oxytocin and vasopressin receptors and species-typical social behaviors
TRANSCRIPT

FOS
LDA
R
ersTitfsrSnibortoattt
haavta
p
dE7
Hormones and Behavior 36, 212–221 (1999)Article ID hbeh.1999.1548, available online at http://www.idealibrary.com on
2
RANK A. BEACH AWARDxytocin and Vasopressin Receptors andpecies-Typical Social Behaviors1
arry J. Young2
epartment of Psychiatry and Behavioral Sciences, Emory University School of Medicine,tlanta, Georgia 30322
eceived June 8, 1999; revised July 20, 1999; accepted July 23, 1999
r1lphdd(
dwPsK1fcite1rfH
fcAm((GsW
Over the past 50 years, a great deal of behavioralndocrinological research has established a centralole for gonadal steroid hormones in the control ofexual behaviors in a wide range of vertebrate taxa.he underlying mechanisms controlling these behav-
ors appear to be well conserved throughout evolu-ion. Both the neural structures regulating male andemale sexual behaviors and the distribution of sexteroid receptors within the brain are similar fromeptiles to mammals (Meisel and Sachs, 1994; Pfaff andchwartz-Giblin, 1994). More recently, the roles of theeuropeptides oxytocin (OT) and vasopressin (AVP)
n modulating reproductive and social behaviors haveegun to be elucidated. Comparative studies focusingn these hormones suggest that unlike gonadal ste-oids, the specific behavioral roles of these neuropep-ides may be quite species specific. Here I review somef the behaviors associated with OT and AVP systemsnd discuss the idea that OT and AVP receptor sys-ems are phylogenetically plastic, perhaps facilitatinghe evolution of species-typical social behavior pat-erns.
Oxytocin and vasopressin are related nonapeptideormones which differ in structure at only two aminocid positions (Gainer and Wray, 1994). Both peptidesre synthesized in magnocellular neurons of the para-entricular (PVN) and supraoptic nuclei of the hypo-halamus, which project to the neurohypophysis andre released into circulation where they modulate pe-
1 This article is based on the 1998 Frank A. Beach Award lecture,resented at the 1998 Society for Neuroscience Meeting.
2 To whom correspondence and reprint requests should be ad-ressed at Department of Psychiatry and Behavioral Sciences,
gp
mory University School of Medicine, Atlanta GA 30322. Fax: (404)27-3233. E-mail: [email protected].
12
ipheral physiology (Brownstein, Russell, and Gainer,980). The peptides are also synthesized in parvocel-uar neurons of the PVN and other limbic nuclei whichroject throughout the brain where they influence be-avior. Vasopressin synthesis by neurons in the amyg-ala and bed nucleus of the stria terminalis is sexuallyimorphic, with males producing more than females
DeVries and Al-Shamma, 1990).Centrally released oxytocin plays a role in the in-
uction of maternal behavior in rats (Pedersen, Cald-ell, Walker, Ayers, and Mason, 1994; Pedersen andrange, 1979), house mice (McCarthy, 1990), andheep (Costa, Guevara-Guzman, Ohkura, Goode, andendrick, 1996; Kendrick, Keverne, and Baldwin,987). Infusion of OT into the brain of virgin animalsacilitates maternal behavior, while blocking OT re-eptors prevents these behaviors. In rats, oxytocin isnvolved in the induction of sexual receptivity by es-rogen and progesterone (Caldwell, Prange, and Ped-rsen, 1986; Schumacher, Coirini, Pfaff, and McEwen,990). In addition, OT appears to play an importantole in the formation of long-lasting pair bonds inemale monogamous prairie voles (Williams, Insel,arbaugh, and Carter, 1994).While many of the roles of OT are associated with
emale-typical behaviors, most of the behaviors asso-iated with AVP have been demonstrated in the male.rginine vasotocin (AVT), the nonmammalian ho-olog of AVP, stimulates sexual behavior in newts
Moore and Miller, 1983) and vocalization in frogsBoyd, 1994) and birds (Goodson, 1998a; Maney,oode, and Wingfield, 1997). Vasopressin facilitates
centmarking behavior in hamsters (Ferris, Albers,esolowski, Goldman, and Leeman, 1984) and ag-
ression in hamsters and voles (Ferris, Melloni, Kop-el, Perry, Fuller, and Delville, 1997; Young, Winslow,
0018-506X/99 $30.00Copyright © 1999 by Academic Press
All rights of reproduction in any form reserved.

NivHpcsziv1snmW1sa
SA
Gwb(rrwB(fa1h1raeaatJsaIitb
tvsbzsaIrvpmprtbpno
OrscshbOOa1fwf1coccsic1tpDbtro
213Oxytocin and Vasopressin Receptors
ilsen, and Insel, 1997) and plays a role in pair bond-ng and paternal care in male monogamous prairieoles (Wang, Ferris, and DeVries, 1994; Winslow,astings, Carter, Harbaugh, and Insel, 1993). Vaso-ressin modulates social behaviors in a species-spe-ific manner. For example, Goodson recently demon-trated that AVT increases aggression in the colonialebra finch (Goodson and Adkins-Regan, 1998) butnhibits aggression in the territorial field sparrow andiolet eared waxbill (Goodson, 1998a; Goodson,998b). Similarly, AVP increases male–male aggres-ion and affiliative behavior in the male monogamous,onterritorial prairie vole, but not in the nonmonoga-ous, territorial montane vole (Young, Nilsen,aymire, MacGregor, and Insel, 1999; Young et al.,
997). These observations suggest that the role of va-opressin in regulating social behavior may varycross species with social organization.
