novel roles of plant retinoblastoma-related (rbr) protein in cell proliferation and asymmetric cell...
TRANSCRIPT

Journal of Experimental Botanydoi:10.1093/jxb/ert411
Review papeR
Novel roles of plant RETINOBLASTOMA-RELATED (RBR) protein in cell proliferation and asymmetric cell division
Bénédicte Desvoyes1,*, Alex de Mendoza2,3, Iñaki Ruiz-Trillo2,3 and Crisanto Gutierrez1,*1 Centro de Biologia Molecular Severo Ochoa, CSIC-UAM, Cantoblanco, Nicolas Cabrera 1, 28049 Madrid, Spain2 Institut de Biologia Evolutiva (CSIC-Universitat Pompeu Fabra), Passeig Marítim de la Barceloneta 37–49, 08003 Barcelona, Spain3 Institució Catalana de Recerca i Estudis Avançats (ICREA), Passeig Lluís Companys 23, 08010 Barcelona, Catalonia, Spain
* To whom correspondence should be addressed. E-mail: [email protected] and [email protected]
Received 27 September 2013; Revised 5 November 2013; Accepted 7 November 2013
Abstract
The retinoblastoma (Rb) protein was identified as a human tumour suppressor protein that controls various stages of cell proliferation through the interaction with members of the E2F family of transcription factors. It was originally thought to be specific to animals but plants contain homologues of Rb, called RETINOBLASTOMA-RELATED (RBR). In fact, the Rb–E2F module seems to be a very early acquisition of eukaryotes. The activity of RBR depends on phospho-rylation of certain amino acid residues, which in most cases are well conserved between plant and animal proteins. In addition to its role in cell-cycle progression, RBR has been shown to participate in various cellular processes such as endoreplication, transcriptional regulation, chromatin remodelling, cell growth, stem cell biology, and differentiation. Here, we discuss the most recent advances to define the role of RBR in cell proliferation and asymmetric cell divi-sion. These and other reports clearly support the idea that RBR is used as a landing platform of a plethora of cellular proteins and complexes to control various aspects of cell physiology and plant development.
Key words: Arabidopsis, asymmetric cell division, cell cycle, gene expression, phosphorylation, plant, Rb plylogenetic tree, retinoblastoma-related (RBR).
Historical and evolutionary overview
Cell-cycle research witnessed a series of key discoveries about 20–25 years ago. Studies in yeast and later in mammalian cells indicated that the G1/S and the G2/M transitions were the major points controlling cell-cycle progression. However, except for the involvement of the Cdc2 kinase required for both the G1/S and the G2/M transitions in fission yeast (Nurse and Bissett, 1981), the molecular framework for these transitions was virtually unknown, in particular in higher eukaryotes. The finding that, in the vast majority of human tumours, the prod-uct of the retinoblastoma (Rb) gene was inactivated (Knudsen and Knudsen, 2006) and the identification and cloning of the human Rb gene constituted major breakthrough discoveries in mammalian cell-cycle research (Friend et al., 1986; Fung et al., 1987; Lee et al., 1987). Soon afterwards, the pivotal roles of Rb as a tumour suppressor, as a negative regulator of the
G1/S transition and as a modulator of the switch from quies-cence to proliferation, were definitely established (Weinberg, 1995; Manning and Dyson, 2012). All these functions rely largely on the interaction of Rb protein with transcription fac-tors (e.g. members of the E2F family, and with CDK/cyclin complexes, in particular CycD-containing complexes). These proteins bind to Rb through the LxCxE amino acid motif pre-sent in CycD. This leads to the phosphorylation and inactiva-tion of Rb, which no longer binds to E2F, thus allowing the expression of E2F targets required for G1/S progression. This normal regulatory circuit can be bypassed by DNA tumour viruses that encode proteins, such as SV40 T-ag or HPV E7, that bind Rb through their LxCxE motifs, mimicking binding of CycD to Rb, inactivating it, and stimulating cell-cycle pro-gression (DeCaprio, 2009).
© The Author 2013. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: [email protected]
Journal of Experimental Botany Advance Access published December 9, 2013 at C
SIC on D
ecember 17, 2013
http://jxb.oxfordjournals.org/D
ownloaded from

Page 2 of 10 | Desvoyes et al.
Nowadays, it is clear that the general principles governing cell-cycle progression are largely conserved in all eukaryotes, including plants. However, back in the early 1990s and based on (i) the absence of Rb in yeast, (ii) the similarity of the first CDK and cyclins identified in plants to yeast proteins, and (iii) the rather naïve assumption that plant cells ‘must’ have a simple and primitive cell-cycle regulatory circuitry, it was widely accepted that the Rb–E2F pathway was an animal invention.
Two major lines of research provided new insights that changed the view on plant cell-cycle regulation. One was the finding that plants contain homologues of the D-type cyc-lins (Dahl et al., 1995; Soni et al., 1995). Although D-type cyclin phylogenetic relationships are not well resolved (Ma et al., 2013) and the degree of similarity of plant CYCD with human proteins is very limited, they share structural and func-tional similarities, such as their expression patterns and, most remarkably, a LxCxE amino acid motif (Soni et al., 1995; Huntley et al., 1998; Murray et al., 1998; Riou-Khamlichi et al., 1999). Another strongly provocative finding came from the field of geminiviruses, a rather unique family of plant DNA viruses. Replication of the virus genome occurs within the nucleus of the infected cell through double-stranded DNA intermediates that are the transcriptionally active tem-plates (reviewed in Gutierrez, 2000; Gutierrez et al., 2002; Jeske, 2009). Interestingly, geminiviruses also trigger nuclear DNA synthesis (Nagar et al., 1995; Hanley-Bowdoin et al., 2000). Remarkably, the early RepA protein of wheat dwarf virus (WDV) contains a LxCxE amino acid motif that is functionally active since it mediates heterologous interaction of RepA with human Rb protein and when mutated reduces significantly virus DNA accumulation in cultured plant cells (Xie et al., 1995; Gutierrez, 2000).
Together, this circumstantial evidence strongly suggested that the Rb pathway might not be a novel and unique acqui-sition of animals but instead an ancient invention shared with plants. Indeed, early studies based on the identification of expressed sequence tag sequences of maize that showed a significant homology with human Rb, in particular in the B domain of its A/B pocket domain, allowed the isolation of a maize cDNA clone encoding a plant homologue of human Rb (Grafi et al., 1996; Xie et al., 1996). Due to the similar-ity between these proteins, the name RETINOBLASTOMA-RELATED (RBR) was adopted for the plant homologues of animal Rb proteins.
