nmda antagonists block restraint-induced increase in extracellular dopac in rat nucleus accumbens
TRANSCRIPT

European Journal of Pharmacology, 162 (1989) 157-166 157 Elsevier
EJP 50642
N M D A antagonists block restraint-induced increase in extracellular DOPAC in rat nucleus accumbens
A n d r 6 Serrano, Maga l i D ' A n g i o and B e r n a r d Sca t ton *
Laboratoires d'Etudes et de Recherches Synthdlabo (L.E.R.S.), Biochemical Pharmacology Group, 31 Avenue Paul Vaillant-Couturier, 92220 Bagneux, France
Received 15 August 1988, revised MS received 24 October 1988, accepted 15 November 1988
The effects of the N-methyl-D-aspartate (NMDA) receptor antagonists CPP, TCP, PK 26124 and ifenprodil, and of the minor tranquillizer diazepam on stress-induced changes of dopamine metabolism in the nucleus accumbens were investigated in the rat. Dopamine metabolism was assessed by measuring the extracellular levels of 3,4-dihydroxyphen- ylacetic acid (DOPAC) by means of in vivo differential pulse voltammetry with electrochemically pretreated carbon fiber electrodes. Physical immobilization of the rats for 4 min caused a marked and long-lasting increase in extracellular DOPAC levels in the nucleus accumbens. A similar, though shorter-lasting, augmentation of extracellular DOPAC was observed in the nucleus accumbens after systemic administration of the anxiogenic agent methyl-/3- carboline-3-carboxylate (fl-CCM) (10 mg/kg s.c.). Pretreatment with CPP (1 mg/kg i.p.), TCP (3 mg/kg i.p.), PK 26124 (3 mg/kg i.p.), ifenprodil (3 mg/kg i.p.) or diazepam (2 mg/kg i.p.) totally antagonized the immobilization-in- duced increase in extracellular DOPAC in the nucleus accumbens. Diazepam and the benzodiazepine (¢0~_ 2) receptor antagonist flumazenil (30 mg/kg i.p.), but not ifenprodil, also antagonized the fl-CCM-induced activation of dopamine metabolism in the nucleus accumbens. Finally, systemic administration of haloperidol (25 /~g/kg i.p.) increased the extracellular concentrations of DOPAC in the nucleus accumbens, but pretreatment with ifenprodil (3 mg/kg i.p.) did not modify this response. These data indicate that NMDA receptor antagonists prevent the activation of dopamine metabolism in the nucleus accumbens caused by immobilization stress but not by /3-CCM-induced anxiogenic stimulation. These results suggest that NMDA receptor antagonists may possess an anxiolytic-like action in the rodent, which is exerted via neuroanatomical circuits distinct from those acted upon by diazepam.
NMDA receptor antagonists; Nucleus accumbens; Dopamine metabolism; Immobilization stress; Methyl-/3-carboline-3-carboxylate (/3-CCM); (In vivo voltammetry)
1. I n t r o d u c t i o n
In the mammalian central nervous system the majority of excitatory neurons use excitatory amino acids (inter alia L-glutamate and L-aspar- tate) as their transmitters (Watkins and Evans, 1981). Receptors for excitatory amino acids are at present classified into three subtypes termed N -
* To whom all correspondence should be addressed.
methyl-D-aspartate (NMDA), kainate and quis- qualate after their preferred agonists (Davies et al., 1980; Watkins and Evans, 1981). To date, only the N M D A class of excitatory amino acid recep- tors has a wide range of selective antagonists e.g. 2-amino-5-phosphonovalerate , CPP (3-[(__+ )-2- carboxypiperazin-4-yl]-propyl- 1 -phosphonic acid, Watkins and Olverman, 1987), TCP (N-(1-[2- thienyl]-cyclohexyl)- 3,4-piperidine), phencyclidine, M K 801 ( (+) -5-methy l -10 ,11-d ihydro-5H-di - benzo[a,d]cyclohepten-5,10-imine, Kemp et al., 1987), PK 26124 (riluzole, 2-amino-6-trifluoro-
0014-2999/89/$03.50 © 1989 Elsevier Science Publishers B.V. (Biomedical Division)

158
methoxy-benzothiazole, Benavides et al., 1985) or ifenprodil (Vadilex ®, (+)-(R,S)-c~-(4-hydroxy- phenyl)- /~-methyl-4-(phenylmethyl)- l-piperidine ethanol, Scatton et aL 1987; Carter et al., 1988a,b).
Most of the current interest in selective NMDA receptor antagonists centres around their demon- strated anticonvulsant, muscle relaxant and anti- ischaemic properties (Meldrum, 1985; Turski et al., 1987). The NMDA receptor antagonists MK 801, 2-amino-7-phosphonoheptanoate and CPP have also been shown to produce discriminative stimuli and anticonflict effects in rats and mice similar to those of clinically active anxiolytics (Clineschmidt et al., 1982; Bennett and Amrick, 1986; 1987; Stephens et al., 1986). This raises the possibility that NMDA receptor antagonists ex- hibit anxiolytic-like activity.
Anxiolytic drugs of various chemical classes are known to antagonize the increase in dopamine (DA) metabolism in the frontal cortex or nucleus accumbens elicited by mild stress in the rodent (Lavielle et al,, 1978; Fadda et al., 1978; Rein- hard, 1982; Claustre et al., 1986; D'Angio et al., 1987). This effect is primarily linked to the anxio- lytic (and not to the sedative or myorelaxant) properties of these drugs (Claustre et al., 1986). In order to further evaluate the anxiolytic potential of NMDA receptor antagonists, we have investi- gated the effects of pretreatment with competitive (CPP) and non-competitive (TCP, PK 26124, ifenprodil) NMDA receptor antagonists on DA metabolism in the nucleus accumbens of rats sub- jected to a short-lasting (4 min) restraint (MK 801 could not be used because in the preliminary studies this compound per se caused a marked enhancement of DA metabolism in the rat nucleus accumbens at doses as low as 0.1 mg/kg i.p.). The effects of the benzodiazepine receptor (redes- ignated ~ receptor, Langer and Arbilla, 1988a,b) agonist diazepam were studied in parallel for com- parison. Moreover, since the preferential ~01 recep- tor inverse agonists e.g. FG 7142 or methyl-/~- carboline carboxylate (/~-CCM) cause behavioural manifestations of anxiety in animal models (Prado de Carvalho et al., 1983; File et al., 1984) and are anxiogenic in man (Dorow et al., 1983), we also studied the effects of ifenprodil and diazepam on the alterations of DA metabolism in the nucleus
accumbens caused by systemic administration of /LCCM. Finally, to assess the specificity of NMDA receptor antagonists to prevent the anxiety-linked increase in DA metabolism, we investigated the effects of ifenprodil on the augmentation of DA metabolism in the nucleus accumbens caused by the neuroleptic agent haloperidol.
