neurotransmitter concentrations and binding at dopamine receptors in rats after maternal exposure to...
TRANSCRIPT

Environmental Toxicology and Pharmacology 9 (2001) 103–115
Neurotransmitter concentrations and binding at dopaminereceptors in rats after maternal exposure to
3,4,3%,4%-tetrachlorobiphenyl: the role of reduced thyroid hormoneconcentrations�
Astrid Roth-Harer a, Hellmuth Lilienthal a,*, Michael Bubser b, Ulrich Kronthaler b,William R. Mundy c, Thomas R. Ward c, Werner Schmidt c, Hilke Winterhoff d,
Gerhard Winneke a
a Department of Biological Psychology, Medical Institute of En6ironmental Hygiene, Aufm Hennekamp 50, D-40225 Dusseldorf, Germanyb Department of Neuropharmacology, Uni6ersity of Tubingen, Mohlstr. 54/1, D-72074 Tubingen, Germany
c Neurotoxicology Di6ision, US En6ironmental Protection Agency, MD-74B, 27711 Research Triangle Park, NC, USAd Institute of Pharmacology and Toxicology, Uni6ersity of Munster, Domagkstr.12, D-48149 Munster, Germany
Received 7 September 2000; received in revised form 20 October 2000; accepted 3 November 2000
Abstract
Polychlorinated biphenyls (PCBs) are environmental contaminants, which accumulate in the food chain and are transferred tothe offspring during prenatal development through the placenta and postnatally via breast milk. It is reported that PCBs exerteffects on thyroid hormone levels and brain neurotransmitter levels. Both actions may alter neuronal development. The aim of thepresent study was to investigate, if PCB-induced effects on concentrations of catecholamines and serotonin can be attributed toPCB-induced reductions in thyroid hormone concentrations. In addition, binding to dopamine D1 and D2 receptors wasexamined. Time-mated Wistar rats were treated prenatally with 1 mg 3,4,3%,4%-tetrachlorobiphenyl (PCB 77)/kg bodyweight or thevehicle. A third group serving as the positive control received perinatally 5 mg propylthiouracil (PTU)/l drinking water. Therewere no overt toxic signs in dams or offspring. Thyroid hormone measurements demonstrated effects in dams and offspring upto postnatal day 40. In particular, total T4 in serum and in the thyroid were decreased in PCB- and PTU-treated dams andoffspring. Only PTU exposed rats exhibited significantly increased concentrations of TSH in the serum and pituitary. Measure-ment of neurotransmitters revealed changes in the PCB-exposed offspring at PND 40, while PTU-treatment was without effect.Dopamine and DOPAC were increased in the medial prefrontal cortex. In adulthood, there were no PCB-related effects onthyroid hormones and neurotransmitters. Binding studies of dopamine D1 and D2 receptors demonstrated that PCB and PTUhad no influence on receptor concentration and affinity. Comparison of PCB 77 exposed offspring to PTU exposed offspringdemonstrated differential effects on TSH and neurotransmitter levels, the latter result indicating that not all PCB-induced effectson the nervous system can be ascribed to decreases in thyroid hormone concentrations. © 2001 Elsevier Science B.V. All rightsreserved.
Keywords: PCB; Propylthiouracil; Thyroid hormones; TSH; Neurotransmitter; Dopamine receptor
www.elsevier.com/locate/etap
� The research described in this article has been reviewed by the National Health and Environmental Effects Research Laboratory, USEnvironmental Protection Agency and approved for publication. Approval does not signify that the contents necessarily reflect the views andpolicies of the Agency nor does mention of trade names or commercial products constitute endorsement or recommendation for use.
* Corresponding author. Tel.: +49-211-3389368; fax: +49-211-3190910.E-mail address: [email protected] (H. Lilienthal).
1382-6689/01/$ - see front matter © 2001 Elsevier Science B.V. All rights reserved.PII: S 1 3 8 2 -6689 (00 )00069 -7

A. Roth-Harer et al. / En6ironmental Toxicology and Pharmacology 9 (2001) 103–115104
1. Introduction
Polychlorinated biphenyls (PCBs) continue to be anenvironmental problem because of their lipophilic prop-erties and their slow degradation. Chronic contamina-tion with PCBs results from residuals present inlandfills (Hansen et al., 1995), old transformers andcapacitors, building materials in houses, and food(DFG, 1988). The accumulation in food chains is im-portant particularly for end consumers, who are ex-posed chronically to non-acutely toxic doses over along period.
Epidemiological studies in the US and Europedemonstrated an influence by PCBs on neurodevelop-ment in children (Jacobson et al., 1984a,b, 1985; Roganet al., 1986; Gladen et al., 1988; Chen et al., 1992;Huisman et al., 1995a,b; Koopman-Esseboom et al.,1996; Winneke et al., 1998; Patandin et al., 1999). Thesestudies found PCB-related alterations in psychomotorfunction or cognitive development, which were ascribedpredominantly to prenatal exposure. This conclusionwas confirmed by a cross-fostering experiment, in whichprenatal, but not postnatal exposure to a technicalPCB-mixture resulted in impairments on two behav-ioral tasks (Lilienthal and Winneke, 1991). In animals,various types of behaviors were found to be affectedfollowing maternal exposure to PCBs, including loco-motor activity, delayed spatial alternation, active andpassive avoidance, operant behavior, and visual rever-sal learning (Tilson and Harry, 1994; overview inBrouwer et al., 1995; Schantz et al., 1995; Seo et al.,1995).
The mechanisms, by which PCBs interfere with neu-ronal development and lead to long-lasting neurobe-havioral changes, are largely unknown. PCBs arereported to decrease the serum concentrations of thy-roid hormones (Byrne et al., 1987; Collins and Capen,1980; Brouwer et al., 1990; Brouwer, 1991) by (a) anincrease in the activity of the phase II enzyme UDP-glucuronosyltransferase, which is coinduced withCYP1A1, resulting in enhanced metabolism of thyroidhormones (Barter and Klaassen, 1992; van Birgelen etal., 1995), (b) blocking the binding sites of thyroidhormones at their serum transport proteins (Brouwerand van den Berg, 1986), and (c) by impaired proteoly-sis of thyroglobulin (Collins and Capen, 1980). Of theseprocesses, the first one is due to non-ortho-substituted,coplanar congeners, while in particular, hydroxylatedPCB metabolites have been implicated in the blockingof thyroxine binding sites. Furthermore, reductions inthyroid hormone concentrations have also been foundin rat offspring following maternal exposure to PCB-mixtures and single congeners (Ness et al., 1993; Morseet al., 1995, 1996a) as well as enhanced metabolism ofthyroxine by fetal hepatic microsomes (Morse et al.,1993). Since thyroid hormones are known regulators of
neuronal development, PCB induced effects on thenervous system may be mediated by an undersupply ofthyroid hormones (Porterfield and Hendrich, 1993;Bernal and Nunez, 1995). Decreases of T4 in maternalserum result in reduced fetal T4 and T3 concentrations,particularly, in the fetal brain. These effects can bereversed by maternal infusions of T4, but not of T3
(Calvo et al., 1990). In accordance with this, hearingdeficits were detected together with hypothyroxinemiain rat offspring after developmental exposure to atechnical PCB-mixture (Goldey et al., 1995b), whichcould — at least partially — be prevented by T4
replacement (Goldey and Crofton, 1998).Other neural effects of PCBs are influences on neuro-
transmitter levels and calcium homeostasis (Seegal,1996). In vitro experiments demonstrated that only theortho-chlorinated PCB influence the dopamine and in-tracellular Ca2+ levels (Shain et al., 1991; Kodavantiand Tilson, 1997) with no effect of coplanar PCBcongeners. In contrast, in vivo experiments revealeddecreases in dopamine concentrations in the frontalcortex and caudate nucleus after perinatal exposure tothe ortho-chlorinated 2,4,2%,4%-tetraCB (PCB 47), butalso increases in dopamine in frontal cortex and sub-stantia nigra by the coplanar 3,4,3%,4%-tetraCB (PCB 77)in rats (Seegal et al., 1997). In addition, the expressionof catalepsy after application of the dopamine D2 re-ceptor blocker haloperidol was increased in rats mater-nally exposed to PCB 77 (Weinand-Harer et al., 1997).PCB-related alterations in serotonin metabolism in sev-eral brain areas of rats by the technical mixture Aroclor1254 (Morse et al., 1996b) and effects on cholinergicreceptors due to neonatal exposure of mice to theortho-chlorinated 2,5,2%,5%-tetraCB (PCB 52) or to thecoplanar PCB 77 (Eriksson et al., 1991; Eriksson andFredriksson, 1996a,b) were also described.
The purpose of this study was to examine if develop-mental decreases in thyroid hormone concentrationsafter exposure to a coplanar PCB can also account forPCB-induced neurochemical effects as they do for hear-ing impairments (Goldey et al., 1995b). Therefore, theeffects of maternal exposure to the coplanar PCB 77were compared with the effects caused by the thyro-static compound propylthiouracil (PTU). Neurobehav-ioral findings of this study related to functionalalterations in neurotransmitter systems have been re-ported previously (Lilienthal et al., 1997).
