physicochemical and kinetic characteristics of rhodanese from the liver of african catfish clarias...
TRANSCRIPT

Physicochemical and kinetic characteristics of rhodanesefrom the liver of African catfish Clarias gariepinus Burchellin Asejire lake
Omolara Titilayo Akinsiku ÆFemi Kayode Agboola Æ Adenike Kuku ÆAdeyinka Afolayan
Received: 9 December 2008 / Accepted: 21 April 2009 / Published online: 18 June 2009
� Springer Science+Business Media B.V. 2009
Abstract Two forms of rhodanese were purified
from the liver of Clarias gariepinus Burchell, desig-
nated catfish rhodanese I (cRHD I) and rhodanese II
(cRHD II), by ion-exchange chromatography on a CM-
Sepharose CL-6B column and gel filtration through a
Sephadex G-75 column. The apparent molecular
weight obtained for cRHD I and cRHD II was
34,500 ± 707 and 36,800 ± 283 Da, respectively.
The subunit molecular weight determined by sodium
dodecyl sulphate–polyacrylamide gel electrophoresis
was 33,200 ± 283 and 35,100 ± 141 Da for cRHD I
and cRHD II, respectively. Atomic absorption spec-
trophotometric analysis revealed that cRHD II con-
tained a high level of iron (Fe), which presumably was
responsible for the brownish colour of the preparation.
In contrast, no Fe was identified in cRHD I, and its
preparation was colourless. Further characterization of
cRHD II gave true Michaelis–Menten constant (Km)
values of 25.40 ± 1.70 and 18.60 ± 1.68 mM for
KCN and Na2S2O3, respectively, an optimum pH of
6.5 and an optimum temperature of 40�C. The
Arrhenius plot of the effects of temperature on the
reaction rate consisted of two linear segments with a
break occurring at 40�C. The apparent activation energy
values from these slopes were 7.3 and 72.9 kcal/mol.
Inhibition studies on the cRHD II enzyme showed that
the activity of the enzyme was not affected by Mn2?,
Co2?, Sn2?, Ni2? and NH4?, but Zn2? inhibited the
enzyme considerably.
Keywords Aquatic organisms � Asejire Lake �Catfish (Clarias gariepinus) � Cyanide �Cyanide detoxification � Cyanogenic plants �Fish � Rhodanese
Introduction
Rhodanese (thiosulphate: cyanide sulphurtransferase,
EC 2.8.1.1) is a sulphur transferase that catalyses, in
vitro, the formation of thiocyanate from cyanide and
thiosulphate or other suitable sulphur donors. Differ-
ent thiosulphonates are able to replace thiosulphate in
the rhodanese reaction (Sorbo 1953a), and sulphite,
sulphanate and persulphide can also serve as donor
substrates (Sorbo 1957; Villarejo and Westley 1963;
Koj 1968; Nagahara et al. 1999). In vivo, however,
the enzyme is multifunctional: it supplies sulphide for
the formation of iron–sulphur centres, maintains the
sulphane pool and participates in selenium metabolism
and thiamine biosynthesis (Smith and Urbanska 1986;
The data presented in this report is part of the research carried
out by O. T. Akinsiku in partial fulfilment for the degree of
Master of Science in Biochemistry at the Obafemi Awolowo
University, Ile-Ife, Nigeria.
O. T. Akinsiku � F. K. Agboola (&) � A. Kuku �A. Afolayan
Department of Biochemistry, Obafemi Awolowo
University, Ile-Ife, Nigeria
e-mail: [email protected]
123
Fish Physiol Biochem (2010) 36:573–586
DOI 10.1007/s10695-009-9328-4

Bordo and Bork 2002). It is a ubiquitous enzyme that is
active in all living organisms, from bacteria to Homo
sapiens and plants (Cosby and Summer 1945; Him-
wich and Saunders 1948; Sorbo 1951; Jarabak and
Westley 1974; Anosike and Ugochukwu 1981; Lee
et al. 1995; Nagahara et al. 1999; Agboola and Okonji
2004; Aminlari et al. 2007). Studies on its intracellular
distribution have revealed that it is present in the
nucleus, mitochondrion and cytosol, with the greatest
concentration in the mitochondrion (Sorbo 1951;
Nagahara et al. 1999; Ali et al. 2001). It is generally
believed that the major function of rhodanese is
cyanide detoxification, especially in mammals in
whom cyanide is converted to the less toxic thiocyanate
and excreted through the kidney (Westley 1980). In
plants, a close relationship exists between rhodanese
activity and cyanogenesis, which suggests that the
enzyme provides a mechanism for cyanide detoxifica-
tion in cyanogenic plants (Smith and Urbanska 1986).
Urbanization and industrial, agricultural and pop-
ulation growth have been accompanied by a tremen-
dous increase in the discharge of a wide diversity of
pollutants to receiving water bodies, causing desirable
effects on different components of the aquatic envi-
ronment (Tolba 1982). Organic pollution of inland
waters in Nigeria, in contrast to the situation in
developed countries of the world, is often the result of
extreme poverty and economic and social under-
development. Fish and marine resources in Nigeria
face total collapse or extinction due to over-fishing and
the destruction of marine life and natural habitats by
pollution of water bodies. Unregulated and excessive
use of pesticides for fishing and the deliberate disposal
and dumping of toxic and hazardous wastes into water
bodies are significant causes of massive fish kills and
the loss of aquatic life and habitats in this country.
Lake Asejire, a man-made lake constructed on the
Osun river in 1972 in Oyo State, Nigeria, has elevated
levels of contaminants due to receiving effluents from
various industries and the breakdown of ecological
balance caused by the widespread destruction of flora
and fauna diversities (Lameed and Obadara 2006).
Despite this high level of pollution, the lake still
supports aquatic life. Cyanide, produced by toxic gases
during the pyrolysis of plastic or nitrile-based polymer
fibres, by extracts of plants containing cyanogenic
glycosides (e.g. cassava) or from industrial waste (e.g.
electroplating), is one of the likely chemical pollutants
present in the water. The African catfish (Clarias
gariepinus), like many other fish, inhabits diverse
water bodies, ranging from lakes, streams, rivers,
swamps to floodplains (Bruton 1979; Clay 1979).
Some of these water bodies are situated in areas where
they are prone to contamination and pollution from
various sources. This paper reports on the purification
and partial characterization of rhodanese isolated in
the liver of Clarias gariepinus. It is assumed that
this enzyme, among other functions, plays a role in
cyanide detoxification in catfish and, consequently, to
its adaptation or survival in such polluted waters.
Materials and methods
Materials
Sodium thiosulphate (pentahydrate), potassium cya-
nide, ammonium sulphate (enzyme grade) and phenyl
methane sulphonyl fluoride (PMSF) were obtained
from BDH Chemical (Poole, UK). Coomassie Brilliant
Blue, bovine serum albumin (BSA) and the standard
proteins contained in the Sigma Molecular Weight
Markers Calibration kit for sodium dodecyl sulphate–
polyacrylamide gel electrophoresis (SDS–PAGE;
molecular weight marker range 14,000–70,000),
including ovalbumin (chicken), pepsin (porcine stom-
ach mucosa), trypsinogen, (bovine, PMSF-treated),
b-lactoglobulin (bovine), lysozyme (egg white) and
a-chymotrypsinogen (bovine pancreas), were obtained
from the Sigma Chemical Company (St. Louis, MO).
CM-Sepharose CL-6B and Sephadex G-100 columns
were obtained from Pharmacia Fine Chemical (Upp-
sala, Sweden), and Bio-Gel P4 was obtained from Bio-
Rad Laboratories (Hercules, CA). All other reagents
were of analytical grade and were obtained from either
Sigma or BDH. All buffers contained 10 mM sodium
thiosulphate to stabilize the enzyme. The catfishes
were collected from Asejire Lake, Ibadan, Nigeria.
