newphy2010 185 701 ca rlk1 superoxide cell death pathogen 이동주
TRANSCRIPT

A novel pepper (Capsicum annuum) receptor-like kinasefunctions as a negative regulator of plant cell death viaaccumulation of superoxide anions
So Y. Yi1*, Dong J. Lee2,3*, Seon-In Yeom2, Joonseon Yoon2, Yun-Hee Kim1, Suk-Yoon Kwon1 and
Doil Choi2,3
1Plant System Engineering Research Center, KRIBB, Daejon 305-600, Korea; 2Department of Plant Science, Plant Genomics and Breeding Institute,
Seoul National University, Seoul, 151-921, Republic of Korea; 3Research Institute for Agriculture and Life Sciences, Seoul National University,
Seoul 151-921, Korea
Author for correspondence:Doil Choi
Tel: +82 2 880 4568
Email: [email protected]
Received: 16 July 2009
Accepted: 22 September 2009
New Phytologist (2010) 185: 701–715doi: 10.1111/j.1469-8137.2009.03095.x
Key words: autophosphorylation, Capsicum
annuum receptor-like kinase, cell death,ectopic expression, lesion stimulating diseasegene, NADPH oxidase, receptor-like kinase,superoxide anion.
Summary
• Plant receptor-like kinases belong to a large gene family. The Capsicum annuum
receptor-like kinase 1 (CaRLK1) gene encodes a transmembrane protein with a
cytoplasmic kinase domain and an extracellular domain.
• The CaRLK1 extracellular domain (ECD)–green fluorescent protein (GFP) fusion
protein was targeted to the plasma membrane, and the kinase domain of the
CaRLK1 protein exhibited autophosphorylation activity. CaRLK1 transcripts were
more strongly induced in treatment with Xag8ra than in treatment with Xag8-13.
Furthermore, infection with incompatible Xanthomonas campestris pv. vesicatoria
race 3 induced expression of CaRLK1 more strongly than in the compatible
interaction.
• Cell death caused by both a disease-forming and an HR-inducing pathogen
was delayed in the CaRLK1-transgenic plants. Ectopic expression of CaRLK1 also
induced transcripts of the lesion stimulating disease (LSD) gene, a negative regula-
tor of cell death. Respiratory burst oxidase homolog (RBOH) genes were up-regu-
lated in the transgenic plants compared with the wild type, as the concentration of
the superoxide anion was increased. In contrast, the concentration of H2O2 did
not differ between the transgenic and wild-type plants.
• These results support the theory that the suppression of plant cell death by
CaRLK1 is associated with consistent production of the superoxide anion and
induction of the RBOH genes and the LSD gene, but not with the concentration of
H2O2. Thus, CaRLK1 may be a receptor of an as yet unidentified pathogen molec-
ular pattern and may function as a negative regulator of plant cell death.
Introduction
The plant receptor-like protein kinase (RLK) genes consti-tute a large gene family in plants, with more than 600 and1000 genes in the Arabidopsis thaliana and rice (Oryza sati-va) genomes, respectively (Shiu et al., 2004). However, inanimals there are only a few receptor tyrosine kinases, which
are similar to the plant RLK genes. Recently, Oh et al.(2009) reported that recombinant cytoplasmic domains ofbrassinosteroid insensitive 1 (BRI1) and its co-receptorBRI1-associated kinase 1 (BAK1) autophosphorylated ontyrosine residues, and a specific tyrosine residue has beenshown to play an important role in vivo in plant receptorkinase function. In plants, RLKs play important roles ingrowth (Clark et al., 1997), development (Matsubayashiet al., 2002), cell wall biosynthesis (Mizuno et al., 2007)and defense responses (Gomez-Gomez & Boller, 2000;
*These authors contributed equally to this work.
GenBank accession number of CaRLK1: EF397556.
NewPhytologist Research
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715 701www.newphytologist.org

Heese et al., 2007). They are defined by the presence of asignal sequence, an amino-terminal domain with a trans-membrane region, and a carboxyl-terminal kinase domain(Torii, 2000; Shiu et al., 2004). The kinase domains ofdifferent RLKs share a high degree of homology (more than45% identity at the amino acid level), whereas their extracel-lular domains are much more divergent. Although the func-tions of most plant RLKs have not been determined, severalmembers of the RLK family have been implicated in con-trolling disease resistance or in interacting with proteins ofmicrobial origin (Zipfel et al., 2006). These genes, contain-ing membrane-bounded and cytoplasmic protein kinasedomains, include rice Xa21, which confers resistance toXanthomonas oryzae pv. oryzae race 6 (Song et al., 1995),wheat (Triticum aestivum) LRK10, a new receptor-likekinase gene encoded at the Lr10 disease resistance locus ofwheat (Feuillet et al., 1997), A. thaliana somatic embryo-genesis receptor kinase 3 (SERK3) ⁄ brassinosteroid-associatedkinase 1 (BAK1) (Heese et al., 2007) and A. thalianaFLAGELLIN-SENSITIVE 2 (FLS2) (Gomez-Gomez &Boller, 2000). For example, SERK3 ⁄ BAK1 rapidly entersan elicitor–dept complex with FLS2 and its peptide deriva-tive flg22 (a peptide spanning a single stretch of 22 aminoacid residues of the most conserved part in the N terminusof flagellin) in A. thaliana (Heese et al., 2007).
A rapid increase in reactive oxygen species (ROS) is ahallmark of the early events in interactions between plantsand pathogens. Plasma membrane-bound NADPH oxidas-es play a significant role in the generation of ROS, particu-larly the superoxide anion (O2
)) (Simon-Plas et al., 2002),which is responsible for a one-electron reduction of oxygenmolecules. The NADPH oxidase gene was identified inphagocytic cells (neutrophils and macrophages), which areresponsible for controlling invasive pathogens (Vulcanoet al., 2004). Plant NADPH oxidases consist of one compo-nent of c. 105 kDa. NADPH oxidase homologs comprisemultigene families in rice, tomato (Solanum lycopersicum),and A. thaliana. The first identified plant NADPH oxidasegenes were the A. thaliana respiratory burst oxidase homo-log (Atrboh) genes (Torres et al., 1998). Arabidopsis thalianahas 10 Atrboh genes (Torres & Dangl, 2005). AtrbohD andAtrbohF are known to produce superoxide following patho-gen invasion (Torres et al., 2002). Recently, Miller et al.(2009) reported that O2
) generated by AtRBOHD plays animportant role in mediating cell-to-cell communication(rapid systemic signaling) over long distances in the plant.NbrbohA and NbrbohB in Nicotiana benthamiana are alsorequired for H2O2 accumulation and for resistance to Phy-tophthora (Yoshioka et al., 2003).
Superoxide dismutase (SOD) catalyzes the dismutationof O2
) to H2O2. There are three classes of SOD in plants:FeSOD (FSD), MnSOD (MSD), and CuZnSOD (CSD)(Conklin & Last, 1995). The lesion stimulating disease 1(LSD1) gene is a part of a signaling pathway for the induc-
tion of the CSD protein (Kliebenstein et al., 1999). Gluta-thione S-transferases (GSTs) (EC 2.5.1.18) are a family ofmultifunctional enzymes that catalyze the conjugation ofreduced glutathione to a range of toxic compounds todetoxify xenobiotics and protect plant tissues against oxida-tive damage (Conklin & Last, 1995). Thus, their expressionis induced by a range of stimuli, such as pathogen attackand oxidative stress. The expression of A. thaliana GST1was found to be strongly induced by ozone treatment(Sharma & Davis, 1997).
In this study, we have identified and functionally charac-terized the Capsicum annuum (pepper) receptor-like kinase1 (CaRLK1) gene. The CaRLK1 gene was induced by treat-ment with pathogen, H2O2, and salicylic acid (SA). Wedetermined the role of CaRLK1 as a critical component ofthe regulation of O2
) production through induction of theRBOH genes.
Materials and Methods
Plant growth and bacterial infection
Plants were grown under standard greenhouse, long-dayconditions with a relative air humidity of 50–60% at25�C. For pepper inoculation, 2 · 108 colony-formingunits (cfu) ml)1 solutions of Xanthomonas axonopodis pv.glycines (Xag) and Xanthomonas campestris pv. vesicatoriarace 3 (Xcv) were pressure-infiltrated into leaf tissuesusing a needleless syringe (Lee et al., 2002; Kim & Kim,2009). Bacterial populations were measured as describedin Thilmony et al. (1995) and Thara et al. (1999). Threeleaf discs of area 0.785 cm2 per plant were collected afterinoculation by syringe infiltration of a serially dilutedsuspension. Bacterial populations were counted after leafdiscs had been ground in 10 mM MgCl2 and serial dilu-tions of bacterial solution were plated on LB agar platescontaining 100 mg l)1 rifampicin. The average numberof bacterial cells is presented as cfu cm)2 of inoculatedleaves.
RNA gel blots analysis and RT-PCR
For northern blot analyses, total RNA was separated onformaldehyde-containing agarose gels and blotted ontonylon membranes following standard procedures (Sam-brook et al., 1989). Blots were hybridized with [a-32P]dCTP-labeled probes. A CaRLK1-specific probe was gener-ated via PCR amplification with the gene-specific primersCaRLK1proLP and CaRLK1proRP. The pepper cDNAprobe for pathogenesis-related 4 (PR4) was isolated previ-ously from pepper (Lee et al., 2002). For RT-PCR, first-strand cDNA was synthesized from 2 lg of total RNA andused for amplification by PCR. Oligonucleotide primerswere made according to the published primer sequences.
702 Research
NewPhytologist
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715
www.newphytologist.org

The nucleotide sequences of other primers are listed in Sup-porting Information Table S1.
Subcellular localization of CaRLK1
Two constructs, CaRLK1-extracellular domain (ECD):smGFP and CaRLK1-ECD-SP:smGFP, were made forCaRLK1 expression in pepper protoplasts. Two primers,RLK(ECD)LP and RLK(ECD)RP, were used to amplifyCaRLK1-ECD:smGFP (amino acids 1-219), andRLK(ECD-SP)LP and RLK(ECD)RP primers were usedto amplify CaRLK1-ECD-SP:smGFP (amino acids19-219, without the putative signal peptide (SP)). Eachconstruct was fused in-frame to the N-terminal region ofthe green fluorescent protein (GFP) under the control ofthe cauliflower mosaic virus (CaMV) 35S promoter andintroduced into pepper protoplasts prepared from youngleaves by polyethylene glycol-mediated transformation(Lee et al., 2001). A DNA construct encoding H+-AT-Pase:RFP, in which H+-ATPase was fused to the C-terminus of RFP (red fluorescence protein), under thecontrol of the CaMV 35S promoter was simultaneouslyintroduced into protoplasts. Expression of the fusion pro-teins was monitored at 40 h after transformation by fluo-rescence microscopy using a Zeiss Axioplan fluorescencemicroscope (Jena, Carl Zeiss, Berlin, Germany), andimages were captured with a cooled charge-coupled devicecamera. The filter sets were used as described previously(Lee et al., 2001). The data were processed using AdobePhotoshop software (Mountain View, CA, USA) and pre-sented in pseudocolor format. The experiments wererepeated independently three times with similar results.