2. Materials and methods
2.1. Housing conditions and treatment schedule
Time-mated Wistar rats (Harlan–Winkelmann,Borchen, Germany) were housed in single macrolonecages with wooden chips on the floor at a 12–12 h light

A. Roth-Harer et al. / En6ironmental Toxicology and Pharmacology 9 (2001) 103–115 105
dark cycle. Light was on from 06:00 to 18:00 h. Food(Altromin, Lage, Germany) and water were availablead libitum.
Thirty-eight time-mated rats and six unmated ratswere exposed per exposure group. One group was in-jected subcutaneously with 1 mg PCB 77/kg body-weight (Promochem, Wesel, Germany; purity\99.8%)from day 7 to 18 of gestation, unmated rats wereinjected on the same days. A positive control group got6-n-propyl-2-thiouracil (PTU) (Sigma, Deisenhofen,Germany) at a concentration of 5 mg/l drinking waterfrom gestational day 7 to postnatal day 21. The PTUexposure lasted longer in comparison to the PCB expo-sure because of the rapid metabolism of PTU. Positivecontrols and the control group were injected with onlythe vehicle (olive oil).
Bodyweights of the dams were measured during ex-posure. Body weights of the offspring were measuredon the day of birth (PND 0) and on postnatal days(PND) 5, 10, 15, and 21. The offsprings were weanedand separated by sex on PND 21. Female siblings andmale siblings were housed in group macrolone cages upto six animals.
The study was conducted in accordance with theregulations of the German animal protection law andthe approval of the experiment was given by theRegierungsprasident of Dusseldorf.
2.2. Measurement of thyroid hormones and TSH
The rats were anesthetized with CO2, and immedi-ately after the breathing stopped, blood was taken byheart puncture from dams and unmated females ongestational day (GD) 19 and PND 21, and from femaleoffspring on PND 21 and male offspring on PND 40and 145. Different PCB congeners do not exert differ-ential effects on thyroid hormones in both sexes (Nesset al., 1993). Thus, different genders were used atdifferent time points in relations with other dissectionsat a given age. Blood was collected in microcentrifugetubes and allowed to clot for about 2 h. Then bloodsamples were centrifuged at 3500 rpm for 10 min.Serum was frozen and stored at −20°C until analysis.The thyroid and the pituitary were also removed fromoffspring animals, weighed and frozen in phosphatebuffer at −80°C until analysis.
The method of thyroid hormone and TSH measure-ment was described by Jekat et al., (1994). Concentra-tions of circulating total T3 and T4 (TT3 and TT4) weredetermined with RIA adapted to rat serum. Radiola-belled hormones, 125I-L-thyroxine and 125I-L-triiodothy-ronine (Du Pont de Nemours, Bad Homburg,Germany), L-thyroxine puriss. and L-triiodothyroninepuriss. (Henning, Berlin, Germany), rabbit-anti-thyrox-ine-BSA-antiserum and rabbit-anti-triiodothyronine-BSA-antiserum (bio Makor Kiryat Weizmann,Rehovot, Israel) were used for analysis.
For the determination of free T4 and T3 (FT3 andFT4) commercial RIA kits were used (Amerlex-MAB,Johnson & Johnson Clinical Diagnostics, Amersham,Braunschweig, Germany and Kodak Diagnostik,Braunschweig, Germany). Intra- and interassay coeffi-cients of variation were B10% for both assays.
For analysis of T3 and T4 in the thyroid, hormoneswere separated from thyroglobulin by enzymatic degra-dation. One lobe of the thyroid was homogenized withan UltraTurrax. Phosphate buffer (250 ml) was addedtogether with pancreatin (50 ml) to 250 ml of the ho-mogenate. Samples were incubated at 37°C for 18 h.Thereafter, they were centrifuged at 4000 rpm for 5 minand the supernatant diluted with barbital buffer (0.08mol) and 0.1% BSA (pH 8.4–8.6). Recovery was about40% with a variation of about 6% between differentanalyses in this laboratory. Concentrations of T3 andT4 were determined with RIA according to the proce-dures described above for TT3 and TT4.
TSH was labeled with the chloramine-T-method us-ing rat-homologous hormone materials, which werekindly provided by the National Institute of Diabetesand Digestive and Kidney Diseases (NIDDK, Balti-more, USA). The TSH concentrations were referred tothe reference preparation RP-1. For determination ofTSH in the pituitary, the whole pituitary was homoge-nized in phosphate buffer and distilled water. Thehomogenate (50 ml) was diluted with BSA buffer (950ml) and measured with RIA. Intra- and interassay vari-ances were 7 and 10%, respectively.
2.3. Analysis of neurotransmitters
The method of dissection and measurement was de-scribed previously by Bubser (1992). Neurotransmitterlevels were measured in dams on PND 21 and maleoffspring on PND 40 and 160. Rats were decapitatedand the brain was removed quickly and cooled inicecold saline for 60 s. Subsequently, the brain was cutin a cutting block into slices of 1.5 mm. The medialprefrontal cortex (PFC), nucleus accumbens (NAC),striatum anterior (SA), and striatum posterior (SP)were removed, weighed, quickly frozen in liquid nitro-gen, and stored at −70°C until analysis. Dopamine,3,4-dihydroxyphenylacetic acid (DOPAC), homovanil-lic acid (HVA), serotonin, and 5-hydroxyindole-3-aceticacid (5-HIAA) were measured with HPLC and electro-chemical detection. The brain regions were homoge-nized with mobile phase after adding the internalstandard 3,4-dihydroxybenzylamine (DHBA). The mo-bile phase was prepared with 6.973 g sodium acetate,7.355 g citric acid H2O, and 0.048 g Na2-EDTA · 2H2Odissolved in 930 ml H2O, vacuum filtrated (membranefilter RC-58), and sonicated in the water bath. To thissolution, 70 ml methanol (filtrated) and 0.035 g Na-oc-tansulfonic acid were added and the mixture sonicated

A. Roth-Harer et al. / En6ironmental Toxicology and Pharmacology 9 (2001) 103–115106
in a water bath. The pH of the mobile phase was set to4.65 with citric acid. The homogenate was centrifugedby 12 000×g for 1 min (Beckman Microfuge ETM).The supernatant was removed with a syringe andfiltered through a teflon filter (0.45 mm). The superna-tants of the NAC and both parts of the striatum werediluted with the mobile phase before analysis. Thefiltrate (20 ml) was injected into the HPLC equippedwith a HPLC pump (Bischoff, Leonberg, Germany),autosampler (CMA/200), analytical cell (ESA Model5011), guard cell (ESA Model 5020), and electrochemi-cal detector (ESA Coulochem, Model 5100A). Theprecolumn (¥ 3 mm) was filled with the same materialas the reversed phase column (Nucleosil 100-5-C18,5mm, 125 x 3.1 mm; Bischoff). The solvent flow ratewas set to 0.8 ml/min.
2.4. Measurement of dopamine receptors
[3H]Spiperone (18.5 Ci/nmol) and [3H]SCH23390(71.3 Ci/nmol) were purchased from DuPont NENResearch Products (Boston, MA) and N-2-hydrox-yethylpiperazine-N’-2-ethanesulfonic acid (HEPES) waspurchased from Sigma Chemical Co (St. Louis, MO).The procedure was based on the method of Mileson etal. (1991). 16 tissue samples per group were prepared atthe age of one year and 3 months in the same manneras for the neurotransmitter measurement and stored at-80°C until analysis. The tissues from two to fouranimals were pooled to obtain the approximately 40 mgwet weight and homogenized in 100 vol of HEPESbuffer (50 mM, pH 7.5) with a Tissues Tearer (BiospecProducts, Racine, WI). The homogenates were cen-trifuged for 20 min at 20,000 x g. The supernatant wasdiscarded and the pellet resuspended and washed oncemore. The final pellet was resuspended in the HEPESbuffer to a protein concentration of 0.3-0.5 mg protein/ml. Binding was assessed by incubating 0.03-0.05 mgprotein in 1 ml volume for 15 min at 35 °C with0.04-5.0 nM 3H SCH23390 (D1) or 0.006-0.70 nM 3HSpiperone (D2) in HEPES buffer. The samples wererapidly filtered and washed with ice cold buffer using aSkatron (Lier, Norway) cell harvester and counted byliquid scintillation. While initial estimates of non-spe-cific binding were made with the addition of 1 mMchlorpromazine (Research Biochemical International,Natick, MA), the final values for BMAX and KD werecalculated from the total binding using the curve fittingprogram RADLIG (Biosoft, Ferguson, MO).
2.5. Statistical analysis
The thyroid hormones and TSH levels were testedwith ANOVA for dams and unmated females and withANOVA with repeated measures for the offspring.Following significant overall effects, differences be-
tween single groups were analyzed with the REGWQpost hoc test (SAS Institute Inc., Cary, NC). Differ-ences in pituitary and thyroid weights were analyzed byANCOVA with body weight as the covariate. Effectson neurotransmitter concentrations were tested with theMann-Whitney U-test. Results of dopamine receptorbinding were analyzed with ANOVA and the Tukey’spost hoc test. Probabilities B0.05 were consideredsignificant.
3. Results
The exposure to PCB 77 or PTU did not cause grosstoxicity in dams and offspring. Number of pups andlitter weights were not significantly affected. Develop-mental data of this experiment are given in Lilienthal etal. (1997). Pituitary weights were not significantly al-tered by exposure to PCB or PTU in dams, untreatedfemales or offspring. Likewise, thyroid weights were notaltered in dams or unmated females. However, therewere exposure-related differences in offspring on PND21 according to ANCOVA with body weight as thecovariate [controls (means + CL95): 13.5 + 2.1 mg;PCB 77: 14.3 + 2.3 mg; PTU: 17.5 + 2.5 mg;F(2,20)=4.79; pB0.05]. Post hoc tests failed to revealsignificant differences between groups.
