novel cck analogs for studying cck-b receptor
TRANSCRIPT

Novel CCK Analogs for Studying CCK-B Receptor
CHIZUKO YANAIHARA$e ATSUKAZU KUWAHARA? MUTSUAKI SUZUK1,C MINORU HOSHIN0,C MIN LI,d LI QIN ZHENG,d KAZUHISA KASHIMOT0,d YASUO
TAKEDA,d KAZUAKI IGUCH1,d TOHORU MOCHIZUKI,d AND NOBORU YANAIHARAd
a Laboratory of Pharmaceutical Sciences Osaka University School of Medicine
Suita-shi, Osaka, Japan bNational Institute for Physiological Sciences
Okazaki-shi, Aichi, Japan CLaboratory of Environmental Metabolism
Graduate School of Nutritional and Environmental Sciences and dLaboratory of Bioorganic Chemistry School of Pharmaceutical Sciences
University of Shizuoka Shizuoka-shi, Shizuoka, Japan
Cholecystokinin (CCK) is a typical brain-gut peptide that localizes not only in the endocrine system but also in the central and peripheral nervous systems and that exhibits various biological activities in different mammalian tissues and organs. Although tissue forms of CCK-related peptides have been identified and sequenced, it has still not been established which forms are released into blood as proper hormone and/or transmitter. CCK-8 possesses the full biological properties of CCK, and structure-activity studies of CCK, which have been carried out in several laboratories worldwide, have been devoted mainly to CCK-8 or CCK-7.
In an earlier study,' we demonstrated dissociation of pancreatic exocrine- stimulating (PZ) activity from gallbladder-contracting (CCK) activity in Na- succinylated [D-Trp3]-CCK-7 (Suc-[D-Trp3]-CCK-7) in anesthetized dogs. Several other synthetic analogs of CCK-7 and CCK-8, in which the Gly residue was substituted, as well as the [D-Trp3] analog were also shown to possess full or almost full PZ activities in anesthetized rats as compared with CCK-8, whereas they were considerably less potent in CCK activity in anesthetized guinea pigs. These observa- tions suggested the possible existence of multi-subtypes of CCK receptor in the gut. We then proposed the usefulness of synthetic analogs of CCK-7 and CCK-8 as tools for distinguishing between receptors associated with CCK and PZ activities. Subse- quently, we succeeded in differentiating CCK binding properties of rat pancreatic and brain receptors with the use of various synthetic CCK COOH-terminal-related peptides and analogs.* Our results, together with recent progress in research on CCK receptors in other laboratories, encouraged us to extend our study on characteriza- tion of CCK receptors in various tissues specifically with the use of synthetic CCK
ePresent address: 2-2 Yamadaoka, Suita-shi, Osaka-fu 565, Japan. 107
108 ANNALS NEW YORK ACADEMY OF SCIENCES
analogs. We thus continued our efforts to develop synthetic analogs of CCK-7 and CCK-8 which are considered appropriate for the study.
CCK-%RELATED PEPTIDES AND RAT BRAIN AND PANCREATIC RECEPTORS
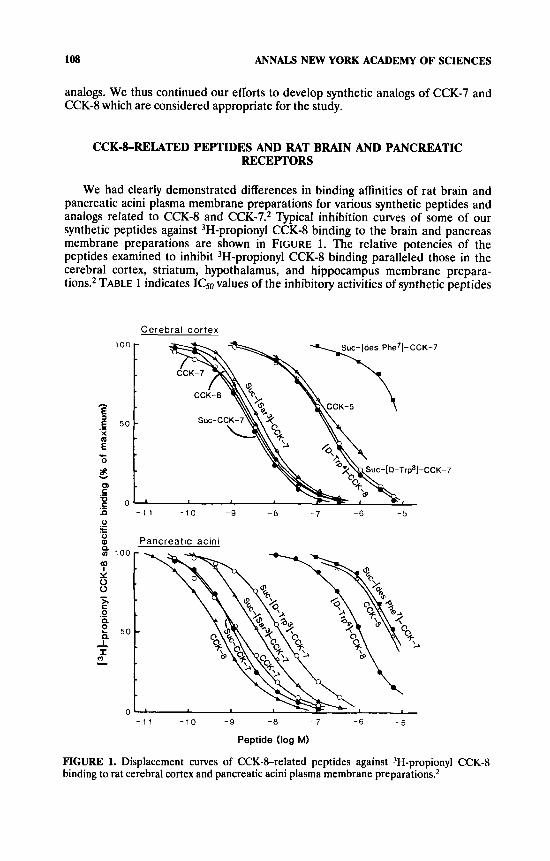
We had clearly demonstrated differences in binding affinities of rat brain and pancreatic acini plasma membrane preparations for various synthetic peptides and analogs related to CCKS and CCK-7.2 Typical inhibition curves of some of our synthetic peptides against 3H-propionyl CCK-8 binding to the brain and pancreas membrane preparations are shown in FIGURE 1. The relative potencies of the peptides examined to inhibit 3H-propionyl CCK-8 binding paralleled those in the cerebral cortex, striatum, hypothalamus, and hippocampus membrane prepara- tions2 TABLE 1 indicates IC,, values of the inhibitory activities of synthetic peptides
