ontogeny of trophic morphology in four sympatric morphs of arctic charr salvelinus alpinus in...
TRANSCRIPT

CHARR
Discrete prey availability promotes foraging segregationand early divergence in Arctic charr, Salvelinus alpinus
Monica V. Garduno-Paz • Colin E. Adams
Received: 22 July 2009 / Revised: 28 November 2009 / Accepted: 14 December 2009 / Published online: 6 January 2010! Springer Science+Business Media B.V. 2010
Abstract Many animal species show individual
foraging specialisms when potential prey requiresprey-specific foraging strategies. Arctic charr are
often found as benthic (macroinvertebrate) or pelagic
(plankton) foraging specialists. Here, we testedspecifically if given a choice of prey with different
characteristics individuals would specialise in asingle prey type and if individuals would chose prey
based on their expressed trophic morphology, in a
laboratory experiment and in a field observation.When offered a choice of benthic and pelagic prey
most individuals (73%) showed that 100% fidelity to
a single foraging source. Naıve individuals (notpreviously exposed to natural prey) with more robust
head and mouth shape were more likely to forage on
a benthic prey source (chironomids). In contrast,individuals with a more fusiform body, larger eye, but
more slender head shape were more likely to
specialise on pelagic prey (Artemia). Field observa-tions of a natural population of Arctic charr from
Loch Doine identified specialists foraging on either
plankton or macrobenthos (on the basis of stomach
contents) and some generalists. Morphological anal-ysis showed that significant differences in shape
reflecting recent foraging history. These results
support the hypothesis that the availability of dis-crete, different prey types results in discrete foraging
specialisms which in turn may result in the expressionof discrete alternative phenotypes through subsequent
plastic ontogenetic process. We conclude that this
provides a partial explanation for why ecologicallydriven evolution processes are particularly prevalent
in fishes from post-glacial lake systems.
Keywords Foraging specialism ! Alternative prey !Trophic phenotype ! Discrete prey
Introduction
It is now clear that ecological processes play a critical
role in the evolutionary divergence that may ultimatelylead to the formation of new species. This is particu-
larly true when divergence occurs in sympatry and in
novel environments (Schluter 1996, 2001). Althoughthe mechanisms through which ecological processes
may drive evolutionary change are far from fully
understood, there have been a number of descriptiveand mathematical models that invoke competition for
resources (particularly food) as the principal a driver
for evolutionary divergence (West-Eberhard, 1989;Dieckmann & Doebeli, 1999; Skulason et al., 1999).
Guest editors: C. Adams, E. Brannas, B. Dempson,R. Knudsen, I. McCarthy, M. Power, I. Winfield /Developments in the Biology, Ecology and Evolution of Charr
M. V. Garduno-Paz (&) ! C. E. AdamsScottish Centre for Ecology and the Natural Environment,University of Glasgow, Rowardennan,Glasgow G630AW, Scotland, UKe-mail: [email protected]
123
Hydrobiologia (2010) 650:15–26
DOI 10.1007/s10750-009-0055-8

In these models, evolutionary divergence begins withbehavioural specialisms in prey choice, which may, in
turn, be shaped by opportunities to use the resources
(Skulason & Smith, 1995). Foraging specialisms maythen result in morphological change, through diet-
induced phenotypic plasticity, that may in turn result in
increased foraging efficiency and therefore reinforcethe foraging specialism (Robinson & Parsons, 2002;
Adams et al., 2003; Michaud et al., 2008). Subsequent
steps require assortative mating through either femalechoice or differential habitat use (Dieckmann &
Doebeli, 1999; Skulason et al., 1999).
Dietary specialisations amongst individuals of thesame species (a pre-requisite for evolutionary diver-
gence under many of these models) are relatively
common (Lu & Bernatchez, 1999; Bolnick et al.,2003; Maerz et al., 2006; Stuart et al., 2006; Michaud
et al., 2008; Woo et al., 2008). In some species
expression of foraging specialisation can be extremeand discrete, taking the form of discontinuous
phenotypes (trophic polymorphisms sensu Skulason
& Smith, 1995) with a functional significance forforaging, prey detection, capture or handling (Adams
& Huntingford, 2002a; Smith & Skulason, 1996;
Schmidt et al., 2006; Januszkiewicz & Robinson,2007; Malaquias et al., 2009).
For freshwater fish inhabiting post-glacial lake
systems, the most abundant potential foragingresources are typically discrete, most usually com-
prising planktonic and macrobenthic prey items
occupying different habitats, the limnetic and littoralbenthic zones, respectively (Ostbye et al., 2005;
Kahilainen & Ostbye, 2006). There are many exam-
ples of individuals within a single species specialisingin different prey types (Baumgartner, 1992; Reilly
et al., 1992; Larson & McIntire, 1993; Snorrason
et al., 1994; Kristjansson et al., 2002; Yonekura et al.,2002; Uchii et al., 2007; Swanson et al., 2008).
Within the three-spined stickleback, Gasterosteusaculeatus many populations contain individuals spe-cialising in benthic or limnetic foraging and express-
ing alternative phenotypes. The limnetic formspecialises in foraging on zooplankton, it has a
slender body, long, numerous and densely spaced gill
rakers, whereas the more robust benthic form isspecialised for feeding on larger food items having
less numerous, shorter and widely spaced gillrakers
(Foster et al., 1992; McPhail, 1992; Bell & Foster,1994; Cresko & Baker, 1996; Baker et al., 2005).
