oxygen and carbon isotopic composition of modern terrestrial gastropod shells from lipari island,...
TRANSCRIPT

Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 119–127
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r .com/ locate /pa laeo
Oxygen and carbon isotopic composition of modern terrestrial gastropodshells from Lipari Island, Aeolian Archipelago (Sicily)
A.C. Colonese a,⁎, G. Zanchetta b,c,d, A.E. Fallick e, G. Manganelli f, P. Lo Cascio g, N. Hausmann a,I. Baneschi c, E. Regattieri b,c
a BioArCh, Department of Archaeology, University of York, Biology S. Block, York YO10 5YW, United Kingdomb Dipartimento di Scienze della Terra, University of Pisa, Via S. Maria, 53, 56126 Pisa, Italyc IGG-CNR, Via Moruzzi 1, 56100 Pisa, Italyd INGV sez. Pisa, Via della Faggiola 32, 56126 Pisa, Italye Scottish Universities Environmental Research Centre, East Kilbride, G75 0QF Glasgow, Scotlandf Dipartimento di Scienze Ambientali, University of Siena, Via P.A. Mattioli 4, 53100 Siena, Italyg Associazione Nesos, Via Vittorio Emanuele n. 24, 98055 Lipari, Italy
⁎ Corresponding author.E-mail addresses: [email protected], andre@p
[email protected] (G. Zanchetta), [email protected]@unisi.it (G. Manganelli), [email protected]@york.ac.uk (N. Hausmann), [email protected]@dst.unipi.it (E. Regattieri).
0031-0182/$ – see front matter © 2013 Elsevier B.V. All rihttp://dx.doi.org/10.1016/j.palaeo.2013.12.003
a b s t r a c t
a r t i c l e i n f oArticle history:Received 26 June 2013Received in revised form 22 November 2013Accepted 3 December 2013Available online 15 December 2013
Keywords:Central MediterraneanLipari IslandTerrestrial gastropod shellsStable isotopesHydrological and environmental proxy
Oxygen (δ18Os) and stable carbon (δ13Cs) isotopic compositions of modern terrestrial gastropod shells fromLipari Island, in the Aeolian Archipelago (Sicily), have been analysed and compared with local meteoric waterδ18O (δ18Op) and vegetation δ13C (δ13Cv) respectively. Results reveal that the δ18Os–δ18Op relationship overthe study area differs from those obtained on continental Europe and Italian shells, implying that even at thescale of the Mediterranean basin different relationships may co-exist. These differences have been interpretedas the increasing influence of Mediterranean vapour water on Tyrrhenian coasts at relatively low altitude(up to ~600 m asl), which compensates for the effect of the δ18Op, and possibly of temperature, on shell δ18O.The steady-state flux balance model (FBM), in agreement with previous studies, suggests that snails are activeprevalently at night. Shell carbon isotope ratios reflect the δ13Cv, as predicted by the metabolic model andrepresent a valuable tool for identifying C3 plants with very distinct isotopic signatures.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
The Mediterranean landscape has been strongly shaped by natural(climatic) and anthropogenic forces at least since the middle Holocene(Blondel and Aronson, 1999). Understanding past evidence of naturalversus human-induced environmental configuration is a challengefor environmental scientists and an essential task for managementand conservation of Mediterranean environments and biodiversity.Palaeoecological records preserved in archaeological archives, althoughdiscontinuous and still under-explored as source of palaeoclimatic in-formation, can offer additional tools for integrating the complex patternof the relation between climate and human activity (Zanchetta et al.,2013). Late Pleistocene and Holocene archaeological deposits fromcentral Mediterranean regions (e.g. Southern Italy and Sicily) containabundant terrestrial gastropod shell remains (Lubell, 2004; Lubelland Barton, 2011), and stable isotope studies (O and C) have provided
alaeo.eu (A.C. Colonese),c.uk (A.E. Fallick),(P. Lo Cascio),.it (I. Baneschi),
ghts reserved.
valuable snapshots on past climate and environment conditions(e.g. Colonese et al., 2010a,b, 2011), parallel to long-termpalaeoclimaticrecords (Zanchetta et al., 2007, 2013; Magny et al., 2011).
These studies were based on empirical observation that atmo-spheric conditions are transmitted to shell oxygen isotopic composi-tion through the 18O/16O ratio of environmental water, itself linked toprecipitation (Heller and Magaritz, 1983; Magaritz and Heller, 1983;Lécolle, 1985; Goodfriend et al., 1987; Goodfriend, 1991; Leng et al.,1998; Goodfriend and Ellis, 2002; Balakrishnan and Yapp, 2004;Zanchetta et al., 2005; Baldini et al., 2007; Yanes et al., 2008, 2012).However the imprinting of atmospheric conditions on the shell oxygenisotope ratio is not straightforward, and distinct regional patterns havebeen observed (Lécolle, 1985; Leone et al., 2000; Zanchetta et al., 2005;Yanes et al., 2009), along with contributions from other variables, suchas the temperature (Lécolle, 1985), relative humidity (Balakrishnanand Yapp, 2004), and other 18O-enriched sources of oxygen (food)(Goodfriend et al., 1987). Isotopic interpretation of fossil assemblagesrequires, therefore, the development of regional baselines, achievedthrough the study of modern analogues, from well-constrained envi-ronmental condition (e.g. Yanes et al., 2008, 2009).
Stable carbon isotopic composition of terrestrial gastropod shellsderives mainly from metabolic CO2 (e.g. from plants and carbonates;DeNiro and Epstein, 1978; Stott, 2002; Metref et al., 2003; ZongXiu

120 A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 119–127
et al., 2007; Yanes et al., 2008) and may depend on feeding behaviour.In environmental studies, 13C/12C has been mainly used for identifyingnot only photosynthetic pathways (e.g. C3, C4, CAM) of consumedvegetation (e.g. Goodfriend and Ellis, 2002; Baldini et al., 2007), butalso differences within the same vegetation photosynthetic category(e.g. Yanes et al., 2011; Stevens et al., 2012; Colonese et al., 2013a,b),although often in a very qualitative way.
The recent growing use of oxygen and carbon isotopic analy-sis on shells of terrestrial gastropods for palaeoclimatic andpalaeoenvironmental reconstructions (Kehrwald et al., 2010; Coloneseet al., 2011, 2013a,b; Yanes et al., 2011, 2012; Paul and Mauldin, 2012;Stevens et al., 2012) requires calibration with modern regional atmo-spheric and environmental conditions. In this study we explore shellisotopic variability in extant terrestrial snails from Lipari Island (AeolianArchipelago, Sicily), a small calc-alkaline volcanic island (Lucchi et al.,2010) at the south-eastern margin of the Tyrrhenian Sea. Precipitationover the study area is dominated by Mediterranean water vapour,thus shell oxygen isotopic compositions of local terrestrial gastropodsprovide the opportunity to investigate this dominant source of precipi-tation on the 18O/16O ratio of their shells in relation to previous studieson other Mediterranean populations (Lécolle, 1985; Zanchetta et al.,2005). Vegetation consists of grasslands and maquis; therefore, wealso investigate their isotopic transmission to the carbon of shells.Assessing regional variability in shell δ18O and δ13C across the Mediter-ranean will certainly improve ecological interpretations of both pastand modern shell assemblages in areas extremely sensitive to climatechange.
2. Environmental setting
The Aeolian volcanic archipelago is located 20 kmoff the NE coast ofSicily and comprises a group of seven islands (Lipari, Vulcano, Salina,Stromboli, Filicudi, Alicudi and Panarea) and small islets (Basiluzzo,Dattilo, Lisca Nera, Bottaro and Lisca Bianca) (Fig. 1). Lipari (37 km2)is the largest island and represents emergent peaks of a volcanic struc-ture up to 602 m above sea level (asl, Monte Chirica), which hasemerged since the Late Quaternary (for detailed geological discussionsee Lucchi et al., 2010). The region exhibits a typical Mediterranean cli-mate, influenced by northward and southwardmigration of subtropicalhigh-pressure cells (Azores high) in summer and winter respectively(Bolle, 2003). As a consequence, air masses from North Africa prevailduring spring and summer, whereas frontal depressions from the
Fig. 1. (A) Geographic position of Aeolian Archipelago (Sicily, Italy) in the central Mediterraneschematic profile of the island (Lucchi et al., 2010).
NorthAtlantic andNorth Europe regions circulate in autumnandwinter.This atmospheric circulation is responsible of a strong seasonality of pre-cipitation,with 75% of the total rainfall occurring fromOctober toMarch(Liotta et al., 2013). In 2009, the meanmonthly temperature at Messina(~40 km SE from Lipari) ranged from 11 °C in winter (February) to28.8 °C in summer (August), the monthly precipitation ranged from4 mm (August) to 309 mm (January), with an annual rainfall amountof 1,255 mm (Fig. 2; data UCEA see later). The mean annual oxygen-isotopic composition of precipitation b100 m asl for the years 2003 to2005 was ~−5.5‰ (δ18Op relative to V-SMOW; Liotta et al., 2006).The δ18Op over Stromboli suggests the absence of a direct contributionof volcanogenic O (Liotta et al., 2006).