The livers were quickly excised and stored in the
freezer until required for analysis.
Methods
Enzyme and protein assays
Rhodanese activity was assayed and estimated
according to Agboola and Okonji (2004). The protein
concentration was determined using a modified
574 Fish Physiol Biochem (2010) 36:573–586
123

method of Gornall et al. (1949) and BSA as the
standard.
Enzyme preparation
A sixty grams wet weight of tissue (obtained from ten
fish) was homogenized in three volumes of 10 mM
sodium thiosulphate containing 1 mM PMSF (Buffer
A) in a Warring Blender. The homogenate was
filtered through a double layer of cheese cloth and the
debris rehomogenized in one volume of buffer A. The
second homogenate was also filtered through cheese
cloth. The combined filtrate was centrifuged at
20,000 rpm for 15 min at 4�C using a Beckman
Optima LE-80K ultracentrifuge (Beckman Coulter,
Fullerton, CA). The supernatant was brought to 65%
ammonium sulphate saturation by the addition of
solid ammonium sulphate and left for 12 h. The
resulting precipitate was collected by centrifugation
at 15,000 rpm for 30 min at 4�C and stored in a
minimal amount of Buffer A.
A CM-Sepharose CL-6B cation-exchanger resin
was washed and equilibrated in 0.2 M sodium
acetate, pH 5.4 (Buffer B) and packed into a
2.5 9 40-cm column. The ammonium sulphate pre-
cipitate was first desalted on a Bio-Gel P4 column
and then layered on the ion exchange column. The
column was first washed with 240 ml of 30 mM
sodium acetate pH 5.4 (Buffer C) followed by elution
with a 200 ml linear gradient of 0–1.0 M KCl in
Buffer C. The fractions were monitored at 280 nm for
protein and assayed for rhodanese activity. The active
fractions in two distinct peaks were pooled and
dialyzed in 70% glycerol in Buffer C. The brown-
coloured fraction was then rechromatographed on the
ion-exchange column under the same conditions.
Sephadex G-75 resin was swollen and equilibrated
in 0.1 M phosphate buffer, pH 7.2 (Buffer D) and
then packed into a 1.5 9 70-cm column. The pre-
ceding two active fractions were run separately.
Three-millilitre fractions were collected at a flow rate
of 15 ml/h. Protein was monitored at 280 nm, and the
fractions were assayed for enzyme activity. The
pooled active fraction was dialyzed in 70% glycerol
in Buffer D. The two enzyme fractions were desig-
nated cRHD I and cRHD II. Polyacrylamide gel
electrophoresis in the absence of SDS was carried out
by the procedure described in the Pharmacia’s
manual (Polyacrylamide Gel Electrophoresis,
laboratory techniques, revised edition, Feb.1983) to
ascertain the purity of the preparation.
Characterizations
Apart from molecular weight determination and the
presence of metal ions in association with the
enzyme, only cRHD II was further characterized.
Molecular weight determinations
The SDS–PAGE was carried out to determine the subunit
molecular weight on an 10% acrylamide gel using the
Sigma Molecular Weight Markers Calibration kit
(molecular weight marker range 14,000–70,000) for the
standards. The native molecular weight was determined
by gel filtration on a Sephadex G-100 column (1.5 9
70 cm). The standard proteins were BSA (67,000;
3 mg/ml) plus a-chymotrypsinogen (25,000; 3 mg/ml)
and ovine albumin (45,000; 3 mg/ml) plus lysozyme
(15,000; 3 mg/ml). A total of 3 ml of each combination
was layered and eluted with 10 mM phosphate buffer,
pH 7.0 at a flow rate of 10 ml/h. Fractions of 3 ml were
collected and monitored for protein at 280 nm.
Determination of kinetic parameters
The concentration of KCN varied between 5 and 50 mM
at different fixed concentrations (ranging from 10 to
50 mM) of Na2S2O3 and vice versa in an assay mixture
containing 10 mM borate buffer, pH 5.4 and 0.1 ml
of the appropriately diluted cRHD II enzyme. The
reciprocal of the initial reaction velocity (1/v) against the
reciprocal of the varied substrates at the fixed concen-
tration of the other substrate was plotted. The vertical
intercepts of these double reciprocal plots (representing
1/Vmax app) and the abscissa intercept (representing
1/Km app) were then replotted, respectively, against the
reciprocal of the fixed concentration of the other
substrate (secondary replots) in accordance with a
ping-pong bi-bi reaction mechanism (Florini and Ves-
tling 1957; Cleland 1970; Segel 1975). The true Km
values of the substrates were then estimated from the
values of the slope and intercept of the secondary replots.
Effect of pH
The effect of pH on cRHD II activity was studied
using the methods of Agboola and Okonji (2004).
Fish Physiol Biochem (2010) 36:573–586 575
123

The enzyme was assayed using the following buffers:
50 mM citrate buffer (pH 4.0–6.5), 10 mM phosphate
buffer (pH 7.0–8.5) and 50 mM borate buffer (pH
9.0–11). A reaction mixture of 1 ml contained 0.5 ml
of the respective buffer, 0.2 ml of 0.25 mM KCN,
0.2 ml of 0.25 mM Na2S2O3 and 0.1 ml of the
enzyme solution.
Effect of cations
The method of Lee et al. (1995) was used to study the
effect of various metal ions on cRHD II activity. The
salts of the cations include CoCl2, MnCl2, NH4Cl,
NiCl2, SnCl2, ZnCl2 and MgCl2 at final concentra-
tions of 0.05 and 0.1 mM in enzyme assay mixture.
Effect of temperature
cRHD II was assayed at temperatures between 0� and
70�C to determine the optimum temperature of the
enzyme. The assay mixture was first incubated at the
indicated temperature for 10 min prior to initiating a
reaction by the addition of an aliquot of the enzyme
which had been previously equilibrated at the same
temperature. The residual activity was then assayed.
The heat stability of the enzyme was determined by
incubating 0.6 ml of the enzyme for 1 h at 30�, 40�, 50�,
60� and 70�C, respectively. A 0.1-ml aliquot was
withdrawn at 10-min intervals and assayed for residual
activity. The activity was expressed as a percentage of
activity of the enzyme incubated at 30�C as the control.
Ultraviolet/visible light spectrophotometric scanning
The post-gel filtration enzyme preparation was dia-
lysed against several changes of distilled water to
remove any ions contributed by the buffer. The
dialysate was scanned at wavelengths between 200
and 600 nm using a Cintra 101 Double Beam UV/
VIS Spectrophotometer (GBC Scientific Equipment
Pty Ltd, Australia).
Atomic absorption spectrophotometry
The dialysate was examined for the presence of
various divalent metal ions: Fe2?, Mg2?, Mn2?,
Ca2?, Co2?, Zn2?, Ni2? and Cu2?. These metal ions
were estimated by atomic absorption spectrophotom-
etry using an air-acetylene flame in the Alpha-4
ChemTech spectrophotometer. The sample was exten-
sively dialyzed against two change of distilled water to
remove any ions contributed by the buffer before it
was digested in a mixture of concentrated HNO3 and
HClO4 (1:1) according to the methods of Kaur et al.
(2006).