Arctic charr, Salvelinus alpinus exhibit similarsympatric trophic specialisations. Most frequently,
this takes the form of a benthic foraging specialist
feeding on relatively large macroinvertebrates and apelagic foraging specialist feeding on pelagic prey
(Skulason et al., 1989; Malmquist et al., 1992; Adams
et al., 1998; Alekseyev et al., 2002; Klemetsen et al.,2003; Fraser et al., 2008). In Arctic charr, as in three-
spined sticklebacks and other fishes, expressed var-
iation in morphology is known to have a functionalsignificance (Smits et al., 1996; Adams & Hunting-
ford, 2002b; Hjelm et al., 2003; West-Eberhard,
2005; Knudsen et al., 2006; Amundsen et al., 2008).In a number of populations of several species, it
has been shown that expression of alternative
phenotypes seen in the wild is wholly or partlyenvironmentally induced through plastic effects on
phenotype during ontogeny (Queral-Regil & King,
1998; Mittelbach et al., 1999; Starck, 1999; Alexander& Adams, 2000; Hegrenes, 2001; Hjelm et al., 2001;
Adams et al., 2003; Hjelm et al., 2003; Wintzer &
Motta, 2005; Olsson et al., 2007; Ruehl & Dewitt,2007; Ke et al., 2008). Thus, although there is good
evidence for the effect of trophic specialism having a
subsequent effect on expressed phenotype throughphenotypic plasticity, little is known about how the
phenotype expressed by an individual may influence
the direction and degree of foraging specialisation.Here, using Arctic charr, a species which is known
to exhibit foraging specialisms and discrete trophic
phenotypes (most notably plankton and macroinver-tebrate feeding specialism), we test in laboratory and
field observations, the degree to which individuals
from a monomorphic population exhibit foragingspecialisations and the extent to which small varia-
tions in morphology determine prey choice in indi-
viduals exposed to alternative prey. Specifically, wetest two hypotheses: (a) given a binary choice of prey
with different characteristics individuals will special-
ise in one prey type, (b) individuals will chose preybased on their expressed trophic morphology.
Methodology
Behavioural observations were carried out using
Arctic charr fry supplied by a commercial hatchery
(John Eccles Hatcheries, Orkney, UK). These fish hadbeen reared in captivity for at least three generations,
16 Hydrobiologia (2010) 650:15–26
123

but with occasional out crossing to first generationwild fish. The stock was originated from two Scottish
Arctic charr populations (Loch Luchart and Loch
Tay). Fish (10 months old, 47–83 mm standardlength) were held in 1 m tangential flow, through-
flow tanks at temperatures between 16 and 18"C and
ambient light, (56"N). They were fed on standardaquaculture pellet food from first feeding until the
experiments started.
Photographs and diet information (stomach con-tents) of specimens of Arctic charr from Loch Doine
collected from an earlier study (Adams et al., 2006)
were used in the present study. Loch Doine is locatedin one arm of the Forth catchment (56"200N,4"200W), which drains a large part of east-central
Scotland eastwards to the North Sea. It has an area of0.54 km2 with 20 m of maximum depth (for more
details, see Adams et al., 2006).
Behavioural trials
A total of 72 juvenile fish were anaesthetized withbenzocaine, marked by Panjet injection in the fins
using Alcian Blue and photographed individually on
the left side for shape analysis. Twelve specimenswereallocated to each of three 500 l (74 cm 9 71 cm 9
95 cm) observation tanks, with no substratum or
vegetation and a constant flow of water. Fish wereinitially acclimatised to the tank and deprived of food
for 3 days to allow them to recover after marking.
At the beginning of each trial, each observationtank was given two discretely different types of prey
at the same time. In order to simulate a pelagic prey
source, Artemia sp. (3–6 mm) were embedded in anice block of 15 9 15 9 4 cm which was floated on
the surface of the water in the right side of the tank.
The ice maintained the Artemia in the surface waterof the tank and allowed a slow release of the Artemiaprey as it defrosted. In order to further prevent
Artemia dropping to the bottom of the tank, atransparent plastic container was fixed 20 cm below
the Artemia food source. The container did notobstruct the movements of the fish and since it was
transparent, the fish were able to observe the Artemiaeasily. In order to simulate a typical benthic prey,chironomid larvae (8–13 mm) were inserted into agar
contained in a Petri dish which was set on the bottom
at the left side of the tank. The agar prevented theprey dispersing in the water current. Prey items were
available throughout the observation trials to preventcompetition between fish due to the lack of food.
Observations were made once a day between 9:00
and 13:00 h over five continuous days. A focalanimal approach was taken, where each individual
fish was observed for 3 min. During each observation
period, the type of prey chosen and number of preyswallowed were recorded.
Diet determination
Information about diet of the fish from Loch Doine
was obtained from Adams et al. (2006). The meth-odology that they used to determine diet was as
follows: stomach contents from approximately 35
charr (mean fork length 169 mm; range 88–251 mm)captured during July using multi-mesh (Norden) gill
nets were removed and placed in 70% ethanol.
Subsequently, prey items were identified, at least tofamily, and counted. Only invertebrate prey items
were found in the stomach contents of fish. In order to
determine the origin of the foraging resource beingexploited, the habitat of origin of each prey group
was categorised as benthic or pelagic (plankton) and
the proportion of prey items originating from benthicand pelagic sources was calculated for each individ-
ual fish (for more details, see Adams et al., 2006).
Geometric morphometrics analysis
In order to determine morphological characteristics,photographs in lateral view of each fish were used.
The photographs of the fish used for experimental
observations included head and body. Meanwhile,photographs of the charr from Loch Doine only
included the head of the fish (see Adams et al., 2006).
Homologous landmarks (22 on fish in the labora-tory experiment and 11 on fish from Loch Doine,
Figs. 1 and 2, respectively) were identified and
placed on the photographs using the software TPS-dig2 (Rohlf, 2006). Landmark configurations for each
specimen were aligned, translated, rotated and scaledto a unit centroid size (CS) using Generalised
Procrustes Analysis superimposition (GPA, Rohlf
and Slice, 1990) using the consensus configuration ofall specimens as the mean shape. Following GPA,
new shape variables, i.e. Partial Warps (PW), were
obtained. In order to explore the overall, within-sample form variability, relative warp analysis,
Hydrobiologia (2010) 650:15–26 17
123

equivalent to principal component analysis for mor-phometric data, was performed on the PW scores,
using the software TPSrelw (Rohlf, 2007).
The CS, which equals the square root of thesummed squared distances of each landmark from the
centroid of that landmark configuration was used as a
measure of fish size. It is determined separately fromshape, and is uncorrelated with shape in the absence
of allometry. This independence from shape is one of
the main reasons why CS is used as a size variable.The other reason is that CS has a crucial role in
defining the metric for a distance between two shapes
(Zelditch et al., 2004). This variable was measured incentimetres, since all the pictures had a scale factor.
Therefore, this variable it is used in the statistical
analyses as any other, normally distributed traditionalvariable.
Statistical analysis
In order to look for general differences between
foraging groups, a MANOVA test was run using all
the PWs scores. The potential effect of size on
morphology of observed fish used during the behav-ioural trials was reduced analysing only individuals in
each of the three foraging groups that overlapped in
size (within the range 8.4–10.7 cm of CS). Discrim-inant analysis of shape data (first five relative warps)
was used to look for the correct assign of individuals
to their correct foraging group (based on behaviouralobservations). In order to compare between foraging
groups, One-way ANOVA tests were carried out on
the three main relative warps (Principal Componentsin geometric morphometrics that explain most of the
variation) and behavioural data (prey consumed).