In this study DA metabolism in the nucleus accumbens of rats was assessed by measuring the extracellular levels of 3,4-dihydroxyphenylacetic acid (DOPAC) by means of in vivo voltammetry with electrochemically pretreated carbon fiber electrodes (Louilot et al., 1985; D'Angio et al., 1987; 1988), In vivo voltammetry enables the di- rect and sequential measurement of the levels of the major dopamine metabolite DOPAC in dis- crete brain areas of conscious, freely moving rats.
2. Materials and methods
Male Sprague Dawley rats (COBS CD strain from Charles River, France) weighing 250-300 g were used. The rats were housed at 22 _+ 0.5 ° C in individual home cages under a 12 h l ight/dark cycle (lights on from 07:00 to 19:00) and had free access to food and water.
Diazepam, PK 26124, /~-CCM, flumazenil (Ro15-1788) and ifenprodil tartrate were syn- thetized by the Chemistry Department, LERS Paris. CPP and haloperidol were purchased from Tocris (UK) and RBI (MA, USA), respectively; TCP was kindly donated by Dr. Chicheportiche (Montpellier, France). CPP, TCP and ifenprodil were dissolved in distilled water. Haloperidol was dissolved in a 2% solution of tartaric acid and /~-CCM in a few drops of 0.1 M hydrochloric acid. PK 26124, flumazenil and diazepam were adminis- tered as a suspension in a 0.1% solution of Tween 80.
Voltammetric measurements were made in un- restrained awake animals using a classical 3 elec- trode system with working carbon fiber, reference and auxiliary electrodes and a PRG5 polarograph (Tacussel, France) (D'Angio et al., 1987; 1988). Working electrodes made of 3 pyrrolytic carbon fibres (diameter 8 /xm, length 500 /~m) were elec- trochemically pretreated before use by applying a

triangular wave potential of + 1.5 V for 20 s as described previously (Gonon et al., 1981). The carbon fiber electrode was positioned in the brain areas with the help of an implantation assembly (Louilot et al., 1987), which was implanted stereo- taxically 7 days prior to the voltammetric record- ings under chloral hydrate (400 mg /kg i.p.) anaesthesia. The stereotaxic coordinates were: 2.2 mm anterior to bregma, 1.5 mm lateral to the midline and 6 mm below the cortex surface. The implantation assembly was anchored to the skull surface with 4 stainless steel screws and acrylic dental cement. The reference electrode (Ag/AgC1- coated silver wire) was placed between the dura mater and the skull, while the auxiliary platinum electrode was soldered to a stainless steel screw implanted in the bone of the skull. The working, auxiliary and reference electrodes were connected to a small three pin plug cemented to the skull. The animals were allowed one week to recover before they were submitted to the stress proce- dure.
For voltammetric recordings, the electrochem- ically pretreated working electrode was positioned via the implantation assembly, and the plug was connected to the polarograph through a flexible cable and swivel connector. The following record- ing parameters were used: ramp potential from - 2 0 0 mV to + 400 mV; scan rate 10 mV/s ; pulse modulation of a square wave form (pulse ampli- tude 50 mV, pulse period 0.2 s, pulse duration 48 ms). Differential pulse voltammograms were re- corded every 2 min. The carbon fibre electrodes were calibrated with a 20 I~M DOPAC/200 /~M ascorbic acid solution in 0.1 M phosphate buffer, pH 7.4 before and after each experiment. Electro- chemical signals were quantified automatically by measuring the height of the oxidation peaks re- corded at - 100 mV (ascorbic acid) and + 100 mV (DOPAC) with a SP 4100 computing integrator (Spectra Physics). There is always some variation in the absolute basal levels of the oxidation cur- rent, which corresponds to DOPAC, between animals. For this reason, the in vivo voltammetric data were expressed as follows: for each individ- ual animal, the heights of the electrochemical sig- nals measured every 2 rain during the 30 min-period preceding the restraint stress or fl-CCM
159
or haloperidol injection were averaged and used as a control baseline value ( = 100%), and individual data (including those of the baseline period) were re-expressed as percentages of this control value. The means with S.E.M. of results obtained from 5 to 6 rats were calculated for the same periods.
For the immobilization stress, the rats were restrained for 4 rain in the hands of the experi- menter. Statistical analysis of the data was per- formed with the non-paired two-tailed Student's t-test; each point was compared with the average _+ S.E.M. of the control period (30 min).
3. Results
As observed previously (D'Angio et al., 1987), an electrochemical signal corresponding to the oxidation potential of DOPAC was recorded at + 100 mV in the rat nucleus accumbens. Earlier pharmacological studies have clearly established the identity of the catechol peak measured from differential pulse voltammograms to be DOPAC (Louilot et al., 1985). Under our experimental conditions, the electrochemical signal recorded at + 100 mV from the nucleus accumbens of control rats was stable over at least a 4 h period (data not shown).
