p2x3-mediated peripheral sensitization of neuropathic pain in resiniferatoxin-induced neuropathy
TRANSCRIPT

Experimental Neurology 235 (2012) 316–325
Contents lists available at SciVerse ScienceDirect
Experimental Neurology
j ourna l homepage: www.e lsev ie r .com/ locate /yexnr
P2X3-mediated peripheral sensitization of neuropathic pain inresiniferatoxin-induced neuropathy
Yu-Lin Hsieh a,b, Hao Chiang a, June-Horng Lue a,⁎, Sung-Tsang Hsieh a,c,d,⁎⁎a Department of Anatomy and Cell Biology, College of Medicine, National Taiwan University, Taipei 10051, Taiwanb Department of Anatomy, School of Medicine, College of Medicine, Kaohsiung Medical University, Kaohsiung 80708, Taiwanc Department of Neurology, National Taiwan University Hospital, Taipei 10002, Taiwand Graduate Institute of Brain and Mind Science, College of Medicine, National Taiwan University, Taipei 10051, Taiwan
Abbreviations: ATF3, Activating transcription factrelated peptide; DRG, Dorsal root ganglion; RTX, Resinifceptor potential vanilloid subtype 1.⁎ Correspondence to: J.-H. Lue, Department of Anatom
Medicine, National Taiwan University, Rm. 609, 1 Jen-Taiwan. Fax: +886 2 23915292.⁎⁎ Correspondence to: S.-T. Hsieh, Department of AnatoMedicine, National Taiwan University, Rm. 638, 1 Jen-Taiwan. Fax: +886 2 23915292.
E-mail addresses: [email protected] (J.-H. Lue), sh
0014-4886/$ – see front matter. Crown Copyright © 20doi:10.1016/j.expneurol.2012.02.013
a b s t r a c t
a r t i c l e i n f oArticle history:Received 17 December 2011Revised 10 February 2012Accepted 20 February 2012Available online 28 February 2012
Keywords:Purinergic receptorP2X3Mechanical allodyniaPeripheral sensitizationSmall-fiber neuropathyResiniferatoxinCalcitonin gene-related peptideActivating transcription factor-3
Patients suffering from sensory neuropathy due to skin denervation frequently have paradoxical manifes-tations of reduced nociception and neuropathic pain. However, there is a lack of satisfactory animal modelsto investigate these phenomena and underlying mechanisms. We developed a mouse system of neuropa-thy induced by resiniferatoxin (RTX), a capsaicin analog, and examined the functional significance ofP2X3 receptor in neuropathic pain. From day 7 of RTX neuropathy, mice displayed mechanical allodynia(pb0.0001) and thermal hypoalgesia (pb0.0001). After RTX treatment, dorsal root ganglion (DRG) neuronsof the peripherin type were depleted (p=0.012), while neurofilament (+) DRG neurons were not affected(p=0.62). In addition, RTX caused a shift in neuronal profiles of DRG: (1) increased in P2X3 receptor(p=0.0002) and ATF3 (p=0.0006) but (2) reduced TRPV1 (p=0.036) and CGRP (p=0.015). The numberof P2X3(+)/ATF3(+) neurons was linearly correlated with mechanical thresholds (p=0.0017). Theperipheral expression of P2X3 receptor in dermal nerves was accordingly increased (p=0.016), and anintraplantar injection of the P2X3 antagonists, A-317491 and TNP–ATP, relieved mechanical allodynia ina dose-dependent manner. In conclusion, RTX-induced sensory neuropathy with upregulation of P2X3 re-ceptor for peripheral sensitization of mechanical allodynia, which provides a new therapeutic target forneuropathic pain after skin denervation.
Crown Copyright © 2012 Published by Elsevier Inc. All rights reserved.
Introduction
Neuropathic pain develops after peripheral nerve degeneration,which presumably reduces nociceptive input. In human peripheralnerve diseases affecting small-diameter nociceptive nerves, some pa-tients could have paradoxical symptoms: neuropathic pain but withreduced sensitivities to noxious stimuli due to skin denervation(Baron et al., 2009; Cheng et al., 2009; Obrosova, 2009; Polydefkiset al., 2004). Similar phenomena are observed after capsaicin-induced skin denervation (Gibbons et al., 2010; Rage et al., 2010). Pe-ripheral sensory nerves are axonal extensions of dorsal root ganglion(DRG) neurons which consist of large-diameter and small-diameter
or-3; CGRP, Calcitonin gene-eratoxin; TRPV1, Transient re-
y and Cell Biology, College ofRi Road, Sec. 1, Taipei 10051,
my and Cell Biology, College ofRi Road, Sec. 1, Taipei 10051,
[email protected] (S.-T. Hsieh).
12 Published by Elsevier Inc. All righ
neurons with corresponding sizes of nerve fibers. It is, however, notclear whether pure injury to small neurons could result in such a par-adoxical combination of symptoms: reduced nociception and neuro-pathic pain. There is also a lack of experimental systems to test thishypothesis. We previously established a neuropathy system inducedby resiniferatoxin (RTX), a capsaicin analog. Skin denervation withdegeneration of unmyelinated nerves is the predominant feature ofRTX-induced neuropathy (Hsieh et al., 2008), which can serve as aprototype of pure small-fiber neuropathy and provide a good oppor-tunity to examine underlying molecular mechanisms of clinical pre-sentations: neuropathic pain and reduced nociception.
Nerve injury causes a cascade of responses in neuronal cell bodies,for example, the upregulation of transcription factors, which leads tothe generation of effector molecules responsible for maladaptive be-haviors of neuropathic pain. Activating transcription factor-3 (ATF3)is a member of the ATF/CREB transcription factor superfamily (Haiand Hartman, 2001) and upregulated in DRG neurons after nerve in-jury, for example, spinal nerve ligation (Fukuoka et al., 2012). Thesefindings raise the possibility of exploring whether or not ATF3 isupregulated in RTX-induced neuropathy, which only selectively de-pletes cutaneous nerves (Avelino and Cruz, 2000; Hsieh et al., 2008;Neubert et al., 2003). If so, will the pattern of ATF3 expression in
ts reserved.

317Y.-L. Hsieh et al. / Experimental Neurology 235 (2012) 316–325
DRG neurons parallel the behavioral manifestations and what are thephenotypes of ATF3(+) neurons?
Puringeric receptors, such as P2X3, play important roles in the de-velopment of neuropathic allodynia (Burnstock, 2009; Kaan et al.,2010). Previous studies have documented upregulation of P2X3 re-ceptor in peripheral neuropathic pain, for example, mechanicalinjury-induced focal neuropathy (Tsuchihara et al., 2009) and chronicvisceral pain model (Banerjee et al., 2009; Xu et al., 2008). It is, how-ever, not clear the phenotypes of P2X3(+) neurons in neuropathicpain, i.e. whether these are injured vs. uninjured neurons or largevs. small-diameter DRG neurons. Thus, P2X3 can potentially providea link between nerve injury and neuropathic pain. However, therole of P2X3 in RTX-induced neuropathy and the contribution ofP2X3 to the pathology and underlying molecular mechanisms of pe-ripheral sensitization have not been systematically explored.
To address the above issues, we used RTX-induced neuropathy toinvestigate (1) neuropathic pain behaviors and their relation to ATF3,(2) the profiles of DRG neurons with induced ATF3 expression, and(3) the pharmacological effects of P2X3 antagonism on RTX-inducedneuropathic pain behaviors.