Vegetation structure and communities at Lipari are strongly influ-enced by its volcanic origin and by human occupation since theNeolithic.Vegetation is typically Mediterranean, dominated by perennial xericgrasslands (with Hyparrhenia hirta or Brachypodium retusum) andmore or less thermophilous maquis, referred to as Oleo-Ceratonion(with Euphorbia arborescens, Pistacia lentiscus, Calicotome infesta, Genistathyrrena) or Cisto-Ericion (with Erica arborea and Arbustus unedo). Plantcommunities now widely occupy neglected fields, due to the decline ofagriculture which has occurred since the second half of 20th century.Nevertheless, the whole archipelago hosts an outstanding biodiversity:900 plant species, which include some endemics, have been reportedfor the Aeolian Islands, representing 17% of Italian flora (Lo Cascio andNavarra, 2003).
3. Material and methods
3.1. Sampling procedure and preparation for stable isotope analysis
Well-preserved empty shells (n = 85) of 6 species of terrestrial gas-tropods were collected in April 2009 from the ground surface in 6 areasaround the island, at elevations between 2 and 560 m (Fig. 1B; Table 1).Vegetation associated with the shells was also collected for isotopicanalysis (Table 2). Collected shells belong to species widespread atLipari and at other islands of the archipelago (Giusti, 1973). Snail speciesare herbivorous and opportunistically omnivorous in the westernMediterranean: Trochoidea pyramidata and Mastus pupa are thermo-philic and xeroresistant species typically inhabiting coastal and inlandareas (Giusti et al., 1995; Manganelli et al., 1995); at Lipari they werecollected from shrubs and perennial–annual grassland habitats, com-posed of Carpobrotus acinaciformis, Limbarda crithmoides, Pelargonium
an; Messina meteorological station (star). (B) Detail of Lipari and sampling areas and the
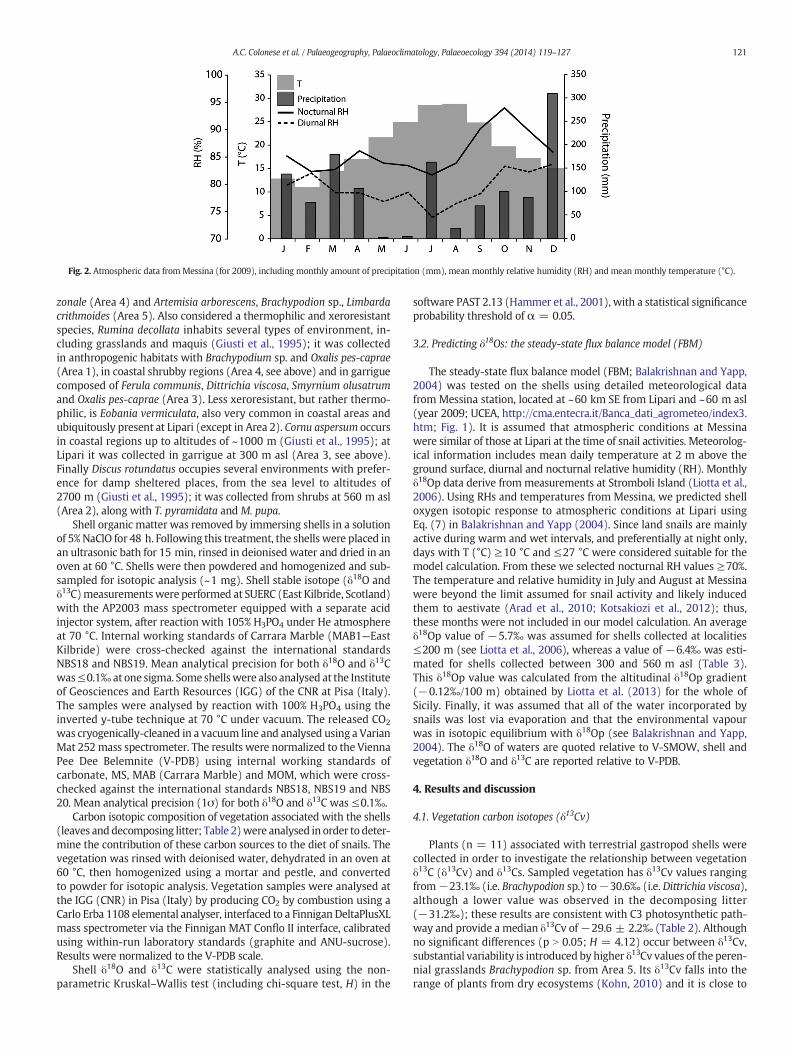
Fig. 2. Atmospheric data from Messina (for 2009), including monthly amount of precipitation (mm), mean monthly relative humidity (RH) and mean monthly temperature (°C).
121A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 119–127
zonale (Area 4) and Artemisia arborescens, Brachypodion sp., Limbardacrithmoides (Area 5). Also considered a thermophilic and xeroresistantspecies, Rumina decollata inhabits several types of environment, in-cluding grasslands and maquis (Giusti et al., 1995); it was collectedin anthropogenic habitats with Brachypodium sp. and Oxalis pes-caprae(Area 1), in coastal shrubby regions (Area 4, see above) and in garriguecomposed of Ferula communis, Dittrichia viscosa, Smyrnium olusatrumand Oxalis pes-caprae (Area 3). Less xeroresistant, but rather thermo-philic, is Eobania vermiculata, also very common in coastal areas andubiquitously present at Lipari (except in Area 2). Cornu aspersum occursin coastal regions up to altitudes of ~1000 m (Giusti et al., 1995); atLipari it was collected in garrigue at 300 m asl (Area 3, see above).Finally Discus rotundatus occupies several environments with prefer-ence for damp sheltered places, from the sea level to altitudes of2700 m (Giusti et al., 1995); it was collected from shrubs at 560 m asl(Area 2), along with T. pyramidata and M. pupa.
Shell organic matter was removed by immersing shells in a solutionof 5% NaClO for 48 h. Following this treatment, the shells were placed inan ultrasonic bath for 15 min, rinsed in deionised water and dried in anoven at 60 °C. Shells were then powdered and homogenized and sub-sampled for isotopic analysis (~1 mg). Shell stable isotope (δ18O andδ13C)measurementswere performed at SUERC (East Kilbride, Scotland)with the AP2003 mass spectrometer equipped with a separate acidinjector system, after reaction with 105% H3PO4 under He atmosphereat 70 °C. Internal working standards of Carrara Marble (MAB1—EastKilbride) were cross-checked against the international standardsNBS18 and NBS19. Mean analytical precision for both δ18O and δ13Cwas≤0.1‰ at one sigma. Some shellswere also analysed at the Instituteof Geosciences and Earth Resources (IGG) of the CNR at Pisa (Italy).The samples were analysed by reaction with 100% H3PO4 using theinverted y-tube technique at 70 °C under vacuum. The released CO2
was cryogenically-cleaned in a vacuum line and analysed using a VarianMat 252 mass spectrometer. The results were normalized to the ViennaPee Dee Belemnite (V-PDB) using internal working standards ofcarbonate, MS, MAB (Carrara Marble) and MOM, which were cross-checked against the international standards NBS18, NBS19 and NBS20. Mean analytical precision (1σ) for both δ18O and δ13C was ≤0.1‰.
Carbon isotopic composition of vegetation associated with the shells(leaves and decomposing litter; Table 2)were analysed in order to deter-mine the contribution of these carbon sources to the diet of snails. Thevegetation was rinsed with deionised water, dehydrated in an oven at60 °C, then homogenized using a mortar and pestle, and convertedto powder for isotopic analysis. Vegetation samples were analysed atthe IGG (CNR) in Pisa (Italy) by producing CO2 by combustion using aCarlo Erba 1108 elemental analyser, interfaced to a Finnigan DeltaPlusXLmass spectrometer via the Finnigan MAT Conflo II interface, calibratedusing within-run laboratory standards (graphite and ANU-sucrose).Results were normalized to the V-PDB scale.
Shell δ18O and δ13C were statistically analysed using the non-parametric Kruskal–Wallis test (including chi-square test, H) in the
software PAST 2.13 (Hammer et al., 2001), with a statistical significanceprobability threshold of α = 0.05.
