noninvasive molecular tracking of colonizing wolf (canis lupus) packs in the western italian alps
TRANSCRIPT

Lucchini, V., Fabbri, E., Marucco, F., Ricci, S., Boitani, L., and Randi, E. (2002). Noninvasive molecular tracking of colonizing wolf (Canis lupus) packs in the western Italian Alps. Molecular Ecology 11: 857-868.
Keywords: 8IT/Alps/Canis lupus/genetics/individual identification/Malme/microsatellite/molecular sexing/mtDNA control region/non-invasive sampling/tracking
Abstract: We used noninvasive methods to obtain genetic and demographic data on the wolf packs (Canis lupus), which are now recolonizing the Alps, a century after their eradication. DNA samples, extracted from presumed wolf scats collected in the western Italian Alps (Piemonte), were genotyped to determine species and sex by sequencing parts of the mitochondrial DNA (mtDNA) control-region and ZFX/ZFY genes. Individual genotypes were identified by multilocus microsatellite analyses using a multiple tubes polymerase chain reaction (PCR). The performance of the laboratory protocols was affected by the age of samples. The quality of excremental DNA extracts was higher in samples freshly collected on snow in winter than in samples that were older or collected during summer. Preliminary mtDNA screening of all samples allowed species identification and was a good predictor of further PCR performances. Wolf, and not prey, DNA targets were preferentially amplified. Allelic dropout occurred more frequently than false alleles, but the probability of false homozygote determinations was always < 0.001. A panel of six to nine microsatellites would allow identification of individual wolf genotypes, also whether related, with a probability of identity of < 0.015. Genealogical relationships among individuals could be determined reliably if the number of candidate parents was 6-8, and most of them had been sampled and correctly genotyped. Genetic data indicate that colonizing Alpine wolves originate exclusively from the Italian source population and retain a high proportion of its genetic diversity. Spatial and temporal locations of individual genotypes, and kinship analyses, suggest that two distinct packs of closely related wolves, plus some unrelated individuals, ranged in the study areas. This is in agreement with field observations.

Molecular Ecology (2002) 11, 857–868
© 2002 Blackwell Science Ltd
Blackwell Science, LtdNoninvasive molecular tracking of colonizing wolf (Canis lupus) packs in the western Italian Alps
V. LUCCHINI ,* E . FABBRI ,* F . MARUCCO,† S . RICCI ,† L . BOITANI‡ and E . RANDI*Istituto Nazionale per la Fauna Selvatica (INFS), Ozzano Emilia (BO), Italy, †Progetto Lupo Interreg, Regione Piemonte, Parco Naturale Alpi Marittime, Valdieri (CN), Italy, ‡Dipartimento di Biologia Animale e dell’Uomo, Università ‘La Sapienza’, Roma, Italy
Abstract
We used noninvasive methods to obtain genetic and demographic data on the wolf packs(Canis lupus), which are now recolonizing the Alps, a century after their eradication. DNAsamples, extracted from presumed wolf scats collected in the western Italian Alps(Piemonte), were genotyped to determine species and sex by sequencing parts of themitochondrial DNA (mtDNA) control-region and ZFX/ZFY genes. Individual genotypeswere identified by multilocus microsatellite analyses using a multiple tubes polymerasechain reaction (PCR). The performance of the laboratory protocols was affected by the ageof samples. The quality of excremental DNA extracts was higher in samples freshlycollected on snow in winter than in samples that were older or collected during summer.Preliminary mtDNA screening of all samples allowed species identification and wasa good predictor of further PCR performances. Wolf, and not prey, DNA targets werepreferentially amplified. Allelic dropout occurred more frequently than false alleles, butthe probability of false homozygote determinations was always < 0.001. A panel ofsix to nine microsatellites would allow identification of individual wolf genotypes, alsowhether related, with a probability of identity of < 0.015. Genealogical relationshipsamong individuals could be determined reliably if the number of candidate parents was6–8, and most of them had been sampled and correctly genotyped. Genetic data indicatethat colonizing Alpine wolves originate exclusively from the Italian source populationand retain a high proportion of its genetic diversity. Spatial and temporal locations ofindividual genotypes, and kinship analyses, suggest that two distinct packs ofclosely related wolves, plus some unrelated individuals, ranged in the study areas. Thisis in agreement with field observations.
Keywords: Canis lupus, individual molecular tracking, microsatellites, molecular sexing, mtDNAcontrol-region, noninvasive genetics
Received 3 September 2001; revision received 21 January 2002; accepted 21 January 2002
Introduction
During the last two centuries wolf populations in Europehave declined strongly because of human persecution,deforestation and a decrease in their natural prey (Delibes1990). Wolves were eradicated from the Alps in the1920s and continued to decline in Italy until the 1970s,at which time ≈ 100 individuals survived in the centralApennines (Zimen & Boitani 1975). Legal protection,
accorded in 1971, and active conservation contributedto the prevention of further population decline. From the1980s onward wolves expanded along the Apennine ridgeand recolonized parts of their historical range (Boitani1992). Wolves crossed the north-western Apennines andreached the south-western Alps in 1992 (Breitenmoser1998; Corsi et al. 1999; Poulle et al. 1999).
The genetic and demographic structure of the colonizingwolf population is unknown. Alpine wolves presumablyoriginate from a natural expansion of the Italian sourcepopulation. However, local shepherds and huntersclaim that illegal releases of captive nonindigenous
Correspondence: Ettore Randi. Fax: 39 051 796 628; E-mail:[email protected]

858 V . L U C C H I N I E T A L .
© 2002 Blackwell Science Ltd, Molecular Ecology, 11, 857–868
wolves have contributed to the fast growth rate of the Alpinepopulation (G. Canavese, personal communication). Thecurrent Alpine wolf population is small and populationdynamics, interpack connectivity and dispersal rates ofwolf packs cannot be estimated using only field observations.
Wolf colonization of the Alps represents an opportunityto apply noninvasive genetic methods (Kohn & Wayne1997; Taberlet et al. 1997; Palsbøll 1999). DNA samplesextracted from scats allow researchers to identify and mapthe presence of individual genotypes, and estimate theirhome ranges and relatedness (Palsbøll et al. 1997; Reedet al. 1997; Kohn et al. 1999; Woods et al. 1999; Ernest et al.2000). However, the low amount and poor quality of targetDNA extracted from noninvasive samples might producecontamination, allelic dropout or scoring of false alleles(Taberlet et al. 1996; Gagneux et al. 1997; Goossens et al.1998; Miller et al. 2002). Therefore, the feasibility of anylarge-scale project should be assessed carefully throughpilot studies (Taberlet et al. 1999).
Here we report the results of a pilot study aimed toexplore the use of excremental DNA as a source of infor-mation for molecular tracking of local wolf packs. DNAsamples were genotyped by: (i) nucleotide sequencingof the hypervariable part of the mitochondrial DNAcontrol region (mtDNA CR), which is diagnostic for theItalian wolf population (Randi et al. 2000); (ii) analyses ofrestriction fragment length polymorphisms (RFLP) insex-linked canine ZFX/ZFY DNA sequences (Garcia-Muroet al. 1997); (iii) genotyping of six to nine microsatellite loci(Fredholm & Winterø 1995; Francisco et al. 1996; Randi &Lucchini 2002). Genetic data were used to: (i) define theorigin of colonizing Alpine wolves; (ii) estimate thenumber and relatedness of the different individual geno-types that were noninvasively sampled; (iii) map thespatiotemporal locations of the individual genotypes andwolf packs within the study areas, in comparison withobservations that are being collected during ongoing fieldresearch.
Materials and methods
Sample collection
We studied two wolf packs, respectively, located in twoareas separated by 40–50 km, in the Ligurian andMaritime Alps (Fig. 1). From May 1999 to August 2000 wecollected 338 scats in the Ligurian Alps and 164 in theMaritime Alps, by following wolf snow tracks in winterand by travelling systematic transects along humantrails/roads in summer. Scats were enclosed separately inclean plastic bags and stored at −18 °C until transferred totest tubes containing 95% ethanol. Before any furthermanipulation, all samples were deep-frozen at −80 °C for10 days to kill any Echinococcus eggs.