Geminiviruses other than WDV and belonging to other genera were found to also encode the early Rep proteins that interact with plant RBR proteins (e.g. Arabidopsis RBR; Kong et al., 2000). Interestingly, in these cases binding occurs through a non-LxCxE amino acid motif, revealing a clear case of convergent evolution to subvert the cell-cycle regulatory machinery of the host (Gutierrez, 2000; Hanley-Bowdoin et al., 2000). Monocotyledonous plants were found to contain various RBR proteins, whereas in dicots RBR appears to be a single gene (Ach et al., 1997; Fountain et al., 1999; Sabelli and Larkins, 2006; Lendvai et al., 2007; Gutzat et al., 2012). After a plant RBR gene was described, E2F transcription factors as well as their dimerization partner
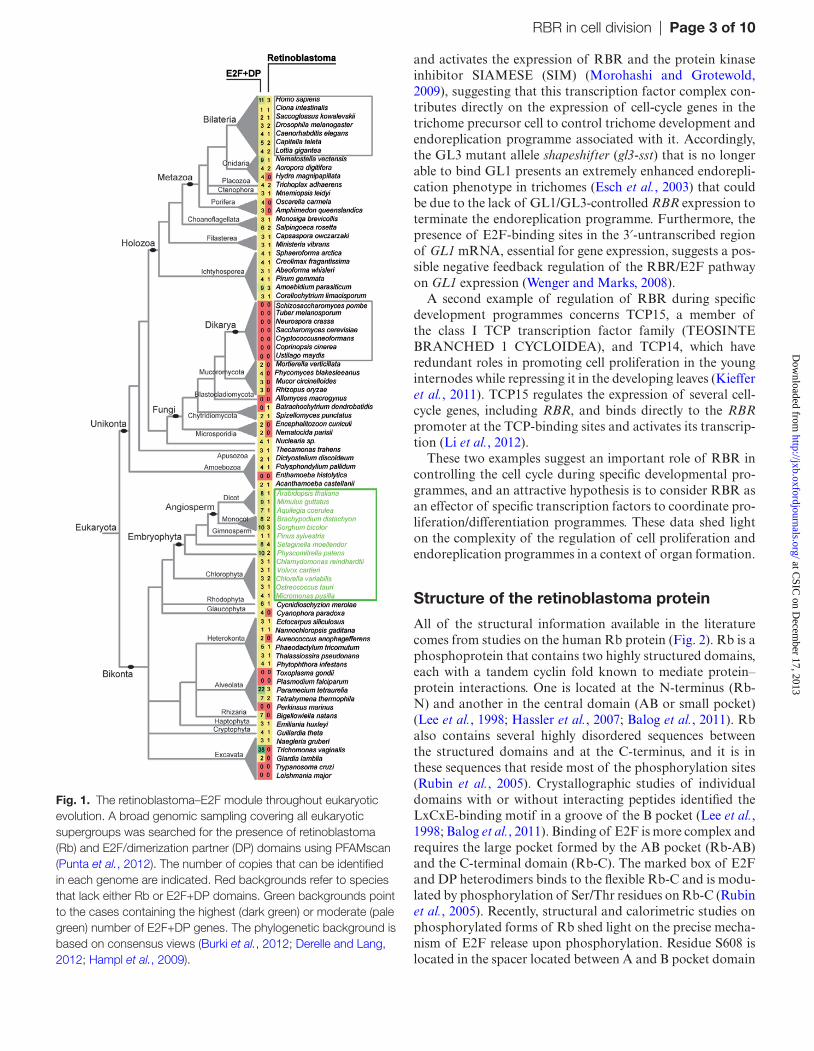
(DP) proteins were identified (Ramirez-Parra et al., 1999; Magyar et al., 2000; Ramirez-Parra and Gutierrez, 2000; Kosugi and Ohashi, 2002; Mariconti et al., 2002; Vlieghe et al., 2005). All these data obtained from both animal and plant cells supported the idea that the presence of the RBR–E2F pathway might be linked to the explosion of multicellular forms of life about 800–1000 Mya. However, RBR was identified as well in some unicellular algae, such as Chlamydomonas reinhardtii and Ostreococcus tauri (Umen and Goodenough, 2001; Moulager et al., 2010). It was also identified in colonial algae, such as Volvox (Kianianmomeni et al., 2008), and in many other eukaryotes, underscoring the ancient origin of the RBR gene. In fact, at least one spe-cies of each of the eukaryotic supergroups has a Rb gene. Interestingly, E2F genes are also present (Fig. 1) in taxa with a Rb gene, except for the cythrid fungi Batrachochytrium dendrobatidis. It is worth noting that some species have sec-ondarily lost both RBR and E2F genes, e.g. yeast, while others have lost just the Rb gene. In the particular case of plants and green algae (Viridiplantae), in agreement with other studies (Gutzat et al., 2012), all the taxa analysed have at least one RBR gene (except the Glaucophyta) and several E2F genes (Fig. 1). Thus, it is clear that the plant ancestor already had the RBR–E2F module. Indeed, the presence of both RBR and E2F genes in the genomes of animals and other eukaryotic lineages, including excavates, ameobozo-ans, and apusozoans indicates that the RBR–E2F pathway appeared before the divergence of the plant and animal line-ages, most likely in the Last Eukaryotic Common Ancestor (Roger and Simpson, 2009; Cao et al., 2010; Cross et al., 2011; Derelle and Lang, 2012).
The decade after the discovery of RBR in plants served to demonstrate unequivocally that RBR is more than just part of the cell-cycle machinery (Gutierrez, 2005). Thus, the RBR–E2F pathway has been shown to have roles in cell proliferation, endoreplication, differentiation, and stem cell biology among other aspects of plant physiology. These have been discussed in other recent comprehensive reviews on the role of RBR during plant development (Dudits et al., 2011; Gutzat et al., 2012) and on the regulatory function of mam-malian Rb (Chinnam and Goodrich, 2011; Henley and Dick, 2012; see also Kuwabara and Gruissem, in press, this issue). Here, we will focus on recent findings not reviewed so far in the context of plant cell cycle and development.