Cerebral cortex
- 9 -a -7 -6 -5 -10 0 'c 0
.- Pancreatic acini :: 100
&
.- s
aD
0 0 - z
P g 50 I I P)
- I
0 -11 -10 - 9 -a - 7 -6 - 5
Peptide (log MI
FIGURE 1. Displacement curves of CCK-brelated peptides against 3H-propionyl CCK-8 binding to rat cerebral cortex and pancreatic acini plasma membrane preparatiom2

YANAIHARA et al.: CCK ANALOGS 109
TABLE I. ICsO Valuesa for Inhibitory Activities of Synthetic CCK-&Related Peptides on 3H-Propionyl CCK-8 Specific Binding to Rat Brain and Pancreatic Plasma Membrane Preparations
Value (nM) Peptide Cerebral Cortex Pancreas
CCK-8 CCK-7 CCK-5 CCK-4 SUC-CCK-7 Suc-[des Phe7]-CCK-7
Suc-[Phe(NHSO3-)']-CCK-7 Sue[ Sar3]-CCK-7 SUC-[ D-Trp3]-CCK-7 Suc-[D-Ala3]-CCK-7 [D-Trp4]-CCK-8 [ D-Ala4]-CCK-8
NS-CCK-8
4.3 f 0.3 3.5 f 0.5 215 f 38 138 f 10 3.5 f 0.3
> 103 39.4 f 3.7 8.2 f 1.2 6.1 f 1.2 152 2 13
47.7 f 7.9 176 2 47
64.7 f 8.4
0.63 f 0.05 1.7 f 0.1
> 103 > 103
1.5 f 0.1 >I03
577 f 50 656 f 57 4.9 f 0.1
18.4 f 1.0 69.6 f 9.9 992 f 17
> 103
aValues are mean f SEM for ICSO values calculated by Hill analysis in independent experiments.
in rat cerebral cortex and pancreatic acini plasma membrane preparations. The binding potencies of CCK-&related analogs in the pancreatic acini preparation are relatively parallel to the in vivo PZ activities of the corresponding peptides.'
Suc-CCK-7 was as potent as CCK-8 in both brain and pancreas assay systems. The NHz-terminal Asp of CCK-8 is not essentially involved in CCK-8 binding to either receptor. Steigerwalt and William3 reported that CCK-4 is the minimum fully active agonist for CCK-B receptor. The relative binding potencies of CCK-5 and CCK-4 against CCK-8 in our systems were two to three orders of magnitude greater in the brain than in the pancreas. In the brain system, ICS0 values of the two shortened peptides were 30-50 times as high as that of CCK-8. It is already known that the potencies of tetragastrin (CCK-4) are about one tenth those of gastrin-17 in various gastrin actions. Suc-[des Phe7]-CCK-7, lacking the COOH-terminal Phe moiety of CCK, showed little activity in both brain and pancreatic receptors, indicating, as already known," the crucial role of the Phe residue for CCK-8 binding to both receptors.
With the use of nonsulfated CCK-8 (NS-CCK-8), the critical role of 0-sulfated Tyr of CCK-8 was confirmed in recognition of the pancreatic binding site, but not the cerebral cortex binding site. We had thus synthesized Suc-CCK-7 analogs in which 0-sulfated Tyr was substituted. In fact, Suc-[Phe(NHSO3-)I]-CCK-7 showed signifi- cantly high selectivity for the brain cortical receptor. The peptide retains substan- tially full binding activity for the brain receptor as compared with CCK-8, whereas the potency of its binding to the pancreatic receptor is four orders of magnitude less than that of CCK-8. More recently, McCort-Tranchepain et aLS reported that replacement of Tyr 0-sulfate by p-carboxylmethyl-Phe in CCK-8 retained consider- ably high affinity for CCK-B receptor.