These samples were not analysed randomly, but wereselected on the basis of field observations to optimize thelaboratory efforts and characterize the individuals in thepacks. Wolves in the Ligurian Alps used an area of ≈ 300 km2
(Valle Pesio, Valle Ellero, Val Corsaglia and other valleys;Fig. 1). We analysed scats collected mainly from VallePesio, where a rendezvous site was found in summer 1999,and where winter snow tracks suggested the simultaneouspresence of several individuals. In summer 2000 wolfsigns were seen mainly in Val Corsaglia where anotherrendezvous site was found 18 km from the summer 1999site. Therefore, scats collected in Val Corsaglia from June toAugust 2000 were analysed to determine whether the samewolf pack used both areas and moved its rendezvous siteto a new location in summer 2000. Samples collected inthe Maritime Alps, from May 1999 to May 2000, wereselected to locate individual wolves in the core area (therendezvous site in Vallone del Piz) and in less frequentlyused peripheral areas (Vallone S. Anna and Vallone dellaValletta; Fig. 1).
Rates of DNA degradation were evaluated in samplescollected in ‘winter’ (November to April) and ‘summer’(May to October). Scats were ranked as ‘fresh’ (1−2 daysold when collected in summer, or 1−7 days old whencollected on snow in winter), or ‘old’ (all the othersamples), based on the time elapsed since the last sam-pling effort, scat appearance, exposure of deposition siteand weather conditions (Ciucci et al. 1997). DNA quality ispoor in aged scats, so samples older than two weeks werenot analysed, except for those collected in Val Corsagliaand Valle Pesio in summer 1999, which were analysedanyway.
DNA analyses
DNA was extracted from ≈ 80 mg of scat material using aguanidinium thiocianate and diatomaceus earth protocol(Gerloff et al. 1995). Extractions were carried out in a roomdedicated to excremental samples and all PCRs wereperformed under a laminar flow hood. Negative controls(no scat material added to the extractions) were used tocheck for contamination.
All DNA samples were initially screened by sequencingthe diagnostic part of the mtDNA CR, which wasamplified using primers l-Pro and H350 (Randi et al.2000), with addition of 1 µg of BSA (Boehringer) to the PCRreactions and 40 amplification cycles. Sequences wereobtained with the ABI (Foster City, USA) Dye-TerminatorKit and an ABI 373A automatic sequencer. ZFX/ZFYsequences (Garcia-Muro et al. 1997) were PCR-amplifiedin a volume of 25 µL using 10 µL of DNA solution, 2.5 µgof BSA and 40 PCR cycles. A sex-specific RFLP pattern wasdetected by overnight digestion of each PCR product with10 units of HaeIII, and by 4% agarose gel electrophoresis in

N O N I N V A S I V E M O L E C U L A R T R A C K I N G O F W O L V E S 859
© 2002 Blackwell Science Ltd, Molecular Ecology, 11, 857–868
TAE buffer. The ZFX product includes one HaeIIIrestriction site that produces two fragments of which onlythe larger (≈ 400 nucleotides) is visible on ethidium bro-mide-stained agarose gels. The HaeIII electrophoretic patternis expected to show two visible bands in males (an uncutpaternal ZFY fragment and a digested maternal ZFXfragment), but only one band in females (generated bydigestion of both ZFX fragments).
Primers used for ZFX/ZFY amplifications (P1-5EZ andP2-3EZ; Aasen & Medrano 1990) are conserved in verte-brates and might amplify DNAs from wolf prey. Thus, wesequenced a subset of the PCR products including allthe distinct individual multilocus genotypes. Thesesequences were aligned to homologous sequences frompotential prey including roe deer (Capreolus capreolus), sheep(Ovis aries), pig (Sus scrofa) and horse (Equus caballus).Relationships among sequences and species assignmentwere obtained by neighbour-joining analysis (NJ; Saitou &Nei 1987) with Kimura 2-parameters (Kimura 1980)
distance matrix and the software paup* Version 4.0b8(Swofford 1998).
Nine loci, including six dinucleotides (CPH2, CPH4,CPH5, CPH8, CPH12; Fredholm & Winterø 1995; andC09.250; Ostrander et al. 1993), and three tetranucleotides(FH2079, FH2088, FH2096; Francisco et al. 1996), were selectedamong 18 canine microsatellites previously used in astudy of Italian wolves (Randi & Lucchini 2002). We usedsix microsatellites (CPH2, CPH8, CPH12, FH2079, FH2088,FH2096) to identify the genotypes, and we added threeloci (CPH4, CPH5 and C09.250) to improve estimatesof kinship. Microsatellites were PCR-amplified (Randi &Lucchini 2002) separately in 10 µL of volume, using5 µL of DNA solution and 1 µg of BSA. Cycling conditionswere optimized for each primer and the number ofcycles varied from 40 to 45. These primers were alsoused to amplify DNAs from possible wolf prey. All micro-satellites were analysed using an ABI 373A automaticsequencer and the ABI software genotyper Version 2.1.
W1
W2W4
W5
W1
W2
W4W4
W4W4
W3
W3W3 W7
W3
W3 W2W7
W2 W1W2W1
W1W4
W2W4
W4
W5
W1W4
W4
W1W3
W2W4
W2
W2
W2W2
W1W1
W1 W1
W1
W3
W1W1
W1
W1W3
W3W6
VinadioAisone
France
V.ne S.
Ber
nolfo
V.ne S
. Ann
a
V.ne
del P
iz
Valle Stura
V.ne d
ella
Val
letta
W12
W12 W12
W13
W9
W9
W12
W14W13
W12W12
W11 W8W11
W12W12
W15
2 4 Km0
0 1.5 3 KmFrance
Val
Cor
sagl
ia
Val
le P
esio
a
b
Val
le E
ller
oV
.ne di
Rio
Fre
ddo
Maritime Alps
Ligurian Alps
Fig. 1 Map of the two study areas in western Italian Alps, showing the geographical locations of wolf individual genotypes (W) in (a) theMaritime Alps and (b) the Ligurian Alps. Shaded areas indicate the approximate distributions of wolves in Italy.

860 V . L U C C H I N I E T A L .
© 2002 Blackwell Science Ltd, Molecular Ecology, 11, 857–868
Data analyses
Protocols for microsatellites were designed to implementthe following multiple tubes approach (Taberlet et al. 1996;Gagneux et al. 1997): (i) all samples were amplified twice;(ii) those loci that were heterozygotes in both replicateswere scored as reliable and genotypes were recorded; (iii)all homozygote and uncertain genotypes (due to failure ofone amplification or to allelic dropout) were additionallyreplicated four times; (iv) all samples that could be notreliably typed at all loci after six amplifications werediscarded. The overall occurrence of PCR uncertainty wascomputed using data from the repetition of 40 scat samplesfor each of the six loci (Table 2). The probability P of falsehomozygotes was computed using the formula P = K(K/2)n−1 (Gagneux et al. 1997), where K is the observed frequencyof false homozygotes averaged over all samples and loci,and n is the number of repeated amplifications.
The probability of different individuals sharing, bychance, an identical genotype (probability of identity, PID;Mills et al. 2000; Waits et al. 2001), and the expected PIDamong full sib dyads (PIDsibs), were estimated in a set of100 Italian wolves using the software prob-id3 (G. Luikartunpublished). Field observations suggested that fewerthan 10 wolves were present in each of the two study areas,so we aimed to achieve PIDsibs values < 0.03, meaning that1 wolf in 30 siblings is expected to share, by chance, anidentical genotype with another wolf. Wolves in a pack areknown to be partially related, sharing alleles which areidentical by descent (Mech 1970; Lehman et al. 1992; Wayneet al. 1995). Moreover, the colonizing population might bea nonrandom sample of the source Italian wolf populationand different genotypes would differ in matching probabilitydue to different allelic frequencies. Therefore, we computedindividual match probabilities (Woods et al. 1999) for randomand sibling individuals. Allelic frequencies were estimatedfrom the Italian wolf population (Randi & Lucchini 2002).Total match probabilities for each multilocus genotype werecomputed by multiplying single locus probabilities, assumingthat loci are independent, as suggested by the microsatellitelinkage map of the domestic dog (Neff et al. 1999).