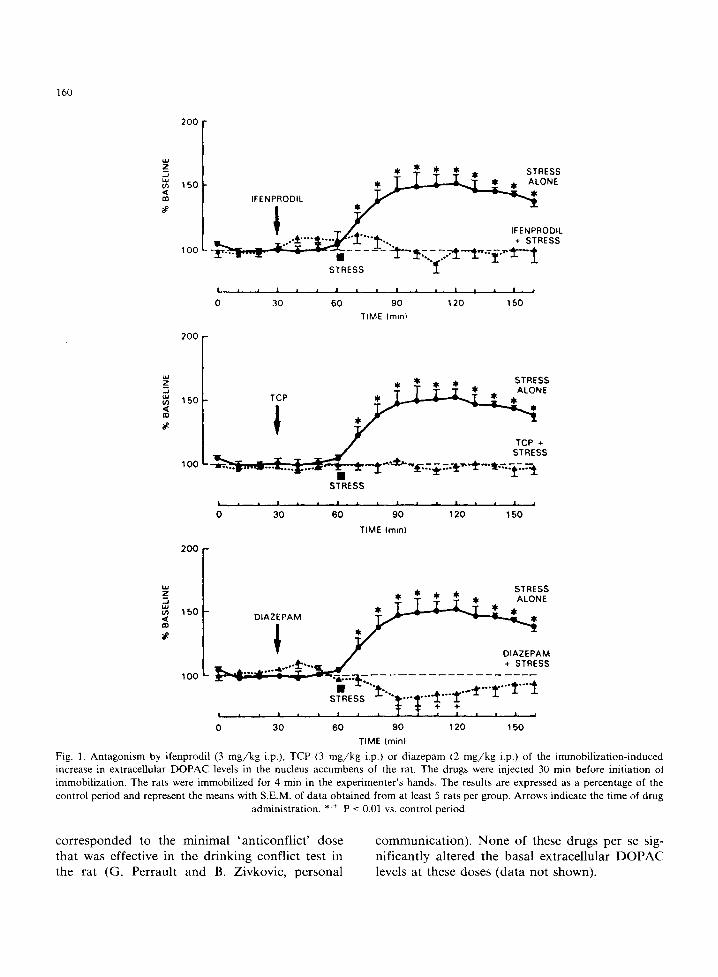
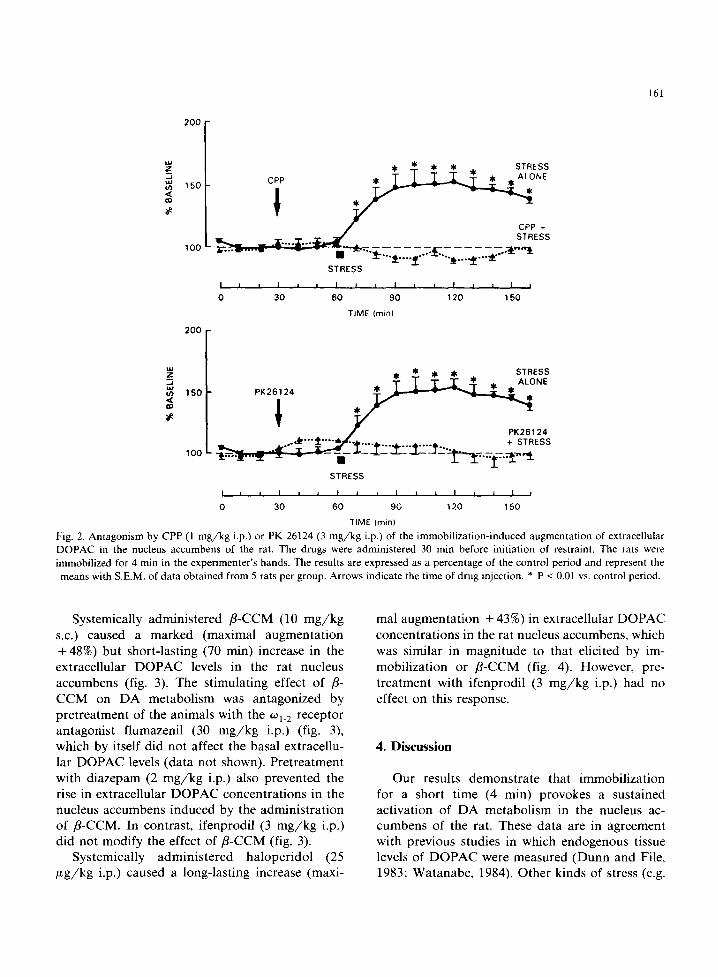
Physical immobilization of the rats for a short time (4 min) in the experimenter's hands caused a marked increase (maximal augmentation +50%) in the amplitude of the DOPAC oxidation peak in the nucleus accumbens (figs. 1 and 2). The DOPAC peak remained elevated for at least 100 min after cessation of the stress. Whereas pretreatment with saline was without an effect on the elevation of extracellular DOPAC concentrations caused by stress, pretreatment with the anxiolytic agent di- azepam (2 mg/kg i.p.) totally antagonized, and even slightly reversed, the immobilization-induced increase in extracellular DOPAC levels in the nucleus accumbens (fig. 1). Similarly, pretreat- ment with ifenprodil (3 m g /k g i.p.), TCP (3 mg/kg i.p.), CPP (1 m g /k g i.p.) or PK 26124 (3 mg/kg i.p.) totally abolished the stress-induced augmen- tation of extracellular DOPAC in the nucleus ac- cumbens (figs. 1 and 2). The doses of NMDA receptor antagonists used in these experiments
160
200
_ a
150
c o
100
200
d= ~ * * ~ STRESS W= ~ ALONE
/ . IFENPRODIL _ _ _ _ _ .':: T'RESS
~ ' " • - - Y~") .:.- . - . t "~--" "~" '~ ' - 'T STRESS
i i i 1 J i i 1 ! • 1 J
0 30 60 90 120 1 SO
TIME (mon)
150
100
200
11= ,~. ~ STRESS I T "I" , ALONE
TCP * ~ 1 $
~ " TCP + _ _ ~ ~ ._,IE=._ _ ~..~.___ S_TR_ ESS
- = , . . - T ~ - - ~ " ~ "-~V: ~ = .~-" "e -- ,--:..]:-._,t. STRESS
| J 1 1 | I | | I i
0 30 60 90 120 150
TIME (rnin)
* ~ * , STRESS -.% '-r 1 ',r T =k ALONE
,~ DIAZEPAM
° 1 Y 2 DIA M ~,. / + STRESS .~. .°- • _ ~ , ~ . = , ~ , . , _ ~ ~ . . . . . . _ - - ~
100 =. ,r - - ~,~ - .~....~ . . . . . . . . . _
ESS t"" ! T s $'"T . . . . .