PECIES DIFFERENCES IN OXYTOCINND VASOPRESSIN RECEPTORS
The central effects of OT and AVP are mediated by-protein coupled, seven transmembrane receptors,hich, like the peptides, share similar structures (Bar-
eris and Tribollet, 1996). A single oxytocin receptorOTR) has been identified and is present in both pe-ipheral and brain tissues. Three subtypes of AVPeceptors, V1a, V1b, and V2, have been identifiedhich differ both structurally and pharmacologically.oth V1a and V1b receptors are expressed in the brain
Barberis, Balestre, Jard, Tribollet, Arsenijevic, Drei-uss, Bankowski, Manning, Chan, Schlosser, Holsboer,nd Elands, 1995; Vaccari, Lolait, and Ostrowski,998), although most of the behavioral effects of AVPave been attributed to the V1a subtype (Wang et al.,994; Winslow et al., 1993). Comparisons of the neu-oanatomical distribution of OTR and V1a receptorsmong species have revealed marked species differ-nces in the pattern of receptor binding in the brain,lthough some generalities have been noted (Barberisnd Tribollet, 1996). For example, rats (DeKloet, Rot-eveel, Voorhuis, and Terlou, 1985; Tribollet, Barberis,ard, Dubois-Dauphin, and Dreifuss, 1988), mice (In-el, Young, Witt, and Crews, 1993), and voles (Inselnd Shapiro, 1992; Young, Huot, Nilsen, Wang, andnsel, 1996) each have a distinct pattern of OTR bind-ng in the brain. Figure 1 illustrates the different pat-
erns of vasopressin (V1a) receptor binding sites in therains of several species. As can be seen in this illus-tV
ration, the distribution and densities of V1a receptorsary quite dramatically even among closely relatedpecies. There is evidence that the pattern of receptorinding in the brain is associated with social organi-ation. For example, among voles, species with similarocial organizations share a common pattern of OTRnd V1a receptor binding (Insel and Shapiro, 1992;nsel, Wang, and Ferris, 1994). The monogamous prai-ie (Microtus ochrogaster) and pine (Microtus pinetorum)oles have similar OTR and V1a receptor bindingatterns which are distinct from the nonmonogamousontane (Microtus montanus) and meadow (Microtus
ennsylvanicus) voles. In these species differences ineceptor distribution and density may contribute tohe species differences in the effects of OT and AVP onehavior. The association between receptor bindingattern and social organization is correlative, but doesot prove a relationship between regional distributionf receptors and social behavior.In addition to species differences in distribution ofT and V1a receptors in the brain, the receptors are
egulated differently by gonadal steroids in differentpecies. This species-specific regulation of peptide re-eptors may have important implications for under-tanding the roles of the peptides in modulating be-avior. In the female rat, the induction of lordosisehavior by estrogen involves the up-regulation ofTR in the VMN. Estrogen increases both the level ofTR binding and gene expression in the VMN (Bale
nd Dorsa, 1995; Johnson, Coirini, Insel, and McEwen,991; Tribollet, Audigier, Dubois-Dauphin, and Drei-uss, 1990) and OTR expression in the VMN varies
ith the ovarian cycle, with peak levels of expressionound during proestrus (Bale, Dorsa, and Johnston,995). The ovulatory surge of progesterone thenauses the OTR to spread to the ventrolateral regionsf the VMN in close proximity to OT fibers (Schuma-her, Coirini, Frankfurt, and McEwen, 1989; Schuma-her et al., 1990). Blockade of the OT receptor withpecific antagonists (Witt and Insel, 1991) or prevent-ng the up-regulation of OTR using antisense oligonu-leotides (McCarthy, Kleopoulos, Mobbs, and Pfaff,994) inhibits sexual receptivity in the female rat. Inhe female prairie vole, which is induced into estrusheromonally and ovulates after mating (Sawrey andewsbury, 1985), estrogen does not influence OTRinding in the VMN (Witt, Carter, and Insel, 1991). Inhe mouse, gonadectomy increases and gonadal ste-oids decrease OTR binding in the VMN, a patternpposite to that of the rat (Insel et al., 1993). Therefore,
he relationship between gonadal steroids, OTR in theMN, and sexual receptivity may differ among rodent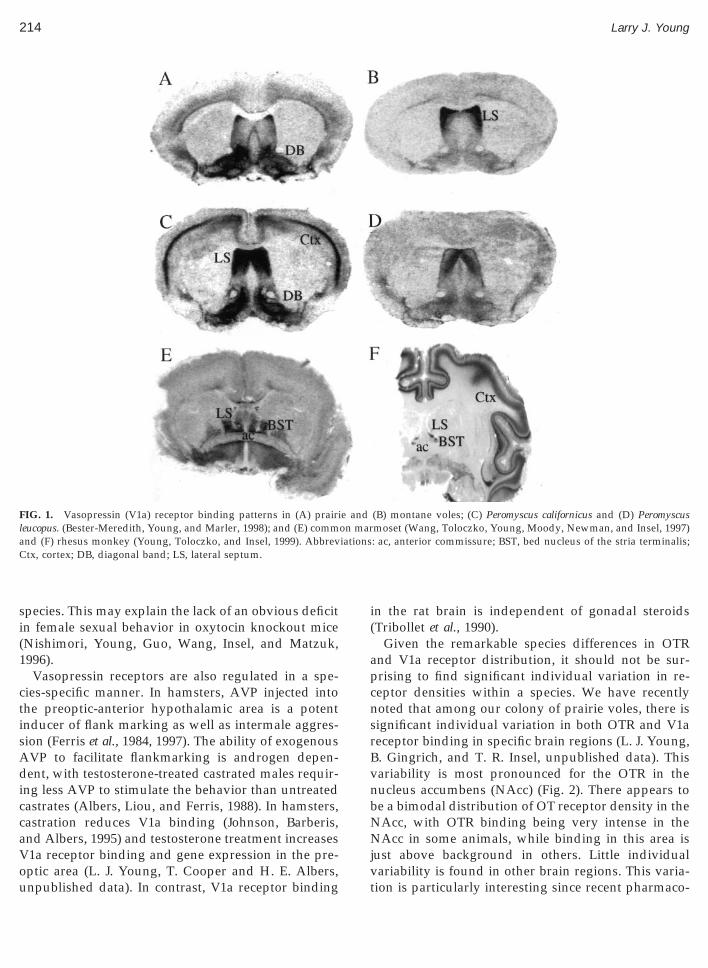
si(1
ctisAdiccaVou
i(
apcnsrBvnbNNj
Fla iationsC
214 Larry J. Young
pecies. This may explain the lack of an obvious deficitn female sexual behavior in oxytocin knockout miceNishimori, Young, Guo, Wang, Insel, and Matzuk,996).Vasopressin receptors are also regulated in a spe-
ies-specific manner. In hamsters, AVP injected intohe preoptic-anterior hypothalamic area is a potentnducer of flank marking as well as intermale aggres-ion (Ferris et al., 1984, 1997). The ability of exogenousVP to facilitate flankmarking is androgen depen-ent, with testosterone-treated castrated males requir-
ng less AVP to stimulate the behavior than untreatedastrates (Albers, Liou, and Ferris, 1988). In hamsters,astration reduces V1a binding (Johnson, Barberis,nd Albers, 1995) and testosterone treatment increases1a receptor binding and gene expression in the pre-
IG. 1. Vasopressin (V1a) receptor binding patterns in (A) prairieucopus. (Bester-Meredith, Young, and Marler, 1998); and (E) commnd (F) rhesus monkey (Young, Toloczko, and Insel, 1999). Abbrevtx, cortex; DB, diagonal band; LS, lateral septum.
ptic area (L. J. Young, T. Cooper and H. E. Albers,npublished data). In contrast, V1a receptor binding
vt
n the rat brain is independent of gonadal steroidsTribollet et al., 1990).
