new ribosomal rna gene locations in gossypium hirsutum mapped by meiotic fish
TRANSCRIPT

Chromosoma (1999) 108:200±207
� Springer-Verlag 1999
New ribosomal RNA gene locations in Gossypium hirsutum mappedby meiotic FISHYuanfu Ji1, Marcos De Donato1, Charles F. Crane1, Wayne A. Raska1, M. Nurul Islam-Faridi1, Thomas D. McKnight2,H. James Price1, David M. Stelly1
1 Department of Soil and Crop Sciences, Texas A&M University, College Station, TX 77843, USA2 Department of Biology, Texas A&M University, College Station, TX 77843, USA
Received: 26 January 1998; in revised form: 28 May 1998 / Accepted: 12 February 1999
Edited by: R. Appels
Correspondence to: D.M. Stellye-mail: [email protected]
Abstract. In this study we have mapped newly identifiedrDNA loci in Gossypium hirsutum. Four new minor 18S-26S rDNA loci, in addition to the sites previously identi-fied, were mapped using fluorescence in situ hybridiza-tion (FISH) to heterozygous translocation (NT) quadriva-lents (IVs). The newly detected 18S-26S rDNA loci weremapped to the right arms of chromosomes 8, 9, 15, 17,19, 20, and 23 and the left arms of chromosomes 5, 11,12, and 14. Using the rDNA loci as common referencepoints, we detected several erroneous arm assignmentsin the previously published map of NT breakpoints. Thedata are summarized in the form of an integrated mapfor all 17 known rDNA loci, relative to centromeres, telo-meres, and NT breakpoints. This information will facili-tate future locus-specific research on rRNA gene evolu-tion and function.
Introduction
Among the most thoroughly investigated repetitive DNAsequences in plant species are rRNA genes, including18S-26S and 5S rDNA tandem repeats (Schmidt et al.1994; Cabrera et al. 1995; Castilho and Heslop-Harrison1995). In several cases, improved in situ hybridization(ISH) techniques have led to identification of new rDNAloci (Alonso-Blanco et al. 1994; Cuadrado et al. 1995;Fominaya et al. 1995). Until recently, this was best exem-plified in wheat. The 18S-26S rDNA sites were originallyfound to be on three chromosome pairs in the commonwheat Chinese Spring by using radioactively detectedISH (Appels et al. 1980; Hutchinson and Miller 1982),and later, five pairs by using enzymatically detectedISH (Mukai et al. 1991). With increasing improvementof the ISH technique and the use of sensitive fluorescenceISH (FISH), two additional minor 18S-26S rDNA sites
were identified in Chinese Spring, and a third new locuswas identified in the common wheat Wichita (Jiang andGill 1994).
Three major 18S-26S rDNA sites (NORs) were detect-ed enzymatically in cotton (Gossypium hirsutum L.) bymeiotic ISH (Bergey et al. 1989). Improved meioticISH methods and development of new methods for mei-otic physical mapping led to the discovery of two more18S-26S rDNA sites, both relatively small, and two 5SrDNA loci (Price et al. 1990; Crane et al. 1993). Allbut one of the smaller 18S-26S rDNA sites were mappedto specific chromosomal arms (Price et al. 1990; Crane etal. 1993). More recently, high-sensitivity mitotic FISH re-vealed at least six additional 18S-26S rDNA loci (Hansonet al. 1996). However, none of the latter loci have beenmapped.
In this study, we mapped and integrated all the rDNAloci into the translocation breakpoint map (Menzel et al.1985) by meiotic FISH to quadrivalents (IVs) of translo-cation heterozygotes (NTs). We expect the integratedmap of the rDNA loci, centromeres, telomeres, and trans-location breakpoints will expedite physical mapping ofunassigned DNA sequences, as well as study of rDNAevolution.
Materials and methods
Plant material. Reciprocal translocation heterozygotes of G. hirsu-tum L. used throughout the study were developed from translocationlines maintained in the Cotton Cytogenetic Collection (Table 1).They were produced by crossing the translocation homozygotes toTM-1, an inbred line that is regarded as a genetic standard (Kohelet al. 1970).
Chromosome preparation. Meiotic chromosome spreads were pre-pared according to the procedures described by Crane et al.(1993) using floral buds from the translocation heterozygotes. Slideswere frozen in liquid nitrogen and then stored in a freezer at±135�C.
Probe labeling and hybridization mixture preparation. PlasmidspGmr3, containing the 4.5 kb EcoRI fragment of the 18S-26S rDNA
201
repeat of Glycine max in pBR325 (kindly supplied by ElizabethZimmer), and pAm033, containing the 470 bp BamHI fragment ofthe 5S rDNA repeat of Acacia melanoxylon in pUC118 (kindly sup-plied by Rudi Appels), were labeled by nick-translation with bothbiotin-14-dATP (BRL BioNick kit) and digoxigenin-11-dUTP witha 1:2 ratio of digoxigenin-11-dUTP to dTTP (Boehringer Mann-heim). For single-color FISH, biotinylated 5S or 18S-26S rDNAprobe (final concentration 2.0 ng/�l) and Escherichia coli DNA (fi-nal concentration 240 ng/�l) were dissolved in a solution containing50% deionized formamide, 20% dextran sulfate, 2� SSC (1� SSC is0.15 M NaCl, 0.015 M sodium citrate). For dual-color FISH, biotin-ylated pGmr3 (final concentration 2.0 ng/�l), digoxigenin-labeledpAm033 (final concentration 2.0 ng/�l) and E. coli DNA (final con-centration 480 ng/�l) were dissolved in the solution mixture as de-scribed above. The probe mixture was denatured in a 90�C waterbath for 10 min, followed by immersion in ice water for 10 min be-fore use.
