optokinetic reactions in man elicited by localized retinal motion stimuli
TRANSCRIPT

OPTOKINETIC REACTIONS IN MAN ELICITED BY LOCALIZED RETINAL MOTION STIMULI
M. F. W. DUBOIS and H. C~LLEWIJN
Department of Physiology, Faculty of Medicine, Erasmus University Rotterdam, Rotterdam, The Netherlands
Abstract-The role of different parts of the human retina in eliciting optokinetic pursuit was investigated with continuously moving random dot or grid patterns (dia up to 30”). The retinal location and velocity of the stimulus could be fixed by servo control of the stimulus position by the eye position, and artificial scotomata could be inserted into the stimulus. Under these conditions (opened optokinetic feedback loop) pursuit velocity became much larger than the stimulus velocity. With a central stimulus, pursuit velocity (gain) in the 8 principal directions showed individual, but no systematic differences. In the periphery, a centrifugal stimulus movement was much more effective than a centripetal move- ment. This directional preference may assist foveation. Pursuit open-loop gain was only moderately diminished by a decrease of the stimulus diameter, but much more severely by deleting small parts in the centre. This indicates that the fovea is more powerfut than the periphery in eliciting optokinetic pursuit. The distribution of fast and slow nystagmic phases with respect to the primary position was unaffected by the use of open-loop conditions or central scotomata. For all peripheral stimuli, the responses were strongly enhanced by the subject’s specific attention.
INTRODUCTION
The response of the human eye to moving objects is highly complex. The most simple form is probably the optokinetic pursuit of a rotatory movement of the entire visual surroundings around the subject. Outside the laboratory, this type of stimutus occurs only due to head rotation and is therefore associated with stimulation of the labyrinths and the perception of self-rotation (circul~v~tjon~. It may be considered as a postural reflex that maintains stability of the vis- ual field and is found in all vertebrate and inverte- brate species with spatially organized vision. Ter Braak (1936) called this Stier-Nystagmus, which he distinguished from Schau-Nystagmus, the smooth pursuit of targets that move with respect to a station- ary background. Smooth pursuit tends to align the fovea or central area with the moving object of inter- est, but it is clear that more than the fovea is involved in the process. The role of the peripheral retina in the selection and foveation of the target is obvious. Foveation is usually achieved by a saccade, but Kom- mereli and Klein (1971) have shown that stationary excentric after-images may elicit smooth eye move- ments with the fovea moving in the direction of the after-image. Also pursuit after fovea1 acquisition of the target is in part peripheral, since smooth pursuit without saccades is slower than target velocity so that fixation errors of several degrees may occur (Pucket and Steinman, 1969; Steinman et al., 1969). Smooth pursuit can even be accomplished for an eccentric target without foveation (Winterson and Steinman, 1978).
For the understanding of global and selective pur- suit it would be important to have information on the power of moving patterns, projected on different parts of the retina, to induce smooth eye movements.
Even for the rabbit, which has only global pursuit, it has recently been established (Dubois and Collewijn, 1979) that the optokinetic effect of a stimulus depends very much upon its retinal location; stimulation of the visual streak was much more effective than of the periphery.
For primates, most available data originate from the work of three groups. Hood {1%7, 1975) has de- scribed a peripheral and a fovea1 type of optokineti~ nys~gmus. The peripheral type was isolated by using scotopic vision or patients with a central scotoma. It was asserted that the peripherai retina was much more powerful in eliciting optokinetic nystagmus than the fovea, as shown by the much higher slow-phase velocity gain in the scotomatous than in the normal eye, and also by the strong decrease of nystagmus seen with progressive masking of the peripheral visual field. Moreover, the fovea1 type had its fast com- ponents directed to the periphery, while the peri- pheral type resembled vestibular nystagmus and had slow phases away from the primary position and fast phases returning the eyes to the mid~sition. Kiirner and colleagues (Korner and Dichgans, 1967; Kiirner and Schiller, 1972; Korner, 1975) presented motion stimuli to one paralysed eye of monkeys and some human patients, and measured the movements elicited in the other (covered) eye. This approach, in which the optokinetic feedback loop is effectively opened, is highly similar to that used by Ter Braak (1936) on rabbits. In agreement with the latter it was found that the slow phase velocity of the elicited nystagmus became much faster than the stimulus velocity. In contrast to Hood’s reports, Kiirner found that the optokinetic effect of stimuli rapidly deteriorated with increase of eccentricity. Largely comparable findings were reported by Cheng and ~terbrjdge (1975), who stimulated humans with computer-generated dot pat-
