phytoliths from archaeological sites in the tropical forest of ituri, democratic republic of congo
TRANSCRIPT

elgium
gwctHtPtb
Quaternary Research54, 102–112 (2000)doi:10.1006/qres.2000.2150, available online at http://www.idealibrary.com on
Phytoliths from Archaeological Sites in the Tropical Forest of Ituri,Democratic Republic of Congo
Julio Mercader
Department of Anthropology, George Washington University, 2110 G Street, NW, Washington, DC 20052
E-mail: [email protected]
Freya Runge
Universitat Paderborn, FB1, Physische Geographie, D-33098, Paderborn, Germany
E-mail: [email protected]
Luc Vrydaghs
Laboratory of Palaeontology, Gent University, Krigslaan, 281/S8, B-9000; and Lab of Wood Biology and Xylarium, RMCA, B-3080, Tervuren, B
E-mail: [email protected]
Hughes Doutrelepont
Lab of Wood Biology and Xylarium, RMCA, B-3080, Tervuren, Belgium
Corneille E. N. Ewango
Cefrecof, P.O. Box 21285, Nairobi, Kenya; and Epulu via Bunia, Democratic Republic of Congo
and
Jordi Juan-Tresseras
Departament de Prehisto`ria, Historia Antiga i Arqueologia, Facultat de Geografia i Histo`ria, Universitat de Barcelona, Baldiri i Reixac,s/n Torre B, p. 11, E-08028, Barcelona, Spain
E-mail: [email protected]
Received November 17, 1999
low-ionalrhave
ctionsedm-s not
8),tane. Sea-s of
andsig-
Phytoliths record late Quaternary vegetation at three archaeo-logical sites in the Ituri rain forest. The oldest deposits, dated to ca.19,000 to 10,000 14C yr B.P., contain abundant phytoliths ofrasses but also enough arboreal forms to show that the landscapeas forested. The late-glacial forests may have had a more open
anopy than today’s. Younger phytolith assemblages show thathe northeast Congo basin was densely forested throughout theolocene. Archaeological materials among the phytoliths show
hat people lived in this region during the Pleistocene. Therefore,leistocene and Holocene prehistoric foragers probably inhabited
ropical forests of the northeast Congo basin many millenniaefore farming appeared in the region. © 2000 University of Washington.
Key Words: Ituri; Democratic Republic Congo; central Africa;phytoliths; Later Stone Age foragers; last glacial maximum;vegetation dynamics; “refugia.”
0033-5894/00 $35.00Copyright © 2000 by the University of Washington.All rights of reproduction in any form reserved.
102
INTRODUCTION
Paleoecological data and climate models indicate thatland tropical forests underwent large climatic and vegetatchanges during the Pleistocene (Kutzbachet al.,1998; Van deKaars and Dam, 1995). However, such changes appear tobeen more complex than the simple cycles of forest contraand expansion resulting from climatic aridity that are propoby refugial theory (Haffer, 1969; Hamilton, 1972). For exaple, some lowland forests contained species associationseen today (Colinvauxet al.,1996; Maley and Brenac, 199such as admixtures of evergreen lowland taxa with monelements and trees characteristic of seasonal formationssonal forests, for their part, were replaced by mixturesavanna and thorny scrub vegetation (Van der HammenAbsy, 1994). These changes were brought about by both

edtaialthuco
reon
t,sisSelex
orarimcooom
rviv
onla
fore
thite,ionhilnthesit
uatheesthemorere%e fa(H
Ham
t ii),ii
( -as eousf onalc
82)s Rift.G of thef rs ofg chh eithern s 10t withv abit-a s.
liths
en-,was
elterto 2last
hadersenniace of
manakenial
arsed tosent
3tolithWThelysisnuru
uralLater
i Turuthisthis
PHYTOLITHS IN THE CONGO 103
nificantly cooler and drier climates and, perhaps, by markreduced CO2 (Jolly et al., 1998). Plant physiological adap-tions to low CO2 (Cowling, 1999) suggest that late glacmaximum tree taxa would produce smaller leaves and,promote tropical forests with a more open canopy andcomitant increase in grasses.
This paper applies phytolith analysis to the forested cothe African continent using phytoliths from three Later StAge archaeological sites (.18,800 6 100 14C yr B.P. to,715 6 45 14C yr B.P.) located in the Ituri rain foresDemocratic Republic of Congo. In Africa, phytolith analyhas been used for paleoenvironmental reconstruction ingal, People’s Republic of Congo, Kenya, and Ethiopia (Aandre et al., 1997; Mworia-Maitima, 1997; Barboniet al.,1999). These studies show that phytolith data from lake cand archaeological sites record both regional and local vtions in climate and vegetation in the grassland belt. The athis paper is to show the various types of environmentaltexts in which ancientHomo sapienscolonized the CongBasin during the late Pleistocene and the Holocene. Sauthors have questioned whether humans could have suin tropical forests without engaging in agriculture (Baileyetal., 1989). Our work suggests that the northeastern CBasin was forested when it was settled by humans in thePleistocene and that the region remained settled while itschanged in structure and composition thereafter.
SETTING
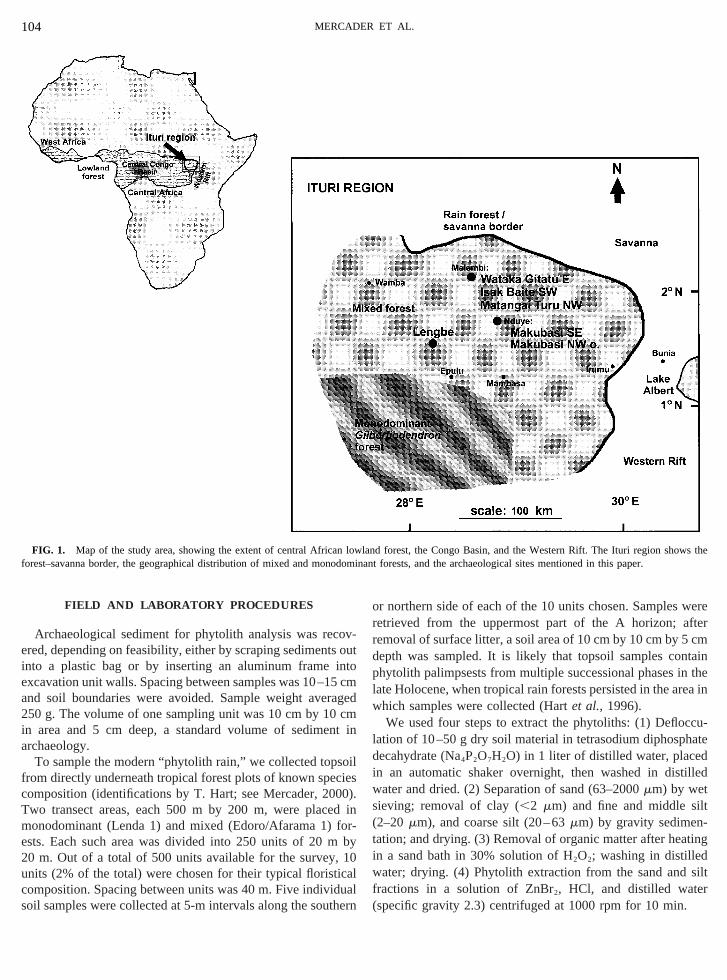
The Ituri region is located in the northeast corner ofGuineo-Congolian tropical forest of central Africa (Wh1983), just north of the equator (Fig. 1). In the Ituri regbetween the Western Rift and the Congo Basin, aregranitic uplands of the north and east, and plains of the cesouth, and west. Altitudes range from 1000 m for the higuplands in the east to 600 m for the plains to the west. Thestudied are 700–800 m above sea level.
