norepinephrine transporter (net), serotonin transporter (sert), vesicular monoamine transporter...
TRANSCRIPT

Norepinephrine Transporter (NET), Serotonin Transporter (SERT),
Vesicular Monoamine Transporter (VMAT2) and Organic Cation
Transporters (OCT1, 2 and EMT) in Human Placenta from
Pre-eclamptic and Normotensive Pregnancies
B. Bottalico, I. Larsson, J. Brodszki, E. Hernandez-Andrade, B. Casslen, K. Marsal and
S. R. Hansson*
Department of Obstetrics and Gynecology, Lund University Hospital, Klinikgatan, 221 85, Lund, Scania, Sweden
Paper accepted 14 October 2003
Pre-eclampsia is one of the most common causes of perinatal and maternal morbidity and mortality. High blood pressure andproteinuria are important clinical signs of pre-eclampsia. Sympathetic overactivity and elevated level of circulating vaso activesubstances, such as monoamines has been shown. Extracellular concentrations of monoamines are normally kept low by specifictransporter proteins of which many are expressed in the placenta. In this study we used in situ hybridization and real-time PCRto study the gene expression of monoamine transporters, such as NET, SERT, VMAT2, EMT and OCT1/2, in normal as wellas in pre-eclamptic placentae. We demonstrated high expression of NET mRNA in the trophoblast cells of the anchoring villi anda lower expression intensity in the chorionic villi. SERT mRNA was mainly detected in chorionic villi. VMAT2 mRNA was notdetected in the central part of the placenta but was present in the spiral arteries of placenta bed biopsies, in cytokeratin positivecells. EMT mRNA was mainly detected in the intra lobular septa and together with OCT1 and OCT2 mRNAs also expressed inscattered cells of placental vessel adventitias. Moreover, quantitative analysis showed a significant lower expression of NET andEMT mRNAs in pre-eclamptic placentae as compared to the control group. A defective gene expression or function of thesemonoamines transporters might explain the elevated concentrations of monoamines in pre-eclamptic patients. Monoaminetransporters may serve as a protective mechanism preventing vasoconstriction in the placental vascular bed and thereby securinga stable blood flow to the fetus.
� 2003 Elsevier Ltd. All rights reserved.Placenta (2004), 25, 518–529
INTRODUCTION
Pre-eclampsia (toxemia) is today still one of the most commoncauses of perinatal and maternal morbidity and mortality. Theoverall incidence is 3–7 per cent of pregnancies, but mainlyprimigravidae are affected. The clinical manifestations ofpre-eclampsia appear after 20 weeks of gestation and arecharacterized by low circulating blood volume, haemoconcen-tration, high blood pressure and proteinuria. Severe cases ofpre-eclampsia furthermore involve pathologic activation of thecoagulation and fibrinolytic systems. Without interventionthe condition may develop into severe epileptic seizures—eclampsia. Only symptomatic treatment of the conditionis available and termination of the pregnancy remains theonly curative intervention [1]. Removal of the placenta isbelieved to be the underlying mechanism for resolution of thesymptoms [2].
The aetiology is still not known, but defective invasion byplacental trophoblast cells into the muscle layers of the spiralarteries seems to be a background key factor. This leads toaltered placental blood flow, which in addition may result inintra uterine growth restriction (IUGR). A generalized in-flammatory process in vascular endothelial cells is a hallmarkof pre-eclampsia [2–5]. Placental derived factors have beensuggested, but so far no specific culprit has been found.The hypertension is due to peripheral vasoconstriction andelevated resistance. Increased activity in the sympatheticnervous system or elevated concentrations of circulatingvaso-active substances, are both important factors in thepathophysiology of hypertension. In fact, sympathetic over-activity is a significant finding in pre-eclampsia [6]. Nor-epinephrine (NE) is the principal neurotransmitter in thesympathetic nervous system and also a hormone released bythe adrenal medulla during stress. Increased blood levels ofmonoamines such as norepinephrine (NE) [7,8] and serotonin(5-HT) [9] have been shown in pre-eclampsia as well as inIUGR.
* To whom correspondence should be addressed. Tel.:+46462223011; E-mail: [email protected]
Placenta (2004), 25, 518–529doi:10.1016/j.placenta.2003.10.017
0143-4004/$–see front matter � 2003 Elsevier Ltd. All rights reserved.

Since monoamines have potent physiological effectsthroughout the body, their extracellular concentrations aretightly regulated. The most important regulators are specificmembrane bond transporter proteins that mediate re-uptake ofmonoamines from the extra cellular fluid [10,11]. In the lastdecade, several genes coding for transporter proteins have beencloned and characterized [12–14]. These include the transport-ers for 5-HT (SERT) [15,16], NE (NET) [10] and DA (DAT)[17,18]. The intracellular fate of monoamines is either degra-dation by intracellular enzymes, catechol-O-methyltransferase(COMT) or monoamine oxidase (MAO), or repacking invesicles in order to again be available for release. Accumulationof monoamines into vesicles involves vesicular monoaminetransporters (VMAT1 and VMAT2) [19].
Additional, non-neuronal corticosterone-sensitive monoam-ine uptake (uptake 2) has been known for many years [20] butthe responsible transporter proteins was only recently cloned[21–26]. The three known organic cation transporters (OCT1,OCT2 and EMT) are mainly expressed in the intestine, liver,kidney and placenta. Unlike the neuronal monoamine trans-porters, which regulate synaptic levels of monoamines, theOCTs play a key role in clearance of monoamines from theblood stream. They have a broad compound affinity transport-ing all biogenic amines as well as exogenous drugs andxenobiotics [27].
Monoamine transporters have been of particular interestbecause of their central role in modern treatment of depressionand anxiety with serotonin specific reuptake inhibition (SSRI)[28,29], and because of their involvement in the mechanismsfor drug addiction [30]. Cocaine and amphetamine blockuptake of monoamines in a way similar to that of the SSRI,leading to increased levels of extracellular monoamines. Co-caine intoxication reportedly gives rise to a pre-eclampsia likecondition in pregnant women [31].
Thus, knowledge of expression patterns of the transportersin normal placentae as compared to placentae from pregnanciescomplicated by pre-eclampsia is of great importance for under-standing and resolving the contribution of monoaminergicmechanisms in the pathogenesis of pre-eclampsia. Thus, we
used in situ hybridization and real-time PCR to study themRNA species for NET, SERT, VMAT2, EMT, OCT1 andOCT2 in placenta tissue from pre-eclamptic and normalpregnancies.
MATERIAL AND METHODS
Collection of placenta tissue
Placental tissue was collected at the Department of Obstetricsand Gynecology, Lund University Hospital. Sampling afterinformed consent was approved by the Ethical CommitteeReview Board for studies in human subjects. Placental tissuefrom ten pre-eclamptic and seven normal pregnancies wasincluded in the study. The controls were matched for maternalage and parity (Table 1). Caesarean section was performed intwo (29 per cent) control and five (50 per cent) pre-eclampticcases. Pre-eclampsia was either defined as blood pressure of>140/90 mmHg and proteinuria of >0.03 g/l, or a rise ofblood pressure >20 mmHg from the first trimester of preg-nancy. Patients with essential hypertension and renal or othersystemic diseases were excluded.