Materials and methods
Systemic RTX treatment
Experiments were performed on 8-week-old adult male ICR mice(35–40 g). RTX (Sigma, St. Louis, MO) was dissolved in the vehicle(10% Tween 80 and 10% ethanol in normal saline). Mice in the RTXgroup received a single dose of RTX (50 μg/kg) by an intraperitonealinjection (Hsieh et al., 2008). The other group received an equal vol-ume of the vehicle (the vehicle group) as the control. Mice werehoused in plastic cages on a 12-h light/12-h dark cycle and wereallowed access water and food ad libitum. All procedures were con-ducted in accordance with ethical guidelines for laboratory animals(Zimmermann, 1983) and the protocol was approved by the AnimalCommittee of National Taiwan University College of Medicine, Taipei,Taiwan and Kaohsiung Medical University, Kaohsiung, Taiwan.
Animal behavior evaluation
The behavior evaluation included thermal (hot-plate test) andmechanical (von Frey filament test) responses. Tests were performedbefore RTX injection (D0) and on day 7 (D7) after RTX treatment, andthen weekly until D84, at the end of the experiment.
Hot-plate testMice were placed on a 52 °C hot plate (IITC, Woodland Hills, CA),
enclosed in a Plexiglas cage. The withdrawal latencies of the hindpawto thermal stimulations were determined to an accuracy of 0.1 s. Eachtest session consisted of three trials separated by 30-min intervals.The criteria of withdrawal included shaking, licking, or jumping onthe hot plate. The mean latency was expressed as the threshold ofan individual animal to the thermal stimulation.
von Frey filament testMechanical thresholds of the hindpaw were assessed using the
up-and-down method with different calibers of von Frey monofila-ments (Somedic Sales AB, Hörby, Sweden). Briefly, a series of mono-filaments was applied to the plantar region of the hindpaw. If pawwithdrawal occurred, a monofilament of a smaller caliber was ap-plied. In the absence of paw withdrawal, a monofilament of a largercaliber was then used. Four additional stimuli with monofilamentsof different calibers based on the preceding responses were applied.The mechanical thresholds were calculated according to a publishedformula (Chaplan et al., 1994).
Double-labeling immunofluorescent staining of DRG and dermal sheets
In double-labeling experiments, the well-established tyramidesignal amplification (TSA) technique was used for primary antiseraraised in the same species (Lin et al., 2008). Sections were sequential-ly incubated with one primary antiserum, a biotinylated-labeled sec-ondary antibody, and streptavidin-horseradish peroxidase (HRP)(1:100, PerkinElmer) for 30 min. Signals were amplified with thefluorescein tyramide reagent (1:50, PerkinElmer) for 3 min. Afterrinsing in 0.5 M Tris buffer (Tris), sections were incubated with thesecond primary antiserum, followed by a Texas red-conjugated sec-ondary antibody for 1 h (1:100, Jackson ImmunoResearch, WestGrove, PA). The concentration of the first primary antiserum wasmuch lower than that for regular immunostaining (1:6000), whichwas beyond the detection limit of conventional immunofluorescence.For the two primary antisera raised in different species, conventionalimmunofluorescent staining was performed, i.e., incubation withTexas red and fluorescein isothiocyanate (FITC)-conjugated second-ary antisera (1:100, Jackson ImmunoResearch) corresponding to ap-propriate primary antisera for 1 h. Sections were mounted usingVectashield (Vector, Burlingame, CA). Cryostat sections of 8-μm inthickness from the lumbar DRGs were used in this study. For system-atic sampling, two ganglia (L4 and L5) per mice were collected forsectioning. Every section of each ganglion with an interval of 80 μmwas immunostained and quantified. The primary antisera includedATF3 (rabbit, 1:6000, Santa Cruz Biotechnology, Santa Cruz, CA), per-ipherin (rabbit, 1:800, Chemicon, Temecula, CA), neurofilamentSMI32 (mouse, 1:600, Covance, Emeryville, CA), P2X3 (rabbit, 1:400,Neuromics, Edina, MN), calcitonin gene-related peptide, (CGRP, rab-bit, 1:800, Sigma), and transient receptor potential vanilloid subtype1 (TRPV1, goat, 1:100, Santa Cruz Biotechnology).
Dermal nerves of different phenotypesWe also investigated the changes in dermal nerves of different
phenotypes. Briefly, the plantar skin of the hindpaw was removedand incubated in an EDTA solution at 37 °C for 30 min. The dermiswas separated from the epidermis and subcutaneous tissues weretrimmed (Tschachler et al., 2004). Dermal sheets were immersed in4% paraformaldehyde overnight and then incubated with differentprimary antisera following the same immunofluorescence proceduresdescribed above.
Quantitation of DRG neurons and dermal nerves of different phenotypes
For DRG neuron quantitation, each DRG section was photo-graphed at 200× under a fluorescence microscope (Axiophot micro-scope, Carl Zeiss, Heidelberg, Germany) in a systematic fashion toproduce a montage of the entire DRG section following establishedprocedures (Hsieh et al., 2008). To avoid density bias, only thearea containing neurons was measured and only neurons with aclear nuclear profile were counted. The diameter of each P2X3(+)DRG neuron was measured with Image J vers. 1.44d software (Na-tional Institutes of Health, Bethesda, MD). The density wasexpressed as neurons/mm2 and a histogram of the diameter ofP2X3(+) DRG neurons was plotted. The quantitation of dermalnerve fibers followed our established protocols (Lin et al., 2008).For each dermal sheet, photographs were taken at 200× under anAxiophot fluorescence microscope (Carl Zeiss) from 10 randomly se-lected areas, covering 30%–40% of the entire dermal sheet. The in-tensities of the immunofluorescence signals were subtracted fromthe background, and each pixel (intensity signal) was equivalentto 0.1445 μm2. The areas of each dermal nerve of different pheno-types were divided by the selected area of the dermis. The dermalnerve density was expressed as μm2/mm2.
318 Y.-L. Hsieh et al. / Experimental Neurology 235 (2012) 316–325
Pharmacological intervention with P2X3 by an intraplantar injection
Pharmacological experiments were performed with single dose ofP2X3 antagonism on D7 after RTX treatment. Two antagonists wereused: (Attal et al., 2010) A-317491 (Sigma), a P2X3-specific antago-nist, at concentrations of 2, 20, and 200 μg/paw and (Avelino andCruz, 2000) TNP–ATP (Sigma), a P2X1-4 antagonist, at concentrationsof 16, 160, and 240 μg/paw (Oliveira et al., 2009). Drugs were dis-solved in normal saline and administrated by an intraplantar injec-tion at one hindpaw (10 μl/paw). The other group of mice receivednormal saline as the control for comparison (the saline group).Changes in mechanical thresholds were assessed at 0.5, 1, 2, 4, and6 h after the intraplantar injection.
Experimental design and statistical analysis
In the set-up of RTX-induced neuropathy, there were two groups:the RTX group and the vehicle group. The pharmacological interven-tion study of P2X3 antagonist, A-317491, consisted of the experimen-tal group and the saline group. The coding information was maskedduring the behavioral tests and the quantification procedures. Therewere 180 mice in total with 5–8 mice in each group at differenttime points. All data are expressed as the mean±standard derivationof the mean and t test was performed for data with a Gaussian dis-tribution. For data which did not follow a Gaussian distribution, anonparametric Mann–Whitney test was conducted. pb0.05 wasconsidered statistically significant.