1105
M. F. W. DUBOIS and H. C~LLEWIJN
down 1 I I I
1 ,
21Fup I L h ,..A.,....,.. L ,
7 1
right 1 i SSC
I I
: eft I
normal \ open loop I \ normal
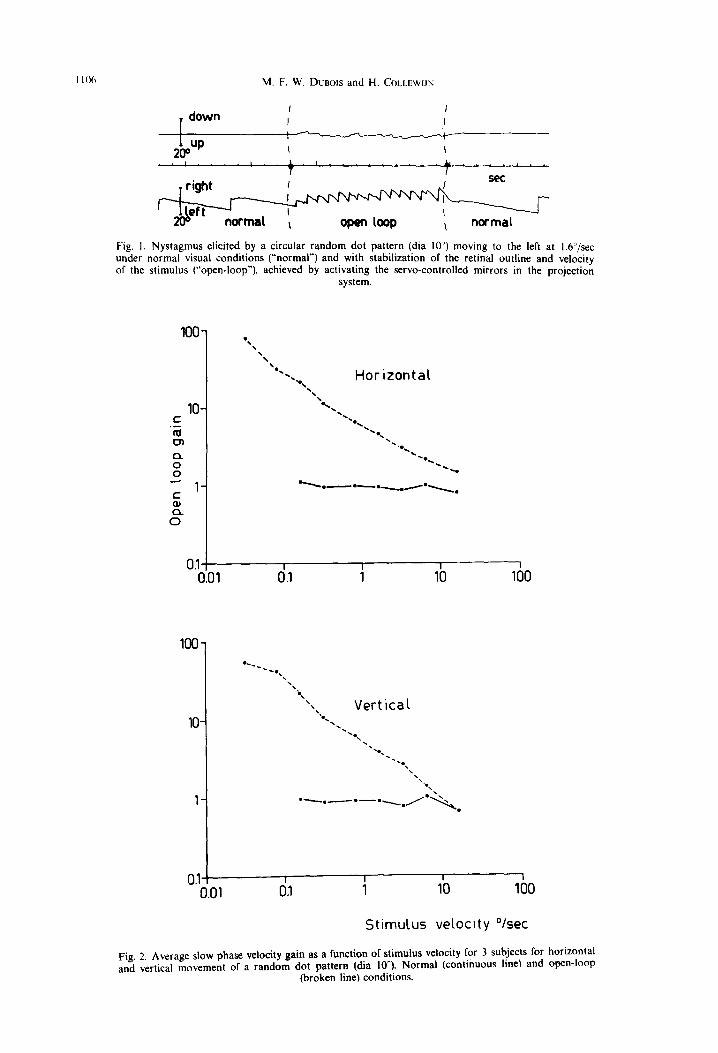
Fig. 1. Nystagmus elicited by a circular random dot pattern (dia IO’) moving to the left at 1.6”/sec under normal visual conditions (“normal”) and with stabilization of the retinal outline and velocity of the stimulus (“open-loop”). achieved by activating the servo-controlled mirrors in the projection
system.
0.0 I I I 1
1 OS 1 10 100
OS I I I I
0.01 OS 1 10 100
Stimulus velocity O/set
Fig. 2. Average slow phase velocity gain as a function of stimulus velocity for 3 subjects for horizontal and vertical movement or a random dot pattern (dia lo”). Normal (continuous line) and open-loop
(broken line) conditions.

Optokinetic nystagmus in man 1107
terns, from which the part projected on the central retina could be deleted.
In view of the incomplete and partly conflicting data in the literature, we reinvestigated smooth eye movements elicited by a moving pattern projected on the retina with different positions, directions and velo- cities, using a new technique to stabilize the retinal stimulus position.
METHODS
Measurements were made on 9 human subjects. None of them showed any visual or oculomotor pathology other than common refraction anomalies. Horizontal and verti- cal eye movements were measured with the technique of a scleral search coil in a magnetic field, developed by Robinson (1963) and modified by Collewijn et al. (1975). The induction coil was embedded in an annulus of silicone rubber, which was shaped in such a way to adhere to the bulbus, concentric with the cornea, when pressed upon the eye. The other eye was covered. Head movements were minimized by supports under the chin and around the skull. The stimulus pattern was projected on a translucent tangential screen at a distance of 230cm from the subject. The screen covered an angle of 65” horizontally and 55” vertically. Subjects could wear their normal spectacles for optimal acuity at the distance of the screen. For calibra- tion. the subject was instructed to fixate a single light point (dia 0.1‘) on the screen which could be displaced in cali-
brated steps. Horizontal and vertical eye position were recorded on a pen-recorder (Grass Polygraph). This tech- nique provided precise and stable recordings with a very low noise level and good linearity up to 20” deviation. The stimulus pattern was printed on an endless film loop, which coutd be moved continuously through a slide projec- tor at 10 fixed velocities (range 0.01616”/sec). The projec- tor could be rotated around its optical axis to provide motion in any direction. Two perpendicular servo-con- trolled mirrors (General Scanning, Watertown, Massachu- setts) were mounted in the light pathway. By connecting the horizontal and vertical eye position signals with proper scaling to the mirror control system, the stimulus outline could be stabilized on the retina in such a way that eye movements did not change the retinal stimulus position. By switching this connection on and off it was very simple to alternate between normal and stabilized conditions. Within the stabilized outline, the pattern, moving at the imposed velocity, was projected. With careful calibration, stabilization was good enough to cause rapid perceptual fading with a stationary pattern. The moving pattern remained visibie. The overall bandwidth of the stabiliza- tion system was 1OOHz (-3dB) for the horizontal and 30 Hz for the vertical position.
The position of the pattern with respect to the fixation point was adjusted by offset voltages. The stimulus outline was changed by inserting different masks in the slide pro- jector; in this way it was also possible to introduce artih- cial scotomata by obscuring a part of the pattern. Either a grid (moving perpendicular to the bars) or a Julesz (1964)
Fig. 3. Vector diagrams of direction and velocity (represented by direction and length of arrows) of smooth eye movements elicited in 5 subjects (A-E) by a circular random dot pattern (dia lo’) moved in the 8 principal directions at I.6 /set (St). The averaged vectors for the same subjects are also
shown (Avg).
110X M. F. W. Dueo~s and H. COLLEWIJN
type random dot pattern (elements 0.43 was used. The latt& had the advantage of a two-dimensional structure and unambiguous direction of motion. The fuminance of the light and dark parts of the pattern was 7 and < 1 cd/m’, respectively. The duration of individual sessions was restricted to a maximum of 40 min to prevent signifi- cant discomfort or degadinp of visual acuity.
