persistence of escherichia coli on injured vegetable plants
TRANSCRIPT

International Journal of Food Microbiology 138 (2010) 232–237
Contents lists available at ScienceDirect
International Journal of Food Microbiology
j ourna l homepage: www.e lsev ie r.com/ locate / i j foodmicro
Persistence of Escherichia coli on injured vegetable plants
Dean Harapas a, Robert Premier b, Bruce Tomkins c, Peter Franz c, Said Ajlouni a,⁎a Melbourne School of Land and Environment, The University of Melbourne, Parkville, Victoria 3010, Australiab Salad Fresh, 75 Northcorp Boulevard, Broadmeadows, Victoria 3047, Australiac Department of Primary Industries, 621 Burwood Highway, Knoxfield, Victoria 3180, Australia
⁎ Corresponding author. Department of AgricultureSchool of Land and Environment, The University of MelbAustralia. Tel.: +61 3 83448620; fax: +61 3 83445037
E-mail address: [email protected] (S. Ajlouni).
0168-1605/$ – see front matter © 2010 Elsevier B.V. Adoi:10.1016/j.ijfoodmicro.2010.01.022
a b s t r a c t
a r t i c l e i n f oArticle history:Received 18 May 2009Received in revised form 8 December 2009Accepted 16 January 2010
Keywords:Escherichia coliPersistenceInjuredVegetableCeleryCos lettuceChiveFoodborne illnessGlasshouseField
Minor shoot injury to glasshouse celery, Cos lettuce and chive plants significantly increased the persistenceof applied Escherichia coli (Pb0.05). After 1 week, mean counts of about 5 log10 CFU/g decreased to fewerthan 0.5 log10 CFU/g on the uninjured plants, compared to 4 log10 CFU/g or more on injured plants. By theend of the 3-week long experiments, counts from the uninjured plants were 0.21 log10 CFU/g or fewer, but2.8, 2.3 and 5.1 log10 CFU/g on injured Cos lettuce, celery and chive plants, respectively. A field experimentusing Cos lettuce also showed that shoot injury increased E. coli persistence. Counts from the injured plants ondays 1, 3, and 7 were, 4.2, 4.1 and 3.3 log10 CFU/g, respectively, whereas the uninjured plants returnedsignificantly (Pb0.05) lower counts on those days, and were 2.8, 2.0 and 1.2 log10 CFU/g, respectively. Thesefindings reveal that increased E. colipersistence on injured tissue is common todifferent vegetables and can occurin the glasshouse and the field. The implications of this study on vegetable production practices are presented.
and Food Systems, Melbourneourne, Parkville, Victoria 3010,.
ll rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
Outbreaks of foodborne illnesses from fresh vegetables contami-nated with bacterial pathogens have been increasing in developednations (Beuchat, 1998; De Roever, 1998; Hoyle, 2008; Lynch et al.,2009), and have caused considerable mortality, morbidity and financiallosses. For example in 2008, Salmonella contamination of peppers andperhaps tomatoes infected 1442 and hospitalised 273 people, and mayhave contributed to two deaths across 43 states of the USA (CDC, 2008).Losses to the US tomato industry were estimated to be over US$100 million (Thompson, 2008). The actual number and extent ofdisease outbreaks from fresh vegetables is probably greater than theliterature indicates, due to the low level of reporting by victims and theavailability of resources to determine the responsible food(s) (Arnoldand Munce, 1997).
Vegetables may become contaminated at any time during theirproduction, harvesting, processing, distribution and preparation forconsumption (Beuchat, 1998). Pre-harvest contamination is consideredlikely to account for many outbreaks, as it is extremely difficult toprevent. Pathogens can arrive via animal manure used for fertilisation
and soil conditioning, faecally tainted irrigation water, wild anddomestic animals, and floodwaters from a contaminated site such as acattle stock yard or sewage treatment facility (Beuchat, 1998; Brackett,1999). Additional sources of human pathogens could includemanure ormanure contaminated soil particles blown onto crops, and poor hygieneof personnel entering the field or glasshouse. Faecally contaminatedfarm equipment, insects or even garden slugs (Sproston et al., 2006)may also transfer human pathogens to crops.
Pre-harvest contamination was demonstrated by Natvig et al.(2002), who in a growth cabinet study isolated Salmonella entericaserovar Typhimurium from radishes and arugula which had grown insoil spiked with this pathogen and manure. Moreover, strong evidenceof pre-harvest contamination was found for a 2006 outbreak caused byEscherichia coliO157:H7 contamination of spinach. E. coliwith the samepulsed-field gel electrophoresis pattern as the outbreak strain wasisolated fromriverwater, cattle andwildpigs close toafield identifiedasa source of contaminated spinach. During this outbreak, at least 205people were infected and three died (CFERT, 2007).
Reducing pre-harvest contamination is critical for improving thesafety of fresh plant produce, because previous studies have foundthat washing and sanitisation of vegetables may not remove allcontaminating bacteria (Behrsing et al., 2000; Han et al., 2000; Seoand Frank, 1999; Solomon et al., 2002; Takeuchi and Frank, 2000).With this in mind, the aim of this study was to help validate practicesfor reducing the risk of foodborne illness from fresh vegetables

Table 1Season, temperature and relative humidity (RH) ranges for the glasshouse experiments.
