peripheral regeneration and central sprouting of sensory neurone axons in aplysia californica...
TRANSCRIPT

2067The Journal of Experimental Biology 198, 2067–2078 (1995)Printed in Great Britain © The Company of Biologists Limited 1995
PERIPHERAL REGENERATION AND CENTRAL SPROUTING OF SENSORYNEURONE AXONS IN APLYSIA CALIFORNICA FOLLOWING NERVE INJURY
ISABELLA STEFFENSEN1, MICHAEL F. DULIN2, EDGAR T. WALTERS2 AND CATHERINE E. MORRIS1,*1Neurosciences, Loeb Institute, University of Ottawa, OCH, Ottawa, Ontario, Canada K1Y 4E9 and
2Department of Integrative Biology, University of Texas – Houston Medical School, PO Box 20708, Houston, TX 77225, USA
Accepted 13 June 1995
Morphological methods were used to examine injury-induced growth of peripheral and central axons ofnociceptive mechanosensory neurones in the ventrocaudal(VC) clusters of the pleural ganglia of Aplysia californica.Pedal nerve crush transected all axons in the nerve whileleaving the overlying sheath largely intact.Immunohistochemical staining was performed with anantibody to a sensory-neurone-specific peptide, sensorin-A.Following bilateral crush of pedal nerve p9, whichinnervates the tail, sensorin-A immunofluorescence waslost distal to the crush site within 2 days. Fineimmunopositive fibres began to invade the crush regionwithin 5 days. These fibres arborized in the crush regionand gradually extended down the crushed nerve.Immunopositive fibres were found near the tail within 3
weeks. Similar results were obtained after injectingindividual sensory neurone somata in the tail/p9 region ofthe VC cluster with biocytin. Biocytin injections andhorseradish peroxidase injections 3 weeks after ipsilateralpedal nerve crush revealed new fibres projecting rostrallyfrom the tail/p9 region of the VC cluster and entering thepleural–cerebral and pleural–abdominal connectives. Suchprojections were never observed in control, uncrushedpreparations. These results demonstrate that nerve injurytriggers extensive growth of both peripheral and centralprocesses of the VC sensory neurones.
Key words: axotomy, nociceptor, recovery of function, sensitization,memory, Aplysia californica.
Summary
Axonal growth is a common reaction to peripheral nerveinjury. Injury-induced growth includes regeneration of severedaxons (e.g. Fawcett and Keynes, 1990) and sprouting of neuritesfrom either damaged or undamaged neurones at a distance fromthe site of injury (e.g. Weddell et al. 1941; Rotshenker, 1988;Bannatyne et al. 1989). The presence of large, identifiedneurones in gastropod molluscs, such as the snails Helisomatrivolvis and Lymnaea stagnalis, has proved useful forexamining cellular mechanisms that control regeneration andsprouting, especially using in vitro preparations (e.g. Bullochand Ridgway, 1989; Carrow and Levitan, 1989; Ridgway et al.1991; Rehder et al. 1992). The gastropod nervous system alsoprovides advantages for linking injury-induced neuronal growthto recovery of function in the whole animal (e.g. Allison andBenjamin, 1985; Moffett and Snyder, 1985; Cohan et al. 1987;Kruk and Bulloch, 1992; Syed et al. 1992).
The occurrence of apparently similar neuritic outgrowth inthe central nervous system (CNS) or central ganglia after nerveinjury (Kelly et al. 1989; Cameron et al. 1992; Woolf et al.
Introduction
*Author for correspondence.
1992) and learning (reviewed by Bailey and Kandel, 1993)raises interesting questions about potential mechanistic andevolutionary relationships between these traditionally separateclasses of plasticity (Walters, 1994; Walters and Ambron,1995). A marine gastropod, Aplysia californica, has beenutilized extensively to study neural mechanisms underlyingbehavior and learning (Kandel, 1979; Walters, 1994; Krasneand Glanzman, 1995). Among these studies have been severalshowing that sensory neurones display growth of new neurites,varicosities and synapses following one form of learning,general sensitization, or following the application ofextracellular or intracellular signals associated withsensitization (Bailey and Chen, 1983, 1988; Nazif et al. 1991;Bailey et al. 1992). Although growth of neurites from A.californica neurones has been described in dissociated cellculture (e.g. Schacher and Proshansky, 1983; Ambron et al.1985; Glanzman et al. 1989), there have been remarkably fewstudies of injury-induced morphological alterations in thewhole animal. Fredman (1988) used retrograde staining and
2068 I. STEFFENSEN AND OTHERS
intracellular dye injection to show regeneration of axons in thepleural–cerebral connectives correlated with recovery ofescape locomotion. Recently Ross et al. (1994) provided adetailed analysis of axonal regeneration and functionalrecovery in an identified buccal motor neurone of A.californica. However, injury-induced morphologicalalterations have not yet been reported in sensory neurones ofA. californica or of any other mollusc.
Wide-dynamic-range, nociceptive mechanosensory neuronesin the ventrocaudal (VC) clusters of A. californica (Walters etal. 1983) offer a particularly interesting opportunity to examinemorphological reactions to nerve injury. These cells are knownto express dramatic alterations after nerve injury, includinglong-term synaptic facilitation and increased excitability oftheir somata (Walters et al. 1991; Clatworthy and Walters,1994; Gunstream et al. 1995). In addition, they displaymorphological alterations after learning-related treatments(Glanzman et al. 1990; Nazif et al. 1991). Moreover, theirperipheral receptive fields display persistent enlargementfollowing noxious stimulation, suggesting the occurrence ofinjury-induced growth in their peripheral arborization (Billyand Walters, 1989). As described in the preceding paper (Dulinet al. 1995), pedal nerve crush transiently abolishes the tail-evoked siphon reflex. Within 3 weeks, some axotomized VCsensory neurones re-establish receptive fields, which led us tohypothesize that one mechanism underlying recovery of thereflex is regeneration of injured VC cell axons. Furthermore,because axotomized VC sensory neurones could be activatedantidromically by stimulation of connectives and nerves intowhich they do not normally project, we hypothesized thataxonal injury causes these cells to sprout new fibres within theCNS that grow into aberrant pathways. Here, we test thesehypotheses with morphological methods and show that nerveinjury triggers both regenerative growth of the peripheral axonsof VC sensory neurones and sprouting of neurites in centralganglia and connectives.