Tissue sampling and handling
Tissue samples were taken immediately after delivery from theplacenta and in the case of Ceasarean section from the uterinewall. Two specific areas within the placenta were sampled: (1)a section, 10�10�4 mm, from the maternal surface, at theentrance of spiral arteries and (2) a cube 10�10�10 mm fromthe central part of the placenta consisting of villi. Placenta bedbiopsies were collected from the uterine wall during Caesareansection in order to study gene expression in the modified spiralarteries. Collected samples were snap frozen on dry ice andstored at �80(C. Tissue sections (12 µm), were thawmounted on to silanized slides and stored at �80(C untilhybridization. Fresh frozen tissue rather than fixative-treated
Table 1. Clinical characteristics of included subjects and offspring
Normal pregnant Pre-eclampsia
N 7 10Maternal age (years) 32.3�1.4 31.4�1.8Parity (nulliparity) 2�0.4 (0) 1.7�0.5 (1 of 10)Gestation length (days/weeks) 274.7�3.9/(39.2) 251.4�8.6 (35.9)*Systolic pressure (mmHg) 118.7�3.8 145.1�4.1*Diastolic pressure (mmHg) 77.1�3.1 100.3�2.5*Proteinuria (g) ND 2.4�0.8*Gestational weight (g)/sex F : M 3723�160.7/3 : 4 2908�305.7/1 : 9*Apgar scores (1 min) 9.1�0.1 9.0�0.1Apgar scores (5 min) 10.0�0 9.7�0.2Apgar scores (10 min) 10�0 9.9�0.1
The results are expressed as mean�. ND=not detected, F=female, M=male.* Significant differences between the groups, P<0.05
Bottalico et al.: Monoamine Transporters in Pre-eclampsia 519

tissue was used in order to maximize the sensitivity for mRNAdetection. Thawing of tissue did not occur prior to sectioningto ensure best possible tissue integrity.
RNA probes
For the human NET mRNA, a probe was used correspondingto 561 NT (316–877), Genbank accession no. NM001043 [28].For the human 5HTT mRNA, a probe was used correspond-ing to 475 NT (1845–2320), Genbank accession no. NM00104[32]. For the vesicular monoamine transporter (VMAT2),corresponding to 486 NT (1300–1786), Genbank accession no.L23205 [12]. For the human OCT1 mRNA, a probe was usedcorresponding to 460 NT (20–480), Genbank accession no.NM_003057 [21–23]. For the human OCT2 mRNA, a probewas used corresponding to 505 NT (50–555), Genbankaccession no. NM_003058 [21,22]. For the human EMT(OCT3) mRNA, a probe was used corresponding to 440 NT(1300–1740), Genbank accession no. NM_021977 [24].
DNA templates were generated by polymerase chain reac-tion (PCR) from the different cDNAs using bipartite primersconsisting of either a T7 RNA promoter and a downstreamgene-specific sequence (anti-sense) or a T3 RNA promoterand an upstream gene-specific primer (sense). PCR reactionsusing 1 ng cDNA, 1 µ primers, 200 µ dNTPs, 3 mMgCl2, 10 m Tris, pH 8.3, 50 m KCl, 2.5 units Taqpolymerase (Boerhinger Mannheim) were amplified at 95(Cfor 1 min, 70(C (NET), 60(C (SERT), 65(C (VMAT2),62(C (OCT1), 66(C (OCT2), 63(C (EMT) for 1 min and72(C for 2.5 min for 30 cycles with a final extension at 72(Cfor 10 min. DNA templates were purified from agarose gelsusing GeneClean (Bio101) and thereafter sequenced using acycle sequencing reaction kit (ABI PRISM, Big dye).
Complementary RNA (cRNA) probes were transcribedfrom 25 ng of gel-purified DNA template using 35S-UTP(Dupont NEN, 1300 Ci/mmol) and either T3 or T7 RNApolymerase according to manufacturer’s instructions (AmbionMAXIscript) to generate sense and antisense probes,respectively.
RNA
Hybridization
Tissue sections were fixed, dehydrated and delipidated aspreviously described [33]. Sections were hybridized (20–24 h,55(C) with 2�106 cpm of denatured 35S-cRNA probe per80 µl hybridization buffer consisting of 20 m Tris–HCl(pH 7.4), 1 m EDTA (pH 8.0), 300 m NaCl, 50 per centformamide, 10 per cent dextran sulphate, 1�Denhardt’s,25 mg/ml yeast tRNA, 100 µg/ml salmon sperm DNA,250 µg/ml total yeast RNA (fraction XI, Sigma), 150 mdithiothreitol (DTT), 0.15 per cent sodium thiosulfate (NTS),and 0.15 per cent sodium dodecyl sulphate (SDS). Following
washes to remove excess probe, slides were apposed to KodakHyperfilm Biomax MR for 3 days, then coated with nucleartrack emulsion (NTB-3, Kodak). After a 4 week exposure at4(C, slides were developed in Dektol (Kodak), fixed andcounterstained with a Giemsa stain.
RNA extractions
Total RNA was extracted from frozen tissue using Trizol�
(Gibco BRL) according to manufacturer’s instructions.Proteoglycan and polysaccharide contaminations were removedby adding isopropanol followed by salt precipitation with 0.8 sodium citrate and 1.2 sodium chloride. The quality ofRNA samples were determined by electrophoresis through a1.5 per cent agaros/2 per cent formalin denaturing gel with a1� MOPS buffer (Intergen company). RNA loading mix(GenHunter) was used to verify the 18S and 28S RNA bandsunder UV light. Only samples with visible 18S/28S bandswere included for further analysis.
cDNA synthesis
RNA was reverse transcribed according to protocols fromapplied biosystems.
In a 50 µl reaction containing: 0.5 µg total RNA, and a finalconcentrations of 1� TaqMan RT buffer, 5.5 m MgCl2,500 µ dNTPs, 2,5 µ random hexamers, 0.4 U/µl RNaseinhibitor, and 1.25 U/µl MultiScribe Reverse Transcriptase.The reactions were incubated at 25(C for 10 min, at 48(C for30 min and then 5 min of inactivation at 95(C. The sampleswere stored at �20(C until further use.
Real-time PCR amplification
Gene transcripts were quantified using real-time PCR on ABIPRISM� 7000 sequence detection system (Applied Biosys-tems). Primers and probes were designed using the PrimerExpress� software program or alternatively ordered fromAssays on-Design/Demande (Applied Biosystems). Eachprimer pair was located on different exons of the investigatedgene in order to avoid genomic DNA contamination (Table 2).Oligonucleotide probes labelled with fluorogenic dye, 6carboxyfluorescein (Fam) and quenched with 6 carboxy-tetramethylrhodamine (Tamra) (Table 2). PCR reactions werecarried out in a 25 µl final volume containing final concen-trations: 1� Universal PCR Master Mix (Applied Biosys-tems), 0.5 µ TaqMan probe, 0.9 µ of forward and reverseprimers respectively, and 1 µl of 10 ng/µl of a DNA aliquot.For transcripts analysed with pre-manufactured probes thereactions were carried out in a 25 µl final volume containingfinal concentrations: 1� Universal PCR Master Mix (AppliedBiosystems), 1� Assaymix (Applied Biosystems), 0.25 µprobe, 0.9 µ of forward and reverse primers respectively, and
520 Placenta (2004), Vol. 25
1 µl of 10 ng/µl of a DNA aliquot. The thermal cyclingconditions were initiated by UNG activation at 50(C for 2 minand an initial denaturation at 95(C for 10 min, then 40 cyclesat 95(C for 15 s, annealing at 60(C. for 1 min. Two negativecontrols, without template, were included in every amplifi-cations. RNA samples were tested for genomic DNA contami-nation prior further investigation. For each reaction, triplicateor duplicate assay was carried out. Transcript of �-actin, asa housekeeping gene, was quantified as endogenous RNA ofreference to normalize each sample. Quantification wasachieved through a calibration curve obtained by serial 10-folddilutions of the template DNA (0.08–80 ng). Results areexpressed as relative values.