Although little rain falls in January and February, an annrainfall of 1600–1900 mm (Bultot, 1971) allows most ofIturi district to support evergreen and semievergreen forSome of the forests are dominated by one taxon, while oare mixed (Hart, 1985). In the monodominant forests, comin the south and west,Gilbertiodendron dewevreiaccounts fomore than 90% of the canopy. In mixed forests, scattthrough the region, no single species forms more than 40the canopy. In the Ituri district, monodominant forests havfewer species per hectare (10) than do mixed forests (53)and Hart, 1986). Evergreen species make up 94%monodominant forests and 86% in mixed environments (and Hart, personal communication, 1995). The most comspecies of trees in the lowland rain forest of Ituri areGilber-iodendron dewevrei(Caesalpiniaceae),Cynometra alexandr(Caesalpiniaceae),Julbernardia seretii (CaesalpiniaceaeKlainedoxa gabonesis(Irvingiaceae),Cleistanthus michelson
ly
s,n-
ofe
ne--
esia-ofn-
eed
gotest
e
,lyer,stes
l
ts.rsn
dofr
artinrt
on
Euphorbiaceae), Erythrophleum suaveolens(Caesalpiniceae), Fagara macrophylla (Rutaceae), andCanariumchweinfurthii (Burseraceae). The most common herbacamily is the Marantaceae (T. Hart and C. Ewango, persommunication).The Ituri forest occupies a granitic domain (Lavreau, 19
haped in part by neotectonics from the neighboringrabens, inselbergs, and tors are common in that section
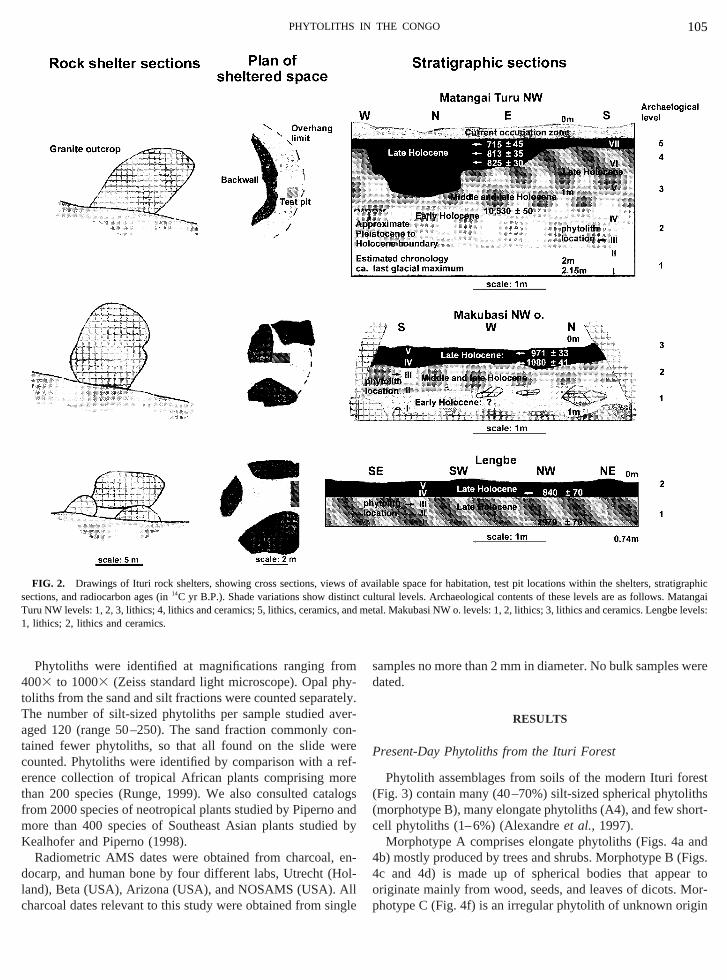
orest nearest the Rift, as are boulders and rock shelteranitoid rocks (Fig. 2) (Gonza´lez, 1997). The shelters, whiave attracted repeated human occupation, developedear inselberg peaks or on hill slopes. Located in outcrop
o 15 m, the caves have their mouths randomly oriented,ariable light and temperature conditions and enclosed hble spaces (average: 51.9 m2), including dry and wet area
The geochemistry of the cave deposits in which phytowere found is described by Mercader (2000).
ARCHAEOLOGY AND CHRONOLOGY
Archaeological surveys in the lowland forest of Ituri idtified over 50 archaeological sites in just 8.5 km2 (Mercader1997). This density of sites suggests that the Ituri forestsettled extensively during prehistoric times. Ten rock shsites were excavated, over a period of 12 months, with upm2 excavated per shelter. The results indicate that by theglacial maximum prehistoric Later Stone Age foragerssettled tropical forest environments. Occupation of divhabitats continued throughout the Holocene, many millebefore the first archaeological indications of the appearanfarming in the region (Mercaderet al., 2000).