Recombinant protein and in vitro phosphorylationassay
The coding region of truncated CaRLK1 was cloned intopRSET-A (Invitrogen, Carlsbad, CA, USA) and exp-ressed in Escherichia coli BL21 cells (Amersham Pharma-cia Biotech, Uppsala, Sweden). Two primers (RLK-KDLP and RLK-KD RP) were used to amplify the CaRLK1kinase domain (CaRLK1-KD; amino acids 306-598).The His-tag fusion protein CaRLK1-KD was purifiedusing Ni-NTA agarose resin, according to the manufac-turer’s instructions (Invitrogen). Phosphorylation was per-formed in a 25-ll assay buffer (50 mM Tris-HCl, pH7.6, 50 mM KCl, 2 mM DTT and 10% (v ⁄ v) glycerol)containing 0.5–1 lg of purified CaRLK1-KD in thepresence of 5 mM MnCl2. Phosphorylation was initiatedby adding 25 lCi of [c-32P] ATP (30 Ci mmol)1), andthe reaction was incubated at 22�C for 60 min and ter-minated by the addition of EDTA to a final concentra-tion of 10 mM. The phosphorylated proteins were
subjected to 10% SDS-PAGE and transferred onto PVDFmembranes (Amersham Pharmacia Biotech). The incorpo-rated phosphate was visualized by autoradiography.Coomassie blue staining of the same membrane was alsoperformed to verify protein loading. The stability of theincorporated phosphate was determined by treating themembranes with water, 1 M HCl or 3 M NaOH for 2 hat room temperature. The treated membranes were thensubjected to autoradiography. The experiments wererepeated independently three times with similar results.
Transformation of N. benthamiana
The pMBP-1:CaRLK1 construct was introduced into Agro-bacterium tumefaciens (GV2260) for plant transformationaccording to Lee & Zeevaart (2005). Leaf discs were incu-bated with A. tumefaciens for 10 min and then transferredto Murashige and Skoog (MS) medium containing1 mg l)1 6-benzylaminopurine (BA) and 0.1 mg l)1 1-naphthaleneacetic acid (NAA) in darkness. After co-culturefor 3 d, leaf discs were transferred to selection medium(MS, 1 mg l)1 BA, 0.1 mg l)1 NAA, 100 mg l)1 kanamy-cin, and 500 mg l)1 carbenicillin). More than 30 individualshoots were cut and transferred to root-inducing medium(MS, 0.1 mg l)1 NAA, 100 mg l)1 kanamycin, and500 mg l)1 carbenicillin). Plantlets (T1) were planted insoil and grown in a greenhouse. The T1 generation was self-pollinated to obtain the T2 generation. Independent trans-genic lines were selected on half-strength MS containing50 mg l)1 kanamycin. All T1 lines showing 3 : 1 segrega-tion (resistant:sensitive) were selected to obtain homozygousT3 plants. Line 3 from these homozygous lines was chosenas a representative line.
Histochemical detection of cell death
Plant cell death was detected by modified lactophenol-trypan blue staining as described by Koch & Slusarenko(1990). The stock solution of lactophenol-trypan bluecontained 10 ml of lactic acid, 10 ml of glycerol, 10 gof phenol, and 10 mg of trypan blue dissolved in 10 mlof deionized distilled water. The working solution wasprepared by diluting the stock solution with ethanol(1 : 2 v ⁄ v). Leaf discs (1 cm in diameter) were stainedwith lactophenol-trypan blue working solution and wereplaced in a heated water bath and boiled for 4 min.The tissues were left overnight, and then destained withchlorohydrate solution (2.5 g of chloral hydrate dissolvedin 1 ml of distilled water). They were mounted in 50%glycerol and photographed under a light microscope (Si-won Optical Technology, Suwon, Republic of Korea).The experiments were repeated independently threetimes with similar results.
NewPhytologist Research 703
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715
www.newphytologist.org

Measurement of ion leakage
Cell death was also assayed by measuring ion leakage fromleaf discs (Liu et al., 2007). Two days after inoculation withPseudomonas syringae pv. tabaci, six leaf discs (1 cm diame-ter) per plant were floated on 6 ml of distilled water for 1 hat room temperature. After incubation, treated tubes weremeasured for electrical conductivity (Model 455C; Istek,Seoul, Republic of Korea). The experiments were repeatedindependently three times with similar results.
Detection of O2) and H2O2
Nitroblue tetrazolium (NBT) staining was performed onfully expanded leaves by vacuum-infiltrating NBT solution,containing 1 mM NBT plus 10 mM sodium azide in50 mM sodium phosphate buffer (pH 7.6) (Achard et al.,2008). Leaves were placed in boiling ethanol for 10 min toremove the chlorophyll and to visualize reduced NBT dye.Use of the luminol-like compound L-012 (8-amino-5-chloro-7-phenylpyridol[3,4-d]pyridazine-1,4(2H,3H)dione)to induce luminescence of reactions is a highly sensitivemeans of measuring superoxide and nitric oxide (NO)production (Imada et al., 1999; Dyke et al., 2007). AnL-012 solution containing 100 lM L-012 in 10 mMsodium phosphate buffer (pH 7.5) was infiltrated intoleaves using a needleless syringe. Diphenylene iodonium(DPI) solution (100 lM) in 50 mM sodium phosphate(pH 7.6) was infiltrated 1 h before injection of L-012(Achard et al., 2008). Intact leaves were removed 30 minafter injection and photographed using the MultiDoc-itDigital Imaging System (UVP, Upland, CA, USA).
We stained peroxides in situ in fully expanded leaves orleaf discs (1 cm diameter) of 6-wk-old plants with 3,3¢-diaminobenzidine tetrahydrochloride (DAB). Fully expan-ded leaves or leaf discs were vacuum-infiltrated with DABsolution (0.1 mg ml)1 DAB in 50 mM Tris acetate buffer,pH 5.0) for 10 min and then placed overnight in darknessat 25�C. Green pigments were cleared in 80% (v ⁄ v) ethanolfor 10 min at 70�C. Control staining was performed in thepresence of 5 mM ascorbic acid. The experiments wererepeated independently three times with similar results.
Superoxide anion-generating activity in leaf discs
Leaf discs of 4 mm diameter were washed thoroughly in50 mM Tris buffer (pH 7.4) for 1 h, blotted dry, and incu-bated at 25�C for 1 h in 50 mM Tris buffer (pH 7.4). Theassay solution consisted of 0.3 mM XTT (2,3-bis(2-meth-oxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilideinner salt) and 2 mM NADPH in 50 mM Tris buffer (pH7.4), containing 5 lM DPI (Achard et al., 2008). The A450
was measured and expressed in micromoles of superoxidegenerated per minute using the molar extinction coefficient
for the XTT formazan product of 21 600 M)1 cm)1 (John-son et al., 2003; Achard et al., 2008). The experiments wererepeated independently three times with similar results.
Enzyme assay for GST and quantification of protein
Mature leaves were harvested, ground with a pestle andmortar in liquid nitrogen and suspended in grinding buffer(50 mM sodium phosphate buffer, pH 7.0, and 0.5% Tri-ton X-100) at 4�C and centrifuged at 13 000g for 5 min.The aqueous phase was used as an enzyme solution. Spec-trophotometric measurements at 340 nm were performedat 25�C in 50 mM potassium phosphate buffer (pH 6.5),containing 5 mM reduced glutathione (GSH) plus 1 mM1-chloro-2,4-dinitrobenzene (CDNB). The specific activityof GST was calculated with an extinction coefficient of9.6 mM)1 cm)1 (Ricci et al., 1984; Ali et al., 2005). Pro-tein assays were carried out using Quick StartTM BradfordDye Reagent (Bio-Rad, Hercules, CA, USA). The experi-ments were repeated independently three times with similarresults.
Results
Isolation of CaRLK1 cDNA
To isolate full-length cDNA, a partial cDNA fragmentobtained from differential display RT-PCR with sequencehomology to the serine ⁄ threonine kinase protein was usedas a probe to screen a cDNA library previously constructedfrom C. annuum (Yi et al., 2006). Twenty-two positiveclones were isolated and further analyzed by restrictionenzyme digestion and DNA sequencing, resulting in theidentification of six clones with > 2.5-kb cDNA inserts.Five of these clones were predicted to encode full-lengthproteins with an open reading frame of 627 aminoacids and a calculated molecular mass of 69 kDa (Fig. 1a).(Pfam is designed to be a comprehensive and accurate col-lection of protein domains and families.) Analyses of thestructural properties of the predicted protein using the Pfamdatabase (Bateman et al., 2004) indicated that this cDNAencodes a putative receptor-like kinase (CaRLK1) with fourdistinctive regions: an N-terminal hydrophobic signal pep-tide (SP), an extracellular domain (ECD), a transmembranedomain (TM), and a cytosolic kinase domain (KD)(Fig. 1b).
CaRLK1 is a putative novel receptor-like kinase
Plant RLKs are classified into multiple subfamilies based ontheir internal kinase domain (IKD) and ECDs. A large num-ber of RLKs do not have any established sequence motifs intheir ECDs (Shiu & Bleecker, 2001; Shiu et al., 2004). TheECD of CaRLK1 has low homology to those of other known
704 Research
NewPhytologist
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715
www.newphytologist.org

plant RLKs (Braun & Walker, 1996; Shiu et al., 2004) orany other proteins in the protein databases. CaRLK1 con-tains 27 leucine residues in its ECD (Fig. 1) which cannotbe organized as a leucine-rich repeat consensus as definedby Kajava (1998), indicating that it contains a new typeof ECD. Two hypothetical proteins from Vitis vinifera(CAN81685 and CAV65965) show the highest homology tothe ECD of CaRLK1 (c. 27% identity and 45% similarity),and one of four wall-associated kinases in the LRK10L-1asubfamily, At1g25390, also shows 45% similarity (Wagner& Kohorn, 2001; Shiu et al., 2004; Lehti-Shiu et al., 2009).Thus, the CaRLK1 gene belongs to the LRK10L-1a sub-family (Shiu et al., 2004; Lehti-Shiu et al., 2009). Twohypothetical proteins from V. vinifera (CAN69599 andCAN76446) show 70% identity with the IKD (amino acids250-627) of CaRLK1. Interestingly, the IKDs of the putativerust resistance kinase Lr10 (BAB17345) from O. sativa andof an RLK from A. thaliana (At1g70250) show 75% similar-ity to the IKD of CaRLK1 (Fig. S2).
Expression of CaRLK1 is induced by pathogen
We examined whether expression of CaRLK1 mRNA isinduced upon pathogen attack in pepper (C. annuum) cv.Bukang. Bukang is a commercial cultivar, and was usedto produce C. annuum expressed sequence tags (http://genepool.kribb.re.kr/new/index.php). Inoculation with the
nonhost pathogen Xag8ra can induce the leaf hypersensitiveresponse (HR) in hot pepper, but inoculation with Xag8-13can not. The expression of CaRLK1 was induced at 4 h postinoculation (hpi) and sustained in both Xag8ra-treated andXag8-13-treated leaves (Fig. 2a). However, the expression ofCaRLK1 was much stronger and of longer duration in bothXag8ra-treated and Xag8-13-treated leaves (Fig. 2a). As apositive control, the expression of PR4 (Park et al., 2001)was not induced in a mock treatment. However, it was firstinduced at 4 hpi with Xag8ra or Xag8-13 and stronglyinduced at 8–12 hpi.