Vegetable Season Temperature (°C) range RH (%) range
Minimum Maximum Minimum Maximum
Celery Late spring 11–19 29–45 19–35 82–100Cos lettuce Autumn 9–14 24–36 21–45 100Chive Winter 13–16 21–28 21–50 75–88
233D. Harapas et al. / International Journal of Food Microbiology 138 (2010) 232–237
contaminated prior to harvest with human bacterial pathogens. UsingE. coli as a model human bacterial pathogen, it was hypothesised thatpreventing shoot injury would decrease the persistence of contam-ination by this bacterium in the glasshouse and field. The reason beingthat the injured tissue would shelter the E. coli from desiccation andUV light, factors that cause the rapid decline of enteric bacteria onplants (Bell, 1976; Brandl and Mandrell, 2002; Cifuentes et al., 2001;O'Brien and Lindow, 1989). This study examined the populationbehaviour of E. coli on injured and uninjured vegetable plants in aglasshouse and in an open field. The proposed null hypothesis was:‘injury to vegetable plant shoots can increase the persistence ofcontaminating enteric bacterial pathogens’.
2. Materials and methods
2.1. E. coli suspension preparation
A single strain of E. coliwas used for the glasshouse experiments andanother for the field experiment. These were provided by The School ofMicrobiology of the University of Melbourne (Australia), and FoodScience Australia, respectively. Both strainswere isolated from humans,and are not known to be pathogenic. The bacteria were thawed fromfrozen stock cultures and incubated overnight at 37 °C, after streakingonto tryptone soya agar plates made from tryptone soya broth (OxoidLtd., Basingstoke, England) and 2% bacteriological agar no.1 (Oxoid).Single colonies were then subcultured onto fresh plates and the growthchecked for purity. For the glasshouse experiments, growth from oneplate was suspended in 4 ml of 0.1% peptone (Oxoid), and then dilutedin 16 l of autoclaved reverse-osmosis water. Whilst for the fieldexperiment, growth from two plates was diluted in double the waterquantity. Blank suspensionswere prepared as per the E. coli suspensionsbut with uninoculated tryptone soya agar (Oxoid) plates. The blanksuspensions were applied to the non-E. coli treated plants. The con-centrationof E. coliused for theglasshouseexperimentwas7.5–7.6 log10CFU/ml and that for the field experiment was 8.0 log10 CFU/ml. Thesewere determined by inoculation onto PetrifilmTM E. coli/coliform countplates (3MMicrobiology, Saint Paul, USA) as described in Section 2.5. Nocolonies grew from the blank suspensions.
2.2. Plant production
Plants were obtained as seedlings at approximately 6 weeks ofage. A local grower provided the Celery (Apium graveolens) cultivars‘Endevor’ and ‘Eltoro’. The Cos or romaine lettuce (Lactuca sativa L. var.longifolia Lam.) cultivar ‘Junior’ was purchased from LeppingtonSpeedy® Seedlings and Supplies Pty. Ltd., NSW, and the green chive(Allium schoenoprasum) was purchased from Floriana Pty. Ltd., Victoria.
For the glasshouse experiments, single plantswere transplanted into10 cm pots filled with a steamed commercial bark/sand potting mixsupplementedwith slow release fertiliser. Overhead sprayerswere usedfor at least 2 weeksuntil 1 day before the start of the experiments,wheneach pot was irrigated via a dripper inserted into the potting mix.Tarpaulinswere deployed underneath the glasshouse benches to collectrunoff for disinfection at the endof the experiments. The temperature inthe glasshouses was regulated by thermostat controlled natural gasheaters and evaporative coolers. The temperature and relative humidityranges during the experiments are summarised in Table 1, which weremeasured using Hobo® Data loggers, (Onset Computer Corp., Bourne,USA).
The field was at the Department of Primary Industries, Knoxfield,Victoria, Australia, approximately 25 km east of Melbourne and wasfallowprior to the incorporationof burnt lime andmineral fertiliser, andthe formation of beds about 20 cm high and 100 cm wide. Cos lettuceseedlingswere transplanted in apatternasdescribed in thenext section.The following pesticides were applied as recommended by their manu-facturers': snail and slug pellets ‘DefenderTM’ (15 g/kg metaldehyde,
Scotts, Australia Pty. Ltd.) just after transplantation, the selectiveherbicide ‘Kerb’ (500 g/kg propyzamide, Rhom and Hass Australia Pty.Ltd.) 2 days post transplantation, and the fungicide ‘ManKocide® DF’(300 g/kg copper and 150 g/kg Mancozeb, Griffin Corporation AustraliaPty. Ltd.) and the insecticide ‘Malathion’ (500 g/l MalathionTM, ArthurYates and Co. Ltd.) 10 days post transplantation. E. coli was applied19 days after transplantation. Irrigation started from 2 days after thisand was performed as required four times in the first week and then atleast one time per week with overhead sprinklers using water from anearby dam, or a hose using tap water. The experiment was conductedin late spring and early summer. Rainfall and temperature data wascollected by theAustralianGovernmentBureauofMeteorologyweatherstation, about 200 m from the field. The minimum and maximumtemperature rangeswere 7 to 9, and 19 to 34 °C, respectively. Therewasa total of 58 mmrainfall during the experiment, 1 mm falling on the dayof E. coli application, a further 3 mm during the first week, and most ofthe balance fell between days 8 to 23.