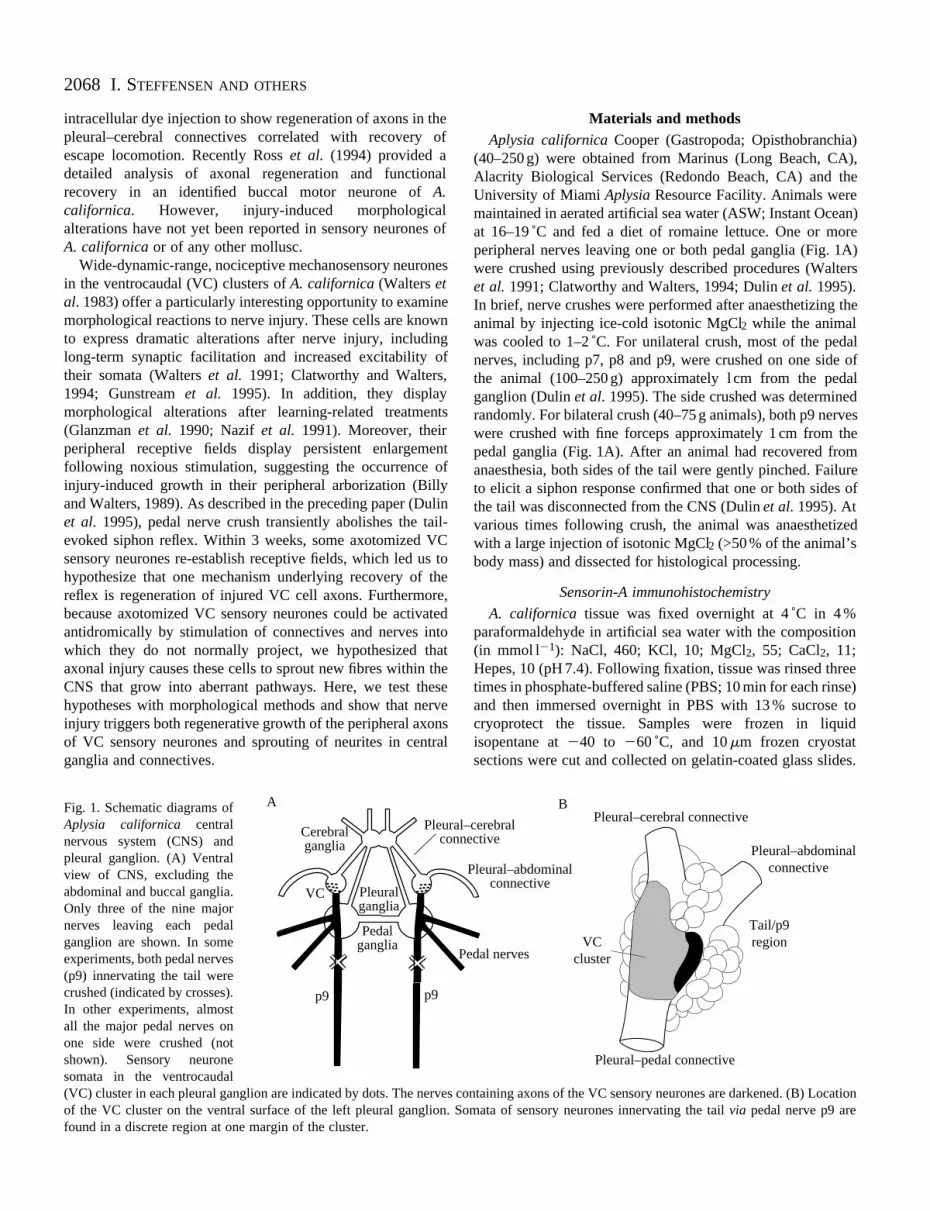
Fig. 1. Schematic diagrams ofAplysia californica centralnervous system (CNS) andpleural ganglion. (A) Ventralview of CNS, excluding theabdominal and buccal ganglia.Only three of the nine majornerves leaving each pedalganglion are shown. In someexperiments, both pedal nerves(p9) innervating the tail werecrushed (indicated by crosses).In other experiments, almostall the major pedal nerves onone side were crushed (notshown). Sensory neuronesomata in the ventrocaudal(VC) cluster in each pleural ganglion are indicated by dots. The nerves cof the VC cluster on the ventral surface of the left pleural ganglion. Sfound in a discrete region at one margin of the cluster.
Cerebral ganglia
Pleural ganglia
Pedal ganglia
Pleuralconn
P
VC
A
p9p9
Materials and methodsAplysia californica Cooper (Gastropoda; Opisthobranchia)
(40–250 g) were obtained from Marinus (Long Beach, CA),Alacrity Biological Services (Redondo Beach, CA) and theUniversity of Miami Aplysia Resource Facility. Animals weremaintained in aerated artificial sea water (ASW; Instant Ocean)at 16–19 ˚C and fed a diet of romaine lettuce. One or moreperipheral nerves leaving one or both pedal ganglia (Fig. 1A)were crushed using previously described procedures (Walterset al. 1991; Clatworthy and Walters, 1994; Dulin et al. 1995).In brief, nerve crushes were performed after anaesthetizing theanimal by injecting ice-cold isotonic MgCl2 while the animalwas cooled to 1–2 ˚C. For unilateral crush, most of the pedalnerves, including p7, p8 and p9, were crushed on one side ofthe animal (100–250 g) approximately l cm from the pedalganglion (Dulin et al. 1995). The side crushed was determinedrandomly. For bilateral crush (40–75 g animals), both p9 nerveswere crushed with fine forceps approximately 1 cm from thepedal ganglia (Fig. 1A). After an animal had recovered fromanaesthesia, both sides of the tail were gently pinched. Failureto elicit a siphon response confirmed that one or both sides ofthe tail was disconnected from the CNS (Dulin et al. 1995). Atvarious times following crush, the animal was anaesthetizedwith a large injection of isotonic MgCl2 (>50 % of the animal’sbody mass) and dissected for histological processing.
Sensorin-A immunohistochemistry
A. californica tissue was fixed overnight at 4 ˚C in 4 %paraformaldehyde in artificial sea water with the composition(in mmol l21): NaCl, 460; KCl, 10; MgCl2, 55; CaCl2, 11;Hepes, 10 (pH 7.4). Following fixation, tissue was rinsed threetimes in phosphate-buffered saline (PBS; 10 min for each rinse)and then immersed overnight in PBS with 13 % sucrose tocryoprotect the tissue. Samples were frozen in liquidisopentane at 240 to 260 ˚C, and 10 mm frozen cryostatsections were cut and collected on gelatin-coated glass slides.
ontaining axons of the VC sensory neurones are darkened. (B) Locationomata of sensory neurones innervating the tail via pedal nerve p9 are
Tail/p9 region
Pleural–cerebral connective
Pleural–abdominal connective
Pleural–pedal connective
VC cluster
–cerebral ective
Pleural–abdominal connective
edal nerves
B

2069Regeneration and sprouting of sensory neurones
Frozen sections were incubated overnight at 4 ˚C in a blockingsolution consisting of 10 % normal goat serum (JacksonImmunoresearch, West Grove, PA), 1 % bovine serum albumin(BSA) and 0.4 % Triton X-100 in PBS. After a single quickrinse in washing buffer (0.1 % Triton X-100 in PBS), sectionswere incubated overnight at 4 ˚C with a rabbit polyclonalantiserum raised against sensorin-A (Brunet et al. 1991; a giftfrom E. Kandel and R. Hawkins) at 1:400 in PBS with 0.4 %Triton X-100. Following the primary antibody incubation,ganglia were rinsed four times in washing buffer (10 min eachrinse) and then incubated overnight at 4 ˚C with adichlorotriazinylaminofluorescein (DTAF)-conjugated goatanti-rabbit secondary antibody (Jackson Immunoresearch) at1:100 in PBS with 0.4 % Triton X-100. After three rinses inwashing buffer (10 min each rinse), sections were mounted ina 3:1 solution of glycerol in PBS with 1 % n-propyl gallate, areagent used to reduce photobleaching. Ganglia were viewedwith a Zeiss Universal microscope equipped withepifluorescence optics (filter set H7909 for fluorescein) andphotomicrographs were taken on Ilford XP2–400 film.