I i i ! • I I J A / 1 I I I I | J
0 30 60 90 120 150
TIME (rain}
Fig. 1. Antagonism by ifenprodil (3 m g / k g i.p,), TCP (3 m g / k g i.p.) or diazepam (2 m g / k g i.p.) of the immobilization-induced increase in extracellular DOPAC levels in the nucleus accumbens of the rat. The drugs were injected 30 min before initiation ot immobilization. The rats were immobilized for 4 min in the experimenter 's hands. The results are expressed as a percentage of the control period and represent the means with S.E,M. of data obtained from at least 5 rats per group. Arrows indicate the time of drug
administration. * '+ P < 0.01 vs. control period.
corresponded to the minimal 'anticonflict ' dose that was effective in the drinking conflict test in the rat (G. Perrault and B. Zivkovic, personal
communication). None of these drugs per se sig- nificantly altered the basal extracellular DOPAC levels at these doses (data not shown).
u~
1/) < oo
200
150
100
200
, ~ * * , STRESS
CPP * I ~ ~ ~ ,ALONE
/ CPP + . . . . . . _ . . . . . . . ESS
~ ' ~ ' = - i . - ~ ~ j - - . - L ~ . a " ~ • ~'"~'"'I . . . . . . . * ' " i " ' ~ "
STRESS I l l l a l l l l l l l , l l l l l
0 30 60 90 120 150 TIME (min)
161
w STRESS -T- T :i- -i- ~ , ALONE
a e'~ 1s0 PK2i24
/ PK26124 ~ ~ - - ~ - - - 4 - - - * ~ + STRESS
100 ~ " ~-~"~]" "" "-:~qj~-=. =7.f..-~-~ , ~
STRESS
I A I I I I I I I 1 J l I = J I i
0 30 60 9(5 120 150 TIME (rain)
Fig. 2. Antagonism by CPP (1 mg/kg i.p.) or PK 26124 (3 mg/kg i.p.) of the immobilization-induced augmentation of extracellular DOPAC in the nucleus accumbens of the rat. The drugs were administered 30 min before initiation of restraint. The rats were immobilized for 4 min in the experimenter's hands. The results are expressed as a percentage of the control period and represent the means with S.E.M. of data obtained from 5 rats per group. Arrows indicate the time of drug injection. * P < 0.01 vs. control period.
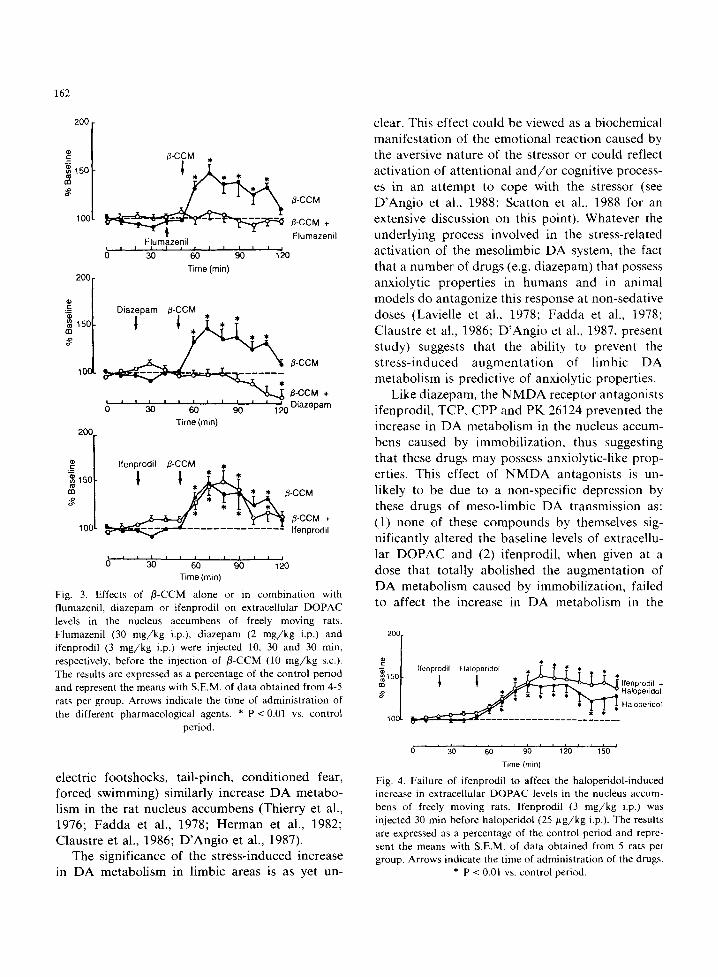
Systemically administered B-CCM (10 mg/kg s.c.) caused a marked (maximal augmentation + 48%) but short-lasting (70 min) increase in the extracellular DOPAC levels in the rat nucleus accumbens (fig. 3). The stimulating effect of /3- CCM on DA metabolism was antagonized by pretreatment of the animals with the c0~_= receptor antagonist flumazenil (30 mg/kg i.p.) (fig. 3), which by itself did not affect the basal extracellu- lar DOPAC levels (data not shown). Pretreatment with diazepam (2 mg/kg i.p.) also prevented the rise in extracellular DOPAC concentrations in the nucleus accumbens induced by the administration of/3-CCM. In contrast, ifenprodil (3 mg /kg i.p.) did not modify the effect of/3-CCM (fig. 3).
Systemically administered haloperidol (25 /~g/kg i.p.) caused a long-lasting increase (maxi-
mal augmentation + 43%) in extracellular DOPAC concentrations in the rat nucleus accumbens, which was similar in magnitude to that elicited by im- mobilization or /3-CCM (fig. 4). However, pre- treatment with ifenprodil (3 mg /kg i.p.) had no effect on this response.