In situ hybridization. The procedures for single-color and dual-colorFISH were as described by Hanson et al. (1996) and Ji et al. (1997),respectively.
Fluorescence microscopy. Slides were screened visually with anOlympus AX-70 microscope equipped with 4',6-diamidino-2-phe-nylindole (DAPI), fluorescein isothiocyanate (FITC)-DAPI, andDAPI-FITC-rhodamine filter sets. Photographs were taken on Fuji-color 400 professional film. Prints were digitally scanned on a con-ventional flatbed scanner, and then processed to build plates, whichwere printed on a dye-sublimation printer.
Chromosomal and subchromosomal localization of rDNA sites. Po-sitions of rDNA FISH signals on metaphase I multivalents of NTswere used for chromosomal localization and subchromosomal local-ization relative to cytogenetic landmarks, i.e., centromeres, telo-meres, and translocation breakpoints (Brown et al. 1981; Menzel
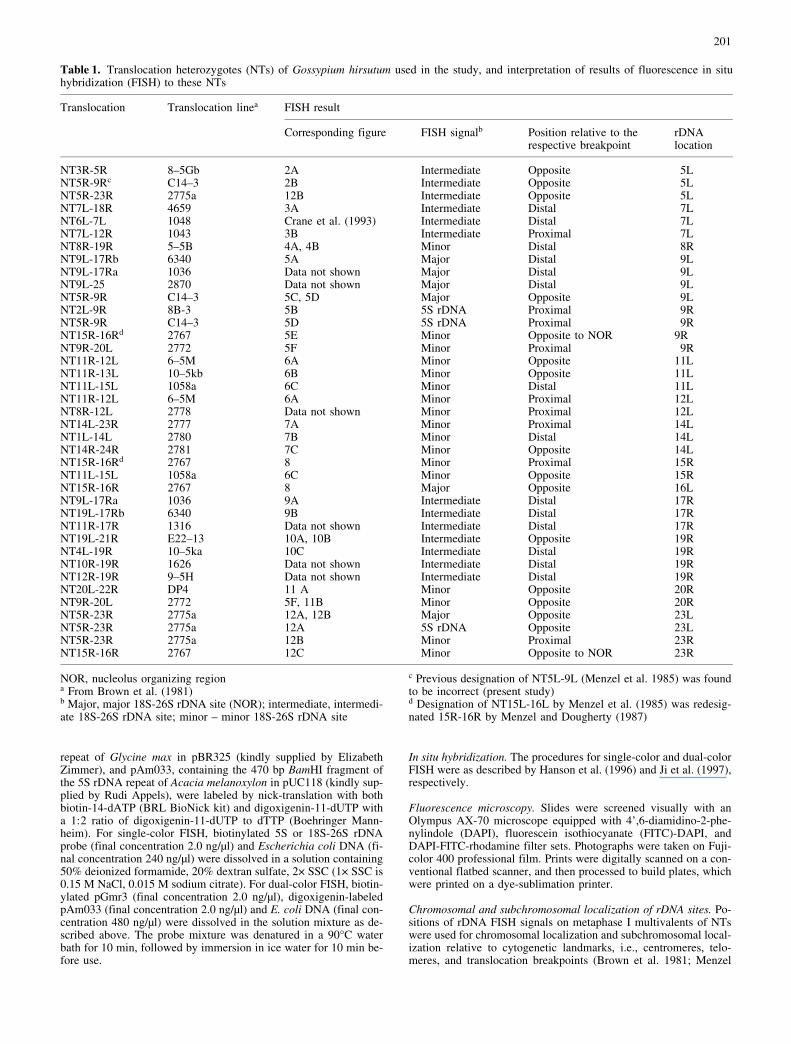
Table 1. Translocation heterozygotes (NTs) of Gossypium hirsutum used in the study, and interpretation of results of fluorescence in situhybridization (FISH) to these NTs
Translocation Translocation linea FISH result
Corresponding figure FISH signalb Position relative to therespective breakpoint
rDNAlocation
NT3R-5R 8±5Gb 2A Intermediate Opposite 5LNT5R-9Rc C14±3 2B Intermediate Opposite 5LNT5R-23R 2775a 12B Intermediate Opposite 5LNT7L-18R 4659 3A Intermediate Distal 7LNT6L-7L 1048 Crane et al. (1993) Intermediate Distal 7LNT7L-12R 1043 3B Intermediate Proximal 7LNT8R-19R 5±5B 4A, 4B Minor Distal 8RNT9L-17Rb 6340 5A Major Distal 9LNT9L-17Ra 1036 Data not shown Major Distal 9LNT9L-25 2870 Data not shown Major Distal 9LNT5R-9R C14±3 5C, 5D Major Opposite 9LNT2L-9R 8B-3 5B 5S rDNA Proximal 9RNT5R-9R C14±3 5D 5S rDNA Proximal 9RNT15R-16Rd 2767 5E Minor Opposite to NOR 9RNT9R-20L 2772 5F Minor Proximal 9RNT11R-12L 6±5M 6A Minor Opposite 11LNT11R-13L 10±5kb 6B Minor Opposite 11LNT11L-15L 1058a 6C Minor Distal 11LNT11R-12L 6±5M 6A Minor Proximal 12LNT8R-12L 2778 Data not shown Minor Proximal 12LNT14L-23R 2777 7A Minor Proximal 14LNT1L-14L 2780 7B Minor Distal 14LNT14R-24R 2781 7C Minor Opposite 14LNT15R-16Rd 2767 8 Minor