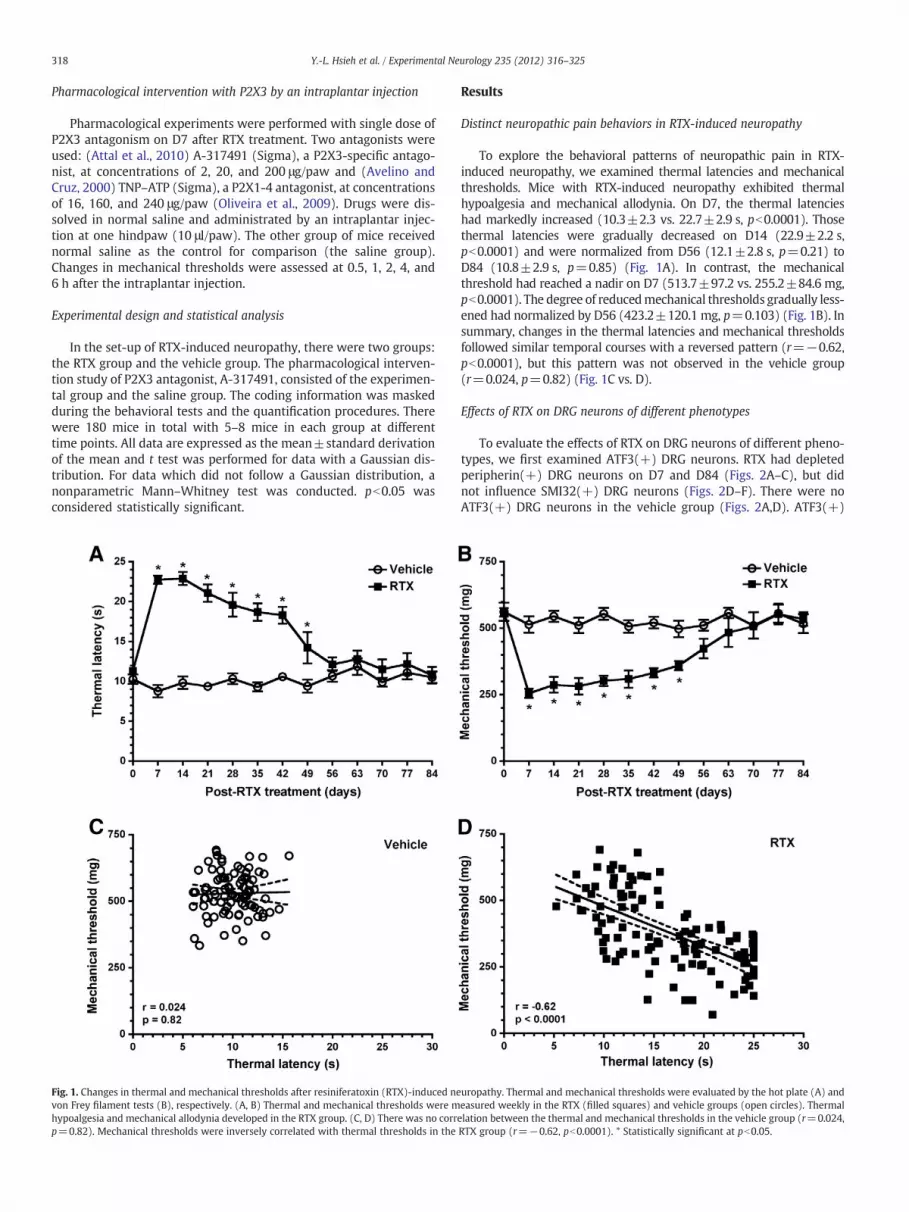
Fig. 1. Changes in thermal and mechanical thresholds after resiniferatoxin (RTX)-induced nevon Frey filament tests (B), respectively. (A, B) Thermal and mechanical thresholds were mhypoalgesia and mechanical allodynia developed in the RTX group. (C, D) There was no corrp=0.82). Mechanical thresholds were inversely correlated with thermal thresholds in the
Results
Distinct neuropathic pain behaviors in RTX-induced neuropathy
To explore the behavioral patterns of neuropathic pain in RTX-induced neuropathy, we examined thermal latencies and mechanicalthresholds. Mice with RTX-induced neuropathy exhibited thermalhypoalgesia and mechanical allodynia. On D7, the thermal latencieshad markedly increased (10.3±2.3 vs. 22.7±2.9 s, pb0.0001). Thosethermal latencies were gradually decreased on D14 (22.9±2.2 s,pb0.0001) and were normalized from D56 (12.1±2.8 s, p=0.21) toD84 (10.8±2.9 s, p=0.85) (Fig. 1A). In contrast, the mechanicalthreshold had reached a nadir on D7 (513.7±97.2 vs. 255.2±84.6 mg,pb0.0001). The degree of reducedmechanical thresholds gradually less-ened had normalized by D56 (423.2±120.1 mg, p=0.103) (Fig. 1B). Insummary, changes in the thermal latencies and mechanical thresholdsfollowed similar temporal courses with a reversed pattern (r=−0.62,pb0.0001), but this pattern was not observed in the vehicle group(r=0.024, p=0.82) (Fig. 1C vs. D).
Effects of RTX on DRG neurons of different phenotypes
To evaluate the effects of RTX on DRG neurons of different pheno-types, we first examined ATF3(+) DRG neurons. RTX had depletedperipherin(+) DRG neurons on D7 and D84 (Figs. 2A–C), but didnot influence SMI32(+) DRG neurons (Figs. 2D–F). There were noATF3(+) DRG neurons in the vehicle group (Figs. 2A,D). ATF3(+)
uropathy. Thermal and mechanical thresholds were evaluated by the hot plate (A) andeasured weekly in the RTX (filled squares) and vehicle groups (open circles). Thermalelation between the thermal and mechanical thresholds in the vehicle group (r=0.024,RTX group (r=−0.62, pb0.0001). * Statistically significant at pb0.05.
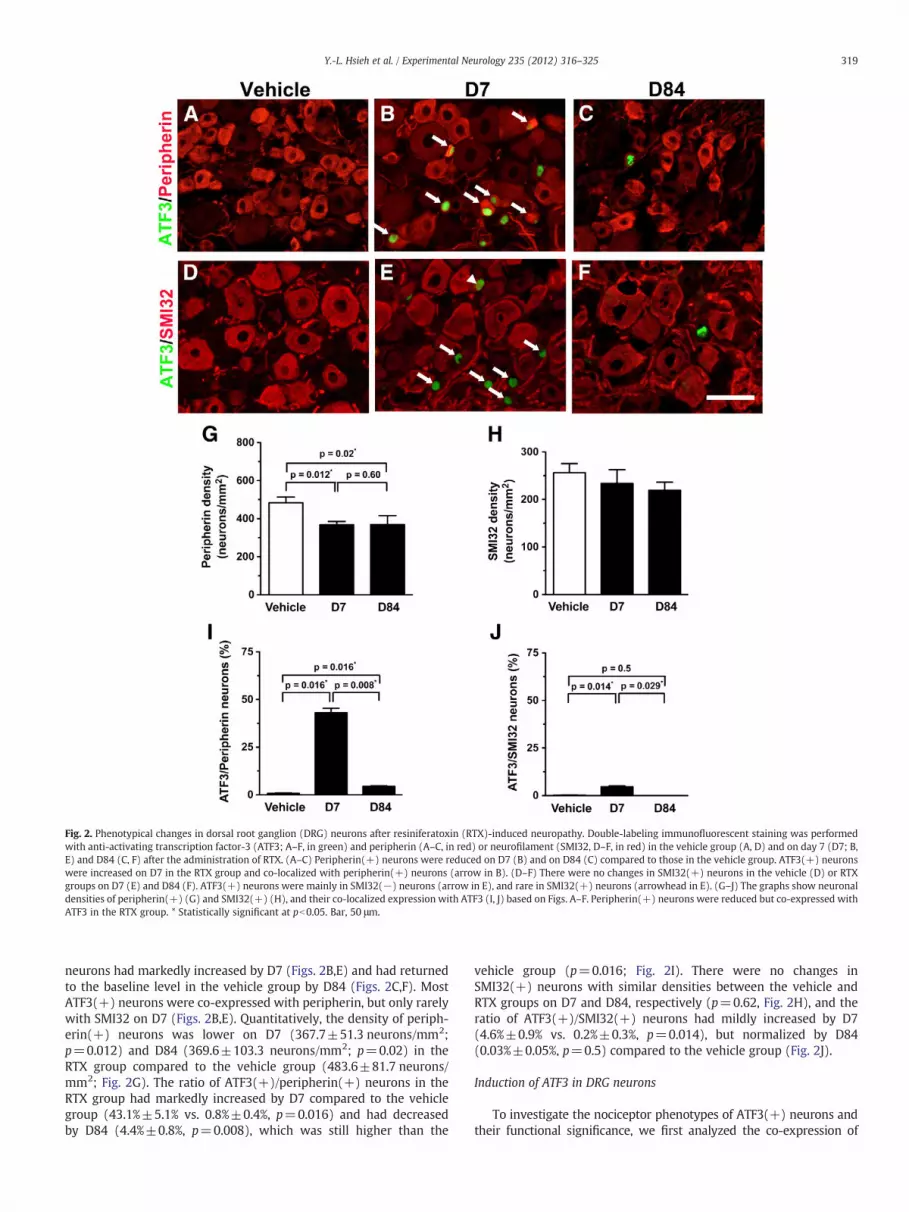
Fig. 2. Phenotypical changes in dorsal root ganglion (DRG) neurons after resiniferatoxin (RTX)-induced neuropathy. Double-labeling immunofluorescent staining was performedwith anti-activating transcription factor-3 (ATF3; A–F, in green) and peripherin (A–C, in red) or neurofilament (SMI32, D–F, in red) in the vehicle group (A, D) and on day 7 (D7; B,E) and D84 (C, F) after the administration of RTX. (A–C) Peripherin(+) neurons were reduced on D7 (B) and on D84 (C) compared to those in the vehicle group. ATF3(+) neuronswere increased on D7 in the RTX group and co-localized with peripherin(+) neurons (arrow in B). (D–F) There were no changes in SMI32(+) neurons in the vehicle (D) or RTXgroups on D7 (E) and D84 (F). ATF3(+) neurons were mainly in SMI32(−) neurons (arrow in E), and rare in SMI32(+) neurons (arrowhead in E). (G–J) The graphs show neuronaldensities of peripherin(+) (G) and SMI32(+) (H), and their co-localized expression with ATF3 (I, J) based on Figs. A–F. Peripherin(+) neurons were reduced but co-expressed withATF3 in the RTX group. * Statistically significant at pb0.05. Bar, 50 μm.