Results
Purification and molecular weight determination
The result of the purification of rhodanese from the
liver of catfish is summarized in Table 1. The elution
profile after ion-exchange chromatography on the
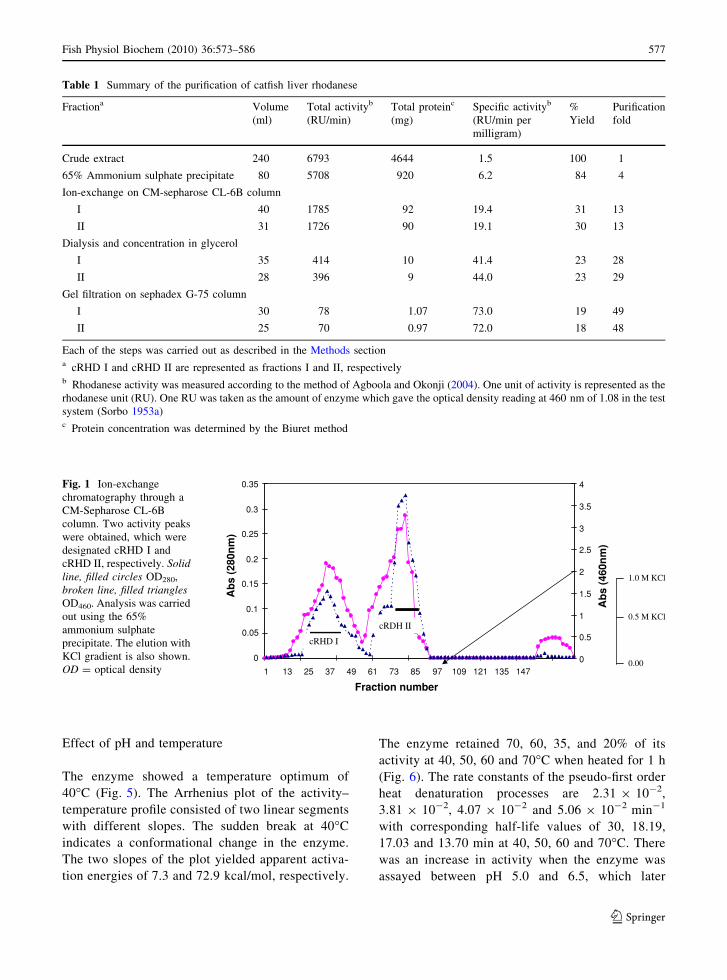
CM-Sepharose CL-6B column is shown in Fig. 1.
Two peaks of enzyme activity were obtained from
this analysis in the breakthrough volume without
the salt gradient elution. These two peaks were also
obtained in two subsequent repetitions of this
experiment, which lead us to consider the peaks as
two different forms of rhodanese, subsequently
designated catfish rhodanese I (cRHD I) and rhoda-
nese II (cRHD II). cRHD II was observed to be
brownish in colour, and this colour could not be
removed by the ion-exchange and gel-filtration
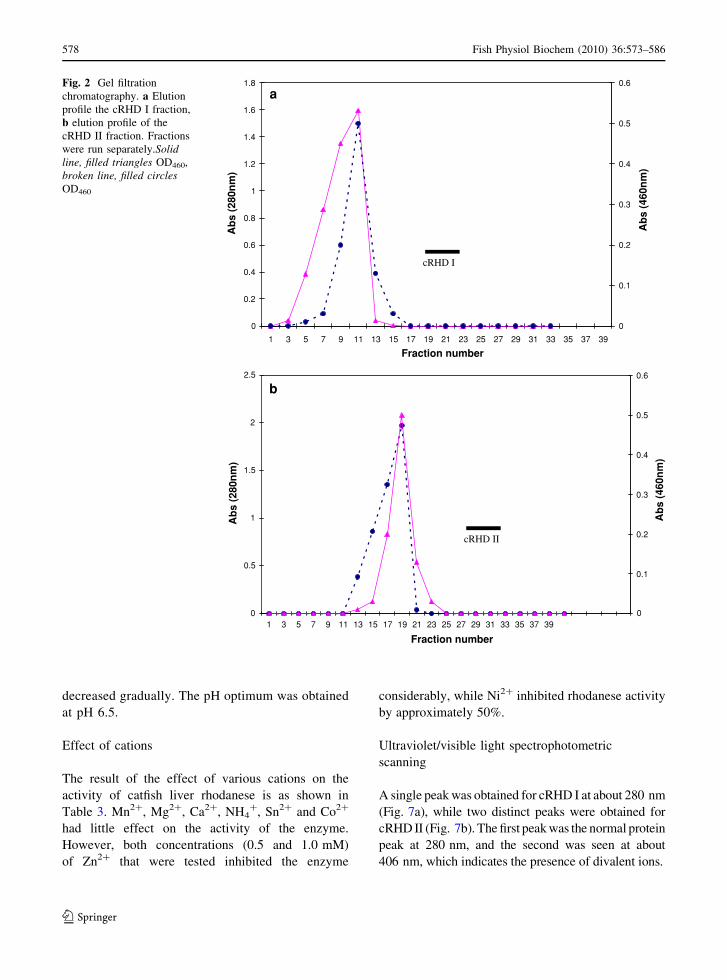
steps. The elution profile on the Sephadex G-75
column is shown in Fig. 2a and b. The photograph
showing the result of PAGE in the absence of SDS is
shown in Fig. 3.
The native molecular weights of the two forms of
the enzyme were estimated by gel filtration to be
34,500 ± 707 and 36,800 ± 283 Da for cRHD I and
cRHD II, respectively, while the subunit molecular
weights estimated by SDS–PAGE were 33,200 ± 283
and 35,100 ± 141 Da for cRHD I and cRHD II,
respectively.
Kinetic parameters
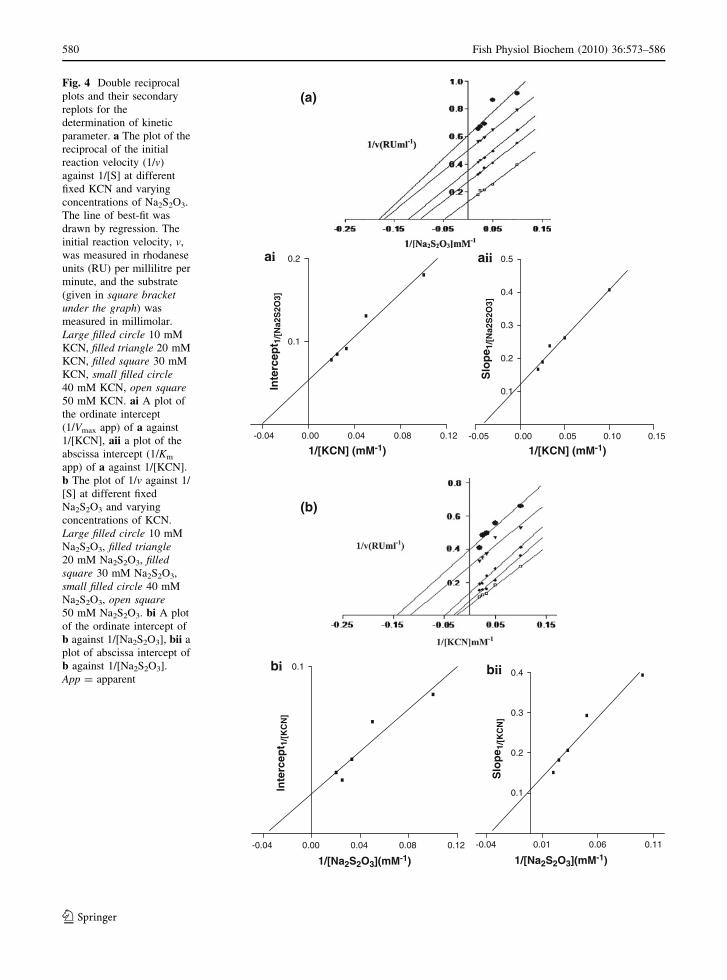
A typical double reciprocal plot of the change in
KCN concentrations at fixed Na2S2O3 concentrations
is shown in Fig. 4a and that of varying concentrations
of Na2S2O3 at fixed KCN concentrations is shown in
Fig. 4b. The lines are almost parallel, depicting an
enzyme ping-pong mechanism (Florini and Vestling
1957; Cleland 1970; Segel 1975). After the secondary
replots, the kinetic parameters were estimated; these
are summarized in Table 2.