Regulation of RBR expression
The RBR promoter contains E2F-binding sites. Consistent with this, its expression is cell-cycle regulated and peaks at G1 in synchronized cell cultures (Menges et al., 2005). Nevertheless, the regulation of RBR expression by other transcription factors involved in different programmes of differentiation has been also reported. Trichome initiation requires the R2-R3 MYB GLABRA1 (GL1) and the bHLH GLABRA3 (GL3) or ENHANCER OF GLABRA 3 (EGL3) transcription factors to control the expression of specific epi-dermal cell fate genes (Schellmann and Hulskamp, 2005). Moreover, this GL1/GL3 complex binds to the promoter
at CSIC
on Decem
ber 17, 2013http://jxb.oxfordjournals.org/
Dow
nloaded from
RBR in cell division | Page 3 of 10
and activates the expression of RBR and the protein kinase inhibitor SIAMESE (SIM) (Morohashi and Grotewold, 2009), suggesting that this transcription factor complex con-tributes directly on the expression of cell-cycle genes in the trichome precursor cell to control trichome development and endoreplication programme associated with it. Accordingly, the GL3 mutant allele shapeshifter (gl3-sst) that is no longer able to bind GL1 presents an extremely enhanced endorepli-cation phenotype in trichomes (Esch et al., 2003) that could be due to the lack of GL1/GL3-controlled RBR expression to terminate the endoreplication programme. Furthermore, the presence of E2F-binding sites in the 3′-untranscribed region of GL1 mRNA, essential for gene expression, suggests a pos-sible negative feedback regulation of the RBR/E2F pathway on GL1 expression (Wenger and Marks, 2008).
A second example of regulation of RBR during specific development programmes concerns TCP15, a member of the class I TCP transcription factor family (TEOSINTE BRANCHED 1 CYCLOIDEA), and TCP14, which have redundant roles in promoting cell proliferation in the young internodes while repressing it in the developing leaves (Kieffer et al., 2011). TCP15 regulates the expression of several cell-cycle genes, including RBR, and binds directly to the RBR promoter at the TCP-binding sites and activates its transcrip-tion (Li et al., 2012).
These two examples suggest an important role of RBR in controlling the cell cycle during specific developmental pro-grammes, and an attractive hypothesis is to consider RBR as an effector of specific transcription factors to coordinate pro-liferation/differentiation programmes. These data shed light on the complexity of the regulation of cell proliferation and endoreplication programmes in a context of organ formation.
Structure of the retinoblastoma protein
All of the structural information available in the literature comes from studies on the human Rb protein (Fig. 2). Rb is a phosphoprotein that contains two highly structured domains, each with a tandem cyclin fold known to mediate protein–protein interactions. One is located at the N-terminus (Rb-N) and another in the central domain (AB or small pocket) (Lee et al., 1998; Hassler et al., 2007; Balog et al., 2011). Rb also contains several highly disordered sequences between the structured domains and at the C-terminus, and it is in these sequences that reside most of the phosphorylation sites (Rubin et al., 2005). Crystallographic studies of individual domains with or without interacting peptides identified the LxCxE-binding motif in a groove of the B pocket (Lee et al., 1998; Balog et al., 2011). Binding of E2F is more complex and requires the large pocket formed by the AB pocket (Rb-AB) and the C-terminal domain (Rb-C). The marked box of E2F and DP heterodimers binds to the flexible Rb-C and is modu-lated by phosphorylation of Ser/Thr residues on Rb-C (Rubin et al., 2005). Recently, structural and calorimetric studies on phosphorylated forms of Rb shed light on the precise mecha-nism of E2F release upon phosphorylation. Residue S608 is located in the spacer located between A and B pocket domain
Fig. 1. The retinoblastoma–E2F module throughout eukaryotic evolution. A broad genomic sampling covering all eukaryotic supergroups was searched for the presence of retinoblastoma (Rb) and E2F/dimerization partner (DP) domains using PFAMscan (Punta et al., 2012). The number of copies that can be identified in each genome are indicated. Red backgrounds refer to species that lack either Rb or E2F+DP domains. Green backgrounds point to the cases containing the highest (dark green) or moderate (pale green) number of E2F+DP genes. The phylogenetic background is based on consensus views (Burki et al., 2012; Derelle and Lang, 2012; Hampl et al., 2009).
at CSIC
on Decem
ber 17, 2013http://jxb.oxfordjournals.org/
Dow
nloaded from

Page 4 of 10 | Desvoyes et al.
where the E2F transactivation domain (E2FTD) binds. When this residue is phosphorylated, the unstructured sequence binds to the pocket interface and the interaction with E2FTD is competitively inhibited. A second phosphorylation event at T373 located in the unstructured region between Rb-N and Rb-AB provokes a major conformational change docking Rb-N and Rb-AB. This converts the elongated architecture of the protein into a globular compact conformation, thus releasing E2F and LxCxE-containing proteins (Burke et al., 2010; Burke et al., 2012; Lamber et al., 2013). Since plant RBRs present a significant sequence similarity and share the same domain organization with their mammalian counter-parts (Fig. 2) it is conceivable that many structural features are also shared between animal and plant Rb proteins.
Multiple interactors of Rb have been identified (Morris and Dyson, 2001; Goodrich, 2006) Thus, an emerging idea is that Rb acts like a platform to bring different cellular proteins and enzymic activities to a specific chromatin location (Chinnam and Goodrich, 2011; Munro et al., 2012). Sequential phos-phorylation by cyclin/CDK complexes, according to a phos-phorylation code that still needs to be uncovered, modulates the binding to different partners (Rubin, 2013). All this structural knowledge gathered for Rb constitutes valuable
information to characterize the function of plant RBR part-ners, although the current list of known RBR partners is not as wide as that for human Rb.
Cell-cycle and endocycle regulation
One of the key points of cell-cycle regulation is the G1/S transition, where cells take the decision to initiate a new round of DNA replication. In animal cells, the role of Rb as a negative regulator of the E2F transcription factor family and recruiting co-repressors proteins is now well established (Henley and Dick, 2012). Upon external stimuli, Rb is inac-tivated by Cyc/CDK-mediated phosphorylation, released from E2F/DP complexes, and transcription of E2F targets is activated to initiate S phase (Chinnam and Goodrich, 2011). Plant cells frequently exit the cell cycle and switch to the endoreplication cycle whereby cells undergo successive full genome replication rounds in the absence of mitosis. The RBR/E2F pathway is conserved in plants and partici-pate both in cell-cycle regulation and endoreplication onset (Fig. 3)(Gutierrez, 2009). Since rbr loss-of-function mutants are gametophytic lethal (Ebel et al., 2004), several strategies,
Fig. 2. Comparative domain organization of human and Arabidopsis thaliana RBR proteins. The major domains (defined by the amino acid positions) are conserved in both proteins. The putative CDK phosphorylation sites in Arabidopsis RBR are also indicated.
