pattern of diversity for morphological and alkaloid yield related traits among the periwinkle...
TRANSCRIPT

Genetic Resources and Crop Evolution 48: 273–286, 2001.© 2001 Kluwer Academic Publishers. Printed in the Netherlands.
273
Pattern of diversity for morphological and alkaloid yield related traitsamong the periwinkle Catharanthus roseus accessions collected from inand around Indian subcontinent
Parul Mishra, G.C. Uniyal, S. Sharma and Sushil Kumar∗Central Institute of Medicinal and Aromatic Plants (CIMAP), Lucknow 226 015, India; ∗Author for correspondence(fax: 555-342666)
Received 11 May 2000; accepted in revised form 6 October 2000
Key words: Catharanthus roseus descriptors, Hierarchical relationships, Madagascar periwinkle accessions,Multivariate analyses, Terpenoid indole alkaloids
Abstract
Thirty two accessions of periwinkle Catharanthus roseus collected from different semitemperate to tropicalgeographical areas of Indian subcontinent, Madagascar, Singapore and Malaysia were characterized under fieldconditions for 53 growth, development, morphogenesis and alkaloid yield related characters over a few seasons atLucknow, India. Large differences were observed among the accessions for each of the characters examined. Thedifferences among the accessions ranged 3, 80 and 15 fold for the alkaloid yield related traits – leaf dry matter yieldand leaf vincristine and vinblastine concentration, respectively. Strong correlations were observed between leaf areaand leaf yield with leaf alkaloid, root and root alkaloid yields, contents of dimeric alkaloid in leaves with yields ofrespective alkaloids, leaf number and root alkaloid content with leaf vinblastine yield and leaf vinblastine contentwith leaf vincristine and vinblastine yields. Multi-variate analyses allowed classification of the accessions into 5to 7 morphologically and presumably genetically distinct groups. Generally, the accessions coming from tropicalagro-climates tended to get separated from those having origins in subtropical to semi-temperate environments.Three clusters were observed to bear complementary characters for possibly cross-parenting high alkaloid yieldingtransgressive segregants. One of the accessions of C. roseus in which the leaf vincristine content was 5 fold andvinblastine content one and a half fold of the respective all accession means appeared suitable for the domesticatedcultivation. It was proposed that accumulation of high amounts of total alkaloids and one or more dimeric alkaloidsin leaves of certain accessions may be a reflection of reproductive fitness achieved under the biotic and abioticstresses imposed on their parent populations, sometime in their evolutionary pathway.
Introduction
The Madagascar periwinkle, Catharanthus roseus (L.)G. Don of the family Apocynaceae has considerableeconomic importance and is becoming a popular sys-tem for the molecular dissection of its secondary meta-bolism. Periwinkle has become naturalised in severalcountries and is widely cultivated in homes, gardensand parks on account of its propensity to bear flowersover most parts of the year in the areas bearing tropicaland sub-tropical agroclimates. The plant accumulatesmore than 125 mono-terpenoid indole alkaloids indifferent organs (Gröger 1985). The roots accumu-
late ajmalicine and serpentine, which are importantcomponents of medicines for controlling high bloodpressure and other types of cardio-vascular maladies.The leaves and stem are the sources of the dimeric al-kaloids vinblastine (VLB) and vincristine (VCR), thatare indispensible parts of most anti-cancer chemother-apies (Son et al. 1990, Jordan et al. 1991, Molner etal. 1995). In C. roseus plants, VLB and VCR occur invery low concentrations of about 1 g and 20 mg, re-spectively, in 1000 kg of leaf and stem material (Tyler1988). The complexity of isolation from plant materialis a major reason for the high cost of these dimericalkaloids. Much of the commercial VLB and VCR are

274
synthesized by oxidative coupling of their monomericprecursor alkaloids vindoline (V) and catharanthine(C) which are either isolated from adult plants or invitro cultured cells, tissues and organs of C. roseus.While V is extracted from leaves, C is resourced fromleaves, roots and in vitro cultures of C. roseus. Itis estimated that 40 to 100 fold increase in alkaloidcontent will make production of VLB and VCR fromC. roseus plant material competitive with the presentsemi-synthetic process (Moreno et al. 1995; Kulkarniet al. 1999).
Recent work on the effect of varying environmentson the growth and expression of secondary meta-bolism in periwinkle has revealed that biotic factorssuch as plant organ age, and their pathogenesis byfungi (Frischknecht et al. 1987; Aerts et al. 1994),and abiotic factors including quality and quantity oflight (Vazquez-Flota and De Luca 1998) temperature,humidity (Talha et al. 1975; Godoy-Harnandez andLoyala-Vargas 1981; Virk and Singh 1990; Saenz etal. 1993), and soil quality (Vazquez-Flota et al. 1994)determined the yield of different alkaloids from vari-ous organs of C. roseus. In this regard, jasmonates,which are known to induce systemic resistance to vir-uses, bacteria and fungi in plants, have been foundto induce synthesis of key enzymes of the terpenoidbiosynthesis in C. roseus resulting in the overproduc-tion of several alkaloids (Menke et al. 1999). Specificcell wall components of yeast and other fungi havealso been observed to elicit transcriptional inductionof alkaloids, like the jasmonates. Wounding stress in-creased indole alkaloids in C. roseus (Frischknecht etal. 1987). Anti-feedant properties have been found tobe associated with the accumulation of strictosidineand V (Luijendijk et al. 1996). The reproduction inBemesia tabaci was found to be arrested on transgenictobacco plants that accumulated C. roseus specific al-kaloid tryptamine (Thomas et al. 1995). These obser-vations indicate that higher levels of indole alkaloids,intermediates and related enzymes somehow improveadaptation of C. roseus plant and provide them protec-tion against stressful conditions caused by a range ofbiotic and abiotic stresses.
The naturalized population of C. roseus occurringin wild in distant geographical locations of tropicaland sub-tropical agro-climates must represent surviv-ors of a variety of location-specific biotic and abioticchallenges over many generations. Here, we examinedthe idea that C. roseus accessions collected from dif-ferent locations may carry variation in the relativeexpression of one or more steps of indole alkaloid
biosynthetic pathway, besides that in growth, devel-opment and morphogenesis dependent morphologicalcharacters. Some of the accessions may accumulatehigh levels of important alkaloid in plant tissues thanothers.
Materials and methods
Periwinkle variants and field experiment design
The origins and characteristics of the 32 germplasmlines of Catharanthus roseus obtained from variouslocations in India and abroad are given in Table 1.To study the relative expression of characters in dif-ferent accessions the field experiments were laid outat the Institute’s field at Lucknow, India, located at26.5◦ N, 80.5◦ E and 120 m above the sea level. Thesoil was sandy-loam in texture with pH about 8.1, lowavailable N (160 kg ha−1), K (105 kg ha−1) and me-dium P (124 kg ha−1). The accessions were croppedtwice over July 1996 to April 1997 and July 1997 toApril 1998 seasons during 1996 monsoon to 1997–98 summer period (monsoon, July to October; winter,November to February; and summer March to June).