Post-hoc tests with Bonferroni corrections wereperformed for the first three relative warps and CS.
A one sample t-test was used to compare the
percentage (arc-sine transformed) of prey itemsconsumed within the generalists foraging group.
Simple regressions were used to describe relation-ships amongst behavioural, diet and morphological
variables.
Results
Behavioural trials
A significant number of individuals showed that a
strong (100%) preference for feeding on only one
prey type. Of the 72 fish observed, 39 chose to feedonly on chironomids and 12 only on Artemia.Chironomid specialists, Artemia specialists and those
that switched foraging sources (hereafter calledforaging generalists) showed that significant differ-
ences in the mean total number of prey consumed for
all fish over all days (F2,71 = 37.8; P = 0.0008).
Fig. 1 Location of 22 landmarks in juvenile Arctic charr fromhatchery stock. 1: Tip of the snout, 2–3: width of inferior lip;4–5: width of maxillary bone; 6: nostril position; 7 and 9:horizontal diameter of the eye; 8 and 10: vertical diameter of
the eye; 11: end of the jaw; 12: junction of the body and theoperculum; 13: division of the operculum; 14: dorsal fin; 15:pelvic fin; 16: adipose fin; 17: anal fin; 18–19: caudal pedunclewidth; 20: end of caudal peduncle
Fig. 2 Location of 11 landmarks on the head of Arctic charrfrom Loch Doine. 1: Tip of the snout; 2–4: horizontal diameterof the eye; 3–5: vertical diameter of the eye; 6: division of theoperculum; 7: pectoral fin position; 8: end of the maxillarybone; 9: end of the jaw; 10: end of the operculum division; 11:upper end of the head
18 Hydrobiologia (2010) 650:15–26
123

Chironomid specialists took the greatest number ofprey (significantly more than both Artemia specialists
and prey generalists, post hoc testing P\ 0.0001).
The number of prey consumed by Artemia specialistsand generalists was not significantly different
(P = 0.9).
A total of 21 individuals fed on both foragingsources at least once (foraging generalists), this group
did not show a difference between the number of prey
items consumed from each source (t20 = 0.61;P = 0.5).
Stomach contents
Based on the stomach contents found in 35 fish from
Loch Doine, three foraging groups were determined.Fifteen individuals were assigned as benthic special-
ists on the basis of 100% benthic prey in their stomach
(chironomid larvae were the most common prey).Fourteen were designated as pelagic specialists,
having 100% pelagic prey in their stomach (Cladocera
and copepods were the most common prey), and sixindividuals were considered as generalists because the
stomach content was represented by both types of prey
[i.e. chironomid larvae, Cladocera, copepod, Dipteraand pea mussel (Pisidium tenuilineatum)].
In contrast to the laboratory experiment, benthic
specialists from Loch Doine had the smallest numberof prey (significantly less than both pelagic specialists
and prey generalists, post-hoc testing P\ 0.0001).
Whereas, the number of prey contained in thestomach of pelagic specialists and generalists was
not significantly different (P = 0.8).
Morphological analysis
In total, 42 fish within the range 8.4–10.7 cm of CSwere used, 12 generalists, 10 Artemia and 20
chironomid consumers. A discriminant analysis of
the first five relative warps (shape variables) derivedfrom geometric morphometric analyses correctly
assigned 88% of individuals to their correct foraginggroups (based on behavioural observations), 16 of 20
(80%) chironomid specialists, 10 of 10 (100%)
Artemia specialists and 11 of 12 (92%) generalists.MANOVA analysis run for all relative warps
showed that there were significant differences
between feeding behaviour groups in relative warpscores (Wilk’s K = 0.27, F2,39 = 11.5; P = 0.001).
Relative warp analysis resulted in three main com-
ponents that together represent the 57.2% of the totalshape variation. One-way ANOVA analysis for each
relative warp showed that significant differences
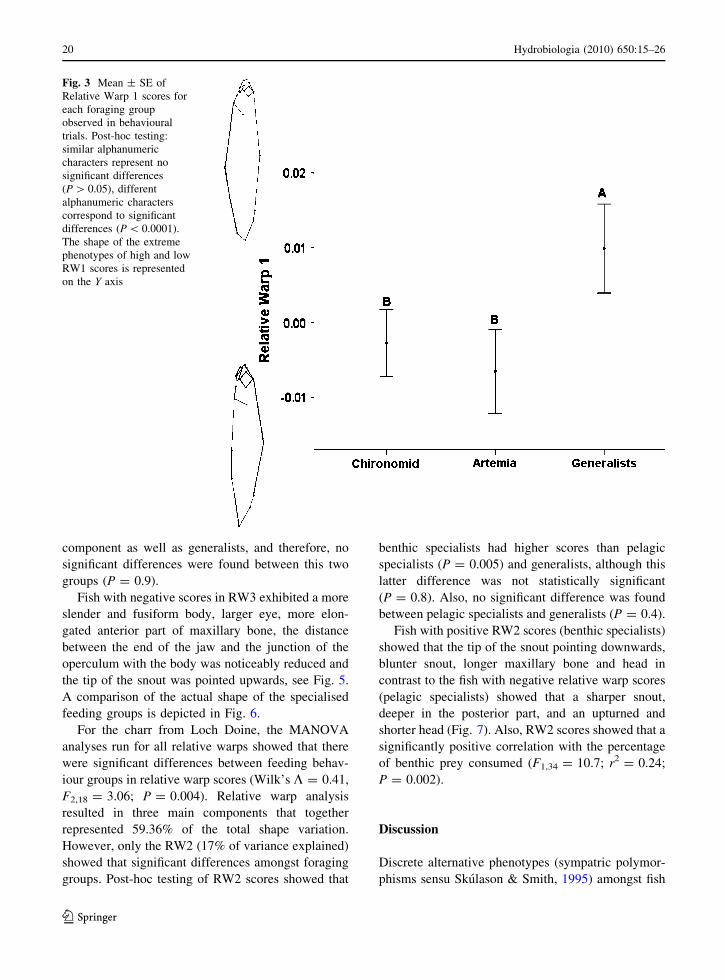
amongst foraging groups (Table 1).Post-hoc testing showed that generalists had
significant higher RW1 scores than both Artemiaspecialists (P = 0.001) and chironomids specialists(P = 0.003), however, the latter two were not
different from each other (P = 0.97).