319Y.-L. Hsieh et al. / Experimental Neurology 235 (2012) 316–325
neurons had markedly increased by D7 (Figs. 2B,E) and had returnedto the baseline level in the vehicle group by D84 (Figs. 2C,F). MostATF3(+) neurons were co-expressed with peripherin, but only rarelywith SMI32 on D7 (Figs. 2B,E). Quantitatively, the density of periph-erin(+) neurons was lower on D7 (367.7±51.3 neurons/mm2;p=0.012) and D84 (369.6±103.3 neurons/mm2; p=0.02) in theRTX group compared to the vehicle group (483.6±81.7 neurons/mm2; Fig. 2G). The ratio of ATF3(+)/peripherin(+) neurons in theRTX group had markedly increased by D7 compared to the vehiclegroup (43.1%±5.1% vs. 0.8%±0.4%, p=0.016) and had decreasedby D84 (4.4%±0.8%, p=0.008), which was still higher than the
vehicle group (p=0.016; Fig. 2I). There were no changes inSMI32(+) neurons with similar densities between the vehicle andRTX groups on D7 and D84, respectively (p=0.62, Fig. 2H), and theratio of ATF3(+)/SMI32(+) neurons had mildly increased by D7(4.6%±0.9% vs. 0.2%±0.3%, p=0.014), but normalized by D84(0.03%±0.05%, p=0.5) compared to the vehicle group (Fig. 2J).
Induction of ATF3 in DRG neurons
To investigate the nociceptor phenotypes of ATF3(+) neurons andtheir functional significance, we first analyzed the co-expression of

320 Y.-L. Hsieh et al. / Experimental Neurology 235 (2012) 316–325
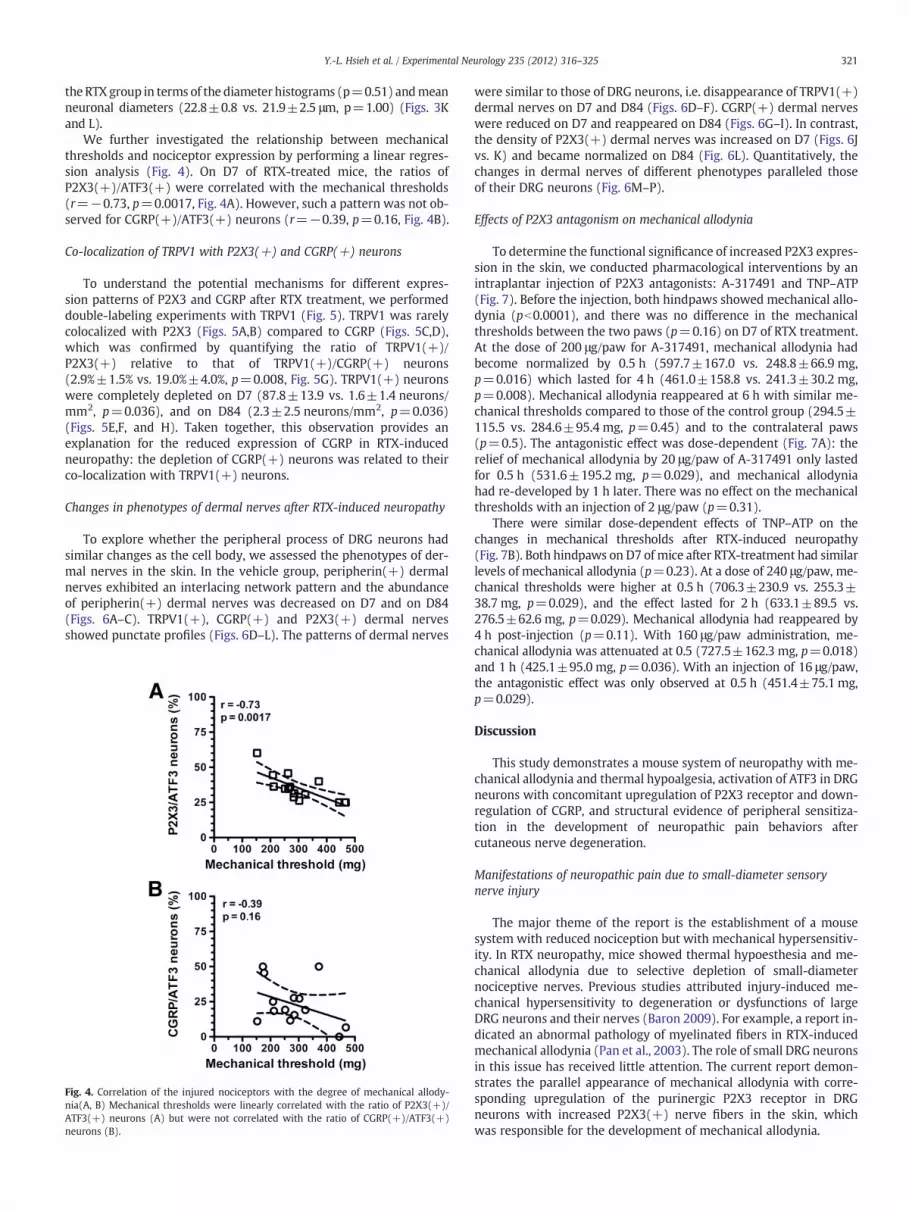
P2X3 and CGRP with ATF3 (Fig. 3). On D7, RTX had induced P2X3expression (234.2±36.4 vs. 125.5±13.4 neurons/mm2; p=0.0002),and the expression in the RTX group had become comparable to the ve-hicle group by D84 (134.6±29.3 neurons/mm2, p=0.27) (Figs. 3A–Cand G). In contrast, the expression of CGRP was lower on D7 (206.7±19.6 vs. 165.8±32.8 neurons/mm2; p=0.015) and had returned tothe baseline by D84 (206.4±32.6 neurons/mm2, p=0.98) (Figs. 3D–F
Fig. 3. Changes in the expression of different nociceptors after resiniferatoxin (RTX)-inducesera against activating transcription factor-3 (ATF3; A–F, in green) and P2X3 (A–C, in red) orday 7 (D7; B, E) and D84 (C, F) after RTX administration. (A–C) There were more P2X3(+)were comparable to those of the vehicle group. ATF3(+) neurons were higher on D7, and soD7 of RTX-induced neuropathy compared to the vehicle group. On D84, the number of CGRneurons also expressed CGRP (arrowhead in E). (G–J) The graphs show neuronal densitiescomparison of co-expression ratio with ATF3(+) neurons. (K, L) The graphs show the mmean neuronal diameter (L). * Statistically significant at pb0.05. Bar, 50 μm.