To evaluate the specificity of CaRLK1 expression, leavesof pepper cultivars ECW-20R (Bs2/Bs2) and ECW (bs2/bs2) were syringe-infiltrated with the pepper bacterial spotpathogen Xanthomonas campestris pv. vesicatoria (Xcv) race3,which expresses the avrBs2 gene. The Bs2 gene specificallyconfers resistance to Xcv that contain the avirulence gene,avrBs2. The compatible host (cv. ECW) did not exhibit anyvisible responses until 24 hpi. In contrast, the incompatiblehost (cv. ECW-20R) developed the HR within 24 hpi (datanot shown). In incompatible interactions, the amount ofCaRLK mRNA detected was significantly increased at 4 hpiand remained at a high level for 36 h, whereas in compati-ble interactions the increase in the amount of CaRLK tran-script was less and was not sustained to the same extentfrom 4 to 24 hpi (Fig. 2b). As expected, the PR4 transcriptwas strongly up-regulated in ECW-20R (BS2 ⁄ BS2) but notin ECW (bs2 ⁄ bs2) (Fig. 2b).
CaRLK1
Mock(a)
(b)
Xag8-13 Xag8ra
(h)
CaPR4
rRNA
CaRLK1
0 4 8 12 0 4 8 12 0 4 8 12
0 4 8 12 24 36 0 4 8 12 24 36
Xcv race 3 (avr Bs2)
(h)
ECW20R (Bs2/Bs2) ECW(bs2/bs2)
CaPR4
rRNA
Fig. 2 Expression of Capsicum annuum receptor-like kinase 1(CaRLK1) mRNA in response to bacterial pathogens. (a) Pepperleaves were infiltrated with either a mock solution (10 mM) or witha solution containing Xanthomonas axonopodis pv. glycines 8ra(nonhost pathogen) and Xanthomonas axonopodis pv. glycines
8-13. (b) Pepper near-isogenic lines (Capsicum annuum ECW-20R,resistant; C. annuum ECW, susceptible) were infiltrated with a pep-per leaf spot pathogen, as described in the Materials and Methods.One blot was hybridized to the CaRLK1 probe, while an identicalblot was hybridized to the PR4b probe as a positive marker for path-ogen infection. Similar results were obtained in three independentexperiments. One representative experiment is shown.
ECD TMD KINASE DOMAINNH2 COOHSP
1 18 249 268 627
(a)
(b)
Fig. 1 Identification and sequence analysis of Capsicum annuumreceptor-like kinase 1 (CaRLK1). (a) Predicted amino acid sequenceof CaRLK1. The signal peptide sequences and the transmembraneregion are denoted by bold letters. Roman numerals indicate the 11characteristic subdomains of protein kinases as defined by Hanks &Quinn (1991). (b) A linear schematic representation of CaRLK1showing the four domains. SP, signal peptide; ECD, extracellulardomain; TMD, transmembrane domain.
NewPhytologist Research 705
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715
www.newphytologist.org
Effect of exogenous H2O2 on expression of CaRLK1
In an effort to elucidate the relationship between oxidativestress signals and the in vivo function of CaRLK1, hot pep-per plants were treated with H2O2 at various time intervalsand at various concentrations. CaRLK1 expression wasinduced 2 h after treatment with 0.25–2 mM H2O2
(Fig. 3a). In addition, expression of CaRLK1 was rapidlyinduced as early as 1 h after treatment and remained at ahigh level until 12 h after treatment with 1 mM H2O2
(Fig. 3b), indicating that the expression of CaRLK1 isregulated by H2O2, and that CaRLK1 expression may beinvolved in oxidative stress signaling or defense signaling.
Subcellular localization of CaRLK1
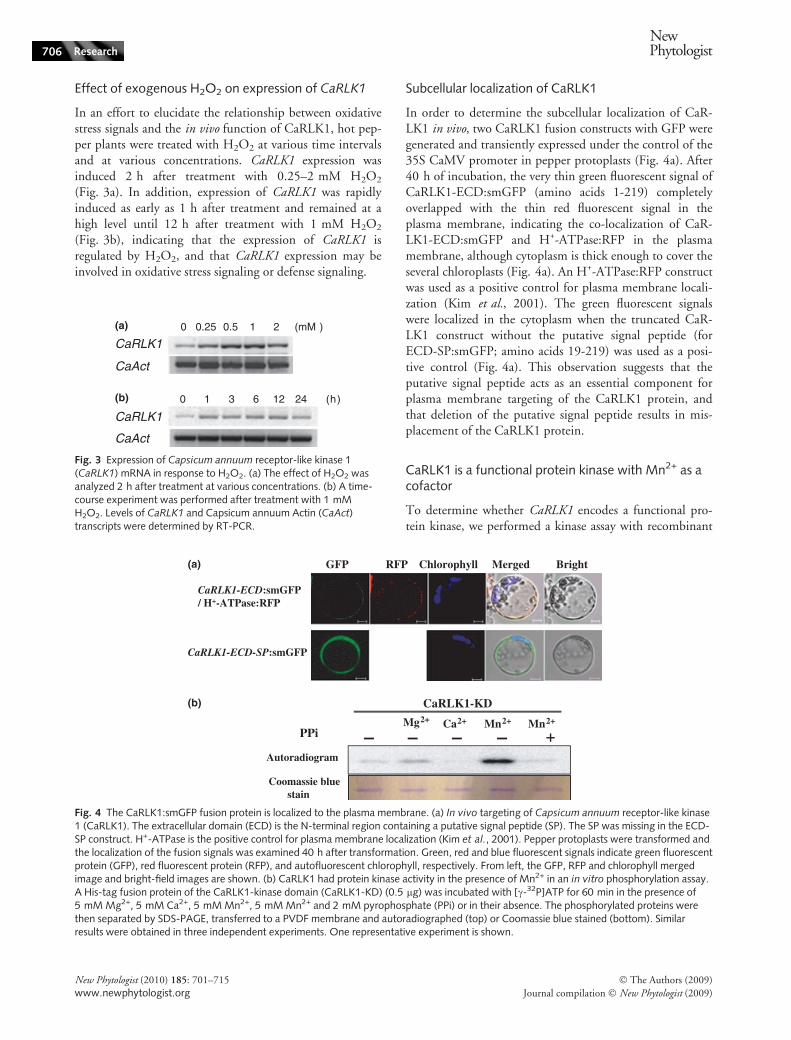
In order to determine the subcellular localization of CaR-LK1 in vivo, two CaRLK1 fusion constructs with GFP weregenerated and transiently expressed under the control of the35S CaMV promoter in pepper protoplasts (Fig. 4a). After40 h of incubation, the very thin green fluorescent signal ofCaRLK1-ECD:smGFP (amino acids 1-219) completelyoverlapped with the thin red fluorescent signal in theplasma membrane, indicating the co-localization of CaR-LK1-ECD:smGFP and H+-ATPase:RFP in the plasmamembrane, although cytoplasm is thick enough to cover theseveral chloroplasts (Fig. 4a). An H+-ATPase:RFP constructwas used as a positive control for plasma membrane locali-zation (Kim et al., 2001). The green fluorescent signalswere localized in the cytoplasm when the truncated CaR-LK1 construct without the putative signal peptide (forECD-SP:smGFP; amino acids 19-219) was used as a posi-tive control (Fig. 4a). This observation suggests that theputative signal peptide acts as an essential component forplasma membrane targeting of the CaRLK1 protein, andthat deletion of the putative signal peptide results in mis-placement of the CaRLK1 protein.
CaRLK1 is a functional protein kinase with Mn2+ as acofactor
To determine whether CaRLK1 encodes a functional pro-tein kinase, we performed a kinase assay with recombinant
CaAct
CaRLK10(a)
(b)
0.25 0.5 1 2 (mM )
0 1 3 6 12 24 (h)
CaAct
CaRLK1
Fig. 3 Expression of Capsicum annuum receptor-like kinase 1(CaRLK1) mRNA in response to H2O2. (a) The effect of H2O2 wasanalyzed 2 h after treatment at various concentrations. (b) A time-course experiment was performed after treatment with 1 mMH2O2. Levels of CaRLK1 and Capsicum annuum Actin (CaAct)transcripts were determined by RT-PCR.
GFP(a)
(b)
RFP BrightMergedChlorophyll
CaRLK1-ECD:smGFP/ H+-ATPase:RFP
CaRLK1-ECD-SP:smGFP
CaRLK1-KD
Autoradiogram
Mg2+ Ca2+ Mn2+ Mn2+
PPi – – – – ++
Coomassie bluestain
Fig. 4 The CaRLK1:smGFP fusion protein is localized to the plasma membrane. (a) In vivo targeting of Capsicum annuum receptor-like kinase1 (CaRLK1). The extracellular domain (ECD) is the N-terminal region containing a putative signal peptide (SP). The SP was missing in the ECD-SP construct. H+-ATPase is the positive control for plasma membrane localization (Kim et al., 2001). Pepper protoplasts were transformed andthe localization of the fusion signals was examined 40 h after transformation. Green, red and blue fluorescent signals indicate green fluorescentprotein (GFP), red fluorescent protein (RFP), and autofluorescent chlorophyll, respectively. From left, the GFP, RFP and chlorophyll mergedimage and bright-field images are shown. (b) CaRLK1 had protein kinase activity in the presence of Mn2+ in an in vitro phosphorylation assay.A His-tag fusion protein of the CaRLK1-kinase domain (CaRLK1-KD) (0.5 lg) was incubated with [c-32P]ATP for 60 min in the presence of5 mM Mg2+, 5 mM Ca2+, 5 mM Mn2+, 5 mM Mn2+ and 2 mM pyrophosphate (PPi) or in their absence. The phosphorylated proteins werethen separated by SDS-PAGE, transferred to a PVDF membrane and autoradiographed (top) or Coomassie blue stained (bottom). Similarresults were obtained in three independent experiments. One representative experiment is shown.
706 Research
NewPhytologist
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715
www.newphytologist.org

CaRLK1 proteins expressed in E. coli. When the recombi-nant CaRLK1-KD (amino acids 306-598) was incubatedwith [c-32P]ATP, 32P was specifically incorporated into theprotein when Mn2+ was used as a cofactor. A lower level ofincorporation was observed in the presence of Ca2+ orMg2+. Some plant protein kinases, including tobacco pro-tein kinase, NPK5 (Muranaka et al., 1994), CBL-interact-ing protein kinase 1 (CIPK1) (Shi et al., 1999) and RLK5(Horn & Walker, 1994), are more active when Mn2+ isused as a cofactor. Sodium pyrophosphate, an inhibitor ofprotein kinase, abolished the kinase activity (Fig. 4b). Thesedata suggest that CaRLK1-KD encodes a functional proteinkinase.