Fig. 3. The gene regulatory network involving RBR in the control of endocreplication onset. Abrogation of mitotic CDK/cyclin activity is a requirement to initiate the endoreplication cycle. This is achieved through various pathways where RBR plays a key role as a regulator of E2F target genes. In addition, direct regulation of the anaphase-promoting complex CCS52A by DEL1/E2Fe and degradation of CDK inhibitors are of primary relevance for the endoreplication onset.
at CSIC
on Decem
ber 17, 2013http://jxb.oxfordjournals.org/
Dow
nloaded from

RBR in cell division | Page 5 of 10
such as viral-induced gene silencing, inactivation of RBR by viral proteins, or inducible RNAi, have been designed to circumvent this experimental drawback. Conditional inac-tivation of RBR leads to an activation of both mitotic and endoreplication cycles that depend on the stage of plant development at the time of inactivation of RBR (Park et al., 2005; Desvoyes et al., 2006; Jordan et al., 2007), a pheno-type reminiscent to that of plants overexpressing E2Fa/DPa (De Veylder et al., 2002). Downregulation of RBR within a window when proliferation has not yet ceased leads to localized hyperproliferation. This suggests that only cells competent for cell division respond in that way to RBR inactivation (Park et al., 2005; Desvoyes et al., 2006; Borghi et al., 2010). In leaves, responsive cells express TMM, a meristemoid marker, indicating that the cells that prolifer-ate are of the stomatal lineage (Borghi et al., 2010). These are indeed known to be the last to exit the cell cycle during leaf development (Gonzalez et al., 2012). Although RBR inactivation in cells that already exit mitotic cycle will trig-ger endoreplication (Park et al., 2005; Desvoyes et al., 2006; Jordan et al., 2007), RBR overexpression in its own domain of expression had the opposite effect and represses both cell proliferation and endoreplication (Magyar et al., 2012). The maize genome possesses four different RBR genes, which can be separated in two different types: RBR1 and RBR2, and RBR3 and RBR4 (Sabelli and Larkins, 2006). RBR1 is the orthologue of Rb, whereas RBR3 shows more similar-ity to the mammalian p107 (Sabelli et al., 2005). RBR3 is a target of the RBR1/E2F pathway and positively regulates the expression of replication genes (Sabelli et al., 2009). Recently, it has been shown that RBR1 regulates cell prolif-eration, endoreplication, and cell death during endosperm development (Sabelli et al., 2013). These phenotypes are reminiscent of the effect of E2Fa-DPa overexpression that also has this dual effect on the mitotic and endocycle (De Veylder et al., 2002; Kosugi and Ohashi, 2003; Magyar et al., 2012). The well-established model of RBR/E2F path-way G1/S regulation was recently questioned by a surpris-ing report regarding this dual role of RBR/E2Fa (Magyar et al., 2012). Quantitative immunoprecipitation experi-ments showed that, in proliferative cells, RBR–E2Fa, but not RBR–E2Fb, forms a stable repressive complex that is resistant to phosphorylation in conditions where CYCD3;1 activity is high. Interestingly, a stable RBR–E2F complex has been also identified in Chlamydomonas (Olson et al., 2010). RBR–E2Fa complexes, together with the atypical E2F DEL1/E2Fe, repress the APC activator and endocycle-promoting genes CCS52A, thus, maintaining cell prolifer-ation status (Lammens et al., 2008; Magyar et al., 2012). The authors suggest that the classic G1/S transition medi-ated by phosphorylation of RBR during mitotic cycle tar-gets RBR–E2Fb complexes. In differentiating cells, E2Fa is released from RBR, by an unknown mechanism triggering the expression of genes, such as CCS52, which will activate endoreplication. This pathway may need the coordinated phosphorylation at other sites on RBR by CDK associated with cyclins other than CYCD3;1. This mechanism can be similar to that found in mammalian cells where Rb/E2F1
complexes, resistant to CDK phosphorylation and present during S phase, have been also reported (Henley and Dick, 2012).
Since RBR function is strictly dependent on its phospho-rylation level by CDK, the status of CDK inhibitors emerges as an important component. In fact, a new layer of complex-ity in the regulation of the G1/S transition has been recently reported and involves the key role of proteasome degrada-tion of members of the KIP-related CDK inhibitors family (KRP). The F-Box protein (FBL17) forms cullin 1-containing E3 ubiquitine ligase complex (SCFFBL17) that targets KRP6 and KRP7 for degradation, an essential step for the prolifera-tion of male germ cells (Kim et al., 2008; Gusti et al., 2009) but also required for the female gamete development (Zhao et al., 2012). The fbl17 mutant resembles cdka loss-of-func-tion alleles in that pollen fails to progress through the mitotic division (Kim et al., 2008; Gusti et al., 2009). Interestingly, FBL17 was identified as an E2F target gene (Vandepoele et al., 2005). It is upregulated in plants overexpressing E2Fa/DPa (Gusti et al., 2009) and regulated directly by E2Fa and RBR (Zhao et al., 2012). The regulation cascade involves four steps of negative regulation and forms a loop where FBL17 inhibits CDKA;1 that in turn inhibits RBR by phosphoryla-tion that inhibits E2Fa that, finally, controls FBL17 and other replication genes’ expression. The negative feedback wiring favours the hysteresis finely controlling the switch to S phase (Zhao et al., 2012). FBL17 is widely expressed in different plant organs (Kim et al., 2008) and mediates the degradation of different members of the KRP family, suggesting a more general role in regulating the entry into S phase not only lim-ited to gametes. (Zhao et al., 2012). Further experimentation will be needed to confirm this hypothesis.
RBR controls asymmetric cell division
Due to the presence of the cell wall, plant cells are not mobile and the plane of cell division that occurred dur-ing cytokinesis in the proliferative area will determine the mature tissue organization and the whole plant architecture. Early events in the formation of a new cell lineage impli-cate the occurrence of an asymmetric cell division (ACD), also known as formative division, of a stem cell initial or of a cell with stem cell potential. This is, for example, the case of the establishment of the stomatal lineage, the initia-tion of lateral roots, or the formation of the ground tissue in the root. Several genes implicated in the specification of asymmetric division during embryogenesis (Ueda and Laux, 2012) or in the formation of different organs or tissues have been identified (De Smet and Beeckman, 2011). However, only recently a direct implication of the cell-cycle machinery has been established for the formative division at the root ground tissue (Sozzani et al., 2010; Cruz-Ramirez et al., 2012; Weimer et al., 2012).
The cortex and endodermis lineages are generated after two successive asymmetric divisions: the cortex/endodermis stem cell (CEI) divides anticlinally to self-renew and form a cortex/endodermis daughter cell (CEID) that, in turn, divides periclinally to initiate the cortex and endodermis layers. The
at CSIC
on Decem
ber 17, 2013http://jxb.oxfordjournals.org/
Dow
nloaded from

Page 6 of 10 | Desvoyes et al.