Dibutyryl (db) cGMP is known to be a potent CCK-A receptor antagonist, and Miller et nL6 described that the db-cGMP might interact directly with CCK, thereby blocking the region of the peptide, probably COOH-terminal portion, that binds to the receptor. We observed a distinct discrepancy in the inhibiting activities of db-cGMP and some other nucleotides against 3H-propionyl CCK-8 binding to the
ANNALS NEW YORK ACADEMY OF SCIENCES
L P
Pancreatic acini
dbc : dibutyryl cyclic GTP
, ,\ATP
dbcGMP 1,. I I 1
1 oc
dbcAMP
cGMP 50
Cerebral cortex
cGMP
GTP
. - . .. ~. c., Y
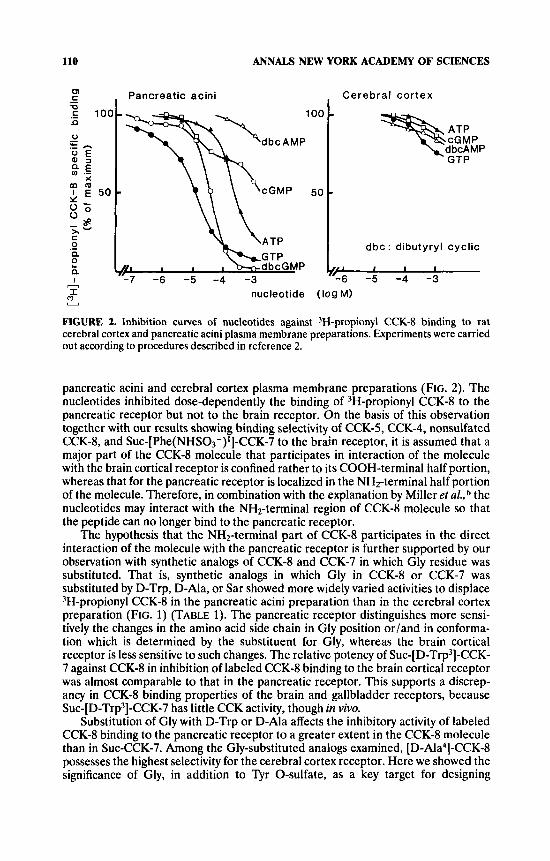
FIGURE 2. Inhibition cuwes of nucleotides against 3H-propionyl CCK-8 binding to rat cerebral cortex and pancreatic acini plasma membrane preparations. Experiments were carried out according to procedures described in reference 2.
pancreatic acini and cerebral cortex plasma membrane preparations (FIG. 2). The nucleotides inhibited dose-dependently the binding of 3H-propionyl CCK-8 to the pancreatic receptor but not to the brain receptor. On the basis of this observation together with our results showing binding selectivity of CCK-5, CCK-4, nonsulfated CCK-8, and Suc-[Phe(NHS03-)*]-CCK-7 to the brain receptor, it is assumed that a major part of the CCK-8 molecule that participates in interaction of the molecule with the brain cortical receptor is confined rather to its COOH-terminal half portion, whereas that for the pancreatic receptor is localized in the NH2-terminal half portion of the molecule. Therefore, in combination with the explanation by Miller et L Z ~ . , ~ the nucleotides may interact with the NH2-terminal region of CCK-8 molecule so that the peptide can no longer bind to the pancreatic receptor.
The hypothesis that the NHz-terminal part of CCK-8 participates in the direct interaction of the molecule with the pancreatic receptor is further supported by our observation with synthetic analogs of CCK-8 and CCK-7 in which Gly residue was substituted. That is, synthetic analogs in which Gly in CCK-8 or CCK-7 was substituted by D-Trp, D-Ala, or Sar showed more widely varied activities to displace 3H-propionyl CCK-8 in the pancreatic acini preparation than in the cerebral cortex preparation (FIG. 1) (TABLE 1). The pancreatic receptor distinguishes more sensi- tively the changes in the amino acid side chain in Gly position or/and in conforma- tion which is determined by the substituent for Gly, whereas the brain cortical receptor is less sensitive to such changes. The relative potency of Suc-[D-Trp3]-CCK- 7 against CCK-8 in inhibition of labeled CCK-8 binding to the brain cortical receptor was almost comparable to that in the pancreatic receptor. This supports a discrep- ancy in CCK-8 binding properties of the brain and gallbladder receptors, because Suc-[D-Trp3]-CCK-7 has little CCK activity, though in vivo.
Substitution of Gly with D-Trp or D-Ala affects the inhibitory activity of labeled CCK-8 binding to the pancreatic receptor to a greater extent in the CCK-8 molecule than in Suc-CCK-7. Among the Gly-substituted analogs examined, [D-ma4]-CCK-8 possesses the highest selectivity for the cerebral cortex receptor. Here we showed the significance of Gly, in addition to Tyr 0-sulfate, as a key target for designing

YANAIHARA el al.: CCK ANALOGS 111
synthetic CCK-8 analogs that possess high selectivity for the brain (CCK-B) receptor and are useful in identifying possible subtypes of the receptor.
ACYLATED CCK-8 AND CCK-7 ANALOGS
Synthetic analogs related to CCK-8 that were recently prepared in several laboratories are mostly in an NHz-terminal acetyl- or t-butyloxycarbonyl (Boc)- blocked form which are expected to be enzyme-resistant (TABLE 2). Stability of biologically active peptides towards proteolytic enzymes is especially required for their in vivo use. When we found that Suc-CCK-8 and Suc-CCK-7 are as active as CCK-8 in binding to the brain receptor,2 we designed synthetic CCK-8 and CCK-7 in which NH2-termini are protected with various acyl or carboxyacyl groups, such as glutaryl (Glt), pyroglutamyl (pGlu), phtharyl (Pht), and benzyloxycarbonyl groups.