Seeds of each accession were sown in shallowearthen pots containing a (1:1) mixture of soil andfarm yard manure (FYM). In the second week of Julyand seedlings of about six weeks of age were trans-planted in the field. The experiment was laid out in thedesign of randomized blocks, replicated three times.The plots of size 2.7 × 1.8 m2 were transplanted withthirty six seedlings in rows 45 cm apart, such thatseedlings were spaced 30 cm in each line. The fieldwas applied N, P and K in the proportion of 80:40:40;the sources of N, P and K were CO(NH2), (NH4)2 H2PO4 and KCl. One third of N and all of the P and Kfertilizers were given as a basal dose at the time ofthe preparation of the field in August. The remain-ing N was applied in equal halves in December andnext April to all the plots at the same rate. The fieldwas irrigated, as and when required. The outbreak ofcollar-rot disease was controlled by a single spray of0.1% bavistin in February.
Method of morphology and alkaloid relatedobservations
The crop was harvested in next late April by dig-ging up all the plants, plot-wise. Qualitative data onshapes, hairiness, extent and distribution of anthocy-anin pigmentation were recorded on leaves, flowers

275
Table 1. Morphological descriptors of 32 accessions of Catharanthus roseus.
Sl. Accession Origin Morphological characteristic(s) a,b,c
no. designation
1. CrC-17 Chandigarh Habit ssp; height sho; branches sma, apa; stem nor; root, thn; petiole
pye, tpr, lon; leaf size, nor, number sma, g(139A), obl, apex acu, base
att, tds; venation idi; flower, sma, sepal g(138C), tpr, nor, petal
rp(74B), orifice y(3B), anther lar, style lon; pedicel pye, tpr; follicle
pyg, tpr, div, nor; fertility hig; seed lar
2. CrC-25 Hissar Habit ere; height nor; branches nor, apa; stem thn; root, thn; petiole
pye, tab, sho; leaf size, lar, number nor, g(137A), ell, apex blu, base
blu, tab; venation dis; flower, nor, sepal g(139D), tab, nor, petal
whi (74B), orifice y(2B), anther lar, style nor; pedicel pyg, tab; follicle
pyg, tab, div, nor; fertility nor; seed nor
3. CrC-28 Hissar Habit ere; height tal; branches low, apa; stem thk; root, thk; petiole
pye, tab, sho; leaf size, nor, number nor, yg(147B), obo, apex att, base
blu, tab; venation dis; flower, sho, sepal g(139A), tab, sho, petal whi
(74B), orifice rp(60A), anther nor, style lon; pedicel pye, tab; follicle
pyg, tab, sho, div; fertility hig; seed nor
4. CrC-4 Jodhpur Habit ere; height tal; branches nor, apn/ste; stem thn; root, thk; petiole
pyg, tab, sho; leaf size, nor, number sma, g(139A), ell, apex att, base
obt, tab; venation idi; flower, lar, sepal g(138C), tab, sho, petal pv
(80C), orifice vb(89B), anther nor, style lon; pedicel pyg, tab; follicle
pyg, tab, div, nor; fertility hig; seed lar
5. CrC-2 Ajmer Habit ere; height sho; branches low, apa; stem thn;root, thn; petiole
pye, tab, lon; leaf size, lar, number sma, yg(139D), obo, apex acu,
base att, tab; venation idi; flower, nor, sepal g(138C), tab, nor, petal
whi, orifice gy(1B), anther nor, style lon; pedicel pye, tab; follicle
pyg, tab, div, sho; fertility hig; seed nor
6. CrC-2 Jhansi Habit ere; height nor; branches low, apl/ste; stem nor; root, nor;
petiole pye, tpr, nor; leaf size, nor, number sma, g(137B), obl, apex
acu, base obt, tds; venation dis; flower, nor, sepal g(139B), tpr, nor,
petal rp(64C), orifice v(88A), anther lon, style lon; pedicel pye, tpr;
follicle pye, tpr, par, nor; fertility hig; seed sma
7. CrC-16 Meerut Habit ssp; height sho; branches sma, apa; stem thn; root, thn; petiole
pye, tpr, lon; leaf size, sma, number sma, g(143B), obl, apex acu, base
obt, tdh; venation dis; flower, sma, sepal g(138C), tpr, nor, petal
rp(74C), orifice p(78A), anther lar, style nor; pedicel pye, tpr; follicle
pye, tpr, div, nor; fertility hig; seed nor
8. CrC-19 Pantnagar Habit ssp; height tal; branches nor, aph/ste; stem thk; root, nor;
petiole pgr, tpr, sho; leaf size, nor, number sma, yg(146C), obo, apex
blu, base obt, tab; venation dis; flower, nor, sepal g(138A), tab, nor,
petal pv(80A), orifice v(86B), anther sma, style nor; pedicel pgr, tpr; follicle
pye, tab, nor, div; fertility low; seed sma

276
Table 1. Continued.
Sl. Accession Origin Morphological characteristic(s) a,b,c
no. designation
9. CrC-3 Nainital Habit ssp; height sho; branches sma, apa; stem nor; root, thn; petiole
pye, tpr, lon; leaf size, nor, number sma, g(139A), obl, apex acu, base
att, tds; venation idi; flower, sma, sepal g(138C), tpr, nor, petal
rp(74B), orifice y(3B), anther lar, style lon; pedicel pye, tpr; follicle
pyg, tpr, div, nor; fertility hig; seed lar
10. CrC-8 Delhi Habit spr; height nor; branches low, apd/ste, pet, ctb, pod; stem nor;
root, thn; petiole pye, tpr, nor; leaf size, nor, number sma, yg(146A),
obo, apex att, base obt, tdh; venation dis; flower, lar, sepal yg(146A),
tpr, nor, petal v(86A), orifice vb(93B), anther sma, style lon; pedicel
pye, tpr; follicle pyg, tpr, div, nor; fertility low; seed lar
11. CrC-13 Delhi Habit ere; height nor; branches nor, apl/ste; stem nor; root, nor;
petiole pye, tpr, nor,lon; leaf size, nor, number nor,yg(147B), obl,
apex acu, base obt, tdn; venation idi; flower, sma, sepal g(138A), tpr,
lon, petal pv(81B), orifice rp(60A), anther nor, style nor; pedicel pye,
tpr; follicle pgr, tpr, div, lon; fertility hig; seed sma
12. CrC-31 Cochin Habit ssp; height nor; branches hig, aph/ste,pet,pod; stem thn; root,
thn; petiole pyg, tpr, lon; leaf size, lar, number hig, g(137A), obo,
apex blu, base obt, tdh; venation dis; flower, lar, sepal yg(146C), tpr,
lon, petal rp(67A), orifice vb(88A), anther lar, style nor; pedicel pyg,
tpr; follicle pye, tpr, par, lon; fertility low; seed lar
13. CrC-30 Puri Habit ere; height tal; branches hig, aph/ste, pet, ctb, pod; stem thn;
root, thk; petiole pyg, tpr, lon; leaf size, lar, number hig, g(137A), obl,
apex acu, base att, tdn; venation dis; flower, lar, sepal yg(148B), tpr,
lon, petal pv(80A), orifice v(83B), anther lar, style nor; pedicel pyg,
tpr; follicle pye, tpr, div, nor; fertility nor; seed lar
14. CrC-5 Darjeeling Habit ere; height nor; branches sma, apa; stem thk; root, nor; petiole
pyg, tpr, sma; leaf size, nor, number sma, yg(146C), lan, apex acu,
base obt, tdh; venation idi; flower, lar, sepal yg(148C), tpr, lon, petal
whi, orifice gy(1B), anther nor, style lon; pedicel pyg, tpr; follicle pye,
tpr, div, nor; fertility nor; seed lar
15. CrC-6 Darjeeling Habit spr; height nor; branches lar, apn/ste, pet, pod; stem nor; root,
nor; petiole pye, tpr, nor; leaf size, lar, number lar, g(137B), obo,
apex blu, base obt, tdh; venation dis; flower, lar, sepal yg(146A), tpr,
nor, petal v(87B), orifice vb(89A), anther nor, style lon; pedicel pye,
tpr; follicle pyg, tpr, par, nor; fertility nor; seed lar
16. CrC-23 Kodaikanal Habit ere; height nor; branches nor, apa; stem nor; root, thn; petiole
pye, tab, sho; leaf size, nor, number nor, g(137B), obl, apex acu, base
blu, tdn; venation dis; flower, sma, sepal yg(148C), tpr, lon, petal whi,
orifice y(3B), anther nor, style nor; pedicel pye, tpr; follicle pye, tpr,
nor, par; fertility nor; seed nor

277
Table 1. Continued.