2.3. Experimental designs
In the glasshouse, an experimental unit consisted of one plant perpot, and a separate experiment was performed for each vegetable(celery, Cos lettuce and chive). Plants were tested for E. coli CFUalmost immediately after the treatment applications and then 1, 3, 7,14 and 21 days later. The treatments were dosed (with E. coli) andinjured, dosed and uninjured, not dosed and injured, and not dosedand uninjured. Replicates of six experimental units were used foreach of the above treatments and times of testing (144 plants perexperiment). Plants were arranged in blocks of 6 by 6 pots, and thetreatments (including times of testing) were applied in a latinised andresolvable manner. Gaps created as pots were removed during theexperiment were re-filled by repositioning the remaining potted plantsto avoid any edge effect.
For the field experiment, Cos lettuce seedlings were planted in16 plots, arranged in a 4 by 4 grid. Each plot was formed from 3 rowsand 31 columns of plants, which were spaced 15 cm apart in the rowsand 10 cm apart in the columns. The same injury and dosing treat-ments as applied to the glasshouse experiments were randomlyapplied to the plots. These treatments were replicated four times andlatinised. Plots were separated by a clear zone of about 3 m to preventcross contamination. The plots were divided into experimental units(subplots) of six plants arranged in three rows and two columns. Theexperimental units were randomly selected for E. coli CFU enumerationat 1, 3, 7, 16, 23 and 37 days post E. coli application. Each experimentalunit was positioned between non-tested experimental units to preventany edge effect.
Latinisationwasperformedusing theCycDesigN software (Version 2,by Department of Statistics, University of Waikatoi, Halmiltion, NewZealand and CSIRO Forestry and Forest Products, Canberra, Australia).
2.4. Injury and E. coli application
Tissue injury was performed just prior to treatment with thebacterial culture. For the glasshouse experiments, the bases of the threelargest and central Cos lettuce and chive leaves earmarked for treatmentand subsequent testing were ‘dotted’ with a permanent marker. That
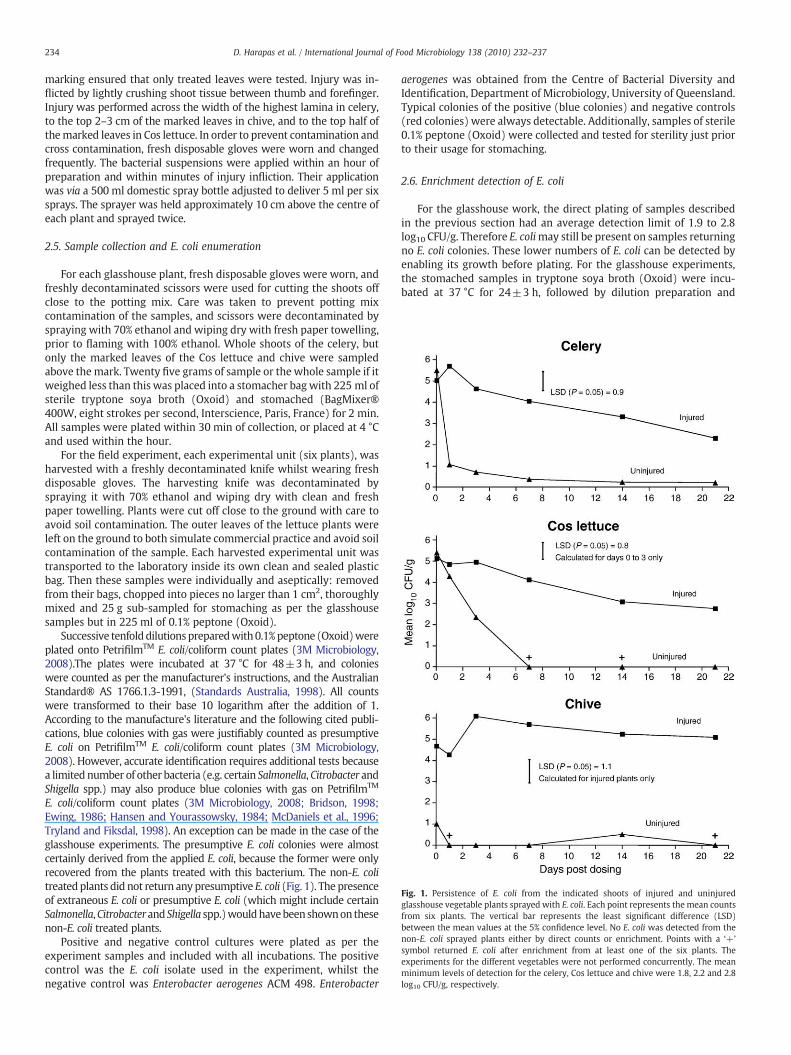
Fig. 1. Persistence of E. coli from the indicated shoots of injured and uninjuredglasshouse vegetable plants sprayed with E. coli. Each point represents the mean countsfrom six plants. The vertical bar represents the least significant difference (LSD)between the mean values at the 5% confidence level. No E. coli was detected from thenon-E. coli sprayed plants either by direct counts or enrichment. Points with a ‘+’
symbol returned E. coli after enrichment from at least one of the six plants. Theexperiments for the different vegetables were not performed concurrently. The meanminimum levels of detection for the celery, Cos lettuce and chive were 1.8, 2.2 and 2.8log10 CFU/g, respectively.
234 D. Harapas et al. / International Journal of Food Microbiology 138 (2010) 232–237
marking ensured that only treated leaves were tested. Injury was in-flicted by lightly crushing shoot tissue between thumb and forefinger.Injury was performed across the width of the highest lamina in celery,to the top 2–3 cm of the marked leaves in chive, and to the top half ofthemarked leaves in Cos lettuce. In order to prevent contamination andcross contamination, fresh disposable gloves were worn and changedfrequently. The bacterial suspensions were applied within an hour ofpreparation and within minutes of injury infliction. Their applicationwas via a 500 ml domestic spray bottle adjusted to deliver 5 ml per sixsprays. The sprayer was held approximately 10 cm above the centre ofeach plant and sprayed twice.