Biocytin fills
For ionophoretic injection into sensory neurones, a 4 %solution of biocytin (Sigma, St Louis, MO) was drawn into thetip of an electrode; the electrode was then backfilled with asolution of 0.33 mol l21 KCl and 10 mmol l21 Tris, pH 7.5.Sensory neurone somata in the tail/p9 region of the VC clusterin surgically desheathed pleural ganglia (Fig. 1B) werealternately hyperpolarized and depolarized by 30 mV fromresting potential (at 0.5 Hz) by passing current through theinjection electrode for 30–40 min. Ganglia were then placed in
Fig. 2. General effects of nerve crush. A–D are phase-contrast images otaken from a different animal. E is a fluorescent image of the same fieltransected fibres and intact sheath. (C) 16 days after nerve crush. The swcrush site. (D) 17 days after nerve crush. The large inclusions were conswith Hoechst 33,258, a fluorescent nuclear stain. Note the absence of n
isotonic L15 culture medium (Schacher and Proshansky, 1983)at 4 ˚C for 20–42 h to allow time for the biocytin to move downthe axons. Tissue was fixed in 4 % paraformaldehyde in ASWovernight at 4 ˚C, rinsed three times in PBS (10 min each rinse)and then cryoprotected in PBS with 15 % sucrose. Frozencryostat sections were prepared as above. Sections wereincubated overnight in blocking solution. Following a singlequick rinse, sections were incubated in Biodipy (MolecularProbes, Eugene, OR) at 1:100 in PBS with 0.4 % Triton X-100.After three rinses in washing buffer, sections were mounted,viewed and photographed as above.
Horseradish peroxidase fills
Sensory neurones in the tail/p9 area of the VC cluster(Fig. 1B) were pressure-injected with a solution of 4 %horseradish peroxidase (HRP) in 1 mmol l21 KCl and6 mmol l21 Fast Green, using procedures similar to thosedescribed by Nazif et al. (1991). Injection electrodes (3–5 MV)were connected to a Picospritzer II (General Valve, Fairfield,NJ) which delivered 2–3 pulses (10–20 ms each) at140–170 kPa. The Fast Green allowed visual monitoring ofeach injection. Injected ganglia were left in culture medium at4 ˚C for 20–24 h. Preparations were then fixed in a solution of2.5 % glutaraldehyde and 30 % sucrose in 0.1 mol l21
phosphate buffer (PB), pH 7.3, for 1 h. The ganglia werewashed with PB and placed in a solution containingdiaminobenzidene (Vector Laboratories). Tissue was thenreacted using 0.003 % peroxide for 15 min, dehydrated in agraded series of ethanol concentrations (20, 50, 75 and 95 %)and cleared in methyl salicylate. Whole-mount preparationswere placed on a glass slide in Permount, viewed on an
f cryostat sections of nerve p9 about 1 cm from the pedal ganglion, eachd as D. (A) Uncrushed p9. (B) Immediately after nerve crush. Note theollen region between the arrowheads is characteristic of the recovering
istently present at the recovering crush site. (E) The same section staineduclei within the large inclusions. Scale bar, 100 mm.

2070 I. STEFFENSEN AND OTHERS
Olympus BH-2 microscope, and photographed with KodakTMAX 100 (black and white) or Ektachrome Tungsten.
ResultsGeneral effects of nerve crush
Nerve crush caused the evident transection of all fibres atthe crush site, while leaving the overlying sheath apparentlyintact, as illustrated by the phase micrograph of Fig. 2B.Transection of the fibres was also indicated by visualinspection of crushed nerves under the dissecting microscope(not shown). Furthermore, in every case that the nerve wastested electrophysiologically within 1 week after nerve crush,sensory neurone action potentials failed to be conductedthrough the crush site (Dulin et al. 1995). Approximately 1–2weeks after nerve crush, the signs of fibre transection at thecrush site disappeared, sensory neurone conduction through the
Fig. 3. Sensorin-A immunofluorescence in control animals.(A) Cryostat section of the pleural (Pl) and pedal (Pd) gangliaand of the posterior pedal nerve p9 (arrow). Sensory neuronesomata in the pleural VC cluster are immunopositive forsensorin-A, as are fibres in the pleural–pedal connective,pedal ganglion and nerve p9. (B,C) Segments of nerve p9from two other animals. Scale bar, 500 mm.
crush site began to be restored (Dulin et al. 1995) and thecrushed region consistently swelled to about twice the diameterof the adjacent nerve regions. The swelling persisted for aslong we examined the nerves, up to 60 days post-crush. Thisswelling provided a useful marker for the crush site in bothmorphological and electrophysiological experiments (seeDulin et al. 1995). Large inclusions (up to 100 mm) wereobserved in the region of the swelling (Fig. 2D) and distally(Fig. 5B–D,H). Failure of the inclusions to stain with thefluorescent nuclear dye Hoechst 33,258 indicated that theseprofiles were acellular (Fig. 2E).
Immunohistochemical evidence for sensory neuroneregeneration
Brunet et al. (1991) discovered a peptide, sensorin-A, thatappears to be expressed in identified clusters ofmechanosensory neurones and in no other neurones in A.