3.2. Predicting δ18Os: the steady-state flux balance model (FBM)
The steady-state flux balance model (FBM; Balakrishnan and Yapp,2004) was tested on the shells using detailed meteorological datafrom Messina station, located at ~60 km SE from Lipari and ~60 m asl(year 2009; UCEA, http://cma.entecra.it/Banca_dati_agrometeo/index3.htm; Fig. 1). It is assumed that atmospheric conditions at Messinawere similar of those at Lipari at the time of snail activities. Meteorolog-ical information includes mean daily temperature at 2 m above theground surface, diurnal and nocturnal relative humidity (RH). Monthlyδ18Op data derive from measurements at Stromboli Island (Liotta et al.,2006). Using RHs and temperatures from Messina, we predicted shelloxygen isotopic response to atmospheric conditions at Lipari usingEq. (7) in Balakrishnan and Yapp (2004). Since land snails are mainlyactive during warm and wet intervals, and preferentially at night only,days with T (°C) ≥10 °C and ≤27 °C were considered suitable for themodel calculation. From these we selected nocturnal RH values ≥70%.The temperature and relative humidity in July and August at Messinawere beyond the limit assumed for snail activity and likely inducedthem to aestivate (Arad et al., 2010; Kotsakiozi et al., 2012); thus,these months were not included in our model calculation. An averageδ18Op value of −5.7‰ was assumed for shells collected at localities≤200 m (see Liotta et al., 2006), whereas a value of −6.4‰ was esti-mated for shells collected between 300 and 560 m asl (Table 3).This δ18Op value was calculated from the altitudinal δ18Op gradient(−0.12‰/100 m) obtained by Liotta et al. (2013) for the whole ofSicily. Finally, it was assumed that all of the water incorporated bysnails was lost via evaporation and that the environmental vapourwas in isotopic equilibrium with δ18Op (see Balakrishnan and Yapp,2004). The δ18O of waters are quoted relative to V-SMOW, shell andvegetation δ18O and δ13C are reported relative to V-PDB.
4. Results and discussion
4.1. Vegetation carbon isotopes (δ13Cv)
Plants (n = 11) associated with terrestrial gastropod shells werecollected in order to investigate the relationship between vegetationδ13C (δ13Cv) and δ13Cs. Sampled vegetation has δ13Cv values rangingfrom−23.1‰ (i.e. Brachypodion sp.) to−30.6‰ (i.e. Dittrichia viscosa),although a lower value was observed in the decomposing litter(−31.2‰); these results are consistent with C3 photosynthetic path-way and provide a median δ13Cv of−29.6 ± 2.2‰ (Table 2). Althoughno significant differences (p N 0.05; H = 4.12) occur between δ13Cv,substantial variability is introduced by higher δ13Cv values of the peren-nial grasslands Brachypodion sp. from Area 5. Its δ13Cv falls into therange of plants from dry ecosystems (Kohn, 2010) and it is close to

Table 1Land snail species, geographical setting of the sample localities, shell δ18O and δ13C and vegetation structure.
Species Alt(m a.s.l.)
Area δ18Os‰ δ13Cs‰ Median δ18Os‰ Median δ13Cs‰ Habitat
Eobania vermiculata (Müller, 1774) 2 4 0.1 −11.3 Coastal shrubEobania vermiculata (Müller, 1774) 2 4 −0.6 −14.3 Coastal shrubEobania vermiculata (Müller, 1774) 2 4 −1.0 −12.2 Coastal shrubEobania vermiculata (Müller, 1774) 2 4 −0.5 −10.8 Coastal shrubEobania vermiculata (Müller, 1774) 2 4 −0.2 −11.1 Coastal shrubEobania vermiculata (Müller, 1774) 2 4 −1.0 −13.2 Coastal shrubEobania vermiculata (Müller, 1774) 2 4 0.0 −13.5 Coastal shrubEobania vermiculata (Müller, 1774) 2 4 0.4 −9.1 Coastal shrubEobania vermiculata (Müller, 1774) 2 4 −0.6 −12.4 Coastal shrubEobania vermiculata (Müller, 1774) 2 4 −0.2 −9.0 Coastal shrubEobania vermiculata (Müller, 1774) 2 4 −0.6 −9.0 Coastal shrubEobania vermiculata (Müller, 1774) 2 4 0.5 −11.3 Coastal shrubRumina decollata (Linnaeus, 1758) 2 4 −2.1 −12.0 Coastal shrubRumina decollata (Linnaeus, 1758) 2 4 −2.1 −12.9 Coastal shrubRumina decollata (Linnaeus, 1758) 2 4 −1.6 −12.0 Coastal shrubRumina decollata (Linnaeus, 1758) 2 4 −1.9 −11.9 Coastal shrubRumina decollata (Linnaeus, 1758) 2 4 −2.6 −11.8 Coastal shrubTrochoidea pyramidata (Draparnaud, 1805) 2 4 −1.2 −12.4 Coastal shrubTrochoidea pyramidata (Draparnaud, 1805) 2 4 −0.4 −12.2 Coastal shrubTrochoidea pyramidata (Draparnaud, 1805) 2 4 −0.8 −10.6 −0.6 ± 0.9 −12.0 ± 1.4 Coastal shrubEobania vermiculata (Müller, 1774) 10 1 −0.1 −11.6 Synanthropic (city centre)Eobania vermiculata (Müller, 1774) 10 1 −0.9 −12.3 Synanthropic (city centre)Eobania vermiculata (Müller, 1774) 10 1 −0.6 −12.8 Synanthropic (city centre)Eobania vermiculata (Müller, 1774) 10 1 −0.8 −11.2 Synanthropic (city centre)Eobania vermiculata (Müller, 1774) 10 1 −1.0 −11.9 Synanthropic (city centre)Eobania vermiculata (Müller, 1774) 10 1 −1.0 −12.0 Synanthropic (city centre)Eobania vermiculata (Müller, 1774) 10 1 0.3 −9.3 Synanthropic (city centre)Eobania vermiculata (Müller, 1774) 10 1 0.0 −13.3 Synanthropic (city centre)Eobania vermiculata (Müller, 1774) 10 1 −0.8 −8.7 Synanthropic (city centr)Rumina decollata (Linnaeus, 1758) 10 1 −0.8 −12.8 Synanthropic (city centre)Rumina decollata (Linnaeus, 1758) 10 1 −1.4 −10.7 Synanthropic (city centre)Rumina decollata (Linnaeus, 1758) 10 1 −1.6 −12.0 Synanthropic (city centre)Rumina decollata (Linnaeus, 1758) 10 1 −0.1 −12.0 Synanthropic (city centre)Rumina decollata (Linnaeus, 1758) 10 1 −1.0 −11.0 Synanthropic (city centre)Rumina decollata (Linnaeus, 1758) 10 1 −1.0 −10.8 Synanthropic (city centre)Rumina decollata (Linnaeus, 1758) 10 1 −1.1 −12.0 Synanthropic (city centre)Rumina decollata (Linnaeus, 1758) 10 1 −1.7 −11.7 Synanthropic (city centre)Rumina decollata (Linnaeus, 1758) 10 1 −0.8 −12.8 −0.9 ± 0.5 −11.9 ± 1.2 Synanthropic (city