Values of relatedness (r; Queller & Goodnight 1989)among genotypes were inferred using nine microsatellites,which were typed in a subset of DNA samples including allthe different genotypes identified in the Alpine wolves,using the software kinship Version 1.2 (Goodnight &Queller 1999). kinship allows estimation of the likelihoodand testing of the likelihood ratios of primary/null hypo-thetical pedigree relationships. In this study we tested aprimary hypothesis of first-order relationship (i.e. dyadsrepresent full sibs or parental-offspring with expectedrelatedness r = 0.5) vs. a null hypothesis of no relationship(r = 0.0). When likelihood ratios are significantly greaterthan 1, we can reject the null hypothesis and avoid a type II
error (that siblings are incorrectly classified as unrelated) ata given level of significance. Equivalent microsatellite datafrom 100 Italian wolves were used to generate the expecteddistributions of pairwise relatedness among parents plusfull sibs (theoretical average r = 0.5) and among unrelatedindividuals (theoretical average r = 0.0).
The proportion of parentages that can be theoreticallyidentified with 80 or 95% confidence, given a set of speci-fied parameters (number and proportion of typed loci,proportion of mistyped loci, number of candidate parentsand proportion of candidate parents sampled), and usingallele frequencies estimated in the source Italian wolfpopulation, was simulated using cervus Version 1.0(Marshall et al. 1998). Hypothetical full sib or parental−offspring dyads were re-examined using the likelihoodapproach implemented in cervus, which computes a log-likelihood of parentage (LOD score) for each candidateparent. cervus was also used to estimate the parametersof genetic diversity (number of alleles, expected HE andobserved HO heterozygosity), and values of the poly-morphic information content (PIC; Hearne et al. 1992) inthe colonizing and source Italian wolf populations.
Departures from Hardy–Weinberg equilibrium (HWE)were tested using the probability test by Guo & Thompson(1992) as implemented in genepop Version 3.2 (Raymond& Rousset 1995) and an estimate of FIS (Weir & Cockerham1984). The origin of the Alpine wolves was inferred by anassignment test of individuals to populations, using aBayesian model implemented in the software structure(Pritchard et al. 2000), and a set of 100 randomly sampleddogs, 100 Italian wolves and 170 wolves from Bulgaria,Croatia, Greece, Turkey, Israel, Latvia, Finland and Spain(unpublished).
Results
Age of the samples and performance of the laboratory protocols
We analysed 130 DNA samples extracted from 104 scatscollected in the Ligurian Alps and 26 collected in theMaritime Alps. We obtained reliable mtDNA sequences in84% of the cases, whereas 64 and 53% of the samples weresuccessfully sexed and genotyped at all six microsatellitesusing the multitubes approach (Table 1). Thus, inde-pendent of the putative age and sampling period, themtDNA CR amplified more frequently than nuclear genes(P < 0.0002; χ2 test). DNA samples extracted from scatscollected in winter performed significantly better thanthose collected in summer (P < 0.0001; χ2 test). The mtDNACR and microsatellites amplified better in DNAs extractedfrom fresh than from old samples (P < 0.0001; χ2 test). Incontrast, amplification of the ZFX/ZFY sequences, whichwere used for sex determination, was not affected by theperiod of sampling or the age of the samples.
N O N I N V A S I V E M O L E C U L A R T R A C K I N G O F W O L V E S 861
© 2002 Blackwell Science Ltd, Molecular Ecology, 11, 857–868
Rates of allelic dropout were different among loci,occurring in the 18% of the heterozygote determinations,on average (Table 2). The probability of false homozygotedeterminations after four positive PCR repetitions wasP < 0.001. Contamination (i.e. the presence of more thantwo alleles in a sample) or false alleles (i.e. alleles fallingoutside the molecular mass range observed at each locus inthe Italian wolves; Randi & Lucchini 2002; see Table 1),were detected in only five amplifications from differentsamples over > 1500 PCRs performed in this study. Theseartefacts did not significantly affect the results.
The microsatellites used in this study did not amplifyDNAs from ungulates, showing that prey DNAs did notaffect genotype determinations. These primers amplifyhomologous loci from dogs and foxes (Vulpes vulpes), which,respectively, showed different allele frequencies or dif-ferent allele sizes (Randi & Lucchini 2002). Thus, theassessment of allele size differences or an assignment test(see below) can be used to identify dog and fox samples.
Mitochondrial sequences and species identification
We obtained 109 mtDNA CR sequences from 130 analysedscats, which were identical to the Italian wolf haplotypeW14 (Randi et al. 2000). Two sequences were identified asa dog and fox, respectively. The dog haplotype joined thedog cluster C1 (see Randi et al. 2000), and the domesticorigin of this sample was confirmed by the assignment test
of its microsatellite genotype. The identification of the foxsample was not obtained by the CR, which was difficult tosequence, but by sequencing part of the mitochondrialcytochrome b gene, and by comparing this sequences withDNA samples extracted from a set of fox tissues (not shown).
Microsatellite variability and genotype identification
Among the 69 scat samples which were successfullygenotyped using six loci, we identified 14 distinctgenotypes, which presumably correspond to at least 14different individuals. In Valle Pesio we identified fourmales (W1, W2, W5 and W6) and three females (W3, W4and W7). In Val Corsaglia we identified two genotypes,identical (also in sex) to wolves W1 and W3 previouslysampled in Valle Pesio. Wolves W1 and W3, sampled nearthe rendezvous site in 1999, were also sampled near therendezvous site in 2000 (Fig. 1). This result supports fieldobservations and suggests that the same wolf pack rangedin Valle Pesio and Val Corsaglia in 1999–2000, and movedrendezvous site during summer 2000. In the Maritime Alpswe identified seven new genotypes (W8, W9, W11, W12,W13, W14 and W15; Fig. 1), sampled from May 1999 toApril 2000, which were different from those found in theLigurian Alps (see Fig. 5). All the genotypes were assignedto the Italian wolf population using structure, with aprobability q > 0.98, and had no significant ancestry in thedog gene pool or in other wolf populations.
Table 2 Rates of allelic dropout observed using replicated PCRs of six microsatellite loci in 40 excremental DNA samples collected fromcolonizing wolves in the western Italian Alps. The number of heterozygotes was estimated after a total of 480 PCRs; the number of correctdeterminations refers to the number of heterozygotes which were correctly determined after the first two PCRs
Microsatellite loci CPH2 CPH8 CPH12 FH2079 FH2088 FH2096 Average
Heterozygotes 30 28 9 26 33 11 137Correct determinations 29 20 6 18 28 11 112Allelic dropout 1 8 3 8 5 0 25Percentage error 3 29 33 31 15 0 18
Table 1 Relative performances of the laboratory protocols used to PCR-amplify and analyse mtDNA CR sequences, sex-linked ZFX/ZFYRFLPs and a panel of six canine microsatellites (the expected lengths of the PCR products are indicated in parentheses), in excremental DNAsamples collected from colonizing wolves in western Italian Alps (Fig. 1). Criteria for ranking the period of sampling and the age of samplesare described in Materials and methods. Percentages are in parentheses
Samples mtDNA (404 bp)
ZFX/ZFY (430 bp)
Microsatellites (92–295 bp)
Period of sampling Winter 54 49 (91) 34 (63) 35 (65)Summer 76 60 (79) 49 (64) 34 (45)
Age of samples Fresh 97 89 (92) 61 (63) 62 (64)Old 33 20 (61) 22 (67) 7 (21)
Total 130 109 (84) 83 (64) 69 (53)
862 V . L U C C H I N I E T A L .
© 2002 Blackwell Science Ltd, Molecular Ecology, 11, 857–868
Recapture rates of genotypes varied in each study area.Wolves W1, W2, W3 and W4 were identified from 10 ormore scats each; W5, W6 and W7 were identified from oneor two scats. Wolf W12 was sampled eight times in theMaritime Alps, the other genotypes one or two times.These genotypes were not sampled during the same sea-son. The highest proportion of new genotypes wassampled in winter. In the Ligurian Alps, the most frequentgenotypes (W1, W2, W3 and W4) occurred almost regu-larly from summer 1999 to spring 2000, except for W4which was sampled regularly in Valle Pesio only from theend of July to December 1999. The other genotypes (W5,W6 and W7) were documented only in winter 2000. Onlywolves W1 and W3 were resampled in summer 2000. In theMaritime Alps we found three different genotypes beforewinter (W9, W12, W13). All the other genotypes werefound in January and February 2000.