Ectopic expression of CaRLK1 in N. benthamiana
To determine whether ectopic expression of CaRLK1 affectsthe plant defense response and cell death in another species,we introduced the 35S::CaRLK1 cDNA construct intoN. benthamiana by Agrobacterium tumefaciens-mediatedtransformation (Lee & Zeevaart, 2005). Eight homozygouslines from more than 30 lines expressing CaRLK1 werefinally selected for resistance against kanamycin (50 mg l)1),and did not show any morphological difference in pheno-type (data not shown). To confirm the presence of theCaRLK1 gene in the transgenic plants, we performed geno-mic PCR amplification with the primer set CaRLK1LP1354and CaRLK1RP1883 (Table S1). The PCR products(0.5 kb) were only detected without any size of other PCRproducts in all transgenic lines (data not shown). Transgenicline 3 (CaRLK1-3) was the main line chosen for furthercharacterization.
Effect of ectopic expression of CaRLK1 on plant celldeath
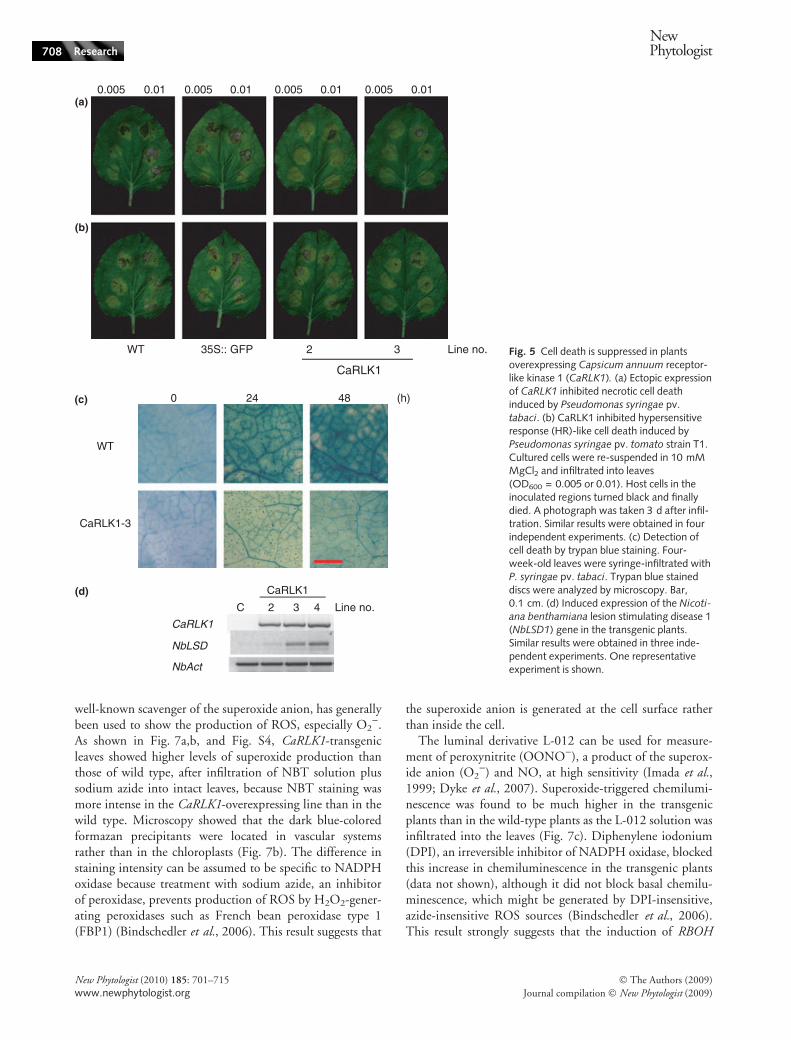
To investigate the possible role of CaRLK1 in inhibiting hostcell death, we challenged at least three fully grown leaves per5- to 6-wk-old plant with a virulent pathogen (P. syringae pv.tabaci strain 11528; As shown in Fig 5a, we used two kindsof concentration of cell, OD600 = 0.01 and 0.005), anddetected cell death using trypan blue exclusion (Fig. 5).Strikingly, at least four of six inoculated spots were turnedblack and finally died in wild-type plants and GFP over-expression plants as a transgenic control, but only one of sixinoculated spots were black to death in CaRLK1 overexpres-sion plants (Fig. 5a). When challenged with P. syringae pv.tomato T1 (OD600 = 0.01 and 0.005), WT and GFP over-expression plants also displayed the normal cell death pheno-type, but CaRLK1 overexpression plants showed only a weakcell death phenotype (Fig. 5b).
Cell death in wild-type leaves was characterized byintense staining with lactophenol-trypan blue (Fig. 5c).Host cell death in the CaRLK1 overexpression line was
found to a much lesser extent than in wild-type leaves atboth 24 and 48 h post inoculation of P. syringae pv. tabaci,indicating that inhibition of cell death is attributable to thespecific expression of CaRLK1 (Fig. 5c, Fig. S3).
LSD1 is well known to be an important factor in the reg-ulation of programmed cell death (PCD) in plants (Die-trich et al., 1997). The lsd1 mutant fails to restrict spread ofcell death under various conditions, such as pathogen infec-tion, strong light, and treatment with SA (Dietrich et al.,1997; Rusterucci et al., 2001). Expression of NbLSD1 tran-scripts was increased in the transgenic plants compared withwild type (Fig. 5d), indicating that constitutive expressionof LSD under the control of CaRLK1 could suppress thespread of cell death in the transgenic plant. The putativeamino acid sequences of two genes (BoLSD1 and BoLSD2)from Brassica oleracea show 85% similarity with that ofNbLSD1 (data not shown) (Coupe et al., 2004). Interest-ingly, the AtLSD1 protein shows 75% similarity withNbLSD1 (data not shown).
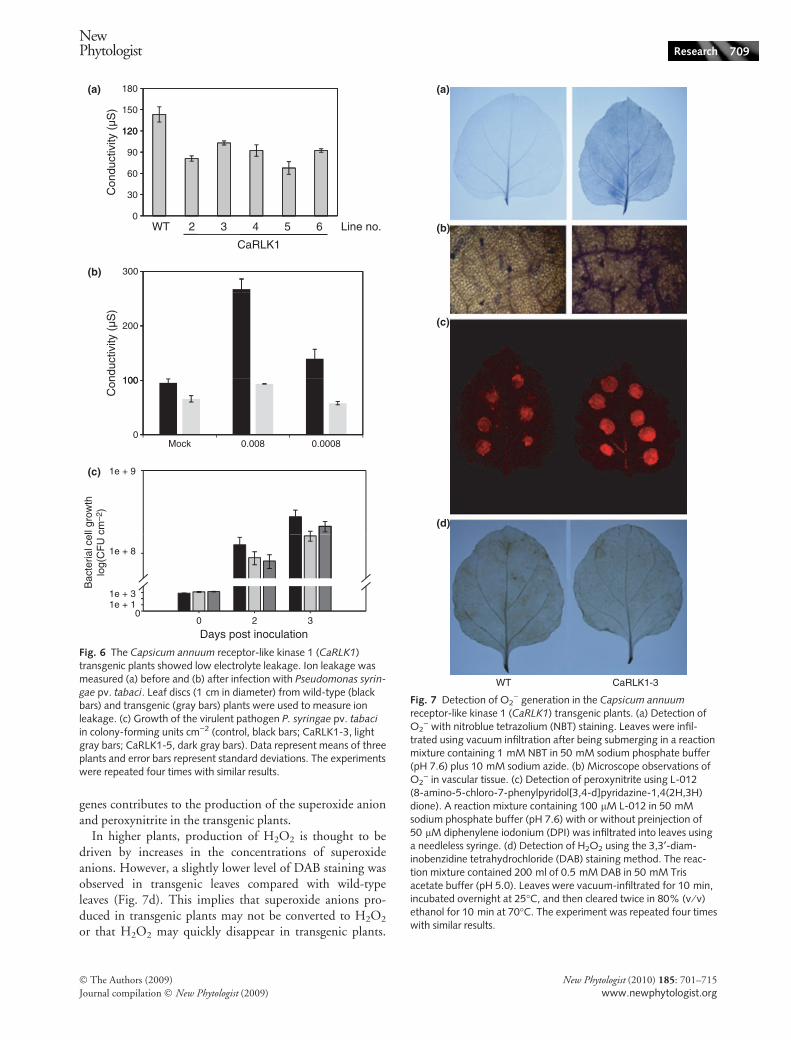
To confirm that cell death was suppressed in transgenicplants, we quantified electrolyte leakage. The conductivityof the wild type was always higher than that of CaRLK1overexpression plants in the absence of pathogen applica-tion, indicating that the transgenic plant may always be pre-paring for self-defense (Fig. 6a). This result was consistentwith observations of reduced electrolyte leakage after patho-gen inoculation; the conductivity of CaRLK1-overexpress-ing plants only changed moderately following infectionwith P. syringae pv. tabaci. However, that of the wild typeincreased threefold (Fig. 6b). This result also suggests thatCaRLK1 is involved in negative regulation of plant celldeath, as shown by lactophenol-trypan blue staining(Fig. 5c).
Interestingly, pathogen colonization was two or threetimes higher in the wild type than in the CaRLK1 over-expression plants at 2–3 d post inoculation (dpi) (Fig. 6c),although the difference was less than we expected. This resultsuggests that the transgenic plants have more tolerance toP. syringae pv. tabaci than wild-type plants because theydelay pathogen growth. In addition, expressions of patho-genesis-related genes (PR1s), systemic acquired resistance 8.2(SAR8.2) and nonexpressor of pathogenesis-related genes(NPR) was also induced in the transgenic plants (Fig. S1c).This suggests that CaRLK1 may be involved in plant basalresistance.