GRAS family of transcription factors SHORTROOT (SHR) and SCARECROW (SCR) are key players for the specifica-tion of these tissues and for the maintenance of the stem cell niche in the root meristem (Bennett and Scheres, 2010). It was reported that RBR interacts genetically with SCR to maintain the stem cell pool in the root meristem (Wildwater et al., 2005). It has been also shown that SCR also physically interacts with RBR through a canonical LxCxE motif and a combination of experimental and modelling studies demon-strated that the ternary complex formed by SCR–SHR–RBR modulates the expression of SHR–SCR targets and inhibits ACD in the endodermis outside CEID (Cruz-Ramirez et al., 2012). Several regulatory loops give robustness to the sys-tem and assure the spatiotemporal specificity of the ACD (Fig. 4). The radial movement of SHR from the stele to the endodermis where it is sequestrated by SCR defines the layer competent for ACD (Cui et al., 2007). CYCLIN D6;1 (CYC D6;1) is a direct target of SCR/SHR, and its expression restricted to the CEID (Sozzani et al., 2010) depends on the interaction of RBR with SCR and also on an apical–basal auxin gradient along the endodermis layer (Cruz-Ramirez et al., 2012). Furthermore, phosphorylation of RBR by CDK/CYCD6;1 in the CEID releases RBR from the SCR–SHR complex triggering ACD. Even though CBKB1 binds and phosphorylates RBR in complex with CYCD6;1 in vitro, it seems that also CDKA;1 could supply this function. Thus, a null mutant of CKDA;1 is severely compromised for cell-cycle progression and is not viable, but reduction of CDKA;1 activity in a series of cdka;1 hypomorphic mutants displayed a delay in the ACD of CEID, indicating that a high CDKA;1 activity is required for ACD (Weimer et al., 2012). Additionally, CDKB;1/CYCD6;1 complexes are able to phosphorylate in vitro the RBR pocket domain (Cruz-Ramirez et al., 2012) but the CEI/CEID phenotype of cdkb1;1 cdkb1;2 double mutant is much milder than that of the cdka;1 mutant, strongly suggesting a predominant role of CDKA;1 in ACD. Furthermore, the delay in ACD shown in the cdka;1 hypomorphic mutant can be partially suppressed when introgressed in a rbr mutant background,
demonstrating that RBR is effectively the target of CDKA;1 (Weimer et al., 2012). However, expression of CDKB1;1 in a cdka;1 null mutant can partially rescue the CEID division, indicating that it must somehow participate in the pathway (Weimer et al., 2012). One possible explanation to the par-ticipation of both CDKA and CDKB1 is that they phospho-rylate RBR sequentially, similarly to mammalian cells where different cyclin/CDK complexes participate in Rb inactiva-tion (Munro et al., 2012). Finally, the last level of regula-tion of ACD is post translational. SHR, SCR, CYCD6;1, and RBR are degraded during or just before mitosis (SCR and RBR by the 26S-proteasome), resetting the system in the daughter cells and confining the ACD at the position of the CEID (Cruz-Ramirez et al., 2012). The interplay of CDKA;1 and RBR is also needed for the ACD that will initiate the sto-matal lineage, targeting the stoma-specific transcription fac-tor SPEECHLESS (Weimer et al., 2012). The role of RBR in these two cell-lineage formative divisions suggests that RBR could be a general regulator of formative divisions and lin-eage commitment along with specific transcription factors. Further research will be needed to challenge this hypothesis but it will be in line with recent findings in mammalian cells where RB together with differentiation-specific transcription factors was shown to influence cell fate choice in vivo (Calo et al., 2010).
Perspectives
The continuous organogenesis programme of plant devel-opment relies in a strict coordination between proliferative potential, cell-cycle exit to differentiation, and endoreplica-tion. A complex gene network is being deciphered that regu-lates these pathways and among them RBR is positioned at a crossroad. In addition to its best-characterized function as a regulator of the G1-S transition and the endoreplica-tion onset, RBR is emerging as a master regulator of many cellular processes, as it is discussed here and in the accom-panying review article in this issue. Part of this multifunc-tional behaviour is mediated by the large amount of RBR
Fig. 4. RBR in asymmetric cell division (ACD), showing the role of the RBR–SCR–SHR complex in regulating ACD for the formation of the root meristem ground tissue and the importance of RBR phosphorylation in disrupting the ternary complex. CEID, cortex/endodermis daughter cell.
at CSIC
on Decem
ber 17, 2013http://jxb.oxfordjournals.org/
Dow
nloaded from

RBR in cell division | Page 7 of 10
interactors. It seems that RBR acts as a platform to recruit and/or deliver cellular factors to certain genome locations, including transcription factors, signalling components, and chromatin remodelling enzymes, among others. These inter-actions depend primarily on two RBR features: one is the modular organization of the protein and another the con-formational changes that take place as a consequence of RBR phosphorylation. Most information on these struc-tural–functional relationships comes from studies on animal Rb protein. However, the amino acid and module conserva-tion between animal and plant Rb proteins supports that some functional activities that depend on certain structural changes are also conserved. In any case, the conservation of potential phosphorylation sites is not complete and there-fore detailed studies on the role of specific phosphorylation sites on different RBR functions are needed. This is impor-tant not only for the cell-cycle E2F-dependent events but also for those involving the interaction with other cellular factors—e.g. chromatin, cell fate, cell division—for which no obvious counterparts with animal Rb can be easily iden-tified. Therefore, we envision a whole set of genomic and proteomic analysis in the future that would shed light on the less-well defined functions of plant RBR during plant growth and development.
Acknowledgements
This work is supported by MINECO (grant BFU2012-34821 to CG and grant BFU2011-23434 to IRT) and by an institu-tional grant of Fundación Ramón Areces to the Centro de Biología Molecular Severo Ochoa.
References
Ach RA, Durfee T, Miller AB, Taranto P, Hanley-Bowdoin L, Zambryski PC, Gruissem W. 1997. RRB1 and RRB2 encode maize retinoblastoma-related proteins that interact with a plant D-type cyclin and geminivirus replication protein. Molecular Cell Biology 17, 5077–5086.
Balog ER, Burke JR, Hura GL, Rubin SM. 2011. Crystal structure of the unliganded retinoblastoma protein pocket domain. Proteins 79, 2010–2014.
Bennett T, Scheres B. 2010. Root development-two meristems for the price of one? Current Topics in Developmental Biology 91, 67–102.
Borghi L, Gutzat R, Futterer J, Laizet Y, Hennig L, Gruissem W. 2010. Arabidopsis RETINOBLASTOMA-RELATED is required for stem cell maintenance, cell differentiation, and lateral organ production. The Plant Cell 22, 1792–1811.
Burke JR, Deshong AJ, Pelton JG, Rubin SM. 2010. Phosphorylation-induced conformational changes in the retinoblastoma protein inhibit E2F transactivation domain binding. Journal of Biological Chemistry 285, 16286–16293.
Burke JR, Hura GL, Rubin SM. 2012. Structures of inactive retinoblastoma protein reveal multiple mechanisms for cell cycle control. Genes and Development 26, 1156–1166.