4. Discussion
Our results demonstrate that immobilization for a short time (4 min) provokes a sustained activation of DA metabolism in the nucleus ac- cumbens of the rat. These data are in agreement with previous studies in which endogenous tissue levels of DOPAC were measured (Dunn and File, 1983; Watanabe, 1984). Other kinds of stress (e.g.
162
200
=o
~ 1 5 0
10¢
200
13-C~M ,
fl-CCM
/~-CCM + Flumazenil
() ' ' Flum~zeni' ' 3 0 60 ' ~)0 ' ' 120
Time (min)
,~ ~5c
100
20(3
=*
iso ~3
10C
Diazepam fl-CCM
1 I
. . . . . . . . fl-CCM
0 30 60 90 120 Dlazepam Time (min)
Ifenprodil ,6'-CC M ,
TM
. , ~ " ' ~ t - ~ ~-CCM+ ~"=~=. - ~ . ~ - ~ - . . . . . . . . . . . . . . ~" Ifenprodil
6 ' ' 0 120 Time (min)
Fig. 3. Effects of fl-CCM alone or in combination with flumazenil, diazepam or ifenprodil on extracellular DOPAC levels in the nucleus accumbens of freely moving rats. Flumazenil (30 mg/kg i.p.), diazepam (2 mg /kg i.p.) and ifenprodil (3 mg/kg i.p.) were injected 10, 30 and 30 min, respectively, before the injection of fl-CCM (10 mg/kg s.c.). The results are expressed as a percentage of the control period and represent the means with S.E.M. of data obtained from 4-5 rats per group. Arrows indicate the time of administration of the different pharmacological agents. * P < 0.01 vs. control
period.
electric footshocks, tail-pinch, conditioned fear, forced swimming) similarly increase DA metabo- lism in the rat nucleus accumbens (Thierry et al., 1976; Fadda et al., 1978; Herman et al., 1982; Claustre et al., 1986; D'Angio et al., 1987).
The significance of the stress-induced increase in IDA metabolism in limbic areas is as yet un-
clear. This effect could be viewed as a biochemical manifestation of the emotional reaction caused by the aversive nature of the stressor or could reflect activation of attentional a n d / o r cognitive process- es in an at tempt to cope with the stressor (see D'Angio et al., 1988; Scatton et al., 1988 for an extensive discussion on this point). Whatever the underlying process involved in the stress-related activation of the mesolimbic DA system, the fact that a number of drugs (e.g. diazepam) that possess anxiolytic properties in humans and in animal models do antagonize this response at non-sedative doses (Lavielle et al., 1978; Fadda et al., 1978; Claustre et al., 1986; D'Angio et al., 1987, present study) suggests that the ability to prevent the stress-induced augmenta t ion of limbic DA metabolism is predictive of anxiolytic properties.
Like diazepam, the N M D A receptor antagonists ifenprodil, TCP, CPP and PK 26124 prevented the increase in DA metabolism in the nucleus accum- bens caused by immobilization, thus suggesting that these drugs may possess anxiolytic-like prop- erties. This effect of N M D A antagonists is un- likely to be due to a non-specific depression by these drugs of meso-limbic DA transmission as: (1) none of these compounds by themselves sig- nificantly altered the baseline levels of extracellu- lar DOPAC and (2) ifenprodil, when given at a dose that totally abolished the augmentation of DA metabolism caused by immobilization, failed to affect the increase in DA metabolism in the
IOOL
Ifenprodil Haloperido[ , ~ ~ * * * .
', , ' , ' "a'°p°"°°' _ _~J]~"~ * ~ J ~ Haloperidol
. . . . . . . . . . . . _ ,__*___
o " " ~ o " ~ o ' ' 9 ' o ' ' 1 ~ o ' ' 4 o '
Time (min)
Fig. 4. Failure of ifenprodil to affect the haloperidol-induced increase in extracellular DOPAC levels in the nucleus accum- bens of freely moving rats. Ifenprodil (3 mg/kg i.p.) was injected 30 min before haloperidol (25 ~g /kg i.p.). The results are expressed as a percentage of the control period and repre- sent the means with S.E.M. of data obtained from 5 rats per group. Arrows indicate the time of administration of the drugs.
* P < 0.01 vs. control period.

nucleus accumbens induced by haloperidol or fl- CCM. A non-specific effect of NMDA antagonists due to sedation or myorelaxation is also unlikely to account for their blockade of the stress re- sponse since at the low doses used in the present study, none of these drugs exhibits sedative or muscle relaxant properties (Bennett and Amrick, 1987; Lehmann et al., 1987; Turski et al., 1987; Stutzmann et al., 1988; Perrault and Zivkovic, personal communication). Finally, a direct inter- action of the NMDA receptor antagonists with ~0 receptors can also be excluded since none of the drugs investigated has affinity for central ~o 1 or ~02 receptors (Benavides et al., 1985; Lehmann et al., 1987; Carter et al., 1988a,b). However, it is still debated whether CPP interacts with ~0 receptors (Lehmann et al., 1987; White et al., 1988). The fact that the doses at which the NMDA receptor antagonists investigated block the immobiliza- tion-induced activation of DA metabolism roughly correspond to those that effectively antagonize NMDA-mediated biochemical responses in vivo (e.g. harmaline-induced increase in cyclic GMP in the mouse cerebellum or NMDA-evoked libera- tion of DA in the rat striatum, see Carter et al., 1988a,b) favours the view that the prevention by these drugs of the stress response is related to NMDA receptor antagonism.
The prevention by NMDA receptor antagonists of the immobilization-induced increase in DA metabolism in the nucleus accumbens (reminiscent of the effects of minor tranquillizers) suggests a clinical application of such agents as novel anxio- lyrics. The fact that CPP (Bennett and Amrick, 1987), PK 26124 and ifenprodil (Perrault and Zivkovic, personal communication) exhibit anti- conflict effects in the rat at doses similar to those used in this study supports this view. These data raise the possibility that neurons that utilize exci- tatory amino-acid neurotransmitters play a part in emotional behavior. That conditioned emotional responses increase the turnover of L-glutamate and L-aspartate in a variety of brain areas in- cluding the nucleus accumbens and ventral teg- mental area (Lane et al., 1982) is consistent with this view.