Fish with positive scores for RW1 showed that areduced head, shorter maxillary bone, a smaller eye,
also a ventral expansion is perceptible, the posterior
section of the body and the head are relativelyupturned in contrast to the fish with extreme negative
relative warp scores (Fig. 3).
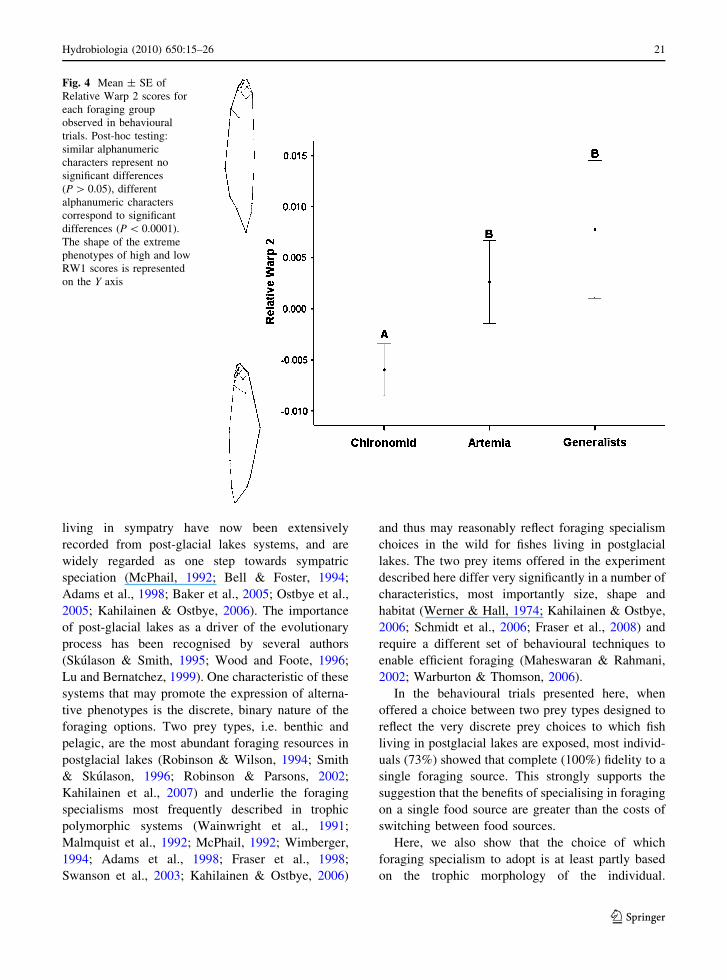
For RW2, post-hoc testing showed that generalistshad significantly higher scores than chironomid
specialists (P = 0.0001) and slightly higher scores
than Artemia specialists, but not significantly so(P = 0.4), whilst significant difference was present
between Artemia and chironomid feeders (P = 0.02).
In the second relative warp, fish with positive scoreshad a more pronounced, deeper body in the posterior
ventral area, the distance from the anal fin to the end
of the caudal peduncle was longer, the head waspointed upwards and the tip of the snout was blunt in
contrast to fish with negative scores which were
dorsally curved, presented an anterior elongation ofthe maxillary bone, the snout was slightly sharp and
the end of the caudal peduncle was turned down
(Fig. 4). Also, only this component, RW2, wassignificantly negatively correlated with the percent-
age of benthic prey consumed (F1,41 = 10; r2 = 0.2;
P = 0.003).Post-hoc testing of RW3 also showed that scores
were significantly lower for Artemia specialists
compared with chironomid specialists (P = 0.03)and generalists (P = 0.04). Chironomid specialists
were distributed in the positive extreme of this
Table 1 General Linear Model comparing relative warpsscores between all foraging groups observed within the fishfrom hatchery stock
RW % Variance explained F SE Sig. (P value)
1 24.3 9.04 0.0099 0.0006
2 17.7 11.8 0.0080 0.0001
3 15.2 4.5 0.0084 0.018
Hydrobiologia (2010) 650:15–26 19
123
component as well as generalists, and therefore, nosignificant differences were found between this two
groups (P = 0.9).
Fish with negative scores in RW3 exhibited a moreslender and fusiform body, larger eye, more elon-
gated anterior part of maxillary bone, the distance
between the end of the jaw and the junction of theoperculum with the body was noticeably reduced and
the tip of the snout was pointed upwards, see Fig. 5.
A comparison of the actual shape of the specialisedfeeding groups is depicted in Fig. 6.
For the charr from Loch Doine, the MANOVA
analyses run for all relative warps showed that therewere significant differences between feeding behav-
iour groups in relative warp scores (Wilk’s K = 0.41,
F2,18 = 3.06; P = 0.004). Relative warp analysisresulted in three main components that together
represented 59.36% of the total shape variation.
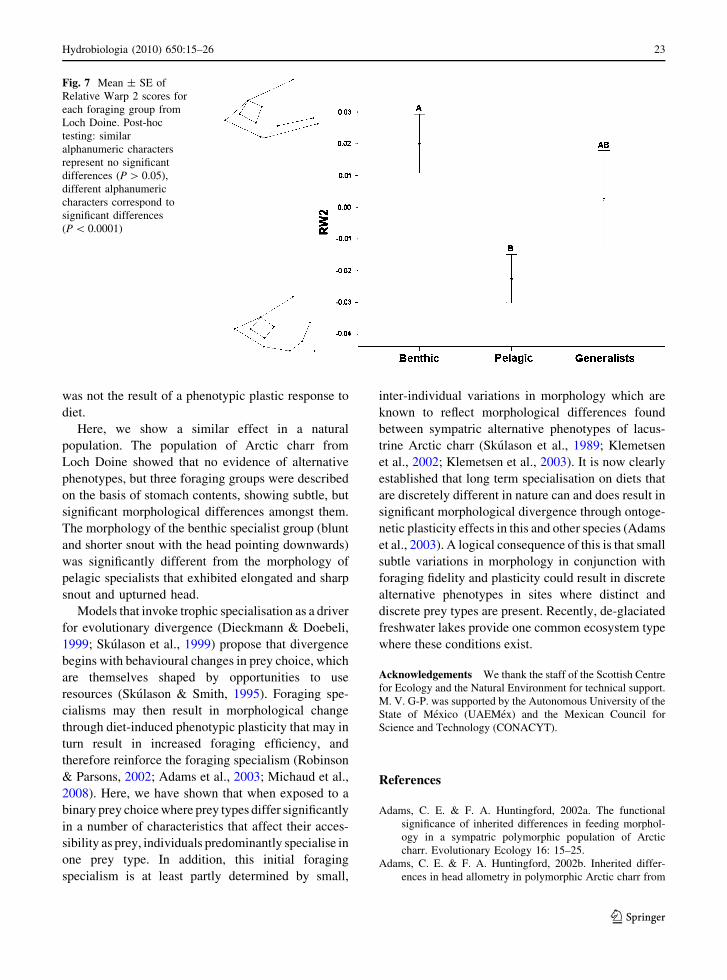
However, only the RW2 (17% of variance explained)showed that significant differences amongst foraging
groups. Post-hoc testing of RW2 scores showed that
benthic specialists had higher scores than pelagicspecialists (P = 0.005) and generalists, although this
latter difference was not statistically significant
(P = 0.8). Also, no significant difference was foundbetween pelagic specialists and generalists (P = 0.4).