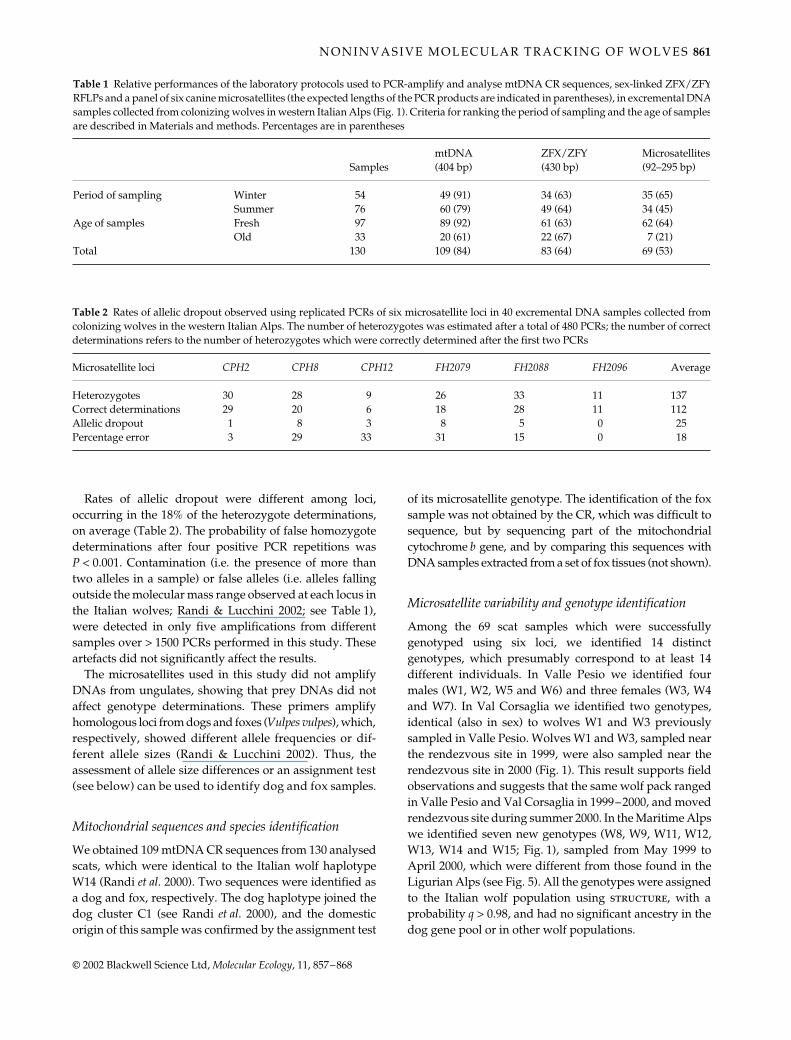
The observed PID was zero, computed using up to fiveloci in a sample of 100 Italian wolves (Fig. 2). The theoreti-cal PID for siblings was 1.50 × 10−2 with six loci, suggestingthat the selected microsatellites should allow detection ofunique genotypes also if related individuals were sampled.Allele frequencies at each locus vary in the Italian wolfpopulation (Randi & Lucchini 2002), and some genotypesare expected to be more frequent than others. However,
match probabilities were usually < 0.025, which means thatonly 1 in 40 sibling individuals would share an identicalgenotype with another wolf by chance.
All microsatellites were polymorphic in the colonizingwolf population, showing 2–4 alleles (average = 2.67 ±0.50). These loci were more variable in the Italian wolfpopulation (3–7 alleles; average = 4.50 ± 1.38). Averagevalues of gene diversity in the colonizing wolves (HO = 0.524,HE = 0.517 and PIC 0.414) were similar to the correspond-ing values in the source population (HO = 0.650, HE = 0.598,and PIC = 0.542), suggesting that the colonizing popula-tion retains a high proportion of the genetic diversity of thesource population. The colonizing population was in HWE(the probability to obtain by chance a value of Fis greaterthan the observed was P = 0.844), whereas the Italianpopulation was in disequilibrium (P = 0.031), likely due togeographical substructuring along the Apennines (see alsoRandi & Lucchini 2002).
Sex-linked sequences and molecular sexing of the colonizing wolves
DNA samples were sexed using diagnostic ZFX/ZFY RFLPs.We validated the sex-specific pattern in 20 sexed Italianwolves. Results of the molecular sexing procedure were100% concordant with a priori known nonmolecular sexing.Using this procedure we identified 45 males and 41 femalesamong the DNA samples collected in the Alpine studyareas. However, these data should not be used to estimatethe sex ratio of the colonizing wolf population, because thesampling probabilities of the different genotypes are veryskewed. Among the 14 different individual microsatellitegenotypes we identified 7 females and 7 males (sex ratioof 1:1).
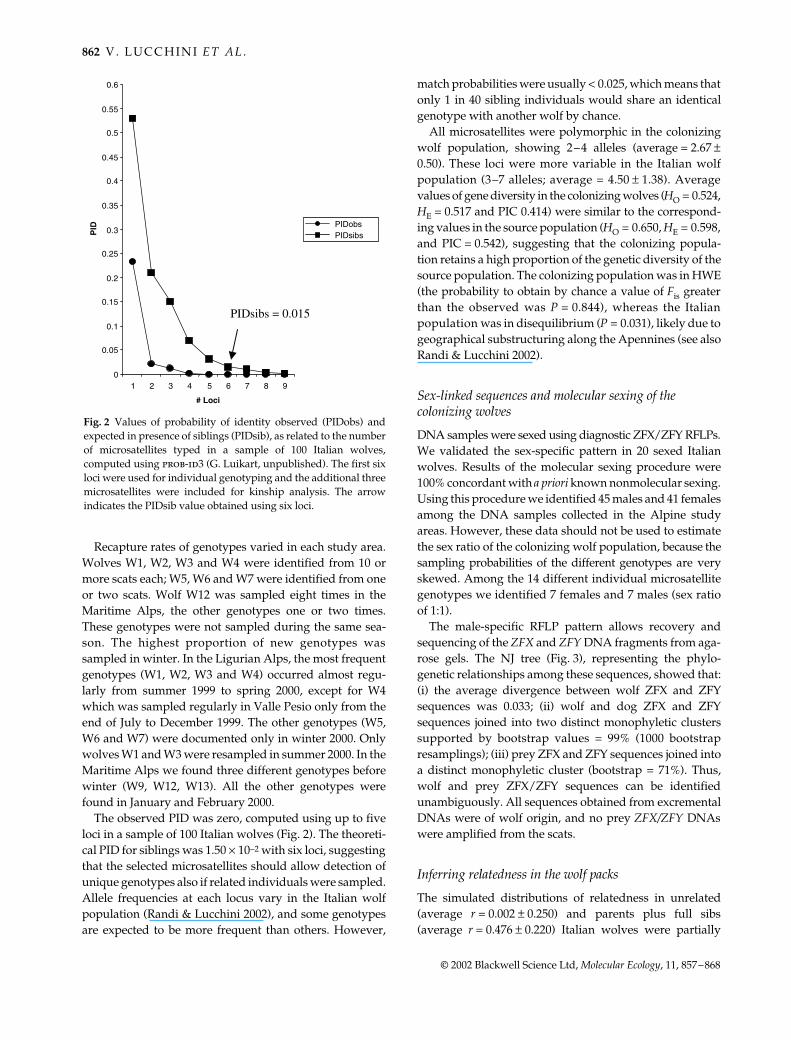
The male-specific RFLP pattern allows recovery andsequencing of the ZFX and ZFY DNA fragments from aga-rose gels. The NJ tree (Fig. 3), representing the phylo-genetic relationships among these sequences, showed that:(i) the average divergence between wolf ZFX and ZFYsequences was 0.033; (ii) wolf and dog ZFX and ZFYsequences joined into two distinct monophyletic clusterssupported by bootstrap values = 99% (1000 bootstrapresamplings); (iii) prey ZFX and ZFY sequences joined intoa distinct monophyletic cluster (bootstrap = 71%). Thus,wolf and prey ZFX/ZFY sequences can be identifiedunambiguously. All sequences obtained from excrementalDNAs were of wolf origin, and no prey ZFX/ZFY DNAswere amplified from the scats.
