oxidative stress, genotoxicity and histopathology biomarker responses in mullet (mugil cephalus) and...
TRANSCRIPT

Marine Pollution Bulletin 64 (2012) 241–251
Contents lists available at SciVerse ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul
Oxidative stress, genotoxicity and histopathology biomarker responsesin mullet (Mugil cephalus) and sea bass (Dicentrarchus labrax) liverfrom Bizerte Lagoon (Tunisia)
Walid Ben Ameur a, Joaquin de Lapuente b,⇑, Yassine El Megdiche a, Badreddine Barhoumi a,Souad Trabelsi a, Lydia Camps b, Joan Serret b, David Ramos-López b, Javier Gonzalez-Linares b,Mohamed Ridha Driss a, Miquel Borràs b
a Laboratory of Environmental Analytical Chemistry (05/UR/12-03), University of Carthage, Faculty of Sciences, Bizerte, 7021 Zarzouna, Tunisiab UTOX-PCB, Unit of Experimental Toxicology and Ecotoxicology, Parc Científic Barcelona, Barcelona, Spain
a r t i c l e i n f o a b s t r a c t
Keywords:PollutionBiomarkerToxicologyFishLiverBizerte Lagoon
0025-326X/$ - see front matter � 2011 Elsevier Ltd. Adoi:10.1016/j.marpolbul.2011.11.026
⇑ Corresponding author. Tel.: +34 934037193; fax:E-mail address: [email protected] (J. de Lapuen
The aim of the study was to evaluate the impact of environmental contaminants on oxidative stress,genotoxic and histopathologic biomarkers in liver of mullet (Mugil cephalus) and sea bass (Dicentrarchuslabrax) collected from a polluted coastal lagoon (Bizerte Lagoon) in comparison to a reference site(the Mediterranean Sea).
Antioxidant enzyme activities were lower in fish from the polluted site compared with fish from thereference site, suggesting deficiency of the antioxidant system to compensate for oxidative stress. DNAdamage was higher in both fish species from the contaminated site indicating genotoxic effects. The liverhistopathological analysis revealed alterations in fish from Bizerte Lagoon. Hepatocytes from both fishspecies featured extensive lipid-type vacuolation and membrane disruption. Results suggest that theselected biomarkers in both fish species are useful for the assessment of pollution impacts in coastalenvironments influenced by multiple pollution sources.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Pollution of the aquatic ecosystems is a serious and growingproblem. Increasing number and amount of industrial, agriculturaland commercial chemicals discharged into the aquatic environ-ment have led to various deleterious effects on the aquatic organ-isms (McGlashan and Hughies, 2001). Aquatic organisms, includingfish, accumulate pollutants directly from contaminated water andindirectly via the food chain. Fish are often used as sentinelsbecause they play a number of roles in the trophic web, bioaccu-mulate toxic substances, and respond to low concentrations ofmutagens (Al-Sabti and Metcalfe, 1995; Stegeman, 2000).
Aquatic organisms are exposed to a wide variety of environmen-tal contaminants and frequently a set of biomarkers is employed toassess the possible biological impacts and environmental quality.Such biomarkers act as an early warning of a specific detrimentalbiological endpoint.
Biomarkers are increasingly worldwide-recognized tools for theassessment of pollution impacts in the marine environment, and
ll rights reserved.
+34 934037109.te).
some are already incorporated in environmental monitoringprograms (Viarengo et al., 2007). Biomarkers can be characterizedas functional measures of exposure to stressors, which are usuallyexpressed at the subcellular level of biologic organization (Adamset al., 2001). Subcellular responses to environmental stressorsoccur before other disturbances, such as disease, mortality, or pop-ulation changes, and thus may offer early warnings of pollutionimpacts (Depledge and Fossi, 1994). Histopathological analysis,DNA integrity and detoxification enzyme status in fish tissues,alone or in combination, have frequently been employed as suchbiomarkers, each regarded as established early warnings of tumourdevelopment, particularly in the liver. This approach is also impor-tant to investigate the mechanisms by which the environmentalinsult causes complex molecular and cellular alterations as wellas their interdependence. Among the numerous ecotoxicologicalbiomarkers proposed in the last decades, those based on the imbal-ance between pro-oxidant and antioxidant status as well as on itsconsequences namely DNA damage, lipid peroxidation (LPO) andenzyme inhibition, represent earlier signals of environmentaldisturbance (Livingstone, 2001; Regoli et al., 2004). Changes inantioxidant defenses and oxidative damage are used as biomarkersof oxidative stress (Livingstone, 2001). Genotoxic pollutants inducechanges in the genetic material of marine biota, including DNA

242 W. Ben Ameur et al. / Marine Pollution Bulletin 64 (2012) 241–251
damage as well as gene and chromosomal alterations. Biomarkersindicating DNA and chromosomal damage are used to assess geno-toxic effects in aquatic organisms (Ohe et al., 2004). With the viewof biomarker application in environmental monitoring pro-grammes, the evaluation and validation of biomarkers in sentinelspecies under different field conditions is crucial. Biomarkers ofoxidative stress and genotoxicity have been applied in several fishspecies for pollution assessment in harbors and coastal areas influ-enced by industrial discharges (McFarland et al., 1999; Stephensenet al., 2000; Ferreira et al., 2005; Amado et al., 2006; Oliveira et al.,2010). Histopathology provides a sensitive indicator of sublethalstress induced by xenobiotics. Histopathological changes havebeen widely used as biomarkers in the evaluation of the healthof fish exposed to contaminants, both in the laboratory (Thophonet al., 2003) and field studies (Teh et al., 1997). Histopathologicbiomarkers in fish offer a number of practical advantages in thefield, ease of sample collection, ability to assess many body sys-tems and cells types from the same fish. Importantly, no other fieldtechnique enables the specific sites of cellular injury to be identi-fied (Hinton et al., 1992). Histopathological examination is widelyrecognized as a valuable method for assessing the injury in fishesdue to the adverse acute and chronic effects of exposure to anthro-pogenic contaminants (Teh et al., 1997).
Fish liver plays a major role in the uptake, biotransformationand excretion of pollutants (Gernhöfer et al., 2001). However, somemetabolites produced are highly reactive and ultimately moretoxic, affecting the structural integrity of DNA (Shugart, 2000).DNA damage has been associated with the carcinogenesis processand perturbations in fecundity, longevity and growth of affectedorganisms that may have repercussions on population-level effects(Theodorakis et al., 2000). The teleost liver has been the focus oftoxicological studies and has indeed been shown to be very sensi-tive to pollutant exposure (Pinkney et al., 2004; Blazer et al., 2007).
In this perspective, the assessment of hepatic defense and dam-age responses in a resident fish species becomes highly relevant inbiomonitoring studies, considering the important role of the liveron numerous vital functions and subsequently, the proneness toface high levels of contaminants/metabolites and ROS (Oliveira etal., 2003, 2010).
The grey mullet (Mugil cephalus) and the sea bass (Dicentrarchuslabrax) have several characteristics required in a sentinel species,such as wide geographic distribution; great abundance, salinityand temperature tolerance; they are common in coastal watersand enter lagoons, estuaries, harbors, and rivers that are frequentlysubjected to pollution. These two species were selected due totheir different characteristics in terms of ecology, biology, trophicposition in the food chain and feeding behavior.
Both species have high economic importance and gastronomicvalue, which means that the information provided by our studymay be important both to people using the lagoon resources andto those responsible for fishery management. Also they havebeen demonstrated to be suitable for biomarker studies (Orbeaet al., 2002; Gravato and Santos, 2003; Ferreira et al., 2004; Gorbiet al., 2005; Pacheco et al., 2005; Maria et al., 2009; Nogueiraet al., 2010).
In Tunisia, the Bizerte Lagoon, located near some industrial unitsand agricultural area, exploited in conchyliculture since 1964 (Beji,2000), represents a receptor of several industrial wastes, pesticidesand chemical fertilizers through soil erosion and runoff, leading to adecrease in bivalves and fish production (ANPE, 1990). In thisaquatic area, many studies have investigated the application ofbiochemical tools, but these biomarker approaches have been re-stricted to certain species such as Mediterranean clam Ruditapesdecussatus (Dellali et al., 2001, 2004), mussels Mytilus galloprovin-cialis (Dellali et al., 2001; Khessiba et al., 2001, 2005), Hexaplex trun-culus (Roméo et al., 2006), Fulvia fragilis (Mahmoud et al., 2010) and
worms Nereis diversicolor (Bouraoui et al., 2010). A previous studyshowed a reproduction disturbance in a fish species (Gobius niger)from Bizerta Lagoon (Louiz et al., 2009). However, no studies haveassessed the use of M. cephalus and D. labrax, abundant fishes inthe Bizerte Lagoon, as sentinel species and as tools for biomonitor-ing this coastal ecosystem.
The aim of the present study was to assess oxidative stress,genotoxicity and histopathologic biomarkers in mullet and seabass liver from animals obtained in the Bizerte Lagoon influencedby multiple pollution sources to evaluate their use for pollutionassessment.
Cellular antioxidant defenses, namely activities of the antioxi-dant enzymes catalase (CAT), superoxide dismutase (SOD), andthe indicative for lipid peroxidation products and oxidativedamage malondialdehyde (MDA), were used as oxidative stressbiomarkers. The comet test indicating DNA damage was used asbiomarker of genotoxicity. The histopathology was used for theevaluation of hepatic histological alterations.
2. Materials and methods
2.1. Study area and sampling
Bizerte Lagoon, a Mediterranean lagoon covering roughly15 km2 represents an economically important body of water dueto a variety of fishing and aquaculture activities. Fifteen animalsfrom the Bizerte Lagoon and five animals from the MediterraneanSea 7 km northwards from the lagoon (reference site) for each spe-cies were caught alive using a net, in June 2010 (Fig. 1). The lowanalyzed number of fish collected from the Mediterranean Sea isdue to difficulty of their fishing in this area. The MediterraneanSea was chosen to serve as reference site based on previous chem-ical analysis of polybrominated diphenyl ethers and their meth-oxylated analogs (Ben Ameur et al., 2011), which indicated thatthe levels of contaminants were low. Immediately after sampling,fish were sacrificed via exsanguination, weighed and measured.Table 1 shows the information about fish biometrical parameters.Livers were removed and a portion fixed in 10% formaldehyde forhistopathological analysis. The remaining liver was divided intwo sets of tissue for oxidative stress and DNA integrity measure-ments and both frozen in liquid nitrogen and stored at �80 �C untilfurther analysis.