Fish with positive RW2 scores (benthic specialists)
showed that the tip of the snout pointing downwards,blunter snout, longer maxillary bone and head in
contrast to the fish with negative relative warp scores
(pelagic specialists) showed that a sharper snout,deeper in the posterior part, and an upturned and
shorter head (Fig. 7). Also, RW2 scores showed that a
significantly positive correlation with the percentageof benthic prey consumed (F1,34 = 10.7; r2 = 0.24;
P = 0.002).
Discussion
Discrete alternative phenotypes (sympatric polymor-
phisms sensu Skulason & Smith, 1995) amongst fish
Fig. 3 Mean ± SE ofRelative Warp 1 scores foreach foraging groupobserved in behaviouraltrials. Post-hoc testing:similar alphanumericcharacters represent nosignificant differences(P[ 0.05), differentalphanumeric characterscorrespond to significantdifferences (P\ 0.0001).The shape of the extremephenotypes of high and lowRW1 scores is representedon the Y axis
20 Hydrobiologia (2010) 650:15–26
123
living in sympatry have now been extensively
recorded from post-glacial lakes systems, and are
widely regarded as one step towards sympatricspeciation (McPhail, 1992; Bell & Foster, 1994;
Adams et al., 1998; Baker et al., 2005; Ostbye et al.,
2005; Kahilainen & Ostbye, 2006). The importanceof post-glacial lakes as a driver of the evolutionary
process has been recognised by several authors
(Skulason & Smith, 1995; Wood and Foote, 1996;Lu and Bernatchez, 1999). One characteristic of these
systems that may promote the expression of alterna-
tive phenotypes is the discrete, binary nature of theforaging options. Two prey types, i.e. benthic and
pelagic, are the most abundant foraging resources in
postglacial lakes (Robinson & Wilson, 1994; Smith& Skulason, 1996; Robinson & Parsons, 2002;
Kahilainen et al., 2007) and underlie the foraging
specialisms most frequently described in trophicpolymorphic systems (Wainwright et al., 1991;
Malmquist et al., 1992; McPhail, 1992; Wimberger,
1994; Adams et al., 1998; Fraser et al., 1998;Swanson et al., 2003; Kahilainen & Ostbye, 2006)
and thus may reasonably reflect foraging specialism
choices in the wild for fishes living in postglacial
lakes. The two prey items offered in the experimentdescribed here differ very significantly in a number of
characteristics, most importantly size, shape and
habitat (Werner & Hall, 1974; Kahilainen & Ostbye,2006; Schmidt et al., 2006; Fraser et al., 2008) and
require a different set of behavioural techniques to
enable efficient foraging (Maheswaran & Rahmani,2002; Warburton & Thomson, 2006).
In the behavioural trials presented here, when
offered a choice between two prey types designed toreflect the very discrete prey choices to which fish
living in postglacial lakes are exposed, most individ-
uals (73%) showed that complete (100%) fidelity to asingle foraging source. This strongly supports the
suggestion that the benefits of specialising in foraging
on a single food source are greater than the costs ofswitching between food sources.
Here, we also show that the choice of which
foraging specialism to adopt is at least partly basedon the trophic morphology of the individual.
Fig. 4 Mean ± SE ofRelative Warp 2 scores foreach foraging groupobserved in behaviouraltrials. Post-hoc testing:similar alphanumericcharacters represent nosignificant differences(P[ 0.05), differentalphanumeric characterscorrespond to significantdifferences (P\ 0.0001).The shape of the extremephenotypes of high and lowRW1 scores is representedon the Y axis
Hydrobiologia (2010) 650:15–26 21
123

Individuals with a more robust, blunter head and
bigger mouth with a more ventral position of the headwere more likely to forage on the benthic prey source
(chironomids) than on pelagic prey. In contrast,
individuals characterised by a slender and fusiformbody, bigger eye, but slightly small body size were
more likely to be pelagic prey (Artemia) than benthic
prey specialists. Overall, generalists did not feedsignificantly more on benthic prey than pelagic prey
and showed that morphological differences from the
two specialist groups: shorter maxillary bone, asmaller eye, ventral expansion, with the posterior
section of the body and the head relatively upturned.
This phenotypic variation amongst individuals fromthis population is the result of a natural, continuous
variation in morphological characteristics, because
morphology was measured before fish were exposed tothe experimental conditions. Consequently, variation
Fig. 5 Mean ± SE ofRelative Warp 3 scores foreach foraging groupobserved in behaviouraltrials. Post-hoc testing:similar alphanumericcharacters represent nosignificant differences(P[ 0.05), differentalphanumeric characterscorrespond to significantdifferences (P\ 0.0001).The shape of the extremephenotypes of high and lowRW1 scores is representedon the Y axis
Fig. 6 Shape of the Arctic charr individuals observed inbehavioural trials. Landmarks indicate the Artemia feeders’mean shape and vectors indicate chironomid feeders’ mean
shape as a deformation from the Artemia feeders’ mean shape.Landmarks are connected by links to facilitate the visualisationshape
22 Hydrobiologia (2010) 650:15–26
123
was not the result of a phenotypic plastic response to
diet.Here, we show a similar effect in a natural
population. The population of Arctic charr fromLoch Doine showed that no evidence of alternative
phenotypes, but three foraging groups were described
on the basis of stomach contents, showing subtle, butsignificant morphological differences amongst them.