Burki F, Okamoto N, Pombert JF, Keeling PJ. 2012. The evolutionary history of haptophytes and cryptophytes: phylogenomic evidence for separate origins. Proceedings of the Royal Society B 279, 2246–2254.
Calo E, Quintero-Estades JA, Danielian PS, Nedelcu S, Berman SD, Lees JA. 2010. Rb regulates fate choice and lineage commitment in vivo. Nature 466, 1110–1114.
Cao L, Peng B, Yao L, Zhang X, Sun K, Yang X, Yu L. 2010. The ancient function of RB–E2F pathway: insights from its evolutionary history. Biology Direct 5, 55.
Chinnam M, Goodrich DW. 2011. RB1, development, and cancer. Current Topics in Developmental Biology 94, 129–169.
Cross FR, Buchler NE, Skotheim JM. 2011. Evolution of networks and sequences in eukaryotic cell cycle control. Philosophical Transactions of the Royal Society B 366, 3532–3544.
Cruz-Ramirez A, Diaz-Trivino S, Blilou I, et al. 2012. A bistable circuit involving SCARECROW-RETINOBLASTOMA integrates cues to inform asymmetric stem cell division. Cell 150, 1002–1015.
Cui H, Levesque MP, Vernoux T, Jung JW, Paquette AJ, Gallagher KL, Wang JY, Blilou I, Scheres B, Benfey PN. 2007. An evolutionarily conserved mechanism delimiting SHR movement defines a single layer of endodermis in plants. Science 316, 421–425.
Dahl M, Meskiene I, Bogre L, Ha DT, Swoboda I, Hubmann R, Hirt H, Heberle-Bors E. 1995. The D-type alfalfa cyclin gene cycMs4 complements G1 cyclin-deficient yeast and is induced in the G1 phase of the cell cycle. The Plant Cell 7, 1847–1857.
DeCaprio JA. 2009. How the Rb tumor suppressor structure and function was revealed by the study of adenovirus and SV40. Virology 384, 274–284.
Derelle R, Lang BF. 2012. Rooting the eukaryotic tree with mitochondrial and bacterial proteins. Molecular Biology and Evolution 29, 1277–1289.
De Smet I, Beeckman T. 2011. Asymmetric cell division in land plants and algae: the driving force for differentiation. Nature Reviews Molecular Cell Biology 12, 177–188.
Desvoyes B, Ramirez-Parra E, Xie Q, Chua NH, Gutierrez C. 2006. Cell type-specific role of the retinoblastoma/E2F pathway during Arabidopsis leaf development. Plant Physiology 140, 67–80.
De Veylder L, Beeckman T, Beemster GT, et al. 2002. Control of proliferation, endoreduplication and differentiation by the Arabidopsis E2Fa-DPa transcription factor. EMBO Journal 21, 1360–1368.
Dudits D, Abraham E, Miskolczi P, Ayaydin F, Bilgin M, Horvath GV. 2011. Cell-cycle control as a target for calcium, hormonal and developmental signals: the role of phosphorylation in the retinoblastoma-centred pathway. Annals of Botany 107, 1193–1202.
Ebel C, Mariconti L, Gruissem W. 2004. Plant retinoblastoma homologues control nuclear proliferation in the female gametophyte. Nature 429, 776–780.
Esch JJ, Chen M, Sanders M, Hillestad M, Ndkium S, Idelkope B, Neizer J, Marks MD. 2003. A contradictory GLABRA3 allele helps define gene interactions controlling trichome development in Arabidopsis. Development 130, 5885–5894.
at CSIC
on Decem
ber 17, 2013http://jxb.oxfordjournals.org/
Dow
nloaded from

Page 8 of 10 | Desvoyes et al.
Fountain M, Murray J, Beck E. 1999. Isolation of a full-length cDNA encoding a retinoblastoma protein (accession no. AJ011681) from suspension-cultured photoautotrophic Chenopodium rubrum L. cells. (99-003). Plant Physiology 119, 363.
Friend SH, Bernards R, Rogelj S, Weinberg RA, Rapaport JM, Albert DM, Dryja TP. 1986. A human DNA segment with properties of the gene that predisposes to retinoblastoma and osteosarcoma. Nature 323, 643–646.
Fung YK, Murphree AL, T’Ang A, Qian J, Hinrichs SH, Benedict WF. 1987. Structural evidence for the authenticity of the human retinoblastoma gene. Science 236, 1657–1661.
Gonzalez N, Vanhaeren H, Inze D. 2012. Leaf size control: complex coordination of cell division and expansion. Trends in Plant Science 17, 332–340.
Goodrich DW. 2006. The retinoblastoma tumor-suppressor gene, the exception that proves the rule. Oncogene 25, 5233–5243.
Grafi G, Burnett RJ, Helentjaris T, Larkins BA, DeCaprio JA, Sellers WR, Kaelin WG. 1996. A maize cDNA encoding a member of the retinoblastoma protein family: involvement in endoreduplication. Proceedings of the National Academy of Sciences, USA 93, 8962–8967.
Gusti A, Baumberger N, Nowack M, Pusch S, Eisler H, Potuschak T, De Veylder L, Schnittger A, Genschik P. 2009. The Arabidopsis thaliana F-box protein FBL17 is essential for progression through the second mitosis during pollen development. PLoS One 4, e4780.
Gutierrez C. 2000. DNA replication and cell cycle in plants: learning from geminiviruses. EMBO Journal 19, 792–799.
Gutierrez C. 2005. Coupling cell proliferation and development in plants. Nature Cell Biology 7, 535–541.
Gutierrez C. 2009. The Arabidopsis cell division cycle. The Arabidopsis Book 7, e0120.
Gutierrez C, Ramirez-Parra E, Castellano MM, del Pozo JC. 2002. G(1) to S transition: more than a cell cycle engine switch. Currents Opinion in Plant Biology 5, 480–486.
Gutzat R, Borghi L, Gruissem W. 2012. Emerging roles of RETINOBLASTOMA-RELATED proteins in evolution and plant development. Trends in Plant Science 17, 139–148.
Hampl V, Hug L, Leigh JW, Dacks JB, Lang BF, Simpson AG, Roger AJ. 2009. Phylogenomic analyses support the monophyly of Excavata and resolve relationships among eukaryotic ‘supergroups’. Proceedings of the National Academy of Sciences, USA 106, 3859–3864.
Hanley-Bowdoin L, Settlage SB, Orozco BM, Nagar S, Robertson D. 2000. Geminiviruses: models for plant DNA replication, transcription, and cell cycle regulation. Critical Reviews in Biochemistry and Molecular Biology 35, 105–140.