2.2. Somatic indexes
Fish were weighed individually, their total length measured andthe liver dissected and weighed. The Condition Factor (CF) was cal-culated as CF = [body weight (g)/length (mm)3] � 100 (Slooff et al.,1983). Hepatosomatic index was calculated using the formulaHSI = [liver weight (g)/body weight] � 100 (Slooff et al., 1983).
2.3. Liver histopathology
The liver samples fixed in formaldehyde solution weredehydrated in a graded series of ethanol bathes and embedded inparaffin. The embedded organs were sectioned at 5 lm thicknesson a Minot microtome (Leica RM 2145 microtome) and stainedwith Harris’ hematoxilin and eosin using an automatic stainer (Lei-ca EG1140H block-forming unit and Varistain 24–4 K, ThermoShandon). Images were captured using a Nikon E400 microscope(with the Leica QWin image analysis software suite). Careful searchwas conducted to check for the presence of portal inflammation,fibrosis, necrosis, regeneration and extensive degeneration.

Fig. 1. Map showing sampling areas.
Table 1Morphometric data of samples from the Bizerte Lagoon (BL) and the Mediterranean Sea (MS).
Common name Scientific name Weight g (±SD) Length cm (±SD) CF (±SD) HSI % (±SD)
BL MS BL MS BL MS BL MS
Mullet Mugil cephalus 273.15 256.95 31.50 29.06 0.94 1.19 2.90 1.50(78.31) (27.29) (2.40) (3.32) (0.16) (0.03) (0.001) (0.005)
Sea bass Dicentrarchus labrax 187.47 197.71 26.03 27.95 0.98 1.06 1.10 0.50(18.54) (24.18) (0.67) (1.56) (0.06) (0.04) (0.001) (0.001)
SD, standard deviation.CF, condition factor.HSI, hepatosomatic index.Values in parentheses represent the standard deviation.
W. Ben Ameur et al. / Marine Pollution Bulletin 64 (2012) 241–251 243
2.4. Biochemical analysis
2.4.1. CatalaseLivers were homogenized in ice-cold 50 mM sodium phosphate
buffer pH 7, containing 1 mM EDTA. Homogenates were centri-fuged at 10,000g for 15 min. All preparation procedures were per-formed at 4 �C. Catalase enzyme activity was assayed in the10,000g-supernant. Catalase activity was measured using a com-mercial kit (Cayman Chemicals). The Catalase Assay Kit utilizesthe peroxidative function of CAT for determination of enzymeactivity. The method is based on the reaction of the enzyme withmethanol in the presence of an optimal concentration of H2O2.
The generated formaldehyde is allowed to react with 4-amino-3-hydrazino-5-mercapto-1,2,4-triazole (as the chromogen); the alde-hydes, upon oxidation, change from colorless to a purple color thatis measured at 540 nm using a microplate reader. One unit (U) ofCAT activity is defined as the amount of enzyme that causes theformation of 1 nmol of formaldehyde per minute at 25 �C.
2.4.2. SODLivers were homogenized in ice-cold 20 mM Hepes buffer, pH
7.2, containing 1 mM EGTA, 210 mM mannitol, and 70 mM sucrose.Homogenates were centrifuged at 1500g for 5 min at 4 �C. Theresulting supernant was centrifuged at 10,000g for 15 min at
0.00
1.00
2.00
3.00
4.00
5.00
6.00
7.00
8.00
MS BL
CA
T a
ctiv
ity (
nmol
/min
/g ti
ssue
)
Sampling Site
Mugil cephalus
Dicentrarchus labrax
*****
0.00
20000.00
40000.00
60000.00
80000.00
100000.00
120000.00
140000.00
160000.00
MS BL
SOD
act
ivity
(U
/g ti
ssue
tiss
ue)
Sampling Site
Mugil cephalus
Dicentrarchus labrax
***#
**
A
B
244 W. Ben Ameur et al. / Marine Pollution Bulletin 64 (2012) 241–251
4 �C. The resulting 10,000g supernant contained the cytosolic SODand the pellet contained the mitochondrial fraction of SOD. SODenzyme activity was measured in the cytosolic fraction.
SOD activity was measured using a commercial kit (CaymanChemicals). The Superoxide Dismutase Assay Kit utilizes a tetrazo-lium salt for detection of superoxide radicals generated byxanthine oxidase and hypoxanthine measured at 540 nm using amicroplate reader. One unit (U) of SOD activity is defined as theamount of enzyme needed to exhibit 50% dismutation of the super-oxide radical.
2.4.3. MDAMDA measure was a modification of the methods described by
Candan and Tuzmen (2008) and by Del Rio et al. (2005). Liverswere homogenized in ice-cold 50 mM sodium phosphate bufferpH 7, containing 1 mM EDTA. Homogenates were centrifuged at10,000g for 15 min. All preparation procedures were performedat 4 �C. MDA level was assayed in the 10,000g supernant.
MDA level was measured after incubation at 95 �C with thiobar-bituric acid (TBA) in acidic conditions. The pink color produced bythese reactions was measured spectrophotometrically at 532 nm.MDA level was expressed as lM/g tissue.
2.5. Evaluation of DNA integrity by comet assay
The alkaline single cell gel electrophoresis (SCGE) or comet as-say was performed as described by Singh et al. (1988) with somemodifications.
About 50 mg of frozen pieces of liver were washed three timeswith chilled phosphate-buffered saline (PBS; Ca++ Mg++ free) to re-move most of the red blood cells, then transferred to an ice-coldhomogenization buffer (1-X HBSS, 20 Mm EDTA, 10% DMSO, pH,7.0–7.5). The tissues were cut into small pieces with a scissors andfinally homogenized to single-cell suspension. The suspension thenwas centrifuged at 3000 rpm for 5 min at 4 �C, after which the cellpellet was resuspended in PBS. Seventy microlitres of cell suspen-sion were added to an Eppendorf tube containing 160 lL of 1% lowmelting point agarose (37 �C). One hundred and ten microlitres ofthat suspension were added to an agarose precoated slide and gentlycovered with a cover slide to make a micro gel. The slides were im-mersed in lysing solution (2.5 M NaCl, 100 mM Na2EDTA, 10 mMTris and freshly added 1% Triton X-100, pH 10) for 1 h. The slideswere then placed in an horizontal gel electrophoresis unit, im-mersed in cold alkaline electrophoresis buffer (300 mM NaOH and1 Mm Na2EDTA, pH > 13.5), and left in solution for 40 min at 4 �C.Alkaline electrophoresis was carried out using the same solutionat 4 �C for 30 min, at 25 V and 300 mA. The slides were neutralizedby washing three times with 0.4 M Tris buffer at (pH 7.5). DNAwas stained with 20 lL of DAPI solution (40,6-diamidino-2-phenyl-indole) and immediately analyzed using a Nikon E600 fluorescencemicroscope. Percent tail DNA was measured using the Comet AssayIV software (Perceptive Instruments, Suffolk, UK).
2.6. Statistics
All values quoted are mean values ± standard deviations. Differ-ences between sites and fish species were tested using a two wayANOVA with a multiple comparison test (LSD) at a 5% significantlevel. All tests were performed using the software SPSS software(SPSS 10.0 for Windows, SPSS Inc.).
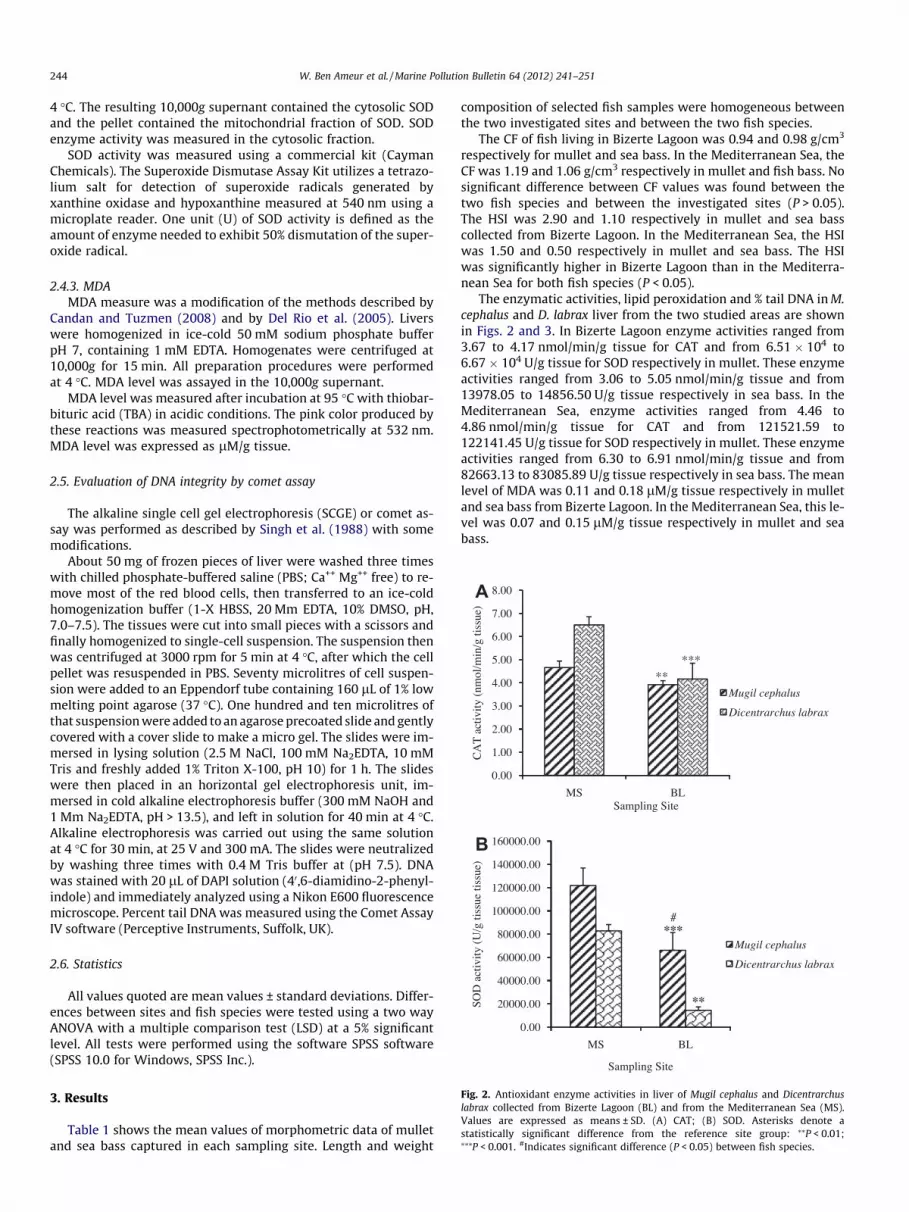
Fig. 2. Antioxidant enzyme activities in liver of Mugil cephalus and Dicentrarchuslabrax collected from Bizerte Lagoon (BL) and from the Mediterranean Sea (MS).Values are expressed as means ± SD. (A) CAT; (B) SOD. Asterisks denote astatistically significant difference from the reference site group: ⁄⁄P < 0.01;⁄⁄⁄P < 0.001. #Indicates significant difference (P < 0.05) between fish species.
