partial purification and kinetic characterization of the microsomal phospholipase a2 from thermally...
TRANSCRIPT

J Comp Physiol B (1985) 155:461M69 Journal of Comparative B,~.~,~, . Systemic,
and Environ-
Physiology B .~ Physiology
�9 Springer-Verlag 1985
Partial purification and kinetic characterization of the microsomal phospholipase A 2 from thermally acclimated rainbow trout (SMmo gMrdnen) N.P. Neas* and J.R. Hazel Department of Zoology, Arizona State University, Tempe, Arizona 85287, USA
Accepted October 14, 1984
Summary. Phospholipase A 2 (PLAz) was extracted from liver microsomal membranes of both 5 and 20 ~ rainbow trout (Salrno gairdneri), using the non-ionic detergent, Triton X-100. Fur- ther purification was achieved by precipitation with 35-65% ammonium sulfate followed by gel filtration chromatography in the presence of 0.1% Triton X-100 on Sephadex G-200. These proce- dures resulted in a 30-fold purification and the re- moval of all traces of phospholipid from the en- zyme of both warm- and cold-acclimated trout. Column elution profiles were similar for both accli- mation groups, yielding a molecular weight esti- mate for the trout liver enzyme of 73,000. Compar- isons of activity levels and kinetic parameters of PLA 2 from warm- and cold-acclimated fish indi- cated that compensation for temperature at non- saturating substrate concentrations was an attri- bute of both the particulate (microsomal) enzyme and the lipid-free protein. Cold acclimation re- sulted in higher activity below Vma x due primarily to decreased apparent K m values. These adapta- tions to temperature could not be attributed to the interaction of the enzyme with the membrane tipids, but were due to qualitative changes in the enzyme that resulted from acclimation. Other adaptive qualities of PLA2, such as reduced K m in response to acute decreases in temperature in warm-acclimated fish, were only apparent in parti- culate preparations, and thus were a function of the protein-lipid complex. These data suggest that an acclimation-induced increase in the activity of
Abbreviations: P A G E polyacrylamide gel electrophoresis; PC phosphatidylcholine; P L A 2 phospholipase A2; S D S sodium dodecylsulfate
* Present address: Department of Biology, Colgate University, Hamilton, NY 13346, USA
PLA 2 may result in the activation of a deacylation- reacylation cycle at cold temperatures.
Introduction
In poikilothermic animals, acclimation to low tem- perature results in alterations in membrane lipid composition. These changes offset the direct effects of temperature on the physicochemical properties of lipids and conserve the vital structural and func- tional properties of cell membranes at cold temper- atures (Cossins 1981; Dickens and Thompson 1981). Among these changes is the substitution of unsaturated fatty acids for saturated and monoen- oic acids on membrane phosphatides of cold-accli- mated animals. In rainbow trout, for example, cold acclimation resulted in a significant increase in the polyenes, 20:5 and 22:6, with a concurrent de- crease in oleate (18: 1) and linoleate (1.8:2) at the sn-two carbon of membrane choline phosphogly- ceride (Hazel J979). Studies on the enzymatic basis for membrane restructuring have traditonally fo- cused on the anabolic pathways of membrane me- tabolism, such as fatty acid desaturation (Sellner and Hazel J982) and de novo synthesis of phos- pholipids (Hazel etal. 1983). Recent evidence, however, has implicated pathways involving se- quential phospholipid deacylation, acyl modifica- tion and subsequent reacylation.
When cultures of Tetrahymena were exposed to chill stress, a rapid retaitoring of molecular spe- cies resulted in equally rapid reestablishment of membrane fluidity with little or no net change in fatty acid composition (Dickens and Thompson 1981, 1982; Ramesha and Thompson 1982). The authors inferred the action of a deacylation-reacy-

462 N.P. Neas and J.R. Hazel : Purification of trout phospholipase A z
lation cycle in achieving this retailoring. An overall increase in unsaturation did occur, however, with prolonged exposure to low temperatures (Fukus- hima et al. 1976).
Deacylation-reacylation pathways have also been implicated in the metabolism of phospholip- ids by liver and gill tissues of cold acclimated rain- bow trout where the temperature-dependent rate of turnover of phospholipid glycerol moieties was significantly different from that of fatty acyl moie- ties (Hazel and Neas 1982). Thus the deacylation- reacylation cycle, well documented in the synthesis of unique molecular species (i.e., different from those produced by de novo synthesis) of mamma- lian phospholipids (van den Bosch 1974; Holub and Kuksis 1978) is implicated in thermally modu- lated restructuring of cell membranes in poikiloth- erms.
The deacylation and reacylation enzymes, phospholipases A 1 and A z (PLA 1 and PLAz) and acyl-CoA: 1-acyl-sn-glycero-3-phosphorylcholine acyltransferase, respectively, have been identified in the microsomes of rainbow trout (Jonas and Bilinski 1967; Neas and Hazel 1984; Holub et al. 1976). Holub et al. (1976) reported no significant effect of acclimation or assay temperature on the activity or specificity of the acyltransferase to- wards palmitoyl- or oleoyl-CoA derivatives. A pre- vious report from our laboratory, however, dem- onstrated that trout liver microsomal phospholi- pase was predominantly of the A 2 type, and that activity of the enzyme from 5 ~ fish exceeded that from 20 ~ fish at assay temperatures of both 5 and 20 ~ (Neas and Hazel 1984). The positional specificity and conservation of activity of the enzyme upon cold acclimation is consistent with the previously noted high levels of thermally induced fatty acid substitution at car- bon sn-two (Hazel 1979) and the differential turn- over rates of glycerol and acyl moieties in cold- acclimated rainbow trout (Hazel and Neas 1982), and suggests a role for PLA 2 in thermally modu- lated membrane restructuring in rainbow trout.
In order to further examine the role of phos- pholipase A 2 in membrane remodeling, the enzyme was partially purified from microsomal mem- branes of rainbow trout and kinetic parameters were related to acclimation history and assay tem- perature. It has been shown that both the activity and kinetic properties of membrane-bound en- zymes are influenced by changes in the lipid micro- milieu (Farias et al. 1975). In addition, the com- pensatory adjustments in activity observed in some membrane-bound enzymes of thermally acclimated poikilotherms may be a function of changes in the
lipid matrix, and not the enzyme protein (Hazel 1972a, b). Consequently, we have investigated the role of membrane lipids in determining the proper- ties of trout liver microsomal phospholipase A 2 by comparing the activity levels and kinetic param- eters of the enzyme from particulate microsomes with those of lipid-free soluble preparations.