Immunohistochemistry
Frozen sections were fixed with 4 per cent paraformaldehydein 0.1 phosphate buffer for 5 min at room temperature (RT).After rinsing with 1�phosphate-buffered saline (PBS) theslides were blocked in 1 per cent bovine serum albumin (BSA)and 0.6 per cent Triton X-100 in PBS for 30 min at RT. Thepresence of cytokeratin, an epithelial cell marker, was detectedusing a 1 : 1000 dilution of a mouse monoclonal anti-humancytokeratin (Dako). Sections were incubated overnight at 4(Cand additional 2 h in RT. After rinsing with PBS, endogenousperoxidase activity was blocked in methanol containing 3 percent H2O2 for 10 min followed by a rinse in PBS for 10 min.Biotin-SP-conjugated anti mouse IgG (Jackson Laboratories)was used as secondary antibody at a 1 : 1000 dilution for 1 h atRT. The primary and secondary antiserum were diluted inPBS containing 1 per cent bovine serum albumin (BSA) and0.6 per cent Triton X-100. After rinsing in PBS the slides wereincubated in streptavidin-HRP (Vectastain elite ABC kit,Vector Laboratories) in PBS containing 5 per cent normal goatserum and 0.6 per cent Triton X-100 for 1 h at RT. After afinal rinse in PBS, followed by 0.1 Tris–HCl (pH 8),immune complexes were visualized in 0.4 mg/ml 3,3’-diaminobenzidine (DAB) containing 0.03 per cent H2O2.Sections were air-dried and coverslipped.
Microphotograph and figure preparation
Microphotographs were prepared using an Axiophot micro-scope (Olympus) equipped for darkfield and brightfieldmicroscopy with a digital camera (Olympus DP50-CU).Captured images were assembled electronically using AdobePhotoshop 7.0. Figures were printed on to matte-finishedpaper by a Fujix Pictrography 3000 (Fuji) printer at 400 dpiresolution.
Statistics
Real-time PCR results are presented as box plots. The Mann–Whitney U test was used to evaluate the significanceof differences between groups. P<0.05 was consideredstatistically significant.
RESULTS
The ISHH probes used in the study were sequenced andhybridized on control tissue to verify their specificity (data notshown). None of the sense probes showed specific hybridiz-ation (data not shown). Hybridizations were repeated at leasttwice with consistent results.
Neuronal monoamine transporters
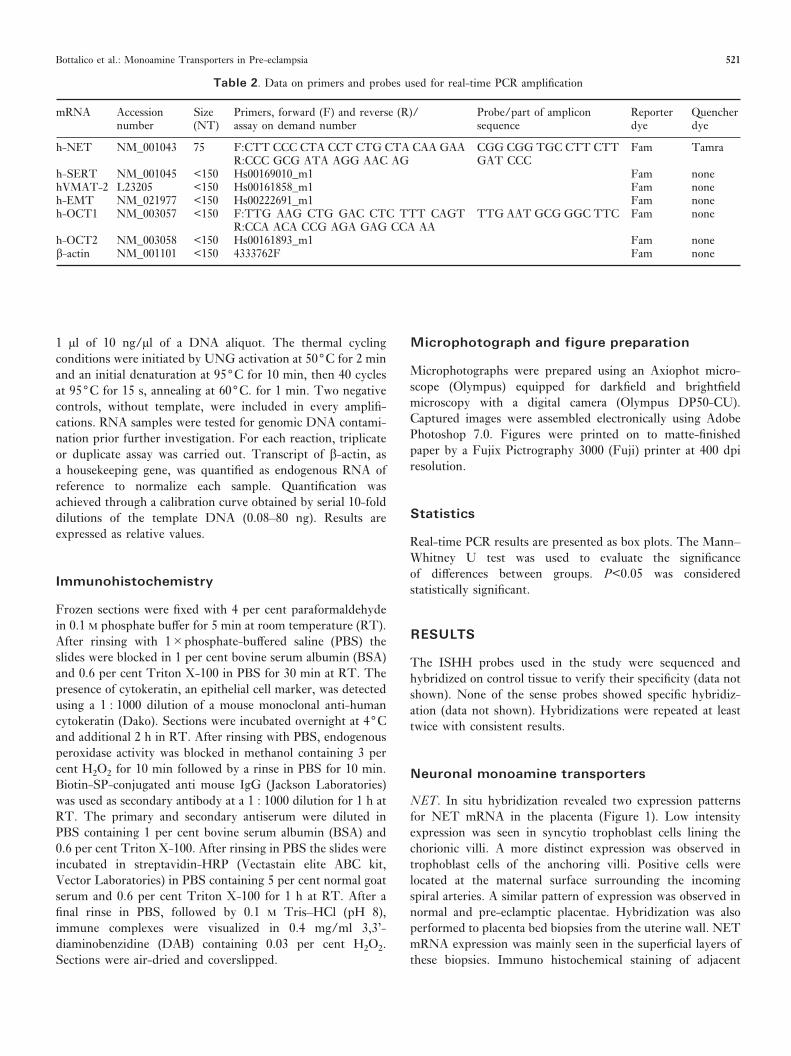
NET. In situ hybridization revealed two expression patternsfor NET mRNA in the placenta (Figure 1). Low intensityexpression was seen in syncytio trophoblast cells lining thechorionic villi. A more distinct expression was observed introphoblast cells of the anchoring villi. Positive cells werelocated at the maternal surface surrounding the incomingspiral arteries. A similar pattern of expression was observed innormal and pre-eclamptic placentae. Hybridization was alsoperformed to placenta bed biopsies from the uterine wall. NETmRNA expression was mainly seen in the superficial layers ofthese biopsies. Immuno histochemical staining of adjacent
Table 2. Data on primers and probes used for real-time PCR amplification
mRNA Accessionnumber
Size(NT)
Primers, forward (F) and reverse (R)/assay on demand number
Probe/part of ampliconsequence
Reporterdye
Quencherdye
h-NET NM_001043 75 F:CTT CCC CTA CCT CTG CTA CAA GAAR:CCC GCG ATA AGG AAC AG
CGG CGG TGC CTT CTTGAT CCC
Fam Tamra
h-SERT NM_001045 <150 Hs00169010_m1 Fam nonehVMAT-2 L23205 <150 Hs00161858_m1 Fam noneh-EMT NM_021977 <150 Hs00222691_m1 Fam noneh-OCT1 NM_003057 <150 F:TTG AAG CTG GAC CTC TTT CAGT
R:CCA ACA CCG AGA GAG CCA AATTG AAT GCG GGC TTC Fam none
h-OCT2 NM_003058 <150 Hs00161893_m1 Fam none�-actin NM_001101 <150 4333762F Fam none
Bottalico et al.: Monoamine Transporters in Pre-eclampsia 521
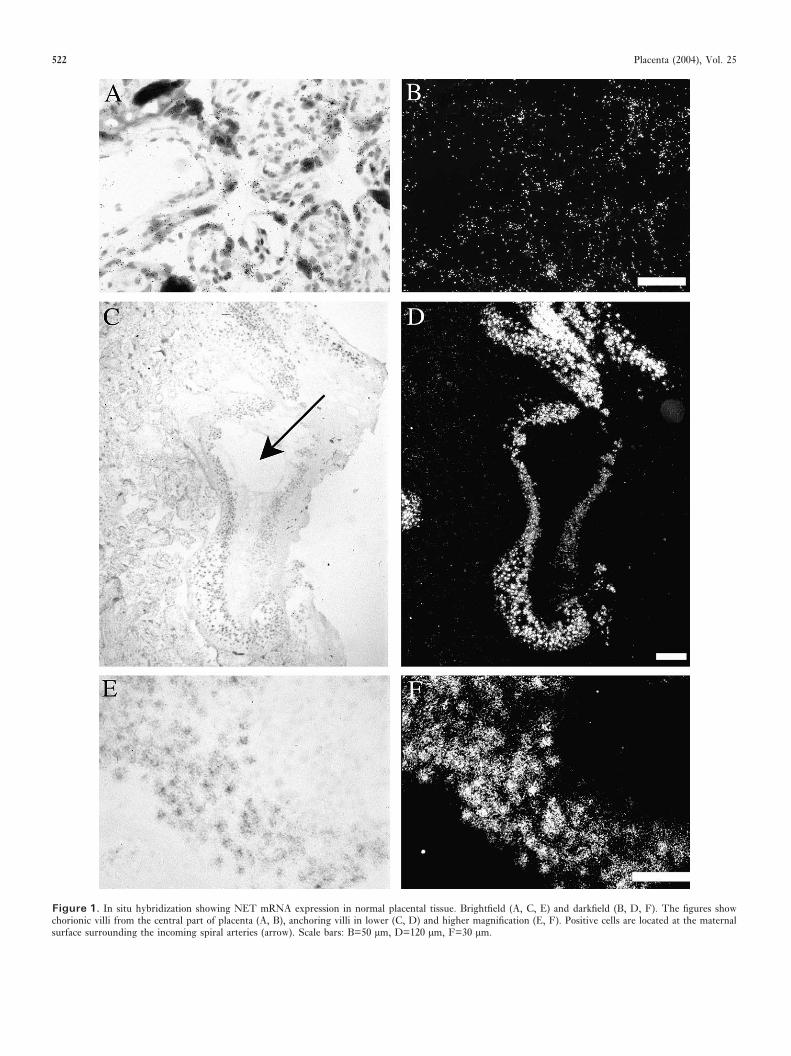
Figure 1. In situ hybridization showing NET mRNA expression in normal placental tissue. Brightfield (A, C, E) and darkfield (B, D, F). The figures showchorionic villi from the central part of placenta (A, B), anchoring villi in lower (C, D) and higher magnification (E, F). Positive cells are located at the maternalsurface surrounding the incoming spiral arteries (arrow). Scale bars: B=50 µm, D=120 µm, F=30 µm.