3. Results
Table 1 shows the mean values of morphometric data of mulletand sea bass captured in each sampling site. Length and weight
composition of selected fish samples were homogeneous betweenthe two investigated sites and between the two fish species.
The CF of fish living in Bizerte Lagoon was 0.94 and 0.98 g/cm3
respectively for mullet and sea bass. In the Mediterranean Sea, theCF was 1.19 and 1.06 g/cm3 respectively in mullet and fish bass. Nosignificant difference between CF values was found between thetwo fish species and between the investigated sites (P > 0.05).The HSI was 2.90 and 1.10 respectively in mullet and sea basscollected from Bizerte Lagoon. In the Mediterranean Sea, the HSIwas 1.50 and 0.50 respectively in mullet and sea bass. The HSIwas significantly higher in Bizerte Lagoon than in the Mediterra-nean Sea for both fish species (P < 0.05).
The enzymatic activities, lipid peroxidation and % tail DNA in M.cephalus and D. labrax liver from the two studied areas are shownin Figs. 2 and 3. In Bizerte Lagoon enzyme activities ranged from3.67 to 4.17 nmol/min/g tissue for CAT and from 6.51 � 104 to6.67 � 104 U/g tissue for SOD respectively in mullet. These enzymeactivities ranged from 3.06 to 5.05 nmol/min/g tissue and from13978.05 to 14856.50 U/g tissue respectively in sea bass. In theMediterranean Sea, enzyme activities ranged from 4.46 to4.86 nmol/min/g tissue for CAT and from 121521.59 to122141.45 U/g tissue for SOD respectively in mullet. These enzymeactivities ranged from 6.30 to 6.91 nmol/min/g tissue and from82663.13 to 83085.89 U/g tissue respectively in sea bass. The meanlevel of MDA was 0.11 and 0.18 lM/g tissue respectively in mulletand sea bass from Bizerte Lagoon. In the Mediterranean Sea, this le-vel was 0.07 and 0.15 lM/g tissue respectively in mullet and seabass.

0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
MS BL
MD
A a
ctiv
ity (µ
M/g
tiss
ue)
Sampling Site
Mugil cephalus
Dicentrarchus labrax*#
0.00
0.50
1.00
1.50
2.00
2.50
MS BL
% ta
il D
NA
Sampling Site
Mugil cephalus
Dicentrarchus labrax
***
**
A
B
Fig. 3. Lipid peroxidation and DNA damage in liver of Mugil cephalus andDicentrarchus labrax collected from Bizerte Lagoon (BL) and from the MediterraneanSea (MS). Values are expressed as means ± SD. (A) MDA activity; (B) DNA damageexpressed as tail length (%). Asterisks denote a statistically significant differencefrom the reference site group: ⁄P < 0.05; ⁄⁄P < 0.01; ⁄⁄⁄P < 0.001. #Indicates signif-icant difference (P < 0.05) between fish species.
W. Ben Ameur et al. / Marine Pollution Bulletin 64 (2012) 241–251 245
Values of the tail DNA in liver of fish from Bizerte Lagoon rangedfrom 1.75% to 2.17% and from 0.82% to 1.77% respectively for mul-let and sea bass. In the Mediterranean Sea, the baseline level ofDNA damage in liver of mullet measured with the comet assay,was between 0.05% and 0.26% tail DNA, while in sea bass liver itwas between 0.07% and 0.08% tail DNA.
The pollutants effects on enzymatic defense system are repre-sented in Fig. 2. Thus fish liver CAT (Fig. 2A), and SOD (Fig. 2B)activities at Bizerte Lagoon were significantly lower whencompared to the Mediterranean Sea (reference site) for both fishspecies (P < 0.05). The catalase and SOD activities were found tobe significantly lower in Bizerte Lagoon liver fish compared toMediterranean Sea counterpart (P < 0.01 for CAT in mullet;P < 0.001 for CAT in sea bass, P < 0.001 for SOD in mullet; P < 0.01 for SOD in sea bass). Moreover when compared to the Mediter-ranean Sea values, 15.92% and 35.95% decreases in CAT activitywere noticed in mullet and sea bass taken from Bizerte Lagoonrespectively. The reduction observed for SOD was 45.73% and82.52% respectively in mullet and sea bass from Bizerte Lagoon.Statistically significant differences in CAT activity was not ob-served between the studied species (P > 0.05).
Damaging effects of pollutants are shown in Fig. 3. A significantincrease in MDA activity (39.61%) was observed for sea basscollected from Bizerte Lagoon. However, no significant increasein MDA activity (31.39%) was noticed for mullet from Bizerte La-goon (Fig. 3A). Only in D. labrax, the MDA activity was found tobe significantly higher in Bizerte Lagoon fishes liver compared totheir Mediterranean Sea counterpart (P < 0.05).
A significant difference in MDA level was found between thetwo fish species only in the Bizerte Lagoon (P < 0.05).
A significant increase in DNA damage (as % tail DNA) 89.23% and93.97% was observed respectively in mullet and sea bass fromBizerte Lagoon (Fig. 3B). Both fish species from Bizerte Lagoon haveDNA damage values significantly higher than those from Mediter-ranean Sea (P < 0.01 for mullet; P < 0.001 for sea bass). Statisticallysignificant differences in DNA damage were not observed betweenthe studied species (P > 0.05).
A histopathological evaluation was done for hepatocytes of fishinhabiting Bizerte Lagoon and the Mediterranean Sea to determinethe existence and extent of liver toxicity during their exposure toenvironmental contaminants. In both fish species from the Medi-terranean Sea, the morphological characteristics of a normal liverparenchymal architecture is as described for other teleost fish(Rabitto et al., 2005), where hepatocytes surround vessels and typ-ical hepatic cells with rounded nuclei, evident nucleoli and delim-ited cytoplasm are observed (Fig. 4). However, fish liver fromBizerte Lagoon featured extensive vacuolation, corresponding toa relatively higher glycogen and/or lipid content. Apart from thiscommon morphologic response, alteration in the size of the hepa-tocytes, visible hepatocellular membrane disintegration and areasof necrosis were observed in mullet hepatocytes caught fromBizerte Lagoon (Fig. 5C and D). Melanomacrophages centres werepresent in both fish species collected from Bizerte Lagoon(Fig. 5B–D). Apoptosis was observed only in mullet from Bizerte La-goon (Fig. 5D). In general, the liver of D. labrax was less damagedthan M. cephalus liver.
4. Discussion
Fish is generally acknowledged as a worthy model for assessingaquatic contamination and is used as an environmental sentinel forwater toxicants. Fish liver can be regarded as the body’s detoxifica-tion organ and hence a target organ of various xenobiotic sub-stances. A first-level screening to identify potential pollutantexposure and effect can be accomplished on the basis of overtand relatively simple measures of condition. Such measures mayserve to identify the most sensitive members of a fish population.In addition, they may provide information on energy reserves andpossibly the ability of animals to tolerate toxicant challenges orother environmental stresses (Mayer et al., 1992). Morphologicalparameters that are often determined in field research are thehepatosomatic index (HSI), to identify potential liver diseases,and the condition factor (CF), to assess the general condition offish.
Some authors propose the use of ecological indexes like hepato-somatic and gonadosomatic indexes and CF in biomonitoring stud-ies to evaluate the influence of biotic processes or as an additionaltool in biomonitoring approaches (Adams and Ryon, 1994). CF isusually related to food quality (Bagnasco et al., 1991) and repro-ductive status. Condition factor is similar in fish from Bizerte La-goon and from the Mediterranean Sea for both species. Thisresult is similar to that found by Tsangaris et al. (in press). The lackof variation of morphometric condition factor between sites for thetwo species most likely due to the narrow size range of analysedindividuals, intentionally selected in order to avoid size effectson biochemical responses. The length, weight and condition factorwere rather homogeneous, indicating similar nutritional condi-tions of fish at the two sampling areas.
Fish from polluted environments usually show an increase inthe HSI (Gül et al., 2004; Azevedo et al., 2009; Karels et al.,1998). This pattern is also observed in both fish species from Biz-erte Lagoon. Increase in liver weight among Bizerte Lagoon fishspecies may be due to liver damage that ranges from increased

Fig. 4. Histological sections of fish liver prepared from Dicentrarchus labrax (A and B) and Mugil cephalus (C and D) inhabiting the Mediterranean Sea showing normal hepatictissue without pathological alterations. HE � 200.
246 W. Ben Ameur et al. / Marine Pollution Bulletin 64 (2012) 241–251
liver weights and fat content to cell necrosis. The low HSI in fishfrom the reference site reflects a lower hepatic stress in thoseindividuals.