centre)Eobania vermiculata (Müller, 1774) 50 5 −1.3 −8.5 Coastal perennial–annual grassesEobania vermiculata (Müller, 1774) 50 5 −0.1 −7.7 Coastal perennial–annual grassesEobania vermiculata (Müller, 1774) 50 5 0.3 −9.4 Coastal perennial–annual grassesEobania vermiculata (Müller, 1774) 50 5 −1.2 −8.6 Coastal perennial–annual grassesEobania vermiculata (Müller, 1774) 50 5 −1.4 −9.2 Coastal perennial–annual grassesEobania vermiculata (Müller, 1774) 50 5 −1.0 −9.4 Coastal perennial–annual grassesEobania vermiculata (Müller, 1774) 50 5 −0.5 −8.8 Coastal perennial–annual grassesMastus pupa (Linnaeus, 1758) 50 5 −1.8 −10.1 Coastal perennial–annual grassesMastus pupa (Linnaeus, 1758) 50 5 −0.9 −8.7 Coastal perennial–annual grassesMastus pupa (Linnaeus, 1758) 50 5 −0.7 −7.2 Coastal perennial–annual grassesTrochoidea pyramidata (Draparnaud, 1805) 50 5 −0.7 −8.2 Coastal perennial–annual grassesTrochoidea pyramidata (Draparnaud, 1805) 50 5 −1.0 −7.1 Coastal perennial–annual grassesTrochoidea pyramidata (Draparnaud, 1805) 50 5 −1.6 −8.7 Coastal perennial–annual grassesTrochoidea pyramidata (Draparnaud, 1805) 50 5 −0.7 −8.1 Coastal perennial–annual grassesTrochoidea pyramidata (Draparnaud, 1805) 50 5 −0.9 −6.9 Coastal perennial–annual grassesTrochoidea pyramidata (Draparnaud, 1805) 50 5 −1.4 −5.1 −1.0 ± 0.5 −8.6 ± 1.2 Coastal perennial–annual grassesEobania vermiculata (Müller, 1774) 200 6 0.1 −13.9 Coastal perennial grasses and shrubsEobania vermiculata (Müller, 1774) 200 6 −0.8 −11.7 Coastal perennial grasses and shrubsEobania vermiculata (Müller, 1774) 200 6 −0.8 −12.7 Coastal perennial grasses and shrubsEobania vermiculata (Müller, 1774) 200 6 0.0 −12.1 Coastal perennial grasses and shrubsEobania vermiculata (Müller, 1774) 200 6 −0.9 −12.5 −0.8 ± 0.5 −12.5 ± 0.9 Coastal perennial grasses and shrubsCornu aspersum (Müller, 1774) 300 3 −1.4 −12.2 GarrigueCornu aspersum (Müller, 1774) 300 3 −1.8 −11.9 GarrigueCornu aspersum (Müller, 1774) 300 3 −1.6 −12.1 GarrigueCornu aspersum (Müller, 1774) 300 3 −1.3 −11.7 GarrigueEobania vermiculata (Müller, 1774) 300 3 −0.7 −10.4 GarrigueEobania vermiculata (Müller, 1774) 300 3 −0.3 −11.1 GarrigueRumina decollata (Linnaeus, 1758) 300 3 −0.7 −10.0 GarrigueRumina decollata (Linnaeus, 1758) 300 3 −1.0 −12.0 GarrigueRumina decollata (Linnaeus, 1758) 300 3 −2.1 −12.2 GarrigueRumina decollata (Linnaeus, 1758) 300 3 −1.0 −10.5 GarrigueRumina decollata (Linnaeus, 1758) 300 3 −2.4 −12.1 −1.3 ± 0.6 −11.9 ± 0.8 GarrigueDiscus rotundatus (Müller, 1774) 560 2 −1.1 −10.8 ShrubsDiscus rotundatus (Müller, 1774) 560 2 −1.1 −10.2 ShrubsDiscus rotundatus (Müller, 1774) 560 2 −1.4 −10.2 ShrubsDiscus rotundatus (Müller, 1774) 560 2 −2.3 −10.0 Shrubs
122 A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 119–127
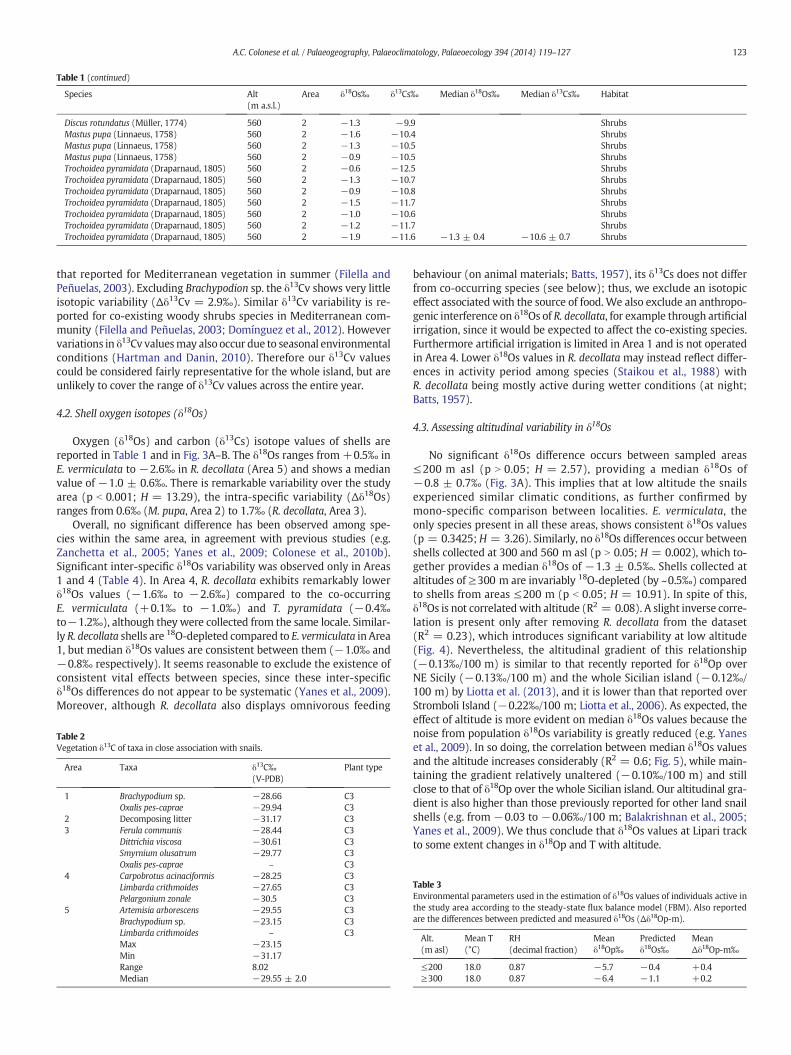
Table 1 (continued)
Species Alt(m a.s.l.)
Area δ18Os‰ δ13Cs‰ Median δ18Os‰ Median δ13Cs‰ Habitat
Discus rotundatus (Müller, 1774) 560 2 −1.3 −9.9 ShrubsMastus pupa (Linnaeus, 1758) 560 2 −1.6 −10.4 ShrubsMastus pupa (Linnaeus, 1758) 560 2 −1.3 −10.5 ShrubsMastus pupa (Linnaeus, 1758) 560 2 −0.9 −10.5 ShrubsTrochoidea pyramidata (Draparnaud, 1805) 560 2 −0.6 −12.5 ShrubsTrochoidea pyramidata (Draparnaud, 1805) 560 2 −1.3 −10.7 ShrubsTrochoidea pyramidata (Draparnaud, 1805) 560 2 −0.9 −10.8 ShrubsTrochoidea pyramidata (Draparnaud, 1805) 560 2 −1.5 −11.7 ShrubsTrochoidea pyramidata (Draparnaud, 1805) 560 2 −1.0 −10.6 ShrubsTrochoidea pyramidata (Draparnaud, 1805) 560 2 −1.2 −11.7 ShrubsTrochoidea pyramidata (Draparnaud, 1805) 560 2 −1.9 −11.6 −1.3 ± 0.4 −10.6 ± 0.7 Shrubs
123A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 119–127
that reported for Mediterranean vegetation in summer (Filella andPeñuelas, 2003). Excluding Brachypodion sp. the δ13Cv shows very littleisotopic variability (Δδ13Cv = 2.9‰). Similar δ13Cv variability is re-ported for co-existing woody shrubs species in Mediterranean com-munity (Filella and Peñuelas, 2003; Domínguez et al., 2012). Howevervariations in δ13Cv valuesmay also occur due to seasonal environmentalconditions (Hartman and Danin, 2010). Therefore our δ13Cv valuescould be considered fairly representative for the whole island, but areunlikely to cover the range of δ13Cv values across the entire year.
4.2. Shell oxygen isotopes (δ18Os)
Oxygen (δ18Os) and carbon (δ13Cs) isotope values of shells arereported in Table 1 and in Fig. 3A–B. The δ18Os ranges from +0.5‰ inE. vermiculata to −2.6‰ in R. decollata (Area 5) and shows a medianvalue of −1.0 ± 0.6‰. There is remarkable variability over the studyarea (p b 0.001; H = 13.29), the intra-specific variability (Δδ18Os)ranges from 0.6‰ (M. pupa, Area 2) to 1.7‰ (R. decollata, Area 3).
Overall, no significant difference has been observed among spe-cies within the same area, in agreement with previous studies (e.g.Zanchetta et al., 2005; Yanes et al., 2009; Colonese et al., 2010b).Significant inter-specific δ18Os variability was observed only in Areas1 and 4 (Table 4). In Area 4, R. decollata exhibits remarkably lowerδ18Os values (−1.6‰ to −2.6‰) compared to the co-occurringE. vermiculata (+0.1‰ to −1.0‰) and T. pyramidata (−0.4‰to−1.2‰), although they were collected from the same locale. Similar-ly R. decollata shells are 18O-depleted compared to E. vermiculata in Area1, but median δ18Os values are consistent between them (−1.0‰ and−0.8‰ respectively). It seems reasonable to exclude the existence ofconsistent vital effects between species, since these inter-specificδ18Os differences do not appear to be systematic (Yanes et al., 2009).Moreover, although R. decollata also displays omnivorous feeding
Table 2Vegetation δ13C of taxa in close association with snails.