Infusion of Glt-CCK-8 or pGlu-CCK-8 into rat suprachiasmatic nucleus resulted in significant suppression of total daily food intake, while CCK-8 showed little effect.' Glt-CCK-8 was significantly more potent than pGlu-CCK-8 in the suppressive activity on food intake.
Close intraarterial injection of CCK-8 into the gastroepiploic artery supplying the antral region of the stomach induced, in nanomolar concentration, dose-dependent stimulation on antral smooth muscle contraction in anesthetized dogs.* The injection technique had the advantage of examining local peptide actions in vivo rather than systemic effects. Atropine abolished the gastric contraction by CCK-8, whereas hexamethonium failed to block the contraction, suggesting that the peptide acts on the intramural cholinergic innervation of the smooth muscle cells and not directly on the muscle. Fox et aL9 suggested that the pentagastrin receptor is present on
TABLE 2. Synthetic CCK-&Related Peptides Recently Reported Synthetic CCK-&-Related Peptides Ref.
A. CCKAnalogs Active to Both CCK-A and CCK-B Receptors Bo~-[Nle~.~~]-CCK(27-33) Ac-[ Nlezx.3 I , SarZ9]-CCK( 26-33) [L-Phe@-CH2S03H)27]-CCK(2&33) [D-P~~@-CHZSO~H)~~]-CCK(~&~~) Ac-[2-NaI3"]-CCK(27-33) A C - [ X - A I ~ ~ ~ ] - C C K ( ~ ~ - ~ ~ )
X = cyclophenyl, cyclohexyl, cyclooctyl, 1-naphthyl B. CCK Analogs for CCK-B Receptor [N-MeNle2X.31]-CCK(2&33) Boc-[NleW, N-MeNle3I]-CCK(27-33) Boc-[gNleZX, mGlyZ9, Nle3']-CCK(27-33) Boc-[gNleX, mGly2', N-MeNle31]-CCK( 27-33) Ac-Tyr(S03-)-Lys-Gly-Trp-Lys-Asp-Phe-NHz
I I CO-( CH2)-CO
Boc-y-D-Gl,u-Tyr( SO3-)-Nle-D-Lys-Trp-Nle-Asp-Phe-NH2
(BDNL) 14 14 15 15 16 17
(SNF8702) 18 19
(BC264) 19 19
(JMV310) 20
21
C. CCKAnalogs with Antagonistic Activiry Boc-Tyr( S03-)-Nle-Gly-D-Trp-Nle-Asp-O-CH2-CH&H~ 22 Boc-Try( SO~-)-Nle-Gly-D-Trp-Nle-Asp-NH-CH2-CHz-p-F-C~H~ 22 Boc-[Phg3I, 1-Nal-N(CH3)233]-CCK(30-33) 23

112 ANNALS NEW YORK ACADEMY OF SCIENCES
TABLE 3. ED% Valuesu for Stimulating Effects of Intraarterially Injected CCK-8 Analogs on Gastric Motility in Anesthetized Dogs6
Peptide EDso(ng/ml) CCK-8 2.97 f 0.63 Glt-CCK-8 4.86 f 0.62 PGIu-CCK-8 4.43 -c 0.55 Suc-[MePhe7]-CCK-7 11.87 2 l . lSb
“Values are mean f SEM (n = 8). *p < 0.05 (VS CCK-8).
cholinergic nerves of the canine gastric corpus. We therefore examined the effects of synthetic CCK-8 analogs in this system. Glt-CCK-8 and pGlu-CCK-8 were less active, but not significantly so, than CCK-8 in the local stimulating effect on gastric motility in anesthetized dogs (TABLE 3). This indicates that NH2-terminal blocking of the CCK-8 molecule does not necessarily cause enhancement of the actions of CCK-8 in in vivo experiments.
It remains to be established if the discrepancy in the relative potencies among CCK-8 and its acylated analogs, Glt-CCK-8 and pGlu-CCK-8, in the aforementioned two assay systems is attributable, at least in part, to differences in ligand recognition mechanisms between CCK receptors in the suprachiasmatic nucleus and those in the stomach muscle cell associated with the suppressive effect on food intake and the local stimulating effect on gastric smooth muscle contraction, respectively; however, the animal species used were different.