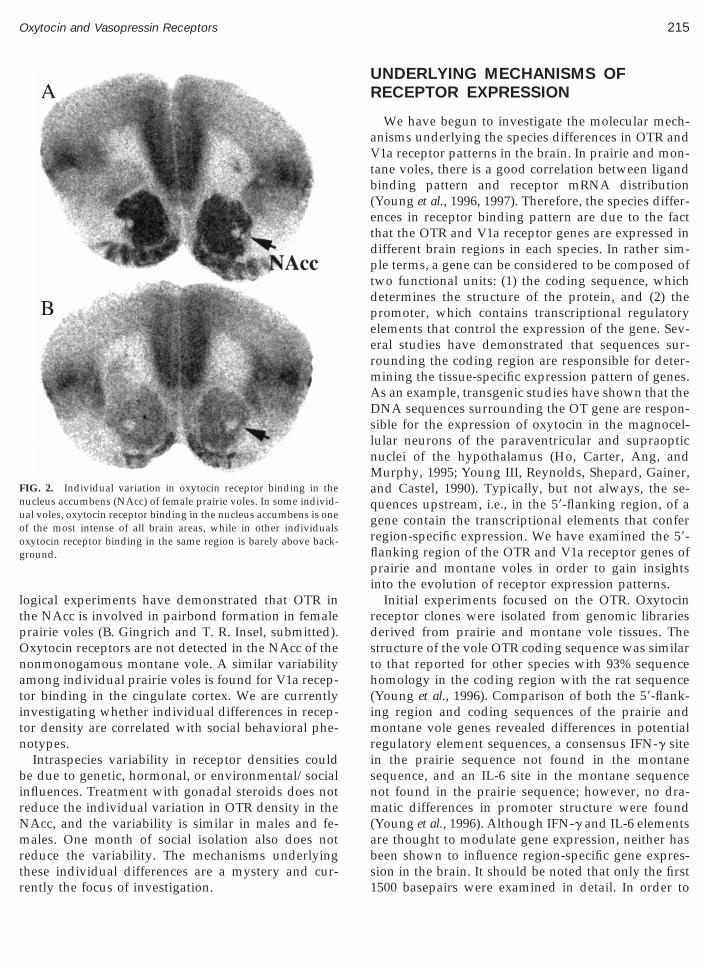
Given the remarkable species differences in OTRnd V1a receptor distribution, it should not be sur-rising to find significant individual variation in re-eptor densities within a species. We have recentlyoted that among our colony of prairie voles, there isignificant individual variation in both OTR and V1aeceptor binding in specific brain regions (L. J. Young,. Gingrich, and T. R. Insel, unpublished data). Thisariability is most pronounced for the OTR in theucleus accumbens (NAcc) (Fig. 2). There appears toe a bimodal distribution of OT receptor density in theAcc, with OTR binding being very intense in theAcc in some animals, while binding in this area is
ust above background in others. Little individual
(B) montane voles; (C) Peromyscus californicus and (D) Peromyscusmoset (Wang, Toloczko, Young, Moody, Newman, and Insel, 1997): ac, anterior commissure; BST, bed nucleus of the stria terminalis;
e andon mar
ariability is found in other brain regions. This varia-ion is particularly interesting since recent pharmaco-
ltpOnatitn
birNmrtr
UR
aVtb(etdptdpeermADslnMaqgrflpi
rdsth(imrisnm(ab
Fnuoog
215Oxytocin and Vasopressin Receptors
ogical experiments have demonstrated that OTR inhe NAcc is involved in pairbond formation in femalerairie voles (B. Gingrich and T. R. Insel, submitted).xytocin receptors are not detected in the NAcc of theonmonogamous montane vole. A similar variabilitymong individual prairie voles is found for V1a recep-or binding in the cingulate cortex. We are currentlynvestigating whether individual differences in recep-or density are correlated with social behavioral phe-otypes.Intraspecies variability in receptor densities could
e due to genetic, hormonal, or environmental/socialnfluences. Treatment with gonadal steroids does noteduce the individual variation in OTR density in theAcc, and the variability is similar in males and fe-ales. One month of social isolation also does not
educe the variability. The mechanisms underlying
IG. 2. Individual variation in oxytocin receptor binding in theucleus accumbens (NAcc) of female prairie voles. In some individ-al voles, oxytocin receptor binding in the nucleus accumbens is onef the most intense of all brain areas, while in other individualsxytocin receptor binding in the same region is barely above back-round.
hese individual differences are a mystery and cur-ently the focus of investigation.
s1
NDERLYING MECHANISMS OFECEPTOR EXPRESSION
We have begun to investigate the molecular mech-nisms underlying the species differences in OTR and1a receptor patterns in the brain. In prairie and mon-
ane voles, there is a good correlation between ligandinding pattern and receptor mRNA distributionYoung et al., 1996, 1997). Therefore, the species differ-nces in receptor binding pattern are due to the facthat the OTR and V1a receptor genes are expressed inifferent brain regions in each species. In rather sim-le terms, a gene can be considered to be composed of
wo functional units: (1) the coding sequence, whichetermines the structure of the protein, and (2) theromoter, which contains transcriptional regulatorylements that control the expression of the gene. Sev-ral studies have demonstrated that sequences sur-ounding the coding region are responsible for deter-ining the tissue-specific expression pattern of genes.s an example, transgenic studies have shown that theNA sequences surrounding the OT gene are respon-
ible for the expression of oxytocin in the magnocel-ular neurons of the paraventricular and supraopticuclei of the hypothalamus (Ho, Carter, Ang, andurphy, 1995; Young III, Reynolds, Shepard, Gainer,
nd Castel, 1990). Typically, but not always, the se-uences upstream, i.e., in the 59-flanking region, of aene contain the transcriptional elements that conferegion-specific expression. We have examined the 59-anking region of the OTR and V1a receptor genes ofrairie and montane voles in order to gain insights
nto the evolution of receptor expression patterns.Initial experiments focused on the OTR. Oxytocin
eceptor clones were isolated from genomic librarieserived from prairie and montane vole tissues. Thetructure of the vole OTR coding sequence was similaro that reported for other species with 93% sequenceomology in the coding region with the rat sequenceYoung et al., 1996). Comparison of both the 59-flank-ng region and coding sequences of the prairie and
ontane vole genes revealed differences in potentialegulatory element sequences, a consensus IFN-g siten the prairie sequence not found in the montaneequence, and an IL-6 site in the montane sequenceot found in the prairie sequence; however, no dra-atic differences in promoter structure were found
Young et al., 1996). Although IFN-g and IL-6 elementsre thought to modulate gene expression, neither haseen shown to influence region-specific gene expres-
ion in the brain. It should be noted that only the first500 basepairs were examined in detail. In order to
dgcg5tbbTwfbwsVaGwlctgcpsimocasHqbgseorwponm
mrvdtgt
rp5DgtRcapnptObanqjssilpecttspiVporindtfpomdh
Vvor
216 Larry J. Young
etermine whether the 59-flanking region of the OTRene contained sequences responsible for tissue-spe-ific expression in the brain, we created a reporterene in which 5000 basepairs of the prairie vole OTR9-flanking region were spliced upstream to the bac-erial lac-Z gene. The lac-Z gene encodes the enzyme-galactosidase, which, when expressed, produces alue precipitate under the appropriate conditions.his transgene was injected into mouse embryoshere it incorporated into the mouse genome. Of the