Effect of CaRLK1 on accumulation of the superoxideanion
It was of interest to determine whether the expression ofCaRLK1 is related to ROS, because CaRLK1 expression wasinduced c. 4 h after infiltration with pathogens (incompati-ble or compatible interaction) (Fig. 2a,b) and was quicklyinduced 1 h after treatment with H2O2 (Fig. 3b). NBT, a
NewPhytologist Research 707
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715
www.newphytologist.org
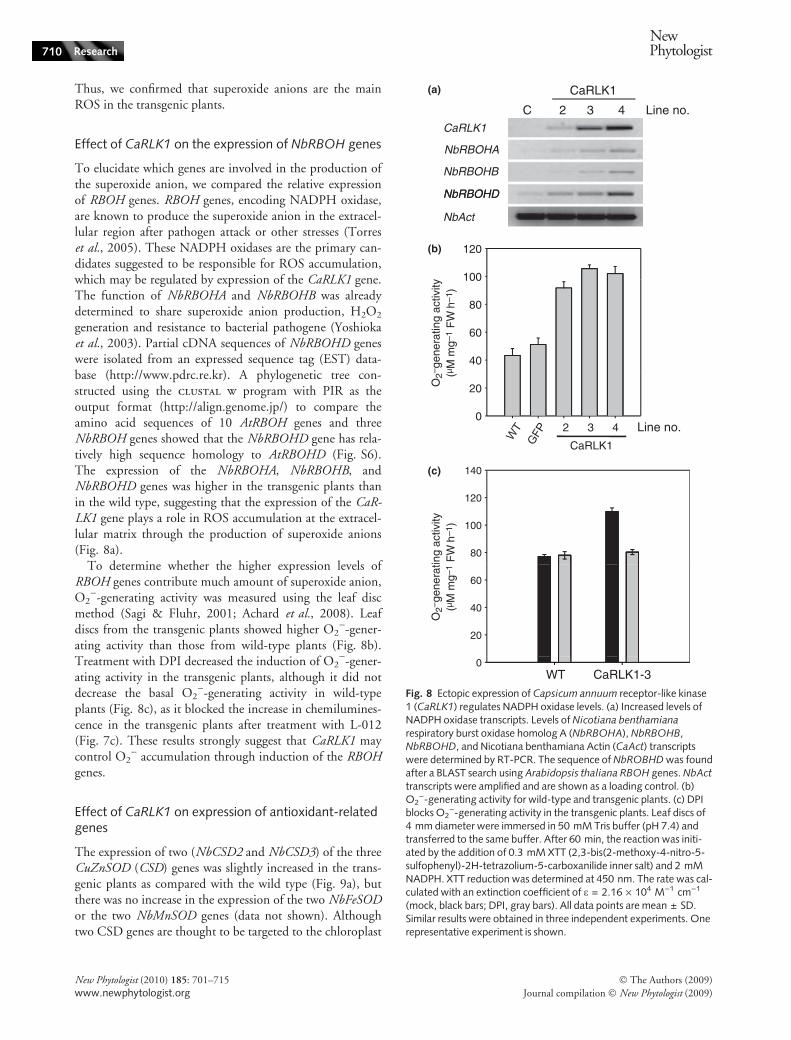
well-known scavenger of the superoxide anion, has generallybeen used to show the production of ROS, especially O2
).As shown in Fig. 7a,b, and Fig. S4, CaRLK1-transgenicleaves showed higher levels of superoxide production thanthose of wild type, after infiltration of NBT solution plussodium azide into intact leaves, because NBT staining wasmore intense in the CaRLK1-overexpressing line than in thewild type. Microscopy showed that the dark blue-coloredformazan precipitants were located in vascular systemsrather than in the chloroplasts (Fig. 7b). The difference instaining intensity can be assumed to be specific to NADPHoxidase because treatment with sodium azide, an inhibitorof peroxidase, prevents production of ROS by H2O2-gener-ating peroxidases such as French bean peroxidase type 1(FBP1) (Bindschedler et al., 2006). This result suggests that
the superoxide anion is generated at the cell surface ratherthan inside the cell.
The luminal derivative L-012 can be used for measure-ment of peroxynitrite (OONO)), a product of the superox-ide anion (O2
)) and NO, at high sensitivity (Imada et al.,1999; Dyke et al., 2007). Superoxide-triggered chemilumi-nescence was found to be much higher in the transgenicplants than in the wild-type plants as the L-012 solution wasinfiltrated into the leaves (Fig. 7c). Diphenylene iodonium(DPI), an irreversible inhibitor of NADPH oxidase, blockedthis increase in chemiluminescence in the transgenic plants(data not shown), although it did not block basal chemilu-minescence, which might be generated by DPI-insensitive,azide-insensitive ROS sources (Bindschedler et al., 2006).This result strongly suggests that the induction of RBOH
CaRLK1
WT 35S:: GFP 2 3 Line no.
WT
CaRLK1-3
0.005(a)
(b)
(c)
(d)
0.01 0.005 0.01 0.005 0.01 0.005 0.01
0 24 48 (h)
C 2 3 4 Line no.
CaRLK1
CaRLK1
NbLSD
NbAct
Fig. 5 Cell death is suppressed in plantsoverexpressing Capsicum annuum receptor-like kinase 1 (CaRLK1). (a) Ectopic expressionof CaRLK1 inhibited necrotic cell deathinduced by Pseudomonas syringae pv.tabaci. (b) CaRLK1 inhibited hypersensitiveresponse (HR)-like cell death induced byPseudomonas syringae pv. tomato strain T1.Cultured cells were re-suspended in 10 mMMgCl2 and infiltrated into leaves(OD600 = 0.005 or 0.01). Host cells in theinoculated regions turned black and finallydied. A photograph was taken 3 d after infil-tration. Similar results were obtained in fourindependent experiments. (c) Detection ofcell death by trypan blue staining. Four-week-old leaves were syringe-infiltrated withP. syringae pv. tabaci. Trypan blue staineddiscs were analyzed by microscopy. Bar,0.1 cm. (d) Induced expression of the Nicoti-ana benthamiana lesion stimulating disease 1(NbLSD1) gene in the transgenic plants.Similar results were obtained in three inde-pendent experiments. One representativeexperiment is shown.
708 Research
NewPhytologist
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715
www.newphytologist.org
genes contributes to the production of the superoxide anionand peroxynitrite in the transgenic plants.
In higher plants, production of H2O2 is thought to bedriven by increases in the concentrations of superoxideanions. However, a slightly lower level of DAB staining wasobserved in transgenic leaves compared with wild-typeleaves (Fig. 7d). This implies that superoxide anions pro-duced in transgenic plants may not be converted to H2O2
or that H2O2 may quickly disappear in transgenic plants.
120
150
180(a)
(b)
(c)
Con
duct
ivity
(µS
)
0
30
60
90
120
Line no.
CaRLK1
300
100
200
0
100
WT 2 3 4 5 6
Mock 0.008 0.0008
Con
duct
ivity
(µS
)
1e + 9
Bac
teria
l cel
l gro
wth
log(
CF
U c
m–2
)
01e + 11e + 3
1e + 8
0 2 3
Days post inoculation
Fig. 6 The Capsicum annuum receptor-like kinase 1 (CaRLK1)transgenic plants showed low electrolyte leakage. Ion leakage wasmeasured (a) before and (b) after infection with Pseudomonas syrin-
gae pv. tabaci. Leaf discs (1 cm in diameter) from wild-type (blackbars) and transgenic (gray bars) plants were used to measure ionleakage. (c) Growth of the virulent pathogen P. syringae pv. tabaci
in colony-forming units cm)2 (control, black bars; CaRLK1-3, lightgray bars; CaRLK1-5, dark gray bars). Data represent means of threeplants and error bars represent standard deviations. The experimentswere repeated four times with similar results.
WT CaRLK1-3
(a)
(b)
(c)
(d)
Fig. 7 Detection of O2) generation in the Capsicum annuum
receptor-like kinase 1 (CaRLK1) transgenic plants. (a) Detection ofO2
) with nitroblue tetrazolium (NBT) staining. Leaves were infil-trated using vacuum infiltration after being submerging in a reactionmixture containing 1 mM NBT in 50 mM sodium phosphate buffer(pH 7.6) plus 10 mM sodium azide. (b) Microscope observations ofO2
) in vascular tissue. (c) Detection of peroxynitrite using L-012(8-amino-5-chloro-7-phenylpyridol[3,4-d]pyridazine-1,4(2H,3H)dione). A reaction mixture containing 100 lM L-012 in 50 mMsodium phosphate buffer (pH 7.6) with or without preinjection of50 lM diphenylene iodonium (DPI) was infiltrated into leaves usinga needleless syringe. (d) Detection of H2O2 using the 3,3¢-diam-inobenzidine tetrahydrochloride (DAB) staining method. The reac-tion mixture contained 200 ml of 0.5 mM DAB in 50 mM Trisacetate buffer (pH 5.0). Leaves were vacuum-infiltrated for 10 min,incubated overnight at 25�C, and then cleared twice in 80% (v ⁄ v)ethanol for 10 min at 70�C. The experiment was repeated four timeswith similar results.
NewPhytologist Research 709
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715
www.newphytologist.org
Thus, we confirmed that superoxide anions are the mainROS in the transgenic plants.
Effect of CaRLK1 on the expression of NbRBOH genes
To elucidate which genes are involved in the production ofthe superoxide anion, we compared the relative expressionof RBOH genes. RBOH genes, encoding NADPH oxidase,are known to produce the superoxide anion in the extracel-lular region after pathogen attack or other stresses (Torreset al., 2005). These NADPH oxidases are the primary can-didates suggested to be responsible for ROS accumulation,which may be regulated by expression of the CaRLK1 gene.The function of NbRBOHA and NbRBOHB was alreadydetermined to share superoxide anion production, H2O2
generation and resistance to bacterial pathogene (Yoshiokaet al., 2003). Partial cDNA sequences of NbRBOHD geneswere isolated from an expressed sequence tag (EST) data-base (http://www.pdrc.re.kr). A phylogenetic tree con-structed using the clustal w program with PIR as theoutput format (http://align.genome.jp/) to compare theamino acid sequences of 10 AtRBOH genes and threeNbRBOH genes showed that the NbRBOHD gene has rela-tively high sequence homology to AtRBOHD (Fig. S6).The expression of the NbRBOHA, NbRBOHB, andNbRBOHD genes was higher in the transgenic plants thanin the wild type, suggesting that the expression of the CaR-LK1 gene plays a role in ROS accumulation at the extracel-lular matrix through the production of superoxide anions(Fig. 8a).
To determine whether the higher expression levels ofRBOH genes contribute much amount of superoxide anion,O2
)-generating activity was measured using the leaf discmethod (Sagi & Fluhr, 2001; Achard et al., 2008). Leafdiscs from the transgenic plants showed higher O2
)-gener-ating activity than those from wild-type plants (Fig. 8b).Treatment with DPI decreased the induction of O2
)-gener-ating activity in the transgenic plants, although it did notdecrease the basal O2
)-generating activity in wild-typeplants (Fig. 8c), as it blocked the increase in chemilumines-cence in the transgenic plants after treatment with L-012(Fig. 7c). These results strongly suggest that CaRLK1 maycontrol O2
) accumulation through induction of the RBOHgenes.
Effect of CaRLK1 on expression of antioxidant-relatedgenes
The expression of two (NbCSD2 and NbCSD3) of the threeCuZnSOD (CSD) genes was slightly increased in the trans-genic plants as compared with the wild type (Fig. 9a), butthere was no increase in the expression of the two NbFeSODor the two NbMnSOD genes (data not shown). Althoughtwo CSD genes are thought to be targeted to the chloroplast
CaRLK1
CaRLK1
NbRBOHA
NbRBOHB
NbRBOHDNbRBOHD
NbAct
100
120
40
60
80
140
O2– g
ener
atin
g ac
tivity
(µM
mg–
1 F
W h
–1)
O2– g
ener
atin
g ac
tivity
(µM
mg–
1 F
W h
–1)
C
(a)
(b)
(c)
2 3 4 Line no.
2 3 4 Line no.