CCK-8 ANALOGS AND GASTRODUODENAL MOTILITY
Our synthetic CCK-8 analogs were also examined for their stimulatory effect on gastroduodenal motility in anesthetized dogs.l0 Peptides were infused through the femoral vein, and the gastric and duodenal mechanical activities were simultaneously recorded. Doyle et nl. l1 obsexved that CCK-&related peptides traverse the liver without significant degradation. The response reached a maximum level in about 5 minutes in the gastric region and immediately in the duodenal region after the initiation of peptide infusion, but the possibility of central effects of CCK-8 cannot be thoroughly excluded in this system. The results are summarized in TABLES 4 and 5. NHz-terminal modification of CCK-8 did not affect the stimulatory activity of CCK-8 on either gastric or duodenal motility. Nonsulfated CCK-8 showed little effect in this
TABLE 4. EDs0 Valuesu for Stimulating Effects of Intravenously Infused CCK-8 Analogs on Gastroduodenal Motility in Anesthetized Dogslo
Peptide EDSO (ng/kg/h)
Stomach Duodenum CCK-8 439 f 58.5 209 f 81.5 Glt-CCK-8 448 f 60.5 202 f 73.5 PGIu-CCK-~ 475 f 60.5 186 f 50.0 Suc-[MePhe7]-CCK-7 653 f 52.P 248 f 50.5
“Values are mean 2SEM (n = 6). ’p <0.05 (VS CCK-8).

YANAIHARA er 01.: CCK ANALOGS 113
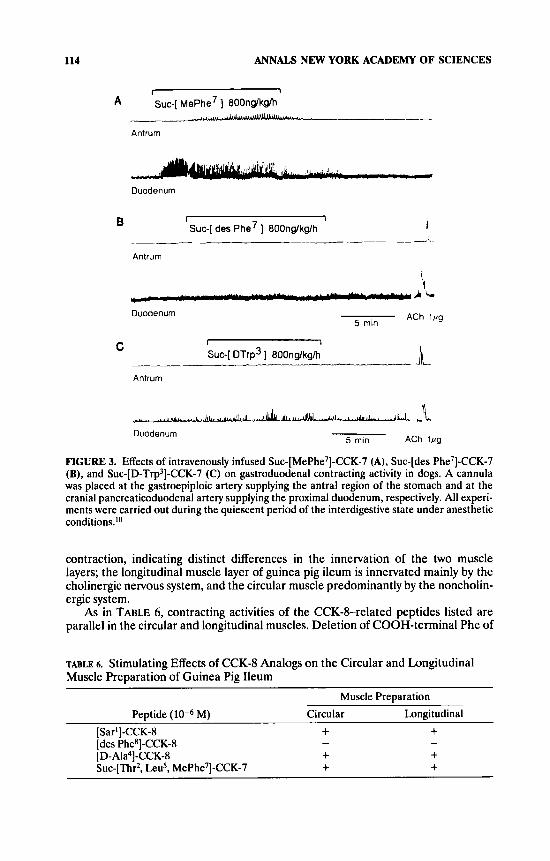
system. Displacement of Phe in Suc-CCK-7 with N-methylated Phe (Suc-[MePhe7]- CCK-7) significantly decreased the stimulating effect on gastric motility as seen in intraarterial injection as mentioned in the preceding section, but its activity on duodenal motility was not changed (TABLE 4) (FIG. 3). This analog, Suc-[MePhe7]- CCK-7, also retained essentially full potency in pancreatic exocrine activity in rats (volume: 91 2 23% [mean ? SEMI vs CCK-8) and in gallbladder contraction activity in guinea pigs (83% [62 - 105%], mean (95% confidence limit], vs CCK-8). On the other hand, removal of COOH-terminal Phe from Suc-CCK-7, namely, Suc-[des Phe7]-CCK-7, disrupted the stimulating activity on gastroduodenal motility (FIG. 3), as seen in most biological assay systems for CCK-8.
Suc-[D-Trp3]-CCK-7 was already mentioned as an interesting analog that selec- tively stimulates pancreatic exocrine secretion in anesthetized dogs.' Moreover, the binding activities of this peptide to rat pancreatic and brain cortical receptors were approximately one thirtieth and one fortieth of those of CCK-8, respectively (TABLE 1). This [D-Trp3] analog was substantially inactive to gastroduodenal motility in the system used (FIG. 3). It is noteworthy that Suc-[Sar3]-CCK-7 showed little activity on gastroduodenal motility (TABLE 5) even though it was almost as potent as CCK-8 in rat cerebral cortex receptor binding assay (TABLE 1).
TABLE 5. Effects of Intravenously Injected CCK-&Related Peptides on Gastroduodenal Motility in Anesthetized Dogs'O
Peptide (800 ng/kg/h)
Motility Stomach Duodenum
CCK-8 GI t-CCK-8 PGIu-CCK-8 NS-CCK-8 SUC-[ MePhe7]-CCK-7 Suc-[des Phe7]-CCK-7 SUC-[ D-Trp'l-CCK-7 SUC-[ Sar3]-CCK-7
These observations suggest possible differences between CCK receptors linked to stimulation of gastric and duodenal smooth muscle contractions, respectively, and also between the gastric and/or duodenal receptors and the cerebral cortex receptor.