Proximal 15RNT11L-15L 1058a 6C Minor Opposite 15RNT15R-16R 2767 8 Major Opposite 16LNT9L-17Ra 1036 9A Intermediate Distal 17RNT19L-17Rb 6340 9B Intermediate Distal 17RNT11R-17R 1316 Data not shown Intermediate Distal 17RNT19L-21R E22±13 10A, 10B Intermediate Opposite 19RNT4L-19R 10±5ka 10C Intermediate Distal 19RNT10R-19R 1626 Data not shown Intermediate Distal 19RNT12R-19R 9±5H Data not shown Intermediate Distal 19RNT20L-22R DP4 11 A Minor Opposite 20RNT9R-20L 2772 5F, 11B Minor Opposite 20RNT5R-23R 2775a 12A, 12B Major Opposite 23LNT5R-23R 2775a 12A 5S rDNA Opposite 23LNT5R-23R 2775a 12B Minor Proximal 23RNT15R-16R 2767 12C Minor Opposite to NOR 23R
NOR, nucleolus organizing regiona From Brown et al. (1981)b Major, major 18S-26S rDNA site (NOR); intermediate, intermedi-ate 18S-26S rDNA site; minor ± minor 18S-26S rDNA site
c Previous designation of NT5L-9L (Menzel et al. 1985) was foundto be incorrect (present study)d Designation of NT15L-16L by Menzel et al. (1985) was redesig-nated 15R-16R by Menzel and Dougherty (1987)

202
et al. 1985). The requisite interpretations were based on principlesestablished by Price et al. (1990), Crane et al. (1993), and Stellyet al. (1996).
Results
Chromosomal and subchromosomal localizationof rDNA sites
An overview of the chromosomal distribution of rDNAsites within G. hirsutum is presented in Fig. 1, and the ev-idence pertaining to each of the rDNA sites is presentedin Figs. 2±12.
Three major 18S-26S rDNA sites were previouslymapped by meiotic ISH to the left arms of chromo-
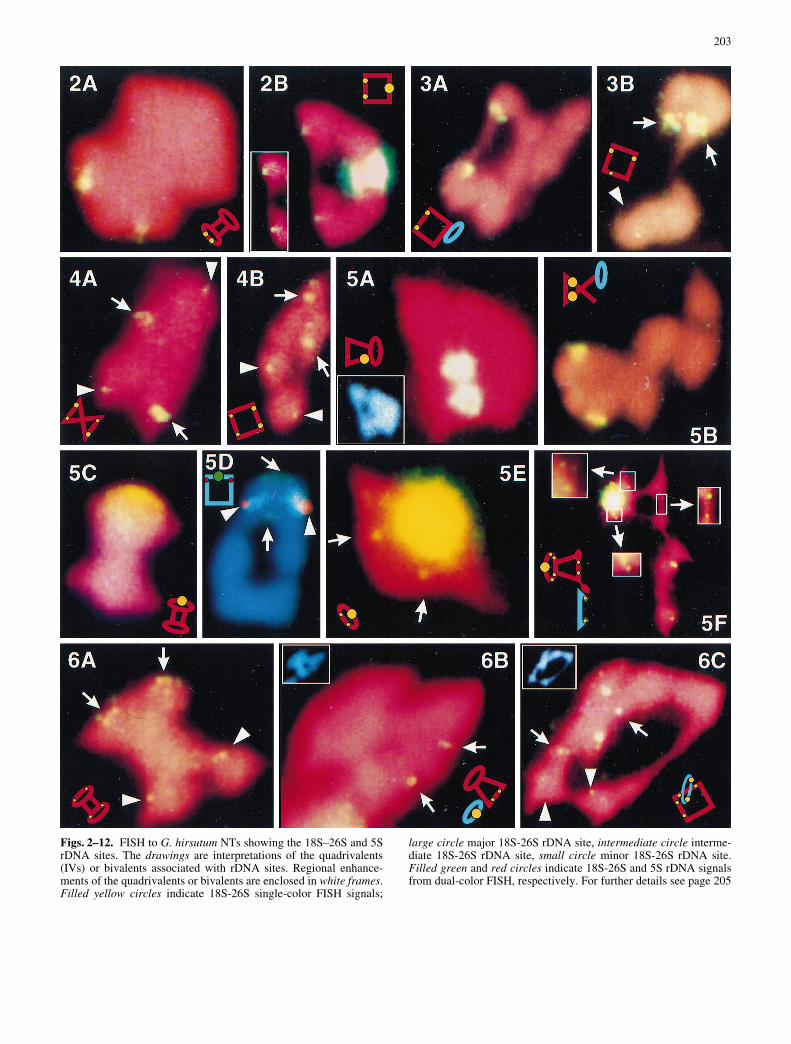
somes 9, 16 and 23 of G. hirsutum (Price et al. 1990;Crane et al. 1993). Two 5S rDNA sites of unequal sizewere also detected in cotton and were found to be on theright arm of chromosome 9 (9R) and left arm of chro-mosome 23 (23L), respectively (Crane et al. 1993).Our FISH analysis further delimited these major rDNAloci relative to the respective translocation breakpoints(Fig. 1). For example, our FISH analysis delimited themajor 18S-26S rDNA locus on chromosome 9 to thesegment distal to the T9L-17Rb breakpoint (Fig. 5A),and the 5S rDNA locus to the segment proximal toT2L-9R (Fig. 5B).