and H). ATF3(+) neurons had increased by D7 (157.4±54.3 vs. 4.1±1.3 neurons/mm2, p=0.0006), but the density had returned to thesame level as the vehicle group by D84 (5.2±1.7 neurons/mm2,p=0.21) (Fig. 3I). The ratio of P2X3(+)/ATF3(+) neurons was higherthan that of CGRP(+)/ATF3(+) neurons (36.5%±10.3% vs. 19.4%±6.2%, p=0.0002) (Fig. 3J). The morphometric analyses indicated thatP2X3(+) DRG neurons were similar in size between the vehicle and
d neuropathy. Double-labeling immunofluorescent staining was performed with anti-calcitonin gene-related peptide (CGRP; D–F, in red) in the vehicle group (A, D), and onneurons on D7 in the RTX group than in the vehicle group. On D84, P2X3(+) neuronsme were co-expressed with P2X3 (arrow in B). (D–F) CGRP(+) neurons were lower onP(+) neurons was similar to that in the vehicle group. A small proportion of ATF3(+)of P2X3(+) (G), CGRP(+) (H) and ATF3(+) (I) according to the Figs. 3A–F, and (J) aorphometry analyses of P2X3(+) neurons including the diameter histogram (K) and
321Y.-L. Hsieh et al. / Experimental Neurology 235 (2012) 316–325
the RTX group in terms of the diameter histograms (p=0.51) andmeanneuronal diameters (22.8±0.8 vs. 21.9±2.5 μm, p=1.00) (Figs. 3Kand L).
We further investigated the relationship between mechanicalthresholds and nociceptor expression by performing a linear regres-sion analysis (Fig. 4). On D7 of RTX-treated mice, the ratios ofP2X3(+)/ATF3(+) were correlated with the mechanical thresholds(r=−0.73, p=0.0017, Fig. 4A). However, such a pattern was not ob-served for CGRP(+)/ATF3(+) neurons (r=−0.39, p=0.16, Fig. 4B).
Co-localization of TRPV1 with P2X3(+) and CGRP(+) neurons
To understand the potential mechanisms for different expres-sion patterns of P2X3 and CGRP after RTX treatment, we performeddouble-labeling experiments with TRPV1 (Fig. 5). TRPV1 was rarelycolocalized with P2X3 (Figs. 5A,B) compared to CGRP (Figs. 5C,D),which was confirmed by quantifying the ratio of TRPV1(+)/P2X3(+) relative to that of TRPV1(+)/CGRP(+) neurons(2.9%±1.5% vs. 19.0%±4.0%, p=0.008, Fig. 5G). TRPV1(+) neuronswere completely depleted on D7 (87.8±13.9 vs. 1.6±1.4 neurons/mm2, p=0.036), and on D84 (2.3±2.5 neurons/mm2, p=0.036)(Figs. 5E,F, and H). Taken together, this observation provides anexplanation for the reduced expression of CGRP in RTX-inducedneuropathy: the depletion of CGRP(+) neurons was related to theirco-localization with TRPV1(+) neurons.
Changes in phenotypes of dermal nerves after RTX-induced neuropathy
To explore whether the peripheral process of DRG neurons hadsimilar changes as the cell body, we assessed the phenotypes of der-mal nerves in the skin. In the vehicle group, peripherin(+) dermalnerves exhibited an interlacing network pattern and the abundanceof peripherin(+) dermal nerves was decreased on D7 and on D84(Figs. 6A–C). TRPV1(+), CGRP(+) and P2X3(+) dermal nervesshowed punctate profiles (Figs. 6D–L). The patterns of dermal nerves
Fig. 4. Correlation of the injured nociceptors with the degree of mechanical allody-nia(A, B) Mechanical thresholds were linearly correlated with the ratio of P2X3(+)/ATF3(+) neurons (A) but were not correlated with the ratio of CGRP(+)/ATF3(+)neurons (B).
were similar to those of DRG neurons, i.e. disappearance of TRPV1(+)dermal nerves on D7 and D84 (Figs. 6D–F). CGRP(+) dermal nerveswere reduced on D7 and reappeared on D84 (Figs. 6G–I). In contrast,the density of P2X3(+) dermal nerves was increased on D7 (Figs. 6Jvs. K) and became normalized on D84 (Fig. 6L). Quantitatively, thechanges in dermal nerves of different phenotypes paralleled thoseof their DRG neurons (Fig. 6M–P).
Effects of P2X3 antagonism on mechanical allodynia
To determine the functional significance of increased P2X3 expres-sion in the skin, we conducted pharmacological interventions by anintraplantar injection of P2X3 antagonists: A-317491 and TNP–ATP(Fig. 7). Before the injection, both hindpaws showed mechanical allo-dynia (pb0.0001), and there was no difference in the mechanicalthresholds between the two paws (p=0.16) on D7 of RTX treatment.At the dose of 200 μg/paw for A-317491, mechanical allodynia hadbecome normalized by 0.5 h (597.7±167.0 vs. 248.8±66.9 mg,p=0.016) which lasted for 4 h (461.0±158.8 vs. 241.3±30.2 mg,p=0.008). Mechanical allodynia reappeared at 6 h with similar me-chanical thresholds compared to those of the control group (294.5±115.5 vs. 284.6±95.4 mg, p=0.45) and to the contralateral paws(p=0.5). The antagonistic effect was dose-dependent (Fig. 7A): therelief of mechanical allodynia by 20 μg/paw of A-317491 only lastedfor 0.5 h (531.6±195.2 mg, p=0.029), and mechanical allodyniahad re-developed by 1 h later. There was no effect on the mechanicalthresholds with an injection of 2 μg/paw (p=0.31).
There were similar dose-dependent effects of TNP–ATP on thechanges in mechanical thresholds after RTX-induced neuropathy(Fig. 7B). Both hindpaws on D7 of mice after RTX-treatment had similarlevels of mechanical allodynia (p=0.23). At a dose of 240 μg/paw, me-chanical thresholds were higher at 0.5 h (706.3±230.9 vs. 255.3±38.7 mg, p=0.029), and the effect lasted for 2 h (633.1±89.5 vs.276.5±62.6 mg, p=0.029). Mechanical allodynia had reappeared by4 h post-injection (p=0.11). With 160 μg/paw administration, me-chanical allodynia was attenuated at 0.5 (727.5±162.3 mg, p=0.018)and 1 h (425.1±95.0 mg, p=0.036). With an injection of 16 μg/paw,the antagonistic effect was only observed at 0.5 h (451.4±75.1 mg,p=0.029).
Discussion
This study demonstrates a mouse system of neuropathy with me-chanical allodynia and thermal hypoalgesia, activation of ATF3 in DRGneurons with concomitant upregulation of P2X3 receptor and down-regulation of CGRP, and structural evidence of peripheral sensitiza-tion in the development of neuropathic pain behaviors aftercutaneous nerve degeneration.