Sl. Accession Origin Morphological characteristic(s) a,b,c
no. designation
17. CrC-22 Mysore Habit ere; height nor; branches nor, apa; stem thk; root, thn; petiole
pye, tpr, nor; leaf size, nor, number nor, yg(146A), obl, apex acu, base
blu, tds; venation dis; flower, sma, sepal yg(146A), tpr, nor, petal whi,
orifice y(4B), anther nor, style nor; pedicel pye, tpr; follicle pyg, tpr,
nor, div; fertility hig; seed nor
18. CrC-29 Calcutta Habit ere; height tal; branches nor, apa; stem thk; root, thn; petiole
pye, tab, sho; leaf size, nor, number nor, yg(147B), obo, apex blu,
base blu, tab; venation dis; flower, lar, sepal yg(148A), tab, sho, petal
rp(66B), orifice pv(82A), anther nor, style lon; pedicel pyg, tab;
follicle pyg, tab, div, nor; fertility low; seed sma
19. CrC-20 Coimbatore Habit ssp; height tal; branches hig, apn/ste; stem thk; root, nor; petiole
pgr, tab, sho; leaf size, lar, number lar, yg(144B), obo, apex blu, base
att, tab; venation dis; flower, nor, sepal g(138A), tab, nor, petal
pv(80A), orifice v(86B), anther sma, style nor; pedicel pgr, tab;
follicle pgr, tab, div, nor; fertility low; seed nor
20. CrC-24 Ootacamund Habit ere; height nor; branches nor, apa; stem thn; root, thk; petiole
pye, tab, sho; leaf size, sma, number nor, g(137A), lan, apex acu, base
blu, tab; venation dis; flower, nor, sepal g(138B), tab, lon, petal whi,
orifice y(4B), anther nor, style nor; pedicel pye, tpr; follicle pye, lon,
div, tab; fertility nor; seed nor
21. CrC-15 Hyderabad Habit ere; height sho; branches sma, apa; stem thn; root, nor; petiole
pye, tpr, sma; leaf size, sma, number sma, g(139A), lan, apex acu,
base att, tdn; venation dis; flower, nor, sepal yg(146A), tpr, lon, petal
whi, orifice y(3B), anther lar, style nor; pedicel pye, tpr; follicle pye,
tpr, par, nor; fertility hig; seed sma
22. CrC-18 Tiruchirapalli Habit ere; height nor; branches nor, apn/ste,pet, pod; stem nor; root,
thn; petiole pyg, tpr, sma; leaf size, sma, number sma, g(138A), obl,
apex acu, base obt, tdh; venation idi; flower, nor, sepal g(138B), tpr,
nor, petal rp(74B), orifice v(83C), anther nor, style lon; pedicel pyg,
tpr; follicle pyg, tpr, div, nor; fertility nor; seed sma
23. CrC-7 Secunderabad Habit ssp; height nor; branches nor, apn/ste; stem thn; root, thn;
petiole pye, tpr, nor; leaf size, nor, number sma, yg(147B), obl, apex
blu, base obt, tdn; venation dis; flower, lar, sepal yg(146B), tpr, lon,
petal v(86C), orifice vb(89A), anther lar, style nor; pedicel pye, tpr; follicle pyg,
tpr, div, nor; fertility low; seed lar
24. CrC-14 Andamaans Habit spr; height sho; branches sma/aph/ste,pet,ctb; stem thn; root,
thn; petiole pyg, tpr, sho; leaf size, lar, number sma, yg(147B), obl,
apex acu, base obt, tdn; venation idi; flower, sma, sepal g(138B), tpr,
lon, petal pv(80C), orifice rp(60B), anther lar, style lon; pedicel pyg,
tpr; follicle pgr, tpr, sho, par; fertility nor; seed sma

278
Table 1. Continued.