Area Taxa δ13C‰(V-PDB)
Plant type
1 Brachypodium sp. −28.66 C3Oxalis pes-caprae −29.94 C3
2 Decomposing litter −31.17 C33 Ferula communis −28.44 C3
Dittrichia viscosa −30.61 C3Smyrnium olusatrum −29.77 C3Oxalis pes-caprae – C3
4 Carpobrotus acinaciformis −28.25 C3Limbarda crithmoides −27.65 C3Pelargonium zonale −30.5 C3
5 Artemisia arborescens −29.55 C3Brachypodium sp. −23.15 C3Limbarda crithmoides – C3Max −23.15Min −31.17Range 8.02Median −29.55 ± 2.0
behaviour (on animal materials; Batts, 1957), its δ13Cs does not differfrom co-occurring species (see below); thus, we exclude an isotopiceffect associated with the source of food. We also exclude an anthropo-genic interference on δ18Os of R. decollata, for example through artificialirrigation, since it would be expected to affect the co-existing species.Furthermore artificial irrigation is limited in Area 1 and is not operatedin Area 4. Lower δ18Os values in R. decollata may instead reflect differ-ences in activity period among species (Staikou et al., 1988) withR. decollata being mostly active during wetter conditions (at night;Batts, 1957).
4.3. Assessing altitudinal variability in δ18Os
No significant δ18Os difference occurs between sampled areas≤200 m asl (p N 0.05; H = 2.57), providing a median δ18Os of−0.8 ± 0.7‰ (Fig. 3A). This implies that at low altitude the snailsexperienced similar climatic conditions, as further confirmed bymono-specific comparison between localities. E. vermiculata, theonly species present in all these areas, shows consistent δ18Os values(p = 0.3425; H = 3.26). Similarly, no δ18Os differences occur betweenshells collected at 300 and 560 m asl (p N 0.05; H = 0.002), which to-gether provides a median δ18Os of −1.3 ± 0.5‰. Shells collected ataltitudes of≥300 m are invariably 18O-depleted (by ~0.5‰) comparedto shells from areas ≤200 m (p b 0.05; H = 10.91). In spite of this,δ18Os is not correlatedwith altitude (R2 = 0.08). A slight inverse corre-lation is present only after removing R. decollata from the dataset(R2 = 0.23), which introduces significant variability at low altitude(Fig. 4). Nevertheless, the altitudinal gradient of this relationship(−0.13‰/100 m) is similar to that recently reported for δ18Op overNE Sicily (−0.13‰/100 m) and the whole Sicilian island (−0.12‰/100 m) by Liotta et al. (2013), and it is lower than that reported overStromboli Island (−0.22‰/100 m; Liotta et al., 2006). As expected, theeffect of altitude is more evident on median δ18Os values because thenoise from population δ18Os variability is greatly reduced (e.g. Yaneset al., 2009). In so doing, the correlation between median δ18Os valuesand the altitude increases considerably (R2 = 0.6; Fig. 5), while main-taining the gradient relatively unaltered (−0.10‰/100 m) and stillclose to that of δ18Op over the whole Sicilian island. Our altitudinal gra-dient is also higher than those previously reported for other land snailshells (e.g. from −0.03 to −0.06‰/100 m; Balakrishnan et al., 2005;Yanes et al., 2009). We thus conclude that δ18Os values at Lipari trackto some extent changes in δ18Op and T with altitude.
Table 3Environmental parameters used in the estimation of δ18Os values of individuals active inthe study area according to the steady-state flux balance model (FBM). Also reportedare the differences between predicted and measured δ18Os (Δδ18Op-m).
Alt.(m asl)
Mean T(°C)
RH(decimal fraction)
Meanδ18Op‰
Predictedδ18Os‰
MeanΔδ18Op-m‰
≤200 18.0 0.87 −5.7 −0.4 +0.4≥300 18.0 0.87 −6.4 −1.1 +0.2
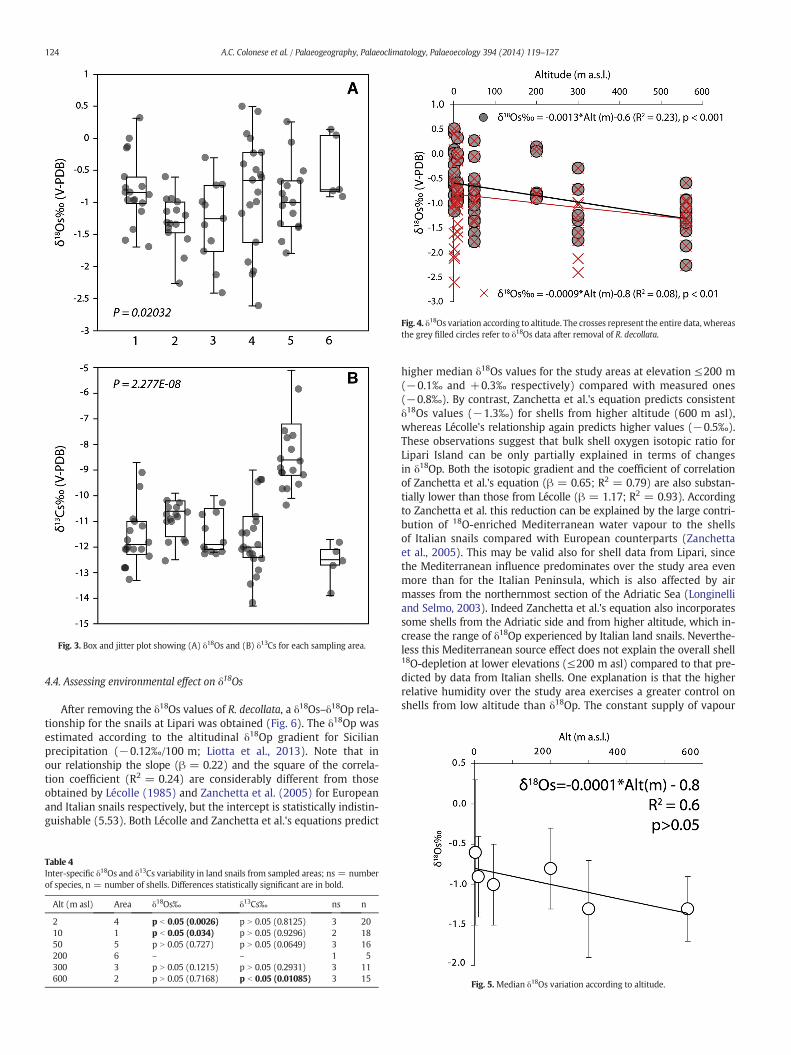
Fig. 3. Box and jitter plot showing (A) δ18Os and (B) δ13Cs for each sampling area.
Fig. 4. δ18Os variation according to altitude. The crosses represent the entire data, whereasthe grey filled circles refer to δ18Os data after removal of R. decollata.
124 A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 119–127
4.4. Assessing environmental effect on δ18Os
After removing the δ18Os values of R. decollata, a δ18Os–δ18Op rela-tionship for the snails at Lipari was obtained (Fig. 6). The δ18Op wasestimated according to the altitudinal δ18Op gradient for Sicilianprecipitation (−0.12‰/100 m; Liotta et al., 2013). Note that inour relationship the slope (β = 0.22) and the square of the correla-tion coefficient (R2 = 0.24) are considerably different from thoseobtained by Lécolle (1985) and Zanchetta et al. (2005) for Europeanand Italian snails respectively, but the intercept is statistically indistin-guishable (5.53). Both Lécolle and Zanchetta et al.'s equations predict
Table 4Inter-specific δ18Os and δ13Cs variability in land snails from sampled areas; ns = numberof species, n = number of shells. Differences statistically significant are in bold.
Alt (m asl) Area δ18Os‰ δ13Cs‰ ns n
2 4 p b 0.05 (0.0026) p N 0.05 (0.8125) 3 2010 1 p b 0.05 (0.034) p N 0.05 (0.9296) 2 1850 5 p N 0.05 (0.727) p N 0.05 (0.0649) 3 16200 6 – – 1 5300 3 p N 0.05 (0.1215) p N 0.05 (0.2931) 3 11600 2 p N 0.05 (0.7168) p b 0.05 (0.01085) 3 15
higher median δ18Os values for the study areas at elevation ≤200 m(−0.1‰ and +0.3‰ respectively) compared with measured ones(−0.8‰). By contrast, Zanchetta et al.'s equation predicts consistentδ18Os values (−1.3‰) for shells from higher altitude (600 m asl),whereas Lécolle's relationship again predicts higher values (−0.5‰).These observations suggest that bulk shell oxygen isotopic ratio forLipari Island can be only partially explained in terms of changesin δ18Op. Both the isotopic gradient and the coefficient of correlationof Zanchetta et al.'s equation (β = 0.65; R2 = 0.79) are also substan-tially lower than those from Lécolle (β = 1.17; R2 = 0.93). Accordingto Zanchetta et al. this reduction can be explained by the large contri-bution of 18O-enriched Mediterranean water vapour to the shellsof Italian snails compared with European counterparts (Zanchettaet al., 2005). This may be valid also for shell data from Lipari, sincethe Mediterranean influence predominates over the study area evenmore than for the Italian Peninsula, which is also affected by airmasses from the northernmost section of the Adriatic Sea (Longinelliand Selmo, 2003). Indeed Zanchetta et al.'s equation also incorporatessome shells from the Adriatic side and from higher altitude, which in-crease the range of δ18Op experienced by Italian land snails. Neverthe-less this Mediterranean source effect does not explain the overall shell18O-depletion at lower elevations (≤200 m asl) compared to that pre-dicted by data from Italian shells. One explanation is that the higherrelative humidity over the study area exercises a greater control onshells from low altitude than δ18Op. The constant supply of vapour
Fig. 5.Median δ18Os variation according to altitude.