Radiometric AMS dating (Table 1) suggests that huoccupation in this sector of the Congo Basin had already tplace by 18,8006 100 14C yr B.P., before the last glacmaximum (LGM). Additional ages (10,5306 50 14C yr B.P.;10,0156 55 14C yr B.P.) show that approximately 8000 yelater, by the start of the Holocene, forager groups continusettle the Ituri region. Hunter-gatherer groups were still prein the area in the middle Holocene (60256 70 14C yr B.P.;29706 70 14C yr B.P.) and in the late Holocene (10806 4114C yr B.P.; 9716 33 14C yr B.P.; 8406 70 14C yr B.P.; 82560 14C yr B.P.; 8136 35 14C yr B.P.; 7156 45 14C yr B.P.).Among ten excavated sites, three were selected for phy
analysis: Matangai Turu NW (northern Ituri), Makubasi N(eastern Ituri), and Lengbe (central Ituri) (Figs. 1 and 2).oldest direct date available in a site for which phytolith anawas carried out is 10,5306 50 14C yr B.P., obtained ocharcoal from a depth of 1.25 m from the site of Matangai TNW. Because this layer is underlain by 0.9 m of cultdeposits that contain chipped stone representative of theStone Age, and because major changes in the MatangaNW paleoenvironmental assemblages occur just belowculturally dated stratum, we infer that the basal layers ofsite are no younger than the LGM.
ovts ointo5agcm
nt
soicie00edfor
, 1icaduthe
wereftercmtain
n theea in
ccu-hatedilled
s ilt-ting
siltr
shows thef
MERCADER ET AL.104
FIELD AND LABORATORY PROCEDURES
Archaeological sediment for phytolith analysis was recered, depending on feasibility, either by scraping sedimeninto a plastic bag or by inserting an aluminum frameexcavation unit walls. Spacing between samples was 10–1and soil boundaries were avoided. Sample weight aver250 g. The volume of one sampling unit was 10 cm by 10in area and 5 cm deep, a standard volume of sedimearchaeology.
To sample the modern “phytolith rain,” we collected topfrom directly underneath tropical forest plots of known specomposition (identifications by T. Hart; see Mercader, 20Two transect areas, each 500 m by 200 m, were placmonodominant (Lenda 1) and mixed (Edoro/Afarama 1)ests. Each such area was divided into 250 units of 20 m20 m. Out of a total of 500 units available for the surveyunits (2% of the total) were chosen for their typical floristcomposition. Spacing between units was 40 m. Five indivisoil samples were collected at 5-m intervals along the sou
FIG. 1. Map of the study area, showing the extent of central Africaorest–savanna border, the geographical distribution of mixed and mon
-ut
cmed
in
ls).in-
by0lalrn
or northern side of each of the 10 units chosen. Samplesretrieved from the uppermost part of the A horizon; aremoval of surface litter, a soil area of 10 cm by 10 cm by 5depth was sampled. It is likely that topsoil samples conphytolith palimpsests from multiple successional phases ilate Holocene, when tropical rain forests persisted in the arwhich samples were collected (Hartet al., 1996).
We used four steps to extract the phytoliths: (1) Deflolation of 10–50 g dry soil material in tetrasodium diphospdecahydrate (Na4P2O7H2O) in 1 liter of distilled water, placein an automatic shaker overnight, then washed in distwater and dried. (2) Separation of sand (63–2000mm) by wetieving; removal of clay (,2 mm) and fine and middle s
(2–20 mm), and coarse silt (20–63mm) by gravity sedimentation; and drying. (3) Removal of organic matter after heain a sand bath in 30% solution of H2O2; washing in distilledwater; drying. (4) Phytolith extraction from the sand andfractions in a solution of ZnBr2, HCl, and distilled wate(specific gravity 2.3) centrifuged at 1000 rpm for 10 min.
wland forest, the Congo Basin, and the Western Rift. The Ituri regionminant forests, and the archaeological sites mentioned in this paper.
n loodo
omhytevecoerreoretalo
ad
enHoll
ing
were
restlithsort-
andFigs.ar to
or-gin
ratigras Matangai
:
PHYTOLITHS IN THE CONGO 105
Phytoliths were identified at magnifications ranging fr4003 to 10003 (Zeiss standard light microscope). Opal ptoliths from the sand and silt fractions were counted separaThe number of silt-sized phytoliths per sample studied aaged 120 (range 50–250). The sand fraction commonlytained fewer phytoliths, so that all found on the slide wcounted. Phytoliths were identified by comparison with aerence collection of tropical African plants comprising mthan 200 species (Runge, 1999). We also consulted cafrom 2000 species of neotropical plants studied by Pipernomore than 400 species of Southeast Asian plants studieKealhofer and Piperno (1998).
Radiometric AMS dates were obtained from charcoal,docarp, and human bone by four different labs, Utrecht (land), Beta (USA), Arizona (USA), and NOSAMS (USA). Acharcoal dates relevant to this study were obtained from s
FIG. 2. Drawings of Ituri rock shelters, showing cross sections, viewections, and radiocarbon ages (in14C yr B.P.). Shade variations show dist
Turu NW levels: 1, 2, 3, lithics; 4, lithics and ceramics; 5, lithics, ceramics1, lithics; 2, lithics and ceramics.
-ly.r-n-ef-
gsndby
-l-
le
samples no more than 2 mm in diameter. No bulk samplesdated.
RESULTS
Present-Day Phytoliths from the Ituri Forest
Phytolith assemblages from soils of the modern Ituri fo(Fig. 3) contain many (40–70%) silt-sized spherical phyto(morphotype B), many elongate phytoliths (A4), and few shcell phytoliths (1–6%) (Alexandreet al., 1997).
Morphotype A comprises elongate phytoliths (Figs. 4a4b) mostly produced by trees and shrubs. Morphotype B (4c and 4d) is made up of spherical bodies that appeoriginate mainly from wood, seeds, and leaves of dicots. Mphotype C (Fig. 4f) is an irregular phytolith of unknown ori
f available space for habitation, test pit locations within the shelters, stphiccultural levels. Archaeological contents of these levels are as follows.d metal. Makubasi NW o. levels: 1, 2, lithics; 3, lithics and ceramics. Lengbe levels
s oinct, an

lithderre-anmo
noocoisehe
in
sstr,) inmuhs
thoryla
zeThlagrb
s.
gbem
M lith
itionromisap-the top
last
t useuruughes in
phy-
Mer-lants
s, treetentlye, andted atntlyrestion’sare
rica
lithsionma-sses,hes
Makubasi
MERCADER ET AL.106
with a distinctive, stippled surface. This stippled phytois represented in very high frequencies (62%) in momonodominantGilbertiodendronforests and has lower repsentation (5%) in mixed forests. Morphotype D (Figs. 4g4h), made up of specific hexagonal and hat bodies, iscommon in mixed than in monodominant forests.