Sl. Accession Origin Morphological characteristic(s) a,b,c
no. designation
25. CrC-10 Andamaans Habit ssp; height sho; branches hig, apl/ste,pet; stem thk; root, nor;
petiole pyg, tpr, sma; leaf size, lar, number hig, g(138A), obl, apex
obt, base acu, tdh; venation dis; flower, sma, sepal g(139D), tpr, nor,
petal pv(81A), orifice v(88B), anther nor, style sho; pedicel pyg, tpr;
follicle pgr, tpr par, nor; fertility hig; seed sma
26. CrC-9 cv. Nirmal Habit ssp; height nor; branches nor, apa; stem nor; root, thn; petiole
pyg, tpr, nor; leaf size, nor, number nor, g(138A), obl, apex acu, base
blu, tdn; venation dis; flower, nor, sepal g(139A), tpr, sma, petal whi,
orifice gy(1B), anther nor, style nor; pedicel pye, tpr; follicle pye, tpr,
div, nor; fertility hig; seed sma
27. CrC-11 cv. Dhawal Habit ere; height nor; branches sma, apa; stem thn; root, nor; petiole
pyg, tpr, sma; leaf size, nor, number sma, yg(144A), obl, apex acu,
base obt, tdh; venation dis; flower, sma, sepal yg(148B), tpr, nor, petal
whi, orifice y(2B), anther lar, style nor; pedicel pye, tpr; follicle pgr,
tpr, sma, div; fertility hig; seed sma
28. CrC-12 mutant of Habit ere; height nor; branches hig, apl/ste; stem thn; root, nor;
Nirmal petiole pye, tpr, nor; leaf size, lar, number nor, g(143B), lan, apex att,
base obt, tab; venation idi; flower, sma, sepal yg(146B), tab, lon, petal
whi, orifice y(2C), anther nor, style nor; pedicel pye, tab; follicle pyg,
tab, sma, div; fertility hig; seed sma
29. CrC-32 Sri Lanka Habit ere; height tal; branches hig, apl/ste; stem thn;
root, thn; petiole pye, tab, lon; leaf size, lar, number nor, g(43A), ell; apex blu,
base blu, tab; venation idi; flower, lar, sepal yg(146B), tab, lon, petal
pv (80C), orifice v(88B), anther nor, style lon; pedicel pye, tab;
follicle pye, tab, div, nor; fertility nor; seed lar
30. CrC-27 Madagascar Habit ere; height sho; branches sma, apa; stem thk; root, nor; petiole
pye, tab, lon; leaf size, nor, number sma, yg(146A), lan, apex acu,
base blu, tab; venation idi; flower, nor, sepal g(139B), tab, sho, petal
p(78B), orifice pv(81A), anther nor, style lon; pedicel pye, tab;
follicle pye, tab, nor, par; fertility hig; seed sma
31. CrC-21 Singapore Habit ssp; height tal; branches lar, aph/ste,pet,ctb,pod; stem thk;
root, nor; petiole pyg, tab, sho; leaf size, lar, number lar, g(138A),
obl, apex acu, base att, tdh; venation idi; flower, nor, sepal yg(148A),
tpr, nor, petal rp(74B), orifice pv(81A), anther sma, style nor;
pedicel pyg, tpr; follicle pgr, tpr, sma, div; fertility hig; seed lar
and follicles. Colour charts from Royal HorticulturalSociety (RHS) were used for classifying the colours.For quantitative characters, five randomly selectedplants per plot were individually measured for plantheight, number of leaves, leaf area and weight of fresh
roots and leaves (Table 3). The leaves and roots werethen dried at 30 ◦C for determining their dry weightand alkaloid contents as given below: To determinealkaloid content, 5 g of finely powdered dry leavesfrom each sample were extracted with 90% ethanol,

279
Table 1. Continued.
Sl. Accession Origin Morphological characteristic(s) a,b,c
no. designation
32. CrC-26 Malaysia Habit ere; height tal; branches lar, apn/ste,pet; stem thn; root, nor;
petiole pyg, nor, tpr; leaf size, nor, number lar, g(137B), obl, apex
acu, base att, tdn; venation idi; flower, nor, sepal yg(146B), tpr, nor,
petal pv(80C), orifice rp(59B), anther nor, style lon; pedicel pyg, tpr;
follicle pye, tpr, sho, div; fertility hig; seed lar
a Usually the quantitative measurements for a character were distributed into 2-5 classes depending uponthe range of differences and called as poor/scanty/thin/low or medium/normal or high/dense/long/thick orvery high.b ere = erect, ssp = semi-spreading, spr = spreading, thn = thin, thk = thick, pye = pale yellow, pgr = palegreen, pyg = pale yellow green, ste = stem, pet = petiole, ctb = corolla tube, pod = follicle, sho = short,lon = long, sma = small, lar = large, nor = normal, tab and tpr = trichome absent or present respectively,tdh, tdn, tds=trichome density high, normal, scanty respectively, hig = high, low = low ell= elliptic, lan =lanceolate, obl = oblong, obo = obovate, att = attenuate, blu = blunt, acu = acute, obt = obtuse, dis= distinct,idi = indistinct, whi= white, par = parallel.c Within the parentheses are the colour codes prescribed by the Royal Horticultural Society: pv = purpleviolet group, v = violet group, yg = yellow green group, g = green group , rp = red purple group.
three times at 2 h interval at room temperature. Theresulting 90 ml suspension was filtered and concen-trated at 35 ◦C in vacuum to 10 ml, diluted with 10 mldistilled water, and acidified with 10 ml of 3% HCl.The solution was thrice extracted with 30 ml ethylacetate at 2 hourly interval. While the ethyl acetateextract was rejected, the aqueous portion was cooled to10 ◦C, alkalinised with ammonia solution and extrac-ted thrice with chloroform (3 × 30 ml). The resultingchloroform extract was washed with 30 ml of dis-tilled water, dried over anhydrous sodium sulphate,filtered and concentrated to dryness at 30 ◦C undervacuum. The obtained brown residue was dried to aconstant weight. The crude extract was analyzed byhigh performance liquid chromatography (HPLC). ForHPLC resolution and quantification, WATERS mod-ular system consisting of two 501 pumps, automatedgradient controller, U6K injector, in-line degasser, 996photodiode array detector and millenium 2010 chro-matography manager was used. The injector, gradientcontroller and 2010 manager were integrated to givereproducible results. A symmetry column of 4 µm(3.9 × 150 mm WATERS, USA) was used. A lineargradient of solvent was used consisting of methanol,acetonitrile 0.025 m ammonium acetate and triethyl-amine in 2 ratios, (A) 13:32:15:2; and (B) 19:46:35:2(Naaranlahti et al. 1987). Ten µl of the extract wasinjected into the pump using a Hamilton syringe.A flow rate of 0.4 ml changing to 1 after 20 minwas maintained, using a UV-detector. The quantit-ative determination of unknown sample of dimeric
indole alkaloids was performed with calibrated ex-ternal standardization. Under above conditions distinctpeaks were obtained at retention times 14.1 and 18.8for VCR and VLB, respectively.
Statistical data analysis
The data was analysed using ANOVA on 20 quant-itative characters (Cochran and Cox 1957). Inter-character correlation coefficients for yield contributingcharacters were calculated. The thirty two accessionswere clustered into similarity groups using principalcomponent analysis (Mahalanobis 1936). Two prin-cipal components were extracted and their relativescores determined and plotted for cluster analysis.Cannonical variable analysis was used to confirm thegroupings, so obtained (Rao 1952). A dendrogramwas constructed using the D2 values obtained in theprincipal component analysis by dividing each valueby the maximum value obtained to attain a scale of0-1. For this purpose a software package, DSQR,developed at this Institute was used (Sharma et al.1996). Metroglyph sketches (Ramanujam et al. 1964)were superimposed on each accession constituting thecluster.
Results
Variation among accessions collected from the wild
A group of 32 accessions of periwinkle C. roseus

280
Table 2. Methods of observation and parameters of variation estimated for the 20 traits investigated in a collectionof 32 accessions of C. roseus.
Sl. Trait Code Mean Range Description of the trait
no.