576 Fish Physiol Biochem (2010) 36:573–586
123
Effect of pH and temperature
The enzyme showed a temperature optimum of
40�C (Fig. 5). The Arrhenius plot of the activity–
temperature profile consisted of two linear segments
with different slopes. The sudden break at 40�C
indicates a conformational change in the enzyme.
The two slopes of the plot yielded apparent activa-
tion energies of 7.3 and 72.9 kcal/mol, respectively.
The enzyme retained 70, 60, 35, and 20% of its
activity at 40, 50, 60 and 70�C when heated for 1 h
(Fig. 6). The rate constants of the pseudo-first order
heat denaturation processes are 2.31 9 10-2,
3.81 9 10-2, 4.07 9 10-2 and 5.06 9 10-2 min-1
with corresponding half-life values of 30, 18.19,
17.03 and 13.70 min at 40, 50, 60 and 70�C. There
was an increase in activity when the enzyme was
assayed between pH 5.0 and 6.5, which later
Table 1 Summary of the purification of catfish liver rhodanese
Fractiona Volume
(ml)
Total activityb
(RU/min)
Total proteinc
(mg)
Specific activityb
(RU/min per
milligram)
%
Yield
Purification
fold
Crude extract 240 6793 4644 1.5 100 1
65% Ammonium sulphate precipitate 80 5708 920 6.2 84 4
Ion-exchange on CM-sepharose CL-6B column
I 40 1785 92 19.4 31 13
II 31 1726 90 19.1 30 13
Dialysis and concentration in glycerol
I 35 414 10 41.4 23 28
II 28 396 9 44.0 23 29
Gel filtration on sephadex G-75 column
I 30 78 1.07 73.0 19 49
II 25 70 0.97 72.0 18 48
Each of the steps was carried out as described in the Methods sectiona cRHD I and cRHD II are represented as fractions I and II, respectivelyb Rhodanese activity was measured according to the method of Agboola and Okonji (2004). One unit of activity is represented as the
rhodanese unit (RU). One RU was taken as the amount of enzyme which gave the optical density reading at 460 nm of 1.08 in the test
system (Sorbo 1953a)c Protein concentration was determined by the Biuret method
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
1 13 25 37 49 61 73 85 97 109 121 135 147
Fraction number
0
0.5
1
1.5
2
2.5
3
3.5
4
Ab
s (4
60n
m)
cRHD I
cRDH II
1.0 M KCl
0.5 M KCl
0.00
Ab
s (2
80n
m)
Fig. 1 Ion-exchange
chromatography through a
CM-Sepharose CL-6B
column. Two activity peaks
were obtained, which were
designated cRHD I and
cRHD II, respectively. Solidline, filled circles OD280,
broken line, filled trianglesOD460. Analysis was carried
out using the 65%
ammonium sulphate
precipitate. The elution with
KCl gradient is also shown.
OD = optical density
Fish Physiol Biochem (2010) 36:573–586 577
123
decreased gradually. The pH optimum was obtained
at pH 6.5.
Effect of cations
The result of the effect of various cations on the
activity of catfish liver rhodanese is as shown in
Table 3. Mn2?, Mg2?, Ca2?, NH4?, Sn2? and Co2?
had little effect on the activity of the enzyme.
However, both concentrations (0.5 and 1.0 mM)
of Zn2? that were tested inhibited the enzyme
considerably, while Ni2? inhibited rhodanese activity
by approximately 50%.
Ultraviolet/visible light spectrophotometric
scanning
A single peak was obtained for cRHD I at about 280 nm
(Fig. 7a), while two distinct peaks were obtained for
cRHD II (Fig. 7b). The first peak was the normal protein
peak at 280 nm, and the second was seen at about
406 nm, which indicates the presence of divalent ions.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39
Fraction number
Ab
s (2
80n
m)
0
0.1
0.2
0.3
0.4
0.5
0.6
Ab
s (4
60n
m)
0
0.5
1
1.5
2
2.5
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39
Fraction number
Ab
s (2
80n
m)
0
0.1
0.2
0.3
0.4
0.5
0.6
Ab
s (4
60n
m)
cRHD I
cRHD II
a
b
Fig. 2 Gel filtration
chromatography. a Elution
profile the cRHD I fraction,
b elution profile of the
cRHD II fraction. Fractions
were run separately.Solidline, filled triangles OD460,
broken line, filled circlesOD460
578 Fish Physiol Biochem (2010) 36:573–586
123

Metal ion content
The divalent metal ion content determined by atomic
absorption spectroscopy showed that the metal ion in
cRHD I was negligible, with the exception of Fe2?
and Mg2?, which were present at 0.53 and 0.34 ppm,
respectively. However, among the metal ions tested
for in cRHD II, Fe2? showed the highest concentra-
tion (11.9 ppm), followed by Ni2? (0.9 ppm), Mg2?
(0.47 ppm), Mn2? (0.45 ppm), Zn2? (0.35 ppm),
Ca2? (0.06 ppm), Cu2? and Co2? (0.00 ppm).
Discussion
The African catfish (Clarias gariepinus) inhabits
calm waters of lakes, streams, rivers, swamps and
floodplains, many of which are subject to contami-
nation from various sources, such as effluents
from manufacturing companies. Studies on the food
composition of C. gariepinus indicate that this fish is
either an omnivore or a predator (Bruton 1979; Clay
1979). Micha (1973) found that C. gariepinus fed
mainly on aquatic insects, fish and higher plant
debris. Some of these food items contain cyanogenic
glycosides which upon hydrolysis release cyanide or
hydrocyanic acid (HCN) (Wokes and Willimott 1951;
Montgomery 1965). Cyanide is a potent cytotoxic
agent that kills the cell by inhibiting cytochrome
oxidase of the mitochondrial electron transport chain
(Warburg 1911; Keilin 1929; Nagahara et al. 1995).
Metal processing and organic chemical industries as
well as iron and steel production are major sources of
cyanide released to aquatic environment (Smith et al.
1978, 1979; ATSDR 1989). Other sources include
hydrogen cyanide used in the fumigation of ships
infested by rodents or insects and cyanide salts used
in ore-extracting processes. Moreover, in the local
traditional settings around the river Osun, from which
Asejire Lake was constructed, it is common for
cassava tubers to be steeped in the flowing river water
for processing into other food products. This is a
major source of cyanide in this water. There are
reports that fishes are the most sensitive aquatic
organisms to cyanide, and cyanide hazards to fish,
wildlife, livestock and man are well documented.
Adverse effects on swimming and reproduction have
been recorded at levels of between 5 and 7.2 lg free
cyanide per litre, with lethal effects usually occurring
at levels of 20–76 lg/l (Eisler 1991). Massive kills of
fresh water fish due to the accidental discharge of
cyanide wastes are common occurrences (Holden and
Marsden 1964; Leduc 1978; Towill et al. 1978; EPA
1980). In one case, cyanide-containing mine effluents
from a Canadian tailings pond released into a nearby
creek killed more 20,000 steelhead (Oncorhynchus
mykiss; Leduc et al. 1982). However, the fact that
various species of catfish and other animals have been
able to adapt and survive in cyanide-contaminated
habitats has lead to the hypothesis that the rhodanese
enzyme, which has been demonstrated to be the
major hepatic enzyme for cyanide detoxification in
animals, provides an inherent safety for catfish and
other aquatic animals against the toxicity of cyanide.
Thus, catfish can be used as a model of survival and
adaptation of aquatic animals in cyanide-polluted
water. Rhodanese catalyses the formation of the
easily excreted thiocyanate from cyanide and thio-
sulphate and acts as general transferase for the
CRUDE A B
Fig. 3 Polyacrylamide gel electrophoresis carried out in the
absence of SDS using the continuous phosphate buffer system.