Morphotypes E and F (Figs. 4j, 4k, and 4l) are not diagtic, except for F4. These can be produced by both monand dicots. Morphotype G2, subtype 1 (Fig. 4i), comprthe distinctive bulliform cell phytolith produced only in tPoaceae. These and other Poaceae phytoliths are scarcemodern profiles from the Ituri forest.
Ancient Phytoliths from Archaeological Sites ofthe Ituri Region
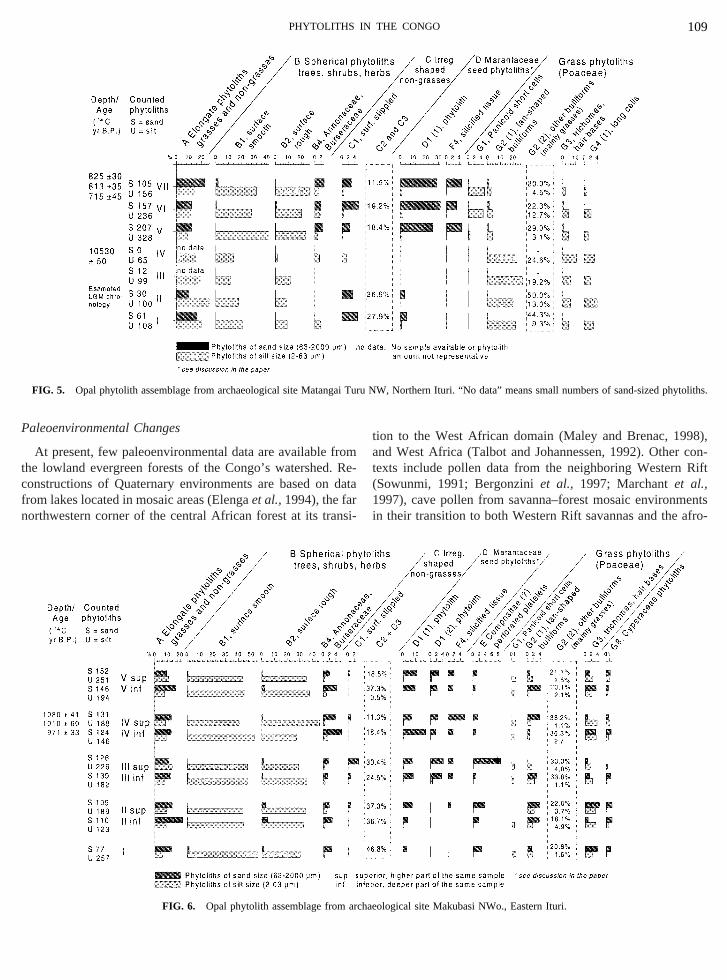
Matangai Turu NW (Malembi). Phytoliths from grasse(e.g., Poaceae, morphotype G2-1) are most common indeposited before 10,53014C yr B.P. (Fig. 5). In contrastphytoliths from rain forest trees and herbs (category D1crease greatly during the Holocene, reaching their maxivalues in the last 200014C yr B.P. Some types of phytolitderived from trees (category B) are most abundant inHolocene but are present throughout the sequence. Categanother arboreal indicator, is common during both thePleistocene and the Holocene.
Makubasi NW (Nduye).Arboreal phytoliths characterithis sequence, which may be entirely Holocene (Fig. 6).sequence lacks major variations in the phytolith assembThe morphotypes include arboreal (B, C, and D) and heceous (e.g., D1) plants typical of rain forest environment
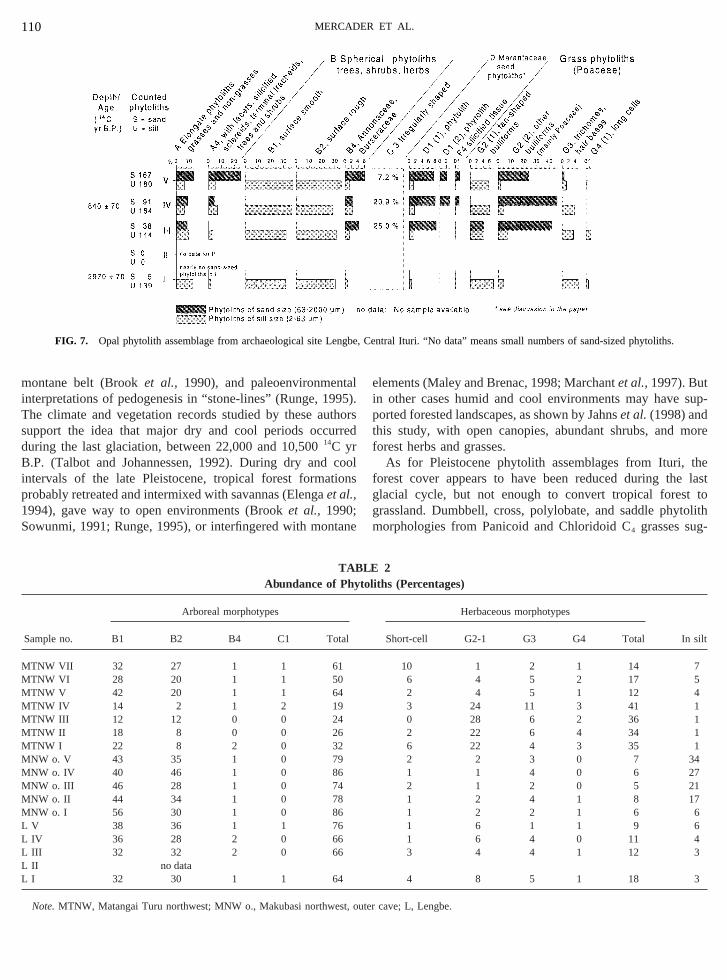
Lengbe (Epulu). The late Holocene sequence of Lenspans the last 300014C yr B.P. As with the sequence fro
akubasi NW, its main feature is the dominance of phyto
TABRadioca
Site Lab no. Archaeological level
MSE Os-21250 basalMTNW UtC Nr5075 2IBSW AA33225 basalWGE AA 33226 basalL Beta 127079 1MNW o. UtC Nr 5076 3MNW o. Os-22251 3MNW o. UtC Nr 5077 3L Beta 127078 1MTNW Os-21235 5MTNW UtC Nr 5074 5MTNW Os-21251 5
Note.MSE, Makubasi southeast; MTNW, Matangai Turu northwest; IBnorthwest outer cave.
n
dre
s-tss
the
ata
-m
eC,
te
ee.
a-
s
typical of rain forests (Fig. 7). Changes in forest composduring the Holocene may be indicated by phytolith C1, funknown trees, which is present low in the sequence, dpears in the middle of the sequence, and reappears nearof the sequence. Phytoliths C3, D, and F peak during themillennium.