1. Plant height (cm) PHT 74 49–100 distance from apex of the plant to
the base
2. Leaf number LNB 591 309–1034 total number of leaves borne on the
plant
3. Leaf area (cm2) LAR 8.30 8.98–12.96 measured directly using a Li-cor
(2000) leaf area meter
4. Root dry yield (kg ha−1) RDY 506 236–1254 total weight of root dry matter per
hectare
5. Leaf dry yield (kg ha−1) LDY 1916 1016–3123 total weight of leaf dry matter per
hectare
6. Root alkaloid RAC 1.12 0.13–2.18 percent concentration of total root
concentration (%) alkaloids on dry weight basis
7. Root alkaloid yield (kg ha−1) RAY 5.59 0.44–13.30 total yield of root alkaloids
per hectare
8. Leaf alkaloid concentration (%) LAC 1.00 0.67–1.29 percent concentration of total leaf
alkaloids on dry weight basis
9. Leaf alkaloid yield (kg ha−1) LAY 18.6 9.0–35.5 total yield of leaf alkaloids per
hectare
10. Vincristine concentration VRC 1.70 0.11–8.45 percent concentration of vincristine
(% X 10−3) on dry weight basis
11. Vinblastine concentration VBC 10.9 1.5–22.7 percent concentration of
(% X 10−3) vinblastine on dry weight basis
12. Vincristine yield VRY 4.71 0.20–62.50 total yield of vincristine per hectare
(g ha−1)
13. Vinblastine yield VBY 22.4 6.9–47.7 total yield of vinblastine per
(g ha−1) hectare
14. Leaf moisture LMC 80.7 72.0–89.0 percent concentration of leaf
concentration (%) moisture
15. Flower diameter (cm) FDI 3.7 2.5–5.0 measured as distance between the
notches of two opposite petals
16. Corolla tube CTL 21.8 12.0–28.0 measured from the base of the
Length (mm) corolla tube to the point of
adhesion of petals
17. Style length (mm) STL 1.90 1.20–2.30 measured from the base to the tip
of the style
18. Pollen fertility (%) POF 81 59–96 acetocarmine staining taken as the
index of fertility
19. Follicle length (mm) FOL 24.5 18.0–33.3 measured from the base to the tip
of the follicle
20. Weight of 100 seeds HSW 112 71–181 weight of hundred dry seeds
having origins in many geographical and climaticallydifferent areas of Indian sub-continent, Madagascar,Singapore and Malaysia were grown under identicalconditions in field and observed for variation in mor-phological, agronomic and biochemical traits. The
description of the accessions in terms of the 53 charac-ters examined, pertaining to the growth, developmentand morphogenesis of different organs and secondaryalkaloid metabolism analysis of the plant is given inTable 1.

281
Table 3. Co-efficients of correlations between pairs of alkaloid yield related characters in 32 accessions of Catharanthus roseus.
Leaf Leaf Root Leaf Leaf Root Leaf Root Vincristine Vinblastine Vincristine Vinblastine
number area yield yield alkaloid alkaloid alkaloid alkaloid concen- concen- yield yield
yield yield concen- concen- tration tration
tration tration
Plant 0.26 0.05 −0.06 0.05 0.04 −0.14 0.04 −0.36∗ −-0.12 −0.07 −0.08 −0.05
height
Leaf 0.56∗∗−0.04 0.38∗ 0.48∗∗ 0.15 0.45∗∗ 0.29 0.25 0.33∗ 0.27 0.58∗∗number
Leaf 0.18 0.31∗ 0.57∗∗ 0.35∗ 0.39∗ 0.37∗ 0.07 0.42∗∗ 0.04 0.58∗∗area
Root 0.66∗∗ 0.56∗∗ 0.74∗∗ −0.05 0.03 −0.11 −0.19 −0.05 0.01
yield
Leaf 0.76∗∗ 0.70∗∗ −0.01 0.24 −0.09 0.04 0.04 0.40∗yield
Leaf 0.67∗∗ 0.49∗∗ 0.37∗ 0.08 0.32∗ 0.08 0.65∗∗alkaloid yield
Root 0.11 0.51∗∗ −0.29 0.16 −0.24 0.30∗alkaloid yield
Leaf 0.33∗ 0.26 0.47∗∗ 0.04 0.41∗alkaloid
concentration
Root −0.25 0.68∗∗ −0.26 0.48∗∗alkaloid
concentration
Vincristine 0.01 0.79∗∗ 0.41∗concentration
Vinblastine −0.14 0.69∗∗concentration
Vincristine 0.64∗∗yield
∗, ∗∗: Significant at 5% and 1% , respectively.
There were large differences among the acces-sions in the pink coloured anthocyanin pigmentationpatterns of different flowering types and other shootorgans; 17 accessions were not pigmented and on theother hand, an accession from Singapore (Crc-21) hadalmost all the shoot organs deeply pigmented. While11 of the accessions did not bear trichomes on theirleaf organs, the trichome intensity varied among otheraccessions.
The characters listed in Table 1 perhaps amplycover the genome in respect of primary metabolismconcerned with for the analysis of variation in them byquantitative uni- and multi-variate analysis proceduresfor defining the genetically distinct groups among C.roseus accessions under examination.
Morphological and agronomic traits
Table 2 presents uni-variate statistics for the 32 acces-sions. The variation for the 6 morphological and 14 ag-ronomic traits showed highly significant and wide dif-ferences between some of the accessions. Remarkabledifferences in the leaf contents of dimeric alkaloids,vincristine and vinblastine, were noteworthy.
Table 3 presents coefficients of correlation betweenthe agronomic traits as a measure of associationbetween them. Some of the strong correlations (r >
0.55) were logical, like that of leaf area and leaf yieldwith leaf alkaloid yield, between root yield and rootalkaloid yield, of leaf vincristine and vinblastine con-centrations with the yields of the respective alkaloids.Such correlations detected between leaf number andleaf area and leaf and root yields appeared to offer

282
promise for simultaneous leaf and root yield improve-ment. The high level correlations between leaf yieldand leaf alkaloid yield on one hand and root alkaloidyield on the other were complementary to those ob-served above. However, some of the observed correla-tions were unexpected, like those between leaf numberand leaf vinblastine yield, root alkaloid concentrationand leaf vinblastine concentration and vincristine andvinblastine yields. These linkages must be indicativeof natural features of plant’s physiology and alkal-oid biosynthetic process. Using these observations, itmay be surmised that synthesis of certain precursorsof dimeric alkaloids may be occurring in roots, al-though the final dimeric products involving them gotsynthesized in the leaves. These correlations appearfavourable for alkaloid yield improvement in C. ros-eus. Certain other correlations were also significant,but weak.
Multi-variate patterns of diversity
Results of the principal component analysis arepresented in Table 4. The percentage of total vari-ation among the accessions accounted for by the firsttwo principal components was 46.3 and 28.7%, re-spectively. In the first principal component (PCI), theloadings of leaf area, leaf alkaloid and vincristineyields and leaf moisture content were high. In thesecond principal component, the leaf area and leafalkaloid yield had significant contribution.
Figure 1 shows the scores of the two principalcomponent axes for all the accessions. Each acces-sion is shown as metroglyph by depicting the degreeof pigmentation, presence or absence of trichomesand pod divergence semi-quantitatively. The 32 acces-sions (metroglyphs) fell into 7 groups (4 clusters ofaccessions and 3 disparate accessions) covering mostparts of the principal component space (PCS). A largecluster of 16 accessions fell into the centre of the PCS(Cluster 2). A large majority of these had origins insemi-tropical to semi-temperate geographical areas.The five accessions of the cluster-1 got placed in theleft upper quadrant of PCS. They had large originsin the tropical areas. Cluster-5 of 4 accessions alsocomprised of accessions from high rainfall and humidareas. Cluster-3 had accessions from semi-tropical tosemi-temperate areas. An accession each from An-daman islands and Singapore and a mutant of theaccession Crc-9 of cluster-2 released as a cultivar, gotseparated from each other and the rest of accessionsby principal component analysis.