2071Regeneration and sprouting of sensory neurones
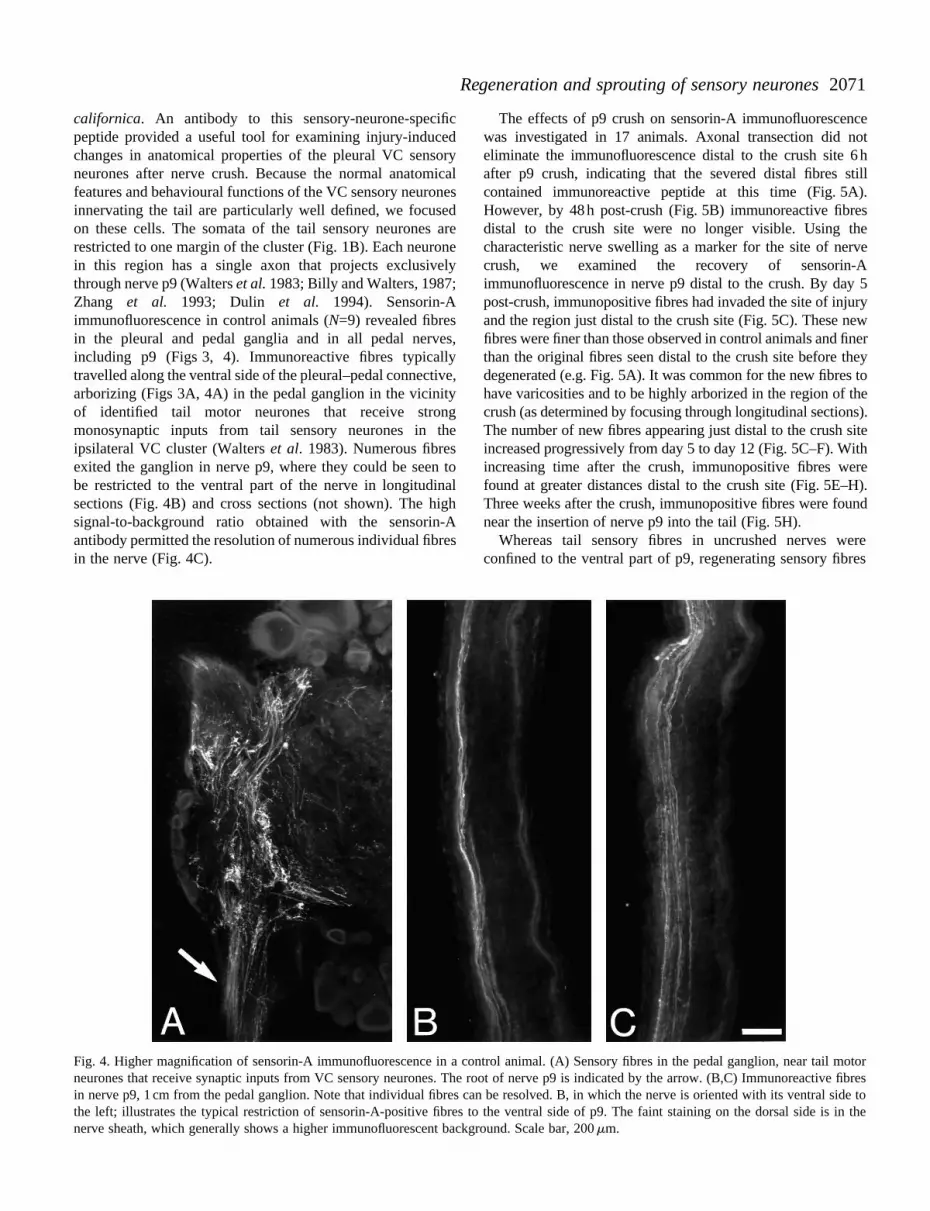
californica. An antibody to this sensory-neurone-specificpeptide provided a useful tool for examining injury-inducedchanges in anatomical properties of the pleural VC sensoryneurones after nerve crush. Because the normal anatomicalfeatures and behavioural functions of the VC sensory neuronesinnervating the tail are particularly well defined, we focusedon these cells. The somata of the tail sensory neurones arerestricted to one margin of the cluster (Fig. 1B). Each neuronein this region has a single axon that projects exclusivelythrough nerve p9 (Walters et al. 1983; Billy and Walters, 1987;Zhang et al. 1993; Dulin et al. 1994). Sensorin-Aimmunofluorescence in control animals (N=9) revealed fibresin the pleural and pedal ganglia and in all pedal nerves,including p9 (Figs 3, 4). Immunoreactive fibres typicallytravelled along the ventral side of the pleural–pedal connective,arborizing (Figs 3A, 4A) in the pedal ganglion in the vicinityof identified tail motor neurones that receive strongmonosynaptic inputs from tail sensory neurones in theipsilateral VC cluster (Walters et al. 1983). Numerous fibresexited the ganglion in nerve p9, where they could be seen tobe restricted to the ventral part of the nerve in longitudinalsections (Fig. 4B) and cross sections (not shown). The highsignal-to-background ratio obtained with the sensorin-Aantibody permitted the resolution of numerous individual fibresin the nerve (Fig. 4C).
Fig. 4. Higher magnification of sensorin-A immunofluorescence in a cneurones that receive synaptic inputs from VC sensory neurones. The rin nerve p9, 1 cm from the pedal ganglion. Note that individual fibres cthe left; illustrates the typical restriction of sensorin-A-positive fibres nerve sheath, which generally shows a higher immunofluorescent back
The effects of p9 crush on sensorin-A immunofluorescencewas investigated in 17 animals. Axonal transection did noteliminate the immunofluorescence distal to the crush site 6 hafter p9 crush, indicating that the severed distal fibres stillcontained immunoreactive peptide at this time (Fig. 5A).However, by 48 h post-crush (Fig. 5B) immunoreactive fibresdistal to the crush site were no longer visible. Using thecharacteristic nerve swelling as a marker for the site of nervecrush, we examined the recovery of sensorin-Aimmunofluorescence in nerve p9 distal to the crush. By day 5post-crush, immunopositive fibres had invaded the site of injuryand the region just distal to the crush site (Fig. 5C). These newfibres were finer than those observed in control animals and finerthan the original fibres seen distal to the crush site before theydegenerated (e.g. Fig. 5A). It was common for the new fibres tohave varicosities and to be highly arborized in the region of thecrush (as determined by focusing through longitudinal sections).The number of new fibres appearing just distal to the crush siteincreased progressively from day 5 to day 12 (Fig. 5C–F). Withincreasing time after the crush, immunopositive fibres werefound at greater distances distal to the crush site (Fig. 5E–H).Three weeks after the crush, immunopositive fibres were foundnear the insertion of nerve p9 into the tail (Fig. 5H).
Whereas tail sensory fibres in uncrushed nerves wereconfined to the ventral part of p9, regenerating sensory fibres
ontrol animal. (A) Sensory fibres in the pedal ganglion, near tail motoroot of nerve p9 is indicated by the arrow. (B,C) Immunoreactive fibresan be resolved. B, in which the nerve is oriented with its ventral side toto the ventral side of p9. The faint staining on the dorsal side is in theground. Scale bar, 200 mm.

2072 I. STEFFENSEN AND OTHERS
Fig. 5. Sensorin-A immunofluorescence at different times after nerve crush. (A) 6 h post-crush. (B) 48 h post-crush. (C) 5 days post-crush. Thearrowhead indicates where a fine fibre has grown through the crush site and bifurcated. (D) 8 days post-crush. (E) 12 days post-crush about0.5 cm distal to the crush site. The arrowhead indicates varicosities within the crush site. (F) Higher magnification of another p9 nerve 12 dayspost-crush about 0.5 cm distal to the crush site. (G) 17 days post-crush about 1 cm distal to the crush site. The arrowhead indicates varicosities.(H) 22 days post-crush about 8 cm distal to the crush site, just proximal to where the main trunk of p9 branches before entering the tail. Someof the large inclusions that were observed by phase contrast (Fig. 2D) are also visible with immunofluorescence (B–D, H). Scale bar, 100 mm(except F, 50 mm).
were found throughout the crush site and emerged distally withan essentially random distribution in p9. We found no evidencein any of these experiments for preferential growth ofregenerating sensorin-A-containing fibres in the ventral part ofp9. At the ‘regenerating front’, sensory fibres were fine andhighly arborized, but behind this front they became lessarborized and thicker with time. At the swollen crush site,however, fine extensively arborized fibres remained even 3
weeks after the crush. It was not possible to determine whetherthese represented persistent, stable structures or processes thatwere still actively extending and retracting more than 2 weeksafter their initial regrowth into the crush site.