The condition of the liver and of the whole body, as measuredwith the HSI and CF values, can provide information on potentialpollution impacts. Although these parameters are not very sensi-tive and may be affected by non-pollutant factors (e.g. season,disease, nutritional level), they may serve as an initial screeningbiomarker to indicate exposure and effects or to provide informa-tion on energy reserves (Mayer et al., 1992). The condition indicesare quite general and non-specific, but their low cost, ease andrapidity still make them valuable environmental risk assessmenttools.
Many pollutants (or their metabolites) may exert toxicitythrough oxidative stress. Oxidative stress occurs when the rate ofROS generation exceeds the antioxidant defense system. Oxidativestress can derive from increased production of reactive oxygenspecies (ROS) mediated by heavy metals and numerous organiccontaminants, including redox cycling compounds, polycyclicaromatic hydrocarbons (PAHs), halogenated hydrocarbons, anddioxins (Livingstone 2001).
For instance, Winston and Di Giulio (1991) found elevated ratesof idiopathic lesions and neoplasia among fish inhabiting pollutedenvironments to be related to the increased oxidative stress asso-ciated with pollutant exposure. Oxygen toxicity is defined as inju-rious effects due to cytotoxic reactive oxygen species, also referredto as reactive oxygen intermediates, oxygen free radicals or oxy-radicals (DiGiulio et al., 1989; Halliwell and Gutteridge, 1999;Winzer, 2001). These reduction products of molecular oxygen(O2) are the superoxide anion radical (O2
��), hydrogen peroxide
(H2O2) and the hydroxyl radical (OH.), an extremely potent oxidantcapable of reacting with critical cellular macromolecules, possiblyleading to enzyme inactivation, lipid peroxidation, DNA damageand, ultimately, cell death (Winston and Di Giulio, 1991). Numer-ous endogenous sources of oxyradical production exist, but theone of more immediate interest regarding environmental biomark-ers is the ability of a number of structurally diverse compounds toenhance intracellular oxyradical production through the process ofredoxcycling. Redox-active compounds include aromatic diols andquinones, nitroaromatic compounds, aromatic hydroxylamines,bipyridyls and certain transition metal chelates (Winston and DiGiulio, 1991). In the redox cycle, the parent compound is typicallyfirst enzymatically reduced by a NADPH dependent reductase(such as cyt P450 RED) to yield a xenobiotic radical. This radical do-nates its unshared electron to molecular O2, yielding O2
�� and theparent compound. Thus, at each turn of the cycle, two potentiallydeleterious events have occurred: a reductant has been oxidizedand an oxyradical has been produced (Winston and Di Giulio,1991; Goeptar et al., 1995). Oxidant-mediated effects with a poten-tial suitability as biomarkers include either adaptive responses,such as increased activities of antioxidant enzymes or concentra-tions of non-enzymatic compounds, or manifestations of oxidant-mediated toxicity such as oxidations of proteins, lipids and nucleicacids, as well as perturbed tissue redox status (Winston and DiGiulio, 1991; Filho Wilhelm, 1996). Defense systems that tend toinhibit oxyradical formation include the antioxidant enzymes suchas SOD, CAT, glutathione-dependent peroxidase (GPX) and gluta-thione reductase (GR). SOD, CAT and GPX are critically importantin the detoxification of radicals to nonreactive molecules. Numer-ous low-molecular-weight antioxidants, such as glutathione

Fig. 5. Histological sections of fish liver prepared from Dicentrarchus labrax (A and B) and Mugil cephalus (C and D) inhabiting the Bizerte Lagoon showing hepatic tissue withpathological alterations; (A) cytoplasmic vacuolization of hepatocytes; (A and B) necrotic liver tissue; (B and D) melanomacrophage center; (C and D) lipid-type vacuolation,and structural disruption. HE � 200.
W. Ben Ameur et al. / Marine Pollution Bulletin 64 (2012) 241–251 247
(GSH), b-carotene (vitamin B), ascorbate (vitamin C), a-tocopherol(vitamin E) and ubiquinol10 have been described (Stegeman et al.,1992; López-Torres et al., 1993).
Antioxidant enzymes are commonly employed biomarkers ofoxidative stress; however, responses to pollution vary for differentspecies, enzymes, and single or mixed contaminants, and evengreater variability is found in field situations (Livingstone, 2001).Hence, literature data on fish species report higher, unchanged orlower activities of antioxidant enzymes for polluted comparedwith cleaner environments (van der Oost et al., 2003). The presentstudy shows overall lower CAT and SOD activities in the liver of M.cephalus and D. labrax from Bizerte Lagoon compared with fishfrom the reference site (Mediterranean Sea), indicating antioxidantenzyme inhibition. Low CAT and SOD activities have also beenreported in M. cephalus from a polluted estuary (Ennore Estuary)in India (Padmini et al., 2009; Padmini and Usha Rani, 2009) andin D. labrax from Aveiro Lagoon in Portugal (Maria et al., 2009).In contrast, previous studies in grey mullet and in sardine (Sardinapilachardus) showed higher CAT and SOD activities in fish frompolluted areas (Rodriguez-Ariza et al., 1993; Peters et al., 1994).Antioxidant enzymes can be induced by enhanced production ofROS as a protection mechanism against oxidative stress or inhib-ited when deficiency of the system occurs, suggesting toxicity(Winston and Di Giulio 1991; Cossu et al., 1997). The enzyme re-sponse to toxic chemicals shows a bell-shaped trend with an initialincrease in activity due to enzyme induction followed by a de-crease in activity due to enhanced catabolic rate and/or direct inhi-bition by toxic chemicals (Viarengo et al., 2007). Accordingly, thelow enzyme activities in fish from Bizerte Lagoon can be associatedwith deficiency to compensate for oxidative stress, possibly due tohigh levels of pollutant exposure. With regard to antioxidant
enzymes, the primary defense against oxygen toxicity involvesthe enzyme SOD which catalyzes the conversion of superoxide an-ion (O2
��) to oxygen and hydrogen peroxide (H2O2) and catalasewhich further degrades H2O2 to water. In our study, the levels ofSOD and CAT were found to be significantly lower in fish liver fromBizerte Lagoon. The decreased activities of SOD and CAT enzymesfavor the accumulation of O2
�� and H2O2 respectively and theseradicals accelerate the conversion of Fe3+ to Fe2+ and the latterserves as a substrate for hydroxyl radical (OH�) to generate a reac-tion leading to enhanced lipid peroxidation (Halliwell and Gutter-idge, 1985). The reduced activity of these enzymes may beattributed their long term exposure to environmental pollutantsand a similar type of response has been demonstrated by severalworkers (Bainy et al., 1996; Lenartova et al., 1997). The low levelsof CAT could be further attributed to high production of superoxideanion radical, which has been reported to inhibit CAT activity incase of excessive production (Pandey et al., 2003). Apart fromsuperoxide anions, nitrites have also been reported to inhibit CATactivity (Arrillo and Melodia, 1991). Comparing the antioxidantdefenses of the two species (Table 2), marked differences wereobserved only in the Bizerte Lagoon for SOD activity. M. cephaluspresented a statistically higher level of SOD than D. labrax(P < 0.05). This result is similar to that reported by Ferreira et al.(2005). This suggests that mullet liver cytosol can deal with theincrease in superoxide anion more efficiently than the sea bass.Antioxidant defenses in fish are dependent on feeding behaviorand nutritional factors. Herbivorous fish species have higher super-oxide dismutase activities than omnivorous fish (Radi et al., 1985).Dietary levels of lipids and some vitamins have been reported toinfluence antioxidant defenses and oxidative status of fish.Mourente et al. (2002) showed that diets containing oxidized oil

248 W. Ben Ameur et al. / Marine Pollution Bulletin 64 (2012) 241–251
significantly affected the activities of liver antioxidant defense en-zymes of gilthead sea bream (Sparus aurata L.) and that dietaryvitamin E partially abrogated these effects. In the Senegalese sole(Solea senegalensis), Rueda-Jasso et al., 2004 reported that activitylevels of the antioxidant enzymes CAT and SOD were higher in liv-ers of fish fed diets with a high lipid level.
The toxicity effect of ROS can produce various damages to thecell, such as DNA damage, lipid peroxidation and lysosomal alter-ation (Viarengo, 1989; Winston et al., 1996; Regoli, 1998, 2000;Frenzilli et al., 2001). Lipid peroxidation is a well-known mecha-nism of cellular injury in vertebrates and invertebrates, and is anindicator of an oxidative damage in cells and tissues. Therefore,measurement of MDA, the end product of lipid peroxidation iswidely used as an indicator of lipid peroxidation (Wheatley,2000). MDA can react readily with amino groups on proteins andother molecules to form a variety of adducts (Esterbauer et al.,1991; Marnett, 1999). A significant increase in MDA concentrationwas found only for D. labrax. The values for MDA in sea bass wereslightly higher than the ones estimated for mullet. This differencemay be related with less efficient defenses, which is consistentwith the lower levels of the SOD as antioxidant primary defensesassessed in sea bass. In addition to that, the difference in fatty acidcomposition of the liver membranes may also be involved(Halliwell and Gutteridge, 1999). The absence of oxidative stressin terms of lipid peroxides in mullet liver indicates that the antiox-idant responses are not overwhelmed. Fish oxidative stress, as wellas its antioxidant potential, differs in relation to species, habitat,and feeding behavior. For example, sole from a polluted harborshowed a high LPO (Di Gulio et al., 1993) and elasmobranchesshowed higher LPO than marine teleosts, which had much highervalues than freshwater teleosts (Filho Wilhelm, 1996). LPO tendsto be lower in herbivorous fish than in omnivorous fish species,which can be correlated to the lower GPX and CAT activities (Radiet al., 1985). The no observed statistical differences in liver LPOmeasurements, suggest that mullet is able to deal with the pollu-tion stress, despite the depletion of several antioxidant defensesat Bizerte Lagoon. This is in agreement with Cossu et al. (2000),who stated that LPO increase cannot be predicted only on the basisof antioxidant depletion. Furthermore, Ferreira-Cravo et al. (2007)demonstrated that the low antioxidant competence against HO.
radicals is not concomitant with high LPO values in estuarine poly-chaeta Laeonereisacuta sampled from a contaminated site, suggest-ing that other macromolecules can also be affected by the higherROS levels.