3.1. Thyroid hormones and TSH
3.1.1. Dams and unmated femalesTable 1 gives the results of thyroid hormone concen-
trations in dams, unmated females, and offspring. Inthe text only overall statistical results of ANOVA aregiven, outcome of post hoc tests is shown on Table 1 incomparison to controls, serum levels of FT4, TT4, andFT3 were significantly lower in dams exposed to PCB77 or PTU on GD 19, the day after the last treatmentwith PCB 77 [FT4: F(2,12)=20.26; pB0.0001; TT4:F(2,12)=31.65; pB0.0001; FT3: F(2,12)=4.90; pB0.05. Reductions in FT4 and TT4 were more pro-nounced in the PTU-group than in the PCB-group,while for FT3 decreases were comparable in bothgroups or even slightly more expressed in PCB dams.Levels of TT3 were not significantly different betweengroups, with the exception of an elevation in PTU-treated dams compared to PCB-exposed dams. Theratio of TT3 to TT4 was significantly higher in bothtreated groups as compared to controls [F(2,12)=33.75; pB0.0001], but this finding was also more ex-pressed in PTU-dams.
Unmated females exhibited slightly higher hormoneconcentrations than dams in all groups with the excep-tion of TT3 in the PTU-group. A remarkable differenceto dams was the lack of significant PCB- or PTU-re-lated changes in level of FT3 as well as in the ratio of

A. Roth-Harer et al. / En6ironmental Toxicology and Pharmacology 9 (2001) 103–115 107
TT3 to TT4. In addition, PTU-induced decreases inFT4 and TT4 levels were less pronounced in unmatedfemales than in dams (about 50% for FT4 and TT4, inunmated females compared to controls versus about70% and 75% for FT4 and TT4, respectively, in PTU-dams). In contrast, the percentages of PCB-inducedreductions in FT4 and TT4 concentrations were similarin unmated females and dams (about 40% for free andtotal T4 levels). Differences due to gestational statuswere significant. However, the interactions of exposurewith gestation were not statistically significant [FT4:gestation: F(1,12)=10.30; pB=0.01; expo-sure*gestation: F(2,12)=0.41; p\0.1; TT4: gestation:F(1,12)=12.09; p\0.005; exposure*gestation:F(2,12)=1.06; p\0.1].
Concentrations of TSH in serum are shown on Table2. Increased serum TSH concentrations were onlyfound in PTU-treated dams when compared to controls(F(2,12)=15.10; pB0.0005; post hoc pB0.05), whilePCB exposure was not effective (post hoc p\0.1).However, in unmated females elevated TSH values weredetected in both treated groups, but post hoc compari-sons revealed a significant difference only in the PTU-group compared to controls (pB0.05) because of thehigh variation in PCB-exposed females (p\0.1). Whencompared to dams, unmated controls had lower serumconcentrations, while in the PCB-group TSH levels in
dams were lower than those in unmated females. Thisinteraction was significant [gestation: F(1,12)=4.44;pB0.06; exposure*gestation: F(2,12)=7.44; pB0.01].
3.1.2. OffspringAt weaning on PND 21, when exposure ended for the
offspring, both FT4 and TT4 levels were reduced byPCB 77 or PTU treatment [Table 1; FT4: F(2,16)=24.53; pB0.0001; TT4: F(2,21)=39.10; pB0.0001],however, decreases by PTU were more pronounced.Significant reductions of serum FT3 values were onlydetected in PTU offspring [F(2,9)=11.88; pB0.005],while there was significant elevation of TT3 in thegroup exposed to PCB 77 as compared to controls[F(2,21)=3.54; pB0.05]. This parameter was notchanged in PTU offspring. As in dams at birth, theratio of TT3 to TT4 was raised in both treated groups[F(2,21)=7.10; pB0.005]. On PND 40, concentrationsof FT4, FT3 and TT3 had normalized in all groups, butthere were still decreases in TT4 in PCB- and PTU-ex-posed offspring [F(2,21)=12.28; pB0.0005]. This alsoresulted in an increased ratio of TT3 to TT4 in bothgroups at this time [F(2,21)=9.16; pB0.005]. On PND145, no alterations by any treatment were found inserum thyroid hormones, showing that treatment ef-fects were transient.
Table 1T4 and T3 concentrations in serum and thyroid of dams, unmated females and offspringa
FT4 (pmol/l) TT4 (nmol/l) FT3 (pmol/l) TT3 (nmol/l) TT3/TT4 *100 TH T3/T4 *100TH T3 (ng/mg)TH T4 (ng/mg)
Dams GD 19 (n=3)n.m.n.m. n.m.2.390.11.4690.104.1490.1553.2893.6012.0792.04Controls
7.2790.26* 30.8990.77* 2.8890.39* 1.2590.04PCB 77 3.490.1* n.m. n.m. n.m.10.090.2* n.m. n.m. n.m.PTU 3.8090.38* 13.1398.75* 3.2090.08* 1.5790.04
Unmated females GD 19 (n=3)2.190.4 n.m.Controls n.m.15.4790.44 n.m.66.2894.12 4.2390.31 1.6390.26
n.m.n.m.n.m.3.591.11.3590.10PCB 77 3.8990.0337.5898.75*9.3791.88*4.090.4 n.m. n.m. n.m.PTU 8.1791.16* 33.4695.02* 4.1590.34 1.5690.12
Offspring PND 21 (n=8)17.5191.46 84.8194.12Controls 4.2690.21 1.5890.06 1.690.1 358.11 9 57.05 15.06 9 1.86 4.4 9 0.17
103.59 9 48.75 *2.790.2*2.2090.11*4.0190.07 7.5 9 0.88 *70.2795.66*13.1491.68*PCB 77 6.39 9 2.71 *5.4990.40* 23.4292.45* 2.9390.19* 1.7090.16PTU 6.691.0* 17.92 9 7.16 * 7.93 9 1.50 * 59.60 9 12.06 *
Offspring PND 40 (n=8)17.5391.83 137.84910.17 4.1890.11 1.8790.09Controls 1.290.1 381.82949.24 23.3392.39 6.490.316.1990.70 96.6593.35* 4.5490.34 1.8390.12PCB 77 1.690.1* 360.54942.88 22.8992.64 6.490.217.2990.76 91.5196.56* 4.4490.20 1.8590.09PTU 1.890.2* 569.84955.66* 25.1192.18 4.590.2
Offspring PND 145 (n=3)16.8991.25 82.3794.51 3.5890.23Controls 1.4090.08 1.490.1 614.88943.61 41.1192.93 6.790.218.2892.12 84.1792.32 3.8190.06 1.5690.23 1.690.3 511.81934.86PCB 77 36.9392.09 7.390.2
3.8490.1691.2593.99 7.390.730.3493.91415.95936.32*1.390.1PTU 1.4390.0719.4490.87
a Means9S.E.M.; TH T3, TH T4=T3, T4 concentrations in the thyroid (ng/mg fresh weight); n.m.=not measured; *PB0.05 vs. controls.

A. Roth-Harer et al. / En6ironmental Toxicology and Pharmacology 9 (2001) 103–115108
Table 2TSH concentrations in serum and pituitary of dams, unmated femalesand offspringa
TSH (ng/ml) P TSH (mg per pituitary)
Dams GD 19 (n=3)Controls 4.6590.82 n.m.PCB 77 4.0490.34 n.m.PTU 13.6991.86 * n.m.
Unmated females GD 19 (n=3)Controls 2.5190.28 n.m.PCB 77 6.7192.08 n.m.PTU 6.7390.83* n.m
Offspring PND 21 (n=8)Controls 1.3790.18 4.8190.24PCB 77 1.2690.14 6.8792.22PTU 5.6590.93* 10.5990.62*
Offspring PND 40 (n=8)Controls 2.0490.59 6.6490.38PCB 77 3.7490.85 6.9190.68PTU 2.4190.29 7.9090.62
Offspring PND 145 (n=3)Controls 4.3190.54 18.1390.99PCB 77 5.6691.22 16.7791.00PTU 7.2791.62 13.5790.95*
a Means9SEM; n.m.=not measured; *PB0.05 vs. controls.
examined in dams on PND 21 (data not shown). Nore-pinephrine was elevated in the SP of the PCB group incomparison to control dams [means9CL95: controls:92944 pg/mg; PCB 77: 152956 pg/mg; PTU: 270956pg/mg; pB0.05]. Serotonin was decreased in the SA ofthe PCB-group compared to controls [means 9 CL95:controls: 6859220 pg/mg; PCB 77: 4089110 pg/mg;PTU: 6319128 pg/mg; pB0.05] and also, in the NACof the PTU group in comparison to controls [means 9CL95; controls: 14889113 pg/mg; PCB 77: 14949466pg/mg; PTU: 11799210 pg/mg; pB0.05].
