interest
1. Richly E, Leister D: An improved prediction of chloroplastproteins reveals diversities and commonalities in thechloroplast proteomes of Arabidopsis and rice. Gene 2004,329:11-16.
2. Jiao Y, Lau OS, Deng XW: Light-regulated transcriptionalnetworks in higher plants. Nat Rev Genet 2007, 8:217-230.
3. Terzaghi WB, Cashmore AR: Light-regulated transcription. AnnuRev Plant Physiol Plant Mol Biol 1995, 46:445-474.
4. Rook F, Hadingham SA, Li Y, Bevan MW: Sugar and ABAresponse pathways and the control of gene expression. PlantCell Environ 2006, 29:426-434.
Current Opinion in Plant Biology 2008, 11:593–599
5. Marın-Navarro J, Manuell AL, Wu J, Mayfield SP: Chloroplasttranslation regulation. Photosynth Res 2007, 94:359-374.
6. Toyoshima Y, Onda Y, Shiina T, Nakahira Y: Plastid transcriptionin higher plants. Crit Rev Plant Sci 2005, 24:59-81.
7. Bradbeer JW, Atkinson YA, Borner T, Hagemann R: Cytoplasmicsynthesis of plastid polypeptide may be controlled by plastid-synthesized RNA. Nature 1979, 279:816-817.
8. Yu F, Fu A, Aluru M, Park S, Xu Y, Liu H, Liu X, Foudree A,Nambogga M, Rodermel S: Variegation mutants andmechanisms of chloroplast biogenesis. Plant Cell Environ 2007,30:350-365.
9. Woodson JD, Chory J: Coordination of gene expressionbetween organellar and nuclear genomes. Nat Rev Genet 2008,9:383-395.
10. Nott A, Jung HS, Koussevitzky S, Chory J: Plastid-to-nucleusretrograde signaling. Annu Rev Plant Biol 2006, 57:739-759.
11. Pesaresi P, Schneider A, Kleine T, Leister D: Interorganellarcommunication. Curr Opin Plant Biol 2007, 10:600-606.
12. Dietzel L, Brautigam K, Pfannschmidt T: Photosyntheticacclimation: state transitions and adjustment of photosystemstoichiometry—functional relationships between short-termand long-term light quality acclimation in plants. FEBS J 2008,275:1080-1088.
13.��
Ruckle ME, DeMarco SM, Larkin RM: Plastid signals remodellight signaling networks and are essential for efficientchloroplast biogenesis in Arabidopsis. Plant Cell 2007,19:3944-3960.
These authors demonstrate that plastid signals can change the responseof photosynthesis-related gene expression to both light quality andquantity and that these interactions contribute to efficient chloroplastbiogenesis.
14. Sullivan JA, Gray JC: Plastid translation is required for theexpression of nuclear photosynthesis genes in the dark and inroots of the pea lip1 mutant. Plant Cell 1999, 11:901-910.
15. Mochizuki N, Brusslan JA, Larkin R, Nagatani A, Chory J:Arabidopsis genomes uncoupled 5 (GUN5) mutant reveals theinvolvement of Mg-chelatase H subunit in plastid-to-nucleussignal transduction. Proc Natl Acad Sci U S A 2001,98:2053-2058.
16. Susek RE, Ausubel FM, Chory J: Signal transduction mutants ofArabidopsis uncouple nuclear CAB and RBCS geneexpression from chloroplast development. Cell 1993,74:787-799.
17. Delannoy E, Stanley WA, Bond CS, Small ID: Pentatricopeptiderepeat (PPR) proteins as sequence-specificity factors in post-transcriptional processes in organelles. Biochem Soc Trans2007, 35:1643-1647.
18.��
Koussevitzky S, Nott A, Mockler TC, Hong F, Sachetto-Martins G,Surpin M, Lim J, Mittler R, Chory J: Signals from chloroplastsconverge to regulate nuclear gene expression. Science 2007,316:715-719.
These authors cloned GUN1, which encodes a chloroplast-localized pen-tatricopeptide repeat protein. They also showed that GUN1 and ABI4, anuclear-localized Apetala 2-type transcription factor, act downstream ofmultiple plastid signals that regulate the expression of nuclear genes.
19. Tyagi AK, Gaur T: Light regulation of nuclear photosyntheticgenes in higher plants. Crit Rev Plant Sci 2003, 22:417-452.
20. Acevedo-Hernandez GJ, Leon P, Herrera-Estrella LR: Sugar andABA responsiveness of a minimal RBCS light-responsive unitis mediated by direct binding of ABI4. Plant J 2005, 43:506-519.