2.5. Sample collection and E. coli enumeration
For each glasshouse plant, fresh disposable gloves were worn, andfreshly decontaminated scissors were used for cutting the shoots offclose to the potting mix. Care was taken to prevent potting mixcontamination of the samples, and scissors were decontaminated byspraying with 70% ethanol and wiping dry with fresh paper towelling,prior to flaming with 100% ethanol. Whole shoots of the celery, butonly the marked leaves of the Cos lettuce and chive were sampledabove the mark. Twenty five grams of sample or the whole sample if itweighed less than this was placed into a stomacher bagwith 225 ml ofsterile tryptone soya broth (Oxoid) and stomached (BagMixer®400W, eight strokes per second, Interscience, Paris, France) for 2 min.All samples were plated within 30 min of collection, or placed at 4 °Cand used within the hour.
For the field experiment, each experimental unit (six plants), washarvested with a freshly decontaminated knife whilst wearing freshdisposable gloves. The harvesting knife was decontaminated byspraying it with 70% ethanol and wiping dry with clean and freshpaper towelling. Plants were cut off close to the ground with care toavoid soil contamination. The outer leaves of the lettuce plants wereleft on the ground to both simulate commercial practice and avoid soilcontamination of the sample. Each harvested experimental unit wastransported to the laboratory inside its own clean and sealed plasticbag. Then these samples were individually and aseptically: removedfrom their bags, chopped into pieces no larger than 1 cm2, thoroughlymixed and 25 g sub-sampled for stomaching as per the glasshousesamples but in 225 ml of 0.1% peptone (Oxoid).
Successive tenfolddilutionspreparedwith0.1%peptone (Oxoid)wereplated onto PetrifilmTM E. coli/coliform count plates (3M Microbiology,2008).The plates were incubated at 37 °C for 48±3 h, and colonieswere counted as per the manufacturer's instructions, and the AustralianStandard® AS 1766.1.3-1991, (Standards Australia, 1998). All countswere transformed to their base 10 logarithm after the addition of 1.According to the manufacture's literature and the following cited publi-cations, blue colonies with gas were justifiably counted as presumptiveE. coli on PetrifilmTM E. coli/coliform count plates (3M Microbiology,2008). However, accurate identification requires additional tests becausea limited number of other bacteria (e.g. certain Salmonella, Citrobacter andShigella spp.) may also produce blue colonies with gas on PetrifilmTM
E. coli/coliform count plates (3M Microbiology, 2008; Bridson, 1998;Ewing, 1986; Hansen and Yourassowsky, 1984; McDaniels et al., 1996;Tryland and Fiksdal, 1998). An exception can be made in the case of theglasshouse experiments. The presumptive E. coli colonies were almostcertainly derived from the applied E. coli, because the former were onlyrecovered from the plants treated with this bacterium. The non-E. colitreatedplants did not return anypresumptive E. coli (Fig. 1). The presenceof extraneous E. coli or presumptive E. coli (which might include certainSalmonella,Citrobacterand Shigella spp.)wouldhavebeenshownon thesenon-E. coli treated plants.
Positive and negative control cultures were plated as per theexperiment samples and included with all incubations. The positivecontrol was the E. coli isolate used in the experiment, whilst thenegative control was Enterobacter aerogenes ACM 498. Enterobacter
aerogenes was obtained from the Centre of Bacterial Diversity andIdentification, Department of Microbiology, University of Queensland.Typical colonies of the positive (blue colonies) and negative controls(red colonies) were always detectable. Additionally, samples of sterile0.1% peptone (Oxoid) were collected and tested for sterility just priorto their usage for stomaching.
2.6. Enrichment detection of E. coli
For the glasshouse work, the direct plating of samples describedin the previous section had an average detection limit of 1.9 to 2.8log10 CFU/g. Therefore E. colimay still be present on samples returningno E. coli colonies. These lower numbers of E. coli can be detected byenabling its growth before plating. For the glasshouse experiments,the stomached samples in tryptone soya broth (Oxoid) were incu-bated at 37 °C for 24±3 h, followed by dilution preparation and

Fig. 2. Persistence of presumptive E. coli colony from shoots of injured and uninjuredfield grown Cos lettuce dosed with E. coli. Each point represents the mean counts fromfour subplots. The vertical bar represents the least significant difference between themean values (LSD) at the 5% level confidence level. The minimum level of detection was1 log10 CFU/g.
235D. Harapas et al. / International Journal of Food Microbiology 138 (2010) 232–237
plating onto PetrifilmTM E. coli/coliform count plates (3MMicrobiology,2008) and examined for E. coli as described in the previous section.
2.7. Statistical analysis
Analysis of variance and the least significance difference betweenthe means (LSD) at the 95% confidence level was calculated using‘GenStat’ software (Fifth Edition, Release 4.2, Copyright 2000 and theSixth Edition, Version-6.1.0.200, Copyright 2002, Lawes AgriculturalTrust, Supplied by VSN International Ltd, Wilkinson House, JordanHill Road, Oxford OX2 8 DR, UK). Means which differed by thecalculated LSD or more were considered statistically significant atα=0.05.