Although all sites suggest diachronic changes in planand, perhaps, in local forest composition, only Matangai TNW, the sole site with evidence for late Pleistocene throearly Holocene human occupation, shows abrupt changtree–grass ratios (Table 2). Sample IV, dated to 10,53014C yrB.P., contains more arboreal phytoliths and fewer grasstoliths than do samples from underlying deposits.
DISCUSSION
Phytolith Representation, Source Area, and Identification
Phytoliths generally abound at all the sites examined (cader, 2000). They appear to come from a diverse set of pand plant structures (e.g., tree and herbaceous seedand herbaceous leaves, bark, woods). Phytoliths consispresent include those from the Annonaceae, BurseraceaMarantaceae. Poaceae phytoliths are also well represencertain time periods. Additional phytoliths, though curreunidentified, probably come from plants of the tropical fobecause they abound in “modern” sediments in the regforests. Such phytoliths include types C1 and C2, whichalso common in tropical forests of central and south Ame(Piperno and Becker, 1996; Piperno, 1998).
In cave sediments of equatorial regions, most phytorepresent local andin situ deposition, as dense vegetatprevents long-distance transport by wind. Anthropogenicterials (e.g., foodstuffs, thatching material, sleeping mattremedicinal and ritual plants, leaves brought in with branc
1n Ages
Material dated Age (14C yr B.P.)
mm charcoal 18,8006 100mm charcoal 10,5306 50docarp ofCanarium schweinfurthii 10,0156 55mm charcoal 60256 70docarp ofCanarium schweinfurthii 29706 70
mm charcoal 10806 41mm charcoal 10106 60mm charcoal 9716 33docarp ofElaeis guineensis 8406 70docarp ofCanarium schweinfurthii 8256 30veral 1- to 5-mm fragments of human bone 8136 35mm charcoal 7156 45
, Isak Baite southwest; WGE, Wataka Gitatu east; L, Lengbe; MNW o.,
LErbo
1,1
en1en222enense2
SW

nteltasht a, in–
enrereen
ock
vidceea
tolireproge98hic
icots.calanderno,lithsdern
ofis
ies in996)l sa-,8).nantor-urser-of ain the
oc-
tf
PHYTOLITHS IN THE CONGO 107
collected for firewood) and colluvial influx (including plamaterial growing and decomposing around the rock shbrought in through the sides of the caves by rain wlikely account for the great majority of phytoliths presenMatangai Turu NW, Makubasi NW, and Lengbe. Todaythe Ituri forest, plants are collected by women within 0.1km of their camps (Ichikawa, 1992; Bailey, 1991). Ancitropical forest foragers probably gathered their plantsources from similarly nearby places. It is likely, therefothat phytoliths represent the plants and the environmthat people exploited within several kilometers of rshelters.
The modern and fossil phytoliths presented here prodirect and indirect evidence for late Pleistocene and Holooccupation of variably dense tropical forests of the northCongo basin by prehistoric Later Stone Age foragers. Phymorphotypes A, B, C, and D may emanate largely from fotaxa. Morphotype A comprises elongate phytoliths mostlyduced by trees and shrubs. It comes from various vestructures, particularly leaves (Piperno, 1988; Postek, 1Morphotype B is made up of spherical rugose bodies w
FIG. 3. Phytolith assemblages from modern mixed tropical rain foresorest (Lenda), Epulu, Central Ituri.
er)
t
4t-,ts
enestthst-
tal1).h
appear to derive mainly from wood, seeds, and leaves of dIt is the most common type of phytolith found in tropiforests, including those of Central and South AmericaSoutheast Asis (Piperno, 1993, 1998; Kealhofer and Pip1998; Runge, 1999). Significantly, spherical rugose phytofrom dicots are scarce in phytolith assemblages from motropical savanna and steppe environments (Barboniet al.,1999; Alexandreet al.,1997). Morphotype C is a phytolithunknown origin with a distinctive, stippled surface. C1produced by several tree taxa and is found in large quantitmodern soils of Amazonian forests (Piperno and Becker, 1but never in phytolith assemblages from modern tropicavannas of Africa (cf. Alexandreet al.,1997; Mworia-Maitima1997; Barboniet al.,1999) or America (Piperno, 1988, 199In the Ituri forest, however, C1 may indicate a monodomiGilbertiodendronstand. Morphotype D abounds in mixed fests and belongs to the Marantaceae, perhaps to the Baceae. Therefore this phytolith type is a potential indicatorclosed canopy. In contrast, Poaceae phytoliths are scarcemodern profiles from the Ituri forest, reflecting their lowcurrence in the shady understory of this habitat today.