3.2.2. Offspring at PND 40 and 160 (Table 3)On PND 40 there were significantly elevated levels of
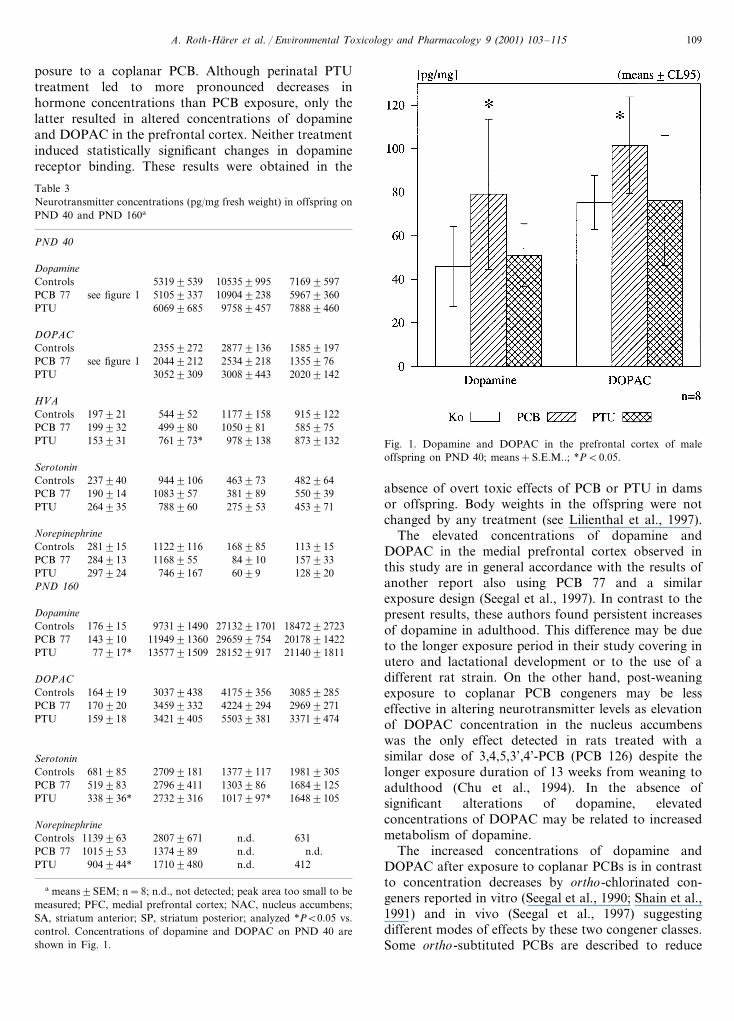
dopamine and DOPAC in the PFC of the PCB-exposedoffspring relative to controls (pB0.05) (Fig. 1). Do-pamine was also increased in the PTU group in the SPin comparison to PCB animals (pB0.01). DOPAC wassignificantly elevated in the NAC (pB0.05), SA (pB0.05), and SP (pB0.005) in the PTU-group in compari-son to PCB-exposed rats. HVA was increased in theNAC of the PTU-group relative to controls, HVA wasdecreased in the SP of the PCB-group in comparison tocontrols (pB0.05). Norepinephrine was decreased inNAC of the PTU animals relative to PCB offspring(pB0.05). Serotonin was increased in the PCB offspringin the NAC in comparison to PTU offspring (pB0.01).At PND 160 there were no changes in dopamine andDOPAC levels between PCB exposed offspring andcontrol offspring, but PTU-exposed rats had significantlydecreased dopamine levels in the PFC in comparison tocontrols and PCB exposed animals (pB0.05). DOPACwas increased in the SA of the PTU group in comparisonto controls and PCB-exposed animals (pB0.05). HVAcould not be evaluated because the peaks in the chro-matograms were too small. The levels of serotonin in thePTU group were decreased in the PFC and the SA incomparison to controls and PCB-exposed offspring (pB0.05). Norepinephrine was decreased only in the PFC ofthe PTU group in comparison to control offspring(pB0.05).
3.3. Dopamine receptors
Despite large differences between groups in somebrain areas, there were no statistically significant devia-tions from controls in D1 and D2 receptor binding.Both, the total concentration (BMAX) and the equi-librium dissociation constant KD failed to reach signifi-cance level (Table 4).
4. Discussion
These findings demonstrate transient reductions inthyroid hormone concentrations by developmental ex-
Measurements of thyroid hormones in the thyroidgenerally corroborated these observations (Table 1).Both T3 and T4 concentrations were reduced in thethyroid on PND 21. T3 was reduced to the same extentby PCB- and PTU-exposure [F(2,20)=4.83; pB0.05],while T4 reductions were more pronounced in thePTU-group [F(2,21)=16.53; pB0.0001]. Also, the ratioof T3 to T4 was increased to a much higher extent in thisgroup [F(2,2)=22.54; pB0.0001]. On PND 40, only anelevation of T4 was detected in PTU offspring, while onPND 145 T4 values in PCB- and PTU exposed rats weredecreased, but this difference was statistically significantonly in the PTU-group according to post hoc tests(pB0.05).
As in dams, increases in serum TSH concentrationswere only detected in PTU-offspring on PND 21 [Table2; F(2,21)=6.50; pB0.01]. However, the PCB-offspringexhibited elevated TSH concentrations in the pituitary bymore than 40% at this age which missed significance bythe high variability. In contrast, the increases in pituitaryTSH in the PTU-group were significant [F(2,20)=5.92;pB0.01]. No group differences were found on PND 40,while on PND 145 PTU-offspring had lower TSH valuesin the pituitary [F(2,20)=5.67; pB0.05].
3.2. Neurotransmitters
3.2.1. Dams on PND 21There were no exposure-related changes in dopamine,
DOPAC, and HVA concentrations in any brain area
A. Roth-Harer et al. / En6ironmental Toxicology and Pharmacology 9 (2001) 103–115 109
posure to a coplanar PCB. Although perinatal PTUtreatment led to more pronounced decreases inhormone concentrations than PCB exposure, only thelatter resulted in altered concentrations of dopamineand DOPAC in the prefrontal cortex. Neither treatmentinduced statistically significant changes in dopaminereceptor binding. These results were obtained in the
Fig. 1. Dopamine and DOPAC in the prefrontal cortex of maleoffspring on PND 40; means+S.E.M..; *PB0.05.
Table 3Neurotransmitter concentrations (pg/mg fresh weight) in offspring onPND 40 and PND 160a
PND 40
Dopamine105359995 71699597Controls 53199539
see figure 1PCB 77 5967936051059337 109049238PTU 788894609758945760699685
DOPAC23559272Controls 28779136 15859197
PCB 77 25349218see figure 1 20449212 1355976PTU 202091423008944330529309
HVA11779158Controls 9159122197921 544952
499980199932 1050981 585975PCB 7787391329789138761973*153931PTU
Serotonin482964Controls 4639739449106237940550939PCB 77 190914 1083957 381989
275953 453971PTU 264935 788960
Norepinephrine11391516898511229116Controls 281915
PCB 77 1168955 84910 157933284913297924PTU 1289207469167 6099
PND 160
Dopamine176915 973191490Controls 2713291701 1847292723
2965997541194991360143910 2017891422PCB 77PTU 77917* 1357791509 281529917 2114091811
DOPAC41759356Controls 30859285164919 30379438
PCB 77 34599332 42249294 2969927117092034219405159918 55039381 33719474PTU
SerotoninControls 27099181 13779117 19819305681985
519983 27969411PCB 77 1303986 1684912516489105338936* 1017997*PTU 27329316
Norepinephrinen.d. 631Controls 1139963 28079671
1015953 n.d.PCB 77 n.d.1374989n.d.17109480904944*PTU 412
a means9SEM; n=8; n.d., not detected; peak area too small to bemeasured; PFC, medial prefrontal cortex; NAC, nucleus accumbens;SA, striatum anterior; SP, striatum posterior; analyzed *PB0.05 vs.control. Concentrations of dopamine and DOPAC on PND 40 areshown in Fig. 1.
absence of overt toxic effects of PCB or PTU in damsor offspring. Body weights in the offspring were notchanged by any treatment (see Lilienthal et al., 1997).
The elevated concentrations of dopamine andDOPAC in the medial prefrontal cortex observed inthis study are in general accordance with the results ofanother report also using PCB 77 and a similarexposure design (Seegal et al., 1997). In contrast to thepresent results, these authors found persistent increasesof dopamine in adulthood. This difference may be dueto the longer exposure period in their study covering inutero and lactational development or to the use of adifferent rat strain. On the other hand, post-weaningexposure to coplanar PCB congeners may be lesseffective in altering neurotransmitter levels as elevationof DOPAC concentration in the nucleus accumbenswas the only effect detected in rats treated with asimilar dose of 3,4,5,3’,4’-PCB (PCB 126) despite thelonger exposure duration of 13 weeks from weaning toadulthood (Chu et al., 1994). In the absence ofsignificant alterations of dopamine, elevatedconcentrations of DOPAC may be related to increasedmetabolism of dopamine.