RESULTS
The response to a moving pattern projected on the central retina could be investigated under normal as well as stabilized conditions. In the first case, the eye was free to fixate elements of the pattern. In the second case the outline of the pattern remained fixed, concentric with the fovea and unchanged by eye movements: the loop between pattern motion and ocular motion was open. Subjects were instructed to watch the pattern with attention. In the normal con- dition the eye pursued the pattern with a slow phase velocity very close to the pattern velocity, i.e. the gain (smooth eye velocity/pattern velocity) was close to 1.0 and the direction of pursuit corresponded exactly to that of the stimulus movement. The effect of switching to the stabilized (open-loop) condition is shown in Fig. I for a random dot stimulus of lo” dia, moving to the left at 1.6”/sec. The pursuit velocity increased rapidly to values far in excess of stimulus velocity, and the direction of the pursuit became less precise,
as shown by the introduction of a vertical component in Fig. 1, centre. Maximum eye velocity was reached within f-5 set after switching to the open-loop condi- tion, and returned immediately to the pattern velocity upon switching back to normai conditions. Fluctua- tions in eye velocity were often seen during open-loop stimulation. Our velocity measurements were made in the periods in which the nysta~mus was maximally developed. No after-nystagmus was seen. even when open-loop stimulation was abruptly replaced by dark- ness. Subjects experienced no circularvection, but did report a marked increase of the apparent velocity of the pattern when the pattern was stabilized, A station- ary stabilized pattern (existing when the transport of the film loop was stopped) was perceived only inter- mittently and caused no significant eye movements.
Average normal and open-loop gains as a function of stimulus velocity are shown for 3 subjects in Fig. 2A for horizontal, and Fig, 28 for vertical motion of a circular random dot pattern (dia IV). Under nor- mal conditions gain was close to 1.0 for the range O.I&16”/sec. For lower stimuius velocities pursuit could no longer be identified in the intersaccadic in- tervals. However. such low velocities elicited a very marked response in open-loop conditions. A stimulus of O.W/sec elicited a response of 5_4’/sec, thus the gain a$proached 100. Gain decreased with the in- crease of stimulus velocity and was close to 1.0 for t6’jsec. the highest stimulus velocity used. Vertical and horizontal stimulation at the average gave rather
0.1 I
Stimulus velocity *WC
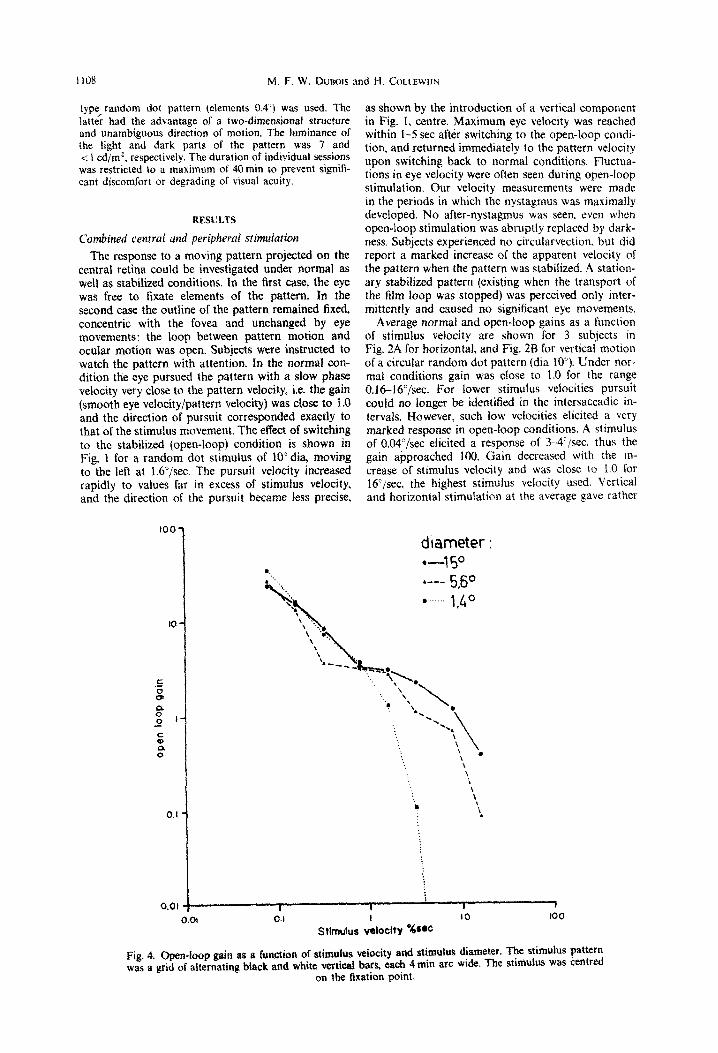
diameter : -4 5O I..-- 5.50 l ._. 1,&O
Fig. 4, Open-loop gain as a function of stimulus vetocity and stimulus diameter. The stimulus pattern was a grid of alternating black and white vettifial bars, each 4 min arc wide, The stbmulus was centred
on the fixation point.

Optokinetic nystagmus in man 1109
similar results, but subjects showed ideosyn- cratic
Peripheral stimulation: central scotomata
For stimulation of the periphery alone, an artificial scotoma was inserted in the projector and stabilized with its centre on the fovea. In this situation, the in- struction to the subject turned out to be crucial. Whereas with central stimulation it was enough that the subject watched and let the eye freely follow the movement of the pattern, for a peripheral stimulus it was essential that the subject directed his attention to the periphery, without really trying to fixate the peripheral image, which was of course impossible. An example is shown in Fig. 5. The stimulus was a grid (alternating black and white vertical bars, each 1” wide) with a size of 20” horizontal x 15” vertical, with
a circular central scotoma of 4” dia. The grid moved at 4”/sec to the right; the outline and scotoma were stabilized on the retina. The subject was inclined to pay attention to the featureless scotoma and in this case the moving periphery elicited only a moderate nystagmus. As soon as the subject switched his atten- tion to the periphery the nystagmus was considerably enhanced. Subjects found it easy to switch their atten- tion and the change in nystagmus appeared consis- tently and with reproducible velocities. In all experi- ments with peripheral stimulation the subject was therefore specifically and repeatedly instructed to attend to the moving pattern, without trying to fixate it.