Four intermediate 18S-26S rDNA loci were mappedto the left arms of A-subgenome chromosomes 5(Fig. 2A, B) and 7 (Fig. 3A, B), and the right arms of
Fig. 1. Map of rDNA sites relative to centromeres, telomeres, andtranslocation breakpoints in G. hirsutum L. The description of18S-26S rDNA FISH sites as major, intermediate and minor sitesfollows Hanson et al. (1996). The following modifications of theprevious translocation map (Menzel et al. 1985) were also included:(i) the left (L) and right (R) arms are reconciled with the long (Lo)and short arms (sh)for chromosomes 14 (Menzel and Dougherty1987; Ji et al. in preparation), and 15 (Menzel and Richmond1985; Menzel and Dougherty 1987); (ii) the left and right armsare reconciled with the short and long arms for chromosomes 5
(Ji et al. in preparation), 16 (Menzel et al. 1987), 17 and 20 (unpub-lished data; Ji et al. in preparation); (iii) the original T5L-9L (lineC14±3), T6L-14L (line AZ-7), T15R-16L (line 8±5Ga) and T15L-16L (line 2767) from Menzel et al. (1985) were redesignated, re-spectively, as T5R-9R (present study), T6L-14R (Menzel andDougherty 1987; Ji et al. in preparation), 15L-16R and 15R-16R(Menzel and Dougherty 1987; Menzel et al. 1987). The sizes andpositions of rDNA sites are relative, but not drawn to scale. sh shortarm; Lo long arm; R right arm; L left arm; M major 18S-26S rDNAsite, I intermediate 18S-26S rDNA site, m minor 18S-26S rDNA site
203
Figs. 2±12. FISH to G. hirsutum NTs showing the 18S±26S and 5SrDNA sites. The drawings are interpretations of the quadrivalents(IVs) or bivalents associated with rDNA sites. Regional enhance-ments of the quadrivalents or bivalents are enclosed in white frames.Filled yellow circles indicate 18S-26S single-color FISH signals;
large circle major 18S-26S rDNA site, intermediate circle interme-diate 18S-26S rDNA site, small circle minor 18S-26S rDNA site.Filled green and red circles indicate 18S-26S and 5S rDNA signalsfrom dual-color FISH, respectively. For further details see page 205
204

205
D-subgenome chromosomes 17 (Fig. 9A, B) and 19(Fig. 10A±C).
Eight minor 18S-26S rDNA loci were detected bymeiotic FISH. Four of the minor sites were associatedwith A-subgenome chromosomes, and the rest with D-
Fig. 2A, B. FISH to chromosome 5 NTs showing an intermediate18S-26S rDNA site on the left arm of chromosome 5 (5L). A A pairof intermediate 18S-26S rDNA FISH signals was located on the ex-terior of NT3R-5R (line 8±5G) barbell IVs, indicating that the FISHsites were located on arm 5L, opposite the T3R-5R breakpoint inchromosome 5. B A pair of intermediate 18S-26S rDNA FISH sig-nals was located on the side of NT5±9 (line C14±3) ring IVs oppositethe major NOR site on 9L. Combining data indicated that the previ-ous assignments of T5±9 breakpoints to 5L and 9L were incorrect;therefore they are redesignated as T5R-9R. Fig. 3A, B. FISH to chro-mosome 7 NTs showing an intermediate 18S-26S rDNA site on theleft arm of chromosome 7 (7L). A Pair of intermediate 18S-26SrDNA FISH signals was on one side of NT7L-18R (line 4659) ringIVs, indicating that the 18S±26S site is distal or opposite to thebreakpoint in chromosome 7. B An intermediate 18S±26S rDNAFISH signal occurred on both sides of NT7L-12R (line1043) ringIVs, indicating that an rDNA site was proximal to the breakpointin chromosome 7, i.e., between the centromere and the breakpointon 7L; in addition, one of the homologous minor loci of chromosome12 is barely visible (arrowhead). In conjunction with the data ofCrane et al. (1993), we can place the 18S±26S rDNA site on 7L be-tween the T6L-7L (line 1048) breakpoint and T7L-12R breakpoint.Fig. 4A, B. FISH to NT8R-19R (line 5±5B) revealed two pairs ofsignals, the relatively brighter signals from chromosome 19 (arrows)and the relatively weaker signals from chromosome 8 (arrowheads).Each pair of signals was located on one side of ring IVs, and oppositethe other pair (A), or (B) on opposite links of chain IVs. Comparisonwith the rDNA map of chromosome 19 indicates that the minorrDNA site was distal to the T8R-19R breakpoint in chromosome 8.Fig. 5A±F. FISH to chromosome 9 NTs and normal bivalents show-ing the relative positions of rDNA sites