peptidyl argininedeiminase 2 cpg island in multiple sclerosis white matter is hypomethylated
TRANSCRIPT

Peptidyl Argininedeiminase 2 CpG Islandin Multiple Sclerosis White Matter IsHypomethylated
Fabrizio G. Mastronardi,1* Abdul Noor,2,3 D. Denise Wood,1 Tara Paton,2
and Mario A. Moscarello1
1Molecular Structure and Function, The Hospital for Sick Children, Toronto, Ontario, Canada2The Centre for Applied Genomics (TCAG), The Hospital for Sick Children, Toronto, Ontario, Canada3Institute of Medical Science, Faculty of Medicine, University of Toronto, Ontario, Canada
In previous studies, we documented increased citrulli-nated myelin basic protein (MBP) was present in MBPisolated from multiple sclerosis (MS) normal appearingwhite matter (NAWM). This increase was due to themyelin enzyme peptidyl argininedeiminase 2 (PAD2). Inthis study, we show that methylation of cytosine of thePAD2 promoter in DNA from MS NAWM was decreasedto one-third of the level of that in DNA from normalwhite matter. The PAD2 promoter in DNA from thymusobtained from the same MS patients and white matterDNA from Alzheimer’s, Huntington’s, and Parkinson’swas not hypomethylated. DNA demethylase activity insupernatants prepared from NAWM of MS patients was2-fold higher than the DNA demethylase from normal,Alzheimer’s, Huntington’s and Parkinson’s diseasewhite matter. The amount of PAD2 enzyme and citrulli-nated MBP was increased in MS NAWM. Thedecreased methylation of cytosines in the PAD2 pro-moter may explain the increased synthesis of PAD2protein that is responsible for the increased amount ofcitrullinated MBP, which in turn results in loss of myelinstability in MS brain. VVC 2007 Wiley-Liss, Inc.
Key words: multiple sclerosis; peptidyl argininedei-minase; DNA methylation; DNA demethylase; CpG island
Multiple sclerosis (MS) is a complex disease of thecentral nervous system (CNS) that involves genetic, envi-ronmental, and immune mediators. Most studies on diseasemechanisms have focused primarily on the autoimmunecomponent and in the characterization of the pathologicallesions. Recently, differences in radiologic parameters ofMS patients and biochemical characteristics of the normalappearing white matter (NAWM) from living MS patientsand post-mortem NAWM are suggestive of an alteredmyelin structure (Mastronardi and Moscarello, 2005). Bio-chemical changes in NAWM before demyelination wereconsistent with radiologic data from MS patients and hasbeen reviewed recently (Ludwin, 2006).
A consistent biochemical change in MS NAWMhas been the presence of elevated citrullinated myelinbasic protein (MBPcit) (Moscarello et al., 1994). Protein
citrullination is mediated by a family of enzymes, thepeptidylarginine deiminases (PAD) (EC 3.5.3.15). Wehave shown previously that PAD2 expression (proteinand mRNA) was elevated before neurologic signs andbefore MBP citrullination in transgenic mice (Moscarelloet al., 2002). PAD2 in brain homogenates from 1-month-old transgenic mice had a specific activity 1.8-fold higher than non-transgenic littermates and had a14.6-fold more PAD2 protein in myelin (Moscarelloet al., 2002). These studies indicated that an increasedamount of PAD2 protein might be the result of elevatedgene expression of this enzyme.
The PAD genes are clustered in a distinct genomicregion in humans (1p36.1) and syntenic locations ofrodent genomes(Vossenaar et al., 2003). The PAD iso-zymes 1–4 are expressed in adult tissue. One of thePAD family members, PAD2 isolated from brain, hasbeen well characterized. It is found in white matter(Lamensa and Moscarello, 1993) where it deiminatesMBP (Finch et al., 1971; Lamensa and Moscarello,1993). An examination of the promoter region of the ratand mouse PAD2 genes showed the presence of a CpGisland (cytosine/guanosine) of 217 base pairs and a CGcontent of 74% (Watanabe et al., 1992; Chavanas et al.,2004), proximal to the transcription start site.
Methylation on CpG islands is an important regula-tory mechanism for gene expression in DNA. TheseCpG islands in the promoter region of some genes havea CG content greater than 60% compared to 40% for
Supplementary Material for this article is available online at http://
www.mrw.interscience.wiley.com/suppmat/0360-4012/suppmat/(www.
interscience.wiley.com).
Contract grant sponsor: MS Society of Canada.
*Correspondence to: Dr. Fabrizio G. Mastronardi, Molecular Structure
and Function, The Hospital for Sick Children, Toronto, ON M5G 1X8,
Canada. E-mail: [email protected]
Received 26 January 2007; Revised 19 February 2007; Accepted 20
February 2007
Published online 27 April 2007 in Wiley InterScience (www.
interscience.wiley.com). DOI: 10.1002/jnr.21329
Journal of Neuroscience Research 85:2006–2016 (2007)
' 2007 Wiley-Liss, Inc.

the bulk DNA. The mammalian genome contains about30,000 such islands, which are found proximal to tran-scription start sites and extend 50 along promoter regions.Methylation of the cytosine in CpG islands turns off pro-moter activity thereby silencing genes (Cedar, 1988; Yei-vin and Razin, 1993). In the methylation reaction, the car-bon on position 5 of the cytosine ring becomes methylatedby a methyltransferase in the presence of S-adenosylmethi-onine. In contrast to protein (arginine) methylation that isnon-reversible, cytosine methylation can be reversed by aDNA demethylase (Eden and Cedar, 1994; Hsieh, 2000;Szyf and Bhattacharya, 2002; Pennings et al., 2005).
Because the human PAD2 gene has 74% CG con-tent in the proximal promoter region, silencing of thePAD2 gene by methylation may represent a mechanismfor controlling the amount of PAD2 formed and subse-quently the amount of citrullinated MBP. We show thatthe proximal upstream region of the PAD2 gene in MSwhite matter is hypomethylated. These results suggestthat the elevated PAD2 protein found in MS brain maybe the result of decreased methylation of its promoter.
MATERIALS AND METHODS
Tissue Samples
Cortical white matter samples from brain and thymustissue from normal and MS patients and cortical white matterfrom Alzheimer’s, Huntington’s, and Parkinson’s patients wereobtained from the Human Brain and Spinal Fluid ResourceCenter (HSB) at UCLA. The MS samples did not containplaque material and each one of them was accompanied byphotographic documentation of the dissected area. The clinicaldiagnosis, pathology, and autolysis times for the frozen whitematter samples are summarized in Supplementary Table I.
Immunofluorescent Labeling of White Matter FromNormal and MS Patients
Formalin-fixed white matter tissue sections from normaland MS patients were examined by immunofluorescence. Af-ter deparaffinization and rehydration, the sections were incu-bated in 0.05% Triton X-100 Tris buffered saline (pH 7.4)overnight at 48C. The primary antibody to MBP (MAb26)(Groome et al., 1988) was used at a 1:500 dilution and poly-clonal antibodies to all PAD isozymes were used at a 1:200dilution and incubated for 1 hr at room temperature in amoisture chamber. Excess primary antibodies were removedand secondary chicken anti-mouse and anti-rabbit IgG anti-bodies, conjugated with Alexa Fluor 488 and 594 (MolecularProbes, Eugene, OR), were diluted 1:200 with 2% normalgoat serum in PBS pH 7.4. After 1 hr, slides were washedwith PBS three times, followed by a 10-min wash with fil-tered deionized water to remove salts, and mounted withDako Cytomation Fluorescent mounting medium (DakoCyto-mation, Glostrup, DK). Pictures were taken using an OlympusBX51 fluorescence microscope equipped with a CCD camera.