Hassler M, Singh S, Yue WW, Luczynski M, Lakbir R, Sanchez-Sanchez F, Bader T, Pearl LH, Mittnacht S. 2007. Crystal structure of the retinoblastoma protein N domain provides insight into tumor suppression, ligand interaction, and holoprotein architecture. Molecular Cell 28, 371–385.
Henley SA, Dick FA. 2012. The retinoblastoma family of proteins and their regulatory functions in the mammalian cell division cycle. Cell Division 7, 10.
Huntley R, Healy S, Freeman D, et al. 1998. The maize retinoblastoma protein homologue ZmRb-1 is regulated during leaf development and displays conserved interactions with G1/S regulators and plant cyclin D (CycD) proteins. Plant Molecular Biology 37, 155–169.
Jeske H. 2009. Geminiviruses. Current Topics in Microbiology and Immunology 331, 185–226.
Jordan CV, Shen W, Hanley-Bowdoin LK, Robertson DN. 2007. Geminivirus-induced gene silencing of the tobacco retinoblastoma-related gene results in cell death and altered development. Plant Molecular Biology 65, 163–175.
Kianianmomeni A, Nematollahi G, Hallmann A. 2008. A gender-specific retinoblastoma-related protein in Volvox carteri implies a role for the retinoblastoma protein family in sexual development. The Plant Cell 20, 2399–2419.
Kieffer M, Master V, Waites R, Davies B. 2011. TCP14 and TCP15 affect internode length and leaf shape in Arabidopsis. The Plant Journal 68, 147–158.
Kim HJ, Oh SA, Brownfield L, Hong SH, Ryu H, Hwang I, Twell D, Nam HG. 2008. Control of plant germline proliferation by SCF(FBL17) degradation of cell cycle inhibitors. Nature 455, 1134–1137.
Knudsen ES, Knudsen KE. 2006. Retinoblastoma tumor suppressor: where cancer meets the cell cycle. Experimental Biology and Medicine (Maywood, NJ) 231, 1271–1281.
Kong LJ, Orozco BM, Roe JL, et al. 2000. A geminivirus replication protein interacts with the retinoblastoma protein through a novel domain to determine symptoms and tissue specificity of infection in plants. EMBO Journal 19, 3485–3495.
Kosugi S, Ohashi Y. 2002. E2Ls, E2F-like repressors of Arabidopsis that bind to E2F sites in a monomeric form. Journal of Biological Chemistry 277, 16553–16558.
Kosugi S, Ohashi Y. 2003. Constitutive E2F expression in tobacco plants exhibits altered cell cycle control and morphological change in a cell type-specific manner. Plant Physiology 132, 2012–2022.
Lamber EP, Beuron F, Morris EP, Svergun DI, Mittnacht S. 2013. Structural insights into the mechanism of phosphoregulation of the retinoblastoma protein. PLoS One 8, e58463.
Kuwabara A, Gruissem W. Arabidopsis retinoblastoma-related and Polycomb group proteins: cooperation during plant cell differentiation and development. Journal of Experimental Botany 65, xxx–xxx.
Lammens T, Boudolf V, Kheibarshekan L, et al. 2008. Atypical E2F activity restrains APC/CCCS52A2 function obligatory for endocycle onset. Proceedings of the National Academy of Sciences, USA 105, 14721–14726.
Lee JO, Russo AA, Pavletich NP. 1998. Structure of the retinoblastoma tumour-suppressor pocket domain bound to a peptide from HPV E7. Nature 391, 859–865.
Lee WH, Bookstein R, Hong F, Young LJ, Shew JY, Lee EY. 1987. Human retinoblastoma susceptibility gene: cloning, identification, and sequence. Science 235, 1394–1399.
Lendvai A, Pettko-Szandtner A, Csordas-Toth E, Miskolczi P, Horvath GV, Gyorgyey J, Dudits D. 2007. Dicot and monocot plants differ in retinoblastoma-related protein subfamilies. Journal of Experimental Botany 58, 1663–1675.
at CSIC
on Decem
ber 17, 2013http://jxb.oxfordjournals.org/
Dow
nloaded from

RBR in cell division | Page 9 of 10
Li ZY, Li B, Dong AW. 2012. The Arabidopsis transcription factor AtTCP15 regulates endoreduplication by modulating expression of key cell-cycle genes. Molecular Plant 5, 270–280.
Ma Z, Wu Y, Jin J, Yan J, Kuang S, Zhou M, Zhang Y, Guo AY. 2013. Phylogenetic analysis reveals the evolution and diversification of cyclins in eukaryotes. Molecular Phylogenetics and Evolution 66, 1002–1010.
Magyar Z, Atanassova A, De Veylder L, Rombauts S, Inze D. 2000. Characterization of two distinct DP-related genes from Arabidopsis thaliana. FEBS Letters 486, 79–87.
Magyar Z, Horvath B, Khan S, Mohammed B, Henriques R, De Veylder L, Bako L, Scheres B, Bogre L. 2012. Arabidopsis E2FA stimulates proliferation and endocycle separately through RBR-bound and RBR-free complexes. EMBO Journal 31, 1480–1493.
Manning AL, Dyson NJ. 2012. RB: mitotic implications of a tumour suppressor. Nature Reviews Cancer 12, 220–226.
Mariconti L, Pellegrini B, Cantoni R, Stevens R, Bergounioux C, Cella R, Albani D. 2002. The E2F family of transcription factors from Arabidopsis thaliana. Novel and conserved components of the retinoblastoma/E2F pathway in plants. Journal of Biological Chemistry 277, 9911–9919.
Menges M, de Jager SM, Gruissem W, Murray JA. 2005. Global analysis of the core cell cycle regulators of Arabidopsis identifies novel genes, reveals multiple and highly specific profiles of expression and provides a coherent model for plant cell cycle control. The Plant Journal 41, 546–566.
Morohashi K, Grotewold E. 2009. A systems approach reveals regulatory circuitry for Arabidopsis trichome initiation by the GL3 and GL1 selectors. PLoS Genetics 5, e1000396.
Morris EJ, Dyson NJ. 2001. Retinoblastoma protein partners. Advances in Cancer Research 82, 1–54.
Moulager M, Corellou F, Verge V, Escande ML, Bouget FY. 2010. Integration of light signals by the retinoblastoma pathway in the control of S phase entry in the picophytoplanktonic cell Ostreococcus. PLoS Genetics 6, e1000957.
Munro S, Carr SM, La Thangue NB. 2012. Diversity within the pRb pathway: is there a code of conduct? Oncogene 31, 4343–4352.
Murray J, Freeman D, Greenwood J, et al. 1998. Plant D cyclins and retinoblastoma (Rb) protein homologues. In: Francis D, Dudits D, Inzé D (eds), Plant cell division . Colchester, UK,: Portland Press. pp. 99–127.