in chromosome 9. A A pairof major 18S±26S FISH signals was distal to an interior breakpoint inmegaphone IVs in chromosome 9, i.e., on the left arm of chromo-some 9 (9L); the 4',6-diamidino-2-phenylindole (DAPI) image ofthe same spread (in white frame) shows the morphology of the mega-phone IV. B A pair of 5S rDNA FISH signals occurred in the inter-stitial regions of NT2L-9R chain IVs, i.e., on the right arm of chro-mosome 9 (9R). C 18S±26S rDNA FISH signal was shown to be op-posite the T5±9 (previously designated T5L-9L) breakpoint in chro-mosome 9. Because the 18S±26S site was previously mapped to 9L,the NT5±9 breakpoint must be located on the right arm of chromo-some 9 (9R), instead of 9L. D Dual-color FISH (5S and 18S-26SrDNA probes) to NT5±9 indicating the 5S rDNA site (red, arrow-heads) in the interstitial region and the 18S-26S rDNA site (green,arrows) opposite the breakpoint. E Meiotic metaphase cell ofNT15R-16R (line 2767) showed a minor 18S±26S rDNA site (ar-rows) was on arm 9R, opposite the major NOR in a chromosome9 ring bivalent. F Weak 18S-26S rDNA FISH signals in chromosome9 (arrows) were proximal to the T9R-20L (line 2772) breakpoint, in-dicating a minor 18S±26S site on right arm of chromosome 9 (9R);another pair of minor 18S±26S rDNA FISH signals (arrowheads)was opposite the breakpoint in chromosome 20, i.e., on 20R.Fig. 6A, B. FISH to chromosomes 11 and 12 NTs showing minor18S±26S rDNA sites on the left arms of chromosomes 11 (11L)and 12 (12L). A Two pairs of 18S±26S rDNA FISH signals wereshown to be associated with NT11R-12L (line 6±5M) barbell IVs.The brighter ones (arrows) were from chromosome 11 and lay oppo-site the breakpoint, i.e., on 11L. The weaker ones (arrowheads) werefrom chromosome 12 and lay proximal to the breakpoint, i.e., on12L. B A pair of FISH signals (arrows) was on arm 11L, oppositethe T11R-13L (line10±5kb) breakpoint in chromosome 11. C Twopairs of 18S±26S rDNA FISH signals were shown to be associated
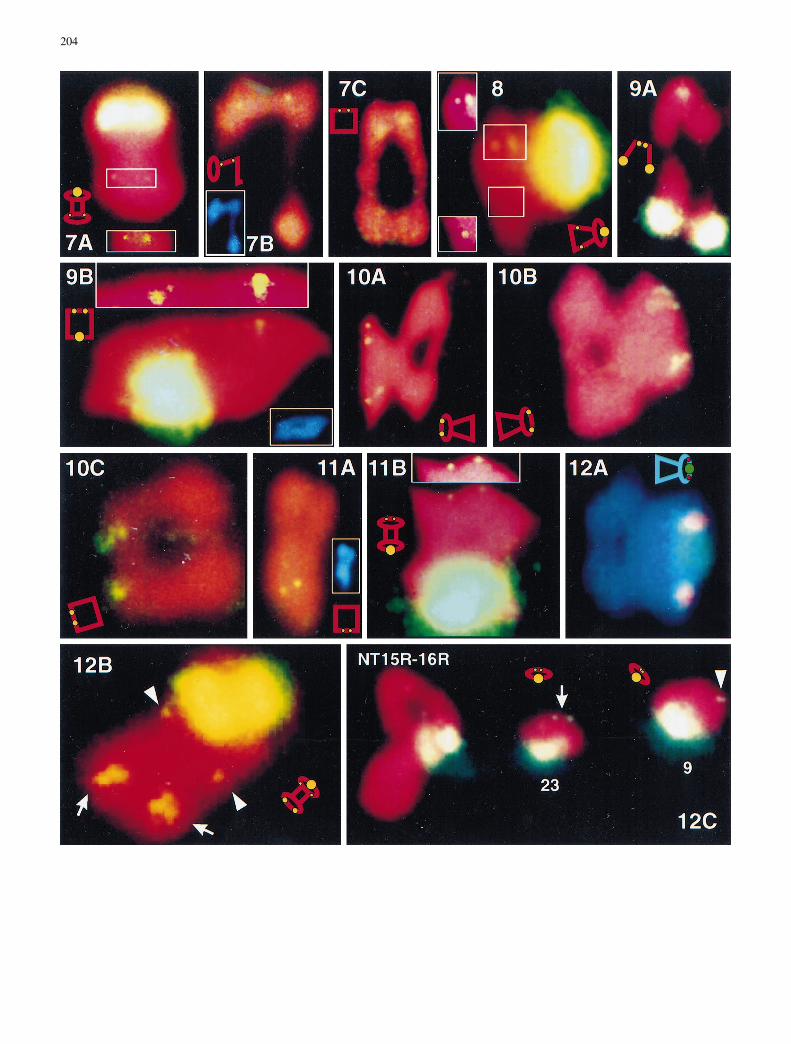
with NT11L-15L (line 1058a) ring IVs. The brighter ones (arrows)were from chromosome 11 and lay distal to the breakpoint, i.e., on11L. The weaker ones (arrowheads) were from chromosome 15and lay proximal to the breakpoint, i.e., on 15L. Fig. 7A±C. FISHto chromosome 14 NTs showing a minor 18S±26S rDNA site onthe left arm of chromosomes 14 (14L). A A pair of 18S±26S FISHsignals (in white frame) was proximal to the T14L-23R (line 2777)breakpoint in the left arm of chromosome 14 (14L). B A pair of FISHsignals was distal to the T1L-14L (line 2780) breakpoint; the DAPIimage of the same NT quadrivalent (in white frame) shows the mor-phology of the megaphone IV. C A pair of FISH signals was distal oropposite to the T14R-24R (line 2781) breakpoint. The data collec-tively indicated that the rDNA minor site is located between theT1L-14L and T14L-23R breakpoints on arm 14L. Fig. 8. FISH toNT15R-16R (line 2767) showing that a minor 18S±26S site (in whiteframe) is located in arm 15R, proximal to the chromosome 15 break-point of T15R-16R. Fig. 9A, B. FISH to chromosome 17 NTs show-ing an intermediate 18S±26S rDNA site on the right arm of chromo-somes 17 (17R). A FISH to NT9L-17Ra (line 1036) chain IVs show-ing intermediate 18S±26S rDNA FISH signals on one side of thechain opposite the major NOR on 9L, indicating an intermediate18S±26S site distal to the NT9L-17Ra breakpoint in arm 17R. BFISH to NT9L-17Rb (line 6340) ring IVs showing intermediate18S±26S rDNA FISH signals on one side opposite the major NORon 9L, indicating an intermediate 18S±26S site distal to the NT9L-17Rb breakpoint in arm 17R. Regional enhancement (inside whiteframe) shows homologous sites, otherwise camouflaged in the digi-tized image owing to the proximity of the major site. Fig. 10A±C.FISH to chromosome 19 NTs showing an intermediate 18S±26SrDNA site on the right arms of chromosome 19 (19R). A FISH toNT19L-21R (line E22±13) megaphone IVs showing FISH signalson all four chromatids of chromosome 19. B A pair of intermediateFISH signals was located to the exterior of NT19L-21R (line E22±13) megaphone IVs, indicating an intermediate 18S±26S rDNA sitein arm 19R, opposite the T19L-21R breakpoint in arm 19L. C A pairof intermediate FISH signals was located on one side of the NT4L-19R (line 10±5Ka) ring IVs, distal to the chromosome 19 breakpointof T19L-21R, assuming that the arm assignments of Menzel et al.