Materials and methods
Materials. Egg phosphatidylcholine and lysophosphatidylcho- line were purchased from Sigma. 1-[3H]-oleic acid was a prod- uct of Amersham Corp., Arlington Heights, I1. Enzymatic grade ammonium sulfate was purchased from Nutritional Biochemi- cals Corp., Cleveland, OH. Sephadex G-200 was a product of Pharmacia Fine Chemicals, Piscataway, NJ. Synthesis of 1-acyl,2-[3H]-oleoyl phosphatidylcholine was as described pre- viously (Neas and Hazel 1984) and according to Conner et al. (1981). Over 99% of the radiolabel in the resulting product was incorporated at the 2-acyl position as determined by snake venom hydrolysis (Dickens and Thompson 1982). Radiola- belled phosphatidylcholine was stored in CHC1jMeOH (2:1, v/v) at - 2 0 ~ under N 2 and was stable for several months. Specific radioactivity was determined periodically by counting aliquots and dividing by molar concentration as determined by phosphorus analysis (Rouser et al. 1970).
Animals. Rainbow trout (Salmo gairdneri) were kindly provided by the Alchesay National Fish Hatchery in Whiteriver, AZ. The animals were acclimated in the laboratory to either 5 or 20 ~ in running water and a 12L: 12D photoregime. Fish were fed once daily with Glencoe Mills Trout Food and were held for at least one month prior to experimentation. At both tem- peratures the trout exhibited normal activity and feeding behav- ior and good health for the duration of the study.
Enzyme purification. For each experiment, three to eight trout livers (0.6-2.4 g) were excised from freshly decapitated fish and homogenized in 6 volumes (w/v) of 0.25 M sucrose, 10 mM Tris-acetate (pH 7.8 at 20 ~ 1 mM EDTA (buffer I) using eight strokes of a teflon tissue homogenizer driven by an electric motor at 1550 rpm. The crude homogenate was centrifuged at 18,000 g for 30 rain, and the decanted supernatant was recen- trifuged at 100,000 g for 60 rain to obtain the microsomal pellet. The surface of this pellet was washed with buffer I and the pellet was resuspended using a ground glass Ten Broeck hand homogenizer in either buffer II (0A M Tris-acetate, pH 8.0, 8 mM CaC12, 0.1% Triton X-100) to a final concentration of 2-3 mg/ml for further purification, or buffer III (0.1 M Tris- acetate, pH 8.0, 8 mM CaCI2) to a final concentration of 20 mg/ml for subsequent enzyme assay. Microsomes were incu- bated in buffer II with constant stirring at 0 ~ for 90 rain, and recentrifuged at 100,000 g for 90 rain. The supernatant, designated 'Tri ton extract', was precipitated with ammonium sulfate; aqueous ammonium sulfate at 100% saturation at 4 ~ was used to raise the concentration to the desired level. The fraction that precipitated between 35 to 65% saturation was centrifuged at 35,000 g for 60 rain, and the fragile pellet was redissolved in 2-3 ml of appropriate buffer (either buffer II or III) at pH 8.0 for Sephadex G-200 column chromatography. Residual ammonium sulfate was removed by dialysis against 500 ml of the column buffer for 12 h.
The dialysed material was filtered through a 1.5 x 60 cm column of Sephadex G-200 at room temperature (23 ~ at

N.P. Neas and J.R. Hazel: Purification of trout phospholipase A 2 463
a flow rate of 0.5 ml/min. Fractions of 5 ml each were collected and assayed for phospholipase A 2 activity.
Enzyme assays. Phospholipase A 2 activity was estimated using a fixed time assay (Neas and Hazel 1984). The assay cocktail was prepared using 0.1 pCi 1-acyl,2-[3H]-oleoyl phosphatidyl- choline in CHC13/MeOH (2:1, v/v) and the appropriate quan- tity of unlabelled egg phosphatidylcholine to yield the desired substrate concentration (100 ~tM, unless otherwise stipulated). Organic solvents were removed under a stream of N 2. For mi- crosomal assays, 0.9 ml of buffer II was added, the mixture was sonicated for two minutes in a bath sonicator and the assays were initiated by the addition of a 0.1 ml aliquot of microsomal suspension (2 mg protein), bringing the final assay volume to 1.0 ml. Fractions from Sephadex G-200 were assayed on a volume-specific basis. The assay cocktail was prepared as above, but in a volume of 0.2 ml for each assay. Aliquots of column fractions of 0.8 ml were added to initiate each assay, again bringing the final assay volume to 1.0 ml. Assays were terminated by the addition of 3 ml CHC13/MeOH (1:2, v/v), and lipids were extracted into CHC13 using the method of Bligh and Dyer (I959) and stored at - 2 0 ~ in biphasic form. Phos- pholipase A 2 activity was determined by separating the prod- ucts of the reaction using thin layer chromatography and mea- suring the amount of radiolabel present in the lipid extract as free oleic acid, lysophosphatidylcholine and unhydrolysed PC from each assay as described previously (Neas and Hazel 1984). All assays were corrected for non-enzymatic hydrolysis as measured under identical conditions using either boiled mi- crosomal protein or an aliquot of the first column fraction (we-void volume eluant).
Experiments reported elsewhere (Neas and Hazel 1984) indi- cated that, while there may be some PLA 1 activity in trout liver microsomes, the PLA 2 reaction is the rate-limiting step of the deacylation of phosphatidylcholine.
For kinetic analysis, Lineweaver-Burk plots were generated by weighted linear regression analyses (Wilkinson 1961) which compensated for the reduced reliability of measurements made at low substrate concentrations. Due to a concentration-depen- dent increase in non-enzymatic hydrolysis at pH 8.0, particular- ly at low substrate levels, assays for kinetic analysis were per- formed at pH 6.5. Atthough the pH optimum was previously determined to be between 8 and 9 (Neas and Hazel 1984), over 85% of the optimal activity was retained at the lower pH, and non-enzymatic hydrolysis did not exceed 5% of the radiola- belled substrate at any substrate concentration. The duration of these assays was 45 rain, which was well within the limits of linearity of hydrolysis with time at all substrate concentra- tions (data not shown).