Both species of fish collected from Bizerte Lagoon showed sig-nificant increases of DNA damage when compared to the referencepopulations (Mediterranean Sea) (Fig. 3B). Our result agrees withprevious data which shows increases in DNA damage in fish fromcontaminated area (Flammarion et al., 2002; Winter et al., 2004;Nogueira et al., 2009). One cause for the significant levels of DNAdamage observed may be the action of highly reactive xenobioticmetabolites following liver metabolism. The relatively higher levelof DNA damage found in M. cephalus may reflect its feeding behav-ior and habitat. It feeds on benthic organisms and is in regular con-tact with the sediments where the majority of the contaminantsaccumulate. It should be stressed that the DNA damage deter-mined through comet assay was paralleled by a lack of CAT in liverfish at the polluted site. Catalase activity is essential to promotethe degradation of H2O2, a precursor of hydroxyl radical, a reactiveoxygen species that induces DNA damage (Halliwell andGutteridge, 1999).
Single-cell gel electrophoresis (SCGE) or comet assay has be-come a widespread technique for detection of DNA damage in-duced by xenobiotics, e.g. Cd (Desai et al., 2006; Fourie et al.,2007), Hg (Tran et al., 2007), organic compounds (Costa et al.,2008). The alkaline version of the assay has proven to be a simple
and reliable method for the quantitation of total DNA fragmenta-tion as a result of the formation of single and double-strand break-age, xenobiotic–DNA adducts and alkali-labile sites (e.g. unstablealtered nucleotides; Singh et al., 1988). The comet assay has beenused in a wide range of aquatic organisms, such as marine diatoms(Desai et al., 2006), bivalve mollusks (e.g. Jha et al., 2005; Desaiet al., 2006) and fish (e.g. Flammarion et al., 2002; Winter et al.,2004; Nogueira et al., 2009), for the biomonitoring of coastalenvironments.
Histopathology provides a rapid method to detect the effect ofirritants in various organs Johnson et al. (1993). The liver is the pri-mary organ for metabolism, detoxification of xenobiotics andexcretion of harmful substances. The liver has the ability to de-grade toxic compounds but its regulating mechanism can beoverwhelmed by elevated concentrations of these compoundswhich could subsequently result in structural damage (Brusleand Gonzalez, 1996). Two of the main alterations observed in bothfish species were vacuolar degeneration changes and areas ofnecrosis. These alterations have been described in other speciesof fish living in contaminated environments (Teh et al., 1997; Neroet al., 2006; Padmini and Usha Rani, 2009; Nogueira et al., 2010)suggesting that these alterations might be related to the exposureto environmental chemicals present. Nevertheless, the presence ofthese changes is less pronounced in the liver of D. labrax than in M.cephalus, which suggests that D. labrax liver is being less affectedby aquatic contamination than M. cephalus. While D. labrax is apredator of small fishes and feeds in the water column, M. cephalusfeeds on benthic organisms and is in regular contact with the sed-iments where the majority of the contaminants accumulate. In thisway, M. cephalus may be in contact with higher concentrations ofcontaminants which may lead to an increased risk of damage tothe liver as observed in this study. Fish livers from Bizerte Lagoontend to be more vacuolated, corresponding to a relatively higherglycogen and/or lipid content. Such vacuolization, which tends tobe uniformly distributed, is often especially apparent in the liversof captive fishes and the fishes that are exposed to toxic contami-nants (Wolf and Wolfe, 2005; Borges et al., 2006). Ferguson (1989)reported that toxic exposure can result in accumulations of fat orglycogen in the liver. It has also been demonstrated that hepaticlipidosis may be associated with toxicant induced lipid peroxida-tion. For example, lipid peroxidation occurred in channel catfishexposed to sediment contaminated with metals (Di Gulio et al.,1993). The melanomacrophage centres, focal accumulations ofpigmented macrophages are involved in the storage of foreign oreffete material and their prevalence and intensity has been pro-posed as a potentially useful biomarker of environmental degrada-tion and pollution (Couillard and Hodson, 1996; in Manera et al.,2000). Comparative studies on the incidence of melanomacro-phage centres in the spleen, kidneys and liver of fish taken fromwaters polluted with toxic chemicals have, however, producedconflicting results (Pulsford et al., 1992). Some authors reportedan increase (Kranz and Peters, 1984; Wolke et al., 1985; Kranzand Gercken, 1987), whilst others recorded the opposite (Kranz,1989). This latter worker observed an increase at low levels of pol-lution, which he ascribed to the capability of the cellular defensesystem to remove debris by increased phagocytic activity, withconsequent increase in melano-macrophage centres.
Apoptosis is due to a complex pathway in which biochemicalmechanisms are responsible for activation of key events. For exam-ple, induction of apoptosis via the release of cytochrome c due tothe effects of heavy metals in mitochondria is well known (Araragiet al., 2003) as are BCL2 inhibition and disturbances of intracellularCa2+ concentrations (Sallas and Buchiel, 1998). Lindane alsoincreases intracellular Ca2+ concentration (Duchiron et al., 2002),which may be associated with cell death. This would partially ex-plain the toxicity of this insecticide to fish (Betoulle et al., 2000).

W. Ben Ameur et al. / Marine Pollution Bulletin 64 (2012) 241–251 249
Also, chlordane (ATSDR, 1994), lindane (Azzalis et al., 1995) andendrin (Bagchi et al., 2002) apparently induced cell death in liversof fish and mammals. Although the mechanisms are still unclear,apparently PAH can induce apoptosis in fish (Weber and Janz,2001).
It was considered, that the induction of antioxidant enzymesrepresents a protective response to eliminate ROS resulting fromcontamination exposure, it has been hypothesised that suchincrease may be related to adaptations to contaminant inducedstress (Livingstone, 2001). In the present study, the significativedecrease of antioxidant enzyme activities, the significative liverDNA damage and liver alterations observed in fish from thepolluted area may reflect signs of effective toxicity and not anadaptation to chronic exposure to contaminants.
It is hard to predict, to what extent the observed alterationsmay eventually influence the health of these native fish popula-tions. Several comprehensive studies using histopathologicalparameters have demonstrated clear relationships betweenenvironmental pollution and fish disease. The relationships, forinstance, between pollution levels, biomarker responses and skinand liver diseases in flounder after long-term exposure to PAH-and PCB-contaminated sediments in a mesocosm study werereported by Vethaak et al. (1996).
Contaminants that induce oxidative stress, genotoxicity, andhistopathological alterations such as metals, PAHs, PCBs, DTTs,and TBT were found in the Bizerte Lagoon. In fact, the BizerteLagoon is subjected to many anthropogenic pressures includingurbanization and industrial activities. The direct and indirect dis-charges of urban and industrial wastes and runoff lead to thechemical contamination of the lagoon by various toxic compoundssuch as heavy metals (Zn, Cd, Ni, Pb and Cu) (Yoshida et al., 2004),organo-chlorinated pesticides (Cheikh et al., 2002), halogenatedaromatic compounds like polychlorobiphenyls (PCBs) (Derouicheet al., 2004), organotins (Mzoughi et al., 2005; Abidli et al., 2011)and polycyclic aromatic hydrocarbons (PAHs) (Trabelsi and Driss,2005; Ben Said et al., 2008). These compounds are likely responsi-ble for the observed effects at Bizerte Lagoon.
Our study did not attempt to define which specific contami-nants in the polluted area were responsible for oxidative stress,genetic damage and histopathological alterations. Our moremodest objective was to detect overall oxidative stress, geno-toxic and histopathologic effects from multiple contaminants,because the Bizerte Lagoon is subjected to multiple sources ofpollution.
5. Conclusion
The liver fish biomarkers evaluated in this study show differ-ences between the area exposed to anthropogenic influence(Bizerte Lagoon) in comparison to the unpolluted zone in the Med-iterranean Sea in front of the Tunisian coast, suggesting distur-bances in the specific cell mechanisms due to the presence ofmultiple xenobiotics in the Bizerte Lagoon. This work reinforcesthe strong anthropogenic influence on Bizerte Lagoon where indus-trial and agricultural activities are intense and shows the suitabilityof M. cephalus and D. labrax as sentinel species for biomonitoringstudies. This study shows oxidative stress, genotoxic effects andhistopathogic alterations by means of biomarkers in mullet ( M.cephalus) and sea bass (D. labrax) from an area influenced by multi-ple pollution sources, thus confirming their usefulness as tools forassessment of pollution impacts in situations where complex mix-tures of contaminants are found. To our knowledge this is the firststudy of enzymatic, histopathologic and genotoxic biomarkersusing biomarkers from fish liver for assessment in Tunisia coastalwaters.
Acknowledgments
We gratefully acknowledge the technical assistance of NatàliaBrull Barco and Constança Porredon Guarch. We thank also theanonymous referees for their helpful comments, which willimprove the paper.
References
Abidli, S., Lahbib, Y., Trigui El Menif, N., 2011. Imposex and butyltin concentrationsin Bolinus brandaris (Gastropoda: Muricidae) from the northern Tunisian coast.Environ. Monit. Assess. 177, 375–384.
Adams, S.M., Giesy, J.P., Tremblay, L.A., Eason, C.T., 2001. The use of biomarkers inecological risk assessment: recommendations from the Christchurch conferenceon Biomarkers in Ecotoxicology. Biomarkers 6, 1–6.
Adams, S.M., Ryon, M.G., 1994. A comparison of health assessment approaches forevaluating the effects of contaminant-related stress on fish populations. J.Aquat. Ecosys. Health 3, 15–25.
Al-Sabti, K., Metcalfe, C.D., 1995. Fish micronuclei for assessing genotoxicity inwater. Mutat. Res. 343, 121–135.
Amado, L.L., Robaldo, R.B., Geracitano, L., Monserrat, J.M., Bianchini, A., 2006.Biomarkers of exposure and effect in the Brazilian flounder Paralichthysorbignyanus (Teleostei: Paralichthyidae) from the Patos Lagoon estuary(Southern Brazil). Mar. Pollut. Bull. 52, 207–213.