CaRLK1
0
20
80
100
120
20
40
60
0WT CaRLK1-3
Fig. 8 Ectopic expression of Capsicum annuum receptor-like kinase1 (CaRLK1) regulates NADPH oxidase levels. (a) Increased levels ofNADPH oxidase transcripts. Levels of Nicotiana benthamiana
respiratory burst oxidase homolog A (NbRBOHA), NbRBOHB,NbRBOHD, and Nicotiana benthamiana Actin (CaAct) transcriptswere determined by RT-PCR. The sequence of NbROBHD was foundafter a BLAST search using Arabidopsis thaliana RBOH genes. NbAct
transcripts were amplified and are shown as a loading control. (b)O2
)-generating activity for wild-type and transgenic plants. (c) DPIblocks O2
)-generating activity in the transgenic plants. Leaf discs of4 mm diameter were immersed in 50 mM Tris buffer (pH 7.4) andtransferred to the same buffer. After 60 min, the reaction was initi-ated by the addition of 0.3 mM XTT (2,3-bis(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide inner salt) and 2 mMNADPH. XTT reduction was determined at 450 nm. The rate was cal-culated with an extinction coefficient of e = 2.16 · 104 M)1 cm)1
(mock, black bars; DPI, gray bars). All data points are mean ± SD.Similar results were obtained in three independent experiments. Onerepresentative experiment is shown.
710 Research
NewPhytologist
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715
www.newphytologist.org
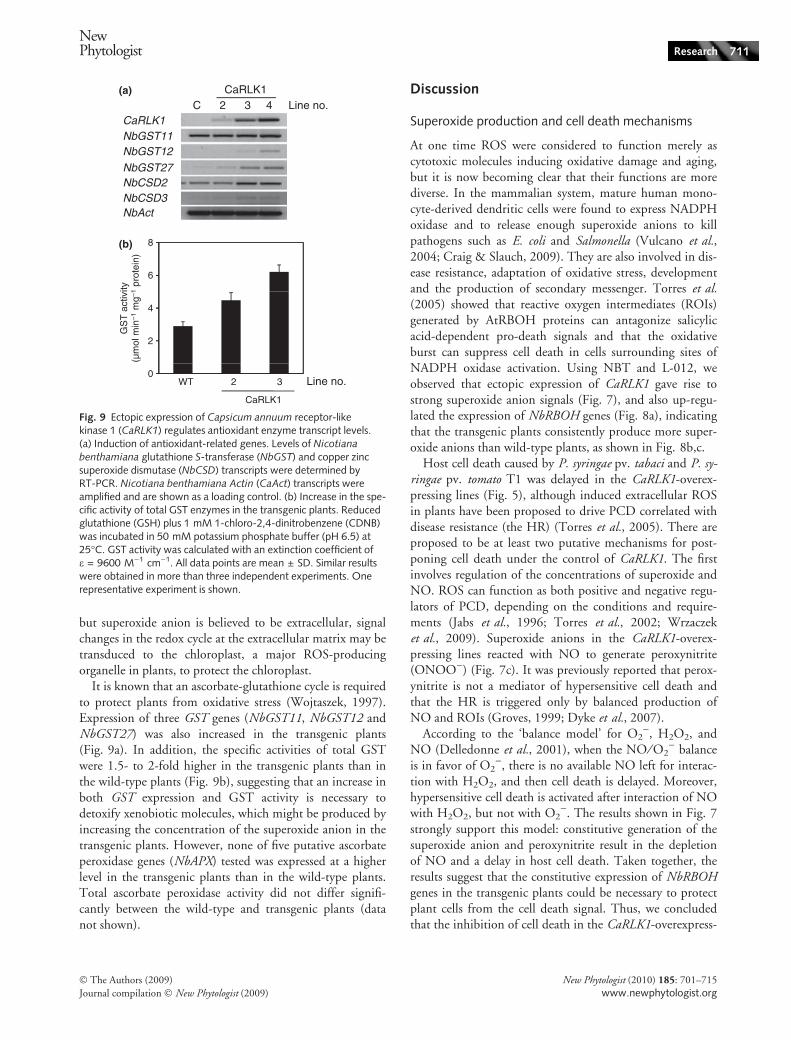
but superoxide anion is believed to be extracellular, signalchanges in the redox cycle at the extracellular matrix may betransduced to the chloroplast, a major ROS-producingorganelle in plants, to protect the chloroplast.
It is known that an ascorbate-glutathione cycle is requiredto protect plants from oxidative stress (Wojtaszek, 1997).Expression of three GST genes (NbGST11, NbGST12 andNbGST27) was also increased in the transgenic plants(Fig. 9a). In addition, the specific activities of total GSTwere 1.5- to 2-fold higher in the transgenic plants than inthe wild-type plants (Fig. 9b), suggesting that an increase inboth GST expression and GST activity is necessary todetoxify xenobiotic molecules, which might be produced byincreasing the concentration of the superoxide anion in thetransgenic plants. However, none of five putative ascorbateperoxidase genes (NbAPX) tested was expressed at a higherlevel in the transgenic plants than in the wild-type plants.Total ascorbate peroxidase activity did not differ signifi-cantly between the wild-type and transgenic plants (datanot shown).
Discussion
Superoxide production and cell death mechanisms
At one time ROS were considered to function merely ascytotoxic molecules inducing oxidative damage and aging,but it is now becoming clear that their functions are morediverse. In the mammalian system, mature human mono-cyte-derived dendritic cells were found to express NADPHoxidase and to release enough superoxide anions to killpathogens such as E. coli and Salmonella (Vulcano et al.,2004; Craig & Slauch, 2009). They are also involved in dis-ease resistance, adaptation of oxidative stress, developmentand the production of secondary messenger. Torres et al.(2005) showed that reactive oxygen intermediates (ROIs)generated by AtRBOH proteins can antagonize salicylicacid-dependent pro-death signals and that the oxidativeburst can suppress cell death in cells surrounding sites ofNADPH oxidase activation. Using NBT and L-012, weobserved that ectopic expression of CaRLK1 gave rise tostrong superoxide anion signals (Fig. 7), and also up-regu-lated the expression of NbRBOH genes (Fig. 8a), indicatingthat the transgenic plants consistently produce more super-oxide anions than wild-type plants, as shown in Fig. 8b,c.
Host cell death caused by P. syringae pv. tabaci and P. sy-ringae pv. tomato T1 was delayed in the CaRLK1-overex-pressing lines (Fig. 5), although induced extracellular ROSin plants have been proposed to drive PCD correlated withdisease resistance (the HR) (Torres et al., 2005). There areproposed to be at least two putative mechanisms for post-poning cell death under the control of CaRLK1. The firstinvolves regulation of the concentrations of superoxide andNO. ROS can function as both positive and negative regu-lators of PCD, depending on the conditions and require-ments (Jabs et al., 1996; Torres et al., 2002; Wrzaczeket al., 2009). Superoxide anions in the CaRLK1-overex-pressing lines reacted with NO to generate peroxynitrite(ONOO)) (Fig. 7c). It was previously reported that perox-ynitrite is not a mediator of hypersensitive cell death andthat the HR is triggered only by balanced production ofNO and ROIs (Groves, 1999; Dyke et al., 2007).
According to the ‘balance model’ for O2), H2O2, and
NO (Delledonne et al., 2001), when the NO ⁄ O2) balance
is in favor of O2), there is no available NO left for interac-
tion with H2O2, and then cell death is delayed. Moreover,hypersensitive cell death is activated after interaction of NOwith H2O2, but not with O2
). The results shown in Fig. 7strongly support this model: constitutive generation of thesuperoxide anion and peroxynitrite result in the depletionof NO and a delay in host cell death. Taken together, theresults suggest that the constitutive expression of NbRBOHgenes in the transgenic plants could be necessary to protectplant cells from the cell death signal. Thus, we concludedthat the inhibition of cell death in the CaRLK1-overexpress-
CaRLK1
NbGST11CaRLK1
NbGST27
NbCSD3NbCSD2
NbGST12
NbAct
6
8
GS
T a
ctiv
ity(µ
mol
min
–1 m
g–1
prot
ein)
2
4
0
C 2 3(a)
(b)
4 Line no.
WT 2 3 Line no.
CaRLK1
Fig. 9 Ectopic expression of Capsicum annuum receptor-likekinase 1 (CaRLK1) regulates antioxidant enzyme transcript levels.(a) Induction of antioxidant-related genes. Levels of Nicotiana
benthamiana glutathione S-transferase (NbGST) and copper zincsuperoxide dismutase (NbCSD) transcripts were determined byRT-PCR. Nicotiana benthamiana Actin (CaAct) transcripts wereamplified and are shown as a loading control. (b) Increase in the spe-cific activity of total GST enzymes in the transgenic plants. Reducedglutathione (GSH) plus 1 mM 1-chloro-2,4-dinitrobenzene (CDNB)was incubated in 50 mM potassium phosphate buffer (pH 6.5) at25�C. GST activity was calculated with an extinction coefficient ofe = 9600 M)1 cm)1. All data points are mean ± SD. Similar resultswere obtained in more than three independent experiments. Onerepresentative experiment is shown.
NewPhytologist Research 711
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715
www.newphytologist.org

ing plants was not attributable to an indirect or pleiotropiceffect as a consequence of transformation, but instead to thefunction of the CaRLK1 gene.
According to the ‘speculative model’ of Beers & McDo-well (2001), PCD, such as the HR, is triggered by H2O2
rather than O2) because the HR is inhibited by expression
of SOD, and antioxidant enzymes are strongly induced byoverproduction of O2
), but not H2O2. Thus, it is veryimportant to elucidate whether the high concentration ofO2
) in the transgenic plants induced a high or low concen-tration of H2O2. Less DAB staining was observed in thetransgenic leaves than in wild-type leaves (Fig. 7d). Treat-ment with ascorbic acid and DAB together completely sup-pressed this DAB staining (Fig. S5). These results suggestthat the delay of cell death in the transgenic plants is associ-ated with a low concentration of H2O2.
Both of these models strongly suggest that constitutivegeneration of O2
) and peroxynitrite and a low concentra-tion of H2O2 lead to the collapse of the NO ⁄ O2
) balanceand a less reaction of NO with H2O2, and finally delay celldeath in the transgenic plants. Thus, O2
) generated in thetransgenic plants seems to be a survival signal.
The second is the levels of LSD. The LSD1 protein func-tions to negatively regulate a pro-death pathway componentby regulating ROS concentrations or to activate a repressorof plant cell death (Torres et al., 2005). The lesion stimulat-ing disease 1 (lsd1) mutant has been shown to develop super-oxide-dependent necrotic cell death in spreading lesions(Dietrich et al., 1997). These spreading legions in the lsdmutant were found to be induced by the application ofsuperoxide (Jabs et al., 1996). Expression of CaRLK1 wassubstantially up-regulated by treatment with 0.1–0.5 mMexogenous SA in pepper leaves (Fig. S1b). SA is requiredfor the function of CaRLK1, and SA accumulates inresponse to pathogen attack. In turn, induction of CaRLK1may increase expression of the putative CaLSD, as the ecto-pic expression of CaRLK1 induced NbLSD1 in N. benth-amiana (Fig. 5d). LSD1 is part of a signaling pathway forthe induction of the CuZnSOD protein to prevent celldeath (Kliebenstein et al., 1999), which reminds us that theexpression of two CSD genes (NbCSD2 and NbCSD3) wasslightly increased in the transgenic plants. Thus, consistentup-regulation of the production of the LSD protein in theCaRLK1 transgenic plants can play a role in inhibiting celldeath caused by pathogens or other factors.