A single band of protein was obtained after staining with
Coomassie Brilliant Blue. A cRHD I, B cRHD II
Fish Physiol Biochem (2010) 36:573–586 579
123
-0.04 0.00 0.04 0.08 0.12
0.1
0.2
Inte
rcep
t 1/[
Na2
S2O
3]
-0.05 0.00 0.05 0.10 0.15
0.1
0.2
0.3
0.4
0.5
1/[KCN] (mM-1)1/[KCN] (mM-1)
Slo
pe 1
/[N
a2S
2O3]
-0.04 0.00 0.04 0.08 0.12
0.1
1/[Na2S2O3](mM-1) 1/[Na2S2O3](mM-1)
Inte
rcep
t 1/[
KC
N]
-0.04 0.01 0.06 0.11
0.1
0.2
0.3
0.4
Slo
pe 1
/[K
CN
]
(a)
ai aii
(b)
bi bii
Fig. 4 Double reciprocal
plots and their secondary
replots for the
determination of kinetic
parameter. a The plot of the
reciprocal of the initial
reaction velocity (1/v)
against 1/[S] at different
fixed KCN and varying
concentrations of Na2S2O3.
The line of best-fit was
drawn by regression. The
initial reaction velocity, v,
was measured in rhodanese
units (RU) per millilitre per
minute, and the substrate
(given in square bracketunder the graph) was
measured in millimolar.
Large filled circle 10 mM
KCN, filled triangle 20 mM
KCN, filled square 30 mM
KCN, small filled circle40 mM KCN, open square50 mM KCN. ai A plot of
the ordinate intercept
(1/Vmax app) of a against
1/[KCN], aii a plot of the
abscissa intercept (1/Km
app) of a against 1/[KCN].
b The plot of 1/v against 1/
[S] at different fixed
Na2S2O3 and varying
concentrations of KCN.
Large filled circle 10 mM
Na2S2O3, filled triangle20 mM Na2S2O3, filledsquare 30 mM Na2S2O3,
small filled circle 40 mM
Na2S2O3, open square50 mM Na2S2O3. bi A plot
of the ordinate intercept of
b against 1/[Na2S2O3], bii a
plot of abscissa intercept of
b against 1/[Na2S2O3].
App = apparent
580 Fish Physiol Biochem (2010) 36:573–586
123
exchange of divalent sulphur in the biosphere (Ogata
et al. 1989). Other enzymes involved in cyanide
detoxification include 3-mercaptopyruvate sulphur-
transferase (3-MST), thiosulphate reductase, cysta-
thionase and albumin (enzyme-like behaviour in
cyanide detoxification) (Lieske et al. 1996).
Our research has documented the existence of two
forms of rhodanese in the liver of the catfish Clarias
gariepinus. These two forms have a similar molecular
size. In an earlier study, Blumenthal and Heinrikson
(1971) obtained two active peaks (rhodanese A and
B) when bovine liver rhodanese was purified on a
DEAE-Sephadex column in combination with PAGE.
Horowitz and DeToma (1970) introduced the ion-
exchange step to replace and improve the yield from
acidic (pH 4.5) and ammonium sulphate precipita-
tions. These researchers also found that these two
forms were identical with respect to kinetic param-
eters, molecular weight and other structural proper-
ties. Based on their data, they reasoned that
rhodanese B may have arisen during the course of
purification by the deamination of the A form. They
also established that rhodanese A and B did not arise
due to population differences of several livers. The
existence of the bovine rhodanese enzyme in a
monomeric–dimer equilibrium of molecular weight
about 18,000–36,000 had earlier been demonstrated
by Volini et al. (1967). Ogata et al. (1989) reported
that the two enzymes of Blumenthal and Heinrikson
(1971) were phospho- and dephosphorhodaneses. In
the study reported here, cRHD II had a brownish
colour that was not removed by rechromatography on
the same ion-exchanger and the subsequent gel
filtration step. However, these preparations were
adjudged to be pure by the presence of a single
protein band after electrophoresis in both the pres-
ence and absence of SDS, respectively. An investi-
gation into the presence of ions in the preparations by
spectrophotometric scanning and atomic absorption
spectrophotometry revealed that cRHD II contains
the metal ion Fe2?, as evidenced by an absorption
Table 2 Summary of kinetic parameters
Parameters KCN Na2S2O3
Km (mM) 25.4 ± 1.70 18.6 ± 1.68
Vmax (RU/ml) 8.22 ± 0.59 9.06 ± 0.51
The kinetic parameters for the two substrates were determined
by the method of Florini and Vestling (1957) and Segel (1975)
20 30 40 50 60 700.0
0.5
1.0
1.5
2.0
Temperature (°C)
Act
ivity
(R
U/m
l/min
)
Fig. 5 Effect of temperature on catfish liver rhodanese
0 10 20 30 40 50 60 700
50
100
Time (min)
% A
ctiv
ity
Fig. 6 Effect of temperature on the stability of catfish liver
rhodanese. The activity at 30�, 40�, 50�, 60� and 70�C was
expressed as a percentage of activity of the enzyme incubated
at 30�C. Open triangle 30�C, filled circle 40�C, open square50�C, filled diamond 60�C, open circle 70�C
Table 3 Effect of cations on catfish liver rhodanese activity
Salt Enzyme residual activity (%)
0.5 mM 1.0 mM
None 100 100
MgCl2 89 89
MnCl2 85 83
NH4Cl 81 79
SnCl2 79 78
CoCl2 78 78
NiCl2 54 53
ZnCl2 41 40
Fish Physiol Biochem (2010) 36:573–586 581
123
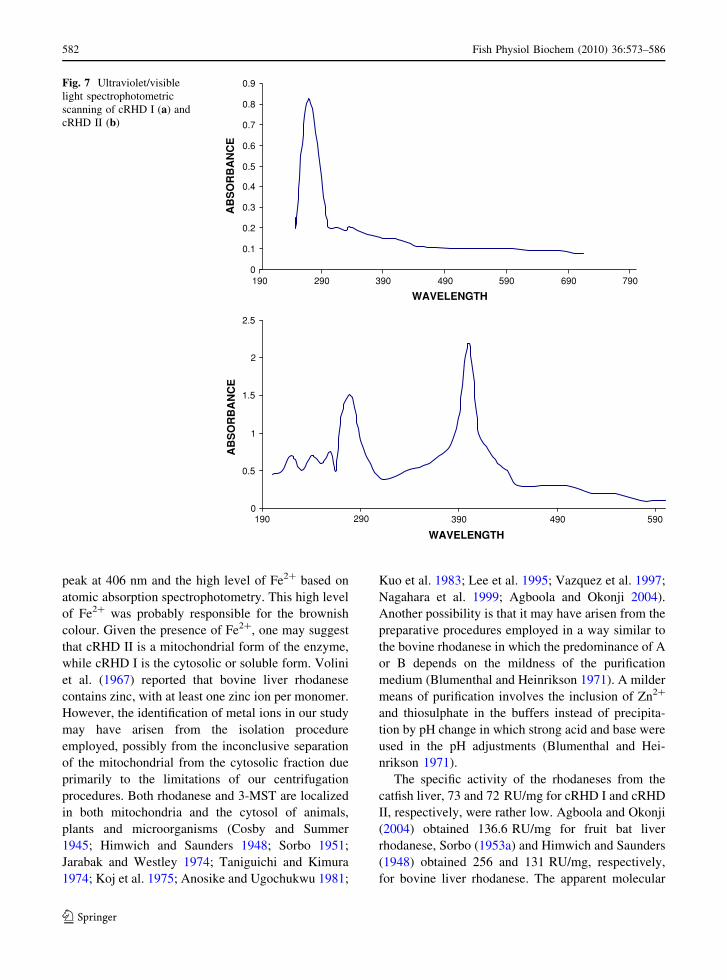
peak at 406 nm and the high level of Fe2? based on
atomic absorption spectrophotometry. This high level
of Fe2? was probably responsible for the brownish
colour. Given the presence of Fe2?, one may suggest
that cRHD II is a mitochondrial form of the enzyme,
while cRHD I is the cytosolic or soluble form. Volini
et al. (1967) reported that bovine liver rhodanese
contains zinc, with at least one zinc ion per monomer.