522 Placenta (2004), Vol. 25
sections for cytokeratin verified that the NET positive cellsco-localized with cytokeratin positive cells (Figure 2).
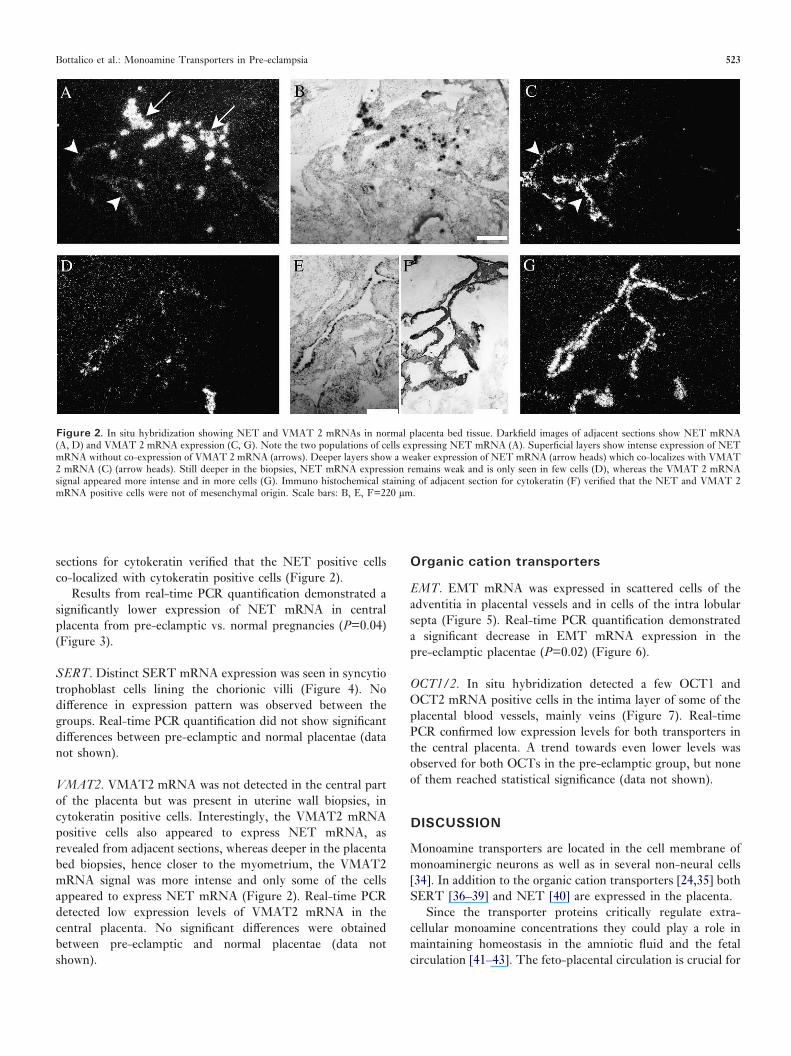
Results from real-time PCR quantification demonstrated asignificantly lower expression of NET mRNA in centralplacenta from pre-eclamptic vs. normal pregnancies (P=0.04)(Figure 3).
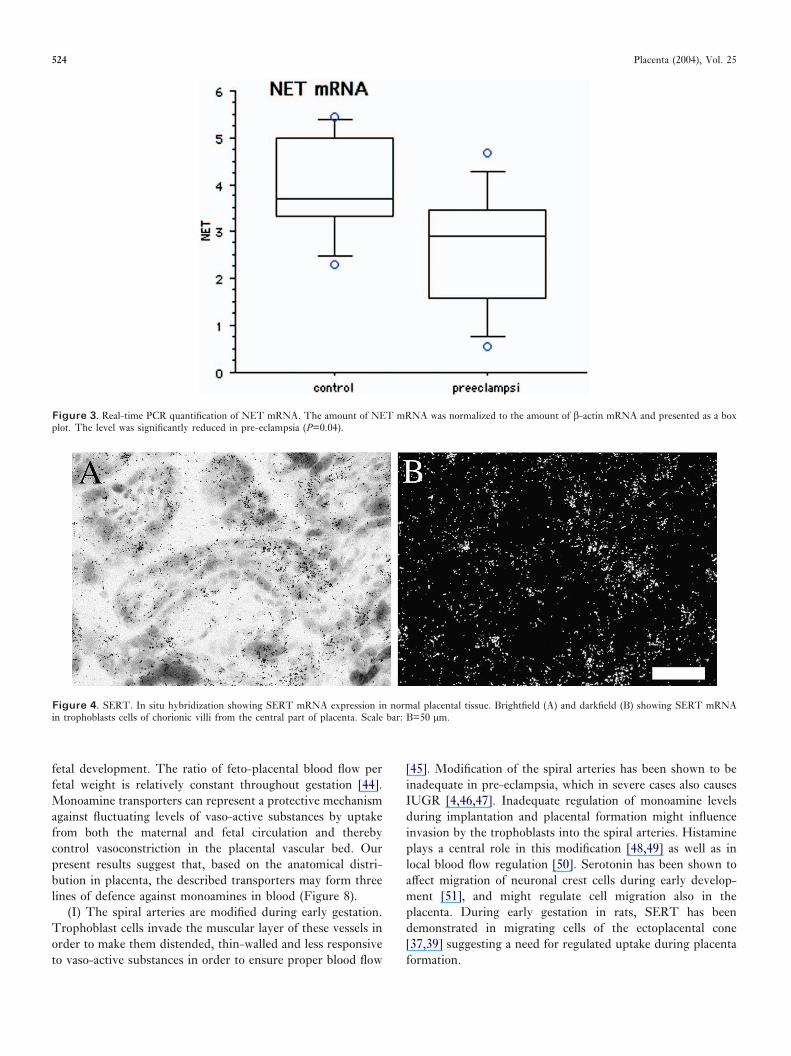
SERT. Distinct SERT mRNA expression was seen in syncytiotrophoblast cells lining the chorionic villi (Figure 4). Nodifference in expression pattern was observed between thegroups. Real-time PCR quantification did not show significantdifferences between pre-eclamptic and normal placentae (datanot shown).
VMAT2. VMAT2 mRNA was not detected in the central partof the placenta but was present in uterine wall biopsies, incytokeratin positive cells. Interestingly, the VMAT2 mRNApositive cells also appeared to express NET mRNA, asrevealed from adjacent sections, whereas deeper in the placentabed biopsies, hence closer to the myometrium, the VMAT2mRNA signal was more intense and only some of the cellsappeared to express NET mRNA (Figure 2). Real-time PCRdetected low expression levels of VMAT2 mRNA in thecentral placenta. No significant differences were obtainedbetween pre-eclamptic and normal placentae (data notshown).