Catalase used to calibrate the G-200 column was assayed according to the procedure of Lfick (1963).
SDS-polyacrylamide gel electrophoresis. Slab SDS-polyacryl- amide gel electrophoresis (SDS-PAGE) was carried out using a modification of the technique of Laemmli (1970). A separat- ing gel of 9.2% total acrylamide in 1.8% crosslinking agent, BIS (N,N'inethylene-bis acrylamide, w/v), and a stacking gel of 1.8% total acrylamide in 2.6% BIS was used. The buffer used for the electrophoretic run was 50 mM Tris-glycine, 0.1% (w/v) SDS, pH 8.8.
Protein samples were treated with equal volumes of denatur- ing buffer (0.125 M Tris-HCl, pH 8.8, 10% fl-mercaptoethanol, 4.6% SDS, 20% glycerol and 0.2% bromphenol blue) at 90 ~ for 10 min and applied to 1 cm wells in the stacking gel in 100 gl aliquots. The electrophoretic run was carried out at 4 ~ using a vertical slab electrophoresis assembly (Model SE 520, Hoefer Scientific Instruments, San Francisco, CA). A constant
current of 18 mA per gel was applied until the bromphenol blue tracking dye reached the running gel; the current was then increased to 40 mA per gel until the end of the run. Each gel was equilibrated to 50% aqueous MeOH for 12 h, stained with ammoniacal silver according to Wray et al. (1981) and photographed immediately.
Protein and lipid determinations. Protein concentrations were determined using the method of Bradford (1976) with concen- trated dye reagent from BioRad Laboratories, Richmond, CA. Protein of fractions eluted from Sephadex G-200 in the presence of detergent was first dialyzed against distilled water for 46 h to minimize the interference of Triton X-100. Appropriate cor- rections for changes in sample volume as a function of dialysis were made.
Phospholipid determinations were done by extracting the samples at the various stages of purification with organic sol- vents (Bligh and Dyer 1959), and assaying for phosphate in acid digests of the extracts according to Rouser et al. (1970).
Results
A previous study from this laboratory showed that the microsomal fraction of rainbow trout liver ac- counted for more than 18% of the total cellular activity ofphospholipase A 2 (PLA2; Neas and Ha- zel 1984). As illustrated in Table 1, extraction of microsomes with buffer alone solubilized 22% of the total microsomal protein, and a comparable amount (24%) of PLA 2 activity. The presence of 0.1% Triton X-100 solubilized 39% of the micro- somal protein, but over 58% of the PLA 2 activity. This resulted in a 1.8- and 1.9-fold purification of PLA 2 from 5- and 20 ~ fish, respec- tively (Table 2). Further purification of the solubi- lized portion of microsomal PLA 2 was accom- plished by ammonium sulfate precipitation (35-65%) of the Triton extract. This resulted in
4 - and 3-fold purifications from warm- and cold- acclimated fish, respectively, and removed over 95% of the phospholipid from the enzyme. All re- maining lipid was removed during gel filtration chromatography on Sephadex G-200. At this
Table 1. Percent of protein and PLA 2 activity extracted from microsomes by detergent"
-Dete rgen t b +Detergent c
Super- Pellet Super- Pellet natant natant
Protein 22 78 39 61 PLA 2 activity 24 76 59 41
Microsomes from cold-acclimated trout were treated for 90 rain with buffer and centrifuged at 100,000 g for 60 rain as described in the text. Total recovery of protein exceeded 85 % in both detergent and detergent-free extractions
b Buffer III (0.1 M TRIS-acetate, pH 8.0, 8 mM CaC12) c Buffer II (0.1 M TRIS-acetate, pH 8.0, 8 mM CaC12, 0.1%
[v/v] Triton-X-100)
464 N.P. Neas and J.R. Hazel: Purification of trout phospholipase A 2