(1985) were correct for this breakpoint. Fig. 11A, B. FISH to chro-mosome 20 NTs showing a minor 18S±26S rDNA site on the rightarms of chromosome 20 (20R). A FISH to NT20L-22R (line DP4)showing FISH signals on one side of NT20L-22R ring IV, oppositeor distal to the chromosome 20 breakpoint of T20L-22R. B FISH toNT9R-20L (line 2772) megaphone IVs showing a minor 18S±26Ssite on 20R, opposite the breakpoint in arm 20L. Fig. 12A±C. FISHto chromosome 9 NTs and normal bivalents showing the relative po-sitions of rDNA sites in chromosome 9. A Dual-color FISH toNT5R-23R (line 2775a) with 18S±26S and 5S rDNAs as probes.Both the major 18S-26S rDNA site (NOR, green) and 5S rDNA site(red) are on 23L, opposite the breakpoint in 23R. The 5S rDNA site(red) is near the centromere, and the 18S±26S site (green) is distal. BThree pairs of 18S±26S rDNA FISH signals were associated withNT5R-23R (line 2775a) barbell IVs. The brightest ones (major site,NOR) and the weakest ones (minor site, arrowheads) were fromchromosome 23, while the intermediate ones (intermediate site, ar-rows) were from chromosome 5. The major NOR was opposite thechromosome 23 breakpoint, i.e., on 23L; the minor site was proximalto the chromosome 23 breakpoint, i.e., on 23R; the intermidate (ar-rows) was opposite the chromosome 5 breakpoint, i.e., on 5L. C Mei-otic metaphase cell of NT15R-16R (line 2767) showed a minor 18S±26S rDNA site was on arm 23R, opposite the major NOR in a chro-mosome 23 ring bivalent
subgenome chromosomes. They were mapped to the rightarms of chromosomes 8 (Fig. 4A, 4B), 9 (Fig. 5E, F), 15(Figs. 6C, 8), 20 (Fig. 11A, B), and 23 (Fig. 12B, C) andthe left arms of chromosomes 11 (Fig. 6A±C), 12(Fig. 6A), and 14 (Fig. 7A±C).

206
Chromosome translocations used in the study
Results of FISH to chromosome translocations are sum-marized in Table 1 and exemplified by a detailed discus-sion of the results involving chromosome 9 transloca-tions. One of the three major 18S-26S rDNA sites(NORs) and one of the two 5S rDNA sites were mappedpreviously by ISH to chromosome 9 (Price et al. 1990;Crane et al. 1993). Price et al. (1990) mapped one NORsite to arm 9L, opposite the T9R-20L breakpoint. OurFISH analysis delimited the major 18S-26S rDNA locusto the segment distal to the T9L-17Rb breakpoint(Fig. 5A). The NOR site was also distal to the T9L-17Ra and T9L-25 breakpoints in chromosome 9, whilethe 5S rDNA site was in arm 9R (data not shown). Thus,we confirmed the tentative conclusion of Crane et al.(1993) that the 5S rDNA sites of chromosome 9 are onopposite sides of the centromere. Crane et al. (1993) ten-tatively mapped the 5S rDNA site to the segment proxi-mal to the T3R-9R breakpoint. Our FISH analysis con-firmed this, and further delimited the 5S rDNA locus tothe segment proximal to the T2L-9R (Fig. 5B) andT9R-20L (data not shown) breakpoints.
The above data confirm the arm assignments of Men-zel et al. (1985) of the chromosome 9 breakpoints forT3R-9R, T2L-9R, T9L-17Rb, and T9R-20L, as well asthe positions of rDNA loci determined by Price et al.(1990) and Crane et al. (1993). In contrast, FISH to thetranslocation heterozygote previously designated asNT5L-9L (Menzel et al. 1985) revealed that the NOR sitewas opposite the breakpoint in chromosome 9 (Fig. 5C).This finding was supported by dual-color FISH to NT5±9IVs, which yielded 5S rDNA signals in the interstitial re-gion, whereas the 18S-26S rDNA signals were in the op-posite arm (Fig. 5D). ISH analyses of monotelodisomictranslocation heterozygotes previously indicated a prob-lem with arm designations for this translocation, but didnot resolve it (Stelly et al. 1996). The new FISH data,however, unequivocally indicated that the T5±9 break-point of chromosome 9 is in arm 9R. FISH also revealedthat the T5±9 breakpoint of chromosome 5 is in arm 5R(Fig. 2B); not 5L as described by Menzel et al. (1985).Therefore, the translocation designated previously asT5L-9L is hereby redesignated as T5R-9R.