Organic cation transporters
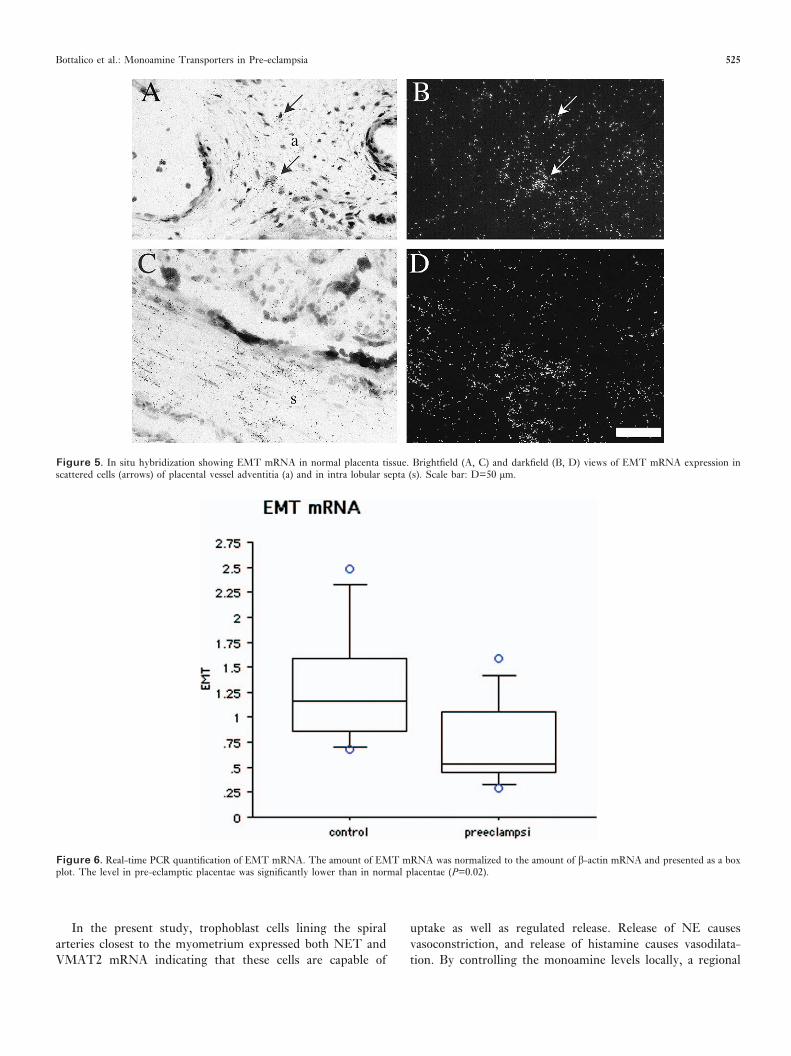
EMT. EMT mRNA was expressed in scattered cells of theadventitia in placental vessels and in cells of the intra lobularsepta (Figure 5). Real-time PCR quantification demonstrateda significant decrease in EMT mRNA expression in thepre-eclamptic placentae (P=0.02) (Figure 6).
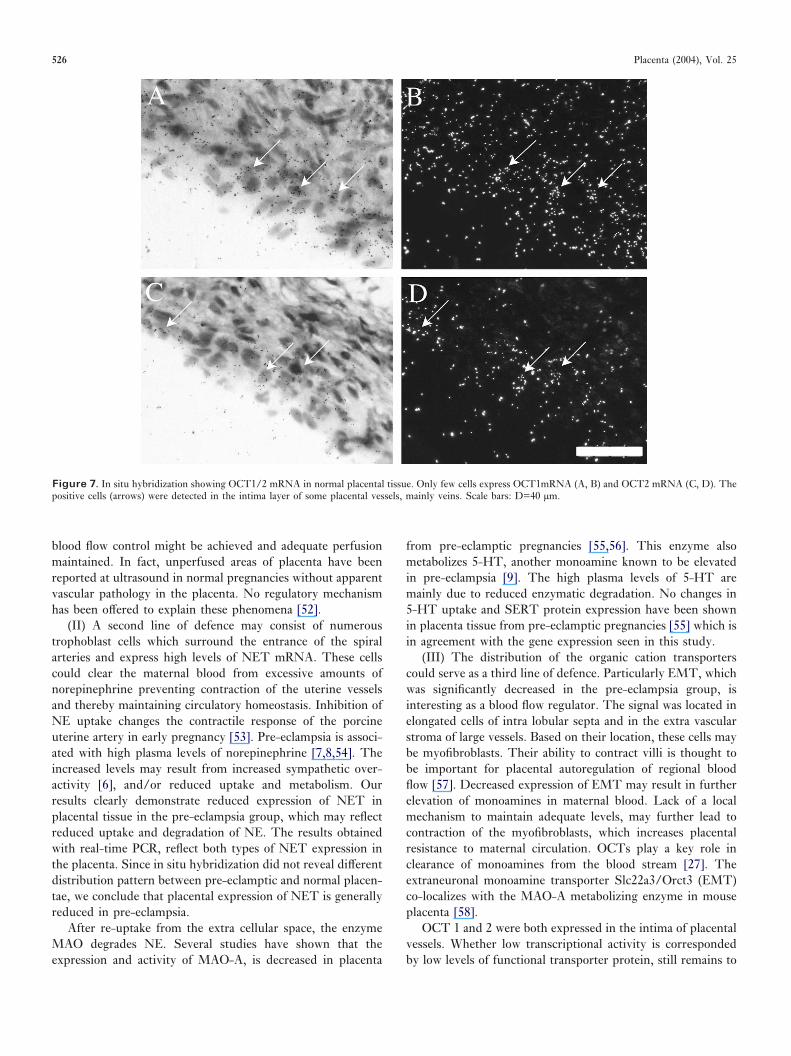
OCT1/2. In situ hybridization detected a few OCT1 andOCT2 mRNA positive cells in the intima layer of some of theplacental blood vessels, mainly veins (Figure 7). Real-timePCR confirmed low expression levels for both transporters inthe central placenta. A trend towards even lower levels wasobserved for both OCTs in the pre-eclamptic group, but noneof them reached statistical significance (data not shown).
DISCUSSION
Monoamine transporters are located in the cell membrane ofmonoaminergic neurons as well as in several non-neural cells[34]. In addition to the organic cation transporters [24,35] bothSERT [36–39] and NET [40] are expressed in the placenta.
Since the transporter proteins critically regulate extra-cellular monoamine concentrations they could play a role inmaintaining homeostasis in the amniotic fluid and the fetalcirculation [41–43]. The feto-placental circulation is crucial for
Figure 2. In situ hybridization showing NET and VMAT 2 mRNAs in normal placenta bed tissue. Darkfield images of adjacent sections show NET mRNA(A, D) and VMAT 2 mRNA expression (C, G). Note the two populations of cells expressing NET mRNA (A). Superficial layers show intense expression of NETmRNA without co-expression of VMAT 2 mRNA (arrows). Deeper layers show a weaker expression of NET mRNA (arrow heads) which co-localizes with VMAT2 mRNA (C) (arrow heads). Still deeper in the biopsies, NET mRNA expression remains weak and is only seen in few cells (D), whereas the VMAT 2 mRNAsignal appeared more intense and in more cells (G). Immuno histochemical staining of adjacent section for cytokeratin (F) verified that the NET and VMAT 2mRNA positive cells were not of mesenchymal origin. Scale bars: B, E, F=220 µm.