Measurement of PAD2 and Citrullinated Protein Levelsin White Matter From Normal and MS Patients
PAD2 protein and citrullinated proteins were quanti-tated by an immunoslot blot method. White matter homoge-
nates were diluted in PBS (pH 7.4) containing 3M urea. Forquantification of the PAD2 protein, 10 lg of homogenateprotein diluted to 100 ng/mL protein were loaded in triplicateonto wetted nitrocellulose membranes in a Biodot (SF) ultrafiltration unit (BioRad, Hercules, CA) under vacuum. Primaryaffinity purified PAD2 antibody (Raijmakers et al., 2005) wasused at 1:1,000 dilution. For citrullinated proteins, 5 lg whitematter homogenate proteins were loaded in each slot. Citrulli-nated proteins were detected using the modified citrullinedetection kit according to the manufacture’s instructions(Upstate Biotechnology, Lake Placid, NY). Slot blots weredeveloped with the ECL Western blotting detection reagent(Amersham Biosciences, Little Chalfont, Buckinghamshire,UK). Blots were scanned using Image SXM image analysis(based on the NIH image software program) software for thePower Mac and the relative amounts of bound antibody wasdetermined for each protein and compared to the respectiveamounts in normal brain.
Western Blot of Myelin Basic Protein
Basic proteins from white matter were prepared asdescribed earlier (Cheifetz and Moscarello, 1985). Peptidyl-citrulline content in white matter basic protein extracts wasdetermined by Western blot (Towbin et al., 1979). Proteinswere separated through 10–20% tricine polyacrylamide gels(Invitrogen, Carlsbad, CA) by electrophoresis and Westernblotted onto nitrocellulose membranes. The membranes wereincubated with primary antibodies (either anti-MBP, mono-clonal MAb26, or with anti-peptidyl citrulline). Peptidyl cit-rulline levels in the basic protein fractions were detected usingthe modified citrulline detection kit according to the manu-facture’s instructions (Upstate Biotechnology). Primary anti-bodies were detected with species specific IgG secondary anti-bodies conjugated with horseradish peroxidase (hrp). Theenzyme linked secondary antibodies were detected using ECLsubstrate after exposure to X-ray film.
Measurement of 5-Methyl Cytosine in White Matterand Thymus Homogenates
Measurement of the bulk 5-methyl cytosine in tissuehomogenates was done by immuno slot blots. Homogenates(5 lg/slot) prepared as above were loaded onto a methanolwetted PVDF membrane. The DNA was cross-linked ontoPVDF membranes with UV light using a DNA Stratalinker(Stratagene, La Jolla, CA). The bound DNA was denaturedby incubating in cold (48C) methanol for 30 min followed byincubation with 7 3 10�3 N NaOH for approximately15 sec. Membranes were incubated with primary anti-5-methyl cytosine polyclonal IgG (Ab1884) (Abcam Inc., Cam-bridge, MA) diluted 1:1,000 in Tris-buffered saline pH 7.4 þ0.05% Tween 20 (TBST) overnight at 48C. Histone H3immunoslot blots were used as loading controls for homoge-nates. Histone H3 was detected with anti-histone H3 antibody(Ab1791) (Abcam Inc.) diluted 10,000 fold in blotto. Boundprimary antibody was detected with secondary anti-sheeppolyclonal antibody linked to hrp (GT-34) (Sigma Aldrich,Oakville, ON) diluted 1,000-fold in TBST. Detection of hrp
DNA Hypomethylation in MS White Matter 2007
Journal of Neuroscience Research DOI 10.1002/jnr

was with ECL reagent as above. X-ray films were scannedand analyzed using Image SXM analysis software.
CpG Island Analysis of the Human PAD2 Locus
The human PAD2 gene was identified by sequencehomology search of NCBI clone RPI-37C10 using a 50 por-tion of the human PAD2 cDNA. This clone contains thecomplete sequence of the human PAD2 gene. The flankingregulatory region spanning 1,100 bp from the Methioninestart codon was analyzed for CG content and was used as areference sequence for PCR primer design.
Genomic DNA Preparation
Genomic DNA was prepared by mincing NAWM orthymus tissue in protease K buffer (200 lg/sample) and digest-ing overnight at 558C. The following day RNA was digestedby adding 200 lg of heat-treated RNase/sample to the diges-tion mixture and incubated for 1 hr at 378C. DNA wasextracted by Trizma (pH 7.4) saturated phenol (Invitrogen):-chloroform (1:1 v/v). The extractions were repeated twicefollowed by precipitation of genomic DNA with one half-vol-ume 5 M ammonium acetate and two volumes 100% anhy-drous ethanol. The DNA was pelleted and solubilized in dou-ble distilled deionized water (dddw). The concentration ofDNA was determined spectrophotometrically by determiningthe absorbance at 260 nm.
Design of Primers for PCR and Sequencing
Genomic sequence containing the predicted CpG islandand 100 bp of 50 and 30 flanking sequence for PAD2 wasobtained from the UCSC Genome Browser, May 2004Human assembly (http://genome.ucsc.edu). Primers foramplification of bisulfite treated DNA were designed from thepredicted sequence using the program MethPrimer (http://www.urogene.org/methprimer/index1.html), using the CpGisland prediction option and allowing product sizes up to500 bp. Two primer sets were selected from the results outputand tested for PCR efficiency. Based on this testing, the for-ward primer (50 GGGTTGGATAAAGGTTGTTTA 30) andreverse primer (50 CACAAAACTAATCCCTAATAACTTC30) were chosen for PCR after bisulfite treatment of genomicDNA. The same primers were used for sequencing of all thePCR amplified DNA samples.
Bisulfite Treatment and PCR of Genomic DNA
To convert non-methylated cytosine residues to uracil,2.0 lg of genomic DNA was treated with bisulfite using theEZ DNA Methylation Kit (Cedarlane, Hornby, ON) follow-ing manufacturer’s instructions. Treated DNA was PCR-amplified with primers designed for bisulfite-modified DNAusing a hot-start protocol (Qiagen, Mississauga, Canada). ThePCR reaction consisted of 3.0 lL of bisulfite-treated DNA,13 Qiagen buffer, 5.0 lL Q-solution, 160 lM dNTP, 0.125lL HotstarTaq DNA polymerase, and 50 ng each primer in a25 lL total volume. Cycling conditions were as follows: 958Cfor 10 min, followed by 35 cycles of 958C for 45 sec, 588Cfor 45 sec, and 728C for 45 sec, with a final extension step at728C for 7 min. PCR products were visualized after separa-
tion through 2.0% agarose gels and purified using Microclean(Microzone, Haywards Heath, UK). Purified PCR productswere sequenced using BigDye 3.1 chemistry and analyzedwith the 3730 XL DNA Analyzer (Applied Biosystems, FosterCity, CA).
Measurement of 5-Methyl Cytosine in White Matter
Methylated cytosines were measured by direct sequenc-ing and analysis of electrophoretograms. Because individualPCR clones were not prepared the sequencing results wereexpected to generate mixed sequences. These sequences werelabeled by the sequencing program ABI (Applied Biosystems)as ‘‘Y’’ indicative as either a T or C residue. Mixed sequenceswere visually scored as half methylated cytosines when at leasthalf the peak corresponded to either residue. The accumulatedcytosines were recorded (Suppl. Table I).
DNA Demethylase Assay
DNA demethylase activity was measured using themethod described by Szyf and Bhattacharya (2002) with mod-ifications for white matter fractions.