Dye-fills of individual sensory neurones demonstrate axonalregeneration
Biocytin is a particularly effective dye for intracellular fills

2073Regeneration and sprouting of sensory neurones
Fig. 6. Biocytin fills of tail sensory neurones21 days after crushing nerve p9. (A) Threebiocytin-filled somata in the VC cluster.Biocytin-filled fibres are seen in the pleural–pedal connective (arrow). (B) Arborization ofbiocytin-filled fibres just proximal to the crushsite. (C) Biocytin-filled fibres in p9 just distalto the crush site. Focusing through the sectionconfirmed that there are two fibres(arrowhead). (D) Biocytin-filled fibre(arrowhead) on the ventral side of p9 about1 cm distal to the crush. (E) Highermagnification of D. Note the globularinclusions at the crush site in B and C anddistal to the site in D. Scale bars, A, 200 mm;B–D, 50 mm; E, 25 mm.
in molluscs (Ewadinger et al. 1994). We used biocytin toexamine the morphology of individual sensory neurones 3weeks after crushing nerve p9. In two animals, we identifiedsensory neurones with presumptive axons in nerve p9 byelectrically stimulating the nerve and recording actionpotentials in the soma (see Dulin et al. 1995). One to threecells activated by p9 stimulation were then filled withbiocytin in each pleural ganglion in each animal (N=8 cellsin total). Each filled sensory neurone had a single axon in thepleural–pedal connective on both the control (not shown) andnerve-crushed sides (Fig. 6A), which passed through thepedal ganglion and entered nerve p9. On the nerve-crushedside, the biocytin-containing fibres traversed the crush siteand continued through p9 for another 1–2 cm to where thenerve had been cut during dissection (Fig. 6B–E). In theregion of the crush, the sensory fibres branched andmeandered (Fig. 6B,C). Once they emerged from the swollenregion, they followed a straight path and arborizations wereno longer observed (Fig. 6D,E).
Morphological evidence for central sprouting by injuredsensory neurones
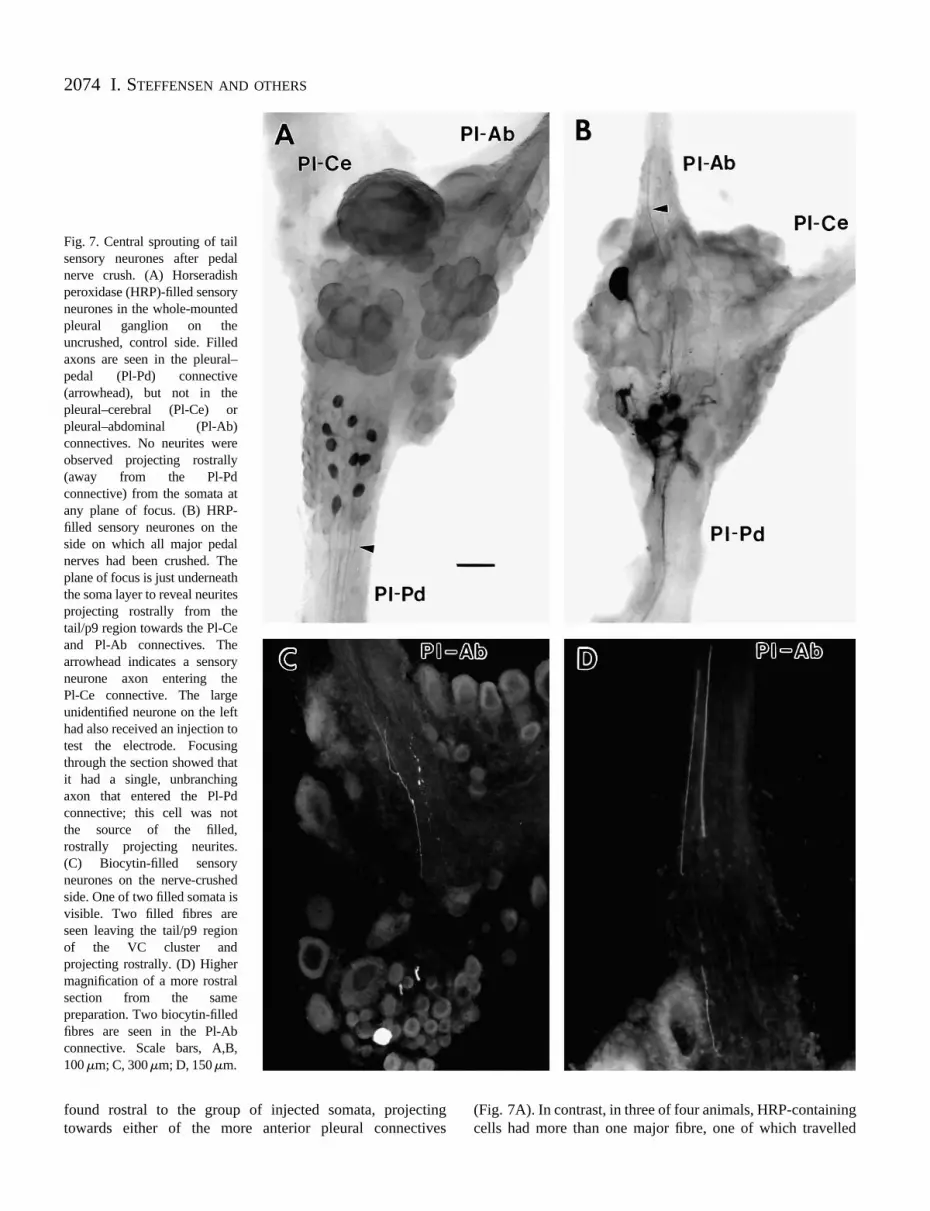
As described in the preceding paper (Dulin et al. 1995), wehave obtained electrophysiological evidence consistent withthe growth of new neurites into central connectives andperipheral nerves by injured VC sensory neurones. To seekanatomical evidence of injury-induced sprouting in the CNSby sensory neurones, we injected either HRP or biocytin intosensory neurone somata in the tail/p9 region of the VC cluster(Fig. 1B). Previous studies indicated that sensory neurones inthis region send single axons directly into the pleural–pedalconnective and never send axons into the pleural–cerebral orpleural–abdominal connectives (Walters et al. 1983; Zhang etal. 1993; Dulin et al. 1994; A. Billy and E. T. Walters,unpublished observations). In initial experiments, we injectedHRP into 5–10 sensory neurone somata in each pleuralganglion 3 weeks after unilateral pedal nerve crush. All stainedcells on the uncrushed side had a single fibre that travelleddirectly into the pleural–pedal connective; no filled fibres were
2074 I. STEFFENSEN AND OTHERS
Fig. 7. Central sprouting of tailsensory neurones after pedalnerve crush. (A) Horseradishperoxidase (HRP)-filled sensoryneurones in the whole-mountedpleural ganglion on theuncrushed, control side. Filledaxons are seen in the pleural–pedal (Pl-Pd) connective(arrowhead), but not in thepleural–cerebral (Pl-Ce) orpleural–abdominal (Pl-Ab)connectives. No neurites wereobserved projecting rostrally(away from the Pl-Pdconnective) from the somata atany plane of focus. (B) HRP-filled sensory neurones on theside on which all major pedalnerves had been crushed. Theplane of focus is just underneaththe soma layer to reveal neuritesprojecting rostrally from thetail/p9 region towards the Pl-Ceand Pl-Ab connectives. Thearrowhead indicates a sensoryneurone axon entering the Pl-Ce connective. The largeunidentified neurone on the lefthad also received an injection totest the electrode. Focusingthrough the section showed thatit had a single, unbranchingaxon that entered the Pl-Pdconnective; this cell was not the source of the filled, rostrally projecting neurites. (C) Biocytin-filled sensoryneurones on the nerve-crushedside. One of two filled somata isvisible. Two filled fibres areseen leaving the tail/p9 region of the VC cluster and projecting rostrally. (D) Highermagnification of a more rostralsection from the samepreparation. Two biocytin-filledfibres are seen in the Pl-Abconnective. Scale bars, A,B, 100 mm; C, 300mm; D, 150mm.
found rostral to the group of injected somata, projectingtowards either of the more anterior pleural connectives
(Fig. 7A). In contrast, in three of four animals, HRP-containingcells had more than one major fibre, one of which travelled

2075Regeneration and sprouting of sensory neurones
towards, and sometimes entered, the pleural–cerebral orpleural–abdominal connective (Fig. 7B). In these animals, thearborization of the filled sensory neurones in the neuropilebeneath the somata appeared denser and the organization of thefibres much less regular on the crushed than on the uncrushedside.