ANPE, 1990. Diagnostic préliminaire pour l’Etude de l’Equilibre Ecologique du lac deBizerte. GIC-NNEA-TECI, ANPE, Tunisie.
Araragi, S., Kondoh, M., Kawase, M., Saito, S., Higashimoto, M., Sato, M., 2003.Mercuric chloride induces apoptosis via a mitochondrial-dependent pathway inhuman leukaemia cells. Toxicology 184, 1–9.
Arrillo, A., Melodia, F., 1991. Nitrite oxidation in Eisema foetida (Savigny): ecologicalimplications. Funct. Ecol. 5, 629–634.
ATSDR, 1994. Toxicological Profile for Chlordane. US Department of Health andHuman Services, Atlanta, USA, 133 pp.
Azevedo, J.S., Serafim, A., Company, R., Braga, E.S., Fávaro, D.I., Bebianno, M.J., 2009.Biomarkers of exposure to metal contamination and lipid peroxidation in thebenthic fish Cathorops spixii from two estuaries in South America. BrazilEcotoxicol. 18, 1001–1010.
Azzalis, L.A., Junqueira, V.B.C., Simon, K., 1995. Prooxidant and antioxidant hepaticfactors in rats chronically fed an ethanol regimen and treated with an acutedose of lindane. Free Radic. Biol. Med. 19, 147–159.
Bagchi, D., Balmoori, J., Bagchi, M., Ye, X., Willians, C.B., Stohs, S.J., 2002.Comparative effects of TCDD, endrin, naphthalene and chromium (VI) onoxidative stress and tissue damage in the liver and brain tissues of mice.Toxicology 175, 73–82.
Bagnasco, M., Camoirano, A., De Flora, S., Melodia, F., Arillo, A., 1991. Enhanced livermetabolism of mutagens and carcinogens in fish living in polluted seawater.Mutat. Res. 262, 129–137.
Bainy, A.C.D., Saito, E., Carvalho, P.S.M., Junqueira, V.B.C., 1996. Oxidative stress ingill, erythrocytes, liver and kidney of Nile tilapia (Oreochromis niloticus) from apolluted site. Aquat. Toxicol. 34, 151–162.
Beji, O., 2000. Les ressources vivantes exportables du lac de Bizerte: Etat actuel etpotentialités (première partie). Bull. Inst. Nat. Sci. Tech. Mer. Salammbô 27, 45–60.
Ben Ameur, W., Ben Hassine, S., Eljarrat, E., El Megdiche, Y., Trabelsi, S., Hammami,B., Barceló, D., Driss, M.R., 2011. Polybrominated diphenyl ethers and theirmethoxylated analogs in mullet (Mugil cephalus) and sea bass (Dicentrarchuslabrax) from Bizerte Lagoon, Tunisia. Mar. Environ. Res. 72, 258–264.
Ben Said, O., Goñi-Urriza, M.S., El Bour, M., Dellali, M., Aissa, P., Duran, R., 2008.Characterization of aerobic polycyclic aromatic hydrocarbondegrading bacteriafrom Bizerte lagoon sediments, Tunisia. J. Appl. Microbiol. 104, 987–997.
Betoulle, S., Duchiron, C., Deschaux, P., 2000. Lindane differently modulatesintracellular calcium levels in two populations of rainbow trout(Oncorhynchus mykiss) immune cells: head kidney phagocytes and peripheralblood leucocytes. Toxicology 145, 203–215.
Blazer, V.S., Fournie, J.W., Wolf, J.C., Wolfe, M.J., 2007. Manual for the diagnosticanalysis of proliferative liver and skin lesions in the brown bullhead Ameiurusnebulosus. Pennsylvania Sea Grant/USGS Publication.
Borges, L.P., Nogueira, C.W., Panatieri, R.B., Rocha, J.B.T., Zeni, G., 2006. Acute liverdamage induced by 2-nitropropane in rats: effect of diphenyl diselenide onantioxidant defenses. Chem. Biol. Interact. 160, 99–107.
Bouraoui, Z., Banni, M., Chouba, L., Ghedira, J., Clerandeau, C., Jebali, J., Narbonne, J.F.,Boussetta, H., 2010. Monitoring pollution in Tunisian coasts using a scale ofclassification based on biochemical markers in worms Nereis (Hediste)diversicolor. Environ. Monit. Assess. 164, 691–700.
Brusle, J., Gonzalez, G., 1996. The structure and function of fish liver. In: Munshi,J.S.D., Dutta, H.M. (Eds.), Fish Morphology. Science Publishers Inc., India.
Candan, N., Tuzmen, N., 2008. Very rapid quantification of malondialdehyde (MDA)in rat brain exposed to lead, aluminium and phenolic antioxidants by high-performance liquid chromatography-fluorescence detection. Neurotoxicology29, 708–713.
Cheikh, M., Derouiche, A., Driss, M.R., 2002. Détermination par (CPG-ECD) desrésidus de pesticides organochlorés dans les sédiments de la lagune de Bizerte.Bull. Inst. Nat. Sci. Technol. Mer. 7, 160–163.

250 W. Ben Ameur et al. / Marine Pollution Bulletin 64 (2012) 241–251
Cossu, C., Doyotte, A., Babut, M., Exinger, A., Vasseur, P., 2000. Antioxidantbiomarkers in freshwater bivalves, unio tumidus, in response to differentcontamination profiles of aquatic sediments. Ecotoxicol. Environ. Saf. 45,106–121.
Cossu, C., Doyotte, A., Jacquin, M.C., Babut, M., Exinger, A., Vasseur, P., 1997.Glutathione reductase, selenium-dependent glutathione peroxidase,glutathione levels, and lipid peroxidation in freshwater bivalves, Uniotumidus, as biomarkers of aquatic contamination in field studies. Ecotoxicol.Environ. Saf. 38, 122–131.
Costa, P.M., Lobo, J., Caeiro, S., Martins, M., Ferreira, A.M., Caetano, M., Vale, C.,DelValls, T.A., Costa, M.H., 2008. Genotoxic damage in Solea senegalensisexposed to sediments from the Sado Estuary (Portugal): effects of metallicand organic contaminants. Mutat. Res. 654, 29–37.
Couillard, C.M., Hodson, P.V., 1996. Pigmented macrophage aggregates: a toxicresponse in fish exposed to bleached-Kraft mill effluent? Environ. Toxicol.Chem. 15, 1844–1854.
Dellali, M., Gnassia-Barelli, M., Roméo, M., Aissa, P., 2001. The use ofacetylcholinesterase activity in Ruditapes decussatus and Mytilusgalloprovincialis in the biomonitoring of Bizerta lagoon. Comp. Biochem.Physiol. C 130, 227–235.
Dellali, M., Roméo, M., Gnassia-Barelli, M., Aissa, P., 2004. A multivariate dataanalysis of the clam Ruditapes decussatus as sentinel organism of the Bizertalagoon (Tunisia). Water, Air, Soil Pollut. 44, 131–156.
Del Rio, D., Stewart, A.J., Pellegrini, N., 2005. A review of recent studies onmalondialdehyde as toxic molecule and biological marker of oxidative stress.Nutr. Metab. Cardiovasc. Dis. 15, 316–328.
Depledge, M.H., Fossi, M.C., 1994. The role of biomarkers in environmentalassessment (2). Invertebrates. Ecotoxicology 3, 161–172.
Derouiche, A., Sanda, Y.G., Driss, M.R., 2004. Polychlorinated biphenyls in sedimentsfrom Bizerte lagoon, Tunisia. Bull. Environ. Contam. Toxicol. 73, 810–817.
Desai, S.R., Verlecar, X.N., Nagarajappa, Goswami, U., 2006. Genotoxicity ofcadmium in marine diatom Chaetoceros tenuissimus using the alkaline Cometassay. Ecotoxicology 15, 359–363.
Di Gulio, R.T., Habig, C., Gallagher, E.P., 1993. Effects of Black Rock Harbor sedimentson indices of biotransformation, oxidative stress, and DNA integrity in channelcatfish. Aquat. Toxicol. 26, 1–22.
DiGiulio, R.T., Washburn, P.C., Wenning, R.J., Winston, G.W., Jewell, C.S., 1989.Biochemical responses in aquatic animals: a review of determinants ofoxidative stress. Environ. Toxicol. Chem. 8, 1103–1123.
Duchiron, C., Betoulle, S., Reynaud, S., Deschaux, P., 2002. Lindane increasesmacrophage-activating factor production and intracellular calcium in rainbowtrout (Oncorhynchus mykiss) leukocytes. Ecotoxicol. Environ. Saf. 53,388–396.
Esterbauer, H., Schaur, R.J., Zollner, H., 1991. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med.11, 81–128.
Ferguson, H.W., 1989. Systemic pathology of fish. A text and atlas of comparativetissue responses in diseases of teleosts. Lowa State, University Press, Ames, IA,p. 263.
Ferreira, M., Moradas-Ferreira, P., Reis-Henriques, M.A., 2005. Oxidative stressbiomarkers in two resident species, mullet (Mugil cephalus) and flounder(Platichthys flesus), from a polluted site in River Douro Estuary, Portugal. Aquat.Toxicol. 71, 39–48.
Ferreira, M., Antunes, P., Gil, O., Vale, C., Reis-Henriques, M.A., 2004. Organochlorinecontaminants in flounder (Platichthys flesus) and mullet (Mugil cephalus) fromDouro estuary and their use as sentinel species for environmental monitoring.Aquat. Toxicol. 69, 347–357.
Ferreira-Cravo, M., Piedras, F.R., Moraes, T.B., Ferreira, J.L., de Freitas, D.P., Machado,M.D., Geracitano, L.A., Monserrat, J.M., 2007. Antioxidant responses and reactiveoxygen species generation in different body regions of the estuarine polychaetaLaeonereis acuta (Nereididae). Chemosphere 66, 1367–1374.
Filho Wilhelm, D., 1996. Fish antioxidant defenses – a comparative approach. Braz.J. Med. Biol. Res. 29, 1735–1742.