Even a small central scotoma reduced the slow phase velocity of nystagmus, notwithstanding the sub- ject’s maximum attention to the moving pattern. Figure 6 shows the velocity of the induced smooth eye movements as a function of stimulus velocity in open-loop condition, for a circular random dot pat- tern with a dia of lo”. The continuous line represents the average results of horizontal stimulation in the right and left direction for 3 subjects. It agrees well with the curve shown in Fig. 2. The broken line was obtained in the same sessions after the introduction of a central scotoma of 3” dia. Except for the lowest stimulus velocities the eye velocity was clearly reduced.
The effect of the diameter of the stimulus and of the central scotoma was systematically investigated in 6 subjects, with a circular random dot pattern mov- ing horizontally at 1.6”/sec. In Fig. 7 the averaged results are shown. Values for similar scotoma dia- meters have been connected by lines. The ,upper line shows the responses elicited by patterns of 3.5-25” in diameter without a scotoma. In agreement with the experiment shown in Fig. 4, the effect of stimulus size in this range is very modest. The introduction of a central scotoma with a diameter of 2.5” (triangles) strongly decreased the response to all stimulus dia- meters. The effect of this small scotoma in a 25” stimulus was even stronger than that of decrease of the diameter of the non-scotomatous stimulus from 25 to 3.5”. Enlargement of the scotoma to 5, 10 and 20” dia (as far as compatible with the stimulus dia- meters) progressively reduced the responses further. Figure 7 shows that the most marked decrease of nys- tagmus was caused by the deletion of the central 2.5” of the stimulus, whereas the further decrease with
200
1 down
I -A
2otp t attention periphery
1 attention scotoma
Fig 5. Effect of shifting the attention (on command) between the centre and the periphery of a stabilized pattern with a central scotoma (dia 4”) positioned on the fovea. The stimulus pattern was a grid (alternating black and white vertical bars, each 1” wide) extending 20” horizontally and 15” vertically,
moving at 1.6”/sec to the right.
1110 M. F. W. Duaoa and H. COLLEWIJN
LJ I 0.0 I
I I -1
I IO 100
Stimulus vetocity Ysec
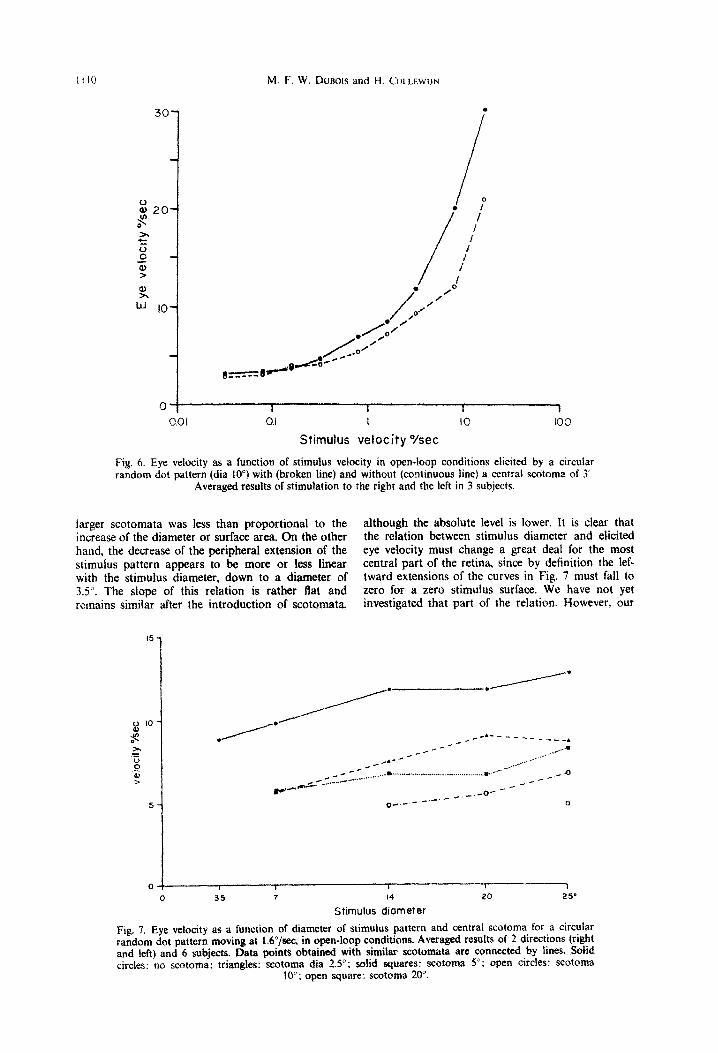
Fig, 6. Eye velocity as a function of stimulus velocity in open-loop conditions elicited by a circular random dot pattern (dia 10”) with (broken line) and without (~ntinuous line) a central scotoma of 3
Averaged results of stimuIat~on to the right and the left in 3 subjects.
larger scotomata was less than proportional to the although the absolute level is lower. It is clear that increase of the diameter or surface area. On the other the relation between stimulus diameter and ehcited hand, the decrease of the peripheral extension of the eye velocity must change a great deal for the most stimulus pattern appears to be more or less linear central part of the retina, since by definition the lef- with the stimulus diameter, down to a diameter of tward extensions of the curves in Fig. 7 must fall to 3.5”. The slope of this relation is rather flat and zero for a zero stimulus surface. We have not yet remains similar after the introduction of scotomata investigated that part of the relation. However, our
0 35 7 14 20 25’
Stimulus diameter
Fig. 7. Eye velocity as a function of diameter of stimulus pattern and central scotoma for a circular random dot pattern moving at l.C”/sec, in open-loop conditions. Averaged results of 2 directions (right and left) and 6 subjects. Data points obtained with similar scotomata are connected by lines. Solid circles: no scotoma: triangles: scotoma dia 2.5”; solid squares: scotoma 5”: open cirdes: scotoma
10”; open square: scotoma 20”.