However, the identification of metal ions in our study
may have arisen from the isolation procedure
employed, possibly from the inconclusive separation
of the mitochondrial from the cytosolic fraction due
primarily to the limitations of our centrifugation
procedures. Both rhodanese and 3-MST are localized
in both mitochondria and the cytosol of animals,
plants and microorganisms (Cosby and Summer
1945; Himwich and Saunders 1948; Sorbo 1951;
Jarabak and Westley 1974; Taniguichi and Kimura
1974; Koj et al. 1975; Anosike and Ugochukwu 1981;
Kuo et al. 1983; Lee et al. 1995; Vazquez et al. 1997;
Nagahara et al. 1999; Agboola and Okonji 2004).
Another possibility is that it may have arisen from the
preparative procedures employed in a way similar to
the bovine rhodanese in which the predominance of A
or B depends on the mildness of the purification
medium (Blumenthal and Heinrikson 1971). A milder
means of purification involves the inclusion of Zn2?
and thiosulphate in the buffers instead of precipita-
tion by pH change in which strong acid and base were
used in the pH adjustments (Blumenthal and Hei-
nrikson 1971).
The specific activity of the rhodaneses from the
catfish liver, 73 and 72 RU/mg for cRHD I and cRHD
II, respectively, were rather low. Agboola and Okonji
(2004) obtained 136.6 RU/mg for fruit bat liver
rhodanese, Sorbo (1953a) and Himwich and Saunders
(1948) obtained 256 and 131 RU/mg, respectively,
for bovine liver rhodanese. The apparent molecular
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
190 290 390 490 590 690 790
WAVELENGTH
AB
SO
RB
AN
CE
0
0.5
1
1.5
2
2.5
WAVELENGTH
AB
SO
RB
AN
CE
190 290 390 490 590
Fig. 7 Ultraviolet/visible
light spectrophotometric
scanning of cRHD I (a) and
cRHD II (b)
582 Fish Physiol Biochem (2010) 36:573–586
123

weight obtained for cRHD I and cRHD II in our study
were 34,500 ± 707 and 36,800 ± 283 Da, respec-
tively, which is in line with reports that rhodanese has
molecular weight value approximately between
33,000 and 37,000 Da (Wang and Volini 1968;
Blumenthal and Heinrikson 1971; Nagahara et al.
1999). Various molecular weights have been obtained
for rhodanese from a variety of different sources:
bovine liver (37,000 Da; Sorbo 1953a, 1955), human
liver (37,000 Da; Jarabak and Westley 1974), mouse
liver (34,800 Da; Lee et al. 1995) and rat liver
(34,000 Da; Nagahara and Nishino 1996). The sub-
unit molecular weights determined by SDS–PAGE
were 33,200 ± 283 and 35,100 ± 141 Da for cRHD
I and cRHD II, respectively. These values are very
close to those obtained by Lee et al. (1995) and
Agboola and Okonji (2004), which were 34,000 and
35,700 Da for fruit bat and mouse rhodanese,
respectively. This result suggests that the enzyme is
a monomeric protein. While Jarabak and Westley
(1974) and Lee et al. (1995) showed that purified
mouse and human liver rhodaneses are monomeric
proteins, other researchers have shown that they are
dimers of two identical subunits of molecular weights
18,000–19,000 Da (Blumenthal and Heinrikson
1971; Volini et al. 1967). Volini et al. (1967)
reported that bovine rhodanese existed as a mono-
mer–dimeric molecule of molecular weight of
18,000–36,000. Russell et al. (1978) and Ploegman
et al. (1978), based on a three-dimensional structure
analysis, showed that bovine liver mitochondrial
rhodanese is a single polypeptide chain with a
molecular weight of 32,000 Da. Since both forms
(cRHD I and cRHD II) of rhodanese isolated in our
study had molecular weights of about 35,000, the
possibility that the two forms are monomeric and
aggregate forms of one of the other need not be
considered (Blumenthal and Heinrikson 1971). How-
ever, given the similar molecular weight for the two
forms, as also obtained for the two bovine rhodanes-
es, which also have other similar structural properties,
one can suggest that the cRHD I and cRHD II may
not be structurally different. Isoenzymes with struc-
tural similarity are not uncommon, as in venom
phospholipase A from Crotalus adamateus and
mushroom tyrosinases (Blumenthal and Heinrikson
1971).
The true Km values, as determined by the equation
of the bireactant enzyme ping-pong mechanism
(Florini and Vestling 1957; Cleland 1970; Segel
1975), for KCN and Na2S2O3 were 25.4 ± 1.70 and
18.6 ± 1.68 mM, respectively, for cRHD II. The
enzyme rhodanese catalyses the transfer of the outer
(sulphane) sulphur of thiosulphate to cyanide, thereby
forming the products thiocyanate and sulphite by way
of a double-displacement (ping-pong) mechanism in
which a covalent enzyme–sulphur (substituted
enzyme) intermediate is formed. Both of the initial
velocity patterns obtained in this experiment are
basically parallel lines (Fig. 4a, b). These patterns are
diagnostic of a double-displacement mechanism
(Cleland 1970; Segel 1975), and they resemble the
patterns observed with bovine liver rhodanese with
thiosulphate and cyanide as the varied substrates
(Schlesinger and Westley 1974) and the kinetics of
prokaryotic sulphurtransferases (Aird et al. 1987).
The ping-pong action is common in reactions
involving the transfer of a functional group between
two different molecules as in transaminases, transac-
ylases or transphosphorylases. Initial velocity studies
with bireactant mechanisms only serve to distinguish
the sequential mechanism from the ping-pong one
and to define certain kinetic constants. The parallel
lines in double-reciprocal plots in this experiment are
indicative of the ping-pong binary complex mecha-
nism. These values are compared to those reported in
earlier studies in Table 4. In Table 4, it can be seen
that the Km values of catfish liver rhodanese are
higher than those of mouse liver, bovine liver, human
liver and Acinetobacter rhodaneses, indicating that
the affinity of the catfish enzyme for these substrates
is less than that of the other enzymes and that it
would catalyze the detoxification reaction with less
efficiency. It should be noted, however, that the
results of the catfish (this work), bovine liver
(Schlesinger and Westley 1974), human liver (Jara-
bak and Westley 1974) and Acinetobacter (Aird et al.
1987) enzymes were determined from the double-
reciprocal plots and the appropriate secondary plots
and, therefore, are more comparable. This less
effective enzymatic system may be due to a lower
exposure to cyanide, in contrast to its mammalian
counterparts which are continually exposed through
their diet. However, it should be noted that cyanide
detoxification is a secondary benefit of rhodanese,
which has a number of major physiological roles,
including the supply of sulphide for the formation of
iron sulphur centres for the electron transport chain.
Fish Physiol Biochem (2010) 36:573–586 583
123

Different optimum temperatures have been
reported for rhodanese from different organisms. Ezzi
et al. (2003) obtained a wide temperature optimum of
35–55�C for rhodanese enzyme in some Trichoderma
strains. A value as high as 50� and 57–59�C was
obtained by Sorbo (1953b) and Chew and Boey
(1972) for bovine liver and tapioca leaves, respec-
tively. Himwich and Saunders (1948) also obtained an
optimum temperature of between 38� and 40�C for
bovine liver rhodanese. Agboola and Okonji (2004)
reported 35�C for the rhodanese in the cytosolic
fraction of fruit bat liver. The optimum temperature
obtained was 40�C for rhodanese from the liver of the
catfish from Asejire Lake. With the level of pollution
in this water, there will be various metabolic activities
going on in this lake which may result in the release of
heat. This ambient temperature may have conditioned
the enzyme to function at higher temperatures.