The morphology of the benthic specialist group (blunt
and shorter snout with the head pointing downwards)was significantly different from the morphology of
pelagic specialists that exhibited elongated and sharp
snout and upturned head.Models that invoke trophic specialisation as a driver
for evolutionary divergence (Dieckmann & Doebeli,
1999; Skulason et al., 1999) propose that divergencebegins with behavioural changes in prey choice, which
are themselves shaped by opportunities to use
resources (Skulason & Smith, 1995). Foraging spe-cialisms may then result in morphological change
through diet-induced phenotypic plasticity that may in
turn result in increased foraging efficiency, andtherefore reinforce the foraging specialism (Robinson
& Parsons, 2002; Adams et al., 2003; Michaud et al.,
2008). Here, we have shown that when exposed to abinary prey choicewhere prey types differ significantly
in a number of characteristics that affect their acces-
sibility as prey, individuals predominantly specialise inone prey type. In addition, this initial foraging
specialism is at least partly determined by small,
inter-individual variations in morphology which are
known to reflect morphological differences foundbetween sympatric alternative phenotypes of lacus-
trine Arctic charr (Skulason et al., 1989; Klemetsenet al., 2002; Klemetsen et al., 2003). It is now clearly
established that long term specialisation on diets that
are discretely different in nature can and does result insignificant morphological divergence through ontoge-
netic plasticity effects in this and other species (Adams
et al., 2003). A logical consequence of this is that smallsubtle variations in morphology in conjunction with
foraging fidelity and plasticity could result in discrete
alternative phenotypes in sites where distinct anddiscrete prey types are present. Recently, de-glaciated
freshwater lakes provide one common ecosystem type
where these conditions exist.
Acknowledgements We thank the staff of the Scottish Centrefor Ecology and the Natural Environment for technical support.M. V. G-P. was supported by the Autonomous University of theState of Mexico (UAEMex) and the Mexican Council forScience and Technology (CONACYT).
References
Adams, C. E. & F. A. Huntingford, 2002a. The functionalsignificance of inherited differences in feeding morphol-ogy in a sympatric polymorphic population of Arcticcharr. Evolutionary Ecology 16: 15–25.
Adams, C. E. & F. A. Huntingford, 2002b. Inherited differ-ences in head allometry in polymorphic Arctic charr from
Fig. 7 Mean ± SE ofRelative Warp 2 scores foreach foraging group fromLoch Doine. Post-hoctesting: similaralphanumeric charactersrepresent no significantdifferences (P[ 0.05),different alphanumericcharacters correspond tosignificant differences(P\ 0.0001)
Hydrobiologia (2010) 650:15–26 23
123

Loch Rannoch, Scotland. Journal of Fish Biology 60:515–520.
Adams, C. E., D. Fraser, F. A. Huntingford, R. B. Greer, C. M.Askew & A. F. Walker, 1998. Trophic polymorphismamongst Arctic charr from Loch Rannoch, Scotland.Journal of Fish Biology 52: 1259–1271.
Adams, C. E., C. Woltering & G. Alexander, 2003. Epigeneticregulation of trophic morphology through feedingbehaviour in Arctic charr, Salvelinus alpinus. Journal ofFish Biology 78: 43–49.
Adams, C. E., J. H. Hamilton, I. McCarthy, J. W. Alastair, A.Grant, G. Alexander, S. Waldron, S. S. Snorrason, M.Ferguson & S. Skulason, 2006. Does breeding site fidelitydrive phenotypic and genetic sub-structuring of a popu-lation of Arctic charr? Evolutionary Ecology 20: 11–26.
Alekseyev, S. S., V. P. Samusenok, A. N. Matveev & M. Y.Pichugin, 2002. Diversification, sympatric speciation, andtrophic polymorphism of Arctic charr, Salvelinus alpinuscomplex, in Transbaikalia. Journal of Fish Biology 64:97–114.
Alexander, G. D. & C. E. Adams, 2000. The phenotypicdiversity of Arctic charr, Salvelinus alpinus, (Salmonidae)in Scotland and Ireland. Aqua Journal of Ichthyology andAquatic Biology 4: 77–88.
Amundsen, P. A., R. Knudsen & A. Klemetsen, 2008. Seasonaland ontogenetic variations in resource use by two sym-patric Arctic charr morphs. Environmental Biology ofFishes 83: 45–55.
Baker, J. A., W. A. Cresko, S. A. Foster & D. C. Heins, 2005.Life-history differentiation of benthic and limnetic eco-types in a polytypic population of three-spine stickleback(Gasterosteus aculeatus). Evolutionary Ecology Research7: 121–131.
Baumgartner, J. V., 1992. Spatial variation of morphology in afreshwater population of the three-spine stickleback,Gasterosteus aculeatus. Canadian Journal of Zoology-Revue Canadienne de Zoologie 70: 1140–1148.
Bell, M. A. & S. A. Foster, 1994. The Evolutionary Biology ofthe Three-Spine Stickleback. Oxford University Press,New York.
Bolnick, D., R. Svanback, J. A. Fordyce, L. H. Yang, J. M.Davis, C. D. Husley & M. L. Forister, 2003. The Ecologyof Individuals: incidence and Implications of IndividualSpecialization. American Naturalist 161: 1–28.
Cresko, W. A. & J. A. Baker, 1996. Two morphotypes oflacustrine three-spine stickleback, Gasterosteus aculeatus,in Benka Lake, Alaska. Environmental Biology of Fishes45: 343–350.
Dieckmann, U. & M. Doebeli, 1999. On the origin of thespecies by sympatric speciation. Nature 400: 354–357.
Foster, S. A., J. A. Baker & M. A. Bell, 1992. Phenotypicintegration of life history and morphology: an examplefrom three-spined stickleback, Gasterosteus aculeatusL. Journal of Fish Biology 41: 21–35.
Fraser, D., C. E. Adams & F. A. Huntingford, 1998. Trophicpolymorphism among Arctic charr Salvelinus alpinus L.,from Loch Ericht, Scotland. Ecology of Freshwater Fish7: 184–191.
Fraser, D., F. A. Huntingford & C. E. Adams, 2008. Foragingspecialisms, prey size and life-history patterns: a test of
predictions using sympatric polymorphic Arctic charr(Salvelinus alpinus). Ecology of Freshwater Fish 17: 1–9.