The anatomical site(s) at which NMDA recep- tor antagonists act to reduce the activation of
163
mesolimbic DA neurons caused by stress remain(s) to be determined. Both the ventral tegmental area and nucleus accumbens are endowed with sub- stantial densities of N MD A receptors (Maragos et al., 1988), and these brain regions could represent possible targets for N MD A receptor antagonists. Moreover, the nucleus accumbens is known to contain intrinsic excitatory amino acidergic cells and to receive glutamatergic afferents from the hippocampal formation (Walaas and Fonnum, 1979), a brain region involved in anxiety (Kuhar, 1986).
The fl-carbolines are a class of pharmacological agents that have been shown not only to antago- nize the actions of the benzodiazepines but also to possess intrinsic actions of their own. These intrin- sic actions are opposite to those of the benzo- diazepines; fl-carbolines have therefore been termed 'inverse agonists' (M/Shler and Richards, 1983). In this regard, fl-carbolines have been re- ported to cause anxiety states in monkeys (Craw- ley et al., 1985; Ninan et al., 1983) and humans (Dorow et al., 1983), to produce proconflict effects in the punished drinking test in the rat (Corda et al., 1983) and to cause behavioural manifestations of anxiety in the rat (File et al., 1984; Novas et al., 1988). These various studies thus point to the anxiogenic effects of fl-carbolines.
Like immobilization stress, systemic adminis- tration of fl-CCM was found to be associated with an activation of DA metabolism in the nucleus accumbens. Previous reports have also shown that systemic administration of fl-CCM, ethyl-fl- carboline-3-carboxylate or FG 7142 increase post- mortem tissue levels of DOPAC in the rat frontal cortex (Tam and Roth, 1985; Claustre et al., 1986). The blockade by flumazenil of the activating ef- fect of fl-CCM on DA metabolism suggests that it is mediated by an ¢0 receptor. The anxiogenic potential of fl-carbolines (see above) together with the fact that the fl-CCM-induced increase in DA metabolism in the nucleus accumbens was antagonized by pretreatment with a non-sedative dose of diazepam strongly suggest that this effect of fl-CCM most likely results from its anxiety-pro- moting action. However, unlike diazepam, ifen- prodil, when given at a dose that totally prevented immobilization-induced increase in DA metabo-

164
lism, failed to prevent the /~-CCM-induced bio- chemical response. This suggests that the 'anxio- lyric action' of NMDA receptor antagonists is exerted at brain sites other than those involved in the action of ~0 receptor agonists.
The mechanism whereby fl-carbolines increase DA turnover in the nucleus accumbens is not yet clearly defined. It has been generally accepted that ~0 receptor agonists exert their pharmacological actions by interacting with the GABA receptor macromolecular complex, resulting in an enhance- ment of GABA neurotransmission (see M~Shler and Richards, 1983). In contrast, inverse agonists such as j3-CCM exert actions opposite to those of agonists, resulting in a reduction of postsynaptic GABA neurotransmission. In view of the evidence for an inhibitory GABAergic control of meso- limbic DA neurons (Fuxe et al., 1975; Walaas and Fonnum, 1980; Scatton et al., 1982), it is possible that the increase in DA metabolism in the nucleus accumbens induced by/~-CCM is subsequent to a diminished GABAergic transmission. Accordingly, diazepam would be able to counteract the effect of /~-CCM on DA neurons in the nucleus accumbens by restoring a normal GABAergic tone.
The lack of blockade by ifenprodil of the activating effects of/~-CCM on DA metabolism in the nucleus accumbens suggests that the interac- tion between NMDA receptor antagonists and /~-CCM does not occur at the level of the dopaminergic neurons themselves. The observa- tion that NMDA receptor antagonists do not in- teract directly with mesolimbic DA neurons is also supported by the inability of ifenprodil to antago- nize the haloperidol-induced increase in DA metabolism in the nucleus accumbens. Together, these data suggest that the site at which NMDA antagonists act to prevent an environmentally elicited activation of DA neurons that project to the nucleus accumbens is upstream of the DA neurons.
In conclusion, the results of the present study indicate that, like minor tranquillizers, competitive and non-competitive NMDA receptor antagonists prevent the restraint stress-induced activation of DA neurons that project to the nucleus accum- bens. These data add further weight to the conten- tion that NMDA receptor antagonists may repre-
sent a novel class of anxiolytic agents and suggest that NMDAergic synapses are involved in en- vironmentally elicited activation of mesolimbic DA neurons. The inability of the NMDA receptor antagonist ifenprodil to prevent the fl-CCM-in- duced increase in DA metabolism indicates that the neuroanatomical circuits involved in the anxio- lytic action of these compounds differ from those involved in the anxiolytic effect of diazepam. NMDA antagonists and diazepam, although acting through different mechanisms, may thus result in a similar end-effect.
Acknowledgement
The authors wish to thank V. Eninger for typing the manuscript.
References
Benavides, J., J.C. Camelin, N. Mitrani, F. Flamand, A. Uzan, J.J. Legrand, C. Gueremy and G. Lefur, 1985, 2-Amino-6- trifluoromethoxy-benzothiazole, a possible antagonist of excitatory amino acid neurotransmission. II. Biochemical properties, Neuropharmacology 24, 1085.
Bennett, D.A. and C.L. Amrick, 1986, 2-Amino-7-phosphono- heptanoic acid (AP7) produces discriminative stimuli and anticonflict effects similar to diazepam, Life Sci. 39, 2455.
Bennett, D.A. and C.L. Amrick, 1987, Antagonists at the N-methyl-D-aspartate receptor produce anticonflict effects, in: Excitatory Amino Acid Transmission, eds. T.P. Hicks, D. Lodge and H. McLennan (Alan R. Liss Inc., New York) p. 213.
Carter, C.J., J. Benavides, P. Legendre, J.D. Vincent, F. Noel, F. Thuret, K.G. Lloyd, S. Arbilla, B. Zivkovic, E.T. Mac- Kenzie, B. Scatton and S.Z. Langer, 1988a, Ifenprodil and SL 82.0715 as cerebral anti-ischaemic agents. II. Evidence for NMDA receptor antagonist properties, J. Pharmacol. Exp. Ther. 247, 1222.