Flammarion, P., Devaux, A., Nehls, S., Migeon, B., Noury, P., Garric, J., 2002.Multibiomarker responses in fish from the Moselle River (France). Ecotoxicol.Environ. Saf. 51, 145–153.
Fourie, F., Reinecke, S.A., Reinecke, A.J., 2007. The determination of earthwormspecies sensitivity differences to cadmium genotoxicity using the comet assay.Ecotoxicol. Environ. Saf. 67, 361–368.
Frenzilli, G., Nigro, M., Scarcelli, V., Gorbi, S., Regoli, F., 2001. DNA integrity and totaloxyradical scavenging capacity in the Mediterranean mussel, Mytilusgalloprovincialis: a field study in a highly eutrophicated coastal lagoon. Aquat.Toxicol. 53, 19–32.
Gernhöfer, M., Pawet, M., Schramm, M., Müller, E., Triebskorn, R., 2001.Ultrastructural biomarkers as tools to characterize the health status of fish incontaminated streams. J. Aqua. Ecosyst. Stress Recov. 8, 241–260.
Goeptar, A.R., Scheerens, H., Vermeulen, N.P.E., 1995. Oxygen and xenobioticreductase activity of cytochrome P450. Crit. Rev. Toxicol. 25, 25–65.
Gorbi, S., Baldini, C., Regoli, F., 2005. Seasonal variability of metallothioneins,cytochrome P450, bile metabolites and oxyradical metabolism in the Europeaneel Anguilla anguilla L. (Anguillidae) and striped mullet Mugil cephalus L.(Mugilidae). Arch. Environ. Contam. Toxicol. 49, 62–70.
Gravato, C., Santos, M.A., 2003. Dicentrarchus labrax biotransformation andgenotoxicity responses after exposure to a secondary treated industrial/urbaneffluent. Ecotoxicol. Environ. Saf. 55, 300–306.
Gül, S., Belge-Kurutas�, E., Yildiz, E., Sahan, A., Doran, F., 2004. Pollution correlatedmodifications of liver antioxidant systems and histopathology of fish(Cyprinidae) living in Seyhan Dam Lake, Turkey. Environ. Int. 30, 605–609.
Halliwell, B., Gutteridge, J.M.C., 1985. Free radicals in Biology and Medicine.Clarendon, Oxford, UK.
Halliwell, B., Gutteridge, J.M.C., 1999. Free radicals in biology and medicine, thirded. Clarendon Press, Oxford.
Hinton, D.E., Baumann, P.C., Gardner, G.R., Hawkins, W.E., Hendricks, J.D.,Murchelano, R.A., Okihiro, M.S., 1992. Histopathologic biomarkers. In: Hugget,R., Kimerle, R., Mehrle, P., Bergman, H. (Eds.), Biomarkers – Biochemical,Physiological and Histological Markers of Anthropogenic Stress. LewisPublishers, Boca Raton, pp. 155–195.
Jha, A.N., Dogra, Y., Turner, A., Millward, G.E., 2005. Impact of low doses of tritiumon the marine mussel, Mytilus edulis: genotoxic effects and tissue-specificbioconcentration. Mutat. Res. 586, 47–57.
Johnson, L.L., Stehr, C.M., Olson, O.P., Myers, M.S., Pierce, S.M., Wigren, C.A., Mccain,B.B., Varnasi, U., 1993. Chemical contaminants and hepatic lesions in winterflounder (Pleuronectes americanus) from the Northeast coast of the UnitedStates. Environ. Sci. Technol. 27, 2759–2777.
Karels, A.E., Soimasuo, M., Lappivaara, J., Leppanen, H., Aaltonen, T., Mellanen, P.,Oikari, A.O.J., 1998. Effects of ECF bleached kraft mill effluent on reproductivesteroids and liver MFO activity in populations of perch and roach. Ecotoxicology7, 123–132.
Kranz, H., 1989. Changes in splenic melano-macrophage centres of dab Limandalimanda during and after infection with ulcer disease. Dis. Aqua. Org. 6, 167–173.
Kranz, H., Gercken, J., 1987. Effects of sublethal concentration of potassiumdichromate on the occurrence of splenic melano-macrophage centres injuvenile plaice, Pleuronectes platessa L. J. Fish. Biol. 31 (Suppl. A), 75–80.
Kranz, H., Peters, N., 1984. Melano-macrophage centres in liver and spleen of ruffe(Gymnocephalus cernuus) from the Elbe Estuary. Helgolander. Meeresun. 37,415–424.
Lenartova, V., Holovska, K., Pedrajas, J.R., Lara, E.M., Peinado, J., Barea, J.L., Rosival, I.,Kosuth, P., 1997. Antioxidant and detoxifying fish enzymes as biomarkers ofriver pollution. Biomarkers 2, 247–252.
Livingstone, D.R., 2001. Contaminant-stimulated reactive oxygen speciesproduction and oxidative damage in aquatic organisms. Mar. Pollut. Bull. 42,656–666.
López-Torres, M., Perez-Campo, R., Cadenas, S., Rojas, C., Barja, G., 1993. Acomparative study of free radicals in vertebrates – II. Non-enzymaticantioxidants and oxidative stress. Comp. Biochem. Physiol. 105, 757–763.
Louiz, I., Ben-Attia, M., Ben-Hassine, O.K., 2009. Gonadosomatic index and gonadhistopathology of Gobius niger (Gobiidea, Teleost) from Bizerta lagoon (Tunisia):evidence of reproduction disturbance. Fish. Res. 100, 266–273.
Mahmoud, N., Dellali, M., El Bour, M., Aissa, P., Mahmoudi, E., 2010. The use of Fulviafragilis (Mollusca: Cardiidae) in the biomonitoring of Bizerta lagoon: amutimarkers approach. Ecol. Indic. 10, 696–702.
Manera, M., Serra, R., Isani, G., Carpenè, E., 2000. Macrophage aggregates in giltheadsea bream fed copper, iron and zinc enriched diets. J. Fish. Biol. 57,457–465.
Maria, V.L., Ahmad, I., Oliveira, M., Serafim, A., Bebianno, M.J., Pacheco, M., Santos,M.A., 2009. Wild juvenile Dicentrarchus labrax L. liver antioxidant and damageresponses at Aveiro Lagoon, Portugal. Ecotoxicol. Environ. Saf. 72, 1861–1870.
Marnett, L.J., 1999. Lipid peroxidation—DNA damage by malondialdehyde. Mutat.Res. 424, 83–95.
Mayer, F.L., Versteeg, D.J., McKee, M.J., Folmar, L.C., Graney, R.L., McCume, D.C.,Rattner, B.A., 1992. Metabolic products as biomarkers. In: Huggett, R.J., Kimerly,R.A., , Mehrle, P.M., Jr., Bergman, H.L. (Eds.), Biomarkers: Biochemical,Physiological and Histological Markers of Anthropogenic Stress. LewisPublishers, Chelsea, MI, USA, pp. 5–86.
McFarland, V.A., Inouye, L.S., Lutz, C.H., Jarvis, A.S., Clarke, J.U., McCant, D.D., 1999.Biomarkers of oxidative stress and genotoxicity in livers of field-collectedbrown bullhead, Ameiurus nebulosus. Arch. Environ. Contam. Toxicol. 37, 236–241.
McGlashan, D.J., Hughies, J.M., 2001. Genetic evidence for historical continuitybetween populations of the Australian freshwater fish Craterocephalusstercusmuscarum (Atherinidae) east and west of the Great Diving Range. J.Fish Biol. 59, 55–67.
Mourente, G., Díaz-Salvago, E., Bell, J.G., Tocher, D.R., 2002. Increased activities ofhepatic antioxidant defence enzymes in juvenile gilthead sea bream (Sparusaurata L.) fed dietary oxidised oil: attenuation by dietary vitamin E. Aquaculture214, 343–361.
Nero, V., Farwell, A., Lister, A., Van Der Kraak, G., Lee, L.E.J., Van Meer, T., MacKinnon,M.D., Dixon, D.G., 2006. Gill and liver histopathological changes in yellow perch(Perca Dixon flavescens) and goldfish (Carassius auratus) exposed to oil sandsprocess-affected water. Ecotoxicol. Environ. Saf. 63, 365–377.
Nogueira, P., Pacheco, M., Pereira, M.L., Mendo, S., Rotchell, J.M., 2010. Anchoringnovel molecular biomarker responses to traditional responses in fish exposed toenvironmental contamination. Environ. Pollut. 158, 1783–1790.
Ohe, T., Watanabe, T., Wakabayashi, K., 2004. Mutagens in surface waters: a review.Mutat. Res. 567, 109–149.
Oliveira, M., Ahmad, I., Maria, V.L., Pacheco, M., Santos, M.A., 2010. Antioxidantresponses versus DNA damage and lipid peroxidation in golden grey mulletliver: a field study at Ria de Aveiro (Portugal). Arch. Environ. Contam. Toxicol.59, 454–463.

W. Ben Ameur et al. / Marine Pollution Bulletin 64 (2012) 241–251 251
Oliveira, M., Santos, M.A., Gravato, C., Pacheco, M., 2003. Chromium effects onAnguilla anguilla liver organ culture. Fresen. Environ. Bull. 12, 349–352.
Orbea, A., Ortiz-Zarragoitia, M., Solé, M., Porte, C., Cajaraville, M.P., 2002.Antioxidant enzymes and peroxisome proliferation in relation to contaminantbody burdens of PAHs and PCBs in bivalve molluscs, crabs and fish from theUrdaibai and Plentzia estuaries (Bay of Biscay). Aquat. Toxicol. 58, 75–98.
Pacheco, M., Santos, M.A., Teles, M., Oliveira, M., Rebelo, J.E., Pombo, L., 2005.Biotranformation and genotoxic biomarkers in mullet species (Liza sp.) from acontaminated coastal lagoon (Ria de Aveiro, Portugal). Environ. Monit. Assess.107, 133–153.
Padmini, E., Usha Rani, M., 2009. Evaluation of oxidative stress biomarkers inhepatocytes of grey mullet inhabiting natural and polluted estuaries. Sci. TotalEnviron. 407, 4533–4541.