The increased concentrations of dopamine andDOPAC after exposure to coplanar PCBs is in contrastto concentration decreases by ortho-chlorinated con-geners reported in vitro (Seegal et al., 1990; Shain et al.,1991) and in vivo (Seegal et al., 1997) suggestingdifferent modes of effects by these two congener classes.Some ortho-subtituted PCBs are described to reduce

A. Roth-Harer et al. / En6ironmental Toxicology and Pharmacology 9 (2001) 103–115110
tyrosine hydroxylase activity in striatal tissue in vitro(Choksi et al., 1997), but in PC12 cells effects ofortho-chlorinated congeners on the activity of L-aro-matic amino acid decarboxylase, converting the pre-cursor DOPA to dopamine, are more pronounced andwere detected at lower treatment levels than reductionsin activity of tyrosine hydroxylase (Angus et al., 1997).At present, the mechanism by which coplanar PCBs leadto increased concentrations of dopamine is not yetknown. The effect may be related to the formation ofmetabolites which are rapidly formed from PCB 77 (e.g.Morse et al., 1995) and are likely to enter the brain bya transthyretin-related mechanism. Since PCB-metabo-lites are known to bind at estrogen receptors (Korach etal., 1988) and since estradiol is reported to modulatedopamine release in the brain (Becker and Beer 1986),this may constitute a further mechanism by which PCB77 leads to influences on the nervous system.
Neither PCB 77 nor PTU caused significant effects onD1 and D2 receptor binding. In some cases there werelarge deviations of values in exposed groups in compari-son to controls, e.g. D1- and D2-related Bmax and KDin the prefrontal cortex, but due to the small amount oftissue in this brain region and the resulting small numberof independent probes (4 samples from different animalshad to be pooled for each of the four different probes)the variability was too high to reach statistical signifi-cance. However, one cannot rule out transient effects onreceptor binding since these samples were taken from ratswith an age of more than one year. Also from functionalstudies there is not yet clear evidence for PCB-inducedeffects on the dopaminergic system. A drug discrimina-tion experiment in littermates of rats used in the presentstudy failed to detect differences between groups using
agonists and antagonists of the D2 receptor (Lilienthalet al., 1997). However, differences were detected in ablocking test with buspirone, an antagonist of the D2receptor and agonist of the serotonin 1A receptor. Both,the PCB 77 and the PTU group exhibited less blockingthan controls. This suggests alterations in the interactionbetween the serotonergic and the dopaminergic system,but PCB effects on processes related to the D1 receptorcannot be precluded from this experiment. Influences onthe interaction between dopamine and serotonin mayalso explain PCB 77 related effects on haloperidol-in-duced catalepsy (Weinand-Harer et al., 1997) sinceserotonin 1A agonists are known to modify catalepsycaused by D2 agonists (Wadenberg, 1996). In generalaccordance with this, gestational exposure to a technicalmixture of PCBs (Aroclor 1254) did not affect levels ofdopamine and its metabolites, but increased themetabolism of serotonin in different brain areas (Morseet al., 1996b). Also, a study in adult rats with exposureto Aroclor 1254 indicated enhanced metabolism ofbiogenic amines because of elevated concentrations ofdopamine metabolite DOPAC and of the serotoninmetabolite 5-HIAA, while dopamine and serotonin werenot altered (Kodavanti et al., 1998). In conclusion, moredetailed studies are needed to elucidate effects on PCBon dopaminergic and serotonergic systems, includingneurotransmitter release, receptor binding and, in partic-ular, resulting functions.
This hold also true for the possible implication ofthyroid hormones in PCB-related neurochemical effects.In contrast to PCB 77, there were no effects on dopamineand DOPAC concentrations in the prefrontal cortex byPTU in offspring on PND 40. However, unlike in PCB77 exposed rats there was a decrease of dopamine in adult
Table 4D1 and D2 Receptors — BMAX (fmole/mg protein) and KD (nM) values for binding of [3H]SCH23390 (D1) and [3H]Spiperone (D2)a
NAC (n=5) SP (n=8)SA (n=8)PFC (n=4)
[3H]SCH 23390BMAX
496953234917145942Controls 3649262739169 359937418947191917PCB 77316986 219918PTU 489956 348925
KD (nM)0.03190.0090.02890.0020.06990.024 0.02090.004Controls
0.07690.058 0.02490.002PCB 77 0.02390.006 0.02790.0040.03790.0060.10390.031 0.02290.006PTU 0.02390.004
[3H ]SpiperoneBMAX
167918 1116997730921Controls 1399991909976213960 616972 1336982PCB 77
677981 1005959182910 1476946PTUKD (nM)
0.24190.012 0.23290.064Controls 0.20990.0230.21790.0480.21590.0130.31190.104 0.23290.019PCB 77 0.22790.013
0.23490.0210.23390.0120.21390.0090.21890.016PTU
a means9SEM; PFC, medial prefrontal cortex; NAC, nucleus accumbens; SA, striatum anterior; SP, striatum posterior.

A. Roth-Harer et al. / En6ironmental Toxicology and Pharmacology 9 (2001) 103–115 111
PTU-exposed offspring on PND 160. Thus, these resultsdemonstrate that, despite reduced thyroid hormone con-centrations by PTU and PCB 77 exposure, they differ intheir effects on neurotransmitter levels, both in the ageat which effects are apparent as well as in the directionof the influence. These differences cannot be attributedto the differences in exposure periods lasting from GD7to GD 18 in the PCB 77 group, while in the PTU groupduration of exposure was extended to PND 21 due to therapid metabolism of this compound. Although half-lifeof PCB 77 is also short in adult rats (McKinley et al.,1993), it is considerably longer in weanling rats rangingfrom 6.33 to 7.75 days (Saghir et al., 1999) and may beeven longer during pre- and postnatal development.Previous results from our laboratory using the same ora similar exposure schedule demonstrated transfer ofPCB 77 to pups across the placenta and via the milk(Weinand-Harer et al., 1997; Hany et al., 1999). Inaddition, weanling offspring of rat dams which weregiven a single treatment with PCB 77 on PND 1 exhibitedinduction of CYP4A isozymes at weaning, showing thatlactational transfer is sufficient to induce biochemicalchanges in offspring (Borlakoglu et al., 1993).
While effects of PTU and PCB 77 on neurotransmitterconcentrations are different, similar influences on theinteraction of dopaminergic and serotonergic processeswere observed in littermates of these rats in the drugdiscrimination study mentioned above (Lilienthal et al.,1997). As an enhanced serotonin metabolism was de-tected after experimentally induced hypothyrosis duringdevelopment in rats (Savard et al., 1984) and decreasedserotonin concentrations were found in the PTU groupin the present experiment on PND 160, a possiblemediation by thyroid hormone-related processes of PCB-induced effects on the serotonergic system requiresfurther attention. The neurochemical findings of Morseet al. (1996b) are compatible with these results. In thepresent study no PCB-related influence on serotonin wasfound, but effects might have been too subtle to bedetected by post mortem analysis of neurotransmitterconcentrations.
In addition to similarities found in drug discrimina-tion, similar effects on auditory functions and behaviorwere reported in rats after developmental exposure toPTU or a technical PCB-mixture (Goldey et al., 1995a,b).Effects by PCB-exposure could at least be partiallyprevented by T4 replacement (Goldey and Crofton 1998),thereby indicating that these PCB-induced effects aremediated by reduced concentrations of thyroid hor-mones.
The effects of PCBs on concentrations of thyroidhormones found in the present study, showing decreasedvalues for FT4, TT4, and FT3 in dams and, with theexception of FT3, also in offspring at the end of theexposure period which was GD19 and PND21 in damsand pups, respectively, are in general accordance with
other studies examining the effects of single PCB con-geners (Morse et al., 1995) and technical mixtures(Goldey et al., 1995b) in pregnant animals and offspring.In comparison to unmated females, dams in our studyexhibited generally lower values of FT4 and TT4, whichis in line with previous observations (Morse et al., 1995).In contrast to unmated females, significantly elevatedratios of TT3/TT4, were detected in PCB- and PTU-ex-posed dams. This indicates an enhanced activation tomeet the increased requirements for thyroid hormones inboth treated groups during gestation (Burrow 1993) tocompensate for lowered T4. The activation may becaused by an increased activity of type II-5’-deiodinasewhich is found in brain, pituitary, brown fat, andplacenta in rats (Larsen et al., 1998), while type I-5’-deio-dinase has been found to be inhibited by metabolites ofPCB 77 (Adams et al., 1990) and after exposure toAroclor 1254 (Hood and Klaassen 2000). The latter studyreported that the resulting net effect of increased activityof type II-5’-deiodinase and decreased type I-5’-deiodi-nase is an overall reduction in serum T3 concentrationsand concluded that activation of the type II-enzymecannot be the reason for normal T3 values which,therefore, remains to be elucidated.
In the offspring a similar pattern of changes in thyroidhormones was detected with the exception of normal FT3values in the PCB 77 group. This may be achieved byenhanced secretion of thyroidal T3, since T3 concentra-tion in the thyroid were reduced by nearly 60% in thisgroup. Similar compensating processes were observed iniodine deficiency (Larsen et al., 1998). As no differencesin thyroid weights between controls and PCB 77 off-spring were observed, the hormone reductions cannot beattributed to altered organ weights. In PTU rats thyroidweight was even raised in comparison to controls. Theactivation as expressed by a raised ratio of T3/T4 anddecreased values of TT4 was still detectable at PND 40in the offspring, indicating that normalization may beachieved only at later ages.