Preparation of single stranded DNA. Methylateddeoxy oligonucleotide (m5-dCdG)24 forward and reverse(dGp-m5-C)24, and non-methylated deoxy oligonucleotide(dCdG)24 forward and reverse (dGdC)24 oligomers were syn-thesized by The Hospital for Sick Children’s DNA facility(Centre for Applied Genomics).
Preparation of the [a-32P] fully methylated5mdCpG and non-methylated DNA templates. Singlestranded DNA templates (non-methylated and methylated)were used in labeling reactions. In the following order, 20 ngof 5mdCpG primer was added to methylated template and 20ng of (CpG) 6 primer (control) (Hospital for Sick Children’sDNA Facility’s Centre for Applied Genomics), 5 ll of 103DNA polymerase1 buffer, 5 ll of [a-32P] dGTP (AmershamBiosciences, Boie d’Urfe, Que), incubated at 378C for 1.5 hr.The unincorporated nucleotides were removed by chromatog-raphy through a NAP-5 column (Amersham Biosciences,Boie d’Urfe, Canada).
Preparation of white matter homogenates forDNA demethylase assays. White matter tissue extracts pre-pared from normal (n ¼ 4) and patients with MS (n ¼ 12)were homogenized in 500 ll of lysis solution (1 M Tris, pH 8;1 M MgCl2; 5 M NaCl; 10% Tween-20; 100% glycerol;dddw). Homogenates were pelleted by centrifuging and thesupernatants were used in the demethylase assays. The superna-tants were diluted in lysis buffer (1:9) and 5ll aliquots wereremoved to determine protein concentrations (Peterson, 1977).
Demethylase assay. One microliter of non-methyl-ated or methylated substrate was added to 30 ll of lysis solu-tion and 5–10 ll of supernatant. The solution was incubatedat 378C for 24 hr, then, 100ll of water, 30 ll of 5 M NaCl,and 2 ll glycogen were added. DNA was recovered by phe-nol extraction as follows: 75 ll of buffer saturated phenol(Invitrogen) and 75 ll of chloroform were added to the mix-tures. The samples were vortexed and then pelleted by centri-fuging for 5 min. The top aqueous phase was recovered andtransferred to a tube containing 300 ll anhydrous ethanol andplaced on dry ice for 15 min. The DNA was pelleted by cen-
2008 Mastronardi et al.
Journal of Neuroscience Research DOI 10.1002/jnr

trifuging for 15 min at full speed (12,000 3 g) and the super-natant removed. The sides of the tubes were dried and theDNAs were solubilized in 8 ll of dddw, 1 ll of 103 micro-coccal nuclease buffer, 1 ll of spleen phosphodiesterase, and1 ll of micrococcal nuclease (Amersham Biosciences). Thesamples were incubated at 378C overnight. Nucleotides in4 ll of each sample were resolved by thin layer chromato-graphy (TLC) using an isobutyric acid, water and ammoniasolution (75, 22.7, 2.3 v/v/v). X-ray film was exposed to theTLC plates and developed either 24 hr or 48 hr later. TheX-ray films were scanned and the ratio of non-methylatedcytidylic acid to 5-methyl cytidylic acid was determined withimage analysis software SXM image (based on NIH Image)for the Power Macintosh.
Statistical Methods
Data were analyzed with two-tailed non-parametrictests. Significance was set at 5%. MS and non-neurologic con-trols as groups were compared using unpaired t-tests withWelch correction or Mann-Whitney tests using Graph PadPrism 4.03 and Instat software for the Power Mac.
RESULTS
PAD2 Protein in MS Myelin Is Elevated
We reported previously that MS NAWM con-tained an elevated myelin basic protein (Moscarelloet al., 1994) component termed C8 (Wood and Moscar-ello, 1989) that contained citrulline instead of arginine atsix positions (Wood and Moscarello, 1989; Moscarelloet al., 1994; Kim et al., 2003). Because there is nocodon that allows for the addition of citrulline duringtranslation, this modification takes place post translation-ally. The enzyme in myelin responsible for this conver-sion is PAD2 (Lamensa and Moscarello, 1993). Todetermine whether PAD2 was affected in MS NAWMwe examined fixed tissues by immunofluorescence andfrozen tissue was used for biochemical measurements.
Immunofluorescent labeling of normal white mat-ter (Fig. 1A) and MS NAWM (Fig. 1B) with PAD2polyclonal antibody (green) and MBP monoclonal anti-body (MAb26) (Groome et al., 1988) (red) resulted in ayellow label where both MBP and PAD labeling over-lapped. The PAD2 staining pattern in the MS brainshowed a pronounced clustering (arrows) around themyelin, whereas the staining pattern in the normal alsoshowed a clustering pattern but had a more discrete dis-tribution. Because the immuno fluorescence data wasqualitative, but suggestive of elevated PAD2 in MSNAWM we conducted quantitative and biochemicalassays in MS NAWM and in white matter from normaland neurodegenerative patients.
To determine whether MS NAWM contained ele-vated amounts of PAD2 protein we used an affinitypurified PAD2 antibody (Raijmakers et al., 2005) inimmuno slot blots to measure the levels of PAD2 in MSand in control white matter. The quantitation showedan elevation of the amount of PAD2 in the NAWMfrom MS individuals (Fig. 1C). The amount in the MS
group (n ¼ 12) was 3-fold higher (P < 0.0001) than thecontrol. The white matter homogenates from the sameindividuals were assayed subsequently for the product ofthe PAD2 enzyme, citrullinated proteins. The results inFigure 1D showed a 2-fold (P < 0.009) increase of pep-tidyl-citrulline in the MS NAWM group (n ¼ 12). Weshowed previously that citrullinated MBP (MBPcit) wasincreased in MS white matter. To determine whetherMBPcit was increased in the MS samples examined inthis study, whole MBP was acid extracted from whitematter as described previously (Cheifetz and Moscarello,1985), the proteins were Western blotted onto nitrocel-lulose membranes, and MBP was detected with the pri-mary monoclonal antibody (MAb26) (Groome et al.,1988) and peptidyl-citrulline with the commercial citrul-line detection kit. The results shown in Figure 1Eshowed elevated anti-citrulline binding in the basic pro-teins from MS individuals. These results were consistentwith the increased citrullination of MBP from MS whitematter reported previously (Moscarello et al., 1994;Wood et al., 1996; Kim et al., 2003).
Amount of 5-Methyl Cytosine in MS WhiteMatter Is Reduced
As mentioned above, the PAD2 gene structure inboth humans and rodents contains CpG islands that areposited to be involved in its regulation. To determine ifglobal cytosine methylation was affected in MS NAWM,we used a 5-methyl cytosine specific antibody (Ab 1884)to measure 5-methyl cytosine in white matter homoge-nates from MS and normal individuals. To normalize theloading in each slot, we took the ratio of 5-methyl cyto-sine/histone H3. The results shown in Figure 2Ashowed that the ratio of 5-methyl cytosine/histone H36 SD was significantly lower by 3-fold in the MS ho-mogenates. Analysis of the data from both the MS andnormal groups with non-parametric statistics using theMann-Whitney test resulted in a two-tailed P-value <0.0001. These results are indicative of a significantreduction of genomic cytosine methylation in NAWMin MS.
The availability of thymus tissue from the sameindividuals allowed us to determine whether the differ-ences observed in white matter were restricted to whitematter or were a generalized change. A similar analysisof thymus showed no significant differences in 5-methylcytosine/histone H3 between normal and MS (P <0.26) (Fig. 2A).