Inferring relatedness in the wolf packs
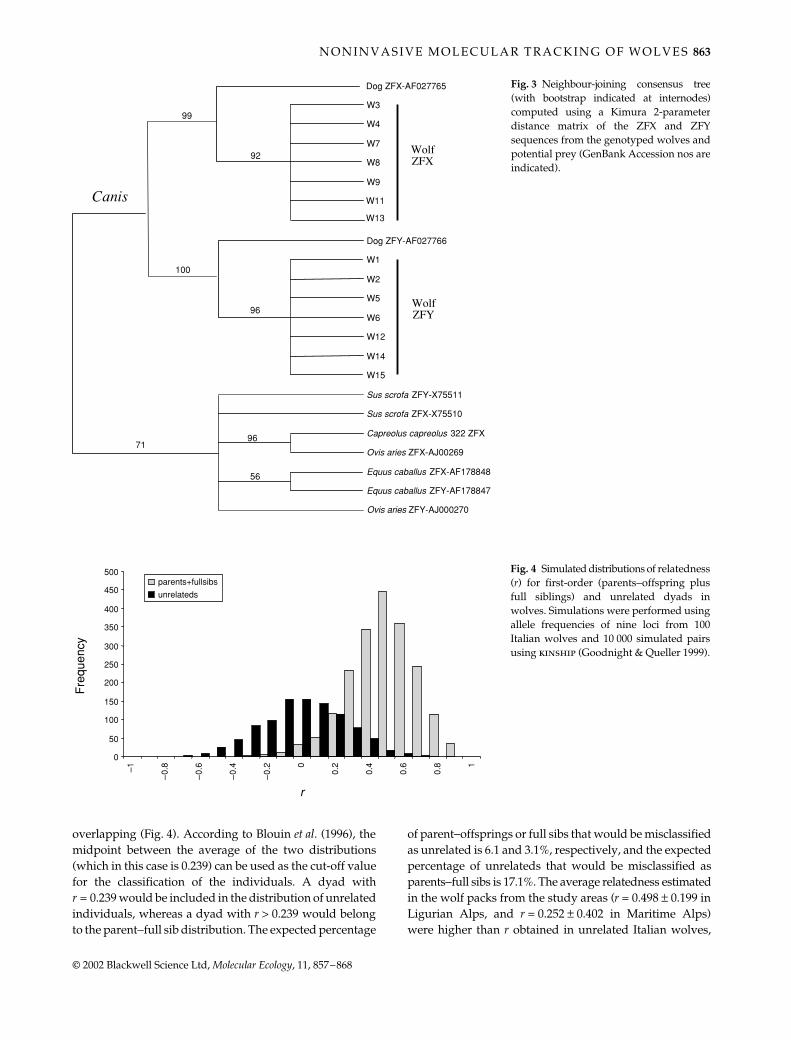
The simulated distributions of relatedness in unrelated(average r = 0.002 ± 0.250) and parents plus full sibs(average r = 0.476 ± 0.220) Italian wolves were partially
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
0.5
0.55
0.6
1 2 3 4 5 6 7 8 9
# Loci
PID PIDobs
PIDsibs
PIDsibs = 0.015
Fig. 2 Values of probability of identity observed (PIDobs) andexpected in presence of siblings (PIDsib), as related to the numberof microsatellites typed in a sample of 100 Italian wolves,computed using prob-id3 (G. Luikart, unpublished). The first sixloci were used for individual genotyping and the additional threemicrosatellites were included for kinship analysis. The arrowindicates the PIDsib value obtained using six loci.
N O N I N V A S I V E M O L E C U L A R T R A C K I N G O F W O L V E S 863
© 2002 Blackwell Science Ltd, Molecular Ecology, 11, 857–868
overlapping (Fig. 4). According to Blouin et al. (1996), themidpoint between the average of the two distributions(which in this case is 0.239) can be used as the cut-off valuefor the classification of the individuals. A dyad withr = 0.239 would be included in the distribution of unrelatedindividuals, whereas a dyad with r > 0.239 would belongto the parent−full sib distribution. The expected percentage
of parent−offsprings or full sibs that would be misclassifiedas unrelated is 6.1 and 3.1%, respectively, and the expectedpercentage of unrelateds that would be misclassified asparents−full sibs is 17.1%. The average relatedness estimatedin the wolf packs from the study areas (r = 0.498 ± 0.199 inLigurian Alps, and r = 0.252 ± 0.402 in Maritime Alps)were higher than r obtained in unrelated Italian wolves,
WolfZFX
WolfZFY
Canis
Dog ZFX-AF027765
W3
W4
W7
W8
W9
W11
W13
Dog ZFY-AF027766
W1
W2
W5
W6
W12
W14
W15
Sus scrofa ZFY-X75511
Sus scrofa ZFX-X75510
Capreolus capreolus 322 ZFX
Ovis aries ZFX-AJ00269
Equus caballus ZFX-AF178848
Equus caballus ZFY-AF178847
Ovis aries ZFY-AJ000270
71
99
92
100
96
96
56
0
50
100
–1
–0.
8
–0.
6
–0.
4
–0.
2 0
0.2
0.4
0.6
0.8 1
150
200
250
300
350
400
450
500parents+fullsibs
unrelateds
Fre
quen
cy
r
Fig. 3 Neighbour-joining consensus tree(with bootstrap indicated at internodes)computed using a Kimura 2-parameterdistance matrix of the ZFX and ZFYsequences from the genotyped wolves andpotential prey (GenBank Accession nos areindicated).
Fig. 4 Simulated distributions of relatedness(r) for first-order (parents−offspring plusfull siblings) and unrelated dyads inwolves. Simulations were performed usingallele frequencies of nine loci from 100Italian wolves and 10 000 simulated pairsusing kinship (Goodnight & Queller 1999).

864 V . L U C C H I N I E T A L .
© 2002 Blackwell Science Ltd, Molecular Ecology, 11, 857–868
showing that colonizing wolf packs likely include mainlyrelated individuals.
Using cervus we simulated the proportion of parentagetests that can be resolved with 80 and 95% confidence usingnine loci, an error rate of 1%, from 2 to 10 candidate par-ents, 100 or 75% candidates sampled, and one or neitherparent known. Results (Table 3) suggested that the typedmicrosatellites would allow identification of all the trueparents from a set of 2–10 candidates if one parent was pre-viously known and if 100% of the candidates were sam-pled. If putative parents are not known, identification ofthe true parents is still highly probable with 2–6 candidates
when all candidates are sampled, whereas it decreaseswhen only 75% of the candidates are sampled. Theseresults suggest that nine microsatellites could produce reli-able parentage assignment in the studied wolf populationsif putative parents were fewer than 6–8, and if most of themwere sampled and genotyped.