Biocytin was injected into tail/p9 sensory neurones thatdisplayed electrophysiological responses to stimulation of thepleural–cerebral or pleural–abdominal connectives 3 weeksafter ipsilateral pedal nerve crush (Dulin et al. 1995). In fouranimals, 1–3 VC sensory neurone somata were successfullyfilled in each pleural ganglion. On the uncrushed side, all sevenfilled cells sent a single fibre posteriorly into the pleural–pedalconnective. No neurites projected anteriorly or entered thepleural–cerebral or pleural–pedal connectives. In contrast, allseven filled cells on the nerve-crushed side displayed one ormore fibres projecting anteriorly (Fig. 7C) towards thepleural–cerebral and/or the pleural–abdominal connectives.Each also sent a fibre posteriorly into the pleural–pedalconnective. As shown in Fig. 7D, some of the anteriorlyprojecting fibres entered an anterior connective. Two cells hadfibres entering both of the anterior connectives, two cells hadfibres entering the pleural–cerebral connective and one cell hada fibre entering the pleural–abdominal connective. The tworemaining cells had fibres that projected anteriorly, but werenot observed to leave the pleural ganglion.
Because biocytin passes through at least some electricalsynapses in A. californica (Hickie, 1994) and other molluscs(Ewadinger et al. 1994), we also looked in these same animalsfor evidence of injury-induced formation of electrical synapsesin the CNS. Biocytin was not found in any neuronal somata inthe pleural or pedal ganglia that had not been injected. Thissuggests that, if novel electrical synapses are formed by injuredVC sensory neurones, they are formed outside the CNS.
DiscussionRegeneration of severed sensory neurone axons
We have found that the VC sensory neurones displayregenerative growth of peripheral axons following nerve crushthat transects their axons but leaves the overlying sheathessentially intact. Axonal regeneration under similarconditions has been described in other mechanosensoryneurones having centrally located somata and peripheralreceptive fields, notably those in the leech (Van Essen andJansen, 1977; Bannatyne et al. 1989) and mammals. Inmammals, sensory neurones with somata in dorsal root gangliahave been utilized to examine a variety of cellular andmolecular concomitants of axonal regeneration (e.g. Wujekand Lasek, 1983; Oblinger and Lasek, 1984; Bisby and Keen,1986; Kanje et al. 1986; Jenkins et al. 1993; Mearow et al.1994). The VC sensory neurones in A. californica, like the N,P and T sensory neurones in the leech, offer specialopportunities to examine molecular and functional aspects ofregeneration because the functions of individual cells are welldefined and intracellular manipulations (e.g. injection of
putative molecular signals or blockers – see Walters andAmbron, 1995) can be performed readily on one or moreidentified cells during various stages of regeneration.
Although we have not yet conducted a systematic analysisof the time course of axonal regeneration in VC sensoryneurones, the appearance of new fibres at the crush site andtheir extension into the distal segment of nerve p9 roughlyparallel the reappearance of spike conduction through the crushsite and the recovery of functioning receptive fields on the tail(Dulin et al. 1995). Thus, some regenerating fibres that labelwith the sensorin-A antibody traverse the crush site 5 days afterinjury, extend farther into p9 as time goes by and are foundnear the tail by 3 weeks after injury, when receptive fields onthe tail are being re-established. Recent results obtained withsensorin-A immunostaining have revealed characteristichelical coil endings in the muscle layer of A. californica bodywall in the tail and siphon (I. Steffensen and C. E. Morris, inpreparation). It will be interesting to examine changes insensorin-A immunoreactivity in body wall disconnected fromthe CNS by nerve injury and in the density or appearance ofthese putative mechanosensory structures during loss andrecovery of function.
Demonstrations by sensorin-A immunofluorescence andbiocytin injections of the extension of VC sensory neuroneaxons well beyond the crush site suggest that regenerating tailsensory neurones from the VC cluster contribute to recoveryof the tail-evoked siphon reflex described in the precedingpaper (Dulin et al. 1995). However, we pointed out that thefailure of the centrally conducted axon responses to nerveshock to be restored earlier than the behavioural responses(even though the crush site was relatively close to the VCsomata) indicates that the earliest stages of reflex recovery arenot due to regeneration of VC sensory neurone axons into thetail. Two mechanisms that, in principle, could producerelatively rapid restoration of sensory input to the CNS fromthe denervated half of the tail are (1) regeneration of the centralaxons of peripheral sensory neurones (or sensoryinterneurones) across the relatively short distance between thecentral nerve crush site and their central targets and (2)reconnection of regenerating VC neurites to surviving axonsin the distal stump of nerve p9. The first mechanism seemslikely to contribute, but has not yet been tested. Some of ourdata are relevant to the second mechanism.
Do proximal and distal stumps of severed axons reconnect?
The possibility that regenerating neurites form electricalsynapses with surviving distal stumps of severed VC sensoryneurone axons needs further examination. The disappearanceof sensorin-A immunoreactivity from axon segments distal tothe crush within several days might be due to degeneration ofthese segments (see below). However, the disappearance ofsensorin-A does not necessarily indicate that the axonsegments are lost. It might simply be that the peptide is lost –perhaps by transport to the periphery. This peptide issufficiently large (Brunet et al. 1991) that it is unlikely to passthrough gap junctions and so should not be replenished in distal

2076 I. STEFFENSEN AND OTHERS
segments if reconnection occurs via electrical synapses. Ourexperiments with biocytin, which readily passes through atleast some electrical synapses in molluscs (Ewadinger et al.1994; Hickie, 1994), argue against novel coupling with otherneurones in the CNS, but have not yet demonstrated either thepresence or the absence of electrical synapses betweenregenerating fibres and surviving distal stumps. Ross et al.(1994) found no evidence for reconnection of proximal anddistal axonal segments in an A. californica buccal motorneurone after nerve crush, but they did find that the distalstumps survived for many weeks. It will be important toperform a systematic series of biocytin injections into VCsensory neurones during the period when reflex responses firststart to recover (our biocytin injections were all 3 weeks post-crush) to see whether there is a sudden appearance of biocytin-filled distal axons extending far beyond the crush site. Suchtests have demonstrated dye-coupling of proximal and distalaxon stumps in the leech (Carbonetto and Muller, 1977; Camhiand Macagno, 1991).