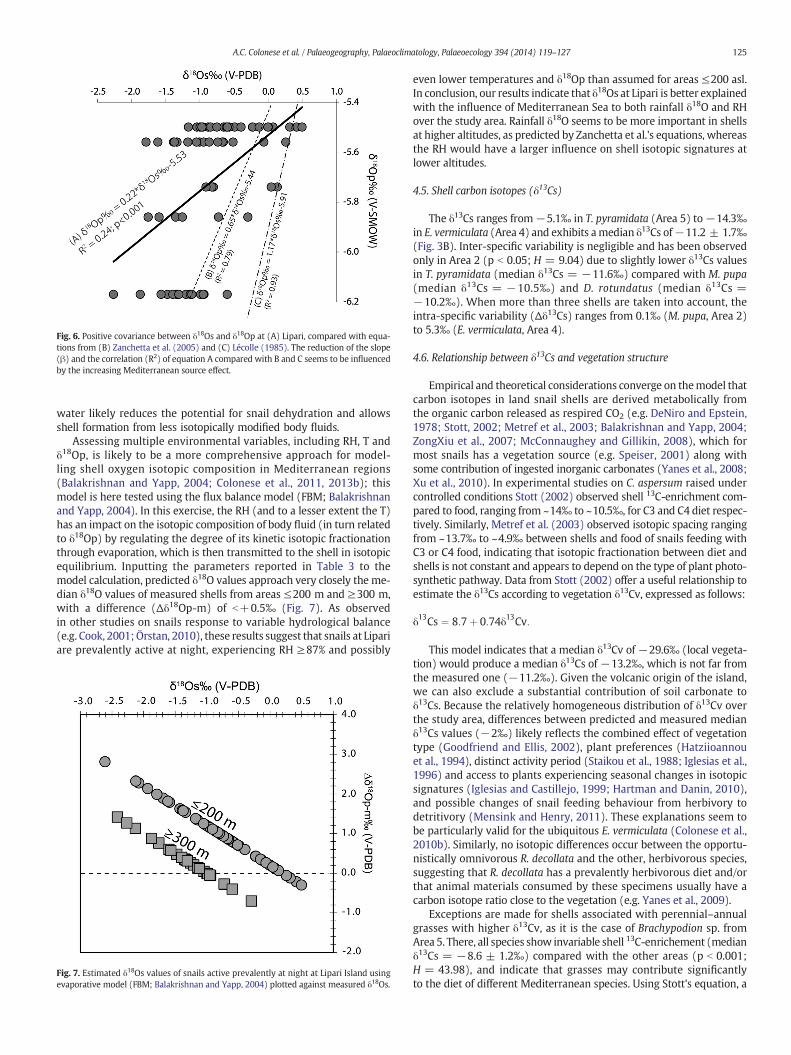
Fig. 6. Positive covariance between δ18Os and δ18Op at (A) Lipari, compared with equa-tions from (B) Zanchetta et al. (2005) and (C) Lécolle (1985). The reduction of the slope(β) and the correlation (R2) of equation A compared with B and C seems to be influencedby the increasing Mediterranean source effect.
125A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 119–127
water likely reduces the potential for snail dehydration and allowsshell formation from less isotopically modified body fluids.
Assessing multiple environmental variables, including RH, T andδ18Op, is likely to be a more comprehensive approach for model-ling shell oxygen isotopic composition in Mediterranean regions(Balakrishnan and Yapp, 2004; Colonese et al., 2011, 2013b); thismodel is here tested using the flux balance model (FBM; Balakrishnanand Yapp, 2004). In this exercise, the RH (and to a lesser extent the T)has an impact on the isotopic composition of body fluid (in turn relatedto δ18Op) by regulating the degree of its kinetic isotopic fractionationthrough evaporation, which is then transmitted to the shell in isotopicequilibrium. Inputting the parameters reported in Table 3 to themodel calculation, predicted δ18O values approach very closely the me-dian δ18O values of measured shells from areas ≤200 m and ≥300 m,with a difference (Δδ18Op-m) of b+0.5‰ (Fig. 7). As observedin other studies on snails response to variable hydrological balance(e.g. Cook, 2001;Örstan, 2010), these results suggest that snails at Lipariare prevalently active at night, experiencing RH ≥87% and possibly
Fig. 7. Estimated δ18Os values of snails active prevalently at night at Lipari Island usingevaporative model (FBM; Balakrishnan and Yapp, 2004) plotted against measured δ18Os.
even lower temperatures and δ18Op than assumed for areas ≤200 asl.In conclusion, our results indicate that δ18Os at Lipari is better explainedwith the influence of Mediterranean Sea to both rainfall δ18O and RHover the study area. Rainfall δ18O seems to be more important in shellsat higher altitudes, as predicted by Zanchetta et al.'s equations, whereasthe RH would have a larger influence on shell isotopic signatures atlower altitudes.
4.5. Shell carbon isotopes (δ13Cs)
The δ13Cs ranges from−5.1‰ in T. pyramidata (Area 5) to−14.3‰in E. vermiculata (Area 4) and exhibits amedian δ13Cs of−11.2 ± 1.7‰(Fig. 3B). Inter-specific variability is negligible and has been observedonly in Area 2 (p b 0.05; H = 9.04) due to slightly lower δ13Cs valuesin T. pyramidata (median δ13Cs = −11.6‰) compared with M. pupa(median δ13Cs = −10.5‰) and D. rotundatus (median δ13Cs =−10.2‰). When more than three shells are taken into account, theintra-specific variability (Δδ13Cs) ranges from 0.1‰ (M. pupa, Area 2)to 5.3‰ (E. vermiculata, Area 4).
4.6. Relationship between δ13Cs and vegetation structure
Empirical and theoretical considerations converge on themodel thatcarbon isotopes in land snail shells are derived metabolically fromthe organic carbon released as respired CO2 (e.g. DeNiro and Epstein,1978; Stott, 2002; Metref et al., 2003; Balakrishnan and Yapp, 2004;ZongXiu et al., 2007; McConnaughey and Gillikin, 2008), which formost snails has a vegetation source (e.g. Speiser, 2001) along withsome contribution of ingested inorganic carbonates (Yanes et al., 2008;Xu et al., 2010). In experimental studies on C. aspersum raised undercontrolled conditions Stott (2002) observed shell 13C-enrichment com-pared to food, ranging from ~14‰ to ~10.5‰, for C3 and C4 diet respec-tively. Similarly, Metref et al. (2003) observed isotopic spacing rangingfrom ~13.7‰ to ~4.9‰ between shells and food of snails feeding withC3 or C4 food, indicating that isotopic fractionation between diet andshells is not constant and appears to depend on the type of plant photo-synthetic pathway. Data from Stott (2002) offer a useful relationship toestimate the δ13Cs according to vegetation δ13Cv, expressed as follows:
δ13Cs ¼ 8:7þ 0:74δ13Cv:
This model indicates that a median δ13Cv of −29.6‰ (local vegeta-tion) would produce a median δ13Cs of −13.2‰, which is not far fromthe measured one (−11.2‰). Given the volcanic origin of the island,we can also exclude a substantial contribution of soil carbonate toδ13Cs. Because the relatively homogeneous distribution of δ13Cv overthe study area, differences between predicted and measured medianδ13Cs values (−2‰) likely reflects the combined effect of vegetationtype (Goodfriend and Ellis, 2002), plant preferences (Hatziioannouet al., 1994), distinct activity period (Staikou et al., 1988; Iglesias et al.,1996) and access to plants experiencing seasonal changes in isotopicsignatures (Iglesias and Castillejo, 1999; Hartman and Danin, 2010),and possible changes of snail feeding behaviour from herbivory todetritivory (Mensink and Henry, 2011). These explanations seem tobe particularly valid for the ubiquitous E. vermiculata (Colonese et al.,2010b). Similarly, no isotopic differences occur between the opportu-nistically omnivorous R. decollata and the other, herbivorous species,suggesting that R. decollata has a prevalently herbivorous diet and/orthat animal materials consumed by these specimens usually have acarbon isotope ratio close to the vegetation (e.g. Yanes et al., 2009).
Exceptions are made for shells associated with perennial–annualgrasses with higher δ13Cv, as it is the case of Brachypodion sp. fromArea 5. There, all species show invariable shell 13C-enrichement (medianδ13Cs = −8.6 ± 1.2‰) compared with the other areas (p b 0.001;H = 43.98), and indicate that grasses may contribute significantlyto the diet of different Mediterranean species. Using Stott's equation, a

126 A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 119–127
median δ13Cs of−8.6‰predicts amedian δ13Cv value of ~−23.4‰. Thisvaluematches verywell the δ13Cv of Brachypodion sp. (−23.15‰), indi-cating that grasses were the main dietary component of snails living inthis area.