FISH also revealed a minor 18S-26S rDNA site in arm9R. A pair of weak 18S-26S rDNA FISH signals werefound opposite the major NOR in chromosome 9 biva-lents (Fig. 5E) and were shown to be proximal to thechromosome 9 breakpoint of T9R-20L (Fig. 5F), indicat-ing that a minor 18S-26S rDNA site was on the right armof chromosome 9 (9R). The weaker FISH signals in thischromosome are therefore infrequently observed becausethey are often obscured by the diffuse halo of signal sur-rounding the major 18S-26S rDNA FISH signals, so itwas difficult to map this minor site relative to all chromo-some 9 translocation breakpoints.
Discussion
This study has used meiotic FISH instead of mitotic chro-mosomes, which allows chromosome translocations to be
analyzed. This demonstrates the feasibility of using mei-otic FISH for physical mapping for crops without reliablemitotic chromosome identification. In cotton, chromo-some translocations are the primary sources for chromo-some identification. Therefore, the use of meiotic FISHconstitutes an important method for physical mapping.
Three major and one intermediate 18S-26S rDNAsites, and two 5S rDNA sites, were previously mappedby meiotic ISH in G. hirsutum L. (Price et al. 1990; Craneet al. 1993), but their positions relative to the transloca-tion breakpoints were not completely determined. Six ad-ditional minor 18S-26S rDNA loci were subsequently re-ported on somatic chromosomes (Hanson et al. 1996). Inthis study, we report four more minor sites detected bymeiotic FISH, and present an integrated cytological andphysical map of all the known rDNA sites (Fig. 1).
Several facts suggest that the observed intermediate andminor FISH signals are real rDNA sites, and that they areactive or potentially active NORs. First, the sensitivity ofour FISH detection system is now very high, with routinedetection of sites below 10 kb, and there are several in-stances where single- or low-copy target fragments in the0.8±1.5 kb range have been detected (unpublished data).Second, recurring FISH sites are usually paired and oftenappear separately on sister chromatids, indicating that theyare not background noise. Third, numerous minor or inter-mediate rDNA sites also appear on mitotic metaphase cells(Hanson et al. 1996). Fourth, multiple rDNA sites are alsoseen in many other species, for example, in wheat (Bada-eva et al. 1996). Fifth, similar improvement in FISH detec-tion has not revealed more 5S rDNA loci. Sixth, there isextensive variation among organisms, including angio-sperms, in the number of 18S-26S rDNA locations andtheir relative sizes (Hanson et al. 1996).
Besides the three major NOR sites, we observed 12 ad-ditional 18S-26S rDNA sites in preparations of meioticmetaphase I cells. The size and brightness of FISH sig-nals among these different rDNA sites differed consis-tently. Therefore, the chromosomes bearing them canbe subjectively classified into three groups. Group 1, in-cluding chromosomes 9, 16 and 23, carries the majorNORs, which can be detected in all cells and are definedas major 18S-26S rDNA sites (M). In addition, the rightarms (R) of chromosomes 9 and 23 also carry minor18S-26S sites. Group 2, including chromosomes 5, 7,17, and 19, produces FISH signals of intermediate sizeand intensity. These intermediate loci are designated I.Within group 2, signals from chromosomes 5 and 7 arebrighter than those from 17 and 19 (data not shown).Group 3, including chromosomes 8, 11, 12, 14, 15, and20, carries the weakest FISH signals, which are definedas minor 18S±26S rDNA sites (m). Within the last group,signals from chromosome 11 are brighter than those from12 (Fig. 6A) and 15 (Fig. 6C) and probably all other mi-nor sites (data not shown).
The multiplicity of rDNA sites and consistent detec-tion of many of them indicate that they will prove usefulin molecular cytogenetic studies of G. hirsutum. First, thethree major NOR sites, four intermediate 18S-26S rDNAloci, and two 5S rDNA loci will provide multiple, consis-tently detectable markers for karyological studies. Crane

207
et al. (1993) indicated that the three major sites and theintermediate site in chromosome 7 could serve to definearm locations and thus facilitate translocation-based map-ping of new loci in 18 of the 26 cotton chromosomes.Analogously, the newly localized intermediate rDNAsites in chromosomes 5, 17, and 19 allow the definitionof arm locations of new loci in all other chromosomes,except chromosomes 22 and 26. Chromosome 22 is asso-ciated with only one translocation, i.e., NT20L-22R, inwhich a minor 18S-26S site marks arm 20R. Chromo-some 26 is not involved in any of the known transloca-tions, so its arms are cytogenetically distinguishable onlyby monotelodisomy.
The distribution of rDNA genes on chromosomal armsis interesting. Two of the three major rDNA loci and twoof the four intermediate rDNA loci are on the short chro-mosomal arms, i.e., 9sh (M), 16sh (M), 5sh (I), and 7sh(I). The major site on chromosome 23 visually appearsto be on the short arm, but there are no known telosomesof chromosome 23 for a more definitive assignment. Thesame holds true for another intermediate site that seemsto be on arm 19sh rather than 19Lo. In contrast, mostof the minor sites are located on the long arms, including9Lo (m), 11Lo (m), 12Lo (m), 14Lo (m), and 20Lo (m).The distribution of rDNA loci suggests that rRNA genesin short arms may be more likely to organize nucleoli.The map of rDNA locations will facilitate site-specificanalysis of rRNA gene function and rDNA evolution, us-ing methods for identification of rDNA-containing bacte-rial artificial chromosomes (Woo 1996).