The ratio of 5-methyl cytosine/histone H3 wasalso measured in the white matter of individuals withAlzheimer’s, Huntington’s, and Parkinson’s diseases (Fig.2B). The results showed a normal range of 5-methyl cy-tosine/histone H3 ratios with no significant differencesamong the three groups. Although there was a smallreduction in the Alzheimer’s group, the results suggestthat a general reduction of cytosine methylation was asignificant feature of MS white matter and not of otherneurologic diseases.
DNA Hypomethylation in MS White Matter 2009
Journal of Neuroscience Research DOI 10.1002/jnr

Sequence Analysis of the ProximalPAD2 Promoter
One possibility for the elevated PAD2 protein inMS NAWM was DNA sequence mutation associatedwith immediate upstream regulatory elements thatenabled enhanced expression. The recent sequencing ofthe entire human genome facilitated the identification ofthe PAD family of enzymes located on chromosome 1(1p36.13) (Fig. 3A, vertical arrow). The CpG island inPAD2 straddles exon 1 and extends distally. Sequence
alignment searches of the NCBI human genome databaseusing fragments of the untranslated region of the humanPAD2 cDNA identified PAD2 genomic sequences. Analy-sis of this sequence showed a high content of CG consistentwith the presence of a CpG island (Suppl. Fig. 2). Wedesigned forward and reverse primers to amplify a portionof this region using the HotStarTaq DNA Polymerase kit(Qiagen, Germantown, MD) (Suppl. Fig. 1). Sequencing900 þ base pairs of flanking sequence of the human PAD2gene from MS NAWM genomic DNA isolated from five
Fig. 1. Immunofluorescent labeling of PAD2 (green) and MBP (red)in normal (A) and MS (B) white matter tissue sections. Asterisksindicate axons. Arrows in the MS section indicate clustered PAD2label. C: PAD2 protein levels in normal (n ¼ 3) and MS (n ¼ 12)white matter 6 SD D: Citrullinated protein levels in normal (n ¼ 3)and MS (n ¼ 12) white matter 6 SD. E: Western blot of MBPextracts from normal and MS with monoclonal anti-MBP (MAb26)(top panel). Lower panel: Western blots of citrullinated MBP from
normal and MS individuals using the modified citrulline detectionmethod. The 18.5 kDa size marker indicates the relative mass ofhuman MBP. Normal individuals from left to right are: HSB 3280,HSB 3322, HSB 3346; The Multiple Sclerosis samples (left to right)were from: HSB 2429, HSB 3502, HSB 3522, HSB 3509, HSB2800, HSB 2485. The neuropathology for each individual is summar-ized in Supplementary Table I.
2010 Mastronardi et al.
Journal of Neuroscience Research DOI 10.1002/jnr

MS and three control white matter samples showed no dif-ferences in sequence (data not shown).
Methylation of DNA
The goal of this study was to determine whether ageneral change in cytosine methylation existed in MSNAWM at the PAD2 CpG island. We next examinedwhether this region showed methylation differences. Weused the procedure described by Clark (1994) and Grigand Clark (1994) to determine the sites of cytosinemethylation in DNA from CNS white matter and thy-mus. Briefly, the procedure involved the treatment of
genomic DNA with sodium bisulfite, which adds to car-bon 4 of the cytosine ring, followed by hydrolyticdeamination, converting cytosine to uracil in the process.The resulting DNA was amplified by PCR with specificprimers (Materials and Methods) and sequenced. ThePCR amplified region (Amplicon) of the CpG (Fig. 3A)consisted of 385 base pairs, of which 71% was CG. Theamplicon contained a portion of intron 1 (53 bp), all ofexon 1 (160 bp), and extended distally (172 bp) (Suppl.Fig. 2). Using this method, we sequenced the PAD2CpG (Fig. 3A) from MS (n ¼ 17), normal individuals(n ¼ 9), and individuals with other neurodegenerativediseases (Alzheimer’s, n ¼ 4; Huntington’s disease, n ¼2; and Parkinson’s disease, n ¼ 2). The results of thesequencing of the PAD2 CpG is summarized in Figure3B. The data showed significantly less 5-methyl cytosinein the PAD promoter of MS NAWM (P < 0.008) com-pared to normal by 2.7-fold. Typical sequence traces ofbisulfite treated DNA from two normal and two MSindividuals are shown in Supplemental Figure 3.
To determine whether the PAD2 CpG islandmethylation pattern in the MS individuals was specificfor white matter or whether it was also altered in othertissues in MS, we analyzed the methylation pattern inthe PAD2 CpG island from thymus obtained from thesame MS patients (Fig. 3B). After bisulfite treatment ofgenomic DNA and amplification of the same stretch ofPAD2 CpG island we found no differences in methyla-tion (MS, n ¼ 3; normal, n ¼ 3). The direct sequencingresult was consistent with the semi-quantitative measureof global 5-methyl cytosine observed with Ab1884 (Fig.2A). These data suggested that a specific methylationchange in MS white matter was associated with elevatedPAD2.
To determine if the specificity of the changes inPAD2 CpG island methylation in DNA from MS whitematter was disease specific we examined the methylationstatus of the PAD2 CpG island in DNA from whitematter from victims of other neurodegenerative diseases(Supplementary Table I) including, Huntington’s, Alz-heimer’s, and Parkinson’s diseases (Fig. 3C). The datawere indicative that white matter methylation wasreduced significantly only in MS and may be responsiblefor the elevated PAD2.