3. Results
Injured celery plant shoots returned significantly (Pb0.05) higherE. coli counts when compared to uninjured celery plants from day 1onwards (Fig. 1). When the bacteria were applied to the uninjuredcelery, their mean colony count declined by more than 99.99%,from 5.5±0.72 to 1.1±1.2 log10 CFU/g within 24 h and remainedlow (b1 log10 CFU/g) for the entire experiment. However, countsfrom the injured celery did not significantly (PN0.05) decrease untilafter day 3, and then declined more slowly and remainedsignificantly higher (Pb0.05) than the corresponding counts fromthe uninjured plants. E. coli counts on injured and uninjured celeryplants at 21 days were 2.31±1.24 and 0.21±0.52 log10 CFU/g,respectively (Fig. 1).
To determine if this increased persistence phenomenon wasspecific only to celery, further glasshouse experiments were per-formed with different vegetable species. Counts from Cos lettuce alsoshowed a clear shoot injury effect (Fig. 1). The initial mean count of5.39±0.28 log10 CFU/g on uninjured Cos lettuce dropped significantly(Pb0.05) by more than 2 logs after 3 days post E. coli applicationand was below the level of detection from 7 days onwards (Fig. 1).On the other hand the initial E. coli counts (5.12±0.66 log10 CFU/g)on injured Cos lettuce declined by less than 1 log10 CFU/g 3 daysafter dosing. Moreover, the injured Cos lettuce still returned countsof 2.8 log10 CFU/g or more from day 7 onwards, whereas there wereno counts from the uninjured plants. The initial mean E. coli count4.7±0.64 log10 CFU/g from injured chive plants increased signifi-cantly (Pb0.05) and reached 6.1 log10 CFU/g 3 days after dosing. Thecount on injured chive did not show any significant (PN0.05) fallafter the initial rise, and the final recorded count was 5.1±1.1 log10CFU/g. The day zero mean count on the uninjured chive (1.0±1.56log10 CFU/g) was much lower than that of the injured plants (4.7±0.64 log10 CFU/g). The counts from the uninjured chive remained atb1.0 log10 CFU/g for the rest of the experiment (Fig. 1).
Although uninjured Cos lettuce from days 7 and 21 and uninjuredchive from days 1 and 21 showed no growth of E coli by direct plating,some of these samples showed detectable E coli counts afterenrichment. However, no E. coli could be detected by direct countsor enrichment from the uninjured Cos lettuce from day 21, and theuninjured chive from days 3 and 7 (Fig. 1).
The field experiment was undertaken to demonstrate that theinjury effect was not restricted to the glasshouse. Similar to theglasshouse experiment, injury of the Cos lettuce tissue increased thepersistence of E. coli (presumptive) in the field (Fig. 2). Counts fromboth injured and uninjured plants declined gradually over the courseof the experiment. The initial presumptive E. coli counts from theinjured and uninjured Cos lettuce were 4.16±0.78 and 2.82±0.62log10 CFU/g, respectively. The rate of bacterial loss was faster for theuninjured plants, which dropped over 2 logs (99%) after the first3 days. During the same time, there was less than a 0.5 log decline incounts from the injured plants, and a 2 log drop did not happen untilday 23 (Fig. 2).
4. Discussion
With the plant production systems used in this study, the resultsprove the hypothesis that preventing shoot injury decreases thepersistence of E. coli in the glasshouse and field.
Although E. coli was only used here, continuous research in ourlaboratory is currently investigating the behaviour of bacteria fromother pathogenic genera. It is anticipated that all vegetative bacteriaentering a recent shoot injury would probably persist longer bygaining protection from UV light and desiccation, and obtainingsustenance from the released nutrients. Injury may also stimulatebiofilm formation which would further protect bacteria from theenvironment. Two growth cabinet studies suggest pathogenicbacteria would have an increased persistence in response to shootinjury in the glasshouse or field. Campylobacter jejuni was found todecline more slowly on spinach plants when their leaves wereinjured, held at 10 °C and sprayed with water every 15 min (Brandlet al., 2004), and E. coli O157:H7 also declined more slowly on injuredCos lettuce plants held under more typical production conditions of25 °C daytime and 20 °C night-time temperatures and relativehumidity of 55% (Aruscavage et al., 2008). In the latter study, injurycaused by disease from the plant pathogen Xanthomonas campestrispv. vitians also increased the persistence E. coli O157:H7.
The extent of persistence would almost certainly vary dependingon the species or even strain of bacterium. Those that have a greatertolerance to desiccation would be expected to better survive oninjured and uninjured shoots under dry conditions, whilst those withgreater adherence could persist longer as they are less likely to bewashed away by rain and irrigation. For example, Salmonella entericaserovar Typhimurium was found to better resist desiccation onanhydrous silica gel, than a strain of E. coli (Janning et al., 1994), and aselection of S. enterica and E. coli strains were found to adhere morestrongly to alfalfa sprouts than E. coli O157: H7 (Barak et al., 2002).