CCK-8 ANALOGS AND GUINEA PIG ILEUM CONTRACTION
A recent study by Forno et aLI2 provides evidence that two CCK receptors, CCK-A and CCK-B/gastrin, mediate contraction of guinea pig isolated ileal longitu- dinal muscle. We used some of our synthetic CCK-8 analogs to characterize neuronal CCK receptors in guinea pig ileum preparation. In the experiment, L-shaped muscle strips were used to simultaneously record circular and longitudinal muscle contrac- t i o n ~ . ' ~ These preparations contain all elements of the gastrointestinal tract (enteric nervous system, muscle layers, and mucosa). CCK-8 showed distinct contractile activities on both circular and longitudinal muscles (FIG. 4). In these preparations, atropine completely blocked CCK-8-induced longitudinal, but not circular, muscle
114 ANNALS NEW YORK ACADEMY OF SCIENCES
A 'Suc-[ MePhe7 ] 800ng/kg/h' .,.,..,.I..I.,I.. -_ Antrum
Duodenum
Duodenum ACh lpg 5 min
I 1
SUC-[ DTrp3 ] 800nglkg/h h C ___ Antrum
----- ACh l r g
Duodenum 5 min
FIGURE 3. Effects of intravenously infused Suc-[MePhe7]-CCK-7 (A), Suc-[des Phe7]-CCK-7 (B), and Suc-[D-Trp3]-CCK-7 (C) on gastroduodenal contracting activity in dogs. A cannula was placed at the gastroepiploic artery supplying the antral region of the stomach and at the cranial pancreaticoduodenal artery supplying the proximal duodenum, respectively. All experi- ments were carried out during the quiescent period of the interdigestive state under anesthetic conditions.1°
contraction, indicating distinct differences in the innervation of the two muscle layers; the longitudinal muscle layer of guinea pig ileum is innervated mainly by the cholinergic nervous system, and the circular muscle predominantly by the noncholin- ergic system.
As in TABLE 6, contracting activities of the CCK-&related peptides listed are parallel in the circular and longitudinal muscles. Deletion of COOH-terminal Phe of
TABLE 6. Stimulating Effects of CCK-8 Analogs on the Circular and Longitudinal Muscle Preparation of Guinea Pig Ileum
Muscle Preparation Peptide M) Circular Longitudinal
[Sar1]-CCK-8 + + [des Phe8]-CCK-8 - - [D-Ala4]-CCK-8 + + Suc-[Thr*, Leus, MePhe7]-CCK-7 + +
YANAIHARA et al.: CCK ANALOGS 115
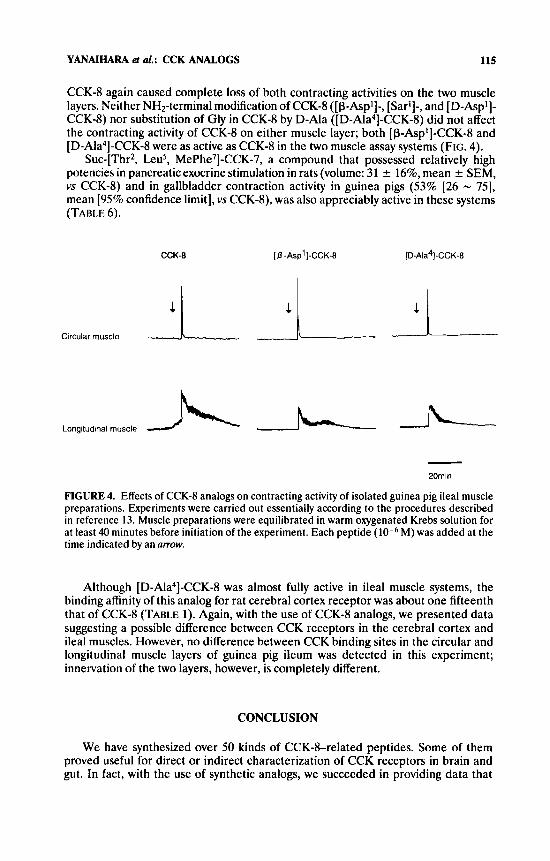
CCK-8 again caused complete loss of both contracting activities on the two muscle layers. Neither NH2-terminal modification of CCK-8 ( [p-Aspi]-, [Sari]-, and [D-Aspl]- CCK-8) nor substitution of Gly in CCK-8 by D-Ala ([D-Ala4]-CCK-8) did not affect the contracting activity of CCK-8 on either muscle layer; both [P-Asp1]-CCK-8 and [D-Ala4]-CCK-8 were as active as CCK-8 in the two muscle assay systems (FIG. 4).