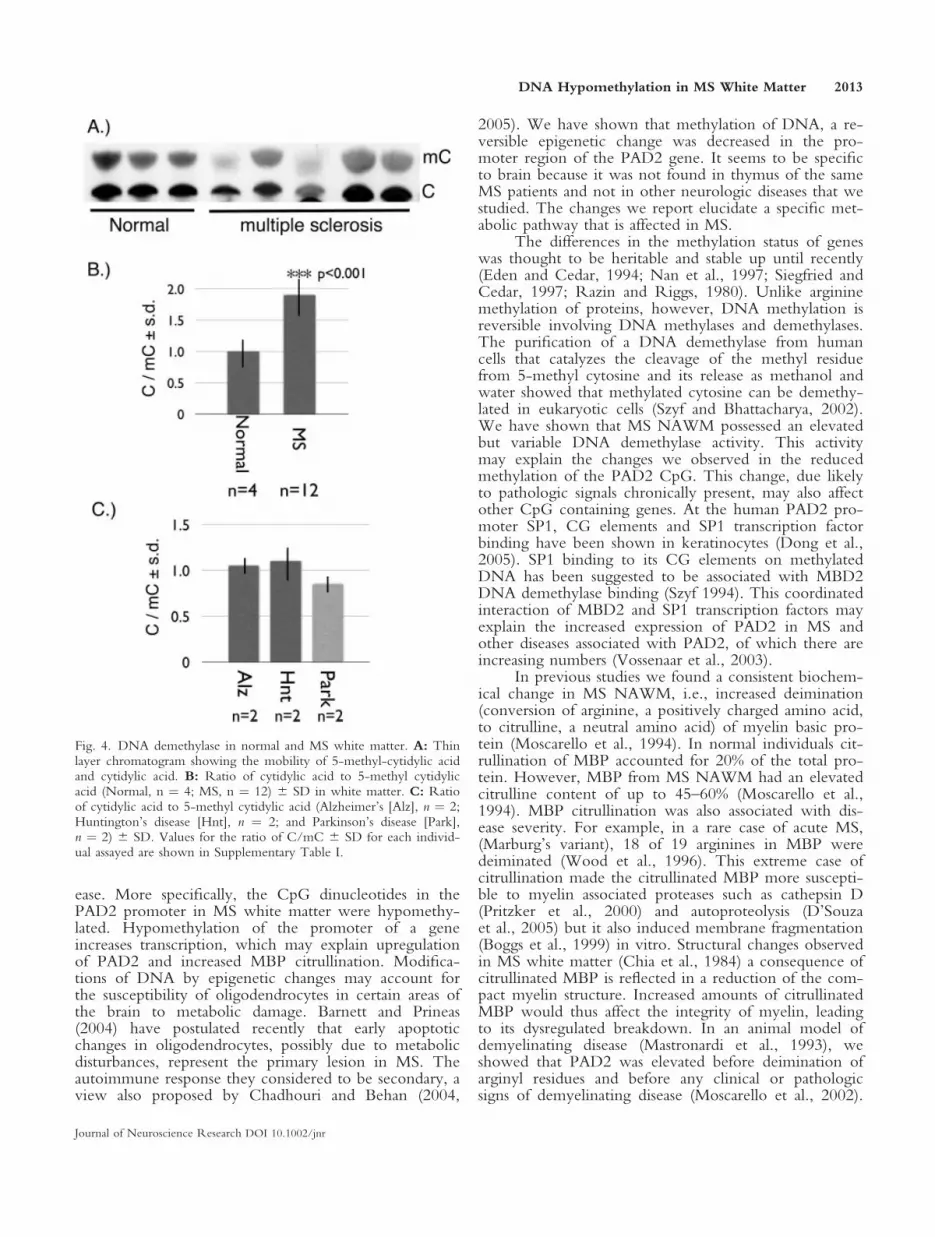
DNA Demethylase in White Matter
To determine if the decreased methylation was theresult of an increase in DNA demethylase activity wemeasured the production of demethylated cytidylic acidfrom a synthetic methylated cytosine oligonucleotidesubstrate (Szyf and Bhattacharya, 2002) as described inMaterials and Methods. Separation of the cytidylic acid(C) from methyl cytidylic acid (mC) by thin layer chro-matography showed a lower amount of mC in MSNAWM (Fig. 4A). Calculation of the ratio of C/mCshow a 2-fold increase in this ratio in MS (Fig. 4B) sug-gesting an increased activity of DNA demethylase. TheDNA demethylase activity in control white matter var-
Fig. 2. 5-Methyl cytosine levels in normal, MS, and neurodegenera-tive diseases by slot blot. A: Level of 5-methyl cytosine/histone H3in normal (n ¼ 4) and MS (n ¼ 12) white matter and thymus 6 SD.B: Level of 5-methyl cytosine/histone H3 in Alzheimer’s (n ¼ 5),Huntington’s (n ¼ 5), and Parkinson’s disease (n ¼ 5) white matter6 SD.
DNA Hypomethylation in MS White Matter 2011
Journal of Neuroscience Research DOI 10.1002/jnr

ied only slightly whereas individual MS patients showeda much higher and more variable activity, possiblyrelated to the severity of the disease.
DISCUSSION
Multiple sclerosis is the most common demyelinat-ing disease of adult humans. Neither the etiology northe pathogenesis is understood, but genetic, immuno-logic, and environmental factors seem to be implicated.The search for genetic factors by screens of MS popula-tions have failed to uncover any strong chromosomal
loci from which it was concluded that several loci, each ofsmall effect, was required for disease (Ebers et al., 1996;Haines et al., 1996; Sawcer et al., 1996; Kuokkanen et al.,1997). Most screens identified the MHC (known fromother studies) but no agreement was obtained for otherloci, prompting Chattaway et al. (1998) to conclude thatthe screens were of ‘‘disappointingly low yield.’’
We have shown that a new genetic complexity inMS white matter involves DNA methylation, an epige-netic change. DNA hypomethylation in MS brain mayexplain some of the biochemical changes observed in theNAWM that contribute to the pathogenesis of this dis-
Fig. 3. PAD2 gene locus and CpG island methylation. A: Ideogramof human chromosome 1 highlighting the position of the PAD2gene (vertical arrow) at position 1p36.13. Sequencing data of ampli-con generated after bisulfite treatment of genomic DNA is encom-passed by the arrows at position �168 to þ226. Thymus and whitematter 5-methyl cytosine amounts are represented by horizontal gra-dient rectangles above and below the proximal region of the PAD2gene. Arrows on intron 1 indicate the direction of transcription. ThePAD2 CpG straddles exon 1 extends into intron 1 and extends along
the flanking promoter proximal region. B: Amount of 5-methyl cy-tosine after bisulfite treatment of genomic DNA and sequence analy-sis of the PAD2 CpG island from normal (n ¼ 9) and MS (n ¼ 17)white matter and normal (n ¼ 3) and MS (n ¼ 3) thymus 6 SD. C:Amount of 5-methyl cytosine of the PAD2 CpG island from Alzhei-mer’s (Alz, n ¼ 4), Huntington’s (Hnt, n ¼ 2), and Parkinson’s dis-ease (Park, n ¼ 2) white matter. Values for the individual patients areshown in Supplementary Table I.
2012 Mastronardi et al.
Journal of Neuroscience Research DOI 10.1002/jnr
ease. More specifically, the CpG dinucleotides in thePAD2 promoter in MS white matter were hypomethy-lated. Hypomethylation of the promoter of a geneincreases transcription, which may explain upregulationof PAD2 and increased MBP citrullination. Modifica-tions of DNA by epigenetic changes may account forthe susceptibility of oligodendrocytes in certain areas ofthe brain to metabolic damage. Barnett and Prineas(2004) have postulated recently that early apoptoticchanges in oligodendrocytes, possibly due to metabolicdisturbances, represent the primary lesion in MS. Theautoimmune response they considered to be secondary, aview also proposed by Chadhouri and Behan (2004,
2005). We have shown that methylation of DNA, a re-versible epigenetic change was decreased in the pro-moter region of the PAD2 gene. It seems to be specificto brain because it was not found in thymus of the sameMS patients and not in other neurologic diseases that westudied. The changes we report elucidate a specific met-abolic pathway that is affected in MS.
The differences in the methylation status of geneswas thought to be heritable and stable up until recently(Eden and Cedar, 1994; Nan et al., 1997; Siegfried andCedar, 1997; Razin and Riggs, 1980). Unlike argininemethylation of proteins, however, DNA methylation isreversible involving DNA methylases and demethylases.The purification of a DNA demethylase from humancells that catalyzes the cleavage of the methyl residuefrom 5-methyl cytosine and its release as methanol andwater showed that methylated cytosine can be demethy-lated in eukaryotic cells (Szyf and Bhattacharya, 2002).We have shown that MS NAWM possessed an elevatedbut variable DNA demethylase activity. This activitymay explain the changes we observed in the reducedmethylation of the PAD2 CpG. This change, due likelyto pathologic signals chronically present, may also affectother CpG containing genes. At the human PAD2 pro-moter SP1, CG elements and SP1 transcription factorbinding have been shown in keratinocytes (Dong et al.,2005). SP1 binding to its CG elements on methylatedDNA has been suggested to be associated with MBD2DNA demethylase binding (Szyf 1994). This coordinatedinteraction of MBD2 and SP1 transcription factors mayexplain the increased expression of PAD2 in MS andother diseases associated with PAD2, of which there areincreasing numbers (Vossenaar et al., 2003).