doro-Afarama), Epulu, Central Ituri, and modernGilbertiodendronmonodominan
t (E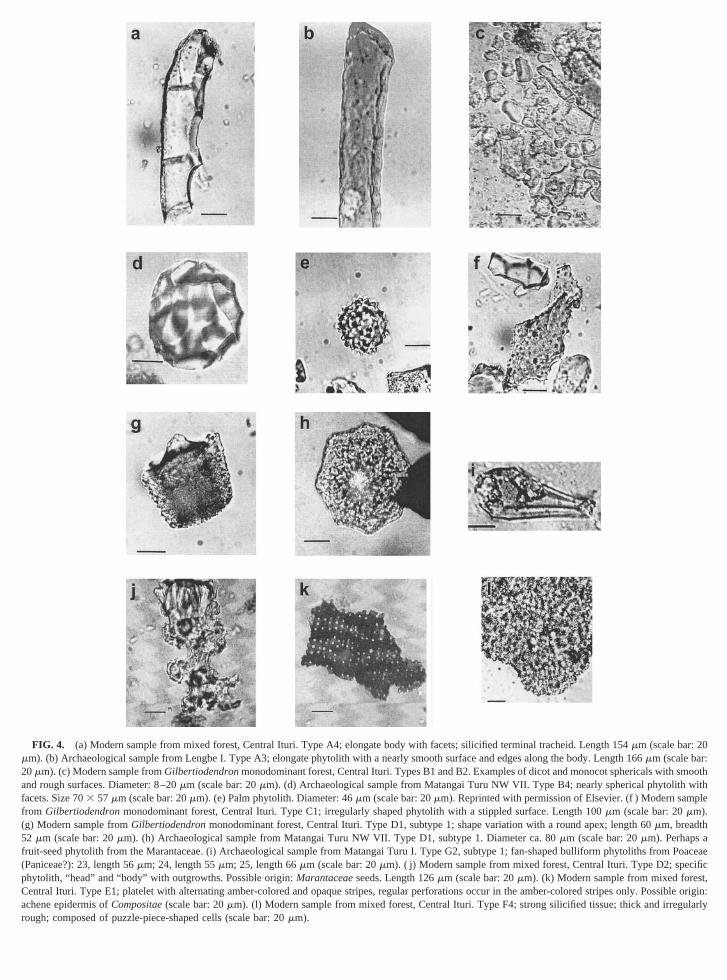
0:smoothwith
f plef(
aecificst,:ularly
r
FIG. 4. (a) Modern sample from mixed forest, Central Ituri. Type A4; elongate body with facets; silicified terminal tracheid. Length 154mm (scale bar: 2mm). (b) Archaeological sample from Lengbe I. Type A3; elongate phytolith with a nearly smooth surface and edges along the body. Length 166mm (scale bar20mm). (c) Modern sample fromGilbertiodendronmonodominant forest, Central Ituri. Types B1 and B2. Examples of dicot and monocot sphericals withand rough surfaces. Diameter: 8–20mm (scale bar: 20mm). (d) Archaeological sample from Matangai Turu NW VII. Type B4; nearly spherical phytolithacets. Size 703 57 mm (scale bar: 20mm). (e) Palm phytolith. Diameter: 46mm (scale bar: 20mm). Reprinted with permission of Elsevier. (f ) Modern samrom Gilbertiodendronmonodominant forest, Central Ituri. Type C1; irregularly shaped phytolith with a stippled surface. Length 100mm (scale bar: 20mm).g) Modern sample fromGilbertiodendronmonodominant forest, Central Ituri. Type D1, subtype 1; shape variation with a round apex; length 60mm, breadth
52 mm (scale bar: 20mm). (h) Archaeological sample from Matangai Turu NW VII. Type D1, subtype 1. Diameter ca. 80mm (scale bar: 20mm). Perhapsfruit-seed phytolith from the Marantaceae. (i) Archaeological sample from Matangai Turu I. Type G2, subtype 1; fan-shaped bulliform phytoliths from Poacea(Paniceae?): 23, length 56mm; 24, length 55mm; 25, length 66mm (scale bar: 20mm). ( j) Modern sample from mixed forest, Central Ituri. Type D2; spephytolith, “head” and “body” with outgrowths. Possible origin:Marantaceaeseeds. Length 126mm (scale bar: 20mm). (k) Modern sample from mixed foreCentral Ituri. Type E1; platelet with alternating amber-colored and opaque stripes, regular perforations occur in the amber-colored stripes only.Possible originachene epidermis ofCompositae(scale bar: 20mm). (l) Modern sample from mixed forest, Central Ituri. Type F4; strong silicified tissue; thick and irregough; composed of puzzle-piece-shaped cells (scale bar: 20mm).
fro. R
dr
n ns
98),con-Rift
1 entsi afro-
phytoliths.
PHYTOLITHS IN THE CONGO 109
Paleoenvironmental Changes
At present, few paleoenvironmental data are availablethe lowland evergreen forests of the Congo’s watershedconstructions of Quaternary environments are based onfrom lakes located in mosaic areas (Elengaet al.,1994), the fa
orthwestern corner of the central African forest at its tra
FIG. 5. Opal phytolith assemblage from archaeological site Matang
FIG. 6. Opal phytolith assemblage from
me-ata
i-
tion to the West African domain (Maley and Brenac, 19and West Africa (Talbot and Johannessen, 1992). Othertexts include pollen data from the neighboring Western(Sowunmi, 1991; Bergonziniet al., 1997; Marchantet al.,
997), cave pollen from savanna–forest mosaic environmn their transition to both Western Rift savannas and the
uru NW, Northern Ituri. “No data” means small numbers of sand-sized
haeological site Makubasi NWo., Eastern Ituri.
ai T
arc
tal95thrre
coion
tan
ti sup-p
more
thelast
t totolith
hs.
MERCADER ET AL.110
montane belt (Brooket al., 1990), and paleoenvironmeninterpretations of pedogenesis in “stone-lines” (Runge, 19The climate and vegetation records studied by these ausupport the idea that major dry and cool periods occuduring the last glaciation, between 22,000 and 10,50014C yrB.P. (Talbot and Johannessen, 1992). During dry andintervals of the late Pleistocene, tropical forest formatprobably retreated and intermixed with savannas (Elengaet al.,1994), gave way to open environments (Brooket al., 1990;Sowunmi, 1991; Runge, 1995), or interfingered with mon
TABAbundance of Phy
Sample no.
Arboreal morphotypes
B1 B2 B4 C1 Total
MTNW VII 32 27 1 1 61MTNW VI 28 20 1 1 50MTNW V 42 20 1 1 64MTNW IV 14 2 1 2 19MTNW III 12 12 0 0 24MTNW II 18 8 0 0 26MTNW I 22 8 2 0 32MNW o. V 43 35 1 0 79MNW o. IV 40 46 1 0 86MNW o. III 46 28 1 0 74MNW o. II 44 34 1 0 78MNW o. I 56 30 1 0 86L V 38 36 1 1 76L IV 36 28 2 0 66L III 32 32 2 0 66L II no dataL I 32 30 1 1 64
Note.MTNW, Matangai Turu northwest; MNW o., Makubasi northwes
FIG. 7. Opal phytolith assemblage from archaeological site Le
).orsd
ols
e
elements (Maley and Brenac, 1998; Marchantet al.,1997). Bun other cases humid and cool environments may haveorted forested landscapes, as shown by Jahnset al.(1998) and
this study, with open canopies, abundant shrubs, andforest herbs and grasses.
As for Pleistocene phytolith assemblages from Ituri,forest cover appears to have been reduced during theglacial cycle, but not enough to convert tropical foresgrassland. Dumbbell, cross, polylobate, and saddle phymorphologies from Panicoid and Chloridoid C4 grasses sug-
2iths (Percentages)
Herbaceous morphotypes
In siltShort-cell G2-1 G3 G4 Total
10 1 2 1 14 76 4 5 2 17 52 4 5 1 12 43 24 11 3 41 10 28 6 2 36 12 22 6 4 34 16 22 4 3 35 12 2 3 0 7 341 1 4 0 6 272 1 2 0 5 211 2 4 1 8 171 2 2 1 6 61 6 1 1 9 61 6 4 0 11 43 4 4 1 12 3
4 8 5 1 18 3
uter cave; L, Lengbe.
e, Central Ituri. “No data” means small numbers of sand-sized phytolit
LEtol
t, o
ngb

ssetyp0%0%the
s gor t thP bs bort i lol ia ot deu cd thc CO
e ps
lity, mltert oclnomangodesta
aps
unog
” orinno
s orongrsiouor
lac
ithricapic
, Rersierr
, L.