Manifestations of neuropathic pain due to small-diameter sensorynerve injury
The major theme of the report is the establishment of a mousesystem with reduced nociception but with mechanical hypersensitiv-ity. In RTX neuropathy, mice showed thermal hypoesthesia and me-chanical allodynia due to selective depletion of small-diameternociceptive nerves. Previous studies attributed injury-induced me-chanical hypersensitivity to degeneration or dysfunctions of largeDRG neurons and their nerves (Baron 2009). For example, a report in-dicated an abnormal pathology of myelinated fibers in RTX-inducedmechanical allodynia (Pan et al., 2003). The role of small DRG neuronsin this issue has received little attention. The current report demon-strates the parallel appearance of mechanical allodynia with corre-sponding upregulation of the purinergic P2X3 receptor in DRGneurons with increased P2X3(+) nerve fibers in the skin, whichwas responsible for the development of mechanical allodynia.

Fig. 5. Phenotypes of dorsal root ganglia (DRG) neurons in resiniferatoxin (RTX)-induced neuropathy. Double-labeling immunofluorescent staining was performed with transientreceptor potential vanilloid subtype 1 (TRPV1; B, D in green) and P2X3 (A in red) or calcitonin gene-related peptide (CGRP, C in red). (A–D) Only rarely P2X3(+) neurons (A, B)were co-expressed with TRPV1 than that of to CGRP(+) neurons (asterisks in C, D). (E, F) TRPV1(+) neurons were completely depleted on D7 (E) and D84 (F) after RTX. (G) Thegraph compares the co-localization ratios according to Fig. 5A–D. (H) The graph showed the quantification of TRPV1(+) neuron density according to Fig. 5B, D, E, and F. * Statis-tically significant at pb0.05. Bar, 50 μm.
322 Y.-L. Hsieh et al. / Experimental Neurology 235 (2012) 316–325
In contrast, large DRG neurons and their nerve fibers in RTX-induced neuropathy remained intact according to normal myelinatednerve morphometry and nerve conduction studies (Hsieh et al.,2008). RTX is an ultrapotent capsaicin analog (Brown et al., 2005;Iadarola and Mannes, 2011; Kissin and Szallasi, 2011). The currentstudy documents that thermal hypoalgesia and mechanical allodyniawere inversely correlated. What is the possible mechanisms underly-ing these two distinct neuropathic behaviors? One putative mecha-nism is the influence of RTX on different populations of ATF3(+)small-diameter neurons independent of TRPV1 phenotype (Braz andBasbaum, 2010). In the present report, mechanical thresholds werelinearly correlated with ratio of P2X3(+)/ATF3(+) but not withCGRP(+)/ATF3(+) neurons, suggesting that abnormal mechanicalsensations could be initiated by injuries to small-diameter sensoryneurons and mediated by enhanced purinergic signaling.
In human small-fiber neuropathy due to skin denervation, mostpatients noticed a loss of thermal nociception. A proportion of pa-tients experienced additional neuropathic pain (Bouhassira et al.,1999). The former symptom of reduced sensitivity is straightforwardas the loss of cutaneous nerve terminals lead to impaired transmis-sion of nociceptive stimuli. However, the pathophysiology andmechanisms of concomitant pain on a background of skin denerva-tion is not fully understood. Furthermore there is no correlation be-tween the degree of pain and the degree of neuropathy (Kalliomakiet al., 2011; Wildgaard et al., 2012). This report provides possiblescenario for the combination of both symptoms: the expression ofATF3 was increased in injured small-diameter neurons and the pref-erential expression of P2X3 led to mechanical allodynia.
The thermal nociception recovered at the late phase of RTX neu-ropathy despite irreversible loss of TRPV1 expression. This observa-tion suggests the existence of compensatory molecules for thermalnociception. Possible candidates for thermal transmission are
peptidergic fibers on the skin. Our previous study indicated the ther-mal latencies was linearly correlated with CGRP(+) cutaneous inner-vation (Hsieh et al., 2008). In the current report, the temporalexpression of CGRP on neuronal soma (DRG) and their peripheralprocesses (dermal nerves) were correlated with withdrawal latenciesto thermal stimuli. Further studies are required to unravel the role ofpeptidergic fibers in the absence of TRPV1.
Taken together, the inverse correlation between thermal with-drawal latencies andmechanical thresholds suggests that two distinctsmall-diameter nociceptors underlie distinct behaviors: CGRP(+) forthermal hypoalgesia and P2X3(+) for mechanical allodynia.
Activation of ATF3 and its consequences in RTX-induced neuropathy
The phenotypic profiles of ATF3(+) DRG neurons after RTX treat-ment provide molecular explanations for the behavioral manifesta-tions. Recently, a comprehensive study demonstrated increasedexpression of ATF3 in DRG neurons after a variety of nerve injuriesand noxious stimuli (Braz and Basbaum, 2010). Those observationsraise intriguing issues: what are the profiles of ATF3(+) DRG neuronsand how do such changes contribute to neuropathic pain behaviors?The current study demonstrates increased ATF3 expression withRTX-induced neuropathy and its link to denervation of cutaneousnerves. The higher percentage of CGRP/TRPV1 neurons compared toP2X3/TRPV1 neurons explains the difference in the degree of cutane-ous nerve denervation after RTX treatment.
In addition to being an injury marker, roles of ATF3 in previousstudies mainly focused on promoting nerve regeneration (Seijfferset al., 2007). That report indicated the association of ATF3 with theexpression of nociceptive molecules in DRG neurons. In the presentstudy, ATF3 was induced after RTX treatment, mainly on periph-erin(+) DRG neurons, and the pattern of ATF3 expression had
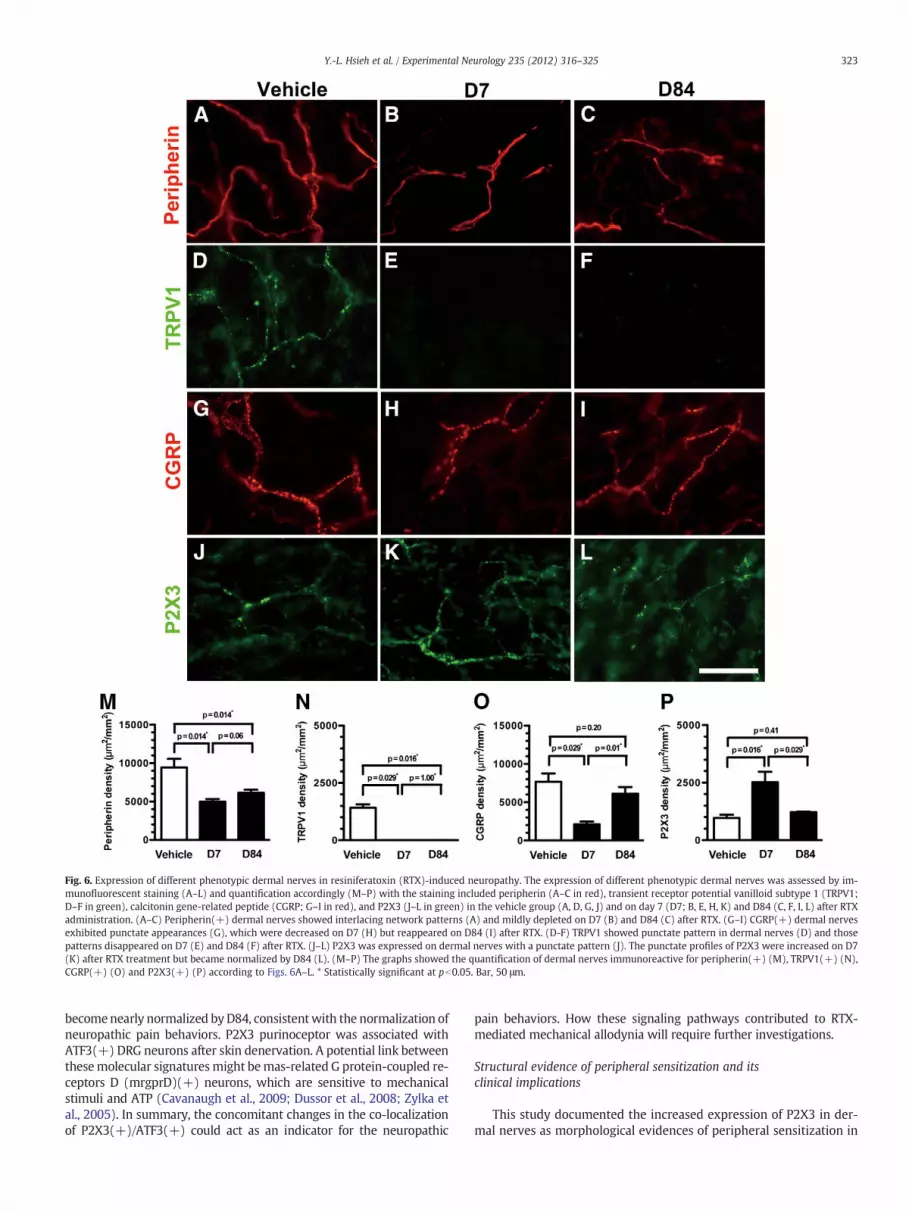