Shells from areas≤200 m (i.e. areas 1, 4 and 6) are also significantly13C-depleted compared with shells collected at ~600 m (p b 0.01;H = 12.73), but no linear correlation with altitude is observed(R2 = 0.0). Small changes inMediterranean C3 vegetationmay accountfor considerable variability in δ13Cs, with similarmagnitude to those ob-served between modern and late Quaternary shell assemblages fromthe western Mediterranean (Colonese et al., 2013a). Although furtherstudies are required in order to understand factors behind C3 plant iso-topic variability at Lipari, the δ13Cs distribution provides a valid meansof identifying plant adaptive strategy and physiological response evenat microhabitat scale (e.g. Area 5).
5. Conclusions
Our study on the oxygen isotopic composition of land snail shellsfrom Lipari Island, in the Aeolian Archipelago, demonstrates thatδ18Os–δ18Op relationships observed in European and Italian land snailshells may not be valid for central Mediterranean populations locatedat low altitudes (≤600 m asl); thus it is mandatory that regional empir-ical relationships are developed. Our results suggest that the high, andfairly constant, relative humidity promoted by water vapour from theMediterranean Sea compensates for the effect of the altitudinal changesof δ18Op, and possibly of temperature, on shell δ18O. An important im-plication is that oxygen isotopic changes in Quaternary shells preservedin geological and archaeological deposits around the Mediterraneanmay have similar amplitude but not necessarily reflect similar changesin climate. The metabolic model points to the vegetation as the main(or exclusive) source of carbon for the shells. Shell δ13C matches withgood accuracy the isotopic composition of sampled plants and providesa valuable tool for identifying C3 plants with very distinct isotopicsignatures.
Acknowledgement
The authors wish to thank T. Donnelly, J. Dougans, and A. Tait fortheir support in stable isotopes analyses. The authors are grateful toSalvatore Iacullo and Vasco Basile for their kind assistance in fieldwork. This work is partially funded by the University of Pisa (ex 60%G. Zanchetta). SUERC is funded by a consortium of Scottish universitiesand NERC. Thanks to the anonymous referees, for their comments andsuggestions that have contributed to improve this research paper.
References
Arad, Z., Mizrahi, T., Goldenberg, S., Heller, J., 2010. Natural annual cycle of heat shock pro-tein expression in land snails: desert versus Mediterranean species of Sphincterochila.J. Exp. Biol. 213, 3487–3495.
Balakrishnan, M., Yapp, C.J., 2004. Flux balance model for the oxygen and carbon isotopecompositions of land snail shells. Geochim. Cosmochim. Acta 68, 2007–2024.
Balakrishnan,M., Yapp, C.J., Theler, J.L., Carter, B.J., Wyckoff, D.G., 2005. Environmental sig-nificance of 13C/12C and 18O/16O ratios of modern land-snail shells from the southerngreat plains of North America. Quat. Res. 63, 15–30.
Baldini, L.M., Walker, S.E., Bruce, R., Baldini, J.U.L., Crowe, D.E., 2007. Isotope ecology ofthe modern land snails Cerion, San Salvador, Bahamas: preliminary advances towardestablishing a low-latitude island palaeoenvironmental proxy. Palaios 22, 174–187.
Batts, J.H., 1957. Anatomy and life cycle of the snail Rumina decollata (pulmonata:Achatinidae). Southwest. Nat. 2 (2/3), 74–82.
Blondel, J., Aronson, J., 1999. Biology and Wildlife of the Mediterranean Regions. OxfordUniversity Press, Oxford.
Bolle, H.-J., 2003. Mediterranean Climate: Variability and Trends. Springer-Verlag, Berlin.Colonese, A.C., Zanchetta, G., Fallick, A.E., Martini, F., Manganelli, G., Drysdale, R.N., 2010a.
Stable isotope composition of Helix ligata (Müller, 1774) from Late Pleistocene–Holocene archaeological record from Grotta della Serratura (Southern Italy):Palaeoclimatic implications. Glob. Planet. Chang. 71, 249–257.
Colonese, A.C., Zanchetta, G., Dotsika, E., Drysdale, R.N., Fallick, A.E., Grifoni Cremonesi, R.,Manganelli, G., 2010b. Early–Middle Holocene Land snail shell stable isotope recordfrom Grotta di Latronico 3 (Southern Italy). J. Quat. Sci. 25 (8), 1347–1359.
Colonese, A.C., Zanchetta, G., Drysdale, R.N., Fallick, A.E., Manganelli, G., Lo Vetro, D.,Martini, F., Di Giuseppe, Z., 2011. Stable isotope composition of Late Pleistocene–Holocene Eobania vermiculata (Müller, 1774) (Pulmonata, Stylommatophora) shellsfrom the Central Mediterranean basin: data from Grotta d'Oriente (Favignana, Sicily).Quat. Int. 244, 76–87.
Colonese, A.C., Zanchetta, G., Perlès, C., Drysdale, R.N., Manganelli, G., Baneschi, I., Dotsika,E., Valladas, H., 2013a. Deciphering late Quaternary land snail shell δ18O and δ13Cfrom Franchthi Cave (Argolid, Greece). Quat. Res. 80, 66–75.
Colonese, A.C., Zanchetta, G., Fallick, A.E., Manganelli, G., Saña, M., Alcade, G., Nebot, J.,2013b. Holocene snail shell isotopic record of millennial-scale hydrological condi-tions in western Mediterranean: data from Bauma del Serrat del Pont (NE IberianPeninsula). Quat. Int. http://dx.doi.org/10.1016/j.quaint.2013.01.019.
Cook, A., 2001. Behavioural ecology: on doing the right thing, in the right place at the righttime. In: Barker, G.M. (Ed.), The Biology of Terrestrial Molluscs. CAB International,Wallingford, pp. 447–487.
DeNiro, M.J., Epstein, S., 1978. Influence of the diet on the distribution of carbon isotopesin animals. Geochim. Cosmochim. Acta 42, 495–506.
Domínguez, M.T., Aponte, C., Pérez-Ramos, I.M., García, L.V., Villar, R., Marañón, T., 2012.Relationships between leaf morphological traits, nutrient concentrations and isotopicsignatures for Mediterranean woody plant species and communities. Plant Soil 357,407–424.
Filella, I., Peñuelas, J., 2003. Partitioning of water and nitrogen in co-occurring Mediterra-nean woody shrub species of different evolutionary history. Oecologia 137, 51–61.
Giusti, F., 1973. Notulae Malacologicae. XVIII. I Molluschi terrestri e salmastri delle IsoleEolie. Lavori della Società Italiana di Biogeografia, 3, pp. 113–306.
Giusti, F., Manganelli, G., Schembri, P.J., 1995. The Non-marine Molluscs of the MalteseIslands. Museo Regionale di Scienze Naturali, Torino.
Goodfriend, G.A., 1991. Holocene trends in 18O in land snail shells from the Negev desertand their implications for changes in rainfall source areas. Quat. Res. 35, 417–426.
Goodfriend, G.A., Ellis, G.L., 2002. Stable carbon and oxygen isotopic variations in modernRabdotus land snail shells in the southern Great Plains, USA, and their relation toenvironment. Geochim. Cosmochim. Acta 66, 1987–2002.
Goodfriend, G.A., Magaritz, M., Gat, J.R., 1987. Stable isotope composition of land snailbody-water and its relation to environmental waters and shell carbonate. Geochim.Cosmochim. Acta 53, 3215–3221.
Hammer, Ø., Harper, D.A.T., Ryan, P.D., 2001. PAST: Paleontological Statistics SoftwarePackage for education and data analysis. Palaeontol. Electron. 4 (1), 9.
Hartman, G., Danin, A., 2010. Isotopic values of plants in relation to water availability inthe Eastern Mediterranean region. Oecologia 162, 837–852.
Hatziioannou, M., Eleutheriadis, N., Lazaridou-Dimitriadou, M., 1994. Food preferencesand dietary overlap by terrestrial snails in Logos Area (Edessa, Macedonia, NorthernGreece). J. Molluscan Stud. 60, 331–341.
Heller, J., Magaritz, M., 1983. From where do land snails obtain the chemicals to buildtheir shells? J. Molluscan Stud. 49, 116–121.
Iglesias, J., Castillejo, J., 1999. Field observations on feeding of the land snails Helix aspersaMüller. J. Mollus. Stud. 65, 411–423.
Iglesias, J., Santos, M., Castillejo, J., 1996. Annual activity cycles of the land snail Helixaspersa Müller in natural populations in north-western Spain. J. Molluscan Stud. 62,495–505.