Table 2. Purification of PLA 2 from liver tissue of thermally acclimated trout. Microsomes were extracted with 0.1% Triton X-100, precipitated with 35-65% ammonium sulfate and filtered on Sephadex G-200. All assays were performed at the respective acclimation temperatures
Fraction Acclimation Total Lipid Phospho l ipase A 2
Temperature Protein (nM/mg (~ (mg) protein)
Specific Total % - Fold activity activity Recovery a Puri- (nM/mg. h) (nM/h) fication
Microsomes 5 189.8 293.1 1.12 212.58 100 1.0 20 181.1 831.3 1.28 231.74 100 1.0
Triton extract 5 50.1 235.7 2.03 101.78 47.9 1.8 20 47.9 125.0 2.46 117.85 50.85 1.9
Triton pellet 5 79.4 - 0.98 77.84 36.62 - 20 60.2 - 1.18 71.08 30.67 -
(NH4)2SO 4 precipitate 5 15.1 11.2 4.55 68.79 32.35 4.1 20 17.4 4.1 3.88 67.43 31.69 3.0
G-200 eluant 5 2.4 nd b 40.36 97.26 45.75 36.0 20 2.5 nd 35.42 89.61 38.67 27.6
Percent recovery is expressed as the proport ion of microsomal PLA 2 activity recovered at each step b None detectable
0.9 ~ ' ~ ~ 5"acclimated
0.7 I.O 5.o
0.6 4.0
o 4 t / / | I', / , . . . ,.o o s ~ / / ,'. / / ' [ ~ - - . ,' ",
-I ~ 0.7
~ 0 . 6
0.5'
0.4'
O3'
0.2'
0.1'
0
20~
A h,
/ I / !, ,.- / ~,, ~ - - ' r - -
50 I00 150 200
25
2O
-15 T
-IO -~
r
-25
-20
-15
-I0
ELUTION VOLUME (ml)
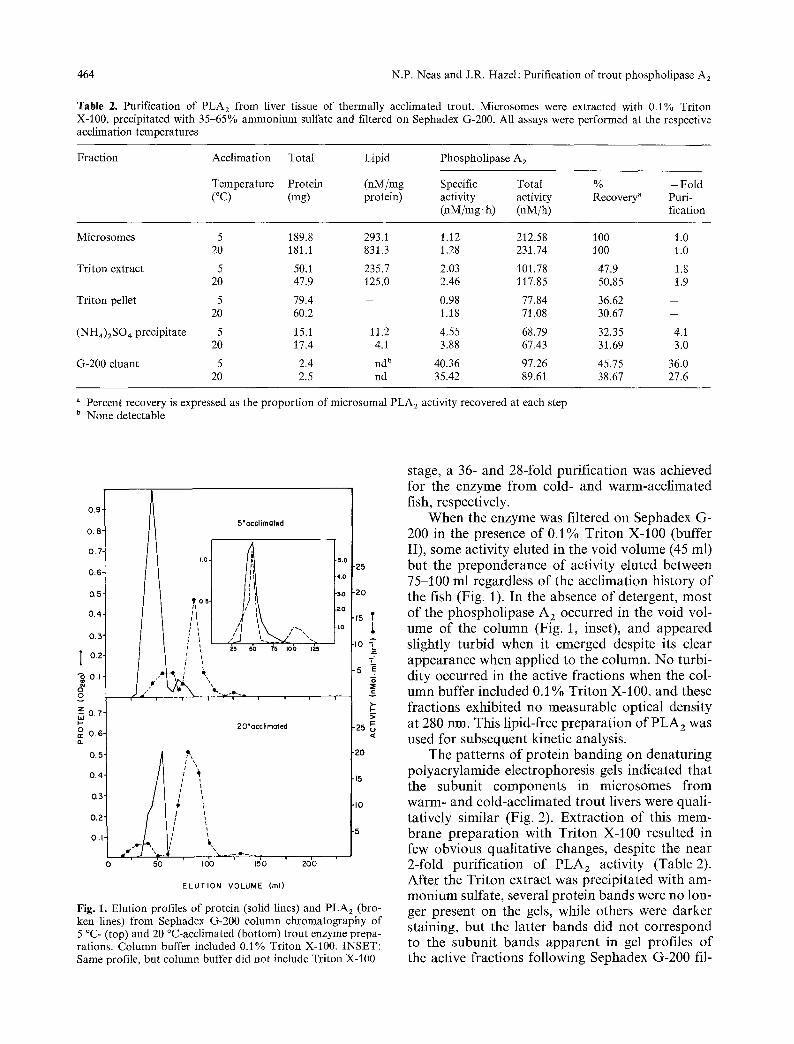
Fig. 1. Elution profiles of protein (solid lines) and PLA 2 (bro- ken lines) from Sephadex G-200 column chromatography of 5 ~ (top) and 20 ~ (bottom) trout enzyme prepa- rations. Column buffer included 0A % Triton X-100. INSET: Same profile, but column buffer did not include Triton X-100
stage, a 36- and 28-fold purification was achieved for the enzyme from cold- and warm-acclimated fish, respectively.
When the enzyme was filtered on Sephadex G- 200 in the presence of 0.1% Triton X-100 (buffer II), some activity eluted in the void volume (45 ml) but the preponderance of activity eluted between 75-100 ml regardless of the acclimation history of the fish (Fig. 1). In the absence of detergent, most of the phospholipase A z occurred in the void vol- ume of the column (Fig. 1, inset), and appeared slightly turbid when it emerged despite its clear appearance when applied to the column. No turbi- dity occurred in the active fractions when the col- umn buffer included 0.1% Triton X-100, and these fractions exhibited no measurable optical density at 280 nm. This lipid-free preparation of PLA 2 was used for subsequent kinetic analysis.
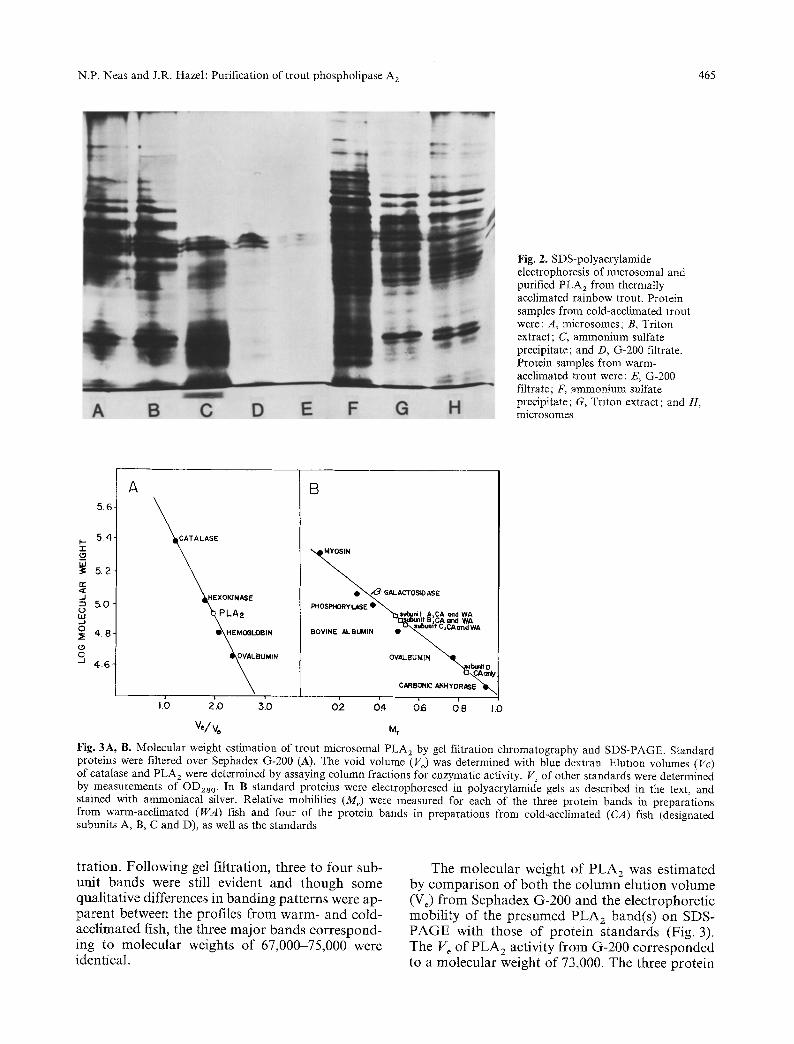
The patterns of protein banding on denaturing polyacrylamide electrophoresis gels indicated that the subunit components in microsomes from warm- and cold-acclimated trout livers were quali- tatively similar (Fig. 2). Extraction of this mem- brane preparation with Triton X-100 resulted in few obvious qualitative changes, despite the near 2-fold purification of PLA 2 activity (Table 2). After the Triton extract was precipitated with am- monium sulfate, several protein bands were no lon- ger present on the gels, while others were darker staining, but the latter bands did not correspond to the subunit bands apparent in gel profiles of the active fractions following Sephadex G-200 ill-
N.P. Neas and J.R. Hazel: Purification of trout phospholipase A 2 465
Fig. 2. SDS-polyacrylamide electrophoresis of microsomal and purified PLA 2 from thermally acclimated rainbow trout. Protein samples from cold-acclimated trout were: A, microsomes; B, Triton extract; C, ammonium sulfate precipitate; and D, G-200 filtrate. Protein samples from warm- acclimated trout were: E, G-200 filtrate; F, ammonium sulfate precipitate; G, Triton extract; and H, microsomes
5 . 6 -
v- 5.4- "1"
~ 5.2- rr"
.A
50 G~ ti3 _J
~ C9 C
4.6
A
SE XOKINASE
LA2
OGLOBIN
I~UMtN
I.() 2'.0 3.0
EB
ALACTOSID ASE
A,CA and WA t BjCA and WA
\=uburm t C~CAand WA BOVINE ALBUMIN �9 ~ . . . .