Optokinetic nystagmus in man 1111
results show that the central few degrees of the visual field contribute far in excess of their relative surface area to the smooth pursuit reponses elicited by a moving pattern.
Spatial distribution of nystugmus with respect to pri- mary position
A series of sessions (in 5 subjects) was specifically devoted to the study of the direction of the fast and smooth components of nystagmus with respect to the primary position. The stimulus was a random dot pattern, 20” wide and 13” high, in which a central scotoma (dia 2.3”) could be inserted. Typical results are shown in Fig. 8 for two subjects, HS and HC. No other instructions were given to the subject than to watch the moving pattern with full attention. When the pattern was moved at lC/sec under normal visual conditions, in the majority of cases the fast com- ponents were directed away from the primary pos- ition, and the slow phases towards it (Fig. 8A, C). As clearly shown in Fig. 8A, reversal of the stimulus di- rection quickly reversed the pursuit direction, fre- quently in the middle of a slow phase. Thus, the change in direction did not typically begin with a fast component.
When the pattern was presented under stabilized (open-loop) conditions and at a lower velocity (l.C/sec) to elicit a nystagmus with comparable vel- ocity, the same spatial organization was maintained (Fig. 8D). Finally, the central scotoma was added to the stimulus. Although a decrease in slow phase vel- ocity occurred, no rearrangement of the nystagmic beats with respect to the primary position was ever observed (Fig. 8B, E). The rule for slow components to be directed towards the primary position was cer- tainly not absolute. One subject behaved in the oppo-
site way, and another showed a symmetrical distribu- tion of nystagmic beats around the midposition in both directions. However, the predominant pattern for a certain subject remained identical under the three circumstances tested.
Eccentric stimulation
In all the experiments described so far, the stimulus pattern was projected symmetrically around the fovea. Although ideosyncratic asymmetries along one or more meridians were revealed in many subjects (Fig. 3), no systematic directional preferences for the sample of subjects as a whole was found. This balance was lost when the stimulus position was no longer symmetrical around the fovea. One general difficulty with the study of the effect of peripheral motion was the tendency of subjects to drift in the direction of the stimulus even when it was stationary, as described by Kommerell and Klein (1971). With the stimulus con- figurations used in our study we found that in the majority of subjects this drift was slower than 1 “/sec.
Figure 9 shows typical examples from a subject with negligible drift when the pattern was stationary. The stimulus was a circular random dot-pattern (dia 15”) moving at 16”/sec to the right and left. With full exposure (Fig. 9A) in stabilized condition the re- sponses were practically symmetrical with slow phase eye velocities of 12S”/sec to the left and 14.4”/sec to the right, as indicated by the arrows. The introduction of a central scotoma (dia 3”) reduced the responses considerably (Fig. 9D), but symmetry was essentially maintained. When the right half of the pattern was obscured with the division line through the fixation point, asymmetrical responses were obtained (Fig. 9B). Movement to the left elicited a stronger response
A Subi. HS normal
Subl.HC normi
open loop + ecotome
Fig. 8. Distribution of nystagmus around the primary position, marked by the interrupted lines. Two subjects: HS (A,B) and HC (C,D,E). Conditions: normal vision of a rectangular random dot pattern (13” vertical x 20” horizontal) moving at 14”/sec (AC); same pattern moving at W/.+x in open-loop conditions (D); same pattern moving at 2.8”/se-c (B) and lA”/sec (E) with a central scotoma of 2.3”
dia. Arrows indicate the moments at which the direction of the stimulus motion was inverted.

1112 M. F‘. W. DLJBOIS and H. COLLEWIJ~
Fig. 9. Responses of one subject to open-loop stimulation to the right and left at 1.6 /set by symmetrical and asymmetrical stimulus configurations in open-loop conditions. The arrows indicate the direction and velocities (also given by the numbers in “/set) of the smooth eye movements. Random dot pattern with dia 15”. Diameter of scotoma (D,E,F) 3”. The centre of the circle was positioned on the fovea.
than movement to the right. This preference was in- verted when the left half of the stimulus was covered (Fig. 9C). In both cases, a pattern movement away from the fovea elicited the strongest response. This directional preference was considerably enhanced by the combination of asymmetry with a central scotoma (Fig. 9E, F). Compared to a symmetrical stimulus with a central scotoma alone (Fig. 9D), the hemifield stimulus elicited a stronger response when moving towards the periphery but a weaker response when moving towards the centre.
The stimulus configuration of Fig. 9E, F (scotoma + half stimulus field) was also rotated in steps of 45” and tested in the vertical and diagonal meri- dians. In all orientations and in all subjects tested it was found that a centrifugal stimulus movement elicited considerably stronger pursuit than a centripe- tal movement. The pursuit was roughly in the direc- tion of the stimulus motion, but errors up to 45” were not unusual. The subjects were often not aware of their asymmetrical responses and after learning about them were unable to influence the asymmetry. In several subjects both eyes were examined. No system- atic differences between the right and left eye were found.