In previous studies we found a consistent biochem-ical change in MS NAWM, i.e., increased deimination(conversion of arginine, a positively charged amino acid,to citrulline, a neutral amino acid) of myelin basic pro-tein (Moscarello et al., 1994). In normal individuals cit-rullination of MBP accounted for 20% of the total pro-tein. However, MBP from MS NAWM had an elevatedcitrulline content of up to 45–60% (Moscarello et al.,1994). MBP citrullination was also associated with dis-ease severity. For example, in a rare case of acute MS,(Marburg’s variant), 18 of 19 arginines in MBP weredeiminated (Wood et al., 1996). This extreme case ofcitrullination made the citrullinated MBP more suscepti-ble to myelin associated proteases such as cathepsin D(Pritzker et al., 2000) and autoproteolysis (D’Souzaet al., 2005) but it also induced membrane fragmentation(Boggs et al., 1999) in vitro. Structural changes observedin MS white matter (Chia et al., 1984) a consequence ofcitrullinated MBP is reflected in a reduction of the com-pact myelin structure. Increased amounts of citrullinatedMBP would thus affect the integrity of myelin, leadingto its dysregulated breakdown. In an animal model ofdemyelinating disease (Mastronardi et al., 1993), weshowed that PAD2 was elevated before deimination ofarginyl residues and before any clinical or pathologicsigns of demyelinating disease (Moscarello et al., 2002).
Fig. 4. DNA demethylase in normal and MS white matter. A: Thinlayer chromatogram showing the mobility of 5-methyl-cytidylic acidand cytidylic acid. B: Ratio of cytidylic acid to 5-methyl cytidylicacid (Normal, n ¼ 4; MS, n ¼ 12) 6 SD in white matter. C: Ratioof cytidylic acid to 5-methyl cytidylic acid (Alzheimer’s [Alz], n ¼ 2;Huntington’s disease [Hnt], n ¼ 2; and Parkinson’s disease [Park],n ¼ 2) 6 SD. Values for the ratio of C/mC 6 SD for each individ-ual assayed are shown in Supplementary Table I.
DNA Hypomethylation in MS White Matter 2013
Journal of Neuroscience Research DOI 10.1002/jnr

More importantly, the amount of PAD2 was elevated at2 months of age and continued to increase during clini-cal disease whereas MBPcit was elevated at 3 months ofage (Mastronardi et al., 1996; Moscarello et al., 2002).Our studies suggest strongly that increased levels ofPAD2 that generates citrullinated MBP plays an impor-tant role in demyelinating disease (Mastronardi et al.,1996; Moscarello et al., 1994, 2002; Mastronardi andMoscarello, 2005).
Methylation changes in DNA have been docu-mented in a number of other processes as well (Gloriaet al., 1996; Liu et al., 2004; Shuto et al., 2006).Chronic inflammation has been correlated with DNAmethylation levels in colorectal tumors (Gloria et al.,1996). In these tumors it was not clear whether thehypomethylated DNA was from tumor cells or wasderived from inflammatory infiltrating cells. In our whitematter samples, infiltrating cells or activated microglia/macrophages in the MS individuals was not different sig-nificantly from the normal individuals. GFAP, an astro-cyte marker, was elevated in the MS NAWM. Thisincrease was associated with an increase of the pro-inflammatory cytokine TNF-a (Mastronardi et al.,2006). TNF-a and possibly other inflammatory mole-cules yet to be determined in MS NAWM may alsocontribute to DNA hypomethylation (Pang et al., 1992;Houde et al., 2004; Liu et al., 2004). The promoter forthe pro inflammatory cytokine interferon-gamma (IFN-
g) contains CpG sequences (Pang et al., 1992; Melvinet al., 1995) methylation that downregulates IFN-g tran-scription (White et al., 2002; Yano et al., 2003). Infec-tion with human immunodeficiency virus Type I stimu-lates DNA methyltransferase that methylates the pro-moter in the presence of S-adenosyl methionine(Mikovits et al., 1998) thereby silencing the gene. Inanother study, the promoter region of the IFN-g genein pro-inflammatory Th1 activated T-cells was hypome-thylated, whereas in Th2 cells the IFN-g promoter wasmethylated. Methylation of the IFN-g promoter in T-cells classified as Th1-positive showed reduced bindingof transcription factors so that IFN-g production wasdownregulated, thereby converting Th1 þ cells intoTh2 cells (Yano et al., 2003). Treatment of activated T-cells with vitamin B12, the universal methyl donor,resulted in a reduction of IFN-g protein and an increasein IFN-b (Mastronardi et al., 2007). IFN-b has beenassociated with methylation (Mowen et al., 2001) andthis function was enhanced with addition of B12 (Mas-tronardi et al., 2004). It is not known, however, if theeffect is at the DNA level. Because IFN-g is a pro-inflammatory cytokine, downregulation of the IFN-gpromoter should benefit disease.
Methylation of genes may cause a localized chro-matin rearrangement that makes the genes inaccessibleand transcriptionally inactive. In the inactive state, chro-matin is bound by the 5-methylcytosine binding protein
Fig. 5. Model of myelin destabilization by increased PAD2. In nor-mal white matter the PAD2 CpG island is methylated (shaded circles)resulting in reduced expression of PAD2. In MS white matter thePAD2 CpG island is hypomethylated (open circles), possibly due toincreased DNA demethylase activity. The non-methylated PAD2promoter is available for increased transcriptional activation in MSwhite matter. The increased hypomethylated PAD2 CpG island leads
to an increase of PAD2 enzyme. Citrullination of MBP (MBPcit6–18)(amount of citrullination depends on disease severity) results in a lossof myelin compaction (rectangle). If MBP citrullination is extreme,then myelin becomes disrupted and breaks down eliciting release ofprotein fragments and the stimulation of an immune response thatcontributes to lesion development and MS.
2014 Mastronardi et al.
Journal of Neuroscience Research DOI 10.1002/jnr

2 (Nan et al., 1997) in combination with Sin 3 (Joneset al., 1998) and histone deacetylase (Jones et al., 1998;Wade et al., 1998). Thus the reduction of DNA methyl-ation may act by preventing the binding of methyl cyto-sine binding proteins. One of these is hypothesized to behistone deacetylase, which when reduced leads to tran-scriptional activation (Bartoli et al., 2003).
This study examines the mechanism by whichPAD2 was upregulated and shows that the methylationstate of the PAD2 promoter from MS white matter wasone-third of normal. The hypomethylation in MSNAWM may be a more general epigenetic phenomenonwhere it may have some biological relevance for geneexpression. These differences may be an indication of amore generalized aberrant DNA methylation. Themodel depicted in Figure 5 summarizes the outcome ofdemethylation of the PAD2 promoter. The moleculespreceding the demethylation of the promoter or inhibi-tion of methylation are not known currently. These mayinclude a host of processes involved in DNA methyla-tion. In the demethylated state, however, the PAD2promoter activity would allow for increased transcriptionof PAD2 resulting in increased amount of enzyme. Thiswould lead to increased citrullination of MBP resultingin the breakdown of myelin with release of myelin pro-tein fragments and subsequent immune response (Mas-tronardi and Moscarello, 2005).
ACKNOWLEDGMENTS
This research was funded by a grant from the MSSociety of Canada to M.A.M. and F.G.M. The authorswould like to acknowledge the sequencing services pro-vided by The Centre for Applied Genomics (www.tcag.ca) at the Hospital for Sick Children in Toronto.We thank Dr. M. Szyf for guidance with the DNAdemethylase assay development and Dr. T.H. Takahara(Japan) for the anti-PAD polyclonal antibody. The tech-nical support from Mrs. T. Miani is greatly appreciated.We are also indebted to Drs. R.M. Nagra and W.W.Tourtellotte for providing the brain and thymus samplesfrom the Human Brain and Spinal Fluid Resource Cen-ter, VA West Los Angeles Healthcare Center (LosAngeles, CA) that is sponsored by NINDS/NIMH,National Multiple Sclerosis Society, and the Departmentof Veterans Affairs.