our transgenic lines produced, one line expressed-galactosidase in several regions of the mouse brain,hich, in the prairie vole brain, expresses the OTR,
uch as the cortex, lateral septum, amygdala, andMN (Young, Waymire, Nilsen, Macgregor, Wang,nd Insel, 1996; Young, Winslow, Wang, Gingrich,uo, Matzuk, and Insel, 1997). The expression patternas not identical to that of the prairie vole OTR since
ittle b-galactosidase was detected in the prelimbicortex and some ectopic expression was found in thehalamus. Caution must be taken in interpreting trans-enic experiments such as this because reporter geneonstructs often result in ectopic expression and inde-endently created lines using identical DNA con-tructs can result in different expression patterns. Thiss likely due to the fact that distant regulatory ele-
ents upstream of the promoter or located in intronsr in the 39-flanking region, which are missing in theonstruct, may also contribute to gene expression. Inddition, transgene expression is influenced by theite of integration of the transgene in the genome.owever, these results do suggest that DNA se-
uences upstream of the OTR coding region are capa-le of conferring tissue-specific expression of the OTRene and therefore could potentially contribute to thepecies-specific pattern of gene expression. A morextensive comparison of the 59- and 39-flanking regionf the OTR gene from montane and prairie voles mayeveal more extensive differences in gene structurehich could contribute to species differences in ex-ression patterns. Nonetheless, this experiment dem-nstrates the ability of transgenic technology to ma-ipulate receptor gene expression in a targetedanner.Comparison of the V1a receptor genes of prairie andontane voles has been more informative. The V1a
eceptor coding sequence is 98% identical between theole species (Young et al., 1999). However, two majorifferences were found between the prairie and mon-
ane V1a receptor loci. First, while the montane vole
enome has a single V1a receptor gene as expected,he prairie vole genome has two distinct loci, likely the5r
esult of a gene duplication (Fig. 3A). One of therairie vole genes has a LINE element located in the9-flanking region. LINE elements are transposableNA elements, making up 5% of the mammalianenome, which encode a reverse transcriptase similaro that of retroviruses (Evans and Palmiter, 1991).ecently, it has been shown that LINEs are capable ofausing surrounding DNA sequences to be duplicatednd translocated to different chromosomal regions,roviding a mechanism for the evolution of the ge-ome (Moran, Deberardinis, and Kazazian, 1999). Therairie vole V1a receptor gene may be an example of
he evolutionary power of LINEs at the genomic level.ne of the prairie vole V1a receptor loci has a singlease mutation, resulting in a premature stop codon,nd therefore a truncated receptor protein which mayot be functional. Both prairie vole loci have a se-uence stretch of approximately 460 basepairs located
ust 723 basepairs upstream of the transcription startite which is absent in the montane vole gene. Thisequence appears to be an expansion of a short repet-tive sequence which is found in the montane voleocus in this area. These data suggest that the ancestralrairie vole V1a receptor gene first accumulated thexpansion in the 59-flanking region and then dupli-ated, perhaps via a LINE dependent mechanism. Ei-her the duplication or the accumulation of the addi-ional promoter sequence near the transcription startite could dramatically impact the neuroanatomicalattern of receptor gene expression. Although at first
t may seem that the duplication of the prairie vole1a gene is not significant since one copy contains aremature stop, this process would result in one copyf the gene translocating to a new chromosomal envi-onment. Since both the regulatory element surround-ng the gene and the position of the gene in the ge-ome influence expression, gene duplication couldramatically alter expression if the remaining func-
ional gene is the translocated locus. Despite the dif-erences in gene structure, we cannot rule out theossibility that species differences in the distributionf the transcription factors which interact with pro-oter elements could alone account for the species
ifferences in V1a receptor expression patterns. Weave not investigated this possibility in the voles.Since the monogamous pine vole has a pattern of
1a receptor binding that is similar to that of prairieoles, while the meadow vole pattern is similar to thatf the montane vole, we also examined the 59-flankingegion of these species. Using PCR to amplify the
9-flanking region in the pine and meadow vole V1aeceptor genes we found that pine voles, like prairie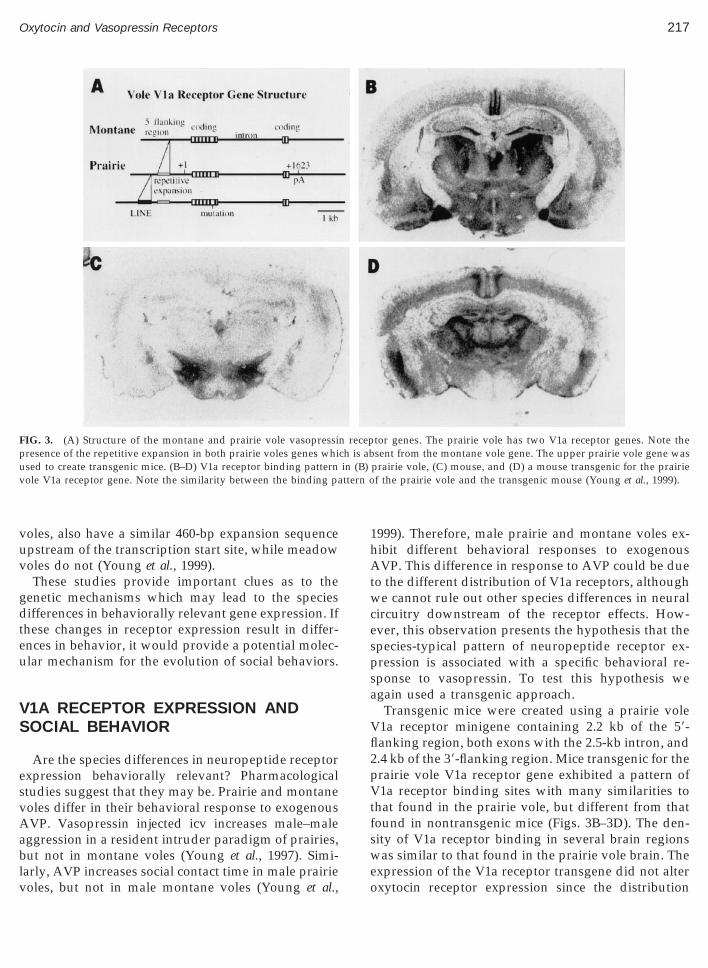
vuv
gdteu
VS
esvAablv
1hAtwcespsa
Vfl2pVtfsw
Fpu in (B)v ttern o
217Oxytocin and Vasopressin Receptors
oles, also have a similar 460-bp expansion sequencepstream of the transcription start site, while meadowoles do not (Young et al., 1999).These studies provide important clues as to the
enetic mechanisms which may lead to the speciesifferences in behaviorally relevant gene expression. If
hese changes in receptor expression result in differ-nces in behavior, it would provide a potential molec-lar mechanism for the evolution of social behaviors.