Bottalico et al.: Monoamine Transporters in Pre-eclampsia 523
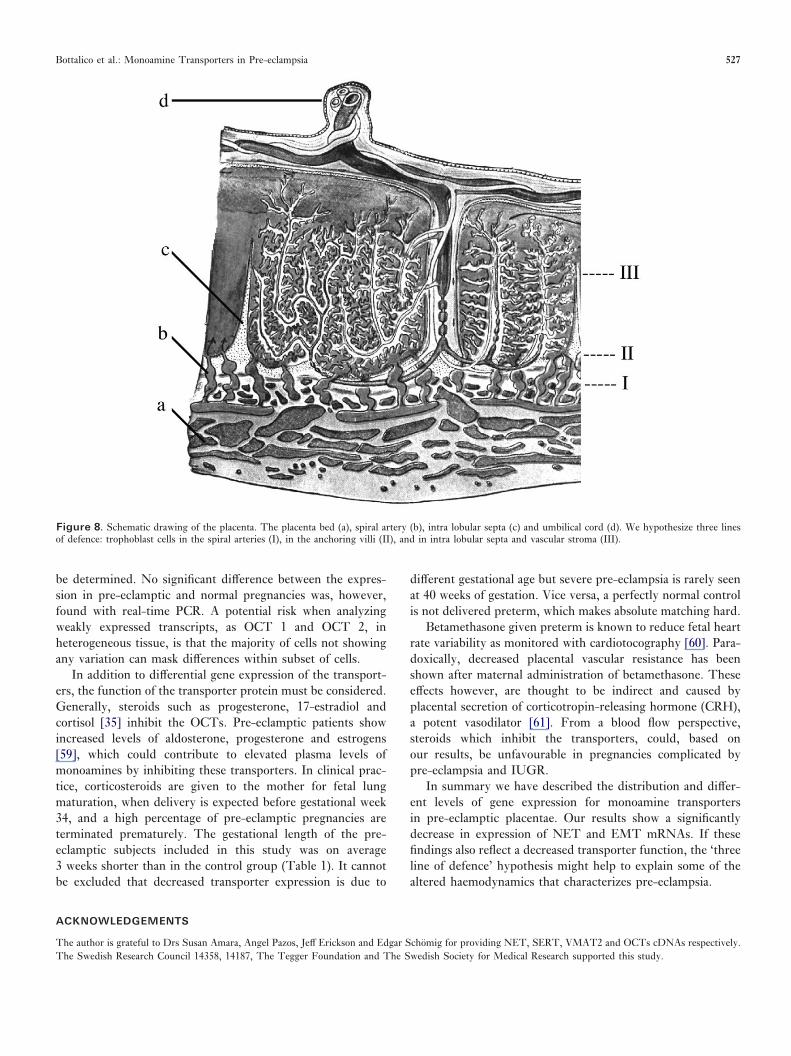
fetal development. The ratio of feto-placental blood flow perfetal weight is relatively constant throughout gestation [44].Monoamine transporters can represent a protective mechanismagainst fluctuating levels of vaso-active substances by uptakefrom both the maternal and fetal circulation and therebycontrol vasoconstriction in the placental vascular bed. Ourpresent results suggest that, based on the anatomical distri-bution in placenta, the described transporters may form threelines of defence against monoamines in blood (Figure 8).
(I) The spiral arteries are modified during early gestation.Trophoblast cells invade the muscular layer of these vessels inorder to make them distended, thin-walled and less responsiveto vaso-active substances in order to ensure proper blood flow
[45]. Modification of the spiral arteries has been shown to beinadequate in pre-eclampsia, which in severe cases also causesIUGR [4,46,47]. Inadequate regulation of monoamine levelsduring implantation and placental formation might influenceinvasion by the trophoblasts into the spiral arteries. Histamineplays a central role in this modification [48,49] as well as inlocal blood flow regulation [50]. Serotonin has been shown toaffect migration of neuronal crest cells during early develop-ment [51], and might regulate cell migration also in theplacenta. During early gestation in rats, SERT has beendemonstrated in migrating cells of the ectoplacental cone[37,39] suggesting a need for regulated uptake during placentaformation.
Figure 3. Real-time PCR quantification of NET mRNA. The amount of NET mRNA was normalized to the amount of �-actin mRNA and presented as a boxplot. The level was significantly reduced in pre-eclampsia (P=0.04).
Figure 4. SERT. In situ hybridization showing SERT mRNA expression in normal placental tissue. Brightfield (A) and darkfield (B) showing SERT mRNAin trophoblasts cells of chorionic villi from the central part of placenta. Scale bar: B=50 µm.
524 Placenta (2004), Vol. 25
In the present study, trophoblast cells lining the spiralarteries closest to the myometrium expressed both NET andVMAT2 mRNA indicating that these cells are capable of
uptake as well as regulated release. Release of NE causesvasoconstriction, and release of histamine causes vasodilata-tion. By controlling the monoamine levels locally, a regional
Figure 5. In situ hybridization showing EMT mRNA in normal placenta tissue. Brightfield (A, C) and darkfield (B, D) views of EMT mRNA expression inscattered cells (arrows) of placental vessel adventitia (a) and in intra lobular septa (s). Scale bar: D=50 µm.
Figure 6. Real-time PCR quantification of EMT mRNA. The amount of EMT mRNA was normalized to the amount of �-actin mRNA and presented as a boxplot. The level in pre-eclamptic placentae was significantly lower than in normal placentae (P=0.02).
Bottalico et al.: Monoamine Transporters in Pre-eclampsia 525
blood flow control might be achieved and adequate perfusionmaintained. In fact, unperfused areas of placenta have beenreported at ultrasound in normal pregnancies without apparentvascular pathology in the placenta. No regulatory mechanismhas been offered to explain these phenomena [52].
(II) A second line of defence may consist of numeroustrophoblast cells which surround the entrance of the spiralarteries and express high levels of NET mRNA. These cellscould clear the maternal blood from excessive amounts ofnorepinephrine preventing contraction of the uterine vesselsand thereby maintaining circulatory homeostasis. Inhibition ofNE uptake changes the contractile response of the porcineuterine artery in early pregnancy [53]. Pre-eclampsia is associ-ated with high plasma levels of norepinephrine [7,8,54]. Theincreased levels may result from increased sympathetic over-activity [6], and/or reduced uptake and metabolism. Ourresults clearly demonstrate reduced expression of NET inplacental tissue in the pre-eclampsia group, which may reflectreduced uptake and degradation of NE. The results obtainedwith real-time PCR, reflect both types of NET expression inthe placenta. Since in situ hybridization did not reveal differentdistribution pattern between pre-eclamptic and normal placen-tae, we conclude that placental expression of NET is generallyreduced in pre-eclampsia.
After re-uptake from the extra cellular space, the enzymeMAO degrades NE. Several studies have shown that theexpression and activity of MAO-A, is decreased in placenta
from pre-eclamptic pregnancies [55,56]. This enzyme alsometabolizes 5-HT, another monoamine known to be elevatedin pre-eclampsia [9]. The high plasma levels of 5-HT aremainly due to reduced enzymatic degradation. No changes in5-HT uptake and SERT protein expression have been shownin placenta tissue from pre-eclamptic pregnancies [55] which isin agreement with the gene expression seen in this study.
(III) The distribution of the organic cation transporterscould serve as a third line of defence. Particularly EMT, whichwas significantly decreased in the pre-eclampsia group, isinteresting as a blood flow regulator. The signal was located inelongated cells of intra lobular septa and in the extra vascularstroma of large vessels. Based on their location, these cells maybe myofibroblasts. Their ability to contract villi is thought tobe important for placental autoregulation of regional bloodflow [57]. Decreased expression of EMT may result in furtherelevation of monoamines in maternal blood. Lack of a localmechanism to maintain adequate levels, may further lead tocontraction of the myofibroblasts, which increases placentalresistance to maternal circulation. OCTs play a key role inclearance of monoamines from the blood stream [27]. Theextraneuronal monoamine transporter Slc22a3/Orct3 (EMT)co-localizes with the MAO-A metabolizing enzyme in mouseplacenta [58].