The directional efficiency of eccentric stimuli was further investigated by positioning a circular random dot pattern (dia 10”) in 8 symmetrical positions around the fovea, with the centre of the stimulus always lo” removed from the fovea. Thus, the most central edge of the pattern was located 5” peripher- ally. The pattern was moved towards or away from the fovea. As shown in Fig. 10 for the same subject as in Fig. 9, the relative superiority of centrifugal motion as a stimulus was confirmed in all meridians.
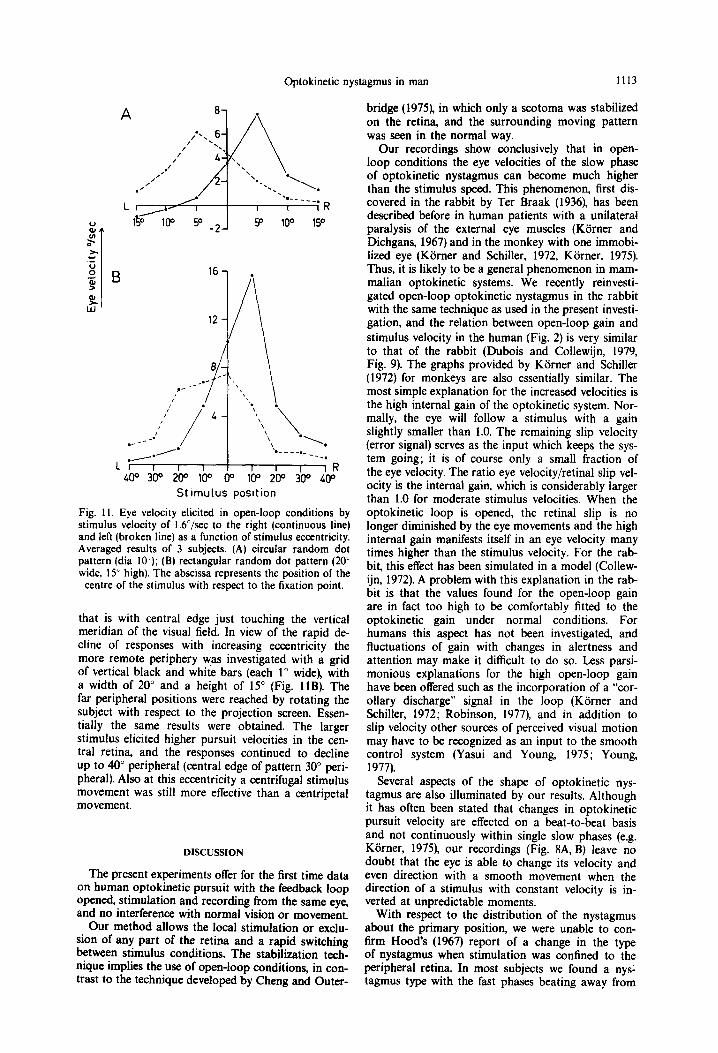
Slow phase velocity as a function of stimulus eccen- tricity was investigated along the horizontal meridian for horizontal stimulus motion. The average results for 3 subjects are shown in Fig. 11. The upper section represents the results obtained with a circular random dot pattern (dia 100) moving at 16”/sec to the right (continuous line). For a central location of the stimu- lus, the responses elicited by movement to the right and left were about equal. In the-periphery, move-
ment to the right elicited a much stronger pursuit when positioned in the right half of the visual field than in the left hemifield. In the last case, responses were negative at 10 and 15“. which means that the eye moved smoothly in the direction of the stimulus location, although the movement of the pattern was in the opposite direction. In this peripheral location, pursuit velocities elicited by the pattern movements were small and could therefore be inverted even by a relatively slow drift. Conversely, pattern movement to the left elicited the best responses when located in the left hemifield, although the average responses happened to be positive in all positions for this direc- tion. Maximum responses for both directions were obtained with the stimulus centred at 5” eccentricity,
42 Pl
Fig. 10. Responses to eccentric stimuli, in open-loop con- ditions. Circular random dot pattern, dia lo”, positioned with centre 10” from the fovea in 8 directions (positions not drawn to scale). Stimulus motion 1.6”/sec towards or away from the fovea. Same subject as in Fig. 9. Smooth eye movement vectors are represented by arrows and
numbers (“/se@.
Optokinetic nystagmus in man 1113
St lmulus posltion
Fig. 1 I. Eye velocity elicited in open-loop conditions by stimulus velocity of l.b”/sec to the right (continuous line) and left (broken line) as a function of stimulus eccentricity. Averaged results of 3 subjects. (A) circular random dot pattern (dia lo’); (B) rectangular random dot pattern (20’ wide, 15” high). The abscissa represents the position of the
centre of the stimulus with respect to the fixation point.
that is with central edge just touching the vertical meridian of the visual field. In view of the rapid de- cline of responses with increasing eccentricity the more remote periphery was investigated with a grid of vertical black and white bars (each 1” wide), with a width of 20” and a height of 15” (Fig. 11s). The far peripheral positions were reached by rotating the subject with respect to the projection screen. Essen- tially the same results were obtained. The larger stimulus elicited higher pursuit velocities in the cen- tral retina, and the responses continued to decline up to 40” peripheral (central edge of pattern 30” peri- pheral). Also at this eccentricity a centrifugal stimulus movement was still more effective than a centripetal movement.
DISCUSSION
The present experiments offer for the first time data on human optokinetic pursuit with the feedback loop opened, stimulation and recording from the same eye, and no interference with normal vision or movement.
Our method allows the local stimulation or excht- sion of any part of the retina and a rapid switching between stimulus conditions. The stabilization tech- nique implies the use of open-loop conditions, in con- trast to the technique developed by Cheng and Outer-
bridge (1975), in which only a scotoma was stabilized on the retina, and the surrounding moving pattern was seen in the normal way.