1A RECEPTOR EXPRESSION ANDOCIAL BEHAVIOR
Are the species differences in neuropeptide receptorxpression behaviorally relevant? Pharmacologicaltudies suggest that they may be. Prairie and montaneoles differ in their behavioral response to exogenousVP. Vasopressin injected icv increases male–male
ggression in a resident intruder paradigm of prairies,ut not in montane voles (Young et al., 1997). Simi-
IG. 3. (A) Structure of the montane and prairie vole vasopressinresence of the repetitive expansion in both prairie voles genes whised to create transgenic mice. (B–D) V1a receptor binding patternole V1a receptor gene. Note the similarity between the binding pa
arly, AVP increases social contact time in male prairieoles, but not in male montane voles (Young et al.,
eo
999). Therefore, male prairie and montane voles ex-ibit different behavioral responses to exogenousVP. This difference in response to AVP could be due
o the different distribution of V1a receptors, althoughe cannot rule out other species differences in neural
ircuitry downstream of the receptor effects. How-ver, this observation presents the hypothesis that thepecies-typical pattern of neuropeptide receptor ex-ression is associated with a specific behavioral re-ponse to vasopressin. To test this hypothesis wegain used a transgenic approach.Transgenic mice were created using a prairie vole
1a receptor minigene containing 2.2 kb of the 59-anking region, both exons with the 2.5-kb intron, and.4 kb of the 39-flanking region. Mice transgenic for therairie vole V1a receptor gene exhibited a pattern of1a receptor binding sites with many similarities to
hat found in the prairie vole, but different from thatound in nontransgenic mice (Figs. 3B–3D). The den-ity of V1a receptor binding in several brain regionsas similar to that found in the prairie vole brain. The
tor genes. The prairie vole has two V1a receptor genes. Note thesent from the montane vole gene. The upper prairie vole gene was
prairie vole, (C) mouse, and (D) a mouse transgenic for the prairief the prairie vole and the transgenic mouse (Young et al., 1999).
recepch is ab
xpression of the V1a receptor transgene did not alterxytocin receptor expression since the distribution
agAgstmapltwhchsttrm
pvwniAsfcspntrttt
ptir(mpNtha
n
atmtpmntaahe
Fimitff
218 Larry J. Young
nd density of OTR binding was identical in trans-enic and nontransgenic mice (Young et al., 1999).lthough the distribution of V1a binding in the trans-enic mouse was similar to the prairie vole, there wereome notable differences. For example, V1a binding inhe hypothalamus and amygdala of the transgenic
ouse was less intense than that of the prairie vole. Inddition, since the transgenic mouse continued to ex-ress its endogenous V1a receptor gene, binding in the
ateral septum of the transgenic mouse was higherhan in the prairie vole, but similar to that of the
ildtype mouse. Interestingly, the V1a binding in theypothalamus was decreased in the transgenic mouseompared to that of the wildtype mouse. With in situybridization, we have detected antisense V1a tran-cripts in the lateral hypothalamus, indicating theransgene is being transcribed in the opposite direc-ion in this region. Antisense mRNA could potentiallyesult in decreased translation of the endogenousRNA and hence a decrease in V1a binding.To determine whether the altered V1a receptor ex-
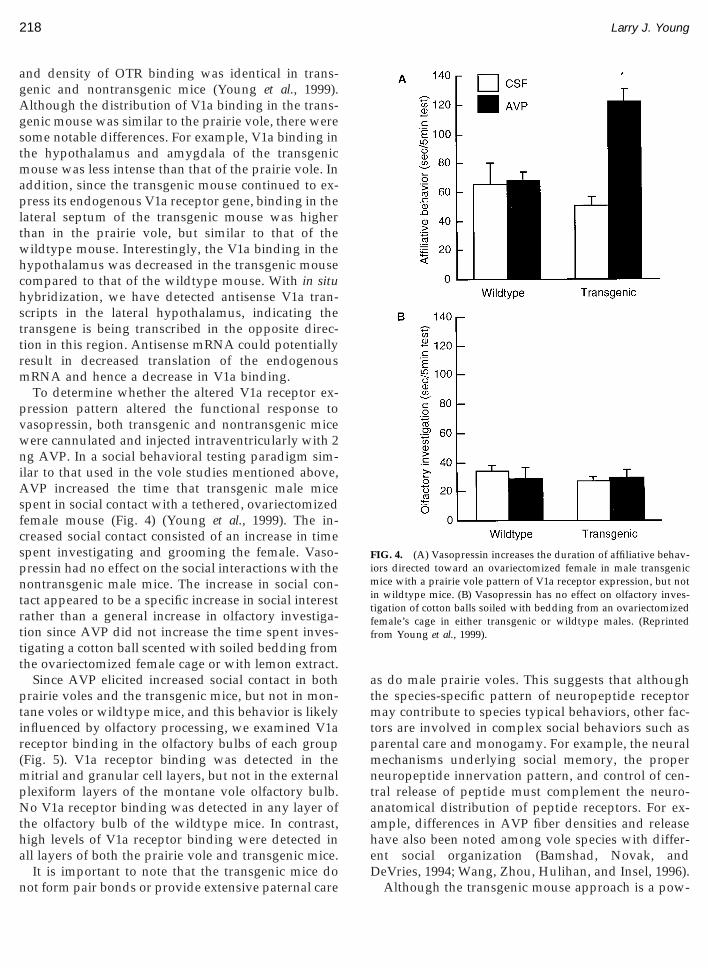
ression pattern altered the functional response toasopressin, both transgenic and nontransgenic miceere cannulated and injected intraventricularly with 2
g AVP. In a social behavioral testing paradigm sim-lar to that used in the vole studies mentioned above,VP increased the time that transgenic male mice
pent in social contact with a tethered, ovariectomizedemale mouse (Fig. 4) (Young et al., 1999). The in-reased social contact consisted of an increase in timepent investigating and grooming the female. Vaso-ressin had no effect on the social interactions with theontransgenic male mice. The increase in social con-
act appeared to be a specific increase in social interestather than a general increase in olfactory investiga-ion since AVP did not increase the time spent inves-igating a cotton ball scented with soiled bedding fromhe ovariectomized female cage or with lemon extract.