21. Richly E, Dietzmann A, Biehl A, Kurth J, Laloi C, Apel K, Salamini F,Leister D: Covariations in the nuclear chloroplasttranscriptome reveal a regulatory master-switch. EMBO Rep2003, 4:491-498.
22. Reed JW, Nagatani A, Elich TD, Fagan M, Chory J: PhytochromeA and phytochrome B have overlapping but distinct functionsin Arabidopsis development. Plant Physiol 1994, 104:1139-1149.
23. Mazzella MA, Cerdan PD, Staneloni RJ, Casal JJ: Hierarchicalcoupling of phytochromes and cryptochromes reconciles
www.sciencedirect.com

Plastid-light interactions Larkin and Ruckle 599
stability and light modulation of Arabidopsis development.Development 2001, 128:2291-2299.
24. Terry MJ, Linley PJ, Kohchi T: Making light of it: the role of planthaem oxygenases in phytochrome chromophore synthesis.Biochem Soc Trans 2002, 30:604-609.
25. Vinti G, Hills A, Campbell S, Bowyer JR, Mochizuki N, Chory J,Lopez-Juez E: Interactions between hy1 and gun mutants ofArabidopsis, and their implications for plastid/nuclearsignaling. Plant J 2000, 24:883-894.
26. Strand A, Asami T, Alonso J, Ecker JR, Chory J: Chloroplast tonucleus communication triggered by accumulation of Mg-protoporphyrin IX. Nature 2003, 421:79-83.
27. Yi C, Deng XW: COP1—from plant photomorphogenesis tomammalian tumorigenesis. Trends Cell Biol 2005, 15:618-625.
28.��
Lee J, He K, Stolc V, Lee H, Figueroa P, Gao Y, Tongprasit W,Zhao H, Lee I, Deng XW: Analysis of transcription factor HY5genomic binding sites revealed its hierarchical role in lightregulation of development. Plant Cell 2007, 19:731-749.
Using chromatin immunoprecipitation, these authors demonstrate thatHY5 binds more than 3000 sites in the Arabidopsis genome and thatphotosynthesis-related genes are overrepresented among genes thatHY5 binds. These authors also show that HY5 contributes to bothinduction and repression of distinct nuclear genes and that HY5 bindingto a gene is not sufficient to regulate transcription.
29. Wanzel M, Herold S, Eilers M: Transcriptional repression byMyc. Trends Cell Biol 2003, 13:146-150.
30. Feige JN, Auwerx J: Transcriptional coregulators in the controlof energy homeostasis. Trends Cell Biol 2007, 17:292-301.
31. Kassel O, Herrlich P: Crosstalk between the glucocorticoidreceptor and other transcription factors: molecular aspects.Mol Cell Endocrinol 2007, 275:13-29.
www.sciencedirect.com
32. Chattopadhyay S, Ang LH, Puente P, Deng XW, Wei N:Arabidopsis bZIP protein HY5 directly interactswith light-responsive promoters in mediating lightcontrol of gene expression. Plant Cell 1998,10:673-683.
33. Ang LH, Chattopadhyay S, Wei N, Oyama T, Okada K,Batschauer A, Deng XW: Molecular interaction betweenCOP1 and HY5 defines a regulatory switch for lightcontrol of Arabidopsis development. Mol Cell 1998,1:213-222.
34. Niyogi KK: Photoprotection revisited: genetic and molecularapproaches. Annu Rev Plant Physiol Plant Mol Biol 1999,50:333-359.
35. Kleine T, Kindgren P, Benedict C, Hendrickson L, Strand A:Genome-wide gene expression analysis reveals a critical rolefor CRYPTOCHROME1 in the response of Arabidopsis to highirradiance. Plant Physiol 2007, 144:1391-1406.
36.��
Danon A, Coll NS, Apel K: Cryptochrome-1-dependentexecution of programmed cell death induced by singletoxygen in Arabidopsis thaliana. Proc Natl Acad Sci U S A 2006,103:17036-17041.
These authors show that a plastid-to-nucleus signaling pathway that istriggered by singlet oxygen stress within the chloroplast requires photo-activated cry1 to induce programmed cell death.
37.�
Hassidim M, Yakir E, Fradkin D, Hilman D, Kron I, Keren N, Harir Y,Yerushalmi S, Green RM: Mutations in chloroplast RNA bindingprovide evidence for the involvement of the chloroplast in theregulation of the circadian clock in Arabidopsis. Plant J 2007,51:551-562.
These authors demonstrate that a chloroplast-localized RNA-bindingprotein can affect the circadian clock. They also demonstrate thatGUN1 and STN7, which were previously shown to participate in plas-tid-to-nucleus signaling, can also affect the circadian clock.
Current Opinion in Plant Biology 2008, 11:593–599