In contrast to PTU-treatment, PCB-induced reduc-tions in thyroid hormones were observed in the absenceof elevated serum concentrations of TSH in dams andoffspring. There was also no elevation of TSH in thepituitary in the offspring. The lack of effect on serumTSH has been described in male as well as pregnant andnon-pregnant female rats after exposure to coplanarcongeners (Seo et al., 1995), Aroclor 1254 (Goldey et al.,1995b; Morse et al., 1996a), or TCDD (Gorski et al.,1988; Schuur et al., 1997). Increases in TSH may bepresent only at higher exposure levels leading to nearlycomplete depletion of T4 in rats (Barter and Klaassen1994; Liu et al., 1995) and marmosets (van den Berg etal., 1988). Other thyroid hormone deficiencies with lowT4, normal or slightly lower T3 and normal TSH havebeen described after phenytoin or salicylate treatment orin central hypothyroidism (Larsen et al., 1998). Centralhypothyroidism can be due to pituitary or hypothalamic

A. Roth-Harer et al. / En6ironmental Toxicology and Pharmacology 9 (2001) 103–115112
failure. To gain insight into the lack of effects on TSHa more detailed analysis of the responsiveness of thehypothalamus-pituitary-thyroid axis to stimulation withTRH, TSH, and T3 in PCB-exposed animals is needed.
In comparison to dams, fetal thyroid hormone levelsappear to be more and for a longer period affected byexposure to Aroclor 1254 (Morse et al., 1996a) and PCB77 (Morse et al., 1995). Particularly, in the latter case thismay be due to the formation of metabolites some ofwhich accumulate to higher levels in fetal plasma andtissues than in dams (Darnerud et al., 1986, 1996; Lanset al. 1993; Morse et al., 1995). Certain metabolites bindto the serum transport protein for thyroid hormonestransthyretin (TTR) with a higher affinity than T4 (Lanset al., 1993; Darnerud et al., 1996). Since TTR issuggested to be essentially involved in the transport ofT4 across the choroid plexus (Lans et al., 1993; Koop-man-Esseboom et al., 1994), a displacement of T4 fromthe binding domain of TTR could lead to a decreasedsupply with T4 even after the start of fetal thyroidhormone production on GD 17 (Porterfield and Hen-drich, 1993). However, recently normal T4 and T3concentrations were detected in the brain tissue ofTTR-null mutant mice, suggesting that other transportmechanisms may be responsible for the transport ofthyroid hormones into the brain (Palha et al., 2000).Future studies should determine T4 and T3 concentra-tions in the brain of PCB-exposed animals.
A diminished supply of the fetal brain with T4 can becounteracted by an increased activity of type II-5’-deio-dinase which results in a higher local conversion of T4into the active form T3. This has been demonstrated bymeasurements in brain tissue from PCB-exposed neona-tal and weanling rats (Morse et al., 1993, 1996a). In fetalrats, it has been shown that type II-5’-deiodinase activityincreases in response to hypothyroidism after the onsetof fetal thyroid hormone production. However, thisprocess may be limited to certain brain regions as typeII-5’-deiodinase activity was reported to be high incerebellum, basal ganglia, brainstem, and hypothalamusat birth, but low in cortex and olfactory bulb (Koddinget al., 1986). Therefore, a region-specific determinationof type II-5’-deiodinase activity in fetal brains of PCB-ex-posed rats would give more insight into the role ofthyroid hormones and effects of lowered hormone levelson brain development.
It has been shown that on GD 15 T3 levels in the fetalbrain are critically dependent on maternal T4 levels inplasma. Supplementation of T4 in thyroid hormone-defi-cient dams restored T3 in fetal brains, while even toxicdoses of T3 given to dams were not effective (Calvo etal., 1990). Since thyroid hormones are known regulatorsduring development of the nervous system, such distur-bances can lead to impaired neural function and distur-bances of behavior, learning and memory (Porterfieldand Hendrich, 1993; Porterfield, 1994; Bernal and
Nunez, 1995). In humans, it has been shown that despiteearly treatment neuropsychological development is af-fected in children with congenital hypothyroidism (Revet1999) and that lower concentrations of T4 in womenduring the second trimester of pregnancy accounted forIQ reductions in their children (Haddow et al., 1999).
The results of this study demonstrate that a coplanarPCB congener, PCB 77, given maternally from gesta-tional day 7 to 18, resulted in decreases of thyroidhormones in dams and in offspring at PND 21 and 40.Thyroid hormone alterations in offspring did not persistinto adulthood. Also, an increase of dopamine andDOPAC in the medial prefrontal cortex was observed atPND 40 but not at PND 160. These neurochemicalchanges were not detected in the PTU-treated positivecontrol group. Changes in thyroid hormones and do-pamine may be associated with behavioral changes seenin adult animals (Lilienthal et al., 1997; Weinand-Hareret al., 1997). We suggest that after maternal PCBexposure some behavioral changes are related to thedecreased supply with thyroid hormones induced bycertain PCB congeners, while other PCB-induced effectson neurobehavioral functions cannot be explained by thismechanism. The exact nature of PCB-induced reductionsin thyroid hormone levels during development and itsrelation to different neuronal endpoints and behavioraltasks remains to be clarified.
Acknowledgements
The authors thank Franzis Boßhammer and BrigitteKorte for excellent technical assistance during hormonemeasurements, Hildegard Huß and Michael Lieverz forexcellent assistance during dissections. Furthermore, wewould like to thank Drs Kevin Crofton and PrasadaKodavanti for their critical comments on an earlierversion of this article.
References
Adams, C., Lans, C., Klasson Weheler, E., van Engelen, J.G.M.,Visser, T.J., Brouwer, A., 1990. Hepatic thyroid hormone 5%-deio-dinase, another target-protein for monhydroxy metabolites of3,3%,4,4%-tetrachlorobiphenyl. In: Hutzinger, O., Fielder, H. (Eds.),Organohalogen Compounds. Ecoinforma Press, Bayreuth, p. 51.
Angus, W.G.R., Mousa, M.A., Vargas, V.M., Quensen, J.F., Boyd,S.A., Contreras, M.L., 1997. Inhibition of L-aromatic amino aciddecarboxylase by polychlorinated biphenyls. Neurotoxicology 18,857.
Barter, R.A., Klaassen, C.D., 1992. UDP-glucuronosyltransferaseinducers reduce thyroid hormone levels in rats by an extrathy-roidal mechanism. Toxicol. Appl. Pharmacol. 113, 36.
Barter, R.A., Klaassen, C.D., 1994. Reduction of thyroid hormonelevels and alteration of thyroid function by four representativeUDP-glucuronosyltransferase inducers in rats. Toxicol. Appl.Pharmacol. 128, 9.

A. Roth-Harer et al. / En6ironmental Toxicology and Pharmacology 9 (2001) 103–115 113
Becker, J.B., Beer, M.E., 1986. The influence of estrogen on nigrostri-atal dopamine activity: Behavioral and neurochemical evidence forboth pre- and postsynaptic components. Behav. Brain Res. 19,27–33.
van den Berg, K.J., Zurcher, C., Brouwer, A., 1988. Effects of3,4,3%,4%-tetrachlorobiphenyl on thyroid function and histology inmarmoset monkeys. Toxicol. Lett. 41, 77.
Bernal, J., Nunez, J., 1995. Thyroid hormones and brain development.Eur. J. Endocrinol. 133, 390.
van Birgelen, A.P.J.M., Smit, E.A., Kampen, I.M., Groeneveld, C.N.,Fase, K.M., van der Kolk, J., Poiger, H., van den Berg, M.,Koeman, J.H., Brouwer, A., 1995. Subchronic effects of 2,3,7,8-TCDD or PCBs on thyroid hormone metabolism: use in riskassessment. Eur. J. Pharmacol. Environ. Toxicol. Pharmacol.Section 293, 77.
Borlakoglu, J.T., Henderson, C.J., Wolf, C.R., 1993. Lactationaltransfer of 2,4,5,2%,4%,5%-hexachlorobiphenyl but not 3,4,3%,4%-tetra-chlorobiphenyl induces neonatal CYP4A1. Biochem. Pharmacol.45, 769.
Brouwer, A., van den Berg, K.J., 1986. Binding of a metabolite of3,4,3%,4%-tetrachlorobiphenyl to transthyretin reduces serum vitaminA transport by inhibiting the formation of protein complex carryingboth retinol and thyroxin. Toxicol. Appl. Pharmacol. 85, 301.
Brouwer, A., Klasson-Wehler, E., Bokdam, M., Morse, D.C., Traag,W.A., 1990. Competitive inhibition of thyroxin binding totransthyretin by monohydroxy metabolites of 3,4,3%,4%-tetrachloro-biphenyl. Chemosphere 20, 1257.
Brouwer, A., 1991. The role of enzymes in regulating the toxicity ofxenobiotics. Biochem. Soc. Trans. 19, 731.
Brouwer, A., Ahlborg, U.G., Van den Berg, M., Birnbaum, L.S.,Boersma, E.R., Bosveld, B., Denison, M.S., Earl Gray, L., Hagmar,L., Holene, E., Huisman, M., Jacobson, S.W., Jacobson, J.L.,Koopman-Esseboom, C., Koppe, J.G., Kulig, B.M., Morse, D.C.,Muckle, G., Peterson, R.E., Sauer, P.J.J., Seegal, R.F., Smits-VanProoije, A.E., Touwen, B.C.L., Weisglas-Kuperus, N., Winneke,G., 1995. Functional aspects of developmental toxicity of polyhalo-genated aromatic hydrocarbons in experimental animals and hu-man infants. Europ. J. Pharmacol. Environ. Toxicol. Section 293,1.