Since AVP elicited increased social contact in bothrairie voles and the transgenic mice, but not in mon-
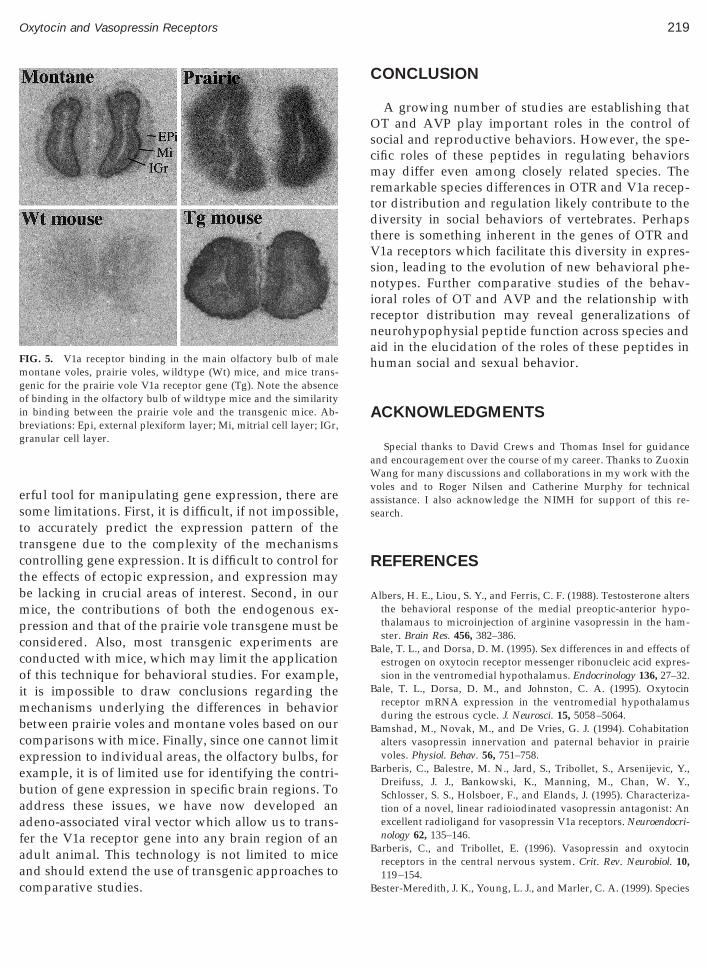
ane voles or wildtype mice, and this behavior is likelynfluenced by olfactory processing, we examined V1aeceptor binding in the olfactory bulbs of each groupFig. 5). V1a receptor binding was detected in the
itrial and granular cell layers, but not in the externallexiform layers of the montane vole olfactory bulb.o V1a receptor binding was detected in any layer of
he olfactory bulb of the wildtype mice. In contrast,igh levels of V1a receptor binding were detected inll layers of both the prairie vole and transgenic mice.
It is important to note that the transgenic mice doot form pair bonds or provide extensive paternal careD
s do male prairie voles. This suggests that althoughhe species-specific pattern of neuropeptide receptor
ay contribute to species typical behaviors, other fac-ors are involved in complex social behaviors such asarental care and monogamy. For example, the neuralechanisms underlying social memory, the proper
europeptide innervation pattern, and control of cen-ral release of peptide must complement the neuro-natomical distribution of peptide receptors. For ex-mple, differences in AVP fiber densities and releaseave also been noted among vole species with differ-nt social organization (Bamshad, Novak, and
IG. 4. (A) Vasopressin increases the duration of affiliative behav-ors directed toward an ovariectomized female in male transgenic
ice with a prairie vole pattern of V1a receptor expression, but notn wildtype mice. (B) Vasopressin has no effect on olfactory inves-igation of cotton balls soiled with bedding from an ovariectomizedemale’s cage in either transgenic or wildtype males. (Reprintedrom Young et al., 1999).
eVries, 1994; Wang, Zhou, Hulihan, and Insel, 1996).Although the transgenic mouse approach is a pow-
esttctbmpccoimbceebaafaac
C
OscmrtdtVsnirnah
A
aWvas
R
A
B
B
B
B
B
Fmgoibg
219Oxytocin and Vasopressin Receptors
rful tool for manipulating gene expression, there areome limitations. First, it is difficult, if not impossible,o accurately predict the expression pattern of theransgene due to the complexity of the mechanismsontrolling gene expression. It is difficult to control forhe effects of ectopic expression, and expression maye lacking in crucial areas of interest. Second, in ourice, the contributions of both the endogenous ex-
ression and that of the prairie vole transgene must beonsidered. Also, most transgenic experiments areonducted with mice, which may limit the applicationf this technique for behavioral studies. For example,
t is impossible to draw conclusions regarding theechanisms underlying the differences in behavior
etween prairie voles and montane voles based on ouromparisons with mice. Finally, since one cannot limitxpression to individual areas, the olfactory bulbs, forxample, it is of limited use for identifying the contri-ution of gene expression in specific brain regions. Toddress these issues, we have now developed andeno-associated viral vector which allow us to trans-er the V1a receptor gene into any brain region of andult animal. This technology is not limited to mice
IG. 5. V1a receptor binding in the main olfactory bulb of maleontane voles, prairie voles, wildtype (Wt) mice, and mice trans-
enic for the prairie vole V1a receptor gene (Tg). Note the absencef binding in the olfactory bulb of wildtype mice and the similarity
n binding between the prairie vole and the transgenic mice. Ab-reviations: Epi, external plexiform layer; Mi, mitrial cell layer; IGr,ranular cell layer.
nd should extend the use of transgenic approaches toomparative studies. B
ONCLUSION
A growing number of studies are establishing thatT and AVP play important roles in the control of
ocial and reproductive behaviors. However, the spe-ific roles of these peptides in regulating behaviorsay differ even among closely related species. The
emarkable species differences in OTR and V1a recep-or distribution and regulation likely contribute to theiversity in social behaviors of vertebrates. Perhaps
here is something inherent in the genes of OTR and1a receptors which facilitate this diversity in expres-
ion, leading to the evolution of new behavioral phe-otypes. Further comparative studies of the behav-
oral roles of OT and AVP and the relationship witheceptor distribution may reveal generalizations ofeurohypophysial peptide function across species andid in the elucidation of the roles of these peptides inuman social and sexual behavior.