Our recordings show conclusively that in open- loop conditions the eye velocities of the slow phase of optokinetic nystagmus can become much higher than the stimulus speed. This phenomenon, first dis- covered in the rabbit by Ter Braak (1936). has been described before in human patients with a unilateral paralysis of the external eye muscles (KSrner and Dichgans, 1967) and in the monkey with one immobi- lized eye (Kiirner and Schiller, 1972, Kiirner. 1975). Thus, it is likely to be a general phenomenon in mam- malian optokinetic systems. We recently reinvesti- gated open-loop optokinetic nystagmus in the rabbit with the same technique as used in the present investi- gation, and the relation between open-loop gain and stimulus velocity in the human (Fig. 2) is very similar to that of the rabbit (Dubois and Collewijn, 1979, Fig. 9). The graphs provided by Kiirner and Schiller (1972) for monkeys are also essentially similar. The most simple explanation for the increased velocities is the high internal gain of the optokinetic system. Nor- mally, the eye will follow a stimulus with a gain slightly smaller than 1.0. The remaining slip velocity (error signal) serves as the input which keeps the sys- tem going; it is of course only a small fraction of the eye velocity. The ratio eye velocity/retinal slip vel- ocity is the internal gain, which is considerably larger than 1.0 for moderate stimulus velocities. When the optokinetic loop is opened, the retinal slip is no longer diminished by the eye movements and the high internal gain manifests itself in an eye velocity many times higher than the stimulus velocity. For the rab- bit, this effect has been simulated in a model (Collew- ijn, 1972). A problem with this explanation in the rab- bit is that the values found for the open-loop gain are in fact too high to be comfortably fitted to the optokinetic gain under normal conditions. For humans this aspect has not been investigated, and fluctuations of gain with changes in alertness and attention may make it difficult to do so. Less parsi- monious explanations for the high open-loop gain have been offered such as the incorporation of a “cor- ollary discharge” signal in the loop (Korner and Schiller, 1972; Robinson, 1977), and in addition to slip velocity other sources of perceived visual motion may have to be recognized as an input to the smooth control system (Yasui and Young, 1975; Young, 1977).
Several aspects of the shape of optokinetic nys- tagmus are also illuminated by our results. Although it has often been stated that changes in optokinetic pursuit velocity are effected on a beat-to-beat basis and not continuously within single slow phases (e.g. Kiirner, 1975), our recordings (Fig. 8A, B) leave no doubt that the eye is able to change its velocity and even direction with a smooth movement when the direction of a stimulus with constant velocity is in- verted at unpredictable moments.
With respect to the distribution of the nystagmus about the primary position, we were unable to con- firm Hood’s (1967) report of a change in the type of nystagmus when stimulation was confined to the peripheral retina. In most subjects we found a nysl tagmus type with the fast phases beating away from

1114 bf. F. w. t&JBwS and H. COLLEWIJN
and the slow phases beating towards the primary pos- ition to be dominant. The introduction of central sco- tomata did not change the nystagmus type in any of our subjects, in agreement with the results of Cheng and Outerbridge (1975).
Our findings with regard to the relative power of the fovea and the periphery to induce optokinetic nys- tagmus agree quite well with those of Kiirner and Schiller (1972), Kijrner (1975) and Cheng and Outer- bridge (1975). The central part of the retina was found to be very powerful in eliciting nystagmus, and dele- tion of the central part of a stimulus considerably reduced optokinetic responses. On the other hand, the periphery was found to be quite capable of induc- ing optokinetic reactions. but the effect of moving patterns declined rapidly with increasing eccentricity. Moreover, specific attention towards the peripheral stimulus was required for optimal responses, Cheng and Outerbridge (1975) found a negative effect of sco- tomata only if their diameter was larger than 5”. Poss- ible reasons for the lack of effect of a 5” scotoma in their experiments might be the relatively course texture of the stimulus pattern, or small inaccuracies in the eye position signal, obtained with electro-ocu- lography. Hoods (1975) assertion that exclusion of the periphery strongly impairs optokinetic responses contrasts with our findings. However, Hood used a stimul,us pattern with white vertical stripes at inter- vals of 15’ and with visual angles smaller than 15” the motion stimulus must have considerably deterior- ated.
We found that stimulus motion away from the fovea elicited much stronger optokinetic responses than motion towards the fovea. This directional pre- ference has not been reported before and was not found in the monkey (Korner and Schiller, 1972). It seems related to the eliciting of a smooth pursuit movement by a stationary target, as described by Kommerell and Klein (1971) for after-images. It may serve a highly meaningful function in the foveation of moving targets. When a target moves away from the fovea, pursuit is useful for fovea1 acquisition of the target. When a target is already moving towards the fovea, pursuit in the direction of the target move- ment would only delay foveation, and is therefore better omitted. Our observation that the effect is strongly enhanced by the introduction of a central scotoma (Fig. 9) may indicate that it is a property of the peripheral retina, more than of the fovea. In our experiments it was not possible to rigorously dis- tinguish between pursuit elicited by position and motion of the target. A better distinction might be achieved in future experiments by projecting patterns symmetrically around the fovea, but moving only a part of the pattern.
The preference for centrifugal pursuit can not be easily related to known properties of the human ner- vous system. Since the right hemifield according to our findings would be especially involved in pursuit to the right, one would expect this to be a function of the left hemisphere. However, clinical experience indicates that the opposite is the case (Hoyt and Dar- off, 1971; Troost et al., 1972). Although in the cat a centrifugal arrangement of direction selective units has been found in the superior colliculus (Straschili and Hoffmann, 1968; Dreher and Hoffmann, 1973),
no such organization has been identified in the mon- key (Cynader and Berman, 1972; Marrocco and Li, 1977). In the rabbit, optokinetic pursuit is probably mediated through the pretectal nucleus of the optic tract (Collewijn, 1975) and each eye (and the contrala- teral nucleus of the optic tract) has a strong prefer- ence to respond to patterns moving in the anterior direction. Since the rabbit’s optokinetic reactions are essentially subcortically organized, and the animal is afoveate, no detailed comparison with the present findings in the human can be made, in spite of some global similarities.