The Arrhenius plot of the effects of temperature on
the reaction rate consists of two linear segments with a
break occurring at 40�C. The apparent activation
energy values from these slopes are 7.3 and 72.9 kcal/
mol, respectively. The 7.3 kcal/mol obtained for
catfish liver rhodanese is close to the 7.5 kcal/mol
reported for bovine liver (Fruton and Simmonds 1963)
and also falls within the physiological activation
energy range (1–25 kcal/mol) for physiological pro-
cesses in living organisms (Fruton and Simmonds
1963; Whitaker 1972). The second value of 72.9
kcal/mol lies within the range (40–100 kcal/mol)
for protein denaturation (Whitaker 1972). The heat
stability experiment showed that the enzyme was
stable up to 50�C for about 30 min.
Pollutants in water include wide spectrum of
chemicals and pathogens which sometimes alters
the acidity, conductivity and temperature of the
water. The water sample collected from Asejire Lake
showed a pH of 5.7, 6.3 and 6.5 at different locations
of the Lake (Lameed and Obadara 2006). This is an
indication of acidic water. An optimum pH of 6.5 was
obtained for rhodanese from the liver of the catfish
from Asejire Lake.
Inhibition studies on the enzyme with a number of
chloride salts showed that the activity of the enzyme
was not affected by Mn2?, Co2? and Sn2? while
NH4?, Zn2? and Ni2? inhibited the enzyme consid-
erably. This is a result of the level of metal ions
present in the water as recorded by Lameed and
Obadara (2006). Metal ions showing a high potential
to inhibit enzyme activity are those that have a strong
affinity for ligands, such as phosphate, cysteinyl and
the histidyl side chains of proteins (Ulmer and Vallee
1972).
In conclusion, the physicochemical and kinetic
properties of rhodanese from the liver of African
catfish Clarias gariepinus are similar to those from
the livers of other animals. In particular, the kinetic
parameters obtained would allow the enzyme to
Table 4 Comparison of Km values of cRHD II with other rhodanese preparations
Substrate Km (mM)
Catfish
liveraFruit bat
liverbMouse
livercBovine
liverdBovine
livereHuman
liverfRat
livergAcinetobacter sulphane
sulphur transferaseh
KCN 25.4 13.36 12.5 19.0 0.06 9.5 NA 0.03
Na2S2O3 18.6 19.15 8.3 6.7 4.0 4.5 4.4 3.2
The Michaelis constant (Km) of KCN and Na2S2O3 of different mammalian liver rhodaneses in comparison with that from cat fish
liver
NA, Data not availablea This workb Agboola and Okonji (2004)c Lee et al. (1995)d Sorbo (1953a)e Schlesinger and Westley (1974)f Jarabak and Westley (1974)g Nagahara and Nishino (1996)h Aird et al. (1987)
584 Fish Physiol Biochem (2010) 36:573–586
123

function in the conversion of cyanide, from various
sources in the water, to thiocyanate and thereby
presumably in the detoxification of the poisonous
cyanide, ultimately improving the survival of the fish
in polluted water.
References
Agboola FK, Okonji RE (2004) Presence of rhodanese in the
cytosolic fraction of the fruit bat (Eidolon helvum) liver.
J Biochem Mol Biol 37(3):275–281
Agency for Toxic Substances and Disease Registry (ATSDR)
(1989) Toxicological profile for cyanide. ATSDR/TP-88/
12; PB90-162058. Prepared by Syracuse Research Cor-
poration for ATSDR, US Public Health Service, under
Contract No. 68-C8- 2004. ATSDR, Atlanta
Aird BA, Heinrikson RL, Westley J (1987) Isolation and
characterization of a prokaryotic sulphurtransferase.
J Biol Chem 262:17327–17335
Ali A, Al-Qarawi HM, Mousa BH (2001) Tissue and intra-
cellular distribution of rhodanese and mercaptopyruvate
sulphurtranferase in ruminants and birds. Vet Res 32:63–
70. doi:10.1051/vetres:2001110
Aminlari M, Malekhusseini A, Akrami F, Ebrahimnejad H
(2007) Cyanide-metabolizing enzyme rhodanese in
human tissues: comparison with domestic animals. Comp
Clin Pathol 16(1):47–51. doi:10.1007/s00580-006-0647-x
Anosike EO, Ugochukwu EN (1981) Characterization of
rhodanese from cassava leaves and tubers. J Exp Bot
32:1021–1027. doi:10.1093/jxb/32.5.1021
Blumenthal KM, Heinrikson RL (1971) Structural studies of
bovine liver rhodanese: I. Isolation and characterization of
two active forms of the enzymes. J Biol Chem 246:2430–
2437
Bordo D, Bork P (2002) The rhodanese/Cdc25 phosphatase
superfamily. Sequence–structure–function relations.
EMBO Rep 3:741–746
Bruton MN (1979) The food and feeding behaviour of Clariasgariepinus (Pisces: Clariidae) in Lake Sibaya, South
Africa, with emphasis on its role as a predator of cichlids.
Trans Zool Soc Lond 35(1):47–114
Chew MY, Boey CG (1972) Rhodanese of tapioca leaf. Phy-
tochemistry 11:167–169. doi:10.1016/S0031-9422(00)899
83-5
Clay D (1979) Sexual maturity and fecundity of the African
catfish (Clarias gariepinus) with an observation on the
spawning behaviour of the Nile catfish (Clarias lazera).
Zool J Linn Soc 65:351–365. doi:10.1111/j.1096-3642.
1979.tb01100.x
Cleland WW (1970) Steady state kinetics. In: Boyer PB (ed)
The enzymes, vol II, 3rd edn. Academic Press, London,
pp 1–65
Cosby EQ, Summer JB (1945) Rhodanese. Arch Biochem
7:457–460
Eisler R (1991) Cyanide hazards to fish, wildlife, and inver-
tebrates: a synoptic review. U.S. Fish Wildl Serv Biol Rep
85(1.23):1–55
Environmental Protection Agency (EPA) (1980) Ambient
water quality criteria for cyanides. U.S. EPA Rep 440/5-
80-037. EPA, Washington D.C.
Ezzi MI, Pascual JA, Gould BJ, Lynch JM (2003) Character-
isation of the rhodanese enzyme in Trichoderma spp.
Enzyme Microb Technol 32(5):629–634. doi:10.1016/
S0141-0229(03)00021-8
Florini JR, Vestling CS (1957) Graphical determination of the
dissociation constants for two substrate enzyme systems.
Biochim Biophys Acta 25:575–578. doi:10.1016/0006-
3002(57)90529-2
Fruton JS, Simmonds S (eds) (1963) Kinetics of enzyme. In:
Fruton JS, Simmonds S (eds) General biochemistry, 2nd
edn, Wiley, New York, pp 244–283
Gornall AG, Bardawill CJ, David MM (1949) Determination of
serum protein by Biuret reaction. J Biol Chem 117:751–
766
Himwich WA, Saunders JB (1948) Enzymic conversion of
cyanide to thiocyanate. Am J Physiol 53:348–354
Holden AV, Marsden K (1964) Cyanide in salmon and brown
trout. Department of Agriculture and Fisheries of Scot-
land. Freshw Salmon Fish Res Ser 33. Department of
Agriculture and Fisheries of Scotland, Edinburgh
Horowitz PM, DeToma F (1970) Improved preparation of
bovine liver rhodanese. J Biol Chem 245(6):984–985
Jarabak R, Westley J (1974) Human liver rhodanese: nonlinear
kinetic behaviour. Double displacement mechanism. Bio-
chemistry 13(16):3233–3236. doi:10.1021/bi00713a006
Kaur M, Singh K, Rup PJ, Kamboj SS, Saxena AK, Sharma M,
Bhagat M, Sood SK, Singh J (2006) A tuber lectin from
Arisaema jacquemontii Blume with anti-insect and anti-
proliferative properties. J Biochem Mol Biol 39(4):432–
440
Keilin D (1929) Cytochrome and respiratory enzymes. Proc R
Soc Lond (Biol Sci) 104:206–251. doi:10.1098/rspb.1929.