OVALBUMtN "~ " , , ~ b ~ t O
C~80NIC/hNH YDR~;E
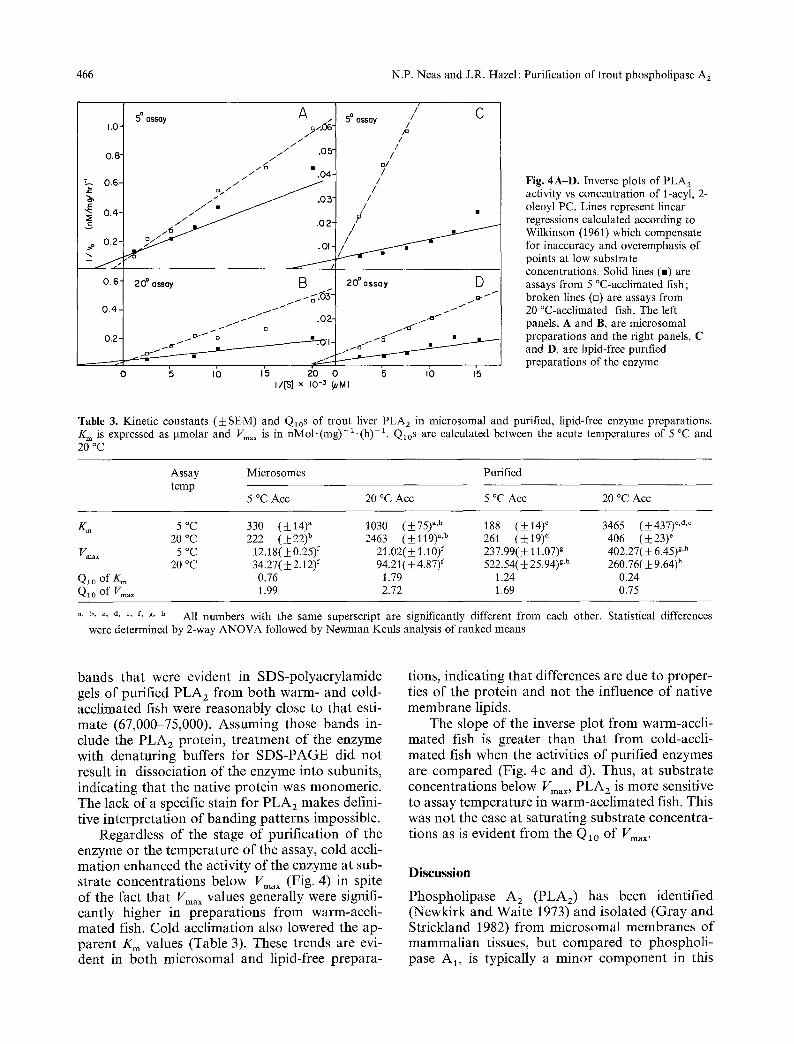
Ve/Vo M~ Fig. 3A, B. Molecular weight estimation of trout microsomal PLA 2 by gel filtration chromatography and SDS-PAGE. Standard proteins were filtered over Sephadex G-200 (A). The void volume (Vo) was determined with blue dextran. Elution volumes (Ve) of catalase and PLA 2 were determined by assaying column fractions for enzymatic activity. V, of other standards were determined by measurements of OD2a o. In B standard proteins were electrophoresed in polyacrylamide gels as described in the text, and stained with ammoniacal silver. Relative mobilities (Mr) were measured for each of the three protein bands in preparations from warm-acclimated (WA) fish and four of the protein bands in preparations from cold-acclimated (CA) fish (designated subunits A, B, C and D), as well as the standards
tration. Following gel filtration, three to four sub- unit bands were still evident and though some qualitative differences in banding patterns were ap- parent between the profiles from warm- and cold- acclimated fish, the three major bands correspond- ing to molecular weights of 67,000-75,000 were identical.