The increased survival of E. coli induced by shoot injury on threedifferent injured vegetable plants (celery, Cos lettuce and chive)suggests this phenomenon is common to most kinds of vegetables.However, the degree of persistence may vary with plant species. Forinstance the remarkable non-declining persistence and even growthof E. coli on injured chive (Fig. 1), may have been caused by the plant'shollow shoots providing better protection from the environment thanthe shoots of the other vegetables. Whereas, the hydrophobic shootsof the uninjured chive plants were probably responsible for thistreatment's very low counts even from day zero (Fig. 1). The E. colisuspension was observed to bead and run off the shoots, and therebyreducing the initial numbers of this bacterium. Such run off was notobserved in the uninjured Cos lettuce and celery, and their initial

236 D. Harapas et al. / International Journal of Food Microbiology 138 (2010) 232–237
counts were similar to each other at about 5.4 log10 CFU/g, and muchhigher than the uninjured chive (Fig. 1). Therefore hydrophobicityprobably does not explain the more rapid loss of E. coli CFU on thecelery compared to the Cos. The reason for this difference may berelated to the more open structure of the celery, which enabled agreater exposure of the E. coli to the lethal effects of UV light anddesiccation. The extremely open structure of the uninjured chiveshoots probably exposed the relatively few retained E. coli cells togreater levels of UV light and desiccation than those in celery.
Relative humidity may also affect the degree of persistence.Certainly, dry conditions (60% relative humidity) caused the decline ofSalmonella enterica, on the leaves of cilantro (Coriandrum sativum)plants, whereas wet conditions enabled the re-growth of thispathogen (Brandl and Mandrell, 2002). For the experiments reportedhere, the initially higher rate of survival of E. coli on Cos lettucecompared to the celery (Fig. 1) may have been caused by the highermaximum relative humidity experienced by Cos lettuce. The relativehumidity never dropped below 100% for Cos lettuce, but ranged from82 to 100% for the chive (Table 1).
It is tempting to attribute different degrees of persistence to theunique characteristics of the vegetables or other variable such asrelative humidity or temperature. However, it should be noted thatthese experiments were not designed to discern any effect except thatcaused by shoot injury on the degree of E. coli persistence, for theparticular plant species under investigation. The observed degrees ofpersistence resulted from the combination of different plant tissues,varying levels of relative humidity, temperature, and other variables.These were different between the experiments, and in order toconfidently ascribe an effect to a particular variable, experiments needto be conducted where the only one variable is changed at a time.
Prior to the field experiment it was thought that the higher fieldUV light level and the flushing effect of rain and overhead irrigationwould prevent the persistence observed for the glasshouse experi-ments (irrigation was directly into the potting mix during theglasshouse experiments). Perhaps the binding of E. coli to injuredplant tissue (Han et al., 2000; Seo and Frank, 1999; Takeuchi andFrank, 2000), prevented it from being washed off the field lettuce.
It is likely that other injured vegetables in the field, particularlycelery and chive, can also protect contaminating bacteria from beingwashed away. Such speculation is in agreement with the results ofBarker-Reid et al. (2009) who reported that the increment in thepersistence of E. coli on injured tissues was not restricted to Coslettuce only, but included injured shoots of iceberg lettuce (Lactucasativa var. Casino) growing in the field.
The initial field counts were made 1 day after E. coli application.Protection of bacteria from the environment during this time by theinjured tissue could account for the significantly higher initial countfrom the injured plants (Fig. 2). Also, the low levels of presumptiveE. coli on non-E. coli treated plants in the field (Fig. 2) may have comefrom the E. coli treated plants, the dam irrigation water, extraneousE. coli laden dust, or by birds and animals defecating in the lettucefield. Certainly, wild birds were often seen amongst the lettuce, andfoxes and feral cats were observed nearby. Moreover, the bulk of thepresumptive E. coli probably arose from the E. coli treatment ratherthan from extraneous sources. If there was significant extraneouscontamination, then comparable presumptive E. coli counts wouldhave been returned from both the E. coli and non-E. coli treated plants.This clearly was not the case (Fig. 2).
E. coli counts declined rapidly on the shoots of uninjured plants,which reached at least 1 log (90%) reduction per day (Figs. 1 and 2).Similar results had been reported for E. coli on glasshouse tomatoplants (Jurkevitch and Shapira, 2000), S. enterica on growth chambercilantro under dry conditions (Brandl and Mandrell, 2002), E. coli,S. enterica and Aeromonas hydrophila on growth chamber bean andcorn under dry conditions (O'Brien and Lindow, 1989), faecal coli-forms on field alfalfa (Bell, 1976), and E. coli and S. enterica high up on
pasture (Taylor and Burrows, 1971). Despite the rapid loss, E. coliwas still detectable from all plants, except uninjured Cos lettuce, atthe end of the experiments (Figs. 1 and 2). This pattern of rapid lossfollowed by an extended period of low numbers is typical for excretedpathogens in the environment (Feachem et al., 1983).
Actual levels of contamination are likely to be much lower thanthose used here, especially for locations with adequate sewagetreatment. The influence of shoot injury on low levels of contamina-tion should therefore be investigated, because low numbers ofbacteria may affect the length and degree of the persistence effect.Low numbers of bacteria may die out sooner than higher numbers,and so shorten the length of persistence. This hypothesis is based onthe finding that Salmonella baildon disappeared faster than highernumbers of this bacterium on shredded lettuce stored in bags at 12 °C.After 12 days of storage S. baildonwas recovered via enrichment fromonly one of six samples originally treated with 0.28 log10 CFU/g of thebacterium, and six out of six samples originally treatedwith 3.28 log10CFU/g (Weissinger et al., 2000). Very low numbers of bacteria mayalso diminish or even negate the injury effect. In this case, there maynot be much difference between the numbers of bacteria on injuredand uninjured plants, because low levels of E. coli were found tosurvive on uninjured plants for most if not all the length of theglasshouse experiments (Fig. 1).