0009
Koj A (1968) Enzymic reduction of thiosulphate in prepara-
tions from beef liver. Acta Biochim Pol 15(2):161–169
Koj A, Frendo J, Wojtczak L (1975) Subcellular distribution
and intramitochondrial localization of the three sulphu-
rtransferases in rat liver. FEBS Lett 57:42–46
Kuo SM, Lea TC, Stiqanuk MH (1983) Developmental pattern,
tissue distribution and subcellular distribution of cysteine:
a-ketoglutarateaminotransferase and 3-mercaptopyruvate
sulphurtransferases activities in the rat. Biol Neonate43:23–32
Lameed GA, Obadara PG (2006) Eco-development impact of
coca-cola industry on biodiversity resources at Asejire
area, Ibadan; Nigeria. J Fish Int 1(4):55–62
Leduc G (1978) Deleterious effects of cyanide on early life
stages of Atlantic salmon (Salmo salar). J Fish Res Board
Can 35:166–174
Leduc G, Pierce RC, McCracken IR (1982) The effects of
cyanides on aquatic organisms with emphasis upon
freshwater fishes. National Research Council of Canada
(NRCC) Publ 19246. NRCC/CNRC, Ottawa
Lee CH, Hwang JH, Lee YS, Cho KS (1995) Purification and
characterization of mouse liver rhodanese. J Biochem Mol
Biol 28:170–176
Lieske CN, Clark CR, Zoeffel LD (1996) Temperature effects
in cyanolysis using elemental sulphur. J Appl Toxicol
Fish Physiol Biochem (2010) 36:573–586 585
123

16:171–175. doi:10.1002/(SICI)1099-1263(199603)16:2
\171::AID-JAT327[3.0.CO;2-R
Micha JC (1973) Etude des populations piscicoles de l’Uban-
gui et tentative de selection et d’adaptation de quelques
especes a l’etang de pisciculture. Centre Technique
Forestier Tropical, Nogent-sur-Marne
Montgomery RD (1965) The medical significance of cyano-
gens in plant foodstuffs. Am J Clin Nutr 17:103–113
Nagahara N, Nishino T (1996) Role of amino acid residues in
the active site of rat liver mercaptopyruvate sulphu-
rtransferases. J Biol Chem 271:27395–27401. doi:10.1074/
jbc.271.44.27395
Nagahara N, Okazaki T, Nishino T (1995) Cytosolic mercap-
topyruvate sulphurtransferase is evolutionarily related to
mitochondrial rhodanese. Striking similarity in active site,
amino acid sequence and the increase in mercaptopyru-
vate sulphurtransferase activity of rhodanese by site
directed mutagenesis. J Biol Chem 270:16230–16235.
doi:10.1074/jbc.270.27.16230
Nagahara N, Ito T, Minam M (1999) Mercaptopyruvate sul-
phurtransferase as a defence against cyanide toxications;
molecular properties and mode of detoxification. Histol
Histopathol 14:1277–1286
Ogata K, Xing D, Volini M (1989) Bovine mitochondrial
rhodanese is a phosphoprotein. J Biol Chem 246(5):2718–
2725
Ploegman JH, Drent G, Kalk KH, Hol WG (1978) Structure of
bovine liver rhodanese. I. Structure determination at 2.5 A
resolution and a comparison of the conformation and
sequence of its two domains. J Mol Biol 123:557–594.
doi:10.1016/0022-2836(78)90207-3
Russell J, Weng L, Kein PS, Heinrikson RL (1978) The
covalent structure of bovine liver rhodanese. J Biol Chem
253:8102–8108
Schlesinger P, Westley J (1974) An expanded mechanism for
rhodanese catalysis. J Biol Chem 249:780–788
Segel IH (ed) (1975) Enzymes. In: Segel IH (ed) Biochemical
calculations; 2nd edn. Wiley, New York, pp 278–281
Smith LL, Broderius SJ, Oseid DM, Kimball GL, Koenst WM
(1978) Acute toxicity of hydrogen cyanide to freshwater
fishes. Arch Environ Contam Toxicol 7:325–337
Smith LL, Broderius SJ Jr, Oseid DM, Kimball GL, Koenst
WM, Lind DT (1979) Acute and chronic toxicity of HCN
to fish and invertebrates. U.S. Environ. Prot. Agency Rep.
600/3-79-009, 129 pp
Smith J, Urbanska KM (1986) Rhodanese activity in Lotuscorniculatus sensu-lato. J Nat Hist 20(6):1467–1476. doi:
10.1080/00222938600770991
Sorbo BH (1951) On the properties of rhodanese. Acta Chem
Scand 5:724–726. doi:10.3891/acta.chem.scand.05-0724
Sorbo BH (1953a) Crystalline rhodanese. Enzyme catalyzed
reaction. Acta Chem Scand 7:1137–1145. doi:10.3891/
acta.chem.scand.07-1137
Sorbo BH (1953b) Crystalline rhodanese. Acta Chem Scand
7:1129–1136. doi:10.3891/acta.chem.scand.07-1129
Sorbo BH (1955) Rhodanese. In: Sidney PL, Kaplan NO (eds)
Methods of enzymology, vol 2. Academic Press, New
York, pp 334–337
Sorbo BH (1957) Enzyme transfer of sulphur from mercapto-
pyruvate to sulphate or sulphonates. Biochim Biophys
Acta 24:324–329. doi:10.1016/0006-3002(57)90201-9
Taniguichi T, Kimura T (1974) Role of 3-mercaptopyruvate
sulphurtransferase in the formation of the iron chromo-
phore of adrenal ferredoxin. Biochim Biophys Acta
364:284–295
Tolba MK (1982) Development without destruction. Evolving
environmental perceptions. Dublin, Tycooly. Nat Resour
Environ Ser 12:197
Towill LE, Drury JS, Whitfield BL, Lewis EB, Galyan EL,
Hammons AS (1978) Reviews of the environmental
effects of pollutants vs. cyanide. U.S. Environmental
Protection Agency (EPA) Rep 600/1-78-027. EPA,
Washington D.C.
Ulmer DD, Vallee BL (1972) Role of metals in sulphu-
rtranferases activity. Annu Rev Biochem 32:86–90
Vazquez E, Gazzaniga S, Polo C, Batlle A (1997) Mitochon-
drial and cytosolic rhodanese from liver of DAB-treated
mice. III. Inhibition kinetic studies. Cancer Biochem
Biophys 15(4):285–293
Villarejo M, Westley J (1963) Mechanism of rhodanese
catalysis of thiosulphate oxidation-relation. J Biol Chem
238:4016–4060
Volini M, DeToma F, Westley J (1967) Dimeric structure and
zinc content of bovine liver rhodanese. J Biol Chem
242:5220–5225
Wang SF, Volini M (1968) Studies on the active site of rho-
danese. J Biol Chem 243:5465–5470
Warburg O (1911) Inhibition of the action of prussic acid in
living cells. Hoppe Seylers Z Physiol Chem 76:331–346
Westley J (ed) (1980) Rhodanese and the sulphane pool. In:
Jakoby WB (ed) Enzymatic basis of detoxification, vol 2.
Academic Press, New York, pp 245–259
Whitaker JR (1972) Effect of temperature on enzyme catalysed
reaction. In: Whitaker JR (ed) Principles of enzymology
for the food science. Marcel Dekker, New York, pp 319–
348
Wokes F, Willimott SG (1951) The determination of cyanide in
seed. J Pharm Pharmacol 3:905–916
586 Fish Physiol Biochem (2010) 36:573–586
123