A upgma clustering of interindividual relatednessshowed that the wolf genotypes form two separate groups(Fig. 5): group A, including wolves W11, W12, W13 andW14 sampled in the Maritime Alps; group B, includingwolves from the Ligurian Alps and W15 and W8 from theMaritime Alps. Wolves W7 (Ligurian Alps) and W9(Maritime Alps) were not closely related to any otherAlpine wolf. Groups A and B correspond in part to thepacks mapped in the two study areas (Fig. 1).
Pedigree analyses were performed to test the primaryhypothesis that dyads include full siblings (expectedr = 0.5) vs. a null hypothesis of unrelatedness (r = 0.0). Inthe Ligurian Alps we identified highly significant relation-ships (P < 0.001) between wolves W1 and W2, and betweenW4 and W5. W1 was also related to W3, W4 and W5(P < 0.01); W3 was also related to W6 (P < 0.01; Fig. 6).Closely related wolves W11, W12, W14 and W13 (P < 0.01–0.001), plus three unrelated wolves (W8, W9 and W15)were identified in the Maritime Alps. Wolves from the twostudy areas were not significantly related to each otherexcept for W15 and W13 (sampled in the Maritime Alps),which were, respectively, related (P < 0.01) to W4−W5 andW3 (sampled in the Ligurian Alps). These data suggest thatgenotypes identified in the study areas should belong totwo distinct packs.
LOD scores generated by cervus were used to identifythe most likely parents in the two packs (Fig. 6). WolvesW1 and W3, sampled at least 10 times from June−July 1999to spring 2000 in the Ligurian Alps, were the likely parentsof W4, sampled 11 times from July to December 1999, andW5 and W6, sampled only in December 1999 and January2000. Wolf W2, sampled 12 times from June 1999 to spring2000, and W1 were significantly related, but they were not
Fig. 5 upgma clustering of pairwise relatedness (1–r) among theAlpine wolf genotypes and 24 unrelated Italian wolves sampled inthe Apennines.
Candidates parents
Candidates sampled
One parent known Neither parent known
80% conf. 95% conf. 80% conf. 95% conf.
2 100 100 100 98 984 100 100 100 99 876 100 100 100 99 768 100 100 99 99 62
10 100 100 97 100 524 75 80 74 82 536 75 84 71 79 388 75 86 67 71 31
10 75 88 60 63 25
Table 3 Proportion of simulated parentagetests (10 000 simulations) resolved with80 and 95% confidence estimated usingcervus (Marshall et al. 1998). Allelic fre-quencies at nine microsatellite loci weredetermined in a sample of 100 Italianwolves. Simulations assumed that 100% ofthe loci were typed for each individual andthe typing error rate was 1%

N O N I N V A S I V E M O L E C U L A R T R A C K I N G O F W O L V E S 865
© 2002 Blackwell Science Ltd, Molecular Ecology, 11, 857–868
identified as a likely parent–offspring dyad, and could bebrothers.
Field observations suggested the presence of at leastthree wolves in the Maritime Alps, and in summer 1999 acore area was identified in Vallone del Piz where puppieswere heard howling. Among the seven genotypes identi-fied in the Maritime Alps, only W11, W12, W13 and W14were significantly related. LOD scores suggested that maleW12 (sampled eight times from July 1999 to March 2000)and female W13 (sampled two times, in September 1999and January 2000), were the likely parents of W11, that wassampled only in January 2000, but not of W14. This wolfcould be a brother of W12, as they appear to be closelyrelated (Fig. 6).
Discussion
The recolonization of the Alps is of paramount importancefor the conservation of the Italian wolf population, whichthrough the Alps could reconnect to other populations,interrupting its long-lasting genetic isolation. Because oftheir elusive nature, direct monitoring of wolf packs in theAlps is problematic. Thus, the use of noninvasive mole-cular methods can add relevant information. However, thequality and power of noninvasive DNA data should becarefully evaluated to avoid pitfalls and optimize samplingprocedures.
Preliminary mtDNA screening allows discarding ofDNA samples that are too degraded for molecular sexingand microsatellite genotyping. Mitochondrial genes maybe typed more easily than nuclear genes, because themtDNA genome is present in multiple copies in cells andcellular extracts. Thus, diluted and degraded DNAs fromexcremental samples might limit the amplification of single-copy genes more than the highly repetitive mitochondrialgenes (Kohn & Wayne 1997; Frantzen et al. 1998).
Species identification by mtDNA typing should be per-formed first, to avoid further analyses of wrong samples(e.g. dogs or foxes). DNA samples that did not amplify formtDNA, also did not amplify for sex-linked or micro-satellite loci. All the samples that were successfully geno-typed with microsatellites also produced clear mtDNAsequences, whereas only 85% of them was successfullysexed. The ZFX/ZFY fragment is longer than microsatel-lites (≈ 430 nucleotides, whereas microsatellites range from92 to 295 nucleotides), and is probably more sensitive toDNA degradation. We suggest targeting shorter fragmentsfor sex determination, with a length comparable with thatof microsatellites. In this study allelic dropout producedinaccurate genotyping in 18% of the PCR experiments,whereas amplification of false alleles was negligible. Weobserved a large per locus variance in dropout rates (Table 2),indicating that some loci are amplified less efficientlythan others using excremental DNA samples. Inefficient
W4
W5
W1
W7
W3
W6
W2
W15
W12
W14
W8W9
W13
W11
Ligurian Alps
Maritime Alps
Fig. 6 Inferred relatedness among wolf microsatellite genotypes identified in the Alpine study areas. Circles represent females and squaresrepresent males. First-order relatives are connected by bold lines (P < 0.001), or double lines (P < 0.01). Arrows indicate the offspring of adeterminate parent as identified by cervus (Marshall et al. 1998).

866 V . L U C C H I N I E T A L .
© 2002 Blackwell Science Ltd, Molecular Ecology, 11, 857–868
loci could be replaced, in future studies, with other loci,which should be equally informative, but less prone toallelic dropout. These findings support the use of the mul-tiple tubes approach (Taberlet et al. 1996) to achieve theprecision required for determining the individual geno-types. However, efficient multiple tubes strategies shouldbe designed using a precise statistical framework, tailoredto detect the most efficient loci and to direct the replicationstowards the problematic ones (Miller et al. 2002).
The presumed ‘age’ of samples and sampling ‘season’significantly affected the performances of DNA analyses.Samples collected in winter performed generally betterthan samples collected in summer. However, obtainingnoninvasive multilocus genotypes is still problematic andlabour intensive. We suggest that field sampling strategiesshould be carefully planned in advance. In an Alpine hab-itat, excremental sampling in winter through snow track-ing might be optimal. Frequent controls of transects insummer, at least once a week, would also improve thequality of laboratory results.
All sequenced DNAs showed the exclusive presence ofthe Italian wolf haplotype W14 (Randi et al. 2000). Theassignment test also showed that all individual microsatel-lite genotypes identified in the Alps belong to the Italianwolf population. Thus, both mtDNA and nuclear markerssuggest that the ongoing recolonization of the Alps is dueto the natural expansion of the Italian wolf population.Occasional cross-breeding of Italian wolves with free-ranging dogs has been feared and documented in Italy(Boitani 1983; Randi & Lucchini 2002). Although the pres-ence of dogs has been directly observed or documented byexcremental DNA analyses, we did not identify any puta-tive case of admixed wolf−dog genotype in the Alpinestudy areas. Moreover, the alleged presence of illegallyreleased nonindigenous wolves was not documented bythis data set. However, the genetic composition of theexpanding Alpine wolf population should be monitoredcontinuously, and eventual cross-breeding with dogs orpresence of nonindigenous wolves should be promptlyassessed.