How to survive in highly oxidative conditions?
When plants are exposed to stress, survival mechanisms areturned on to reduce damage. The balance between stressand survival signals determines the degree of damage suf-fered by the plant. Recent studies have demonstrated thatROS function as important signaling molecules involved inthe control of processes such as pathogen defense, hormonal
signaling, stress responses, plant growth, and development(Torres et al., 2002; Foreman et al., 2003; Kwak et al.,2003). ROS may play different or even opposite roles underdifferent plant growth and developmental conditions. Dur-ing the HR, they are actively produced to trigger cell death.However, the production of ROS under some abiotic stressconditions is thought to be a byproduct of stress metabo-lism for survival of the cell (Neill et al., 2002; Mittler et al.,2004). Rusterucci et al. (2001) suggested that the nature ofROIs produced by cells undergoing the HR is differentfrom that of ROIs associated with signaling from thosecells, and monitored by LSD1. Interestingly, CaRLK1 hasbeen suggested to play a role in delaying cell death (Fig. 5).
Superoxide production was enhanced by elicitors,mechanical factors, or pathogens. Thus, pathogens such asXag8ra and Xcv or their elicitors can induce NADPH oxi-dase expression regulated by CaRLK1 to produce superoxideanions. In turn, GST genes, one component of the ROSscavenging system, were also induced. Expression of GST11,GST12 and GST27 was increased in the transgenic plants(Fig. 9a). There is a interesting report that O2
) rather thanH2O2 is the primary ROI signal for induction of GSTbecause sodium diethyldithiocarbamate, a SOD inhibitor,substantially up-regulated expression of GST after treatmentof soybean (Glycine max) cells with P. syringae pv. glycineacarrying avrA (Delledonne et al., 2001). Thus, the induc-tion of GST11, GST12 and GST27 transcripts and theincrease in the specific activity of GST are attributable tosuperoxide anions accumulated through ectopic expressionof CaRLK1. Although their spatial expression was not eluci-dated in N. benthamiana, these three GST genes can protecttransgenic plants from the superoxide anion by removingxenobiotic material or other oxidized material, becauseGSTs can catalyze the cross-linking of reduced glutathioneand xenobiotic material to remove toxic molecules. In addi-tion, increased expression of NbLSD1 may also induce otherantioxidant molecules, because LSD1 is required for the SA-dependent induction of antioxidant CuZnSOD or otherantioxidant molecules (Kliebenstein et al., 1999).
Glutathione reductase (GR) catalyzes the reduction ofoxidized glutathione disulfide (GSSG) to glutathione(GSH) using NADPH as a substrate. This is an essentialreaction that maintains the GSH:GSSG ratio in the cyto-plasm. Unfortunately, expressions of GR1 and GR2 werenot induced in the transgenic plants (data not shown),reflecting that normal expressions of GR1 and GR2 areenough to protect plant from oxidative stress.
Ectopic expression of CaRLK1 did not affect overall phe-notype of N. benthamiana (data not shown). The overallphenotypes of CaRLK1-overexpressing plants, includingplant height, leaf size, root length, flower shape, fruit size,and flowering time, were not distinguishable from that ofthe wild type, although all the transgenic plants were undermild oxidative stress conditions at all times (data not shown).
712 Research
NewPhytologist
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715
www.newphytologist.org

This indicates that the concentration of the superoxide anionin the transgenic plants is not high enough to be cytotoxic,whereas in the lsd mutant the accumulation of the superox-ide anion is high enough to cause runaway cell death understress conditions (Jabs et al., 1996; Dietrich et al., 1997).
Further studies on function of CaRLK1 using moleculargenetics and biochemical approaches, including the deter-mination of its ligand in pathogen-challenged conditions,are warranted. Research on the downstream target proteinswill also provide insights into the role of CaRLK1 in patho-gen-induced cell death signaling, as the CaRLK1-KDdomain showed autophosphorylation activity in hot pepper(Fig. 4b).
Ectopic expression of CaRLK1 in agricultural plants maybe useful, in light of the finding that superoxide anions weremainly produced in the vascular system in the transgenicplants (Fig. 7b). This observation suggests that ectopicexpression of CaRLK1 may protect the plant from vasculartissue-invading pathogens such as Ralstonia solanacearum(Park et al., 2007) and Fusarium spp., similarly to the wayin which macrophages produce superoxide and kill infectingbacteria in mammalian systems (Vulcano et al., 2004; Craig& Slauch, 2009).
Acknowledgements
We would like to thank Dr Inhwan Hwang for theRFP:H+ATPase plasmid (Postech, Pohang, Korea). We alsothank Dr Choong-Min Ryu (Kribb, Daejon, Korea) forcritical reading of the manuscript. This work was supportedfinancially by grants from CFGC (CG1431) and the 21stCentury Frontier Research Program funded by MOST ofthe Korean Government. This work was also supported inpart by a grant from KRF (KRF-2008-313-C00853). SYYwas supported by the MOEHRD program of KRF.
References
Achard P, Renou JP, Berthome R, Harberd NP, Genschik P. 2008. Plant
DELLAs restrain growth and promote survival of adversity by reducing
the levels of reactive oxygen species. Current Biolology 18: 656–660.
Ali MB, Hahn EJ, Paek KY. 2005. Effects of light intensities on antioxi-
dant enzymes and malondialdehyde content during short-term acclima-
tization on micropropagated Phalaenopsis plantlet. Environmental andExperimental Botany 54: 109–120.
Bateman A, Coin L, Durbin R. 2004. The Pfam protein families database.
Nucleic Acids Research Database 32: D138–D141.
Beers EP, McDowell JM. 2001. Regulation and execution of programmed
cell death in response to pathogens, stress and developmental cues. Cur-rent Opinion in Plant Biology 4: 561–567.
Bindschedler LV, Dewdney J, Blee KA, Stone JM, Asai T, Plotnikov J,
Denoux C, Hayes T, Gerrish C, Davies DR et al. 2006. Peroxidase-
dependent apoplastic oxidative burst in Arabidopsis required for patho-
gen resistance. Plant Journal 47: 851–863.
Braun DM, Walker JC. 1996. Plant transmembrane receptors: new
pieces in the signaling puzzle. Trends in Biochemical Sciences 21:
70–73.
Clark SE, Williams RW, Meyerowitz EM. 1997. The CLAVATA1 gene
encodes a putative receptor kinase that controls shoot and floral meri-
stem size in Arabidopsis. Cell 89: 575–585.
Conklin PL, Last RL. 1995. Differential accumulation of antioxidant
mRNAs in Arabidopsis thaliana exposed to ozone. Plant Physiology 109:
203–212.
Coupe SA, Watson LM, Ryan DJ, Pinkney TT, Eason JR. 2004. Molecu-
lar analysis of programmed cell death during senescence in Arabidopsisthaliana and Brassica oleracea: cloning broccoli LSD1, Bax inhibitor and
serine palmitoyltransferase homologues. Journal of Experimental Botany55: 59–68.
Craig M, Slauch JM. 2009. Phagocytic superoxide specifically damages an
extracytoplasmic target to inhibit or kill Salmonella. PLoS ONE 4:
e4975.
Delledonne M, Zeier J, Marocco A, Lamb C. 2001. Signal interactions
between nitric oxide and reactive oxygen intermediates in the plant
hypersensitive disease resistance response. Proceedings of the NationalAcademy of Sciences, USA 98: 13454–13459.
Dietrich RA, Richberg MH, Schmidt R, Dean C, Dangl JL. 1997. A
novel zinc finger protein is encoded by the Arabidopsis LSD1 gene and
functions as a negative regulator of plant cell death. Cell 88: 685–694.
Dyke KV, Ghareeb E, Dyke MV, Thiel DHV. 2007. Ultrasensitive
peroxynitrite-based luminescence with L-012 as a screening system for
antioxidative ⁄ antinitrating substances, e.g. Tylenol (acetaminophen),
4-OH tempol, quercetin and carboxy-PTIO. Luminescence 22: 267–274.
Feuillet C, Schachermayr G, Keller B. 1997. Molecular cloning of a new
receptor-like kinase gene encoded at the Lr10 disease resistance locus of
wheat. Plant Journal 11: 45–52.
Foreman J, Demidchik V, Bothwell JH, Mylona P, Miedema H, Torres
MA, Linstead P, Costa S, Brownlee C, Jones JDG et al. 2003. Reactive
oxygen species produced by NADPH oxidase regulate plant cell growth.
Nature 422: 442–446.
Gomez-Gomez L, Boller T. 2000. FLS2: an LRR receptor-like
kinase involved in the perception of the bacterial elicitor flagellin in
Arabidopsis. Molecules and Cells 5: 1003–1011.
Groves JT. 1999. Peroxynitrite: reactive, invasive and enigmatic. CurrentOpinion in Chemical Biology 3: 226–235.
Hanks SK, Quinn AM. 1991. Protein kinase catalytic domain sequence
database: identification of conserved features of 1� structure and classifi-
cation of family members. Methods In Enzymology 200: 38–62.
Heese A, Hann DR, Gimenez-Ibanez S, Jones AM, He K, Li J, Schroeder
JI, Peck SC, Rathjen JP. 2007. The receptor-like kinase SERK3 ⁄ BAK1is a central regulator of innate immunity in plants. Proceedings of theNational Academy of Sciences, USA 104: 12217–12222.
Horn MA, Walker JC. 1994. Biochemical properties of the autophospho-
rylation of RLK5, a receptor-like protein kinase from Arabidopsis thali-
ana. Biochimica et Biophysica Acta 1208: 65–74.
Imada I, Sato EF, Miyamoto M, Ichimori Y, Minamiyama Y, Konaka R,
Inoue M. 1999. Analysis of reactive oxygen species generated by neu-
trophils using a chemiluminescence probe L-012. Analytical Biochemistry271: 53–58.
Jabs T, Dietrich RA, Dangl JL. 1996. Initiation of runaway cell death in
an Arabidopsis mutant by extracellular superoxide. Science 273: 1853–
1856.
Johnson SM, Doherty SJ, Croy RRD. 2003. Biphasic superoxide genera-
tion in potato tubers. A self-amplifying response to stress. Plant Physiol-ogy 131: 1440–1449.
Kajava AV. 1998. Structural diversity of leucine-rich repeat proteins.
Journal of Molecular Biology 277: 519–527.
Kim SG, Kim YH. 2009. Histological and cytological changes associated
with susceptible and resistant responses of chili pepper root and stem to
Phytophthora capsici infection. Plant Pathology Journal 25: 113–120.
Kim DH, Eu YJ, Yoo CM, Kim YW, Pih KT, Jin JB, Kim SJ, Stenmark
H, Hwang I. 2001. Trafficking of phosphatidylinositol 3-phosphate
NewPhytologist Research 713
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715
www.newphytologist.org

from the trans-Golgi network to the lumen of the central vacuole in
plant cells. Plant Cell 13: 287–301.
Kliebenstein DJ, Dietrich RA, Martin AC, Last RL, Dangl JL. 1999.
LSD1 regulates salicylic acid induction of copper zinc superoxide dismu-
tase in Arabidopsis thaliana. Molecular Plant-Microbe Interaction 12:
1022–1026.