Table 4. Contribution of Eigenvectors of 20 plant vigour and al-kaloid yield related characters tothe two principal components ina collection of 32 accessions ofCatharanthus roseus.
Sl. Trait Eigen vector
no. PC1 PC2
1. PHT −0.04 0.08
2. LNB 0.12 0.14
3. LAR 0.36 0.54
4. RDY −0.02 0.04
5. LDY 0.07 0.14
6. RAC 0.19 0.14
7. RAY 0.17 0.18
8. LAC 0.07 0.05
9. LAY 0.70 −0.56
10. VRC 0.05 −0.22
11. VRY 0.31 0.24
12. VBC 0.01 0.08
13. VBY 0.14 −0.22
14. LMC 0.28 0.09
15. FDI −0.14 0.08
16. CTL −0.15 −0.18
17. STL − 0.13 −0.19
18. POF −0.13 −0.08
19. FOL −0.04 −0.18
20. HSW 0.05 0.03
Table 5. Mean intra- and inter-cluster distances among theseven clusters formed by 32 accessions of Catharanthusroseus based on 20 plant vigour and alkaloid yield relatedcharacters.
Cluster 1 2 3 4 5 6 7
1 3045
2 7688 1988
3 9679 6819 3148
4 17136 13975 5934 –
5 13126 5138 5237 5946 2332
6 26806 11953 13711 9563 4948 –
7 24638 10002 19214 20649 8494 5841 –
The distances between the clusters are presentedin Table 5. The mutual differences were very largebetween the clusters/groups 1 and 6, 1 and 7, 3 and7, and 4 and 7. The clusters 2,3 and 5 occupiedintermediate positions with respect to the others.
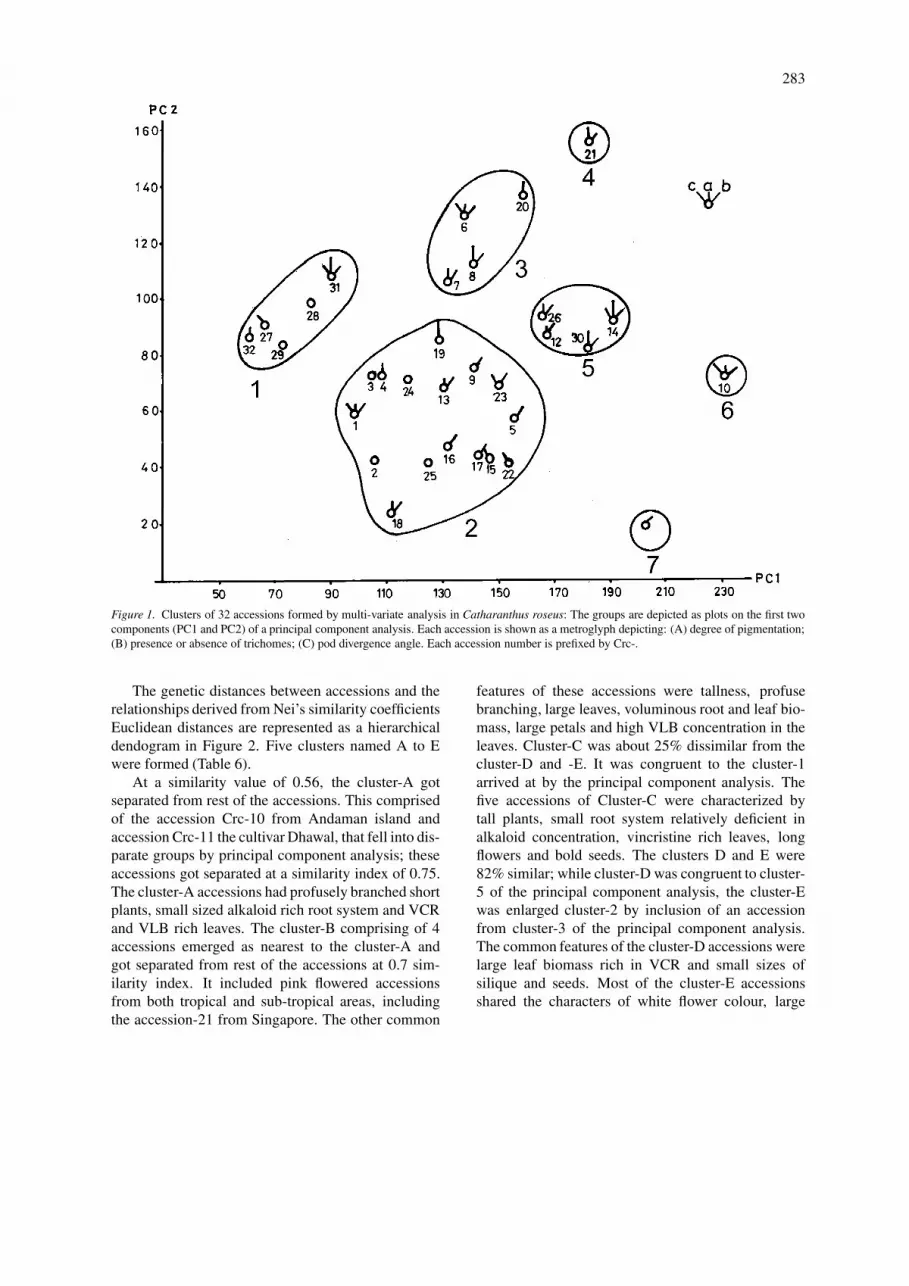
283
Figure 1. Clusters of 32 accessions formed by multi-variate analysis in Catharanthus roseus: The groups are depicted as plots on the first twocomponents (PC1 and PC2) of a principal component analysis. Each accession is shown as a metroglyph depicting: (A) degree of pigmentation;(B) presence or absence of trichomes; (C) pod divergence angle. Each accession number is prefixed by Crc-.
The genetic distances between accessions and therelationships derived from Nei’s similarity coefficientsEuclidean distances are represented as a hierarchicaldendogram in Figure 2. Five clusters named A to Ewere formed (Table 6).