The molecular weight of PLA 2 was estimated by comparison of both the column elution volume (V~) from Sephadex G-200 and the electrophoretic mobility of the presumed PLA 2 band(s) on SDS- PAGE with those of protein standards (Fig. 3). The V e of PLA 2 activity from G-200 corresponded to a molecular weight of 73,000. The three protein
466 N.P. Neas and J.R. Hazel: Purification of trout phospholipase A 2
[ .0-
0.8-
v._. 0.6- "b
0.4- c
0,2- >o
0.6-
0.4--
0.2-
o g 20 o ~ ,b 1'5 tips] x 10-3 ~M)
5 ~ assay A 9/.(~_
f /
/ " ,05- / / "
/ .04- / . ' /
ft.-"
ZO ~ assay B
~ .02-
~ee ~ _ _ _ - - - - - - - - - - - - - ~ I-
IO 15
/ C 5 ~ assay / /
/ /
/ a/ /
/ /
/ /
/ /
20~ assay D
i
Fig. 4A-D. Inverse plots of PLA 2 activity vs concentration of 1-acyl, 2- oleoyl PC. Lines represent linear regressions calculated according to Wilkinson (1961) which compensate for inaccuracy and overemphasis of points at low substrate concentrations. Solid lines (R) are assays from 5 ~ fish; broken lines (n) are assays from 20 ~ fish. The left panels, A and B, are microsomal preparations and the right panels, C and D, are lipid-free purified preparations of the enzyme
Table 3. Kinetic constants ( • and Qlo s of trout liver PLA2 in microsomal and purified, lipid-free enzyme preparations. K m is expressed as gmolar and Vma x is in n M o l . ( m g ) - ~ ' ( h ) 1. Qlos are calculated between the acute temperatures of 5 ~ and 20 ~
Assay Microsomes Purified temp
5 ~ Acc 20 ~ Acc 5 ~ Acc 20 ~ Acc
K~. 5 ~ 330 ( • a 1030 (-k75) a,b 188 (+__14) c 3465 ( • c,a,~ 20 ~ 222 ( • b 2463 ( • ",b 261 ( • d 406 ( + 2 3 ) e
Vm. x 5 ~ 12 .18(_0 .25) f 21 .02( • 1.10) f 237 .99(_ 11.07) g 402 .27( • 6.45) g'h 20 ~ 34 .27(• 2.12) r 94 .21( • f 522 .54(• 25.94) g,h 260 .76(• 9.64) h
Qlo o f K m 0.76 1.79 1.24 0.24 Qlo of Vma x 1.99 2.72 1.69 0.75
a, b, c, a, e, f, g, h All numbers with the same superscript are significantly different from each other. Statistical differences were determined by 2-way A N O V A followed by Newman Keuls analysis of ranked means
bands that were evident in SDS-polyacrylamide gels of purified PLA 2 from both warm- and cold- acclimated fish were reasonably close to that esti- mate (67,000-75,000). Assuming those bands in- clude the PLA 2 protein, treatment of the enzyme with denaturing buffers for SDS-PAGE did not result in dissociation of the enzyme into subunits, indicating that the native protein was monomeric. The lack of a specific stain for PLA 2 makes defini- tive interpretation of banding patterns impossible.
Regardless of the stage of purification of the enzyme or the temperature of the assay, cold accli- mation enhanced the activity of the enzyme at sub- strate concentrations below Vma x (Fig. 4) in spite of the fact that Vma x values generally were signifi- cantly higher in preparations from warm-accli- mated fish. Cold acclimation also lowered the ap- parent Km values (Table 3). These trends are evi- dent in both microsomal and lipid-free prepara-
tions, indicating that differences are due to proper- ties of the protein and not the influence of native membrane lipids.
The slope of the inverse plot from warm-accli- mated fish is greater than that from cold-accli- mated fish when the activities of purified enzymes are compared (Fig. 4c and d). Thus, at substrate concentrations below Vmax, PLA 2 is more sensitive to assay temperature in warm-acclimated fish. This was not the case at saturating substrate concentra- tions as is evident from the Qlo of Vm,x.
Discussion
Phosphol ipase A 2 (PLA2) has been identified (Newkirk and Waite 1973) and isolated (Gray and Strickland 1982) from microsomal membranes of mammalian tissues, but compared to phospholi- pase A1, is typically a minor component in this

N.P, Neas and J.R. Hazel: Purification of trout phospholipase A 2 467
subcellular compartment (van den Bosch 1980). Its occurrence in microsomes often is correlated with a specialized cellular function, such as synthesis of dipalmitoyl lecithin for pulmonary surfactant in lung tissue (Longmore et al. 1979) or prosta- glandin synthesis in brain (Gray and Strickland 1982), or, in the case of rainbow trout, tempera- ture-dependent phospholipid remodeling (Neas and Hazel 1984).
Homogeneous preparations of microsomal PLA 2 have been prepared from bovine brain (Gray and Strickland 1982) by Triton extraction, ammo- nium sulfate precipitation, Sephadex G-200 filtra- tion, ion exchange chromatography and prepara- tive gel electrophoresis. The trout liver enzyme was solubilized by extraction with Triton X-100 (Ta- ble 1) in the same manner as microsomal PLA 2 from bovine brain (Gray and Strickland 1982), but most of the affiliated phosphotipid was not re- moved until the enzyme was precipitated with am- monium sulfate (35-65%; Table 2). Final traces of phospholipid (phosphorus extractable with or- ganic solvents) were removed from the preparation when it was filtered on Sephadex G-200. At this stage, the enzyme was purified about 30-fold rela- tive to crude microsomes, and although not entire- ly homogeneous, as indicated by the appearance of several protein bands following polyacrylamide electrophoresis (Fig. 2), the preparation was con- sidered to be sufficiently purified and free of lipid to warrant preliminary kinetic characterization and an examination of the role of membrane lipids in regulating the activity of the enzyme.
The microsomal form of the trout liver enzyme appeared to form high molecular weight aggre- gates in detergent-free aqueous solution which were too large to be retained by Sephadex G-200. Waite and Sisson (1971) reported similar observa- tions in aqueous solutions of rat mitochondrial PLA 2. Gel filtration in the presence of 0.5 M KC1 did not change the elution behavior of the rat liver enzyme. In detergent solutions, however, trout mi- crosomal PLA 2 was retained by G-200 beyond the void volume of the column (Fig. 1). Membrane- bound proteins have hydrophobic sites where they are in contact with the lipid core of the membrane. These sites are thermodynamically unstable at the water interface in aqueous solutions and tend to seek other areas of hydrophobicity. Thus, hydro- phobic interactions between proteins at these sites are most likely responsible for aggregation. These interactions apparently were disrupted by Triton X-100. The estimated molecular weight of 73,000 is high relative to molecular weights of PLA 2 re- ported from other sources. Bovine brain micro-
soma1 PLA 2 was estimated at 18,500 (Gray and Strickland 1982), rat liver mitochondrial PLA 2 at between 10,000 and 15,000 (de Winter et al. 1982) and snake venom phospholipases A 2 a r e dimers with native molecular weights near 30,000 (Saito and Hanahan 1962). These comparisons imply that trout liver PLA 2 is several times larger than other native enzymes.