97).ne inlogy
rest,y of
ter, E.
lithsiopia.
paleo-from
ta for
az,
onal
). Acial
dum.
Late-
delom-
H .
n bypper
ixedEast
ubsis-y
96).asin,
en,pp.
buti,
istory(site
PHYTOLITHS IN THE CONGO 111
gestive of savannas are rare in the Pleistocene phytolith ablages from the Ituri region. Indeed, the presence of thisof aridity marker in the Ituri assemblages is very low (5–1Figs. 5–7, Table 2), while they largely abound (22–4Alexandreet al., 1997) in the phytolith assemblages fromavannas and forest–savanna mosaics of the Guineo-Conegion (Alexandreet al.,1997). These changes suggest thaleistocene to Holocene transition in the area was markedhift to a denser forest cover. Nevertheless, because araxa are represented throughout the sequence, the Iturands probably remained forested during the final millennhe late Pleistocene. Perhaps the forests contained anderstory of many shrubs and herbaceous plants. Suchitions are predicted by models of vegetational changesonsider the effects on plant growth of lower atmospheric2
(e.g., Cowling, 1999). The decrease in phytolith occurrencunit of a silt fraction suggests fewer occupation episodescarcer use of plants by Pleistocene foragers. The variabithe Holocene phytolith assemblages, on the other handreflect food gathering and processing at the rock shechanging spatial distribution of plant resources in this pathe Ituri region, or changes in forest composition due tomatic factors that altered the abundance and distributioplant resources available for exploitation. Tropical forest cmunities were not stable even after the vegetational rearrments that marked the close of the Pleistocene. In fact, mclimate and vegetation patterns in central Africa were elished only after 3,00014C yr B.P. (Elengaet al.,1994; Hartetal., 1996; Jollyet al.,1998; Maley and Brenac, 1998), perheven centuries ago (Alexandreet al., 1997, p. 226).
Our phytolith analysis also suggests that the spatial boaries between forest and grasslands, as shown in the palegraphical depiction of the northeastern Congo “refugium“core area” (Hamilton, 1972), should be reconsidered. Duthe LGM the core area may have consisted of heterogevegetational formations, including forests (Ituri), mosaicforests and grasslands (Matupi Cave: Van Noten, 1977; Bet al., 1990), and, perhaps, parkland environments (Ru1995), rather than a homogenous forest block. This diveimplies that proposed areas of forest refugia probably shnot be depicted as monolithic havens in space and time,stable zones unaffected by a dryer and cooler late gmaximum.
As for the antiquity of human colonization in Ituri, phytoland archaeological data indicate that Later Stone Age Afforagers lived in and exploited plant resources of the troforest (Mercader, 1997) by perhaps 19,00014C yr B.P.
ACKNOWLEDGMENTS
We thank the Spanish Ministry of Education and Culture for fundsBailey and the Department of Anthropology at George Washington Univfor support and guidance, R. Marti, O. Carreras, I. Gonzalez, and J. Gu
m-e,;
lianey aealw-fnseon-at
erorinayrs,f
i-of-e-rn
b-
d-eo-rgusfoke,tyldasial
nal
.tyero
for help with field work, and T. Hart, J. Hart, A. Brooks, D. PipernoKealhofer, J. Runge, and J. Maley for discussions.
REFERENCES
Alexandre, A., Meunier, J., Lezine, A., Vincens, A., and Schwartz, D. (19Phytoliths: Indicators of grassland dynamics during the Late Holoceintertropical Africa.Palaeogeography, Palaeoclimatology, Palaeoeco136,213–229.
Bailey, R. (1991). The behavioral ecology of Efe pygmy men in the Ituri foZaire, Anthropological Paper 86, Museum of Anthropology, UniversitMichigan, Ann Arbor.
Bailey, R., Head, G., Jenike, M., Owen, B., Rechtman, R., and Zechen(1989). Hunting and gathering in tropical rain forest: Is it possible?Amer-ican Anthropologist91, 59–82.
Barboni, D., Bonnefille, R., Alexandre, A., and Meunier, J. (1999). Phytoas paleoenvironmental indicators, west side Middle Awash Valley, EthPalaeogeography, Palaeoclimatology, Palaeoecology152,87–100.
Bergonzini, L., Chalie, F., and Gasse, F. (1997). Paleoevaporation andprecipitation in the Tanganyika Basin at 18,000 years B.P. inferredhydrologic and vegetation proxies.Quaternary Research47, 295–305.
Brook, G., Burney, D., and Cowart, J. (1990). Paleoenvironmental daIturi, Zaire, from sediments in Matupi Cave, Mount Hoyo.In “Evolution ofEnvironments and Hominidae in the African Western Rift Valley” (N. BoEd.), pp. 49–70. Virginia Museum of Natural History, Martinsville.
Bultot, F. (1971). “Atlas Climatique du Bassin Congolase.” Institute Natipour l’Etudes Agronomique du Congo, Brussels.
Colinvaux, P., Oliveira, P., Moreno, E., Miller, C., and Bush, M. (1996long pollen record from lowland Amazonia: Forest and cooling in glatimes.Science274,85–88.
Cowling, S. (1999). Simulated effects of low atmospheric CO2 on structure ancomposition of North American vegetation at the Last Glacial MaximGlobal Ecology and Biogeography8, 81–93.
Elenga, H., Schwartz, D., and Vincens, A. (1994). Pollen evidence ofQuaternary vegetation and inferred climate changes in Congo.Palaeogeography, Palaeoclimatology, Palaeoecology109,345–356.
Gonzalez, I. (1997). Estudio petrolo´gico de la regio´n noreste del Ituri, Zaire.In“Bajo el Techo Forestal. La Evolucio´n del Poblamiento en el BosqueIturi, Zaire” (J. Mercader, unpublished Ph.D. thesis, Universidad Cplutense, Madrid), pp. 372–385.
affer, J. (1969). Speciation in Amazonian forest birds.Science165,131–136
Hamilton, A. (1972). The significance of patterns of distribution showforest plants and animals in tropical Africa for the reconstruction of UPleistocene palaeoenvironments.Palaeoecology of Africa9, 63–97.