REFERENCES
Barnett MH, Prineas JW. 2004. Relapsing and remitting multiple sclero-
sis: pathology of the newly forming lesion. Ann Neurol 55:458–468.
Bartoli A, Fettucciari K, Fetriconi I, Rosati E, Ianni MD, Tabilio A,
Delfino DV, Rossi R, Marconi P. 2003. Effect of trichostatin a and 50-azacytidine on transgene reactivation in U937 transduced cells. Pharma-
col Res 48:111–118.
Boggs JM, Rangaraj G, Koshy KM, Ackerley C, Wood DD, Moscarello
MA. 1999. Highly deiminated isoform of myelin basic protein from
multiple sclerosis brain causes fragmentation of lipid vesicles. J Neurosci
Res 57:529–535.
Cedar H. 1988. DNA methylation and gene activity. Cell 53:3–4.
Chataway J, Feakes R, Coraddu F, Gray J, Deans J, Fraser M, Robertson
N, Broadley S, Jones H, Clayton D, Goodfellow P, Sawcer S, Comp-
ston A. 1998. The genetics of multiple sclerosis: principles, background
and updated results of the United Kingdom systematic genome screen.
Brain 121:1869–1887.
Chaudhuri A, Behan PO. 2004. Multiple sclerosis is not an autoimmune
disease. Arch Neurol 61:1610–1612.
Chaudhuri A, Behan PO. 2005. Multiple sclerosis: looking beyond auto-
immunity. J R Soc Med 98:303–306.
Chavanas S, Mechin MC, Takahara H, Kawada A, Nachat R, Serre G,
Simon M. 2004. Comparative analysis of the mouse and human pepti-
dylarginine deiminase gene clusters reveals highly conserved non-coding
segments and a new human gene, PADI6. Gene 330:19–27.
Cheifetz S, Moscarello MA. 1985. Effect of bovine basic protein charge
microheterogeneity on protein-induced aggregation of unilamellar
vesicles containing a mixture of acidic and neutral phospholipids. Bio-
chemistry 24:1909–1914.
Chia LS, Thompson JE, Moscarello MA. 1984. Alteration of lipid-phase
behavior in multiple sclerosis myelin revealed by wide-angle x-ray dif-
fraction. Proc Natl Acad Sci U S A 81:1871–1874.
Clark SJ, Harrison J, Paul CL, Frommer M. 1994. High sensitivity map-
ping of methylated cytosines. Nucleic Acids Res 22:2990–2997.
D’Souza CA, Wood DD, She YM, Moscarello MA. 2005. Autocatalytic
cleavage of myelin basic protein: an alternative to molecular mimicry.
Biochemistry 44:12905–12913.
Dong S, Kojima T, Shiraiwa M, Mechin MC, Chavanas S, Serre G,
Simon M, Kawada A, Takahara H. 2005. Regulation of the expression
of peptidylarginine deiminase type II gene (PADI2) in human keratino-
cytes involves Sp1 and Sp3 transcription factors. J Invest Dermatol
124:1026–1033.
Ebers GC, Kukay K, Bulman DE, Sadovnick AD, Rice G, Anderson C,
Armstrong H, Cousin K, Bell RB, Hader W, Paty DW, Hashimoto S,
Oger J, Duquette P, Warren S, Gray T, O’Connor P, Nath A, Auty
A, Metz L, Francis G, Paulseth JE, Murray TJ, Pryse-Phillips W, Risch
N, et al. 1996. A full genome search in multiple sclerosis. Nat Genet
13:472–476.
Eden S, Cedar H. 1994. Role of DNA methylation in the regulation of
transcription. Curr Opin Genet Dev 4:255–259.
Finch PR, Wood DD, Moscarello MA. 1971. The presence of citrulline
in a myelin protein fraction. FEBS Lett 15:145–148.
Gloria L, Cravo M, Pinto A, de Sousa LS, Chaves P, Leitao CN, Quina
M, Mira FC, Soares J. 1996. DNA hypomethylation and proliferative
activity are increased in the rectal mucosa of patients with long-standing
ulcerative colitis. Cancer 78:2300–2306.
Grigg G, Clark S. 1994. Sequencing 5-methylcytosine residues in
genomic DNA. Bioessays 16:431–436.
Groome N, Dawkes A, Barry R, Hruby S, Alvord E Jr. 1988. New
monoclonal antibodies reactive with defined sequential epitopes in
human myelin basic protein. J Neuroimmunol 19:305–315.
Haines JL, Ter-Minassian M, Bazyk A, Gusella JF, Kim DJ, Terwedow
H, Pericak-Vance MA, Rimmler JB, Haynes CS, Roses AD, Lee A,
Shaner B, Menold M, Seboun E, Fitoussi RP, Gartioux C, Reyes C,
Ribierre F, Gyapay G, Weissenbach J, Hauser SL, Goodkin DE, Lin-
coln R, Usuku K, Oksenberg JR, et al. 1996. A complete genomic
screen for multiple sclerosis underscores a role for the major histocom-
patability complex. The Multiple Sclerosis Genetics Group. Nat Genet
13:469–471.
Houde C, Li Y, Song L, Barton K, Zhang Q, Godwin J, Nand S, Toor
A, Alkan S, Smadja NV, Avet-Loiseau H, Lima CS, Miele L, Coignet
LJ. 2004. Overexpression of the NOTCH ligand JAG2 in malignant
plasma cells from multiple myeloma patients and cell lines. Blood
104:3697–3704.
Hsieh CL. 2000. Dynamics of DNA methylation pattern. Curr Opin
Genet Dev 10:224–228.
DNA Hypomethylation in MS White Matter 2015
Journal of Neuroscience Research DOI 10.1002/jnr

Jones PL, Veenstra GJ, Wade PA, Vermaak D, Kass SU, Landsberger N,
Strouboulis J, Wolffe AP. 1998. Methylated DNA and MeCP2 recruit
histone deacetylase to repress transcription. Nat Genet 19:187–191.
Kim JK, Mastronardi FG, Wood DD, Lubman DM, Zand R, Moscarello
MA. 2003. Multiple sclerosis: an important role for post-translational
modifications of myelin basic protein in pathogenesis. Mol Cell Proteo-
mics 2:453–462.
Kuokkanen S, Gschwend M, Rioux JD, Daly MJ, Terwilliger JD, Tie-
nari PJ, Wikstrom J, Palo J, Stein LD, Hudson TJ, Lander ES, Peltonen
L. 1997. Genome wide scan of multiple sclerosis in Finnish multiplex
families. Am J Hum Genet 61:1379–1387.
Lamensa JW, Moscarello MA. 1993. Deimination of human myelin basic
protein by a peptidylarginine deiminase from bovine brain. J Neuro-
chem 61:987–996.
Liu C, Xu D, Sjoberg J, Forsell P, Bjorkholm M, Claesson HE. 2004.
Transcriptional regulation of 15-lipoxygenase expression by promoter
methylation. Exp Cell Res 297:61–67.
Ludwin SK. 2006. The pathogenesis of multiple sclerosis: relating human pa-
thology to experimental studies. J Neuropathol Exp Neurol 65:305–318.
Mastronardi FG, Ackerley CA, Arsenault L, Roots BI, Moscarello MA.
1993. Demyelination in a transgenic mouse: a model for multiple scle-
rosis. J Neurosci Res 36:315–324.