Prey or contaminant DNA did not affect our laboratoryprotocols, thus molecular sexing, species and populationassignment were reliable. The identification of a dog and afox DNA using both mtDNA sequences and microsatel-lite alleles suggests that these two samples were errone-ously identified as wolf scats. Alternatively, it is possiblethat wolf scats were marked by dog or fox urine. How-ever, in this case, we suspect that true wolf mtDNAand/or microsatellite alleles would have been amplifiedand typed concomitantly with the contaminant dog andfox DNAs, leading to anomalous sequence or allelicpatterns.
Values of genetic diversity in Alpine and Italian wolveswere similar, suggesting that the colonizing population
retained a relevant portion of the genetic variability of thesource population (see also Forbes & Boyd 1996; whichobtained similar results in a study of colonizing wolves inMontana). The smaller number of alleles in colonizing wolvescould have been inflated by sample size differences, thatis 100 Italian wolves vs. the 14 different genotypes thatwere identified in the study areas. However, the averagerelatedness in colonizing wolf packs was higher thanamong a sample of Italian wolves, indicating that coloniz-ing wolf packs likely include mainly related individuals.
Six microsatellites were sufficient to achieve reliableidentification of individual genotypes in the Alpinewolves, with PIDsibs < 0.015. We identified 14 distinct gen-otypes, with a sex ratio of 1:1. Individual genotypesshowed values of match probability < 0.025, which is lowenough to avoid randomly typing two identical genotypesin a sample of 40 siblings. Recapture rates were variable: afew genotypes were collected several times, whereas oth-ers were collected only once or twice. Genotypes sampledonly once were mainly from scats collected in winter alongwolf snow tracks. Following a pack travel route in thesnow it is possible to collect scats of each individual asidefrom the individual marking behaviour. Instead, scatscollected in summer along human trails/roads, were likelysamples from dominant individuals that frequently markthe territory. These findings are consistent with wolf mark-ing behaviour which affects the patterns of defecation ontrails (Peters & Mech 1975; Vilà et al. 1994; but see Kohnet al. 1999 in the coyote).
Wolves sampled within each area were more related toeach other than wolves between the two study areas, andwere subdivided into two distinct groups of relatedness(Fig. 5), mostly concordant with sampling locations (Fig. 1)and with the inferred patterns of kinship (Fig. 6). Wolves ingroup A likely constitute a single pack including the twoparents (male W12 and female W13) one offspring (femaleW11) and a relative (male W14). These results are concord-ant with field data: wolves W12 and W13 were sampledfrom July−September 1999 to winter 2000, and could bethe candidates parents. Group B includes mainly wolvesfrom the Ligurian Alps, but also wolves W15 and W8 fromthe Maritime Alps. Wolves W7 (Ligurian Alps) and W9(Maritime Alps) were not closely related to any other wolf.Wolves W1, W2 and W3 were sampled from June 1999 andtheir movements in the study area (Fig. 1) suggested thatthey were adults and likely candidate parents. Wolves W1and W3 were sampled together near the rendezvous site inVal Corsaglia (Fig. 1). Wolves W4 and W5 were sampledaround the rendezvous site in 1999 (Fig. 1) and their smallscat sizes suggest that they were pups. Thus, the resultsof kinship analyses are mostly concordant with fieldobservations.
The genealogical relationships are preliminary and needto be reassessed when additional data are available. The

N O N I N V A S I V E M O L E C U L A R T R A C K I N G O F W O L V E S 867
© 2002 Blackwell Science Ltd, Molecular Ecology, 11, 857–868
microsatellite data generated in this study might allow fordetailed pedigree inference only if the number of candidateparents in the study area is small (fewer than 6–8), and allor most of them are sampled. The number of parents−off-spring in a study area can be inferred approximately byfield observations (snow tracks and wolf howling). How-ever, the reliability of field data would be uncertain due tothe unknown structure of packs, turnover rates and immi-gration of dispersing wolves into the study areas. Underthese conditions, inferring kinship from genetic data mightbe problematic, mainly because of the unknown propor-tion of sampled and genotyped candidate parents (Mar-shall et al. 1998).
The typed microsatellites showed fewer alleles in thecolonizing wolves than in the Italian wolf population.Moreover, the average relatedness in the Alpine wolfpacks was higher than the average relatedness amongunrelated Italian wolves thus showing that colonizingwolves likely include related individuals. The simu-lated distributions of relatedness among Italian wolvessuggested 17.1% of unrelateds could be misclassified asparents−full sibs. Low allelic diversity and high related-ness might reduce the likelihood to identify the pedi-gree relationships in packs, also because it is usuallydifficult to discriminate parent−offspring from full sibsdyads (Marshall et al. 1998). These results suggest thatincreasing the number of loci or including loci withmore alleles in the colonizing population would bebeneficial.
Conclusions
The information obtained from these analyses should beconsidered preliminary and needs to be supported byadditional genetic and field data. However, it is apparentthat noninvasive genetic sampling methods can addressissues that could not be addressed in any other way. Thescreening of a limited number of genetic markers producesinformation which can reliably be used to identify species,origin of local populations, identity of individuals, areasand movement of packs, as well as to hypothesize detailedgenealogical relationships and describe the fine structureof wolf packs. An important prediction of this study is thata limited improvement of field sampling procedures,which should focus on collecting fresh scats in winter alongwolf snow tracks, could allow significant improvement inthe quality of the laboratory data. However, a variety offield sampling schemes could be defined accordingly to thekind of genetic data which one aims to obtain. Intensivesample collection, randomized across the entire wolf packrange and concentrated in short period (a few months),before and after the reproductive period, should beobtained to use noninvasive genetic data for populationsize estimations.
Acknowledgements
This study would not have been possible without the help of col-leagues who collected and kindly provided wolf samples, and inparticular Tommaso Galli and the rangers of Valle Pesio and AlpiMarittime Natural Parks, Alessandra Tropini and Lorenzo Manghi.We thank two anonymous referees for constructive comments andfor helping with manuscript revision and editing. This study wassupported by the Interreg Project and INFS.
References
Aasen E, Medrano JF (1990) Amplification of the ZFY and ZFXgenes for sex identification in humans, cattle, sheep and goat.Bio /Technology, 8, 1279–1281.
Blouin MS, Parson M, Lacaille V, Lotz S (1996) Use of microsat-ellite loci to classify individuals by relatedness. Molecular Ecol-ogy, 5, 393–401.
Boitani L (1983) Wolf and dog competition in Italy. Acta ZoologicaFennica, 174, 259–264.
Boitani L (1992) Wolf research and conservation in Italy. BiologicalConservation, 61, 125–132.
Breitenmoser U (1998) Large predators in the Alps: the fall and riseof man’s competitors. Biological Conservation, 83, 279–289.
Ciucci P, Boitani L, Francisci F, Andreoli G (1997) Home range,activity and movements of a wolf pack in central Italy. Journal ofZoology (London), 243, 803–819.
Corsi F, Duprè E, Boitani L (1999) A large-scale model of wolfdistribution in Italy for conservation planning. ConservationBiology, 13, 150–159.
Delibes M (1990) Status and conservation needs of the wolf in theCouncil of Europe member states. Nature and Environment Series,Strasbourg, 47, 1–46.
Ernest HB, Penedo MCT, May BP, Syvanen M, Boyce WM (2000)Molecular tracking of mountain lions in the Yosemite Valleyregion in California: genetic analysis using microsatellites andfaecal DNA. Molecular Ecology, 9, 433–441.
Forbes SH, Boyd DK (1996) Genetic variation of naturally colonizingwolves in the central Rocky Mountains. Conservation Biology, 10,1082–1090.
Francisco LV, Langston AA, Mellersh CS, Neal CL, OstranderEA (1996) A class of highly polymorphic tetranucleotiderepeats for canine genetic mapping. Mammalian Genome, 7,359–362.