Kehrwald, N.M., McCoy, W.D., Thibeault, J., Burns, S.J., Oches, E.A., 2010. Paleoclimaticimplications of the spatial patterns of modern and LGM European land snail shellδ18O. Quat. Res. 74, 166–0176.
Kohn, M.J., 2010. Carbon isotope compositions of terrestrial C3 plants as indicators of(paleo)ecology and (paleo)climate. Proc. Natl. Acad. Sci. U. S. A. 107, 19691–19695.
Kotsakiozi, P., Pafilis, P., Giokas, S., Valakos, E., 2012. A comparison of the physiologicalresponses of two land snail species with different distributional ranges. J. MolluscanStud. 78 (2), 217–224.
Lécolle, P., 1985. The oxygen isotope composition of land snail shells as a climatic indicator:applications to hydrogeology and paleoclimatology. Chem. Geol. (Isot. Geosci. Sect.)58, 157–181.
Leng, M.J., Heaton, T.H.E., Lamb, H.F., Naggs, F., 1998. Carbon and oxygen isotope varia-tions within the shell of an African land snail (Limicolaria kambeul chudeauiGermani): a high-resolution record of climate seasonality. The Holocene 8 (4),407–412.
Leone, G., Bonadonna, F.P., Zanchetta, G., 2000. Stable isotope record inmollusca and ped-ogenic carbonate from late Pliocene soils of Central Italy. Palaeogeogr. Palaeoclimatol.Palaeoecol. 163, 115–131.
Liotta, M., Brusca, L., Grassa, F., Inguaggiato, S., Longo, M., Madonia, P., 2006. Geochemistryof rainfall at Stromboli volcano (Aeolian Islands): isotopic composition andplume–raininteraction. Geochem. Geophys. Geosyst. 7. http://dx.doi.org/10.1029/2006GC001288.
Liotta, M., Grassa, F., D'Alessandro, W., Favara, R., Gagliano Candela, E., Pisciotta, A.,Scaletta, C., 2013. Isotopic composition of precipitation and groundwater in Sicily,Italy. Appl. Geochem. http://dx.doi.org/10.1016/j.apgeochem.2013.03.012.
Lo Cascio, P., Navarra, E., 2003. Guida naturalistica alle Isole Eolie. La vita in un arcipelagovulcanico. L'Epos, Palermo.
Longinelli, A., Selmo, E., 2003. Isotopic composition of precipitation in Italy: a first overallmap. J. Hydrol. 270, 75–88.
Lubell, D., 2004. Are land snails a signature for the Mesolithic/Neolithic transition in thecircum-Mediterranean? In: Budja, M. (Ed.), The Neolithization of Eurasia—Paradigms.Models and Concepts Involved Neolithic Studies, 11, pp. 1–24.
Lubell, D., Barton, N., 2011. Gastropods and humans in the late Palaeolithic andMesolithicof the western Mediterranean basin. Quat. Int. 244, 1–4.
Lucchi, F., Tranne, C.A., Rossi, P.L., 2010. Stratigraphic approach to geological mapping ofthe late Quaternary volcanic island of Lipari (Aeolian archipelago, southern Italy).In: Groppelli, G., Goette, L.V. (Eds.), Stratigraphy and Geology of Volcanic Areas,GSA Special Papers, pp. 1–32.

127A.C. Colonese et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 119–127
Magaritz, M., Heller, J., 1983. Annual cycle of 18O/16O and 13C/12C isotope ratios in landsnail shells. Chem. Geol. (Isot. Geosci. Sect.) 1, 243–255.
Magny, M., Vannière, B., Calo, C., Millet, L., Leroux, A., Peyron, O., Zanchetta, G., La Mantia,T., Tinner, W., 2011. Holocene hydrological changes in south-western Mediterraneanas recorded by lake-level fluctuations at Lago Preola, a coastal lake in southern Sicily,Italy. Quat. Sci. Rev. 30, 2459–2475.
Manganelli, G., Bodon, M., Favilli, L., Giusti, F., 1995. Gastropoda Pulmonata. In: Minelli, A.,Ruffo, S., La Posta, S. (Eds.), Checklist delle specie della fauna italiana. Calderini,Bologna, p. 60.
McConnaughey, T.A., Gillikin, D.P., 2008. Carbon isotopes in mollusk shell carbonates.Geo-Mar. Lett. 28, 287–299.
Mensink, P.J., Henry, H.A.L., 2011. Rain events influence short-term feeding preferences inthe snail Cepaea nemoralis. J. Molluscan Stud. 77, 241–247.
Metref, S., Rousseau, D.-D., Bentaleb, I., Labonne, M., Vianey-Liaud, M., 2003. Study of thediet effect on δ13Cs of shell carbonate of the land snail Helix aspersa in experimentalconditions. Earth Planet. Sci. Lett. 211, 381–393.
Örstan, A., 2010. Activities of four species of land snails at low temperatures. J. Conchol. 40(2), 245–246.
Paul, D., Mauldin, R., 2012. Implications for Late Holocene climate from stable carbon andoxygen isotopic variability in soil and land snail shells from archaeological site41KM6 9 in Texas, USA. Quat. Int. http://dx.doi.org/10.1016/j.quaint.2012.08.006.
Speiser, B., 2001. Food and feeding behaviour. In: Barker, G.M. (Ed.), The Biology ofTerrestrial Molluscs. CAB International, Wallingford, pp. 259–288.
Staikou, A., Lazaridou-Dimitriadou, M., Farmakis, N., 1988. Aspects of the life cycle, popu-lation dynamics, growth and secondary production of the edible snail Helix lucorumLinnaeus, 1758 (Gastropoda, Pulmonata) in Greece. J. Molluscan Stud. 54, 139–155.
Stevens, R.E., Metcalfe, S.E., Leng, M.J., Lamb, A.L., Sloane, H.J., Naranjo, E., González, S.,2012. Reconstruction of late Pleistocene climate in the Valsequillo Basin (CentralMexico) through isotopic analysis of terrestrial and freshwater snails. Palaeogeogr.Palaeoclimatol. Palaeoecol. 319–320, 16–27.
Stott, L.D., 2002. The influence of diet on the δ13Cs of shell carbon in the pulmonate snailHelix aspersa. Earth Planet. Sci. Lett. 195, 249–259.
Xu, B., Gu, Z., Han, J., Liu, Z., Pei, Y., Lu, Y., Wu, N., Chen, C., 2010. Radiocarbon and stablecarbon isotope analyses of land snails from the Chinese loess plateau: environmentaland chronological implications. Radiocarbon 52 (1), 149–156.
Yanes, Y., Delgado, A., Castillo, C., Alonso, M.R., Ibáñez, M., De la Nuez, J., Kowalewski, M.,2008. Stable isotope (δ18O, δ13C, and δD) signatures of recent terrestrial communitiesfrom a low-latitude, oceanic setting: endemic land snails, plants, rain, and carbonatesediments from the eastern Canary Islands. Chem. Geol. 249, 277–292.
Yanes, Y., Romanek, C.S., Delgado, A., Brant, H.A., Noakes, J.E., Alonso, M.R., Ibáñez, M.,2009. Oxygen and carbon stable isotopes of modern land snail shells as environmen-tal indicators from a low-latitude oceanic island. Geochim. Cosmochim. Acta 73 (14),4077–4099.
Yanes, Y., Romanek, C.S.,Molina, F., Cámara, J., Delgado, A., 2011.Holocenepaleoenvironment(7200–4000 cal BP) of the Los Castillejos archaeological site (SE Spain) inferred from thestable isotopes of land snail shells. Quat. Int. 244, 67–75.
Yanes, Y., Gutiérrez-Zugasti, I., Delgado, A., 2012. Late-glacial to Holocene transition innorthern Spain deduced from land-snail shelly accumulations. Quat. Res. 78, 373–385.
Zanchetta, G., Leone, G., Fallick, A.E., Bonadonna, F.P., 2005. Oxygen isotope compositionof living land snail shells: data from Italy. Palaeogeogr. Palaeoclimatol. Palaeoecol.223, 20–33.
Zanchetta, G., Drysdale, R.N., Hellstrom, J.C., Fallick, A.E., Gagan, M.K., Pareschi, M.T., 2007.Enhanced rainfall in the Western Mediterranean during deposition of Sapropel S1:stalagmite evidence from Corchia cave (Central Italy). Quat. Sci. Rev. 26, 279–286.
Zanchetta, G., Bini, M., Cremaschi, M., Magny, M., Sadori, L., 2013. The transition from nat-ural to anthropogenic-dominated environmental change in Italy and the surroundingregions since the Neolithic: an introduction. Quat. Int. http://dx.doi.org/10.1016/j.quaint.2013.05.009.
ZongXiu, L., ZhaoYan, G., NaiQin,W.U., Bing, X., 2007. Diet control on carbon isotopic com-position of land snail shell carbonate. Chin. Sci. Bull. 52 (3), 388–394.