CKNOWLEDGMENTS
Special thanks to David Crews and Thomas Insel for guidancend encouragement over the course of my career. Thanks to Zuoxinang for many discussions and collaborations in my work with the
oles and to Roger Nilsen and Catherine Murphy for technicalssistance. I also acknowledge the NIMH for support of this re-earch.
EFERENCES
lbers, H. E., Liou, S. Y., and Ferris, C. F. (1988). Testosterone altersthe behavioral response of the medial preoptic-anterior hypo-thalamaus to microinjection of arginine vasopressin in the ham-ster. Brain Res. 456, 382–386.
ale, T. L., and Dorsa, D. M. (1995). Sex differences in and effects ofestrogen on oxytocin receptor messenger ribonucleic acid expres-sion in the ventromedial hypothalamus. Endocrinology 136, 27–32.
ale, T. L., Dorsa, D. M., and Johnston, C. A. (1995). Oxytocinreceptor mRNA expression in the ventromedial hypothalamusduring the estrous cycle. J. Neurosci. 15, 5058–5064.
amshad, M., Novak, M., and De Vries, G. J. (1994). Cohabitationalters vasopressin innervation and paternal behavior in prairievoles. Physiol. Behav. 56, 751–758.
arberis, C., Balestre, M. N., Jard, S., Tribollet, S., Arsenijevic, Y.,Dreifuss, J. J., Bankowski, K., Manning, M., Chan, W. Y.,Schlosser, S. S., Holsboer, F., and Elands, J. (1995). Characteriza-tion of a novel, linear radioiodinated vasopressin antagonist: Anexcellent radioligand for vasopressin V1a receptors. Neuroendocri-nology 62, 135–146.
arberis, C., and Tribollet, E. (1996). Vasopressin and oxytocinreceptors in the central nervous system. Crit. Rev. Neurobiol. 10,
119–154.ester-Meredith, J. K., Young, L. J., and Marler, C. A. (1999). Species

B
B
C
D
D
D
E
F
F
G
G
G
G
H
I
I
I
J
J
K
M
M
M
M
M
M
N
P
P
P
S
S
S
T
T
V
W
220 Larry J. Young
differences in paternal behavior and aggression in Peromyscusand their associations with vasopressin immunoreactivity andreceptors. Horm. Behav. 36, 25–38.
oyd, S. K. (1994). Arginine vasotocin facilitation of advertisementcalling and call phonotaxis in bullfrogs. Horm. Behav. 28, 232–240.
rownstein, M., Russell, J., and Gainer, H. (1980). Synthesis, trans-port and release of posterior pituitary hormones. Science 207,373–387.
aldwell, J. D., Prange, A. F., and Pedersen, C. A. (1986). Oxytocinfacilitates the sexual receptivity of estrogen-treated female rats.Neuropeptides 7, 175–189.a Costa, A. P. C., Guevara-Guzman, R. G., Ohkura, S., Goode, J. A.,and Kendrick, K. M. (1996). The role of oxytocin release in theparaventricular nucleus in the control of maternal behaviour inthe sheep. J. Neuroendocrinol. 8, 163–177.e Kloet, E. R., Rotteveel, F., Voorhuis, T. A. M., and Terlou, M.(1985). Topography of binding sites for neurohypophyseal hor-mones in rat brain. Eur. J. Pharmacol. 110, 113–119.e Vries, G. J., and Al-Shamma, H. S. (1990). Sex differences inhormonal responses of vasopressin pathways in the rat brain.J. Neurobiol. 21, 686–693.
vans, J. P., and Palmiter, R. D. (1991). Retrotransposition of amouse L1 element. Proc. Natl. Acad. Sci. USA 88, 8792–8795.
erris, C. F., Albers, H. E., Wesolowski, S. M., Goldman, B., andLeeman, S. (1984). Vasopressin injected into the hypothalamustriggers a stereotypic behavior in golden hamsters. Science 224,521–523.
erris, C. F., Melloni, R. H., Koppel, G., Perry, K. W., Fuller, R. W.,and Delville, Y. (1997). Vasopressin/serotonin interactions in theanterior hypothalamus control aggressive behavior in goldenhamsters. J. Neurosci. 17, 4331–4340.ainer, H., and Wray, W. (1994). Cellular and molecular biology ofoxytocin and vasopressin. In E. Knobil and J. D. Neill (Eds.), ThePhysiology of Reproduction, pp. 1099–1129. Raven, New York.oodson, J. (1998a). Territorial aggression and dawn song are mod-ulated by septal vasotocin and vasoactive intestinal polypeptidein male field sparrows (Spizella pusilla). Horm. Behav. 34, 67–77.oodson, J. L. (1998b). Vasotocin and vasoactive intestinal polypep-tide modulate aggression in a territorial songbird, the violet-earedwaxbill (Estrildidae: Uraeginthus granatina). Gen. Comp. Endocrinol.11, 233–244.oodson, J. L., and Adkins-Regan, E. (1998). Effect of intraseptalvasotocin and vasoactive intestinal polypetide infusions on court-ship song and aggression in the male zebra finch (Taeniopygiaguttata). J. Neuroendocrinol. 11, 19–25.o, M. Y., Carter, D. A., Ang, H. L., and Murphy, D. (1995). Bovineoxytocin transgenes in mice. J. Biol. Chem. 270, 27199–27205.
nsel, T. R., and Shapiro, L. E. (1992). Oxytocin receptor distributionreflects social organization in monogamous and polygamousvoles. Proc. Natl. Acad. Sci. 89, 5981–5985.
nsel, T. R., Wang, Z., and Ferris, C. F. (1994). Patterns of brainvasopressin receptor distribution associated with social organiza-tion in microtine rodents. J. Neurosci. 14, 5381–5392.
nsel, T. R., Young, L., Witt, D. M., and Crews, D. (1993). Gonadalsteroids have paradoxical effects on brain oxytocin receptor.J. Neuroendocrinol. 5, 619–628.
ohnson, A. E., Barberis, C., and Albers, H. E. (1995). Castrationreduces vasopressin receptor binding in the hamster hypothala-mus. Brain Res. 674, 153–158.
ohnson, A. E., Coirini, H., Insel, T. R., and McEwen, B. S. (1991). Theregulation of oxytocin receptor binding in the ventromedial hy- W
pothalamic nucleus by testosterone and its metabolites. Endocri-nology 128, 891–896.