Carter, C.J., F. Noel, J. Benavid+s, J.P. Rivy, F. Thuret, K.G. Lloyd, S. Arbilla, S.Z. Langer, B. Zivkovic and B. Scatton, 1988b, |fenprodil, a potent non-competitive NMDA antagonist with a novel mechanism of action, Br. J. Pharmacol. 93, 88P.
Claustre, Y., J.P. Rivy, T. Dennis and B. Scatton, 1986, Pharmacological studies on stress-induced increase in fron- tal cortical dopamine metabolism in the rat, J. Pharmacol. Exp. Ther. 238, 693.
Clineschmidt, B.V., M. Williams, J.J. Witoslawski, P.B. Bunt- ing, E.A. Risley and J.A, Totaro, 1982, Restoration of shock-suppressed behavior by treatment with ( + )5-methyl-

10,11 -dihydro-5 H-dibenzo[a,d]cyclohepten-5,10-imine (MK 801), a substance with potent anticonvulsant, central sym- pathomimetic, and apparent anxiolytic properties, Drug. Dev. Res. 2, 147.
Corda, M.G., W.D. Blacker, W.B. Mendelson, A. Guidotti and E. Costa, 1983, Beta-carboline enhance shock-induced sup- pression of drinking in rats, Proc. Natl. Acad. Sci. 80, 2072.
Crawley, J., P. Ninan, D. Pickar, G. Chrousos, M. Linnoila, P. Skolnick and S. Paul, 1985, Neuropharmacological antagonism of the beta-carboline-induced anxiety response in rhesus monkeys, J. Neurosci. 5, 477.
D'Angio, M., A. Serrano, P. Driscoll and B. Scatton, 1988, Stressful environmental stimuli increase extracellular DOPAC levels in the prefrontal cortex of hypoemotional (Roman high-avoidance) but not hyperemotional (Roman low-avoidance) rats. An in vivo voltammetric study, Brain Res. 451, 237.
D'Angio, M., A. Serrano, J.P. Rivy and B. Scatton, 1987, Tail-pinch stress increases extracellular DOPAC levels (as measured by in vivo voltammetry) in the rat nucleus ac- cumbens but not frontal cortex: antagonism by diazepam and zolpidem, Brain Res. 409, 169.
Davies, J., R.H. Evans, A.A. Francis, A.W. Jones and J.C. Watkins, 1980, Excitatory amino acid receptors in the vertebrate central nervous system, in: Neurotransmitters and Their Receptors, eds. U.Z. Littaner, Y. Dudai, I. Silman, V.I. Teichberg and Z. Vogel (John Wiley and Sons Ltd.) p. 333.
Dorow, R., R. Horowski, G. Paschelke, M. Amin and C. Braestrup, 1983, Severe anxiety induced by FG 7142, a fl-carboline ligand for benzodiazepine receptors, Lancet ii, 98.
Dunn, A..I. and S.E. File, 1983, Cold restraint alters dopamine metabolism in frontal cortex, nucleus accumbens and neostriatum, Physiol. Behav. 31, 511.
Fadda, F., A. Argiolas, M.R. Melis, A.H. Tissari, P.L. Onali and G.L. Gessa, 1978, Stress-induced increase in 3,4-dihy- droxyphenylacetic acid (DOPAC) levels in the cerebral cortex and in nucleus accumbens: reversal by diazepam, Life Sci. 23, 2219.
File, S.E., R.G. Lister, R. Maninov and J.C. Tucker, 1984, Intrinsic behavioral actions of propyl-B-carboline- 3-carboxylate, Neuropharmacology 23, 463.
Fuxe, K., T. H~kfelt, A. Ljungdahl, L. Agnati, O. Johansson and M. Perez de la Mora, 1975, Evidence for an inhibitory gabaergic control of the mesolimbic dopamine neurons: possibility of improving treatment of schizophrenia by combined treatment with neuroleptic and gabaergic drugs, Med. Biol. 53, 177.
Gonon, F., C. Fombarlet, M. Buda and J.F. Pujol, 1981, Improvement of pyrolytic carbon fiber electrodes by elec- trochemical treatments, Anal. Biochem. 53, 1386.
Herman, .I.P., D. Guillonneau, R. Dantzer, B. Scatton, L. Semerdjian-Rouquier and M. Le Moal, 1982, Differential effects of inescapable footshocks and of stimuli previously paired with inescapable footshocks on dopamine turnover in cortical and limbic areas of the rat, Life Sci. 30, 2207.
165
Kemp, J.A., A.C. Foster and E. Wong, 1987, Non-competitive antagonists of excitatory amino acid receptors, Trends Neurosci. 10, 294.
Kuhar, M.J., 1986, Neuroanatomical substrates of anxiety: a brief survey, Trends Neurosci. 6, 307.
Lane, J.D., M.P. Sands, M.E. Freeman, D.R. Cherek and .I.E. Smith, 1982, Amino acid neurotransmitter utilization in discrete rat brain regions is correlated with conditioned emotional response, Pharmacol. Biochem. Behav. 16, 329.
Langer, S.Z. and S. Arbilla, 1988a, Imidazopyridines as a tool for the characterization of benzodiazepine receptors: a pro- posal for a pharmacological classification as omega sub- types, Pharmacol. Biochem. Behav. 29, 763.
Langer, S.Z. and S. Arbilla, 1988b, Limitations of the benzodi- azepine receptor nomenclature: a proposal for a pharmaco- logical classification as omega receptor subtypes, Fundam. Clin. Pharmacol. 2, 159.
Lavielle, S., J.P. Tassin, A.M. Thierry, G. Blanc, D. Herv& C. Barthrlrmy and .I. Glowinski, 1978, Blockade by benzodi- azepines of the selective high increase in dopamine turnover induced by stress in mesocortical dopaminergic neurons of the rat, Brain Res. 168, 585.