Bubser, M., 1992. Verhaltenspharmakologische und neurochemischeCharakterisierung der Funktion von Dopamin im medialenprafrontalen Cortex der Ratte. Dissertation, Biologisches Institutder Universitat Stuttgart.
Burrow, G.N., 1993. Thyroid function and hyperfunction duringgestation. Endocr. Rev. 14, 194.
Byrne, J.J, Carbone, J.P., Hanson, E.A., 1987. Hypothyroidism andabnormalities in the kinetics of thyroid hormone metabolism in ratstreated chronically with polychlorinated biphenyl and polybromi-nated biphenyl. Endocrinology 121, 520.
Calvo, R., Obregon, M.J., Ruiz de Ona, C., Escobar del Rey, F.,Morreale de Escobar, G., 1990. Congenital hypothyroidism, asstudied in rats. J. Clin. Invest. 86, 889.
Chen, Y.-C.J., Guo, Y.-L., Hsu, C.-C., Rogan, W.J., 1992. Cognitivedevelopment of Yu-Cheng (‘oil disease’) children prenatally ex-posed to heat-degraded PCBs. J. Am. Med. Assoc. 268, 3213.
Choksi, N.Y., Kodavanti, P.R.S., Tilson, H.A., Booth, R.G., 1997.Effects of polychlorinated biphenyls (PCBs) on brain tyrosinehydroxylase activity and dopamine synthesis in rats. Fund. Appl.Toxicol. 39, 76.
Chu, I., Villeneuve, D.C., Yagminas, A., LeCavalier, P., Poon, R.,Feeley, M., Kennedy, S.W., Seegal, R.F., Hakansson, H., Ahlborg,U.G., Valli, V.E., 1994. Subchronic toxicity of 3,3%,4,4%,5-pen-tachlorobiphenyl in the rat. Fundam. Appl. Toxicol. 22, 457.
Collins, W.T., Capen, C.C., 1980. Fine structural lesions and hormonalalterations in thyroid glands of perinatal rats exposed in utero andby the milk to polychlorinated biphenyls. Am. J. Pathol. 99, 125.
Darnerud, P.O., Brandt, I., Klasson-Wehler, E., Bergman, A., D’Ar-gry, R., Dencker, L., Sperber, G.O., 1986. 3,3%,4,4%-tetra-chloro[14C]biphenyl in pregnant mice: enrichment of phenol andmethyl sulphone metabolites in late gestational fetuses. Xenobiotica16, 295.
Darnerud, P.O., Morse, D., Klasson-Wehler, E., Brouwer, A., 1996.Binding of a 3,3%,4,4%-tetrachlorobiphenyl (CB-77) metabolite tofetal transthyretin and effects on fetal thyroid hormone levels inmice. Toxicology 106, 105.
DFG (Deutsche Forschungsgemeinschaft) 1988. PolychlorierteBiphenyle, Bestandsaufnahme uber Analytik, Vorkommen, Kinetikund Toxikologie. VCH Verlagsgesellschaft mbH, Weinheim.
Eriksson, P., Fredriksson, A., 1996a. Developmental neurotoxicity offour ortho-substituted polychlorinated biphenyls in the neonatalmouse. Environ. Toxicol. Pharmacol. 1, 155.
Eriksson, P., Fredriksson, A., 1996b. Neonatal exposure to 2,2%,5,5%-te-trachlorobiphenyl causes increased susceptibility in the cholinergictransmitter system at adult age. Environ. Toxicol. Pharmacol. 1,217.
Eriksson, P., Lundkvist, U., Fredriksson, A., 1991. Neonatal exposureto 3,3%,4,4%-tetrachlorobiphenyl: changes in spontaneous behaviourand cholinergic muscarinic receptors in the adult mouse. Toxicol-ogy 69, 27.
Gladen, B.C., Rogan, W.J., Hardy, P., Thullen, J., Tingelstad, J., Tully,M., 1988. Development after exposure to polychlorinated biphenylsand dichlorodiphenyl dichloroethene transplacentally and throughhuman milk. J. Pediatr. 113, 991.
Goldey, E.S., Crofton, K.M., 1998. Thyroxine replacement attenuateshypothyroxinemia, hearing loss, and motor deficits following devel-opmental exposure to Aroclor 1254 in rats. Toxicol. Sci. 45, 94.
Goldey, E.S., Kehn, L.S., Rehnberg, G.L., Crofton, K.M., 1995a.Effects of developmental hypothyroidism on auditory and motorfunction in the rat. Toxicol. Appl. Pharmacol. 135, 67.
Goldey, E.S., Kehn, L.S., Lau, C., Rehnberg, G.L., Crofton, K.M.,1995b. Developmental exposure to polychlorinated biphenyls (Aro-clor 1254) reduces circulating thyroid hormone concentrations andcauses hearing deficits in rats. Toxicol. Appl. Pharmacol. 135, 77.
Gorski, J.R., Muzi, G., Weber, L.W.D., Pereira, D.W., Arceo, R.J.,Iatropoulos, M.J., Rozman, K., 1988. Some endocrine and mor-phological aspects of acute toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Toxicol. Pathol. 16, 313.
Haddow, J.E., Palomaki, G.E., Allan, W.C., Williams, J.R., Knight,G.J., Gagnon, J., O’Heir, C.E., Mitchell, M.L., Hermos, R.J.,Waisbren, S.E., Faix, J.D., Klein, R.Z., 1999. Maternal thyroiddeficiency during pregnancy and subsequent neuropsychologicaldevelopment of the child. New Engl. J. Med. 341, 549.
Hansen, L.G., Li, M.-H., Saeed, A., Bush, B., 1995. Environmentalpolychlorinated biphenyls: acute toxicity of landfill soil extract tofemale prepubertal rats. Arch. Environ. Contam. Toxicol. 29, 334.
Hany, J., Lilienthal, H., Roth-Harer, A., Ostendorp, G., Heinzow, B.,Winneke, G., 1999. Behavioral effects following single and com-bined exposure to PCB 77 (3,4,3%,4%-tetrachlorobiphenyl) and PCB47 (2,4,2%,4%-tetrachlorobiphenyl) in rats. Neurotoxicol. Teratol. 21,147.
Hood, A., Klaassen, C.D., 2000. Effects of microsomal enzymeinducers on outer-ring deiodinase activity toward thyroid hormonesin various rat tissues. Toxicol. Appl. Pharmacol. 163, 240.
Huisman, M., Koopman-Esseboom, C., Fidler, V., Hadders-Algra, M.,van der Paauw, C.G., Tuinstra, L.G.M.T., Weisglas-Kuperus, N.,Sauer, P.J.J., Touwen, B.C.L., Boersma, E.R., 1995a. Perinatalexposure to polychlorinated biphenyls and dioxins and its effect onneonatal neurological development. Early Hum. Develop. 41, 111.
Huisman, M., Koopman-Esseboom, C., Lanting, C.I., van der Paauw,C.G., Tuinstra, L.G.M.Th., Fidler, V., Weisglas-Kuperus, N.,Sauer, P.J.J., Boersma, E.R., Touwen, B.C.L., 1995b. Neurologicalcondition in 18-month-old children perinatally exposed to polychlo-rinated biphenyls and dioxins. Early Hum. Dev. 43, 165.

A. Roth-Harer et al. / En6ironmental Toxicology and Pharmacology 9 (2001) 103–115114
Jacobson, J.L., Fein, G., Jacobson, S.W., Schwartz, P.M., Dowler,J.K., 1984a. The transfer of polychlorinated biphenyls (PCBs) andpolybrominated biphenyls (PBBs) across the human placenta andinto maternal milk. J. Public Health 74, 378.
Jacobson, J.L., Fein, G., Schwartz, P.M., Dowler, J.K., 1984b.Prenatal exposure to an environmental toxin: a test of the multi-ple effects model. Dev. Psychol. 20, 523.
Jacobson, S.W., Fein, G., Jacobson, J.L., Schwartz, P.M., Dowler,J.K., 1985. The effect of intrauterine PCB exposure on visualrecognition memory. Child Dev. 56, 853.
Jekat, F.W., Meisel, M.L., Eckard, R., Winterhoff, H., 1994. Effectsof pentachlorophenol (PCP) on the pituitary and thyroidal hor-mone regulation in the rat. Toxicol. Lett. 71, 9.
Kodavanti, P.R.S., Tilson, H.A., 1997. Structure-activity relation-ships of potentially neurotoxic PCB congeners in the rat. Neuro-toxicology 18, 425.
Kodavanti, P.R.S., Derr-Yellin, E.C., Mundy, W.R., Shafer, T.J.,Herr, D.W., Barone, S.Jr., Choksi, N.Y., MacPhail, R.C., Tilson,H.A., 1998. Repeated exposure of adult rats to Aroclor 1254causes brain region-specific changes in intracellular Ca2+ buffer-ing and protein kinase C activity in the absence of changes intyrosine hydroxylase. Toxicol. Appl. Pharmacol. 153, 186.
Kodding, R., Fuhrmann, H., von zur Muhlen, A., 1986. Investiga-tions on iodothyronine deiodinase activity in the maturing ratbrain. Endocrinology 118, 1347.