It is well documented that the lipid matrix of membrane-bound enzymes can influence catalytic behavior (Razin 1972; Farias etal. 1975). Most studies have focused on the enzymes of membrane transport ( Brasitus et al. 1979; Yuli et al. 1981; Stadtlander et al. 1982), but those involved in me- tabolism of membrane lipids themselves also are subject to modulation (Smith 1983). With regard to temperature adaptation in Tetrahymena, Martin et al. (1976) and Kasai et al. (1976) have proposed that the rigidifying effect of cold temperatures on membrane acyl chains caused the membrane- bound desaturases to be partially exuded from the bilayer, thus exposing the active sites and increas- ing the accessibility of the substrate. Fatty acid desaturation was thus enhanced by the passive ef- fect of temperature on the lipid matrix. Sellner (1981) proposed a similar model to account for perfect compensation in the rates of A 4 desatura- tion in hepatocytes from thermally acclimated trout.
Hazel (1972a) found that succinic dehydroge- nase activity exhibited perfect compensation for temperature in thermally acclimated goldfish, i.e., rates from cold-acclimated fish were comparable to those from warm-acclimated fish when mea- sured at the respective acclimation temperatures, but only when the enzyme was examined in whole mitochondria. Lipid-free preparations exhibited no compensation. Thus, perfect compensation was due to changes in membrane composition, and not qualitative or quantitative changes in the protein itself (Hazel 1972b). Trout liver PLA 2 in particu- late form (microsomes) also exhibited perfect com- pensation for temperature at sub-saturating sub- strate concentrations (Neas and Hazel 1984), but lipid-free preparations retained this characteristic (Fig. 4) implying that factors other than membrane composition are responsible for the adaptation.
Cold acclimation invariably lowered the appar- ent K m (Table 3) of the enzyme in both membrane- bound and soluble forms, accounting for the in- creased specific activity observed for cold-accli- mated fish at nonsaturating substrate concentra- tions (Fig. 4). In contrast, in particulate prepara- tions the maximal velocity was higher in warm- than cold-acclimated fish, whereas perfect compen-

468 N.P. Neas and J.R. Hazel: Purification of trout phospholipase A 2
sation of PLA2 activity was observed for the solu- ble form of the enzyme (5 ~ assayed at 5 ~ Vm, x = 2 3 8 + l l nM/mg.h ; 20 ~ mated assayed at 20 ~ Vm~ x = 261 _+ 10 nM/mg, h; Table 3) due, in large part, to the inverse tempera- ture sensitivity of the reaction in warm-acclimated fish. Thus, while lipid composition may somewhat modify the activity of trout liver PLA2, it is largely qualitative differences in the PLA 2 (evident as sig- nificant differences in K m and Q10 s) at different acclimation temperatures that ensure that phos- pholipid deacylation occurs at low temperatures, most notably the significantly lower K m of the en- zyme from cold-acclimated fish. These qualitative differences may entail changes in primary structure (i.e., isozymes), secondary structure, or tertiary structure of the enzyme protein that affect kinetic properties and occur following prolonged exposure to differing thermal regimes.
Apparent K m values for PLA 2 from cold-accli- mated fish were not significantly affected by assay temperature (i.e., Qao of K m = 1), regardless of the nature of the enzyme preparations. In warm-accli- mated fish, the apparent K m was significantly lower when assays were performed at 5 ~ as opposed to 20 ~ but this decrease was only apparen~ in microsomaI preparations. The opposite was true when the enzyme was assayed in lipid-free form. For the enzyme assays the pH of the TRIS-acetate assay system was adjusted to 8.0 at 20 ~ and, similar to the in vivo situation, the pH of TRIS- acetate varies inversely with temperature. Thus, these shifts in K~ observed in vitro reflect in vivo changes, not an artifact of the assay. A positive correlation between assay temperature and K m was observed in studies of muscle LDH, and has been termed 'positive thermal modulation' (Yancey and Somero 1978). In vivo, the lowering of K m with an acute decrease in temperature serves to conserve the catalytic function of the enzyme, assuming that the substrate concentration is not significantly af- fected. Since it is unlikely that the amount of phos- phatidylcholine changes acutely with temperature, positive thermal modulation may be a factor in maintaining the activity of PLA 2 in warm-accli- mated fish. Due to the absence of the phenomenon in lipid-free preparations, it is probably not an at- tribute of the protein itself, but a function of the intramembrane lipid-protein complex. The oppo- site trend (although not statistically significant) in the pattern of K,, vs assay temperature in micro- somat and soluble preparations of enzyme from cold-acclimated fish is further evidence that ther- mal acclimation effects qualitative changes in trout microsomal PLA 2.
Acknowledgements. This work was supported by National Science Foundation Grant PCM-800-3454 to J.R.H.