At a similarity value of 0.56, the cluster-A gotseparated from rest of the accessions. This comprisedof the accession Crc-10 from Andaman island andaccession Crc-11 the cultivar Dhawal, that fell into dis-parate groups by principal component analysis; theseaccessions got separated at a similarity index of 0.75.The cluster-A accessions had profusely branched shortplants, small sized alkaloid rich root system and VCRand VLB rich leaves. The cluster-B comprising of 4accessions emerged as nearest to the cluster-A andgot separated from rest of the accessions at 0.7 sim-ilarity index. It included pink flowered accessionsfrom both tropical and sub-tropical areas, includingthe accession-21 from Singapore. The other common
features of these accessions were tallness, profusebranching, large leaves, voluminous root and leaf bio-mass, large petals and high VLB concentration in theleaves. Cluster-C was about 25% dissimilar from thecluster-D and -E. It was congruent to the cluster-1arrived at by the principal component analysis. Thefive accessions of Cluster-C were characterized bytall plants, small root system relatively deficient inalkaloid concentration, vincristine rich leaves, longflowers and bold seeds. The clusters D and E were82% similar; while cluster-D was congruent to cluster-5 of the principal component analysis, the cluster-Ewas enlarged cluster-2 by inclusion of an accessionfrom cluster-3 of the principal component analysis.The common features of the cluster-D accessions werelarge leaf biomass rich in VCR and small sizes ofsilique and seeds. Most of the cluster-E accessionsshared the characters of white flower colour, large

284
Table 6. Mean and standard errors for 20 traits for hierarchical groups derived from the dendrogram linking the 32 examinedaccessions of Catharanthus roseus.
Hierarchical group (subgroups or/accession(s)) forming the group as per the dendrogram given in Figure 2
Trait A B C D E
(Crc-10,11) (Crc-20,21,6,8) (Crc-27,28,29, (Crc-12,14,26,30) (Crc-1,2,3,4,5,7,9,
31,32) 13,15,16, 17,
18,19,22,23,24,25)
PHT 62.2 ± 7.5 85.9 ± 6.1 79.4 ± 6.9 68.1 ± 9.2 71.4 ± 2.3
LNB 739 ± 193 771 ± 103 556 ± 73 669 ± 67 545 ± 7
LAR 9.6 ± 1.3 11.0 ± 0.9 7.2 ± 0.4 10.6 ± 0.6 10.6 ± 3.2
RDY 397 ± 132 846 ± 226 370 ± 51 475 ± 56 532 ± 42
LDY 1871 ± 257 2566 ± 296 1662 ± 133 2037 ± 155 1816 ± 117
RAC 1.9 ± 0.26 1.2 ± 0.49 0.2 ± 0.63 1.6 ± 0.11 1.2 ± 0.06
RAY 3.3 ± 2.3 9.7 ± 2.2 0.8 ± 0.2 7.1 ± 1.6 6.0 ± 0.6
LAC 1.2 ± 0.06 1.1 ± 0.57 1.0 ± 0.05 1.0 ± 0.08 1.0 ± 0.04
LAY 22.6 ± 2.2 24.5 ± 6.9 15.9 ± 1.3 21.0 ± 2.1 16.0 ± 1.1
VCC 4.9 ± 3.3 0.9 ± 0.3 3.2 ± 1.4 2.5 ± 1.3 0.9 ± 0.1
VCY 8.15 ± 4.85 2.02 ± 0.67 15.80 ± 11.74 6.12 ± 3.80 1.36 ± 0.20
VBC 19.6 ± 1.1 15.8 ± 3.5 2.6 ± 0.7 12.6 ± 3.0 10.8 ± 1.2
VBY 36.8 ± 7.40 32.5 ± 4.03 19.32 ± 5.27 32.4 ± 6.54 9.43 ± 2.29
LMC 80 ± 6.5 76 ± 1.5 80 ± 2.0 84 ± 2.8 81 ± 1.1
FDI 2.9 ± 0.05 4.1 ± 0.28 4.1 ± 0.39 3.6 ± 0.54 3.7 ± 0.16
CTL 22.5 ± 1.50 25.7 ± 0.75 26.0 ± 0.68 23.2 ± 0.85 21.9 ± 1.42
STL 1.55 ± 0.35 1.87 ± 0.07 1.98 ± 0.58 1.95 ± 0.13 1.96 ± 0.53
PFT 93 ± 3.0 73 ± 8.3 78 ± 5.4 83 ± 4.3 81 ± 1.9
FOL 24.5 ± 6.2 24.6 ± 1.2 26.2 ± 2.8 21.8 ± 1.6 25.3 ± 0.9
HSW 103.5 ± 7.5 100.0 ± 10.8 115.8 ± 4.2 98.2 ± 5.3 110.9 ± 4.5
leaves and low concentrations of alkaloids in leafbiomass.
Discussion
C. roseus is believed to have the centre of origin inMadagascar from where it has spread world wide asan ornamental garden plant. A founding population ofthis plant was introduced into southern parts of Indiaby Portuguese missionaries about 200 years ago; be-sides there might have been subsequent augmentationsof this base population by new introductions into In-dia. In time, C. roseus became a common garden plantin most parts of Indian subcontinent. Since introduc-tion, the plant has become naturalized and has beenfound growing in different plant communities andvarious soil conditions of tropical to semi-temperateagro-climates of the subcontinent. Genetic drift res-ulting from cross-pollination mode of sexual repro-duction would have been responsible for the observed
wide establishment of the species in nature. Thepresent characterization of variability in more than 50morphological and biochemical characters among theaccession samples representing the spread of C. roseuspopulations in Indian subcontinent and neighbouringcountries has led to categorization of accessions intofive hierarchically related morphological and presum-ably genetically distinguishable groups. Those comingfrom the tropical agro-climates tended to got separatedfrom those having origins in sub-tropical to semi-temperate environments (Cluster-E). The mixing upof accessions coming from variable agro-climates incommon clusters (Cluster-B, -D and -E) was perhapsrelated to the nature of reproductive system in C. ros-eus (Souze and Sorrells, 1991). The flower structureof C. roseus is suitable for insect mediated pollina-tion, both self and cross-pollinations would occur inthe flowers borne on a plant. The allogamy relatedgene flow should be considered responsible for mak-ing populations from different edaphic areas similar.

285
Figure 2. Dendrogram showing hierarchical relationships between32 accessions of periwinkle Catharanthus roseus, based on variationin 20 plant vigour and alkaloid yield related characters. The degreesof differences between the accessions are depicted using 100 as theuppermost level.
However, the genetic base of the populations of C.roseus in Indian subcontinent and the neighbouringAsian region must be narrow, as half of the accessionscoming from wide geographical distances got placedtogether in clusters formed by multivariate principalcomponent and heirarchical UPGMA analysis. Thisclustering together of subtropical accessions substan-tiated the view of Zhong and Qualset (1995), that insuch accessions co-adaptive associations of the fitnesscharacters got similarly selected.
The present analysis of variability in C. roseus ac-cessions has allowed identification of certain cluster-A, -B, and -D members as bearing some of the comple-mentary characters for cross-parenting transgressivesegregants (Beer et al. 1993) from which high yieldsof dimeric alkaloids could be obtained. While therewere about 3, 80 and 15 fold differences between thelowest and highest values of leaf dry matter yield andVCR and VLB concentrations in leaves, 8 of the ac-cessions contained VLB+VCR at 5 fold higher level
than the one in which the level was minimum. In oneof the accessions (Crc-31), VCR was more than 74fold higher and VLB 12 fold higher than the respectivelowest values among all the accessions, this accessionCrc-31 appears to be suitable for direct cultivation.This accession and Crc-11 and accessions Crc-10 and-21 appear suitable as explant resource for developinghairy root cultures for obtaining high yields of dimericalkaloids vincristine and vinblastine, respectively.