Nagar S, Pedersen TJ, Carrick KM, Hanley-Bowdoin L, Robertson D. 1995. A geminivirus induces expression of a host DNA synthesis protein in terminally differentiated plant cells. The Plant Cell 7, 705–719.
Nurse P, Bissett Y. 1981. Gene required in G1 for commitment to cell cycle and in G2 for control of mitosis in fission yeast. Nature 292, 558–560.
Olson BJ, Oberholzer M, Li Y, Zones JM, Kohli HS, Bisova K, Fang SC, Meisenhelder J, Hunter T, Umen JG. 2010. Regulation of the Chlamydomonas cell cycle by a stable, chromatin-associated retinoblastoma tumor suppressor complex. The Plant Cell 22, 3331–3347.
Park JA, Ahn JW, Kim YK, Kim SJ, Kim JK, Kim WT, Pai HS. 2005. Retinoblastoma protein regulates cell proliferation, differentiation, and endoreduplication in plants. The Plant Journal 42, 153–163.
Punta M, Coggill PC, Eberhardt RY, et al. 2012. The Pfam protein families database. Nucleic Acids Research 40, D290–301.
Ramirez-Parra E, Gutierrez C. 2000. Characterization of wheat DP, a heterodimerization partner of the plant E2F transcription factor which stimulates E2F–DNA binding. FEBS Letters 486, 73–78.
Ramirez-Parra E, Xie Q, Boniotti MB, Gutierrez C. 1999. The cloning of plant E2F, a retinoblastoma-binding protein, reveals unique and conserved features with animal G(1)/S regulators. Nucleic Acids Research 27, 3527–3533.
Riou-Khamlichi C, Huntley R, Jacqmard A, Murray JA. 1999. Cytokinin activation of Arabidopsis cell division through a D-type cyclin. Science 283, 1541–1544.
Roger AJ, Simpson AG. 2009. Evolution: revisiting the root of the eukaryote tree. 19, R165–R167.
Rubin SM. 2013. Deciphering the retinoblastoma protein phosphorylation code. Trends in Biochemical Sciences 38, 12–19.
Rubin SM, Gall AL, Zheng N, Pavletich NP. 2005. Structure of the Rb C-terminal domain bound to E2F1-DP1: a mechanism for phosphorylation-induced E2F release. Cell 123, 1093–1106.
Sabelli PA, Dante RA, Leiva-Neto JT, Jung R, Gordon-Kamm WJ, Larkins BA. 2005. RBR3, a member of the retinoblastoma-related family from maize, is regulated by the RBR1/E2F pathway. Proceedings of the National Academy of Sciences, USA 102, 13005–13012.
Sabelli PA, Hoerster G, Lizarraga LE, Brown SW, Gordon-Kamm WJ, Larkins BA. 2009. Positive regulation of minichromosome maintenance gene expression, DNA replication, and cell transformation by a plant retinoblastoma gene. Proceedings of the National Academy of Sciences, USA 106, 4042–4047.
Sabelli PA, Larkins BA. 2006. Grasses like mammals? Redundancy and compensatory regulation within the retinoblastoma protein family. Cell Cycle 5, 352–355.
Sabelli PA, Liu Y, Dante RA, et al. 2013. Control of cell proliferation, endoreduplication, cell size, and cell death by the retinoblastoma-related pathway in maize endosperm. Proceedings of the National Academy of Sciences, USA 110, E1827–1836.
Schellmann S, Hulskamp M. 2005. Epidermal differentiation: trichomes in Arabidopsis as a model system. International Journal of Developmental Biology 49, 579–584.
Soni R, Carmichael JP, Shah ZH, Murray JA. 1995. A family of cyclin D homologs from plants differentially controlled by growth regulators and containing the conserved retinoblastoma protein interaction motif. The Plant Cell 7, 85–103.
Sozzani R, Cui H, Moreno-Risueno MA, Busch W, Van Norman JM, Vernoux T, Brady SM, Dewitte W, Murray JA, Benfey PN. 2010. Spatiotemporal regulation of cell-cycle genes by SHORTROOT links patterning and growth. Nature 466, 128–132.
Ueda M, Laux T. 2012. The origin of the plant body axis. Currents Opinion in Plant Biology 15, 578–584.
Umen JG, Goodenough UW. 2001. Control of cell division by a retinoblastoma protein homolog in Chlamydomonas. Genes and Development 15, 1652–1661.
Vandepoele K, Vlieghe K, Florquin K, Hennig L, Beemster GT, Gruissem W, Van de Peer Y, Inze D, De Veylder L. 2005.
at CSIC
on Decem
ber 17, 2013http://jxb.oxfordjournals.org/
Dow
nloaded from

Page 10 of 10 | Desvoyes et al.
Genome-wide identification of potential plant E2F target genes. Plant Physiology 139, 316–328.
Vlieghe K, Boudolf V, Beemster GT, Maes S, Magyar Z, Atanassova A, de Almeida Engler J, De Groodt R, Inze D, De Veylder L. 2005. The DP-E2F-like gene DEL1 controls the endocycle in Arabidopsis thaliana. Current Biology 15, 59–63.
Weimer AK, Nowack MK, Bouyer D, Zhao X, Harashima H, Naseer S, De Winter F, Dissmeyer N, Geldner N, Schnittger A. 2012. Retinoblastoma related1 regulates asymmetric cell divisions in Arabidopsis. The Plant Cell 24, 4083–4095.
Weinberg RA. 1995. The retinoblastoma protein and cell cycle control. Cell 81, 323–330.
Wenger JP, Marks MD. 2008. E2F and retinoblastoma related proteins may regulate GL1 expression in developing Arabidopsis trichomes. Plant Signaling and Behavior 3, 420–422.
Wildwater M, Campilho A, Perez-Perez JM, Heidstra R, Blilou I, Korthout H, Chatterjee J, Mariconti L, Gruissem W, Scheres B. 2005. The RETINOBLASTOMA-RELATED gene regulates stem cell maintenance in Arabidopsis roots. Cell 123, 1337–1349.
Xie Q, Sanz-Burgos AP, Hannon GJ, Gutierrez C. 1996. Plant cells contain a novel member of the retinoblastoma family of growth regulatory proteins. EMBO Journal 15, 4900–4908.
Xie Q, Suarez-Lopez P, Gutierrez C. 1995. Identification and analysis of a retinoblastoma binding motif in the replication protein of a plant DNA virus: requirement for efficient viral DNA replication. EMBO Journal 14, 4073–4082.
Zhao X, Harashima H, Dissmeyer N, et al. 2012. A general G1/S-phase cell-cycle control module in the flowering plant Arabidopsis thaliana. PLoS Genetics 8, e1002847.
at CSIC
on Decem
ber 17, 2013http://jxb.oxfordjournals.org/
Dow
nloaded from