OCT 1 and 2 were both expressed in the intima of placentalvessels. Whether low transcriptional activity is correspondedby low levels of functional transporter protein, still remains to
Figure 7. In situ hybridization showing OCT1/2 mRNA in normal placental tissue. Only few cells express OCT1mRNA (A, B) and OCT2 mRNA (C, D). Thepositive cells (arrows) were detected in the intima layer of some placental vessels, mainly veins. Scale bars: D=40 µm.
526 Placenta (2004), Vol. 25
be determined. No significant difference between the expres-sion in pre-eclamptic and normal pregnancies was, however,found with real-time PCR. A potential risk when analyzingweakly expressed transcripts, as OCT 1 and OCT 2, inheterogeneous tissue, is that the majority of cells not showingany variation can mask differences within subset of cells.
In addition to differential gene expression of the transport-ers, the function of the transporter protein must be considered.Generally, steroids such as progesterone, 17-estradiol andcortisol [35] inhibit the OCTs. Pre-eclamptic patients showincreased levels of aldosterone, progesterone and estrogens[59], which could contribute to elevated plasma levels ofmonoamines by inhibiting these transporters. In clinical prac-tice, corticosteroids are given to the mother for fetal lungmaturation, when delivery is expected before gestational week34, and a high percentage of pre-eclamptic pregnancies areterminated prematurely. The gestational length of the pre-eclamptic subjects included in this study was on average3 weeks shorter than in the control group (Table 1). It cannotbe excluded that decreased transporter expression is due to
different gestational age but severe pre-eclampsia is rarely seenat 40 weeks of gestation. Vice versa, a perfectly normal controlis not delivered preterm, which makes absolute matching hard.
Betamethasone given preterm is known to reduce fetal heartrate variability as monitored with cardiotocography [60]. Para-doxically, decreased placental vascular resistance has beenshown after maternal administration of betamethasone. Theseeffects however, are thought to be indirect and caused byplacental secretion of corticotropin-releasing hormone (CRH),a potent vasodilator [61]. From a blood flow perspective,steroids which inhibit the transporters, could, based onour results, be unfavourable in pregnancies complicated bypre-eclampsia and IUGR.
In summary we have described the distribution and differ-ent levels of gene expression for monoamine transportersin pre-eclamptic placentae. Our results show a significantlydecrease in expression of NET and EMT mRNAs. If thesefindings also reflect a decreased transporter function, the ‘threeline of defence’ hypothesis might help to explain some of thealtered haemodynamics that characterizes pre-eclampsia.
ACKNOWLEDGEMENTS
The author is grateful to Drs Susan Amara, Angel Pazos, Jeff Erickson and Edgar Schomig for providing NET, SERT, VMAT2 and OCTs cDNAs respectively.The Swedish Research Council 14358, 14187, The Tegger Foundation and The Swedish Society for Medical Research supported this study.
Figure 8. Schematic drawing of the placenta. The placenta bed (a), spiral artery (b), intra lobular septa (c) and umbilical cord (d). We hypothesize three linesof defence: trophoblast cells in the spiral arteries (I), in the anchoring villi (II), and in intra lobular septa and vascular stroma (III).
Bottalico et al.: Monoamine Transporters in Pre-eclampsia 527

REFERENCES
[1] Schroeder BM. ACOG practice bulletin on diagnosing and managingpreeclampsia and eclampsia. Am Fam Physician 2002;66:330–1.
[2] Roberts JM, Cooper DW. Pathogenesis and genetics of preeclampsia.Lancet 2001;357:53–6.
[3] Roberts JM, Taylor RN, Musci TJ, Rodgers GM, Hubel CA,McLaughlin MK. Preeclampsia: An endothelial cell disorder. Am JObstet Gynecol 1989;161:1200–4.
[4] de Groot CJM, Taylor RN. New insights into the etiology ofpre-eclampsia. Ann Med 1993;25:243–9.
[5] Granger JP, Alexander BT, Bennett WA, Khalil RA. Pathophysiology ofpregnancy-induced hypertension. Am J Hypertens 2001;14:178S–185S.
[6] Schobel HP, Fischer T, Heuszer K, Geiger H, Schmieder RE.Preeclampsia—a state of sympathetic overactivity. N Engl J Med 1996;335:1480–5.
[7] Manyonda IT, Fenske C, Hole D, Choy MY, Wilson C. A role fornoradrenaline in pre-eclampsia: towards a unifying hypothesis for thepathophysiology. Br J Obstet Gynaecol 1998;105:641–8.
[8] Kaaja RJ, Moore PM, Yandle TG, Ylikorkala O, Frampton CM,Nicholls GM. Blood pressure and vasoactive hormones in mild pre-eclampsia and normal pregnancy. Hypertens Pregnancy 1999;18:173–87.
[9] Middelkoop CM, Dekker GA, Kraayenbrink AA, Popp-Snijders C.Platelet-poor plasma serotonin in normal and preeclamptic pregnancy.Clin Chem 1993;39:1675–8.
[10] Amara SG, Kuhar MJ. Neurotransmitter transporters: recent progress.Annu Rev Neurosci 1993;16:73–93.
[11] Brownstein MJ, Hoffman BJ. Neurotransmitter transporters. RecentProg Horm Res 1994;49:27–42.
[12] Erickson JD, Eiden LE. Functional identification and molecular cloningof a human brain vesicle monoamine transporter. J Neurochem 1993;61:2314–7.
[13] Borowsky B, Hoffman BJ. Neurotransmitter transporters: molecularbiology, function, and regulation. Int Rev Neurobiol 1995;38:139–99.
[14] Chang AS, Chang SM, Starnes DM, Schroeter S, Bauman AL, BlakelyRD. Cloning and expression of the mouse serotonin transporter. BrainRes Mol Brain Res 1996;43:185–92.
[15] Blakely RD, Berson HE, Fremeau RT Jr, Caron MG, Peek MM, PrinceHK et al. Cloning and expression of a functional serotonin transporterfrom rat brain. Nature 1991;354:66–70.
[16] Hoffman BJ. Expression cloning of a serotonin transporter: a new way tostudy antidepressant drugs. Pharmacopsychiatry 1994;27:16–22.
[17] Kilty JE, Lorang D, Amara SG. Cloning and expression of a cocaine-sensitive rat dopamine transporter. Science 1991;254:578–9.
[18] Usdin TB, Mezey E, Chen C, Brownstein MJ, Hoffman BJ. Cloning ofthe cocaine-sensitive bovine dopamine transporter. Proc Natl Acad SciU S A 1991;88:11168–71.
[19] Peter D, Liu Y, Sternini C, de Giorgio R, Brecha N, Edwards RH.Differential expression of two vesicular monoamine transporters. JNeurosci 1995;15:6179–88.
[20] Iversen LL. The uptake of catecholamine at high perfusion concen-trations in the rat isolated heart: a novel catecholamine uptake process.Br J Pharmacol 1965;25:18–33.
[21] Gorboulev V, Ulzheimer JC, Akhoundova A, Ulzheimer-Teuber I,Karbach U, Quester S et al. Cloning and characterization of two humanpolyspecific organic cation transporters. DNA Cell Biol 1997;16:871–81.
[22] Koehler MR, Wissinger B, Gorboulev V, Koepsell H, Schmid M. Thetwo human organic cation transporter genes SLC22A1 and SLC22A2 arelocated on chromosome 6q26. Cytogenet Cell Genet 1997;79:198–200.
[23] Zhang L, Dresser MJ, Gray AT, Yost SC, Terashita S, Giacomini KM.Cloning and functional expression of a human liver organic cationtransporter. Mol Pharmacol 1997;51:913–21.
[24] Grundemann D, Schechinger B, Rappold GA, Schomig E. Molecularidentification of the corticosterone-sensitive extraneuronal catecholaminetransporter. Nat Neurosci 1998;1:349–51.
[25] Kekuda R, Prasad PD, Wu X, Wang H, Fei Y-J, Leibach FH et al.Cloning and functional characterization of a potential-sensitive, poly-specific organic cation transporter (OCT3) most abundantly expressed inplacenta. J Biol Chem 1998;273:15971–9.