endrick, K. M., Keverne, E. B., and Baldwin, B. A. (1987). Intrace-rebroventricular oxytocin stimulates maternal behaviour insheep. Neuroendocrinology 46, 56–61.aney, D., Goode, C., and Wingfield, J. (1997). Intraventricularinfusion of arginine vasotocin induces singing in a female song-bird. J. Neuroendocrinol. 9, 487–491.cCarthy, M. M. (1990). Oxytocin inhibits infanticide in house mice(Mus domesticus). Horm. Behav. 24, 365–375.cCarthy, M. M., Kleopoulos, S. P., Mobbs, C. V., and Pfaff, D. W.(1994). Infusion of antisense oligonucleotides to the oxytocin re-ceptor in the ventromedial hypothalamus reduces estrogen-in-duced sexual receptivity and oxytocin receptor binding in thefemale rat. Neuroendocrinology 59, 432–440.eisel, R. L., and Sachs, B. D. (1994). The physiology of male sexualbehavior. In E. Nobil and J. D. Neill (Eds.), The Physiology ofReproduction, Vol. 2, pp. 3–105. Raven Press, New York.oore, F. L., and Miller, L. J. (1983). Arginine vasotocin inducessexual behavior of newts by acting on cells in the brain. Peptides 4,97–102.oran, J. V., Deberardinis, R. J., and Kazazian, H. H. (1999). Exonshuffling by L1 retrotransposition. Science 283, 1530–1534.ishimori, K., Young, L. J., Guo, Q., Wang, Z., Insel, T. R., andMatzuk, M. M. (1996). Oxytocin is required for nursing but is notessential for partuition or reproductive behavior. Proc. Natl. Acad.Sci. USA 93, 11699–11704.
edersen, C. A., Caldwell, J. D., Walker, C., Ayers, G., and Mason,G. A. (1994). Oxytocin activates the postpartum onset of rat ma-ternal behavior in the ventral tegmental and medial preoptic area.Behav. Neurosci. 108, 1163–1171.
edersen, C. A., and Prange, A. J. (1979). Induction of maternalbehavior in virgin rats after intracerebroventricular administra-tion of oxytocin. Proc. Natl. Acad. Sci. USA 76, 6661–6665.
faff, D. W., and Schwartz-Giblin, S. (1994). Cellular and molecularmechanisms of female reproductive behaviors. In E. Knobil andJ. D. Neill (Eds.), The Physiology of Reproduction, 2nd ed., Vol. 2, pp.107–220. Raven Press, New York.
awrey, D. K., and Dewsbury, D. A. (1985). Control of ovulation,vaginal estrus, and behavioral receptivity in voles (Microtus).Neurosci. Biobehav. Rev. 9, 563–571.
chumacher, M., Coirini, H., Frankfurt, M., and McEwen, B. S.(1989). Localized actions of progesterone in hypothalamus in-volve oxytocin. Proc. Natl. Acad. Sci. USA 86, 6798–6801.
chumacher, M., Coirini, H., Pfaff, D. W., and McEwen, B. S. (1990).Behavioral effects of progesterone associated with rapid modula-tion of oxytocin receptors. Science 250, 691–694.
ribollet, E., Audigier, S., Dubois-Dauphin, M., and Dreifuss, J. J.(1990). Gonadal steroids regulate oxytocin receptors but not va-sopressin receptors in the brain of male and female rats. Anautoradiographical study. Brain Res. 511, 129–140.
ribollet, E., Barberis, C., Jard, S., Dubois-Dauphin, M., and Dreifuss,J. J. (1988). Localization and parmacological characterization of highaffinity binding sites for vasopressin and oxytocin in the rat brain bylight microscopic autoradiography. Br. Res. 442, 105–118.
accari, C., Lolait, S. J., and Ostrowski, N. L. (1998). Comparativedistribution of vasopressin V1b and oxytocin receptor messengerribonucleic acids in brain. Endocrinol. 139, 5015–5033.ang, Z., Ferris, C. F., and De Vries, G. J. (1994). Role of septalvasopressin innervation in paternal behavior in prairie voles (Mi-
crotus ochrogaster). Proc. Natl. Acad. Sci. 91, 400–404.ang, Z., Toloczko, D., Young, L. J., Moody, K., Newman, J. D., and
W
W
W
W
W
Y
Y
Y
Y
Y
221Oxytocin and Vasopressin Receptors
Insel, T. R. (1997). Vasopressin in the forebrain of common mar-mosets (Callithrix jacchus): studies with in situ hybridization, im-munocytochemistry and receptor autoradiography. Brain Res.768, 147–156.ang, Z., Zhou, L., Hulihan, T. J., and Insel, T. R. (1996). Immuno-reactivity of central vasopressin and oxytocin pathways in micro-tine rodents: A quantitative comparative study. J. Comp. Neurol.366, 726–737.illiams, J. R., Insel, T. R., Harbaugh, C. R., and Carter, C. S. (1994).Oxytocin administered centrally facilitates formation of a partnerpreference in prairie voles (Microtus ochrogaster). J. Neuroendocrin.6, 247–250.inslow, J., Hastings, N., Carter, C. S., Harbaugh, C., and Insel, T.(1993). A role for central vasopressin in pair bonding in monog-amous prairie voles. Nature 365, 545–548.itt, D. M., Carter, C. S., and Insel, T. R. (1991). Oxytocin receptorbinding in female pairie voles: Endogenous and exogenous estra-diol stimulation. J. Neuroendocrinol. 3, 155–161.itt, D. M., and Insel, T. R. (1991). A selective oxytocin antagonist
attenuates progesterone facilitation of female sexual behavior.Endocrinology 128, 3269–3276.oung, L. J., Huot, B., Nilsen, R., Wang, Z., and Insel, T. R. (1996).Species differences in central oxytocin receptor gene expression:Comparative analysis of promoter sequences. J. Neuroendocrinol.8, 777–783.
oung, L. J., Nilsen, R., Waymire, K. G., MacGregor, G. R., and Insel,T. R. (1999). Increased affiliative response to vasopressin in miceexpressing the vasopressin receptor from a monogamous vole.Nature 400, 766–768.
oung, L. J., Toloczko, D., and Insel, T. R. (1999). Localization ofvasopressin (V1a) receptor binding and mRNA in the rhesusmonkey brain. J. Neuroendocrinol. 11, 291.
oung, L. J., Waymire, K. G., Nilsen, R., Macgregor, G. R.,Wang, Z., and Insel, T. R. (1996). The 59 flanking region ofthe monogamous prairie vole oxytocin receptor gene directstissue specific expression in mice. Ann. N.Y. Acad. Sci. 807,514 –517.
oung, L. J., Winslow, J. T., Nilsen, R., and Insel, T. R. (1997). Speciesdifferences in V1a receptor gene expression in monogamous and
non-monogamous voles: Behavioral consequences. Behav. Neuro-sci. 111, 599–605.