References
Alonso-Blanco C, Pendas AM, Garcia-Suarez R, Roca A, Goicoe-chea PG, Giraldez R (1994) Physical mapping of 5S rDNA re-veals a new locus on 3R and unexpected complexity in a ryetranslocation used in chromosome mapping. Chromosoma103:331±337
Appels R, Gerlach WL, Dennis ES, Swift H, Peacock WJ (1980)Molecular and chromosomal organization of DNA sequencescoding for the ribosomal RNAs in cereals. Chromosoma 78:293±311
Badaeva ED, Friebe B, Gill BS (1996) Genome differentiation inAegilops. 2. Physical mapping of 5S and 18S-26S ribosomalRNA gene families in diploid species. Genome 39:1150±1158
Bergey DR, Stelly DM, Price HJ, McKnight TD (1989) In situ hy-bridization of biotinylated DNA probes to cotton meiotic chro-mosomes. Stain Technol 64:25±37
Brown MS, Menzel MY, Hasenkampf CA, Naqi S (1981) Chromo-some configurations and orientations in 58 heterozygous trans-locations in Gossypium hirsutum. J Hered 72:161±168
Cabrera A, Friebe B, Jiang J, Gill BS (1995) Characterization ofHordeum chilense chromosomes by C-banding and in situ hy-bridization using highly repeated DNA probes. Genome 38:435±442
Castilho A, Heslop-Harrison JS (1995) Physical mapping of 5S and18S±25S rDNA and repetitive DNA sequences in Aegilops um-bellulata. Genome 38:91±96
Crane CF, Price HJ, Stelly DM, Czeschin DG (1993) Identificationof a homeologous chromosome pair by in situ DNA hybridiza-tion to ribosomal RNA loci in meiotic chromosomes of cotton(Gossypium hirsutum). Genome 36:1015±1022
Cuadrado A, Jouve N, Heslop-Harrison JS (1995) Physical mappingof the 5S rDNA multigene family in 6� triticale and rye: iden-tification of a new rye locus. Genome 38:623±626
Fominaya A, Hueros G, Loarce Y, Ferrer E (1995) Chromosomaldistribution of a repeated DNA sequence from C-genome het-erochromatin and the identification of a new ribosomal DNA lo-cus in the Avena genus. Genome 38:548±557
Hanson RE, Islam-Faridi MN, Percival EA, Crane CF, Ji Y,McKnight TD, Stelly DM, Price HJ (1996) Distribution of 5Sand 18S rDNA loci in a tetraploid cotton (Gossypium hirsutumL.) and its putative diploid ancestors. Chromosoma 105:55±61
Hutchinson J, Miller TE (1982) The nucleolar organizers of tetra-ploid and hexaploid wheats revealed by in situ hybridization.Theor Appl Genet 61:285±288
Ji Y, Raska DA, McKnight TD, Islam-Faridi NM, Crane CF, ZwickMS, Hanson RE, Price HJ, Stelly DM (1997) Use of meiotic flu-orescence in situ hybridization for identification of a new mono-some in Gossypium hirsutum L. Genome 40:34±40
Jiang J, Gill BS (1994) New 18 S±26 S ribosomal RNA gene loci:chromosomal landmarks for the evolution of polyploid wheats.Chromosoma 103:179±185
Kohel RJ, Richmond TR, Lewis CF (1970) Texas Marker-1. De-scription of a genetic standard for Gossypium hirsutum L. CropSci 10:670±671
Menzel MY, Richmond KL (1985) A revised map of chromosome15 in Gossypium hirsutum. Can J Genet Cytol 28:272±277
Menzel MY, Dougherty BJ (1987) Transmission of duplication-de-ficiencies from cotton translocations is unrelated to map lengthsof the unbalanced segments. Genetics 116:321±330
Menzel MY, Richmond KL, Dougherty BJ (1985) A chromosometranslocation breakpoint map of the Gossypium hirsutum ge-nome. J Hered 76:406±414
Menzel MY, Richmond KL, Dougherty BJ (1987) A revised map ofchromosome 16 in Gossypium hirsutum. Genome 29:823±827
Mukai Y, Endo TR, Gill BS (1991) Physical mapping of the18 S.26S rDNA multigene family in common wheat: identifica-tion of a new locus. Chromosoma 100:71±78
Price HJ, Stelly DM, McKnight TD, Scheuring CF, Raska D,Michaelson MJ, Bergey D (1990) Molecular cytogenetic mappingof a nucleolar organizer region in cotton. J Hered 81: 365±370
Schmidt T, Schwarzacher T, Heslop-Harrison JS (1994) Physicalmapping of rRNA genes by fluorescent in-situ hybridizationand structural analysis of 5S rRNA genes and intergenic spacersequences in sugar beet (Beta vulgaris). Theor Appl Genet88:629±636
Stelly DM, Crane CF, Hanson RE, McKnight TD, Price HJ (1996)Molecular-meiotic analysis of cotton. In: Jauhar PP (ed) Meth-ods of genome analysis in plants: their merits and pitfalls. CRCPress, Boca Raton, Florida, pp 99±111
Woo, S-S (1996) Development of a map-based cloning system inSorghum bicolor: 1) Isolation of megabase-size DNA and con-struction of a bacterial artificial chromosome library and 2) ge-netic and physical mapping of the 5S rDNA locus. Ph.D. disser-tation, Texas A&M University, College Station, Tex