Suc-[Thr2, Leu5, MePhe7]-CCK-7, a compound that possessed relatively high potencies in pancreatic exocrine stimulation in rats (volume: 31 ? 16%, mean -c SEM, vs CCK-8) and in gallbladder contraction activity in guinea pigs (53% [26 - 751, mean [95% confidence limit], vs CCKd), was also appreciably active in these systems (TABLE 6).
CCK-a [ B -Asp1]-CCK-8 [D-Ala4]-CCK-8
Longitudinal muscle A L L 20min
FIGURE 4. Effects of CCK-8 analogs on contracting activity of isolated guinea pig ileal muscle preparations. Experiments were carried out essentially according to the procedures described in reference 13. Muscle preparations were equilibrated in warm oxygenated Krebs solution for at least 40 minutes before initiation of the experiment. Each peptide M) was added at the time indicated by an arrow.
Although [D-Ala4]-CCK-8 was almost fully active in ileal muscle systems, the binding affinity of this analog for rat cerebral cortex receptor was about one fifteenth that of CCK-8 (TABLE 1). Again, with the use of CCK-8 analogs, we presented data suggesting a possible difference between CCK receptors in the cerebral cortex and ileal muscles. However, no difference between CCK binding sites in the circular and longitudinal muscle layers of guinea pig ileum was detected in this experiment; innervation of the two layers, however, is completely different.
CONCLUSION
We have synthesized over 50 kinds of CCK-%related peptides. Some of them proved useful for direct or indirect characterization of CCK receptors in brain and gut. In fact, with the use of synthetic analogs, we succeeded in providing data that

116 ANNALS NEW YORK ACADEMY OF SCIENCES
support the existence of subtypes of neuronal CCK receptors (CCK-B) in the gut. These data will be clues for further detailed study of CCK-B/gastrin receptor.
From pharmacological and pharmaceutical points of view, several recently developed elegant nonpeptide CCK receptor antagonists are most promising in CCK research. Molecular biology is disclosing amino acid sequences of CCK receptor molecules. However, for differential analysis of CCK receptors, especially in ligand- binding states, various CCK analogs are indispensable, and development of peptide analogs that can differentiate each subtype receptor, if any, in the CCK-B family is an important target for synthesis of CCK-8 analogs.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
REFERENCES
YANAIHARA, C., N. SUGIURA, K. KASHIMOTO, M. KONDO, M. KAWAMURA, S. NARUSE, A. YASUI & N. YANAIHARA. 1985. Dissociation of pancreozymin (PZ) activity from cholecystokinin (CCK) activity by Nu-carboxyacyl CCK7 and CCK8 analogues with a substituted glycine. Biomed. Res. 6 111-1 15.
TAKEDA, Y., M. HOSHINO, N. YANAIHARA, C. YANAIHARA, J. ISOBE, N. SUGIURA, K. KASHIMOTO, Y. TAKANO & H. KAMIYA. 1989. Comparison of CCK-8 receptors in the pancreas and brain of rats using CCK-8 analogues. Japan. J. Pharmacol. 4 9 471481.
STEIGERWALT, R. W. & J. A. WILLIAMS. 1981. Characterization of cholecystokinin receptors on rat pancreatic membranes. Endocrinology 109 1746-1753.
GALAS, M.-C., M.-F. LIGNON, M. RODRIGUEZ, C. MENDRE, P. FULCKAND, J. LAUR & J. MARTINEZ. 1988. Structure-activity relationship studies on cholecystokinin: Analogues with partial agonist activity. Am. J. Physiol. 254: G176G182.
MCCORT-TRANCHEPAIN, I., D. FICHEUX, C. DURIEUX & B. P. ROGUES. 1992. Replacement of Tyr-SO3H by a p-carboxymethyl-phenylalanine in a CCK8-derivative preserves its high affinity for CCK-B receptor. Int. J. Peptide Protein Res. 3 9 48-57.
MILLER, L. J., W. M. REILLY, S. A. ROSENZWEIG, J. D. JAMIESON & V. L. W. Go. 1983. A soluble interaction between dibutyryl cyclic guanosine 3'5'-monophosphate and chole- cystokinin: A possible mechanism for the inhibition of cholecystokinin activity. Gastro- enterology 84: 1505-1511.
MORI, T., K. NAGAI, H. NAKAGAWA & N. YANAIHARA. 1986. Intracranial infusion of CCK-8 derivatives suppresses food intake in rats. Am. J. Physiol. 251: R718-R723.
KUWAHARA, A., K. OZAWA & N. YANAIHAKA. 1986. Effects of cholesystokinin-octapeptide on gastric motility of anesthetized dogs. Am. J. Physiol. 251: G678-G681.