Hart, T. (1985). “The Ecology of a Single-Species Dominant Forest and MForest in Zaire.” Unpublished Ph.D. thesis, Michigan State University,Lansing.
Hart, T., and Hart, J. (1986). The ecological basis of hunter-gatherer stence in African rain forests: The Mbuti of eastern Zaire.Human Ecolog14, 29–55.
Hart, T., Hart, J., Dechamps, M., Fournier, M., and Ataholo, M. (19Changes in forest composition over the last 4000 years in the Ituri BZaire. In “The Biodiversity of African Plants” (L. J. G. van der MaesX. M. van der Burgt, and J. M. van Medenback de Kooy, Eds.),545–563. Kluwer Academic, Dordrecht.
Ichikawa, M. (1992). Traditional use of tropical rain forest by the Mhunter-gatherers in central Africa.In “Topics in Primatology” (N. ItoigawaEd.), pp. 305–317. University of Tokyo Press, Tokyo.
Jahns, S., Huls, M., and Sarnthein, M. (1998). Vegetation and climate hof west equatorial Africa based on a marine pollen record off Liberia

nd
atewit
flor
rif, F
,
M mene la.
M an
M niver
M DaAAoaa
M inde
M stetolit
P ica
P in tns
.), pp.del-
P ssils:pical
P entraltural
P ricancs and
P
R a—nd
R etatione
S rica:
T maticand
-
V acial.
V tional
V
W anyrces
MERCADER ET AL.112
GIK 16776) covering the last 400,000 years.Review of Palaeobotany aPalynology102,277–288.
Jolly, D., Harrison, S., Damnati, B., and Bonnefille, B. (1998). Simulclimate and biomes of Africa during the late Quaternary: Comparisonpollen and lake status data.Quaternary Science Reviews17, 629–658.
Kealhofer, L., and Piperno, D. (1998). Opal phytoliths in Southeast AsianSmithsonian Contributions to Botany88, 1–39.
Kutzback, J., Gallimore, R., Harrison, S., Behling, P., Selin, R., and Laa(1998). Climate and biome simulations for the past 21,000 years.Quater-nary Science Reviews17, 473–506.
Lavreau, J. (1982). Etude geologique du Haut-Zaı¨re. Gene`se et e´volution d’unsegment lithosphe´rique Archeen, Annales, Sciences ge´ologiques, no. 88MuseeRoyal de l’Afrique Centrale, Tervuren, Belgium.
aley, J., and Brenac, P. (1998). Vegetation dynamics, palaeoenvironand climatic changes in the forests of western Cameroon during th28,000 years B.P.Review of Palaeobotany and Palynology1998, 157–187
archant, R., Taylor, D., and Hamilton, A. (1997). Late PleistoceneHolocene history at Mubwindi Swamp, southwest Uganda.QuaternaryResearch47, 316–328.
ercader, J. (1997). “Bajo el Techo Forestal. La Evolucio´n del Poblamiento eel Bosque Ecuatorial del Ituri, Zaire.” Unpublished Ph.D. thesis, Unsidad Complutense, Madrid.
ercader, J. (2000). Congo contextual data for phytolith analysis. Worldcenter-A for paleoclimatology data contribution series 1999-044 NONGDC Paleoclimatology Program, Boulder, CO. [http://www.ngdc.ngov/paleo.html]
ercader, J., Garcı´a-Heras, M., and Gonza´lez, I. (2000). Ceramic traditionthe African rain forest: Characterization analysis of ancient and mopottery from Ituri, D.R. Congo.Journal of Archaeological Science27,163–182.
woria-Maitima, J. (1997). Prehistoric fires and land cover change in weKenya: Evidences from pollen, charcoal, grass cuticles, and grass phyThe Holocene7, 409–417.
iperno, D. (1988). “Phytolith Analysis. An Archaeological and GeologPerspective.” Academic Press, London.
iperno, D. (1993). Phytolith and charcoal records from deep lake coresAmerican tropics.In “Current Research in Phytolith Analysis: Applicatio
dh
a.
.
ts,st
d
-
ta/.
rn
rnhs.
l
he
in Archaeology and Paleoecology” (D. Pearsall and D. Piperno, Eds58–71. University Museum of Archaeology and Anthropology, Philaphia.
iperno, D. (1998). Paleoethnobotany in the neotropics from microfoNew insights into ancient plant use and agricultural origins in the troforest.Journal of World Prehistory12, 393–449.
iperno, D., and Becker, P. (1996). Vegetational history of a site in the cAmazon basin derived from phytolith and charcoal records from nasoils.Quaternary Research45, 202–209.
iperno, D., and Pearsall, D. (1998). The silica bodies of tropical Amegrasses: Morphology, taxonomy, and implications for grass systematifossil phytolith identification.Smithsonian Contributions to Botany85,1–40.
ostek, M. (1981). The occurrence of silica in the leaves ofMagnolia gran-diflora. Botanical Gazette124,124–134.
unge, F. (1999). The opal phytolith inventory of soils in Central AfricQuantities, shapes, classification, and spectra.Review of Palaeobotany aPalynology107,23–53.
unge, J. (1995). New results on Late Quaternary landscape and vegdynamics in eastern Zaire (Central Africa).Zeitschrift fur Geomorphologi99, 65–74.
owunmi, M. (1991). Late Quaternary environments in equatorial AfPalynological evidence.Palaeoecology of Africa22, 213–238.
albot, M., and Johannessen, T. (1992). A high resolution palaeoclirecord for the last 27,500 years in tropical West Africa from the carbonnitrogen isotopic composition of lacustrine organic matter.Earth and Planetary Science Letters110,23–37.
an der Hammen, T., and Absy, M. (1994). Amazonia during the last glPalaeogeography, Palaeoclimatology, Palaeoecology109,247–261.
an der Kaars, W., and Dam, M. (1995). A 135,000 year record of vegetaand climatic change from the Bandung area, West Java, Indonesia.Palaeo-geography, Palaeclimatology, Palaeoecology117,55–72.
an Noten, F. (1977). Excavations at Matupi Cave.Antiquity 51, 35–40.
hite, F. (1983). Vegetation of Africa: A descriptive memoir to accompthe UNESCO/AETFAT/UNSO vegetation map of Africa, Natural ResouResearch, no. 20. UNESCO, Paris.