Padmini, E., Usha Rani, M., Vijaya Geetha, B., 2009. Studies on antioxidant status inMugil cephalus in response to heavy metal pollution at Ennore estuary. Environ.Monit. Assess. 155, 215–225.
Pandey, S., Parvez, S., Sayeed, I., Haque, R., Bin-Hafeez, B., Raisuddin, S., 2003.Biomarkers of oxidative stress: a comparative study of river Yamuna fishWallago attu (Bl. & Schn.). Sci. Total Environ. 309, 105–115.
Peters, L.D., Porte, C., Albaigés, J., Livingstone, D.R., 1994. 7-Ethoxyresorufin Odeethylase (EROD) and antioxidant enzyme activities in larvae of sardine(Sardina pilchardus) from the North coast of Spain. Mar. Pollut. Bull. 28, 299–304.
Pinkney, A.E., Harshbarger, J.C., May, E.B., Reichert, W.L., 2004. Tumor prevalenceand biomarkers of exposure and response in brown bullhead (Ameiurusnebulosus) from the Anacostia river, Washington, DC and Tuckahoe river,Maryland, USA. Environ. Toxicol. Chem. 323, 638–647.
Pulsford, A.L., Ryan, K.P., Nott, J.A., 1992. Metals and melano-macrophages inflounder Platichthys flesus spleen and kidney. J. Mar. Biol. Assoc. UK 72,483–498.
Rabitto, I., Alves Costa, J.R.M., Akaishi, F.M., Silva de Assis, H.C., Pelletier, E., OliveiraRibeiro, C.A., 2005. Dietary Pb(II) and TBT (tributyltin) exposures to neotropicalfish hoplias malabaricus: histopatological and biochemical findings. Ecotoxicol.Environ. Saf. 60, 147–156.
Radi, A.A.R., Hai, D.Q., Matkovics, B., Gabrielak, T., 1985. Comparative antioxidantenzyme study in freshwater fish with different types of feeding behaviour.Comp. Biochem. Physiol. C 81, 395–399.
Regoli, F., 1998. Trace metals and antioxidant enzymes in gills and digestive glandof the Mediterranean mussel, Mytilus galloprovincialis. Arch. Environ. Contam.Toxicol. 34, 48–63.
Regoli, F., 2000. Total oxyradical scavenging capacity (TOSC) in polluted andtranslocated mussels: a predictive biomarker of oxidative stress. Aquat. Toxicol.50, 351–361.
Regoli, F., Frenzilli, G., Bocchetti, R., Annarumma, F., Scarcelli, V., Fattorini, D., Nigro,M., 2004. Time-course variations of oxyradical metabolism, DNA integrity andlysosomal stability in mussels, Mytilus galloprovincialis, during a fieldtranslocation experiment. Aquat. Toxicol. 68, 167–178.
Rodriguez-Ariza, A., Peinado, J., Pueyo, C., Lopez-Barea, J., 1993. Biochemicalindicators of oxidative stress in fish from polluted littoral areas. Can. J. Fish.Aquat. Sci. 50, 2568–2573.
Roméo, M., Gharbi-Bouraoui, S., Gnassia-Barelli, M., Dellali, M., Aissa, P., 2006.Responses of Hexaplex (Murex) trunculus to selected pollutants. Sci. Total.Environ. 359, 135–144.
Rueda-Jasso, R., Conceicao, L.E.C., Dias, J., De Coen, W., Gomes, E., Rees, J.F., Soares, F.,Dinis, M.T., Sorgeloos, P., 2004. Effect of dietary non-protein energy levels oncondition and oxidative status of Senegalese sole (Solea senegalensis) juveniles.Aquaculture 231, 417–433.
Sallas, V.M., Buchiel, S.W., 1998. Apoptosis in Daudi Human B cells in response tobenzo[a]pyrene and benzo[a]pyrene-7,8-dihydrodiol. Toxicol. Appl. Pharmacol.151, 367–376.
Singh, N.P., McCoy, M.T., Tice, R.R., Schneider, E.L., 1988. A simple technique forquantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 175,184–191.
Slooff, W., Van Kreijl, C.F., Baars, A.J., 1983. Relative liver weights and xenobiotic-metabolizing enzymes of fish from polluted surface water in the Netherlands.Aquat. Toxicol. 4, 1–14.
Stegeman, J.J., 2000. Cytochrome P450 gene diversity and function in marineanimals: past, present, and future. Mar. Environ. Res. 50, 61–62.
Stegeman, J.J., Brouwer, M., Richard, T.D.G., Förlin, L., Fowler, B.A., Sanders, B.M., vanVeld, P.A., 1992. Molecular responses to environmental contamination: enzyme
and protein systems as indicators of chemical exposure and effect. In: Huggett,R.J., Kimerly, R.A., Mehrle, P.M., Jr., Bergman, H.L. (Eds.), Biomarkers:Biochemical, Physiological and Histological markers of Anthropogenic Stress.Lewis Publishers, Chelsea, MI, USA, pp. 235–335.
Stephensen, E., Svavarsson, J., Sturve, J., Ericson, G., Adolfsson-Erici, M., Förlin, L.,2000. Biochemical indicators of pollution exposure in shorthorn sculpin(Myoxocephalus scorpius), caught in four harbours on the southwest coast ofIceland. Aquat. Toxicol. 48, 431–442.
Teh, S.J., Adams, S.M., Hinton, D.E., 1997. Histopathological biomarkers in feralfreshwater fish populations exposed to different types of contaminant stress.Aquat. Toxicol. 37, 51–70.
Theodorakis, C.W., Swartz, C.D., Rogers, W.J., Bickham, J.W., Donnelly, K.C., Adams,S.M., 2000. Relationship between genotoxicity, mutagenicity, and fishcommunity structure in a contaminated stream. J. Aqua. Ecosyst. StressRecov. 7, 131–143.
Thophon, S., Kruatrachue, M., Upatham, E.S., Pokethitiyook, P., Sahaphong, S.,Jaritkhuan, S., 2003. Histopathological alterations of white seabass, Latescalcarifer in acute and subchronic cadmium exposure. Environ. Pollut. 121,307–320.
Trabelsi, S., Driss, M.R., 2005. Polycyclic aromatic hydrocarbons in superficialcoastal sediments from Bizerte Lagoon, Tunisia. Mar. Pollut. Bull. 50,344–348.
Tran, D., Moody, A.J., Fisher, A.S., Foulkes, M.E., Jha, A.N., 2007. Protective effects ofselenium on mercury-induced DNA damage in mussel haemocytes. Aquat.Toxicol. 84, 11–18.
Tsangaris, C., Vergolyas, M., Fountoulaki, E., Nizheradze, K., in press. Oxidative stressand genotoxicity biomarker responses in grey mullet (Mugil cephalus) from apolluted environment in Saronikos Gulf, Greece. Arch. Environ. Contam. Toxicol.
van der Oost, R., Beyer, J., Vermeulen, N.P.E., 2003. Fish bioaccumulation andbiomarkers in environmental risk assessment: a review. Environ. Toxicol.Pharmacol. 13, 57–149.
Vethaak, A.D., Jol, J.G., Meijboom, A., Eggens, M.L., Reinallt, T., Wester, P.W., van deZande, T., Bergman, A., Dankers, N., Ariese, F., Baan, R.A., Everts, J.M.,Opperhuizen, A., Marquenie, J.M., 1996. Skin and liver diseases induced inflounder (Platichthys flesus) after long-term exposure to contaminatedsediments in large-scale mesocosms. Environ. Health Perspect. 104, 1218–1229.
Viarengo, A., 1989. Heavy metals in marine invertebrates, mechanisms ofregulation and toxicity at the cellular level. CRC Crit. Rev. Aquat. Sci. 1,295–317.
Viarengo, A., Lowe, D., Bolognesi, C., Fabbri, E., Koehler, A., 2007. The use ofbiomarkers in biomonitoring: a 2-tier approach assessing the level of pollutant-induced stress syndrome in sentinel organisms. Comp. Biochem. Physiol. C.Toxicol. Pharmacol. 146, 281–300.
Weber, L.P., Janz, D.M., 2001. Effect of b-dimethylbenz[a]-anthracene on apoptosisand HSP70 expression in juvenile channel catifish (Ictalurus punctatus) ovary.Aquat. Toxicol. 54, 39–50.
Wheatley, R.A., 2000. Some recent trends in the analytical chemistry of lipidperoxidation. Trends Anal. Chem. 19, 617–628.
Winston, G.W., Di Giulio, R.T., 1991. Prooxidant and antioxidant mechanisms inaquatic organisms. Aquat. Toxicol. 19, 137–161.
Winston, G.W., Moore, N.M., Kirchin, M.A., Soverchia, C., 1996. Production ofreactive oxygen species by hemocytes from the marine mussel, Mytilus edulis:lysosomal localization and effect of xenobiotics. Comp. Biochem. Physiol. C.Pharmacol. Toxicol. Endocrinol. 113, 221–229.
Winter, M., Day, N., Hayes, R.A., Taylor, E.W., Butler, P.J., Chipman, J.K., 2004. DNAstrand breaks and adducts determined in feral and caged chub (Leuciscuscephalus) exposed to rivers exhibiting variable water quality aroundBirmingham, UK. Mutat. Res. 552, 163–175.
Winzer, K., 2001. Oxidative stress in the marine environment-prognostic tools fortoxic injury in fish liver cells. Academic Thesis, University of Amsterdam.
Wolf, J.C., Wolfe, M.J., 2005. A brief overview of nonneoplastic hepatic toxicity infish. Toxicol. Pathol. 33, 75–85.
Wolke, R.E., Murchelano, R.A., Dickstein, C.D., George, C.J., 1985. Preliminaryevaluation of the use of macrophage aggregates (MA) as fish health monitors.Bull. Environ. Contam. Toxicol. 35, 222–227.
Yoshida, M., Hamdi, H., Abdulnasser, I., Jedidi, N., 2004. Contamination ofpotentially toxic elements (PTEs) in Bizerte lagoon bottom sediments, surfacesediment and sediment repository. In: Ghrabi, A., Yoshida, M. (Eds.), Study onEnvironmental Pollution of Bizerte Lagoon. INRST-JICA Publishers, Tunis, pp.31–54.