Hegrenes, S., 2001. Diet-induced phenotypic plasticity offeeding morphology in the orangespotted sunfish, Lepomishumilis. Ecology of Freshwater Fish 10: 35–42.
Hjelm, J., R. Svanback, P. Bystrom, L. Persson & E.Wahlstrom,2001. Diet-dependent body morphology and onto-genetic reaction norms in Eurasian perch. Oikos 95:311–323.
Hjelm, J., G. H. van de Weerd & F. A. Sibbing, 2003. Func-tional link between foraging performance, functionalmorphology, and diet shift in roach (Rutilus rutilus).Canadian Journal of Fisheries and Aquatic Sciences 60:700–709.
Januszkiewicz, A. J. & B. W. Robinson, 2007. Divergentwalleye (Sander vitreus) mediated inducible defenses inthe centrarchid pumpkinseed sunfish (Lepomis gibbosus).Journal of Fish Biology 90: 25–36.
Kahilainen, K. & K. Ostbye, 2006. Morphological differenti-ation and resource polymorphism in three sympatricwhitefish Coregonus lavaretus (L.) forms in a subarcticlake. Journal of Fish Biology 68: 63–79.
Kahilainen, K. K., T. Malinen, A. Tuomaala, E. Alajaervi, A.Tolonen & H. Lehtonen, 2007. Empirical evaluation ofphenotype-environment correlation and trait utility withallopatric and sympatric whitefish. Coregonus lavaretus(L.), populations in subarctic lakes. Journal of Fish Biology92: 561–572.
Ke, Z. X., P. Xie & L. G. Guo, 2008. Phenotypic plasticity ingut length in the planktivorous filter-feeding silver carp(Hypophthalmichthys molitrix). Scientific World Journal8: 169–175.
Klemetsen, A., J. M. Elliott, R. Knudsen & P. Sorensen, 2002.Evidence for genetic differences in the offspring of twosympatric morphs of Arctic charr. Journal of Fish Biology60: 933–950.
Klemetsen, A., R. Knudsen, F. J. Staldvik & P. A. Amundsen,2003. Habitat, diet and food assimilation of Arctic charrunder the winter ice in two subarctic lakes. Journal of FishBiology 62: 1082–1098.
Knudsen, R., A. Klemetsen, P. A. Amundsen & B. Hermansen,2006. Incipient speciation through niche expansion: anexample from the Arctic charr in a subarctic lake. Pro-ceedings of the Royal Society B-Biological Sciences 273:2291–2298.
Kristjansson, B., S. Skulason & D. L. Noakes, 2002.Morphological segregation of Icelandic threespine stickle-back (Gasterosteus aculeatus L). Journal of Fish Biology76: 247–257.
Larson, G. L. & C. D. McIntire, 1993. Food-habits of differentphenotypes of threespine stickleback in Paxton Lake,British Columbia. Transactions of the American FisheriesSociety 122: 543–549.
Lu, G. Q. & L. Bernatchez, 1999. Correlated trophic special-ization and genetic divergence in sympatric lake whitefishecotypes (Coregonus clupeaformis): support for the eco-logical speciation hypothesis. Evolution 53: 1491–1505.
Maerz, J. C., E. M. Myers & D. C. Adams, 2006. Trophicpolymorphism in a terrestrial salamander. EvolutionaryEcology Research 8: 23–35.
24 Hydrobiologia (2010) 650:15–26
123

Maheswaran, G. & A. Rahmani, 2002. Foraging behaviour andfeeding success of the black-necked stork (Ep-hippiorhynchus asiaticus) in Dudwa National Park, UttarPradesh, India. Journal of Zoology 258: 189–195.
Malaquias, M. A. E., E. Berecibar & D. G. Reid, 2009.Reassessment of the trophic position of Bullidae(Gastropoda: Cephalaspidea) and the importance of diet inthe evolution of cephalaspidean gastropods. Journal ofZoology 277: 88–97.
Malmquist, H. J., S. S. Snorrason, S. Skulason, B. Jonsson,O. T. Sandlund & P. M. Jonasson, 1992. Diet differenti-ation in polymorphic Arctic charr in Thingvallavatn,Iceland. Journal of Animal Ecology 61: 21–35.
McPhail, J., 1992. Ecology and evolution of sympatric stick-lebacks (Gasterosteus): evidence for a species-pair inPaxton Lake, Texada Island, British Columbia. CanadianJournal of Zoology-Revue Canadienne de Zoologie 70:361–369.
Michaud, W. K., M. Power & M. T. Kinnison, 2008. Trophi-cally mediated divergence of Arctic charr (Salvelinusalpinus L.) populations in contemporary time. Evolu-tionary Ecology Research 10: 1051–1066.
Mittelbach, G. C., C. W. Osenberg & P. C. Wainwright, 1999.Variation in feeding morphology between pumpkinseedpopulations: phenotypic plasticity or evolution? Evolu-tionary Ecology Research 1: 111–128.
Olsson, J., M. Quevedo, C. Colson & R. Svanback, 2007. Gutlength plasticity in perch: into the bowels of resourcepolymorphisms. Journal of Fish Biology 90: 517–523.
Ostbye, K., T. F. Naesje, L. Bernatchez, O. T. Sandlund &K. Hindar, 2005. Morphological divergence and origin ofsympatric populations of European whitefish (Coregonuslavaretus L.) in Lake Femund, Norway. Journal ofEvolutionary Biology 18: 683–702.
Queral-Regil, A. & R. B. King, 1998. Evidence for phenotypicplasticity in snake body size and relative head dimensions inresponse to amount and size of prey.Copeia 1998: 423–429.
Reilly, S. M., G. V. Lauder & J. P. Collins, 1992. Performanceconsequences of a trophic polymorphism: feedingbehavior in typical and cannibal phenotypes of Ambys-toma tigrinum. Copeia 1992: 672–679.
Robinson, B. W. & K. J. Parsons, 2002. Changing times,spaces, and faces: tests and implications of adaptivemorphological plasticity in the fishes of northern post-glacial lakes. Canadian Journal of Fisheries and AquaticSciences 59: 1819–1833.
Robinson, B. W. & D. S. Wilson, 1994. Character release anddisplacement in fishes: a neglected literature. AmericanNaturalist 144: 596–627.
Rohlf, F. J., 2006. TpsDig, version 2.10. Department of Ecol-ogy and Evolution, State University of New York at StonyBrook. Available at http://life.bio.sunysb.edu/morph.