E. coli has been found to adhere to injured plant tissues (Han et al.,2000; Seo and Frank, 1999; Takeuchi and Frank, 2000). This ad-herence was strong enough to resist the washing effect of the stainingprocesses used in these studies. This may mean that for this study, theactual numbers of E. coli CFU on the injured plants were higher thanrevealed by the colony counts. The stomaching process may not havedislodged all the E .coli from the injured plants samples, and asa consequence fewer E. coli cells were included in the inoculumsfor the PetrifilmTM E. coli/coliform count plates (3M Microbiology).Enumeration of such strongly attached E. coli cells may be possibleby thoroughly blending the plant samples, so that particles of planttissue with attached E. coli will be included in the inoculums for thegrowth medium. Another way to ascertain the number of E. coli cellson plant tissue after stomaching is via microscopy. The E. coli cellscould be counted after specific and viability staining with the methodof Seo and Frank (1999).
5. Conclusions
Vegetable plant shoots may become contaminated with entericbacterial pathogens through many factors including, animals, people,animal manures, and contaminated irrigation water. In general thesecontaminating bacteria can succumb quickly to desiccation and UVlight when exposed on the surface of the plant. However, the resultsof this investigation indicate that shoot injury can increase the per-sistence of contaminating bacteria on vegetable plants. All glasshousetested vegetables, celery, Cos lettuce and chive confirmed this as wellas Cos lettuce grown in the field. This has implications for on-farmfood safety plans and good agricultural practices (GAP).
Growers should avoid or modify production practices, which cancause even minor shoot injuries at the time the crop could also beexposed to human pathogenic bacteria. For example, untreated animalmanure or faecally tainted water should not be applied to crops injuredby factors such as harsh weather, plant disease or pests, and productionpractices such as spraying or partial harvests.
Acknowledgements
This research was supported by funding from the VictorianDepartment of Primary Industries, Victorian Department of HumanServices, and Horticulture Australia Limited, VegFed (The New ZealandVegetable and Potato Growers Federation), the Australian vegetablegrowers through the AUSVEG levy, CL & AK Warlan, Lightowler Fowl

237D. Harapas et al. / International Journal of Food Microbiology 138 (2010) 232–237
Manure Pty Ltd, and TD & EC Ould Pty Ltd. The authors wish to thankGraham Hepworth for his assistance in the experimental designand statistical analysis; Helen Harapas for her proof reading of themanuscript; and Janyce Truett, Pam Rogers, Siegfried Engleitner PhilipBall, Alan Noon, Trevor Davy, John Howard, Norman Morrison, andAdam Shaw for their help with plant production.
References
3MMicrobiology, Australia, 2008. 3M™ Petrifilm™ E. coli/Coliform Count Plates. http://solutions.3m.com.au/wps/portal/3M/en_AU/Microbiology/FoodSafety/ accessedMay 2008.
Arnold, G.J., Munce, B.A., 1997. Chapter 3: investigation of foodborne disease outbreaks.In: Hocking, A.D., Arnold, G., Jenson, I., Newton, K., Sutherland, P. (Eds.), FoodborneMicroorganisms of Public Health Significance. Australian Institute of Food Scienceand Technology Inc., North Sydney, pp. 31–70.
Aruscavage, D., Miller, S.A., lvey, M.L.L., Lee, K., LeJeune, J.T., 2008. Survival anddissemination of Escherichia coli on physically and biologically damaged lettuceplants. Journal of Food Protection 71, 2384–2388.
Barak, J.D., Whitehand, L.C., Charkowski, A.O., 2002. Differences in attachment ofSalmonella enterica serovars and Escherichia coli O157:H7 to alfalfa sprouts. Appliedand Environmental Microbiology 68, 4758–4763.
Barker-Reid, F., Harapas, D., Engleitner, S., Kreidl, S., Holmes, R., Faggian, R., 2009.Persistence of Escherichia coli on injured iceberg lettuce in the field, overheadirrigated with contaminated water. Journal of Food Protection 72, 458–464.
Behrsing, J., Winkler, S., Franz, P., Premier, R., 2000. Efficacy of chlorine for inactivationof Escherichia coli on vegetables. Postharvest Biology and Technology 19, 187–192.
Bell, R.G., 1976. Persistence of fecal coliform indicator bacteria on alfalfa irrigated withmunicipal sewerage effluent. Journal of Environmental Quality 5, 39–42.
Beuchat, L.R., 1998. Surface Decontamination of Fruits and Vegetables Eaten Raw: AReview. Food Safety Unit, World Health Organisation. WHO/FSF/FOS/98.2. 42.
Brackett, R.E., 1999. Incidence, contributing factors, and control of bacterial pathogensin produce. Postharvest Biology and Technology 15, 305–311.
Brandl, M.T., Mandrell, R.E., 2002. Fitness of Salmonella enterica serovar Thompson in thecilantro phyllosphere. Applied and Environmental Microbiology 68, 3614–3621.
Brandl, M.T., Haxo, A.F., Bates, A.H., Mandrell, R.E., 2004. Comparison of survival ofCampylobacter jejuni in the phyllosphere with that of the rhizosphere of spinachand radish plants. Applied and Environmental Microbiology 70, 1182–1189.
Bridson, E.Y., 1998. The Oxoid Manual, Eighth ed. Oxoid Limited, Basingstoke.CDC (Centers for Disease Control and Prevention), 2008. Outbreak of Salmonella
serotype Saintpaul infections associated with mutilple raw produce items— UnitedStates. MMWR 57, 929–934.