Fox, J. E. T., E. E. DANIEL, J. JURY, A. E. Fox & S. M. COLLINS. 1983. Sites and mechanisms of action of neuropeptides on canine gastric motility differ in vivo and in vitro. Life Sci. 33: 817-825.
KUWAHARA, A., N. SUGIURA & N. YANAIHARA. 1989. Structure-function relationship of cholecystokinin to canine gastroduodenal motility. Biomed. Res. 1 0 123-130.
DOYLE, J. W., M. M. WOLFE & J. E. MCGUIGAN. 1984. Hepatic clearance of gastrin and cholecystokinin peptides. Gastroenterology 87: 60-68.
FORNO, G. D., C. PIETRA, M. URCIUOLI, F. T. M. V. AMSTERDAM, G. TOSON, G. GAVIARAGHI & D. TRIST. 1992. Evidence for two cholecystokinin receptors mediating the contraction of the guinea pig isolated ileum longitudinal muscle myenteric plexus. J. Pharmacol. Exp. Ther. 261: 1056-1063.
KUWAHARA, A,, T. OZAKI & N. YANAIHARA. 1989. Galanin suppresses the neurally- evoked circular muscle contractions in the guinea-pig ileum. Eur. J. Pharmacol.
RON, D., C. GILON, M. HANANI, A. VROMEN, Z. SELINGER & M. CHOREV. 1992. N-methylated analogs of A~[Nle~*.~~]CCK(26-33): Synthesis, activity, and receptor selectivity. J. Med. Chem. 35 2806-2811.
& B. P. ROQUES. 1991. Solid phase synthesis of a fully active analogue of cholecystoki- nin using the acid-stable Boc-Phe @-CHz) SOsH as a substitute for Boc-Tyr(SO3H) in CCK8. Int. J. Peptide Protein Res. 37: 331-340.
164: 175-178.
GONZALEZ-MUNIZ, R., F. CORNILLE, F. BERGERON, D. FICHEUX, J. POTHIER, C. DUR~EUX

YANAIHARA et al.: CCK ANALOGS 117
16.
17.
18.
19.
20.
21.
22.
23.
DANHO, W., J. W. TILLEY, S.-J. SHIUEY, 1. KULESHA, J. SWISTOK, R. MAKOFSKE, J. MICHALEWSKY, R. WAGNER, J. TRISCARI, D. NELSON, F. Y. CHIRUUO & s. WEATHER- FORD. 1992. Structure activity studies of tryptophanM modified analogs of Ac-CCK-7. Int. J. Peptide Protein Res. 39 337-347.
TILLEY, J. W., W. DANHO, S.-J. SHIUEY, I. KULESHA, R. MAKOFSKE, G. L. OLSON, E.
CHIRUZZO & S. WEATHERFORD. 1992. Structure activity of C-terminal modified analogs of Ac-CCK-7. Int. J. Peptide Protein Res. 39 322-336.
Cholecystokinin analogues with high affinity and selectivity for brain membrane recep- tors. Int. J. Peptide Protein Res. 35: 566573.
B. P. ROGUES. 1988. Enzyme-resistant CCK analogs with high affinities for central receptors. Peptides 9 835-841.
1990. Synthesis of cyclic analogues of cholecystokinin highly selective for central receptors. Int. J. Peptide Protein Res. 35: 441-451.
CHARPENTIER, B., A. DOR, P. ROY, P. ENGLAND, H. PHARM, C. DURIEUX & B. P. ROQUES. 1989. Synthesis and binding affinities of cyclic and related linear analogues of CCK8 selective for central receptors. J. Med. Chem. 32: 1184-1190.
MARTINEZ. 1988. 2-Phenylethyl ester and 2-phenylethyl amide derivative analogues of the C-terminal hepta- and octapeptide of cholecystokinin. Int. J. Peptide Protein Res. 3 2 384-395.
CORRINGER, P. J., J. H. WENG, B. Ducos, C. DURIEUX, P. BOUDEAU, A. BOHME & B. P. ROQUES. 1993. CCK-B agonist or antagonist activities of structurally hindered and peptidase-resistant Boc-CCK4 derivatives. J. Med. Chem. 3 6 166172.
CHIANG, V. K. RUSIECKI, R. WAGNER, J. MICHALEWSKY, J. TRISCARI, D. NELSON, F. Y.
HRUBY, V. J., s. FANG, R. KNAPP, W. KAZMIERSKI, G. K. LUl & H. I. YAMAMURA. 1990.
CHARPENTIER, B., C. DURIEUX, D. PELAPRAT, A. DOR, M. REIBAUD, J.-C. BLANCHARD &
RODRIGUEZ, M., M. AMBLARD, M.-C. GALAS, M.-F. LIGNON, A. AUMELAS & J. MARTINEZ.
FULCRAND, P., M. RODRIGUEZ, M.-C. GALAS, M.-F. LIGNON, J. LAUR, A. AUMELAS & J.