If the formation of electrical synapses plays a role inreconnecting the tail to the CNS, this mechanism for functionalrecovery is likely to be temporary. Injury-induced formation ofnovel electrical synapses has been shown to be transient inother molluscs (e.g. Cohan et al. 1987). Furthermore, the largeglobular inclusions we observed distal to the crush resemblethose seen in other injured A. californica axons and these havebeen linked to axonal degeneration, which occurs over a periodof weeks (Ross et al. 1994). Regenerative growth of newaxonal segments and slow degeneration of old axonal segmentsdistal to the crush would indicate that over several weeks newgrowth into the receptive field gradually takes over sensoryfunctions that might be temporarily maintained by electricalsynapses between proximal and distal segments of severedsensory axons.
Central sprouting by sensory neurones after peripheral axoninjury
In the preceding paper, we report the unexpected discoverythat previously injured VC sensory neurones can sometimes beactivated by stimulating central connectives or peripheralnerves that do not normally contain axons of the tested sensoryneurones (Dulin et al. 1995). As shown in Fig. 7, dye-fills ofsensory neurone somata in the tail/p9 region of the VC clusteripsilateral to crushed pedal nerves reveal fibres entering tracts(the pleural–abdominal and pleural–cerebral connectives) intowhich the sensory neurones in this region do not normallyproject. These observations, and the lack of fibres enteringthese tracts on the uncrushed side, indicate that peripheral axoninjury can trigger the sprouting of new fibres from these cellswithin the CNS. We do not yet know the destination of the newfibres, but it will be interesting to test (1) whether they followalternative paths back to the original receptive field, and (2)whether they form synapses with other neurones that may beinvolved in recovery of function or protection of the peripheralregion affected by the nerve injury (see Dulin et al. 1995).
Central sprouting of mechanosensory neurones in response
to peripheral axotomy has also been described in the leech(Bannatyne et al. 1989) and mammals (Cameron et al. 1992;Woolf et al. 1992). In the latter case, the new growth was nearthe region of the synaptic terminals of the sensory neurones,suggesting the formation of new synapses in response to injury(see also Walters, 1994). Interestingly, in A. californica,unilateral nerve crush causes facilitation of sensory neuronesynapses that lasts for weeks (Walters et al. 1991; Clatworthyand Walters, 1994). Long-term behavioural sensitization (asimple form of learning) in A. californica and long-termsynaptic facilitation induced by application of 5-hydroxytryptamine or cyclic AMP are associated with anincreased arborization and growth of new synapses bymechanosensory neurones (Bailey and Chen, 1983, 1988;Nazif et al. 1991; Bailey et al. 1992). To what extent do themorphological and physiological similarities in cellularreactions to peripheral injury and cellular responses duringlearning reflect shared mechanisms? Explicit comparativestudies of injury-induced and learning-induced alterations inmechanosensory neurones in A. californica and other animalsmay provide insight, not only into mechanisms of regenerationbut perhaps also into fundamental mechanisms of plasticitythat are also utilized in some forms of learning and memory.
The authors are grateful to J. Pastore and K. Hensley for aidin the preparation of illustrations. This work was supported byan NSERC Canada research grant to C.E.M., and grants IBN-9210268 from the NSF and MH38726 from the NIMH toE.T.W. I.S. was the recipient of an Ontario GraduateScholarship.
ReferencesALLISON, P. AND BENJAMIN, P. R. (1985). Anatomical studies of
central regeneration of an identified molluscan interneuron. Proc.R. Soc. Lond. 226, 135–157.
AMBRON, R. T., DEN, H. AND SCHACHER, S. (1985). Synaptogenesisby single identified neurons in vitro: Contribution of rapidlytransported and newly synthesized proteins. J. Neurosci. 5,2857–2865.
BAILEY, C. H. AND CHEN, M. (1983). Morphological basis of long-term habituation and sensitization in Aplysia. Science 220, 91–93.
BAILEY, C. H. AND CHEN, M. (1988). Long-term memory in Aplysiamodulates the total number of varicosities of single identifiedsensory neurons. Proc. natn. Acad. Sci. U.S.A. 85, 2373–2377.
BAILEY, C. H. AND KANDEL, E. R. (1993). Structural changesaccompanying memory storage. A. Rev. Physiol. 55, 397–426.
BAILEY, C. H., MONTAROLO, P., CHEN, M., KANDEL, E. R. AND
SCHACHER, S. (1992). Inhibitors of protein and RNA synthesisblock structural changes that accompany long-term heterosynapticplasticity in Aplysia. Neuron 9, 749–758.
BANNATYNE, B. A., BLACKSHAW, S. E. AND MCGREGOR, M. (1989).New growth elicited in adult leech mechanosensory neurones byperipheral axon damage. J. exp. Biol. 143, 419–434.
BILLY, A. J. AND WALTERS, E. T. (1987). Receptive field plasticity andsomatotopic organization of pleural mechanosensory/nociceptiveneurons of Aplysia. Soc. Neurosci. Abstr. 13, 1393.
BILLY, A. J. AND WALTERS, E. T. (1989). Long-term expansion and

2077Regeneration and sprouting of sensory neurones
sensitization of mechanosensory receptive fields in Aplysia supportan activity-dependent model of whole-cell sensory plasticity. J.Neurosci. 9, 1254–1262.
BISBY, M. A. AND KEEN, P. (1986). Regeneration of primary afferentneurons containing substance P-like immunoreactivity. Brain Res.365, 85–95.
BRUNET, J. F., SHAPIRO, E., FOSTER, S. A., KANDEL, E. R. AND IINO,Y. (1991). Identification of a peptide specific for Aplysia sensoryneurons by PCR-based differential screening. Science 252,856–859.
BULLOCH, A. G. M. AND RIDGWAY, R. L. (1989). Neuronal plasticityin the adult invertebrate nervous system. J. Neurobiol. 20, 295–311.
CAMERON, A. A., POVER, C. M., WILLIS, W. D. AND COGGESHALL, R. E. (1992). Evidence that fine primary afferent axons innervate awider territory in the superficial dorsal horn following peripheralaxotomy. Brain Res. 575, 151–154.
CAMHI, J. M. AND MACAGNO, E. R. (1991). Using fluorescencephotoablation to study the regeneration of singly cut leech axons.J. Neurobiol. 22, 116–129.
CARBONETTO, S. AND MULLER, K. J. (1977). A regenerating neuronein the leech can form an electrical synapse on its severed axonsegment. Nature 267, 450–452.
CARROW, G. M. AND LEVITAN, I. B. (1989). Selective formation andmodulation of electrical synapses between cultured Aplysianeurons. J. Neurosci. 9, 3657–3664.
CLATWORTHY, A. L. AND WALTERS, E. T. (1994). Comparativeanalysis of hyperexcitability and synaptic facilitation induced bynerve injury in two populations of mechanosensory neurones ofAplysia californica. J. exp. Biol. 190, 217–238.
COHAN, C. S., HAYDON, P. G., MERCIER, A. J. AND KATER, S. B.(1987). Formation, maintenance and functional uncoupling ofconnections between identified Helisoma neurons in situ. J.Neurobiol. 18, 329–341.
DULIN, M. F., BILLY, A. J., STEFFENSEN, I. AND WALTERS, E. T. (1994).Somatotopic organization and injury-induced sprouting in pleuralsensory neurons of Aplysia. Soc. Neurosci. Abstr. 20, 230.