Rohlf, F. J., 2007, tpsRelw, version 1.45. Department of Ecologyand Evolution, State University of New York at StonyBrook. Available at http://life.bio.sunysb.edu/morph.
Rohlf, F. J. & D. E. Slice, 1990. Extensions of the Procrustesmethod for the optimal superimposition of landmarks.Systematic Zoology 39: 40–59.
Ruehl, C. B. & T. J. Dewitt, 2007. Trophic plasticity andforaging performance in red drum, Sciaenops ocellatus
(Linnaeus). Journal of Experimental Marine Biology andEcology 349: 284–294.
Schluter, D., 1996. Ecological speciation in postglacial fishes.Philosophical Transactions of the Royal Society of Lon-don, Series B 351: 807–814.
Schluter, D., 2001. Ecology and the origin of species. Trends inEcology & Evolution 16: 372–380.
Schmidt, B. R., N. Ramer & J. Van Buskirk, 2006. A trophicpolymorphism induced by both predators and prey. Evo-lutionary Ecology Research 8: 1301–1309.
Skulason, S. & T. B. Smith, 1995. Resource polymorphisms invertebrates. Trends in Ecology & Evolution 10: 366–370.
Skulason, S., D. L. G. Noakes & S. S. Snorrason, 1989.Ontogeny of trophic morphology in 4 sympatric morphsof Arctic charr Salvelinus alpinus in Thingvallavatn,Iceland. Journal of Fish Biology 38: 281–301.
Skulason, S., S. S. Snorrason & B. Jonsson, 1999. Sympatricsmorphs, populations and speciation in freshwater fish withemphasis on Arctic charr. In Magurran, A. E. & R. May(eds), Evolution of Biological Diversity. Oxford Univer-sity Press, Oxford, UK: 70–92.
Smith, T. B. & S. Skulason, 1996. Evolutionary significance ofresource polymorphism in fishes, amphibians and birds.Annual Review of Ecology and Systematics 27: 111–133.
Smits, J. D., F. Witte & F. G. VanVeen, 1996. Functionalchanges in the anatomy of the pharyngeal jaw apparatusof Astatoreochromis alluaudi (Pisces, Cichlidae), andtheir effects on adjacent structures. Journal of Fish Biol-ogy 59: 389–409.
Snorrason, S. S., S. Skulason, B. Jonsson, H. J. Malmquist, P.M. Jonasson, J. T. Sandlund & T. Lindem, 1994. Trophicspecialization in Arctic charr Salvelinus alpinus (Pisces;Salmonidae): morphological divergence and ontogeneticniche shifts. Journal of Fish Biology 52: 1–18.
Starck, J. M., 1999. Structural flexibility of the gastro-intestinaltract of vertebrates – implications for evolutionary mor-phology. Zoologischer Anzeiger 238: 87–101.
Stuart, B., A. P. Richard, M. Rona, C. Yves, A. D. Deborah &P. C. John, 2006. Stable isotopes indicate sex-specific andlong-term individual foraging specialisation in divingseabirds. Marine Ecology Progress Series 311: 157–164.
Swanson, B. O., A. C. Gibb, J. C. Marks & D. A. Hendrickson,2003. Trophic polymorphism and behavioral differencesdecrease intraspecific competition in a cichlid, Herichthysminckleyi. Ecology 84: 1441–1446.
Swanson, B., A. Gibb, J. Marks & D. Hendrickson, 2008.Variation in foraging behavior facilitates resource parti-tioning in a polymorphic cichlid, Herichthys minckleyi.Environmental Biology of Fishes 83: 147–154.
Uchii, K., N. Okuda, R. Yonekura, Z. Karube, K. Matsui &Z. Kawabata, 2007. Trophic polymorphism in bluegillsunfish (Lepomis macrochirus) introduced into LakeBiwa: evidence from stable isotope analysis. Limnology8: 59–63.
Wainwright, P. C., C. W. Osenberg & G. G. Mittelbach, 1991.Trophic polymorphism in the pumpkinseed sunfish (Lep-omis gibbosus Linnaeus): effects of environment onontogeny. Functional Ecology 5: 40–55.
Warburton, K. & C. Thomson, 2006. Costs of learning: thedynamics of mixed-prey exploitation by silver perch,
Hydrobiologia (2010) 650:15–26 25
123

Bidyanus bidyanus (Mitchell, 1838). Animal Behaviour71: 361–370.
Werner, E. E. & D. J. Hall, 1974. Optimal foraging and the sizeselection of prey by the bluegill sunfish (Lepomis mac-rochirus). Ecology 55: 1042–1052.
West-Eberhard, M. J., 1989. Phenotypic plasticity and theorigins of diversity. Annual Review of Ecology andSystematics 20: 249–278.
West-Eberhard, M. J., 2005. Developmental plasticity and theorigin of species differences. Proceedings of the NationalAcademy of Sciences 102: 6543–6549.
Wimberger, P. H., 1994. Trophic polymorphisms, plasticityand speciation in vertebrates. In Stouder, D. J., K. L. Fresh& R. J. Feller (eds), Theory and Application of FishFeeding Ecology. Bellebaruch Library in Marine ScienceNo. 18, Columbia S.C. University of South CarolinaPress, Colombia, USA: 19–43.
Wintzer, A. P. & P. J. Motta, 2005. Diet-induced phenotypicplasticity in the skull morphology of hatchery-reared
Florida largemouth bass, Micropterus salmoides florid-anus. Ecology of Freshwater Fish 14: 311–318.
Woo, K. J., K. H. Elliott, M. Davidson, A. J. Gaston & G. K.Davoren, 2008. Individual specialization in diet by a gen-eralist marine predator reflects specialization in foragingbehaviour. Journal of Animal Ecology 77: 1082–1091.
Wood, C. & C. J. Foote, 1996. Evidence for sympatric geneticdivergence of anadromous and nonanadromous morphs ofsockeye salmon (Oncorhynchus nerka). Evolution 50:1265–1279.
Yonekura, R., K. Nakai & M. Yuma, 2002. Trophic poly-morphism in introduced bluegill in Japan. EcologicalResearch 17: 49–57.
Zelditch, M. L., D. L. Swiderski, H. D. Sheets & W. L. Fink,2004. Geometric Morphometrics for Biologists: A Primer.Elsevier, New York.
26 Hydrobiologia (2010) 650:15–26
123