Koch E, Slusarenko A. 1990. Arabidopsis is susceptible to infection by a
downy mildew fungus. Plant Cell 2: 437–445.
Kwak JM, Mori IC, Pei ZM, Leonhardt N, Torres MA, Dangl JL, Bloom
RE, Bodde S, Jones JD, Schroeder JI. 2003. NADPH oxidase AtrbohDand AtrbohF genes function in ROS-dependent ABA signaling in Ara-
bidopsis. EMBO Journal 22: 2623–2633.
Lee DJ, Zeevaart JAD. 2005. Molecular cloning of GA 2-cxidase3 from
spinach and its ectopic expression in Nicotiana sylvestris. Plant Physiology138: 243–254.
Lee YJ, Kim DH, Kim Y-W, Hwang I. 2001. Identification of a signal that
distinguishes between the chloroplast outer envelope membrane and the
endomembrane system in vivo. Plant Cell 13: 2175–2190.
Lee SJ, Lee MY, Yi SY, Oh SK, Choi SH, Her NH, Choi D, Min BW,
Yang SG, Harn CH. 2002. PPI1: a novel pathogen-induced basic
region-leucine zipper (bZIP) transcription factor from pepper. MolecularPlant-Microbe Interaction 15: 540–548.
Lehti-Shiu MD, Zou C, Hanada K, Shiu S-H. 2009. Evolutionary history
and stress regulation of plant receptor-like kinase ⁄ pelle genes. PlantPhysiology 150: 12–26.
Liu Y, Ren D, Pike S, Pallardy S, Gassmann W, Zhang S. 2007. Chloro-
plast-generated reactive oxygen species are involved in hypersensitive
response-like cell death mediated by a mitogen-activated protein kinase
cascade. Plant Journal 51: 941–954.
Matsubayashi Y, Ogawa M, Morita A, Sakagami Y. 2002. An LRR recep-
tor kinase involved in perception of a peptide plant hormone, phy-
tosulfokine. Science 296: 1470–1472.
Miller G, Schlauch K, Tam R, Cortes D, Torres MA, Shulaev V, Dangl JL,
Mittler R. 2009. The plant NADPH cxidase RBOHD mediates rapid
systemic signaling in response to diverse stimuli. Science Signaling 2: ra45.
Mittler R, Vanderauwera S, Gollery M, Van Breusegem F. 2004. The reac-
tive oxygen gene network of plants. Trends in Plant Science 9: 490–498.
Mizuno S, Osakabe Y, Maruyama K, Ito T, Osakabe K, Sato T, Shinozaki
K, Yamaguchi-Shinozaki K. 2007. Receptor-like protein kinase 2
(RPK2) is a novel factor controlling anther development in Arabidopsisthaliana. Plant Journal 50: 751–766.
Muranaka T, Banno H, Machida Y. 1994. Characterization of tobacco
protein kinase NPK5, a homolog of Saccharomyces cerevisiae SNF1 that
constitutively activates expression of the glucose-repressible SUC2 gene
for a secreted invertase of S. cerevisiae. Molecular and Cellular Biology 14:
2958–2965.
Neill S, Desikan R, Hancock J. 2002. Hydrogen peroxide signalling. Cur-rent Opinion in Plant Biology 5: 388–395.
Oh M-H, Wang X, Kota U, Goshe MB, Clouse SD, Huber SC. 2009.
Tyrosine phosphorylation of the BRI1 receptor kinase emerges as a com-
ponent of brassinosteroid signaling in Arabidopsis. Proceedings of theNational Academy of Sciences, USA 106: 658–663.
Park CJ, Shin R, Park JM, Lee GJ, Yoo TH, Paek KH. 2001. A hot pep-
per cDNA encoding a pathogenesis-related protein 4 is induced during
the resistance response to tobacco mosaic virus. Molecules and Cells 28:
122–127.
Park EJ, Lee SD, Chung EJ, Lee MH, Um HY, Murugaiyan S, Moon BJ,
Lee S-W. 2007. MicroTom – a model plant system to study bacterial
wilt by Ralstonia solanacearum. Plant Pathology Journal 23: 239–244.
Ricci G, Bello ML, Caccuri AM, Galiazzo F, Federici G. 1984. Detection
of glutathione transferase activity on polyacrylamide gels. Analytical Bio-chemistry 143: 226–230.
Rusterucci C, Aviv DH, Holt BF III, Dangl JL, Parker JE. 2001. The dis-
ease resistance signaling components EDS1 and PAD4 are essential regu-
lators of the cell death pathway controlled by LSD1 in Arabidopsis.
Plant Cell 13: 2211–2224.
Sagi M, Fluhr R. 2001. Superoxide production by plant homologues of
the gp91phox NADPH oxidase. Modulation of activity by calcium and
by tobacco mosaic virus infection. Plant Physiology 126: 1281–1290.
Sambrook J, Fritrsch EF, Maniatis T 1989. Molecular cloning: a laboratorymanual, 2nd Edn. Plainview, NY, USA: Cold Spring Harbor Laboratory
Press.
Sharma YK, Davis KR. 1997. The effects of ozone on antioxidant
responses in plants. Free Radical Biology and Medicine 23: 480–488.
Shi J, Kim K-N, Ritz O, Albrecht V, Gupta R, Harter K, Luan S, Kudla
J. 1999. Novel protein kinases associated with calcineurin B-like calcium
censors in Arabidopsis. Plant Cell 11: 2393–2405.
Shiu SH, Bleecker AB. 2001. Plant receptor-like kinase gene family: diver-
sity, function, and signaling. Science’s STKE 18: RE22.
Shiu SH, Karlowski WM, Pan R, Tzeng YH, Mayer KF, Li WH. 2004.
Comparative analysis of the receptor-like kinase family in Arabidopsis
and rice. Plant Cell 16: 1220–1234.
Simon-Plas F, Elmayan T, Blein J-P. 2002. The plasma membrane oxi-
dase NtrbohD is responsible for AOS production in elicited tobacco cells.
Plant Journal 31: 137–148.
Song W-Y, Wang G-L, Chen L-L, Kim H-S, Pi L-Y, Holsten T, Gadner
J, Wang B, Zhai W-X, Zhu L-H et al. 1995. A receptor kinase-like pro-
tein encoded by the rice disease resistance gene, Xa21. Science 270:
1804–1806.
Thara VK, Tang X, Gu YQ, Martin GB, Zhou JM. 1999. Pseudomonas sy-ringae pv. tomato induces the expression of tomato EREBP-like genes
pti4 and pti5 independent of ethylene, salicylate and jasmonate. PlantJournal 20: 475–483.
Thilmony RL, Chen Z, Bressan RA, Martin GB. 1995. Expression of the
tomato Pto gene in tobacco enhances resistance to Pseudomonas syringaepv. tabaci expressing avrPto. Plant Cell 7: 1529–1536.
Torii KU. 2000. Receptor kinase activation and signal transduction in
plants: an emerging picture. Molecular Plant-Microbe Interaction 3: 361–
367.
Torres MA, Dangl JL. 2005. Functions of the respiratory burst oxidase in
biotic interactions, abiotic stress and development. Molecular Plant-Microbe Interaction 8: 397–403.
Torres MA, Onouchi H, Hamada S, Machida C, Hammond-Kosack KE,
Jones JD. 1998. Six Arabidopsis thaliana homologues of the human
respiratory burst oxidase (gp91phox). Plant Journal 14: 365–370.
Torres MA, Dangl JL, Jones JD. 2002. Arabidopsis gp91phox homologues
AtrbohD and AtrbohF are required for accumulation of reactive oxygen
intermediates in the plant defense response. Proceedings of the NationalAcademy of Sciences, USA 99: 517–522.
Torres MA, Jones JD, Dangl JL. 2005. Pathogen-induced, NADPH oxi-
dase-derived reactive oxygen intermediates suppress spread of cell death
in Arabidopsis thaliana. Nature Genetics 37: 1130–1134.
Vulcano M, Dusi S, Lissandrini D, Badolato R, Mazzi P, Riboldi E, Bor-
roni E, Calleri A, Donini M, Plebani A et al. 2004. Toll receptor-medi-
ated regulation of NADPH oxidase in human Dendritic cells. Journal ofImmunology 173: 5749–5756.
Wagner TA, Kohorn BD. 2001. Wall-associated kinases are expressed
throughout plant development and are required for cell expansion. PlantCell 13: 303–318.
Wojtaszek P. 1997. Oxidative burst: an early plant response to pathogen
infection. Biochemical Journal 322: 681–692.
Wrzaczek M, Brosche M, Kollist H, Kangasjarvi J. 2009. Arabidopsis
GRI is involved in the regulation of cell death induced by extracellular
ROS. Proceedings of the National Academy of Sciences, USA 106: 5412–
5417.
Yi G, Lee JM, Lee S, Choi D, Kim BD. 2006. Exploitation of pepper
EST-SSRs and an SSR-based linkage map. Theoretical and AppliedGenetics 114: 113–130.
714 Research
NewPhytologist
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715
www.newphytologist.org

Yoshioka H, Numata N, Nakajima K, Katou S, Kawakita K, Rowland
O, Jones JD, Doke N. 2003. Nicotiana benthamiana gp91phox
homologs NbrbohA and NbrbohB participate in H2O2 accumulation
and resistance to Phytophthora infestans. Plant Cell 15: 706–718.
Zipfel C, Kunze G, Chinchilla D, Caniard A, Jones JD, Boller T,
Felix G. 2006. Perception of the bacterial PAMP EF-Tu by the
receptor EFR restricts Agrobacterium-mediated transformation. Cell125: 749–760.
Supporting Information
Additional supporting information may be found in theonline version of this article.
Fig. S1 (a) The spatial expression of the Capsicum annuumreceptor-like kinase 1 (CaRLK1) transcript in pepper. (b)Induction of CaRLK1 transcripts after salicylic acid (SA)treatment. (c) Ectopic expression of CaRLK1 up-regulatedthe expression of Nicotiana benthamiana nonexpressor ofpathogenisis-related genes (NbNPR), Nicotiana benthami-ana systemic acquired resistance 8.2 (NbSAR8.2), patho-genesis-related 1a (PR1a), and PR1b.
Fig. S2 Comparison of the putative amino acid sequencesof receptor-like kinase (RLK) genes.
Fig. S3 Detection of host cell death after treatment withPseudomonas syringae pv. tabaci.
Fig. S4 Detection of superoxide anions with nitrobluetetrazolium (NBT).
Fig. S5 There was less 3,3¢-diaminobenzidine tetrahydro-chloride (DAB) staining in the transgenic plant leaves.
Fig. S6 Comparison of the putative amino acid sequencesof the respiratory burst oxidase homolog (RBOH) genes.
Table S1 Primers used in this study.
Please note: Wiley-Blackwell are not responsible for the con-tent or functionality of any supporting information suppliedby the authors. Any queries (other than about missing mate-rial) should be directed to the New Phytologist Central Office.
NewPhytologist Research 715
� The Authors (2009)
Journal compilation � New Phytologist (2009)
New Phytologist (2010) 185: 701–715
www.newphytologist.org