DULIN, M. F., STEFFENSEN, I., MORRIS, C. E. AND WALTERS, E. T.(1995). Recovery of function, peripheral sensitization and sensoryneurone activation by novel pathways following axonal injury inAplysia californica. J. exp. Biol. 198, 2055–2066.
EWADINGER, N. M., SYED, N. I., LUKOWIAK, K. AND BULLOCH, A. G.M. (1994). Differential tracer coupling between pairs of identifiedneurones of the mollusc Lymnaea stagnalis. J. exp. Biol. 192,291–297.
FAWCETT, J. W. AND KEYNES, R. J. (1990). Peripheral nerveregeneration. A. Rev. Neurosci. 13, 43–60.
FREDMAN, S. M. (1988). Recovery of escape locomotion following aCNS lesion in Aplysia. Behav. neural Biol. 49, 261–279.
GLANZMAN, D. L., KANDEL, E. R. AND SCHACHER, S. (1989). Identifiedtarget motor neuron regulates neurite outgrowth and synapseformation of Aplysia sensory neurons in vitro. Neuron 3, 441–50.
GLANZMAN, D. L., KANDEL, E. R. AND SCHACHER, S. (1990). Target-dependent structural changes accompanying long-term synapticfacilitation in Aplysia neurons. Science 249, 799–802.
GUNSTREAM, J. D., CASTRO, G. A. AND WALTERS, E. T. (1995).Retrograde transport of plasticity signals in Aplysia sensoryneurons following axonal injury. J. Neurosci. 15, 439–448.
HICKIE, C. P. (1994). Functional and motor neuronal analysis ofdefensive siphon responses in Aplysia californica. PhD thesis,University of Texas Houston Graduate School of BiomedicalSciences.
JENKINS, R., MCMAHON, S. B., BOND, A. B. AND HUNT, S. P. (1993).Expression of c-Jun as a response to dorsal root and peripheralnerve section in damaged and adjacent intact primary sensoryneurons in the rat. Eur. J. Neurosci. 5, 751–759.
KANDEL, E. R. (1979). Behavioral Biology of Aplysia. San Francisco:W. H. Freeman & Co.
KANJE, M., FRANSSON, I., EDSTROM, A. AND LOWKVIST, B. (1986).Ornithine decarboxylase activity in dorsal root ganglia ofregenerating frog sciatic nerve. Brain Res. 381, 24–28.
KELLY, M. E. M., BULLOCH, A. G. M., LUKOWIAK, K. AND BISBY, M. A. (1989). Regeneration of frog sympathetic neurons isaccompanied by sprouting and retraction of intraganglionicneurites. Brain Res. 477, 363–368.
KRASNE, F. B. AND GLANZMAN, D. L. (1995). What we can learn frominvertebrate learning. A. Rev. Psychol. 46, 585–624.
KRUK, P. J. AND BULLOCH, A. G. M. (1992). Axonal regeneration ofan identified Helisoma neuron depends on the site of axotomy. J.Neurosci. Res. 31, 401–412.
MEAROW, K. M., KRIL, Y., GLOSTER, A. AND DIAMOND, J. (1994).Expression of NGF receptor and GAP-43 mRNA in DRG neuronsduring collateral sprouting and regeneration of dorsal cutaneousnerves. J. Neurobiol. 25, 127–142.
MOFFETT, S. AND SNYDER, K. (1985). Behavioral recovery associatedwith central nervous system regeneration in the snail Melampus. J.Neurobiol. 16, 193–209.
NAZIF, F. A., BYRNE, J. H. AND CLEARY, L. J. (1991). cAMP induceslong-term morphological changes in sensory neurons of Aplysia.Brain Res. 539, 324–327.
OBLINGER, M. M. AND LASEK, R. J. (1984). A conditioning lesion ofthe peripheral axons of dorsal root ganglion cells acceleratesregeneration of only their peripheral axons. J. Neurosci. 4,1736–1744.
REHDER, V., JENSEN, J. R. AND KATER, S. B. (1992). The initial stagesof neural regeneration are dependent upon intracellular calciumlevels. Neuroscience 51, 565–574.
RIDGWAY, R. L., SYED, N. I., LUKOWIAK, K. AND BULLOCH, A. G.(1991). Nerve growth factor (NGF) induces sprouting of specificneurons of the snail, Lymnaea stagnalis. J. Neurobiol. 22, 377–390.
ROSS, T. L., GOVIND, C. K. AND KIRK, M. D. (1994). Neuromuscularregeneration by buccal motoneuron B15 after peripheral nervecrush in Aplysia californica. J. Neurophysiol. 72, 1897–1910.
ROTSHENKER, S. (1988). Multiple modes and sites for the induction ofaxon growth. Trends Neurosci. 11, 363–366.
SCHACHER, S. AND PROSHANSKY, E. (1983). Neurite regeneration byAplysia neurons in dissociated cell culture: modulation by Aplysiahemolymph and the presence of the initial axonal segment. J.Neurosci. 12, 2403–2413.
SYED, N. I., RIDGWAY, R. L., LUKOWIAK, K. AND BULLOCH, A. G. M.(1992). Transplantation and functional integration of an identifiedrespiratory interneuron in Lymnaea stagnalis. Neuron 8, 767–774.
VAN ESSEN, D. AND JANSEN, J. K. (1977). The specificity of re-innervation by identified sensory and motor neurons in the leech.J. comp. Neurol. 171, 433–454.
WALTERS, E. T. (1994). Injury-related behavior and neuronalplasticity: an evolutionary perspective on sensitization,hyperalgesia and analgesia. Int. Rev. Neurobiol. 36, 325–427.
WALTERS, E. T., ALIZADEH, H. AND CASTRO, G. A. (1991). Similarneuronal alterations induced by axonal injury and learning inAplysia. Science 253, 797–799.
WALTERS, E. T. AND AMBRON, R. T. (1995). Long-term alterationsinduced by injury and by 5-HT in Aplysia sensory neurons:

2078 I. STEFFENSEN AND OTHERS
Convergent pathways and common signals? Trends Neurosci. 18(in press).
WALTERS, E. T., BYRNE, J. H., CAREW, T. J. AND KANDEL, E. R.(1983). Mechanoafferent neurons innervating tail of Aplysia. I.Response properties and synaptic connections. J. Neurophysiol. 50,1522–1542.
WEDDELL, G., GUTTMANN, L. AND GUTTMANN, E. (1941). The localextension of nerve fibers into denervated areas of skin. J. Neurol.Neurosurg. 4, 206–225.
WOOLF, C. J., SHORTLAND, P. AND COGGESHALL, R. E. (1992).Peripheral nerve injury triggers central sprouting of myelinatedafferents. Nature 355, 75–78.
WUJEK, J. R. AND LASEK, R. J. (1983). Correlation of axonalregeneration and slow component B in two branches of a singleaxon. J. Neurosci. 3, 243–251.
ZHANG, H., BYRNE, J. H. AND CLEARY, L. J. (1993). Topographicalorganization of sensory neurons in the ventrocaudal cluster of thepleural ganglion of Aplysia. Soc. Neurosci. Abstr. 19, 813.