CFERT (California Food Emergency Response Team), 2007. Investigation of an Escherichiacoli O157:H7 outbreak associated with Dole pre-packaged spinach 50. http://www.marlerclark.com/2006_Spinach_Report_Final_01.pdf. Accessed May 2008.
Cifuentes, R.A., Harris, L.J., Hernandez, A.M., Suslow, T.V., 2001. Assessment ofproduction and retail handling practices of peaches, plum and nectarines onmicrobial food safety risk reduction. Central Valley Postharvest Newsletter 10, 12.
De Roever, C., 1998. Microbiological safety evaluations and recommendations on freshproduce. Food Control 9, 321–347.
Ewing, W.H., 1986. Edwards and Ewing's Identification of Enterobacteriaceae, Fourthed. Elsevier Science Publishing Co., Inc., New York.
Feachem, R.G., Bradley, D.J., Garelick, H., Mara, D.D., 1983. 4: detection, survival, andremoval of pathogens in the environment. In: Feachem, R.G., Bradley, D.J., Garelick, H.,Mara, D.D. (Eds.), Sanitation and Disease, Health Aspects of Excreta and WastewaterManagement. John Wiley & Sons, Chichester, pp. 53–66.
Han, Y., Linton, R.H., Nielsen, S.S., Nelson, P.E., 2000. Inactivation of Escherichia coliO157:H7 on surface-uninjured and -injured green pepper (Capsicum annuum L.) bychlorine dioxide gas as demonstrated by confocal laser scanning microscopy. FoodMicrobiology 17, 643–655.
Hansen, W., Yourassowsky, E., 1984. Detection of beta-glucuronidase in lactose-fermenting members of the family Enterobacteriaceae and its presence in bacterialurine cultures. Journal of Clinical Microbiology 20, 1177–1179.
Hoyle, B., 2008. Rise in foodborne outbreaks from leafy greens proves puzzling. Microbe 3,269–270.
Janning, B., in 't Veld, P.H., Notermans, S., Krämer, J., 1994. Resistance of bacterial strainsto dry conditions: use of anhydrous silica gel in a desiccation model system. Journalof Applied Bacteriology 77, 319–324.
Jurkevitch, E.J., Shapira, G., 2000. Structure and colonization dynamics of epiphyticbacterial communities and of selected component strains on tomato (Lycopersiconesculentum) leaves. Microbial Ecology 40, 300–308.
Lynch, M.F., Tauxe, R.V., Hedberg, C.W., 2009. The growing burden of foodborneoutbreaks due to contaminated fresh produce: risks and opportunities. Epidemi-ology and Infection 137, 307–315 Review article.
McDaniels, A.E., Rice, E.W., Reyes, A.L., Johnson, C.H., Haugland, R.A., Stelma Jr, G.N.,1996. Confirmational identification of Escherichia coli, a comparison of genotypicand phenotypic assays for glutamate decarboxylase and ß-D-glucuronidase.Applied and Environmental Microbiology 62, 3350–3354.
Natvig, E.E., Ingham, S.C., Ingham, B.H., Cooperband, L.R., Roper, T.R., 2002. Salmonellaenterica Serovar Typhimurium and Escherichia coli contamination of root and leafvegetables grown in soils with incorporated bovine manure. Applied andEnvironmental Microbiology 68, 2737–2744.
O'Brien, R.D., Lindow, S.E., 1989. Effect of plant species and environmental conditionson epiphytic populations sizes of Pseudomonas syringae and other bacteria.Phytopathology 79, 619–627.
Seo, K.H., Frank, J.F., 1999. Attachment of Escherichia coli O157:H7 to lettuce leaf surfaceand bacterial viability in response to chlorine treatment as demonstrated by usingconfocal scanning laser microscopy. Journal of Food Protection 62, 3–9.
Solomon, E.B., Potenski, C.J., Mathews, K.R., 2002. Effect of irrigation method ontransmission to and persistence of Escherichia coli O157:H7 on lettuce. Journal ofFood Protection 65, 673–676.
Sproston, E.L., Macrae, M., Ogden, I.D., Wilson, M.J., Strachan, N.J.C., 2006. Slugs:potential novel vectors of Escherichia coli O157:H7. Applied and EnvironmentalMicrobiology 72, 144–149.
Standards Australia Committee on Food Microbiology, 1998. Collection 27, FoodMicrobiology. Standards Australia collections. Standards Australia Publishing,Strathfield, NSW.
Takeuchi, K., Frank, J.F., 2000. Penetration of Escherichia coli O157:H7 into lettucetissues as affected by inoculum size and temperature and the effect of chlorinetreatment on cell viability. Journal of Food Protection 63, 434–440.
Taylor, R.J., Burrows, M.R., 1971. The survival of Escherichia coli and Salmonella dublin inslurry on pasture and the infectivity of S. dublin for grazing calves. British VeterinaryJournal 127, 536–543.
Thompson, S.A. 2008. Feeling the squeeze tomato growers take big hit from food scare,Rural Cooperatives, vol. 75. 13–15, 41.
Tryland, I., Fiksdal, L., 1998. Enzyme characteristics of ß-D-galactosidase and ß-D-glucuronidase positive bacteria and their interference in rapid methods for thedetection of waterborne coliforms and Escherichia coli. Applied and EnvironmentalMicrobiology 64, 1018–1023.
Weissinger, W.R., Chantarapanont, W., Beuchat, L.R., 2000. Survival and growth ofSalmonella baildon in shredded lettuce and diced tomatoes, and effectiveness ofchlorinated water as a sanitizer. International Journal of Food Microbiology 62,123–131.