[26] Verhaagh S, Schweifer N, Barlow PD, Zwart R. Cloning of the mouseand human solute carrier 22a3 (Slc22a3/SLC22A3) identifies a conservedcluster of three organic cation transporters on mouse chromosome 17 andhuman 6q26–q27. Genomics 1999;55:209–18.
[27] Eisenhofer G. The role of neuronal and extraneuronal plasma membranetransporters in the activation of peripheral catecholamines. PharmacolTher 2001;91:35–62.
[28] Pacholczyk T, Blakely RD, Amara SG. Expression cloning of a cocaine-and antidepressant-sensitive human noradrenaline transporter. Nature1991;350:350–4.
[29] Tatsumi M, Groshan K, Blakely RD, Richelson E. Pharmacologicalprofile of antidepressants and related compounds at human monoaminetransporters. Eur J Pharmacol 1997;340:249–58.
[30] Schuldiner S, Steiner-Mordoch S, Yelin R, Wall SC, Rudnick G.Amphetamine derivatives interact with both plasma membrane andsecretory vesicle biogenic amine transporters. Mol Pharmacol 1993;44:1227–31.
[31] Towers CV, Pircon RA, Nageotte MP, Porto M, Garite TJ. Cocaineintoxication presenting as preeclampsia and eclampsia. Obstet Gynecol1993;81:545–7.
[32] Ramamoorthy S, Bauman AL, Moore KR, Han H, Yang-Feng T, ChangAS et al. Antidepressant- and cocaine-sensitive human serotonin trans-porter: molecular cloning, expression, and chromosomal localization.Proc Natl Acad Sci U S A 1993a;90:2542–6.
[33] Bradley DJ, Towle HC, Young WS III. Spatial and temporal expressionof �- and �-thyroid hormone receptor mRNAs, including the �2-subtype,in the developing mammalian nervous system. J Neurosci 1992;12(6):2288–302.
[34] Hoffman BJ, Hansson SR, Mezey E, Palkovits M. Distribution ofmonoamine neurotransmitter transporters in the rat brain. FrontNeuroendocrinol 1998.
[35] Hayer-Zillgen M, Bruss M, Bonisch H. Expression and pharmacologicalprofile of the human organic cation transporters hOCT1, hOCT2 andhOCT3. Br J Pharmacol 2002;136:829–36.
[36] Balkovetz DF, Tiruppathi C, Leibach FH, Mahesh VB, Ganapathy V.Evidence for an imipramine-sensitive serotonin transporter in humanplacental brush-border membranes. J Biol Chem 1989;264:2195–8.
[37] Yavarone MS, Shuey DL, Sadler TW, Lauder JM. Serotonin uptake inthe ectoplacental cone and placenta of the mouse. Placenta 1993;14:149–61.
[38] Prasad PD, Hoffmans BJ, Moe AJ, Smith CH, Leibach FH, GanapathyV. Functional expression of the plasma membrane serotonin trans-porter but not the vesicular monoamine transporter in human placentaltrophoblasts and choriocarcinoma cells. Placenta 1996;17:201–7.
[39] Hansson SR, Mezey E, Hoffman BJ. Serotonin transporter mRNAexpression in neuronal crest-derived structures and sensory pathways ofthe developing rat embryo. Neuroscience 1998.
[40] Ramamoorthy S, Prasad PD, Kulanthaivel P, Leibach FH, Blakely RD,Ganapathy V. Expression of a cocaine-sensitive norepinephrine trans-porter in the human placental syncytiotrophoblast. Biochemistry 1993b;32:1346–53.
[41] Ganapathy V, Ramamoorthy S, Leibach FH. Transport and metabolismof monoamines in the human placenta. Trophoblast Res 1993;7:35–51.
[42] Ganapathy V, Leibach FH. Placental biogenic amines and theirtransporters. In: Ramasastry BV, editor. Placental Toxicology. BocaRaton (FL): CRC; 1995, p. 161–74.
[43] Prasad PD, Leibach FH, Mahesh VB, Ganapathy V. Human placenta asa target organ for cocaine action: interaction of cocaine with the placentalserotonin transporter. Placenta 1994;15:267–78.
[44] Lingman G, Marsal K. Fetal central blood circulation in the thirdtrimester of normal pregnancy. Early Hum Dev 1986;13:137–50.
[45] Robertson WB, Khong TY, Brosens I, De Wolf F, Sheppard BL, BonnarJ. The placental bed biopsy: review from three European centers. Am JObstet Gynecol 1986;155:401–12.
[46] Khong TY, De Wolf F, Robertson WB, Brosens I. Inadequate maternalvascular response to placentation in pregnancies complicated by pre-eclampsia and small-for-gestational age infants. Br J Obstet Gynaecol1986;93:1049–59.
[47] Pijnenborg R, Anthony J, Davey DA, Rees A, Tiltman A, Vercruysse Let al. Placental bed spiral arteries in the hypertensive disorders ofpregnancy. Br J Obstet Gynaecol 1991;98:648–55.
[48] Dey SK. Role of histamine in implantation:inhibition of histidinedecarboxylase induced delayed implantation in the rabbit. Biol Reprod1981;24:867–9.
[49] Hatanaka K, Kitamura Y, Maeyama K, Watanabe T, Matsumoto K.Deciduoma formation in uterus of genetically mast cell-deficient W/Wmice. Biol Reprod 1982;27:25–8.
528 Placenta (2004), Vol. 25

[50] Barkai U, Kraicer PF. Intrauterine signaling and embryonicimplantation. Biological Signals 1996;5:111–21.
[51] Moiseiwitsch JR, Lauder JM. Serotonin regulates mouse cranial neuralcrest migration. Proc Natl Acad Sci U S A 1995;92:7182–6.
[52] Gudmundsson S, Marsal J, Dubiel P, Maly P, Marsal K, Laurini R.Power doppler, x-ray contrast angiography and morphological imaging ofthe human placenta. Prenat Neonat Med 2000;5:27–34.
[53] Laporte R, DeRoth L. Modulation of the effects of norepinephrineuptake inhibitors on the norepinephrine-induced contractile response ofthe porcine uterine artery during early pregnancy. Can J Vet Res 1997;61:214–20.
[54] Ferreira-de-Almeida JA, Amenta F, Cardoso F, Polonia JJ. Association ofcirculating endothelin and noradrenaline with increased calcium-channelbinding sites in the placental bed in pre-eclampsia. Br J Obstet Gynaecol1998;105:1104–12.
[55] Carrasco G, Cruz MA, Dominguez A, Gallardo V, Miguel P, GonzalezC. The expression and activity of monoamine oxidase A, but not of theserotonin transporter, is decreased in human placenta from pre-eclampticpregnancies. Life Sci 2000a;67:2961–9.
[56] Sivasubramaniam SD, Finch CC, Billett MA, Baker PN, Billett EE.Monoamine oxidase expression and activity in human placentae frompre-eclamptic and normotensive pregnancies. Placenta 2002;23:163–71.
[57] Demir R, Kosanke G, Kohnen G, Kertschanska S, Kaufmann P.Classification of human placental stem villi: review of structural andfunctional aspects. Microsc Res Tech 1997;38.
[58] Verhaagh S, Barlow PD, Zwart R. The extraneuronal monoaminetransporter Slc22a3/Orct3 co-localizes with the Maoa metabolizingenzyme in mouse placenta. Mech Dev 2001;100:127–30.
[59] Redman CW. Immunology of preeclampsia. Semin Perinatol 1991;15:257–62.
[60] Senat MV, Minoui S, Multon O, Fernandez H, Frydman R, Ville Y.Effect of dexamethasone and betamethasone on fetal heart rate variabilityin preterm labour: a randomised study. Br J Obstet Gynaecol 1998;105:749–55.
[61] Wallace EM, Baker LS. Effect of antenatal betamethasone administrationon placental vascular resistance. Lancet 1999;353.
Bottalico et al.: Monoamine Transporters in Pre-eclampsia 529