Finally, it should be emphasized that the optokine” tic responses in human subjects are far from mechani- cal and show considerable variation in time and between subjects. The importance of attention has been reported before (Kiirner and Dichgans, 1967) also in particular for the reponse to peripheral targets (Cheng and Outerbridge, 1975). The required atten- tion apparently involves the selective processing of localized parts of the visual input. The importance of selection in the response to peripheral targets should hardly come as a surprise, since it must be fundamental to all voluntary eye movements. At the same time it is a serious limitation of the control that an investigator can exert on the experimental situation, and of the precision and reliability of the data obtained.
Acknowledgements-The investigations were supported in Dart bv The Foundation for Medical Research FUNGO which-is subsidized by the Netherlands Organization for the Advancement of Pure Research (Z.W.O.). We thank all the subjects who participated in the experiments.
REFERENCES
Cheng M. and Outerbridge J. S. (1975) Optokinetic nys- tagmus during selective retinal stimulation. Expl. Bruin Res. 23, 129-139.
Collewijn H. (1972) An analog model of the rabbit’s opto- kinetic system. Brain Res. 36, 71-88.
Colbwijn H., Van der Mark F. and Jansen T. C. (1975) Precise recording of human eye movements. Vision Res. 15,447-&o.
Cynader M. and Berman N. (1972) Receptive-field organ- ization of monkey superior colliculus. J. Neurophysiol. 35, 187-201. _ -
Dreher B. and Hoffman K. P. (1973) Properties of excita- tory and inhibitory regions‘in the receptive fields of single units in the cat’s superior colliculus. Expl Bruin Res. 16, 333-353.
Dubois M. F. W. and Collewijn H. (1979) The optokinetic reactions of the rabbit: relation to the visual streak. Vision Res. 19, 9-17.
Hood J. D. (1967) Observations upon the neurological mechanism of optokinetic nystagmus with especial refer- ence to the contribution of peripheral vision. Acra oto- lar. 63 208-215.
Hood J. b. (1975) Observations upon the role of the peri- pheral retina iu the execution of eye movements. J. Oto- rhino-lar. Borderlands 37, 65-73.
Hoyt W. F. and Daroff R. B. (1971) Supranuclear disorders of ocular control systems in man. In The control of eye mouemenrs (edited by Bach-y-Rica Y. P. and Collins C. C.). pp. 175-235. Academic Press, New York.
Julesz B. (1964) Binocular depth perception without fami- liarity cues. Science 145, 356-362.

Optokinetic nystagmus in man 1115
Kommerell G. and Klein U. (1971) ober die visuelle Rege- R. and Berthoz A.), pp. 49-58. Elsevier-North Holland, Amsterdam. _ lung der Okulomotorik: die optomotorische Wirkuig
exzentrischer Nachbilder. Vision Res. 11, 905-920. KGrner F. (1975) Untersuchungen ilber die nichtvisuelle
Kontrolle von Augenbewegungen. Adv. Ophtha1. 31, 100-158.
KBrner F. and Dichgans J. (1967) Bewegungswahrneh- mung, optokinetischer Nystagmus und retinale Bildwan- derung. Albrecht v. Graefes Arch. klin. exp. Ophthal. 174, 34-48.
Kiirner F. and Schiller P. H. (1972) The optokinetic re- sponse under open and closed loop conditions in the monkey. Expf Bruin Res. 14, 318-330.
Marrocco R. T. and Li R. H. (1977) Monkey superior colli- culus: properties of single cells and their afferent inputs. J. Neurophys~o~. 40, 844-860.
Puckett J. W. and Steinman R. M. (1969) Tracking eye movements with and without saccadic correction. Vision Res. 9, 695-703.
Robinson D. A. (1963) A method of measuring eye move- ment using a scleral search coil in a magnetic field. IEEE Trans. Homed. Electron. BME-10, 137-145.
Robinson D. A. (1977) Vestibular and optokinetic sym- biosis: an example of explaining by modelling. In Con- trol of Gaze by Bruin Stem Neurons (edited by Baker
Steinman R. M.. Skavenski A. A. and Sansburv R. V. 111969) Voluntary control of smooth pursuit veiociiy. Vision Res: 9, 1167-1171.
Straschill M. and Hoffman K. P. (1968) Relationship between localization and functional properties of move- ment-sensitive neurons of the cat’s tectum opticum. Brain Res. 8, 382-385.
Ter Braak J. W. G. (1936) Untersuchungen iiber optokine- tischen Nystagmus. Archs net;& Physiol. 21, 309-376.
Troost B. T., Daroff R. B., Weber R. B. and Dell’Osso L. F. (1972) Hemispheric control of eye movements. II. Quantitative analysis of smooth pursuit in a hemispher- ectomy patient. Archs Neural. 27, 449-452.
Winterson B. J. and Steinman R. M. (1978) The effect of luminance on human smooth pursuit of perifoveal and fovea1 targets. Vision Res. 18, 1165-l 172.
Yasui S. and Young L. R. (1975) Perceived visual motion as effective stimulus to pursuit eye movement system. Science 190, 906-908.
Young L. R. (1977) Pursuit ‘eye movement-what is being pursued? In Control ,of Gaze by Brain Stem Neurons (edited by Baker R. and Berthoz A.), pp. 29-36. Elsevier- North Holland, Amsterdam.