Frantzen MAJ, Silk JB, Ferguson JWH, Wayne RK, Kohn MH(1998) Empirical evaluation of preservation methods for faecalDNA. Molecular Ecology, 7, 1423–1428.
Fredholm M, Winterø AK (1995) Variation of short tandem repeatswithin and between species belonging to the Canidae family.Mammalian Genome, 6, 11–18.
Gagneux P, Boesch C, Woodruff DS (1997) Microsatellitescoring errors associated with noninvasive genotyping onnuclear DNA amplified from shed hair. Molecular Ecology, 6,861–868.
Garcia-Muro E, Aznar MP, Rodellar C, Zaragoza P (1997) Sex-specific PCR/RFLPs in the canine ZFX/ZFY loci. AnimalGenetics, 28, 156.
Gerloff U, Schlotterer C, Rassmann K et al. (1995) Amplification ofhypervariable simple sequence repeats (microsatellites) fromexcremental DNA of wild living Bonobos (Pan paniscus).Molecular Ecology, 4, 515–518.

868 V . L U C C H I N I E T A L .
© 2002 Blackwell Science Ltd, Molecular Ecology, 11, 857–868
Goodnight KF, Queller DC (1999) Computer software forperforming likelihood tests of pedigree relationship usinggenetic markers. Molecular Ecology, 8, 1231–1234.
Goossens B, Waits LP, Taberlet P (1998) Plucked hair samples as asource of DNA: reliability of dinucleotide microsatellite geno-typing. Molecular Ecology, 7, 1237–1241.
Guo SW, Thompson EA (1992) Performing the exact test ofHardy–Weinberg proportion for multiple alleles. Biometrics, 48,361–372.
Hearne CM, Ghosh S, Todd JA (1992) Microsatellite for linkageanalysis of genetic traits. Trends in Genetics, 8, 288–294.
Kimura M (1980) A simple method for estimating evolutionaryrate of base substitutions through comparative studies ofnucleotides sequences. Journal of Molecular Evolution, 16,111–120.
Kohn MH, Wayne RK (1997) Facts from feces revisited. Trends inEcology and Evolution, 12, 223–227.
Kohn MH, York EC, Kamradt DA, Haught G, Sauvajot RM,Wayne RK (1999) Estimating population size by genotypingfaeces. Proceedings of the Royal Society of London, Series B, 266,657–663.
Lehman N, Clarckson P, Mech LD, Meyer TJ, Wayne RK (1992) Astudy of the genetic relationships within and among wolf packsusing DNA fingerprinting and mitochondrial DNA. BehavioralEcology and Sociobiology, 30, 83–94.
Marshall TC, Slate J, Kruuk LEB, Pemberton JM (1998) Statisticalconfidence for likelihood-based paternity inference in naturalpopulations. Molecular Ecology, 7, 639–655.
Mech LD (1970) The Wolf: The Ecology and Behaviour of an Endan-gered Species. Natural History Press, Garden City, NY.
Miller C, Joyce P, Waits LP (2002) Assessing allelic dropout andgenotype reliability using maximum likelihood. Genetics, 160,357–366.
Mills LS, Citta JJ, Lair KP, Schwartz MK, Tallmon DA (2000)Estimating animal abundance using noninvasive DNAsampling: promise and pitfalls. Ecological Applications, 10,283–294.
Neff MW, Broman KW, Mellersh CS et al. (1999) A second-generation linkage map of the domestic dog, Canis familiaris.Genetics, 151, 803–820.
Ostrander EA, Sprague GF, Rine J (1993) Identification andcharacterization of dinucleotide repeat (CA)n markers forgenetic-mapping in dog. Genomics, 16, 207–213.
Palsbøll PJ (1999) Genetic tagging: contemporary molecularecology. Biological Journal of the Linnean Society, 68, 3–22.
Palsbøll PJ, Allen J, Bérubé M et al. (1997) Genetic tagging ofhumpback whales. Nature, 388, 767–769.
Peters RP, Mech LD (1975) Scent-marking in wolves: a field study.American Scientist, 63, 628–637.
Poulle ML, Lequette B, Dahier T (1999) Le recolonisation des Alpespar le loup de 1992 à 1998. Bulletin Mensuel de l’Office National dela Chasse, 242, 4–13.
Pritchard JK, Stephens M, Donnelly PJ (2000) Inference of popula-tion structure using multilocus genotype data. Genetics, 155,945–959.
Queller DC, Goodnight KF (1989) Estimating relatedness usinggenetic markers. Evolution, 43, 258–275.
Randi E, Lucchini V (2002) Detecting rare introgression of
domestic dog genes into wild wolf (Canis lupus) populations byBayesian admixture analyses of microsatellite variation. Conserva-tion Genetics, 3, 31–45.
Randi E, Lucchini V, Christensen MF et al. (2000) MitochondrialDNA variability in Italian and east European wolves: detectingthe consequence of small population size and hybridization.Conservation Biology, 14, 464–473.
Raymond M, Rousset F (1995) genepop Version 1.2: populationgenetic software for exact tests and ecumenicism. Journal ofHeredity, 86, 248–249.
Reed JZ, Tollit DJ, Thompson PM, Amos W (1997) Molecular sca-tology: the use of molecular genetic analysis to assign species,sex and individual identity to seal faeces. Molecular Ecology, 6,225–234.
Saitou N, Nei M (1987) The neighbor-joining method: a newmethod for reconstructing phylogenetic trees. Molecular Biologyand Evolution, 4, 406–425.
Swofford DL (1998) PAUP. Phylogenetic Analysis Using Parsimony (andOther Methods), Version 4.0. Sinauer Associates, Sunderland, MA.
Taberlet P, Camarra JJ, Griffin S et al. (1997) Noninvasive genetictracking of the endangered Pyrenean brown bear population.Molecular Ecology, 6, 869–876.
Taberlet P, Griffin S, Goossens B et al. (1996) Reliable genotypingof samples with very low DNA quantities using PCR. NucleicAcids Research, 24, 3189–3194.
Taberlet P, Waits LP, Luikart G (1999) Noninvasive genetic sam-pling: look before you leap. Trends in Ecology and Evolution, 14,323–327.
Vilà C, Urios V, Castroviejo J (1994) Use of faeces for scentmarking in Iberian wolves (Canis lupus). Canadian Journal ofZoology, 72, 374–377.
Waits LP, Luikart G, Taberlet P (2001) Estimating the probabilityof identity among genotypes in natural populations: cautionsand guidelines. Molecular Ecology, 10, 249–256.
Wayne RK, Lehman N, Fuller TK (1995) Conservation genetics ofthe gray wolf. In: Ecology and Conservation of Wolves in aChanging World (eds Carbyn LN, Fritts SH, Seip DR), 399–407.Canadian Circumpolar Institute, Occasional Publication no.35. Edmonton, Alberta.
Weir BS, Cockerham CC (1984) Estimating F-statistics for theanalysis of population structure. Evolution, 38, 1358–1370.
Woods JC, Paetkau D, Lewis D et al. (1999) Genetic tagging offree-ranging black and brown bears. Wildlife Society Bulletin,27, 616–627.
Zimen E, Boitani L (1975) Number and distribution of Wolves inItaly. Zeitschrift für Säugetierkunde, 40, 102–112.
This research was conducted at the Conservation Geneticslaboratory at Italian Institute of Wildlife Biology (INFS), and it ispart of an ongoing project on wolf recolonization of the westernAlps (Progetto Lupo Interreg, Regione Piemonte). V. Lucchini andE. Fabbri are, respectively, researcher and PhD fellow at INFS andUniversity of Ferrara. This work is part of the PhD thesis byF. Marucco and S. Ricci under the supervision of Prof LuigiBoitani, University of Rome. E. Randi is head of ConservationBiology and Genetics at INFS.
All in-text references underlined in blue are linked to publications on ResearchGate, letting you access and read them immediately.