Lehmann, .I., J. Schneider, S. McPherson, D.E. Murphy, P. Bernard, C. Tsai, D.A. Bennett, G. Pastor, D.J. Steel, C. Boehm, D.L. Cheney, J.M., Liebman, M. Williams and P.L. Wood, 1987, CPP, a selective N-methyl-D-aspartate (NMDA) type receptor antagonist: characterization in vitro and in vivo, J. Pharmacol. Exp. Ther. 240, 737.
Louilot, A., M. Buda, F. Gonon, H. Simon, M. LeMoal and J.F. Pujol, 1985, Effect of haloperidol and sulpiride on dopamine metabolism in nucleus accumbens and olfactory tubercle: a study by in vivo voltammetry, Neuroscience 14, 775.
Louilot, A., A. Serrano and M. D'Angio, 1987, A novel carbon fiber implantation assembly for cerebral voltammetric mea- surements in freely moving rats, Physiol. Behav. 41,227.
Maragos, W.F., J.B. Penney and A.B. Young, 1988, Anatomic correlation of NMDA and 3H-TCP-labelled receptors in rat brain, J. Neurosci. 8, 493.
Meldrum, B., 1985, Possible therapeutic applications of antagonists of excitatory amino acid neurotransmitters, Clin. Sci. 68, 113.
Mt~hler, I.T. and G. Richards, 1983, Anxiolytics: neurochem- ical, behavioral and clinical perspectives, in: Receptors for Anxiolytic Drugs, eds. S. Malick, S. Enna and H. Yamamura (Raven Press, New York) p. 15.
Ninan, P.T., T. Insel, R.M. Cohen, J.M. Cook, P. Skolnick and S.M. Paul, 1983, Benzodiazepine receptor mediated anxiety in primates, Science 218, 1332.
Novas, M.L., C. Wolfman, J.H. Medina and E. DeRobertis, 1988, Proconvulsant and 'anxiogenic' effects of n-butyl-fl- carboline-3-carboxylate, an endogenous benzodiazepine binding inhibitor from brain, Pharmacol. Biochem. Behav. 30, 331.
Prado de Carvalho, L., G. Grecksch, G. Chapouthier and J. Rossier, 1983, Anxiogenic and non-anxiogenic benzodi- azepine antagonists, Nature (London) 301, 64.

166
Reinhard, J.F., M.J. Bannon and R.H. Roth, 1982, Accelera- tion by stress of dopamine synthesis and metabolism in prefrontal cortex: antagonism by diazepam, Naunyn- Schmiedeb. Arch. Pharmacol. 318, 374.
Scatton, B., C. Carter, Y. Claustre, R. L'Heureux, S. Arbilla, S.Z. Langer, B. Gotti, D. Duverger and E.T. MacKenzie, 1987, The cerebral anti-ischaemic agents, ifenprodil and SL 82.0715, antagonise the effects of NMDA, Xth Interna- tional Congress of Pharmacology (Sydney), Abstr. No. 012.064.
Scatton, B., M. D'Angio, P. Driscoll and A. Serrano, 1988, An in vivo voltammetric study of the response of mesocortical and mesoaccumbens dopaminergic neurons to environmen- tal stimuli in strains of rats with differing levels of emotion- ality, in: The Mesocorticolimbic Dopamine System, eds. P.W, Kalivas and C.B. Nemeroff, Ann. New York Acad. Sci. Vol. 357, p. 214.
Scatton, B., B. Zivkovic, J. Dedek, K.G. Lloyd, J. Constan- tinidis, R. Tissot and G. Bartholini, 1982, "/-Aminobutyric (GABA) receptor stimulation. III. Effect of progabide (SL 76002) on norepinephrine, dopamine and 5-hydroxy- tryptamine turnover in rat brain areas, J. Pharmacol. Exp. Ther. 220, 678.
Stephens, D.N., B.S. Meldrum, R. Weidmann, C. Schneider and M. Griatzner, 1986, Does the excitatory amino acid receptor antagonist 2-APH exhibit anxiolytic activity, Psy- chopharmacology 90, 166.
Stutzmann, J,M., M. Lucas, J.C. Blanchard and P.M. Laduron, 1988, Riluzole, a glutamate antagonist, enhances slow wave and REM sleep in rats, Neurosci. Lett. 88, 195.
Tam, S.Y. and R.H. Roth, 1985, Selective increase in dopamine
metabolism in the prefrontal cortex by the anxiogenic beta-carboline FG 7142, Biochem. Pharmacol. 34, 1595.
Thierry, A.M., J.P. Tassin, G. Blanc and J. Glowinski, 1976, Selective activation of the mesocortical DA system by stress, Nature (London) 263, 242.
Turski, L., T, Klockgether, K.H. Sontag, P.L Herrling and J.C. Watkins, 1987, Muscle relaxant and anticonvulsant activity of 3-( +_ )-2-carboxypiperazin-4-yl)propyl-1-phosphonic acid, a novel N-methyl-D-aspartate antagonist, in rodents, Neu- rosci. Lett. 73, 143.
Walaas, I. and F. Fonnum, 1979, The effects of surgical and chemical lesions on neurotransmitter candidates in the nucleus accumbens of the rat, Neuroscience 4, 209.
Walaas, I. and F. Fonnum, 1980, Biochemical evidence for 7-aminobutyrate containing fibers from the nucleus ac- cumbens to the substantia nigra and ventral tegmental area in the rat, Neuroscience 5, 63.
Watanabe, H., 1984, Activation of dopamine synthesis in mesolimbic dopamine neurons by immobilization stress in the rat, Neuropharmacology 23, 1335.
Watkins, ].C. and R.H. Evans, 1981, Excitatory amino acid transmitters, Ann. Rev. Pharmacol. Toxicol. 21, 165.
Watkins, J.C. and H.J. Olverman, 1987, Agonists and antagonists for excitatory amino acid receptors, Trends Neurosci. 10, 265.
White, H.S., A.S. Bender and E.A. Swinyard, 1988, Effect ot the selective N-methyI-D-aspartate receptor antagonist 3- (2-carboxypiperazin-4-yl)propyl-l-phosphonic acid on [3H]flunitrazepam binding, European J. Pharmacol. 147, 149.