References
Bligh EG, Dyer WJ (1959) A rapid method of total lipid extrac- tion and purification. Can J Biochem Physiol 37:911-917
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254
Brasitus TT, Schachter D, Mamouneas TG (1979) Functional interactions of lipids and proteins in rat intestinal microvil- lus membranes. Biochemistry 18:4136~144
Conner AM, Brimble PD, Choy PC (1981) A simple method for the preparation of phosphatidylcholine labelled at the 2-acyl position. Prep Biochem 11:91-97
Cossins AR (1981) The adaptation of membrane dynamic struc- ture to temperature. In: Morris GJ, Clarke A (eds) Effects of low temperatures on biological membranes. Academic Press, New York, pp 83-106
de Winter JM, Vianen GM, van den Bosch H (1982) Purifica- tion of rat liver mitochondrial phospholipase A 2. Biochim Biophys Acta 712:332-341
Dickens BF, Thompson GA Jr (1981) Rapid membrane re- sponse during low-temperature acclimation. Correlation of early changes in the physical properties and lipid composi- tion of Tetrahymena microsomal membranes. Biochim Bio- phys Acta 664:211-218
Dickens BF, Thompson GA Jr (1982) Phospholipid molecular species akerations in microsomat membranes as an initiaI key step during cellutar acclimation to low temperatures. Biochemistry 21 : 3604-36I 1
Farias RN, Bloj B, Morero RD, Sineriz F, Trucco RE (1975) Regulation of allosteric membrane-bound enzymes through changes in membrane lipid composition. Biochim Biophys Acta 415:231-250
Fukushima H, Martin CE, Iida H, Kitajima Y, Thompson GA Jr, Nozawa Y (1976) Changes in membrane lipid composi- tion during temperature adaptation by a thermotolerant strain of Tetrahymena pyriformis. Biochim Biophys Acta 43] : 165-179
Gray NCC, Strickland KP (1982) The purification and charac- terization of a phospholipase A 2 activity from the 106,000 x g pellet (microsomal) fraction of bovine brain act- ing on phosphatidylinositol. Can J Biochem 60:108-117
Hazel JR (1972a) The effect of temperature acclimation upon succinic dehydrogenase activity from the epaxial muscle of the common goldfish (Carassius auratus L.) - I. Properties of the enzyme and the effect of lipid extraction. Comp Bio- chem Physiol 43B: 837-861
Hazel JR (1972b) The effect of temperature acclimation upon succinic dehydrogenase activity from the epaxial muscle of the common goldfish (Carassius auratus L.) - II. Lipid reac- tivation of the soluble enzyme. Comp Biochem Physiol 43B : 863-882
Hazel JR (1979) Influence of thermal acclimation on membrane lipid composition of rainbow trout liver. Am J Physiol 236: R9~-R101
Hazel JR, Neas NP (1982) Turnover of the fatty acyl and gly- cerol moieties of microsomal membrane Iipids from liver, gill and muscle tissue of thermally acclimated rainbow trout, Salmo gazrdneri. J Comp Physiol 149:11 18
Hazel JR, Sellner PA, Hagar AF, Neas NP (1983) Thermal adaptation in biological membranes: The acylation of sn- glycerol-3-phosphate by liver microsomes of thermally accli- mated trout (Salmo gairdnerO. Molec Physiol 4:125-140

N.P. Neas and J.R. Hazel : Purification of trout phospholipase A 2 469
Holub BJ, Kuksis A (1978) Metabolism of molecular species of diacylglycerophospholipids. Adv Lipid Res 16:1-125
Holub BJ, Piekarski J, Cho CY, Slinger SJ (1976) Incorporation of fatty acids into phosphatidylcholine by acyl-CoA : l-acyl- sn-glycero-3-phosphorytcholine acyltransferase in liver of rainbow trout, Salmo gairdneri. J Fish Res Board Can 33:2821 2826
Jonas REE, Bilinski E (1967) Phospholipase A activity in rain- bow trout muscle. J Fish Res Board Can 24:2555-2562
Kasai R, Kitajima Y, Martin CE, Nozawa Y, Skriver L, Thompson GA Jr (1976) Molecular control of membrane properties during temperature acclimation. Membrane flui- dity regulation of fatty acid desaturase action? Biochemistry 15 : 5228-5233
Laemmli UK (1970) Cleavage of structural proteins during as- sembly of the head of bacteriophage T4. Nature 227: 680-685
Longmore WJ, Oldenborg V, van Golde LMG (1979) Phospho- l i pa se A 2 in rat-lung microsomes: Substrate specificity to- wards endogenous phosphatidylcholines. Biochim Biophys Acta 572:452-460
Lfick H (1963) Catalase. In: Bergmeyer HU (ed) Methods in enzymatic analysis. Academic Press, New York, pp 885-894
Martin CE, Hiramitsu K, Kitajima Y, Nozawa Y, Skriver L, Thompson GA Jr (1976) Molecular control of membrane properties during temperature acclimation. Fatty acid desa- turase regulation of membrane fluidity in acclimating Tetra- hymena cells. Biochemistry 15 : 5218-5227
Neas NP, Hazel JR (1984) Phospholipase A 2 from liver micro- somal membranes of thermally acclimated rainbow trout. J Exp Zool 233 : 51-60
Newkirk JD, Waite M (1973) Phospholipid hydrolysis by phos- photipases A 1 and A 2 in plasma membranes and micro- somes of rat liver. Biochim Biophys Acta 298 : 562-576
Ramesha CS, Thompson GA Jr (1982) Changes in the lipid composition and physical properties of Tetrahymena ciliary membranes following low-temperature acclimation. Bio- chemistry 21 : 3612-3617
Razin S (1972) Reconstitution of biological membranes. Bio- chim Biophys Acta 265:241-296
Rouser G, Fleischer S, Yamamoto A (1970) Two dimensional thin layer chromatographic separation of polar lipids and determination of phospholipids by phosphorus analysis of spots. Lipids 5: 494-496
Saito K, Hanahan DJ (1962) A study of the purification and properties of the phospholipase A of Crotalus adamanteus venom. Biochemistry 1 : 521-532
Sellner PA (1981) Unsaturated fatty acid metabolism in hepato- cytes from thermaIly-acclimated rainbow trout. PhD disser- tation, Arizona State University, Tempe
Sellner PA, Hazel JR (1982) Desaturation and elongation of unsaturated fatty acids in hepatocytes from thermally accli- mated rainbow trout. Arch Biochem Biophys 213:58-66
Smith JD (1983) Effect of modification of membrane phospho- lipid composition on the activity of phosphatidylethanol- amine N-methyltransferase of Tetrahymena. Arch Biochem Biophys 223 : 193-201
Stadtlander K, Rade S, Ahlers J (1982) Influence of growth conditions on the composition of the plasma membrane from yeast and on kinetic properties of two membrane func- tions. J Cell Biochem 20:369-380
van den Bosch H (1974) Phosphoglyceride metabolism. Annu Rev Biochem 43 : 243-277
van den Bosch H (1980) Intracellular phospholipases A. Bio- chim Biophys Acta 604:191-246
Waite M, Sisson P (1971) Partial purification and characteriza- tion of the phospholipase A 2 from rat liver mitochondria. Biochemistry 10 : 237%2383
Wilkinson GN (i 961) Statistical estimations in enzyme kinetics. Biochem J 80 : 324-332
Wray W, Boulikas T, Wray VP, Hancock R (1981) Silver stain- ing of proteins in polyacrylamide gels. Anal Biochem 118:197-203
Yancey PH, Somero GN (1978) Temperature dependence of intracellular pH; its role in the conservation of pyruvate apparent K m values of lactate dehydrogenases. J Comp Physiol 125 : 129-134
Yuli I, Wilbrandt W, Shinitzky M (198I) Glucose transport through cell membranes of modified lipid fluidity. Biochem- istry 20: 4230M256