Fig. 6. Expression of different phenotypic dermal nerves in resiniferatoxin (RTX)-induced neuropathy. The expression of different phenotypic dermal nerves was assessed by im-munofluorescent staining (A–L) and quantification accordingly (M–P) with the staining included peripherin (A–C in red), transient receptor potential vanilloid subtype 1 (TRPV1;D–F in green), calcitonin gene-related peptide (CGRP; G–I in red), and P2X3 (J–L in green) in the vehicle group (A, D, G, J) and on day 7 (D7; B, E, H, K) and D84 (C, F, I, L) after RTXadministration. (A–C) Peripherin(+) dermal nerves showed interlacing network patterns (A) and mildly depleted on D7 (B) and D84 (C) after RTX. (G–I) CGRP(+) dermal nervesexhibited punctate appearances (G), which were decreased on D7 (H) but reappeared on D84 (I) after RTX. (D-F) TRPV1 showed punctate pattern in dermal nerves (D) and thosepatterns disappeared on D7 (E) and D84 (F) after RTX. (J–L) P2X3 was expressed on dermal nerves with a punctate pattern (J). The punctate profiles of P2X3 were increased on D7(K) after RTX treatment but became normalized by D84 (L). (M–P) The graphs showed the quantification of dermal nerves immunoreactive for peripherin(+) (M), TRPV1(+) (N),CGRP(+) (O) and P2X3(+) (P) according to Figs. 6A–L. * Statistically significant at pb0.05. Bar, 50 μm.
323Y.-L. Hsieh et al. / Experimental Neurology 235 (2012) 316–325
becomenearly normalized byD84, consistentwith the normalization ofneuropathic pain behaviors. P2X3 purinoceptor was associated withATF3(+) DRG neurons after skin denervation. A potential link betweenthesemolecular signatures might bemas-related G protein-coupled re-ceptors D (mrgprD)(+) neurons, which are sensitive to mechanicalstimuli and ATP (Cavanaugh et al., 2009; Dussor et al., 2008; Zylka etal., 2005). In summary, the concomitant changes in the co-localizationof P2X3(+)/ATF3(+) could act as an indicator for the neuropathic
pain behaviors. How these signaling pathways contributed to RTX-mediated mechanical allodynia will require further investigations.
Structural evidence of peripheral sensitization and itsclinical implications
This study documented the increased expression of P2X3 in der-mal nerves as morphological evidences of peripheral sensitization in

Fig. 7. Effects of P2X3 antagonism on mechanical allodynia after resiniferatoxin (RTX)-induced neuropathy. Antagonists against P2X3, A-317491 (A), and TNP–ATP (B) wereadministrated into the hindpaw by an intraplantar injection. Mechanical thresholdswere evaluated at 0.5, 1, 2, 4, and 6 h by the von Frey filament test. (A) A-317491 re-lieved mechanical allodynia for up to 4 h in a dose-dependent pattern. (B) TNP–ATPnormalized mechanical thresholds in a dose-dependent fashion but with a shorter du-ration of effects than A-317491. * Statistically significant at pb0.05.
324 Y.-L. Hsieh et al. / Experimental Neurology 235 (2012) 316–325
RTX-induced neuropathic pain. Previously, most studies mainlydepended on a pharmacological intervention of peripheral tissuesto document peripheral sensitization, for example, demonstratingchanges in neuropathic pain behaviors by an intraplantar injection.A gap exists between structural data and pharmacological results,i.e., a lack of evidence showing that P2X3 expression increased incutaneous nerve terminal regions. The current report shows struc-tural evidence that increased P2X3 expression at dermal nerveterminals is responsible for injury-induced mechanical allodynia.Such observations were supported by an intraplantar injection ofP2X3 antagonists to normalize mechanical allodynia. Currently,most treatment guidelines for neuropathic pain recommend theuse of antidepressants and anticonvulsants, for example, the block-ing of the α2δ subunit of the calcium channel (Attal et al., 2010;O'Connor and Dworkin, 2009) and treatment responses are notsatisfactory (Attal et al., 2010; Finnerup et al., 2010). Taken to-gether with a recent report on cancer pain (Kaan et al., 2010),the present study provides not only a system to screen medica-tions of mechanical hypersensitivity for preclinical studies butalso a new therapeutic strategy to design pharmacologic interven-tions for neuropathic pain, particularly at the level of cutaneousnerve terminals.
Acknowledgments
This work was supported by grants from National Health ResearchInstitute (NHRI-EX99-9736NI), National Science Council (NSC97-2320-B-002-042-MY3, NSC100-2320-B-037-018), Kaohsiung MedicalUniversity Research Foundation (KMU-Q099005, KMU-M100004),and the Excellent Translational Medicine Research Projects ofNTUMC and NTUH (98C101-201).
References
Attal, N., Cruccu, G., Baron, R., Haanpaa, M., Hansson, P., Jensen, T.S., Nurmikko, T., 2010.EFNS guidelines on the pharmacological treatment of neuropathic pain: 2010 revi-sion. Eur. J. Neurol. 17, e1113–e1188.
Avelino, A., Cruz, F., 2000. Peptide immunoreactivity and ultrastructure of rat urinarybladder nerve fibers after topical desensitization by capsaicin or resiniferatoxin.Auton. Neurosci. 86, 37–46.
Banerjee, B., Medda, B.K., Schmidt, J., Zheng, Y., Zhang, Z., Shaker, R., Sengupta, J.N.,2009. Altered expression of P2X3 in vagal and spinal afferents following esophagi-tis in rats. Histochem. Cell Biol. 132, 585–597.
Baron, R., 2009. Neuropathic pain: a clinical perspective. Handb. Exp. Pharmacol. 3–30.Baron, R., Tolle, T.R., Gockel, U., Brosz, M., Freynhagen, R., 2009. A cross-sectional cohort
survey in 2100 patients with painful diabetic neuropathy and postherpetic neural-gia: differences in demographic data and sensory symptoms. Pain 146, 34–40.
Bouhassira, D., Attal, N., Willer, J.C., Brasseur, L., 1999. Painful and painless peripheralsensory neuropathies due to HIV infection: a comparison using quantitative senso-ry evaluation. Pain 80, 265–272.
Braz, J.M., Basbaum, A.I., 2010. Differential ATF3 expression in dorsal root ganglionneurons reveals the profile of primary afferents engaged by diverse noxious chem-ical stimuli. Pain 150, 290–301.