Koopman-Esseboom, C., Morse, D.C., Weisglas-Kuperus, N.,Lutkeschipholt, I.J., van der Paauw, C.G., Tuinstra, L.G.M.T.,Brouwer, A., Sauer, P.J.J., 1994. Effects of dioxins and polychlo-rinated biphenyls on thyroid hormone status of pregnant womenand their infants. Pediatr. Res. 36, 468.
Koopman-Esseboom, C., Weisglas-Kuperus, N., de Riddert, M.A.J.,van der Paauw, C.G., Tuinstra, L.G.M.T., Sauer, P.J.J., 1996.Effects of polychlorinated biphenyl/dioxin exposure and feedingtype on infants’ mental and psychomotor development. Pediatrics97, 700.
Korach, K.S., Sarver, P., Chae, K., McLachlan, J.A., McKinney,J.D., 1988. Estrogen receptor-binding activity of polychlorinatedhydroxybiphenyls: conformationally restricted structural probes.Mol. Pharmacol. 33, 120.
Lans, M.C., Klasson-Wehler, E., Willemsen, M., Meussen, E., Safe,S., Brouwer, A., 1993. Structure-dependent, competitive interac-tion of hydroxy-polychlorobiphenyls, -dibenzo-p-dioxins and -dibenzofurans with human transthyretin. Chem. Biol. Interact. 88,7.
Larsen, P.R., Davies, T.F., Hay, I.D., 1998. The thyroid gland. In:Wilson, J.D., Foster, D.W., Kronenberg, H.M., Larsen, P.R.(Eds.), Williams Textbook of Endocrinology, Ninth ed. Saunders,Philadelphia, p. 389.
Lilienthal, H., Winneke, G., 1991. Sensitive periods for behavioraltoxicity of polychlorinated biphenyls: determination by cross-fos-tering in rats. Fundam. Appl.Toxicol. 17, 368.
Lilienthal, H., Weinand-Harer, A., Winterhoff, H., Winneke, G.,1997. Effects of maternal exposure to 3,3%,4,4%-tetrachlorobiphenylor propylthiouracil in rats trained to discriminate apomorphinefrom saline. Toxicol. Appl. Pharmacol. 146, 162.
Liu, J., Liu, Y., Barter, R.A., Klaassen, C.D., 1995. Alteration ofthyroid homeostasis by UDP-glucuronosyltransferase inducers inrats: a dose-response study. J. Pharmacol. Exp. Ther. 273, 977.
McKinley, M.K., Kedderis, L.B., Birnbaum, L.S., 1993. The effect ofpretreatment on the biliary excretion of 2,3,7,8-tetra-chlorodibenzo-p-dioxin, 2,3,7,8-tetrachlorodibenzofuran and3,3%,4,4%-tetrachlorobiphenyl in rats. Fundam. Appl. Toxicol. 21,425.
Mileson, B.E., Lewis, M.H., Mailman, R.B., 1991. Dopamine recep-tor ‘supersensitivity’ occuring without receptor up-regulation.Brain Res. 561, 1.
Morse, D.C., Groen, D., Veerman, M., van Amerongen, C.J., Koeter,H.B.W.M., Smits van Prooije, A.E., Visser, T.J., Koeman, J.H.,Brouwer, A., 1993. Interference of polychlorinated biphenyls inhepatic and brain thyroid hormone metabolism in fetal andneonatal rats. Toxicol. Appl. Pharmacol. 122, 27.
Morse, D.C, Klasson Wehler, E., van de Pas, M., de Bie, A.Th.H.J.,van Bladeren, P.J., Brouwer, A., 1995. Metabolism and biochem-ical effects of 3,3%,4,4%-tetrachlorobiphenyl in pregnant and fetalrats. Chem. Biol. Interact. 95, 41.
Morse, D.C., Seegal, R.F., Borsch, K.O., Brouwer, A., 1996b. Long-term alterations in regional brain serotonin metabolism followingmaternal polychlorinated biphenyl exposure in the rat. Neurotox-icology 17, 631.
Morse, D.C., Klasson Wehler, E., Wesseling, W., Koeman, J.H.,Brouwer, A., 1996a. Alterations in rat brain thyroid hormonestatus following pre- and postnatal exposure to polychlorinatedbiphenyls (Aroclor 1254). Toxicol. Appl. Pharmacol. 136, 269.
Ness, D.K., Schantz, S.L., Moshtaghian, J., Hansen, L.G., 1993.Effects of perinatal exposure to specific PCB congeners on thyroidhormone concentrations and thyroid histology in the rat. Toxicol.Lett. 68, 311.
Palha, J.A., Fernandes, R., de Escobar, G.M., Episkopou, V., Got-tesman, M., Saraiva, M.J., 2000. Transthyretin regulates thyroidhormone levels in the choroid plexus, but not in the brainparenchyma: study in a transthyretin-null mouse model. En-docrinology 141, 3267.
Patandin, S., Lanting, C.I., Mulder, P.G.H., Boersma, E.R., Sauer,P.J.J., Weisglas-Kuperus, N., 1999. Effects of environmental ex-posure to polychlorinated biphenyls and dioxins on cognitiveabilities in Dutch children at 42 months of age. J. Pediatr.
Porterfield, S.P., Hendrich, C.E., 1993. The role of thyroid hormonesin prenatal and neonatal neurological development-current per-spectives. Endocr. Rev. 14, 94.
Porterfield, S.P., 1994. Vulnerability of the developing brain to thy-roid abnormalities: environmental insults to the thyroid system.Environ. Health Perspect. 102 (Suppl. 2), 125.
Revet, J.F., 1999. Long-term neuropsychological sequelae of early-treated congenital hypothyroidism: effects in adolescence. ActaPaediatr. Suppl. 432 88.
Rogan, W.J., Gladen, B.C., McKinney, J.D., Carreras, N., Hardy, P.,Thullen, J., Tinglestad, J., Tully, M., 1986. Neonatal effects oftransplacental exposure to PCBs and DDE. J. Pediatr. 109, 335.
Saghir, S.A., Koritz, G.D., Hansen, L.G., 1999. Short-term distribu-tion, metabolism, and excretion of 2,2’,5-tri-, 2,2’,4,4’-tetra-, and3,3’,4,4’-tetrachlorobiphenyl in prepubertal rats. Arch. Environ.Contam. Toxicol. 36, 213.
Savard, P., Merand, Y., Di Paolo, T., Dupont, A., 1984. Effect ofneonatal hypothyroidism on the serotonin system of the rat brain.Brain Res. 292, 99.
Schantz, S., Moshtaghian, J., Ness, D.K., 1995. Spatial learningdeficits in adult rats exposed to ortho-substituted PCB congenersduring gestation and lactation. Fundam. Appl. Toxicol. 26, 117.
Schuur, A.G., Boekhorst, F.M., Brouwer, A., Visser, T.J., 1997.Extrathyroidal effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin onthyroid hormone turnover in male Sprague–Dawley rats. En-docrinology 138, 3727.
Seegal, R.F., Bush, B., Shain, W., 1990. Lightly chlorinated ortho-substituted PCB congeners decrease dopamine in nonhuman pri-mate brain and in tissue culture. Toxicol. Appl. Pharmacol. 106,136.
Seegal, R.F., 1996. Epidemiological and laboratory evidence of PCB-induced neurotoxicity. Crit. Rev. Toxicol. 26, 709.
Seegal, R.F., Brosch, K.O., Okoniewski, R.J., 1997. Effects of inutero and lactational exposure of the laboratory rat to 2,4,2’,4’-and 3,4,3’,4’-tetrachlorobiphenyl on dopamine function. Toxicol.Appl. Pharmacol. 146, 95.

A. Roth-Harer et al. / En6ironmental Toxicology and Pharmacology 9 (2001) 103–115 115
Seo, B.-W., Li, M.-H., Hansen, L.G., Moore, R.W., Peterson, R.E.,Schantz, S.L., 1995. Effects of gestational and lactational exposureto coplanar polychlorinated biphenyl (PCB) congeners or 2,3,7,8-te-trachlorodibenzo-p-dioxin (TCDD) on thyroid hormone concen-trations in weanling rats. Toxicol. Lett. 78, 253.
Shain, W., Bush, B., Seegal, R., 1991. Neurotoxicity of polychlorinatedbiphenyls: structure–activity relationship of individual congeners.Toxicol. Appl. Pharmacol. 111, 33.
Tilson, H.A., Harry, G.J., 1994. Developmental neurotoxicology ofpolychlorinated biphenyls and related compounds. In: Isaacson,R.L., Jensen, K.F. (Eds.), The Vulnerable Brain and EnvironmentalRisks, Toxins in Air and Water, vol. 3. Plenum Press, New York.
Wadenberg, M.-L., 1996. Serotonergic mechanisms in the neuro-leptic-induced catalepsy in the rat. Neurosci. Biobehav. Rev. 20,325.
Weinand-Harer, A., Lilienthal, H., Bucholski, K.-A., Winneke, G.,1997. Behavioral effects of maternal exposure to an ortho-chlori-nated or a coplanar PCB congener in rats. Environ. Toxicol.Pharmacol. 3, 97.
Winneke, G., Bucholski, A., Heinzow, B., Kramer, U., Schmidt, E.,Walkowiak, J., Wiener, J.-A., Steingruber, H.-J., 1998. Develop-mental neurotoxicity of polychlorinated biphenyls (PCBs): cogni-tive and psychomotor functions in seven months old children.Toxicol. Lett. 102-103, 423.
.