Mastronardi FG,Mak B, Ackerley CA, Roots BI, MoscarelloMA. 1996. Mod-
ifications of myelin basic protein in DM20 transgenic mice are similar to those
in myelin basic protein from multiple sclerosis. J Clin Invest 97:349–358.
Mastronardi FG, Min W, Wang H, Winer S, Dosch M, Boggs JM,
Moscarello MA. 2004. Attenuation of experimental autoimmune en-
cephalomyelitis and nonimmune demyelination by IFN-beta plus vita-
min B12: treatment to modify notch-1/sonic hedgehog balance.
J Immunol 172:6418–6426.
Mastronardi FG, Moscarello MA. 2005. Molecules affecting myelin sta-
bility: a novel hypothesis regarding the pathogenesis of multiple sclero-
sis. J Neurosci Res 80:301–308.
Mastronardi FG, Wood DD, Mei J, Raijmakers R, Tseveleki V, Dosch
HM, Probert L, Casaccia-Bonnefil P, Moscarello MA. 2006. Increased
citrullination of histone H3 in multiple sclerosis brain and animal mod-
els of demyelination: a role for tumor necrosis factor-induced peptidy-
larginine deiminase 4 translocation. J Neurosci 26:11387–11396.
Mastronardi FG, Tsui H, Winer S, Wood DD, Selvanantham T, Galligan
C, Fish EN, Dosch H-M, Moscarello MA. 2007. Synergy between
paclitaxel plus an exogenous methyl donor in the suppression of murine
demyelinating diseases. Mult Scler (in press).
Melvin AJ, McGurn ME, Bort SJ, Gibson C, Lewis DB. 1995. Hypome-
thylation of the interferon-gamma gene correlates with its expression by
primary T-lineage cells. Eur J Immunol 25:426–430.
Mikovits JA, Young HA, Vertino P, Issa JP, Pitha PM, Turcoski-Cor-
rales S, Taub DD, Petrow CL, Baylin SB, Ruscetti FW. 1998. Infec-
tion with human immunodeficiency virus type 1 upregulates DNA
methyltransferase, resulting in de novo methylation of the gamma inter-
feron (IFN-gamma) promoter and subsequent downregulation of IFN-
gamma production. Mol Cell Biol 18:5166–5177.
Moscarello MA, Pritzker L, Mastronardi FG, Wood DD. 2002. Peptidy-
larginine deiminase: a candidate factor in demyelinating disease. J Neu-
rochem 81:335–343.
Moscarello MA, Wood DD, Ackerley C, Boulias C. 1994. Myelin in mul-
tiple sclerosis is developmentally immature. J Clin Invest 94:146–154.
Mowen KA, Tang J, Zhu W, Schurter BT, Shuai K, Herschman HR,
David M. 2001. Arginine methylation of STAT1 modulates IFNalpha/
beta-induced transcription. Cell 104:731–741.
Nan X, Campoy FJ, Bird A. 1997. MeCP2 is a transcriptional repressor
with abundant binding sites in genomic chromatin. Cell 88:471–481.
Pang Y, Norihisa Y, Benjamin D, Kantor RR, Young HA. 1992. Inter-
feron-gamma gene expression in human B-cell lines: induction by
interleukin-2, protein kinase C activators, and possible effect of hypo-
methylation on gene regulation. Blood 80:724–732.
Pennings S, Allan J, Davey CS. 2005. DNA methylation, nucleosome
formation and positioning. Brief Funct Genomic Proteomic 3:351–361.
Peterson GL. 1977. A simplification of the protein assay method of
Lowry et al., which is more generally applicable. Anal Biochem
83:346–356.
Pritzker LB, Joshi S, Gowan JJ, Harauz G, Moscarello MA. 2000. Deimi-
nation of myelin basic protein. 1. Effect of deimination of arginyl resi-
dues of myelin basic protein on its structure and susceptibility to diges-
tion by cathepsin D. Biochemistry 39:5374–5381.
Raijmakers R, Vogelzangs J, Croxford JL, Wesseling P, van Venrooij
WJ, Pruijn GJ. 2005. Citrullination of central nervous system proteins
during the development of experimental autoimmune encephalomyeli-
tis. J Comp Neurol 486:243–253.
Razin A, Riggs AD. 1980. DNA methylation and gene function. Science
210:604–610.
Sawcer S, Jones HB, Feakes R, Gray J, Smaldon N, Chataway J, Robert-
son N, Clayton D, Goodfellow PN, Compston A. 1996. A genome
screen in multiple sclerosis reveals susceptibility loci on chromosome
6p21 and 17q22. Nat Genet 13:464–468.
Shuto T, Furuta T, Oba M, Xu H, Li JD, Cheung J, Gruenert DC,
Uehara A, Suico MA, Okiyoneda T, Kai H. 2006. Promoter hypome-
thylation of Toll-like receptor-2 gene is associated with increased
proinflammatory response toward bacterial peptidoglycan in cystic fi-
brosis bronchial epithelial cells. FASEB J 20:782–784.
Siegfried Z, Cedar H. 1997. DNA methylation: a molecular lock. Curr
Biol 7:R305–307.
Szyf M. 1994. DNA methylation properties: consequences for pharma-
cology. Trends Pharmacol Sci 15:233–238.
Szyf M, Bhattacharya SK. 2002. Measuring DNA demethylase activity in
vitro. Methods Mol Biol 200:155–161.
Towbin H, Staehelin T, Gordon J. 1979. Electrophoretic transfer of pro-
teins from polyacrylamide gels to nitrocellulose sheets: procedure and
some applications. Proc Natl Acad Sci U S A 76:4350–4354.
Vossenaar ER, Zendman AJ, van Venrooij WJ, Pruijn GJ. 2003. PAD, a
growing family of citrullinating enzymes: genes, features and involve-
ment in disease. Bioessays 25:1106–1118.
Wade PA, Jones PL, Vermaak D, Veenstra GJ, Imhof A, Sera T, Tse C,
Ge H, Shi YB, Hansen JC, Wolffe AP. 1998. Histone deacetylase
directs the dominant silencing of transcription in chromatin: association
with MeCP2 and the Mi-2 chromodomain SWI/SNF ATPase. Cold
Spring Harb Symp Quant Biol 63:435–445.
Watanabe K, Nomoto M, Nagata S, Itoh Y, Hikichi K, Maruyama N,
Mita T, Senshu T. 1992. The rat peptidylarginine deiminase-encoding
gene: structural analysis and the 50-flanking sequence. Gene 114:261–
265.
White GP, Watt PM, Holt BJ, Holt PG. 2002. Differential patterns of
methylation of the IFN-gamma promoter at CpG and non-CpG sites
underlie differences in IFN-gamma gene expression between human
neonatal and adult CD45RO- T cells. J Immunol 168:2820–2827.
Wood DD, Bilbao JM, O’Connors P, Moscarello MA. 1996. Acute mul-
tiple sclerosis (Marburg type) is associated with developmentally imma-
ture myelin basic protein. Ann Neurol 40:18–24.
Wood DD, Moscarello MA. 1989. The isolation, characterization, and
lipid-aggregating properties of a citrulline containing myelin basic pro-
tein. J Biol Chem 264:5121–5127.
Yano S, Ghosh P, Kusaba H, Buchholz M, Longo DL. 2003. Effect of
promoter methylation on the regulation of IFN-gamma gene during in
vitro differentiation of human peripheral blood T cells into a Th2 pop-
ulation. J Immunol 171:2510–2516.
Yeivin A, Razin A. 1993. Gene methylation patterns and expression.
EXS 64:523–568.
2016 Mastronardi et al.
Journal of Neuroscience Research DOI 10.1002/jnr