Brown, D.C., Iadarola, M.J., Perkowski, S.Z., Erin, H., Shofer, F., Laszlo, K.J., Olah, Z.,Mannes, A.J., 2005. Physiologic and antinociceptive effects of intrathecal resinifer-atoxin in a canine bone cancer model. Anesthesiology 103, 1052–1059.
Burnstock, G., 2009. Purinergic receptors and pain. Curr. Pharm. Des. 15, 1717–1735.Cavanaugh, D.J., Lee, H., Lo, L., Shields, S.D., Zylka, M.J., Basbaum, A.I., Anderson, D.J.,
2009. Distinct subsets of unmyelinated primary sensory fibers mediate behavioralresponses to noxious thermal and mechanical stimuli. Proc. Natl. Acad. Sci. U.S.A.106, 9075–9080.
Chaplan, S.R., Bach, F.W., Pogrel, J.W., Chung, J.M., Yaksh, T.L., 1994. Quantitative assess-ment of tactile allodynia in the rat paw. J. Neurosci. Methods 53, 55–63.
Cheng, H.T., Dauch, J.R., Hayes, J.M., Hong, Y., Feldman, E.L., 2009. Nerve growth factormediates mechanical allodynia in a mouse model of type 2 diabetes. J. Neuro-pathol. Exp. Neurol. 68, 1229–1243.
Dussor, G., Zylka, M.J., Anderson, D.J., McCleskey, E.W., 2008. Cutaneous sensory neu-rons expressing the Mrgprd receptor sense extracellular ATP and are putative noci-ceptors. J. Neurophysiol. 99, 1581–1589.
Finnerup, N.B., Sindrup, S.H., Jensen, T.S., 2010. The evidence for pharmacological treat-ment of neuropathic pain. Pain 150, 573–581.
Fukuoka, T., Yamanaka, H., Kobayashi, K., Okubo, M., Miyoshi, K., Dai, Y., Noguchi, K.,2012. Re-evaluation of the phenotypic changes in L4 dorsal root ganglion neuronsafter L5 spinal nerve ligation. Pain 153, 68–79.
Gibbons, C.H., Wang, N., Freeman, R., 2010. Capsaicin induces degeneration of cutane-ous autonomic nerve fibers. Ann. Neurol. 68, 888–898.
Hai, T., Hartman, M.G., 2001. The molecular biology and nomenclature of the activatingtranscription factor/cAMP responsive element binding family of transcription fac-tors: activating transcription factor proteins and homeostasis. Gene 273, 1–11.
Hsieh, Y.L., Chiang, H., Tseng, T.J., Hsieh, S.T., 2008. Enhancement of cutaneous nerve re-generation by 4-methylcatechol in resiniferatoxin-induced neuropathy. J. Neuro-pathol. Exp. Neurol. 67, 93–104.
Iadarola, M.J., Mannes, A.J., 2011. The vanilloid agonist resiniferatoxin for interventional-based pain control. Curr. Top. Med. Chem. 11, 2171–2179.
Kaan, T.K., Yip, P.K., Patel, S., Davies, M., Marchand, F., Cockayne, D.A., Nunn, P.A.,Dickenson, A.H., Ford, A.P., Zhong, Y., Malcangio, M., McMahon, S.B., 2010. Systemicblockade of P2X3 and P2X2/3 receptors attenuates bone cancer pain behaviour inrats. Brain 133, 2549–2564.
Kalliomaki, M., Kieseritzky, J.V., Schmidt, R., Hagglof, B., Karlsten, R., Sjogren, N.,Albrecht, P., Gee, L., Rice, F., Wiig, M., Schmelz, M., Gordh, T., 2011. Structural andfunctional differences between neuropathy with and without pain? Exp. Neurol.231, 199–206.
Kissin, I., Szallasi, A., 2011. Therapeutic targeting of TRPV1 by resiniferatoxin, from pre-clinical studies to clinical trials. Curr. Top. Med. Chem. 11, 2159–2170.
Lin, Y.Y., Tseng, T.J., Hsieh, Y.L., Luo, K.R., Lin, W.M., Chiang, H., Hsieh, S.T., 2008. Deple-tion of peptidergic innervation in the gastric mucosa of streptozotocin-induced di-abetic rats. Exp. Neurol. 213, 388–396.
Neubert, J.K., Karai, L., Jun, J.H., Kim, H.S., Olah, Z., Iadarola, M.J., 2003. Peripherally in-duced resiniferatoxin analgesia. Pain 104, 219–228.
Obrosova, I.G., 2009. Diabetic painful and insensate neuropathy: pathogenesis andpotential treatments. Neurotherapeutics 6, 638–647.
O'Connor, A.B., Dworkin, R.H., 2009. Treatment of neuropathic pain: an overview of re-cent guidelines. Am. J. Med. 122, S22–S32.
Oliveira, M.C., Pelegrini-da-Silva, A., Tambeli, C.H., Parada, C.A., 2009. Peripheral mech-anisms underlying the essential role of P2X3,2/3 receptors in the development ofinflammatory hyperalgesia. Pain 141, 127–134.

325Y.-L. Hsieh et al. / Experimental Neurology 235 (2012) 316–325
Pan, H.L., Khan, G.M., Alloway, K.D., Chen, S.R., 2003. Resiniferatoxin induces paradox-ical changes in thermal and mechanical sensitivities in rats: mechanism of action. J.Neurosci. 23, 2911–2919.
Polydefkis, M., Hauer, P., Sheth, S., Sirdofsky, M., Griffin, J.W., McArthur, J.C., 2004. Thetime course of epidermal nerve fibre regeneration: studies in normal controls andin people with diabetes, with and without neuropathy. Brain 127, 1606–1615.
Rage, M., Van Acker, N., Facer, P., Shenoy, R., Knaapen, M.W., Timmers, M., Streffer, J.,Anand, P., Meert, T., Plaghki, L., 2010. The time course of CO2 laser-evoked re-sponses and of skin nerve fibre markers after topical capsaicin in human volun-teers. Clin. Neurophysiol. 121, 1256–1266.
Seijffers, R., Mills, C.D., Woolf, C.J., 2007. ATF3 increases the intrinsic growth state of DRGneurons to enhance peripheral nerve regeneration. J. Neurosci. 27, 7911–7920.
Tschachler, E., Reinisch, C.M., Mayer, C., Paiha, K., Lassmann, H.,Weninger,W., 2004. Sheetpreparations expose the dermal nerve plexus of human skin and render the dermalnerve end organ accessible to extensive analysis. J. Invest. Dermatol. 122, 177–182.
Tsuchihara, T., Ogata, S., Nemoto, K., Okabayashi, T., Nakanishi, K., Kato, N.,Morishita, R., Kaneda, Y., Uenoyama, M., Suzuki, S., Amako, M., Kawai, T.,Arino, H., 2009. Nonviral retrograde gene transfer of human hepatocyte growthfactor improves neuropathic pain-related phenomena in rats. Mol. Ther. 17,42–50.
Wildgaard, K., Ringsted, T.K., Hansen, H.J., Petersen, R.H., Werner, M.U., Kehlet, H.,2012. Quantitative sensory testing of persistent pain after video-assisted thoracicsurgery lobectomy. Br. J. Anaesth. 108, 126–133.
Xu, G.Y., Shenoy, M., Winston, J.H., Mittal, S., Pasricha, P.J., 2008. P2X receptor-mediated visceral hyperalgesia in a rat model of chronic visceral hypersensitivity.Gut 57, 1230–1237.
Zimmermann, M., 1983. Ethical guidelines for investigations of experimental pain inconscious animals. Pain 16, 109–110.
Zylka, M.J., Rice, F.L., Anderson, D.J., 2005. Topographically distinct epidermal nocicep-tive circuits revealed by axonal tracers targeted to Mrgprd. Neuron 45, 17–25.