Many secondary metabolites, including some ofthe alkaloids of C. roseus, have been found to provideprotection against biotic stresses when sprayed on orsynthesized in other plants (Bennet and Wallsgrove1994; Grayer and Harborne 1994; Rhodes 1994).These appear to be especially effective against a vari-ety of insect pests (Wink 1993). Synthesis and accu-mulation of high amounts of total alkaloids and/or oneor both of dimeric alkaloids by certain accessions maybe a reflection of reproductive fitness achieved in themunder heavy loads of insect pests and fungal diseases,in the tropical agro-climatic conditions of growth andevolution that this species underwent.
References
Aerts R.J., Gise P., Decarolis E., De Luca V. and Baumann T.W.1994. Methyl jasmonate vapors increase the developmentallycontrolled synthesis of alkaloids in Catharanthus roseus andCinchona seedlings. Plant J. 5: 635–643.
Beer S.C., Goffreda J.G., Phillips T.D., Murphy J.P. and SorrelsM.E. 1993. Assessment of genetic variation in Avena sterilesusing morphological traits, isozymes and RFLPs. Crop Sci. 33:1386–1393.
Bennet R.N. and Wallsgrove R.M. 1994. Secondary metabolites inplant defence mechanism. New Phytol. 127: 617–633.
Cochran W.G. and Cox G.M. 1957. Experimental Designs, 2nd ed.J. Wiley, New York.
Frischknecht P.M., Battig M. and Baumann T.W. 1987. Effect ofdrought and wounding stress on indole alkaloid formation inCatharanthus roseus. Phytochem. 26: 707–710.
Godoy-Harnandez G. and Loyola-Vargas V.M. 1991. Effect offungal homogenate, enzyme-inhibitors and osmotic stress on al-kaloid content of Catharanthus roseus cell suspension cultures.Plant Cell Rep. 10: 537–540.
Grayer R.J and Harborne J.B. 1994. A survey of antifungal com-pounds from higher plants. Phytochem. 37: 19–42.
Gröger D. 1985. Alkaloids derived from tryptophan. In: Mothes K.,Schütte H.R. and Luckner M. (Eds.), Biochemistry of Alkaloids,pp. 272–331. VEB Deutscher Verlag der Wissenschaften, Berlin.
Jordan M.A., Thrower D. and Wilsen L. 1991. Mechanism of in-hibition of cell proliferation by Vinca alkaloids. Cancer Res. 51:2212–2222.
Kulkarni R.N., Baskaran K., Chandrashekhara R.S. and Kumar S.1999. Inheritance of morphological traits of periwinkle mutantswith modified contents and yields of leaf and root alkaloid. PlantBreed. 118: 71–74.

286
Luijendijk T.J.C., Vander Heijden R. and Verpoorte R. 1996. In-volvement of strictosidine as a defensive chemical in Cathar-anthus roseus. J. Chem. Ecol. 22: 1355–1366.
Mahalanobis P.C. 1936. On the generalised distance in statistics.Proc. of National Institute of Science 2: 49–55.
Menke F.L.H., Parchmann S., Mueller M.J., Kijne J.W. andMemelink J. 1999. Involvement of the octadecanoid pathwayand protein phosphorylation in fungal elicitor induced expressionof terpenoid indole alkaloid biosynthetic genes in Catharanthusroseus. Plant Physiol. 119: 1289–1296.
Molner A., Liliom K., Firosz F., Vertessy B.G. and Ovadi J. 1995.Anti-Calimodulin potency of indole alkaloids in in vitro systemsEur. J. Pharmakol. 291: 73–82.
Moreno R.H., Van Der Heijden R. and Verpoorte R. 1995. Celland tissue culture of Catharanthus roseus: A literature surveyII Updating 1988 to 1993. Plant Cell Tiss. Org. Cult. 42: 1–25.
Naaranlahti T., Nordstorm M. and Huhtikangas A. 1987. De-termination of Catharanthus alkaloids by reversed phase highperformance liquid chromatography. J. Chromat. 410: 488–493.
Ramanujam S. and Kumar S. 1964. Metroglyph analysis of geo-graphical complexes in Indian Vetiver. Ind. J. Genet. Breed. 24:144–150.
Rao C.R. 1952. Advanced Statistical Methods in Biometrical Re-search, p. 390. John Wiley and Sons Inc., New York.
Rhodes M.J.C. 1994. Physiological roles of secondary metabolitesin plants: some progress, many outstanding problems. Plant Mol.Biol. 24: 1–20.
Saenz K., Santamaria J.M., Villaneuva M.A., Loyala Vargas V.M.and Oropeza C. 1993. Change in alkaloid content of plant ofCatharanthus roseus (L.) G. Don as a result of water stress andtreatment with abscisic acid. J. Plant Physiol. 142: 244–247.
Sharma S., Tyagi B.R., Mandal S., Singh V. and Singh H. 1996.Cluster analysis of 38 genotypes of peppermint (Mentha piper-
ita) based on essential oil yield and quality traits. J. Med. Arom.Plant Sci. 18: 280–286.
Son J.K., Rosazza J.P.N. and Duffel M.W. 1990. Vinblastine andvincristine inhibitors of monoamine oxidase B. J. Med. Chem.33: 1845–1848.
Souze E. and Sorrelo M.E. 1991. Relationships among 70 AmericanOat Germplasm I. Cluster analysis using quantitative characters.Crop Sci. 31: 599–605.
Talha M., Radwan A.S. and Negm S. 1975. The effect of soil mois-ture deficit on growth and alkaloidal content of Catharanthusroseus G. Don. Curr. Sci. 44(17): 614–616.
Thomas J.C., Adams D.G., Nessler C.L., Brown J.K. and Bo-hnert H.J. 1995. Tryptophan decarboxylase, tryptamine andreproduction of white fly. Plant Physiol. 115: 717–720.
Tyler V.E. 1988. Medicinal plant research. Planta Med. 54: 95–100.Vazquez-Flota F. and Deluca V. 1998. Development and light reg-
ulation of desacetoxy-vindoline 4-hydroxylase in Catharanthusroseus (L.) Don. Plant Physiol. 117: 1351–1361.
Vazquez-Flota F., Moreno-Velenzuela O., Miranda-Ham M.L.,Coello-Coello J. and Loyola-Vargas V.M. 1994. Catharanth-ine and ajmalicine synthesis in Catharanthus roseus hairy rootcultures. Plant Cell Tiss. Org. Cult. 38: 273–279.
Virk S.S. and Singh O.S. 1990. Osmotic properties of droughtstressed perwinkle (Catharanthus roseus) genotype. Ann. Bot.66: 23–30.
Wink M. 1993. Allelochemic properties or the raison d’etre of al-kaloids. In: Brossi, A. (Ed.), The Alkaloids, 43, pp. 1–118.Academic Press, San Diego.
Zhong G.Y. and Qualsit C.O. 1995. Quantitative genetic diversityand conservation strategies for an allogamous annual species –Dasypyruune villosum (L.) Candargy (Poaceae). Theor. Appl.Genet. 91: 1064–1073.