neuronal network coherent with hand kinematics during fast repetitive hand movements
TRANSCRIPT
This article appeared in a journal published by Elsevier. The attached
copy is furnished to the author for internal non-commercial research
and education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling or
licensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of the
article (e.g. in Word or Tex form) to their personal website or
institutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies are
encouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Neuronal network coherent with hand kinematics during fast repetitivehand movements
Mathieu Bourguignon a,b,!, Veikko Jousmäki a,b, Marc Op de Beeck a, Patrick Van Bogaert a,Serge Goldman a, Xavier De Tiège a
a Laboratoire de Cartographie Fonctionnelle du Cerveau, ULB-Hôpital Erasme, 808 route de Lennik, 1070 Brussels, Belgiumb Brain Research Unit, Low Temperature Laboratory, Aalto University School of Science, Puumiehenkuja 2B, Otaniemi, Espoo, Finland
a b s t r a c ta r t i c l e i n f o
Article history:Received 18 April 2011Revised 9 August 2011Accepted 12 September 2011Available online 22 September 2011
Keywords:AccelerometerHand movementsKinematicsMagnetoencephalographyMotor cortex
We quanti!ed the coupling between magnetoencephalographic (MEG) cortical signals and the kinematics offast repetitive voluntary hand movements monitored by a 3-axis accelerometer. Ten healthy right-handedadults performed self-paced "exion–extension movements of right-hand !ngers at ~3 Hz with either touch-ing the thumb during "exions (TOUCH) or not (noTOUCH). At the sensor level, we found in all subjects andconditions signi!cant coherence at the movement frequency (F0) and its !rst harmonic (F1). Coherencevalues were signi!cantly higher in TOUCH compared to noTOUCH. At the group level, dynamic imaging of co-herent sources localized the main source of coherent activity at the left primary motor (M1) hand area, ex-cept at F0 TOUCH were the main source was localized at the left primary sensory (S1) hand area. Othercoherent brain areas were also identi!ed at right S1-M1 cortices (F0), left dorsolateral prefrontal cortex(F1), left posterior parietal cortex (F0 TOUCH and F1 noTOUCH) and left medial S1-M1 areas (TOUCH).This study highlights the prominent role of rhythmic neuronal activity phase-locked to movements for theencoding and the integration of key sensori-motor features of limb kinematics. This study also suggeststhat somatosensory afferences play a key role to sustain a high synchronization level between the neuronalactivity in coherent brain areas and hand acceleration. Some coherent brain regions differed between F0 andF1 in both conditions, suggesting that distinct cortical areas are involved in different features of handkinematics.
© 2011 Elsevier Inc. All rights reserved.
Introduction
Most coordinated voluntary movements require highly synchro-nized and synergistic neuronal control of antagonistic musclesgroups. Unraveling how these movements are generated at the neu-ronal level represents a major issue in neuroscience and a prerequi-site to better understand movement disorders pathophysiology orto improve brain computer interface (BCI) approaches.
Previous human and non-human primate studies have shown thatmany movement's kinematics parameters correlate with the neuro-nal activity in the primary motor (M1) cortex (Ebner et al., 2009). In-deed, single neuron studies performed in non-human primates haveshown that neurons in M1 cortex encode various movement parame-ters such as direction, position, rotation, and movement velocity(Ashe and Georgopoulos, 1994; Caminiti et al., 1990; Carmena et al.,2003; Georgopoulos et al., 1988; Mehring et al., 2003; Moran andSchwartz, 1999; Reina et al., 2001). In addition, other studies haveshown that hand movement target and velocity can be successfully
inferred from M1 local !eld potentials (Mehring et al., 2003). At thecoarsest recording scale, human electric potentials on the scalp andmagnetic !elds outside the head demonstrated a robust relationshipbetween time-varying M1 neuronal activity and movement velocity(Kelso et al., 1998; O'Suilleabhain et al., 1999).
In non-human primates, limb kinematics correlates with neuronalactivity recorded at the single-cell or multiunit levels in several non-primary brain areas such as premotor cortex, posterior parietal cortex(PPC), and cerebellum (Averbeck et al., 2005; Carmena et al., 2003;Coltz et al., 1999; Moran and Schwartz, 1999; Stark et al., 2007).This !nding has been extended in humans by positron emission to-mography (PET) or functional magnetic resonance imaging (fMRI)studies, which have shown that the activity in multiple primary andnon-primary sensori-motor cortical areas correlates with movementvelocity (Lewis et al., 2003; Turner et al., 1998).
High-temporal resolution magnetoencephalographic (MEG) re-cordings have been used in conjunction with tomographic source re-construction methods to characterize non-invasively the relationshipbetween neuronal activity throughout the human brain and move-ment kinematics (Bradberry et al., 2009; Jerbi et al., 2007). Thesestudies used complex visuomotor tasks and identi!ed a large neuro-nal network involved in the visuomotor control of hand speed. In
NeuroImage 59 (2012) 1684–1691
! Corresponding author at: 808 route de Lennik, 1070 Bruxelles, Belgium. Fax: +3225556631.
E-mail address: [email protected] (M. Bourguignon).
1053-8119/$ – see front matter © 2011 Elsevier Inc. All rights reserved.doi:10.1016/j.neuroimage.2011.09.022
Contents lists available at SciVerse ScienceDirect
NeuroImage
j ourna l homepage: www.e lsev ie r .com/ locate /yn img

Author's personal copy
particular, Jerbi et al. (2007) found signi!cant phase-locking betweenslow (2- to 5-Hz) neuronal activity at handM1 cortex, contralateral tohand movement, and time-varying hand movement velocity. HandM1 cortical activity also showed task-related coupling in the samefrequency band with the neuronal activity in a large-scale visuomotornetwork including frontoparietal circuits and cerebello-thalamo-cortical pathways (Jerbi et al., 2007).
In addition, Pollok et al. (2004a, 2005) investigated the neuronalnetwork involved in externally triggered hand movements usingMEG and dynamic imaging of coherent sources (DICS; Gross et al.,2001), a spatial !lter used for source reconstruction. By mappingthe synchronization between MEG oscillations and movement fre-quency as recorded by electromyography (EMG), they showed thatthe source with the strongest coherence to EMG was localized inthe primary sensori-motor (S1-M1) hand area contralateral to themoving hand. In addition, hand S1-M1 cortical activity also showedtask-related coupling with premotor areas, the PPC, the thalamusand the cerebellum.
Previous functional cerebral imaging studies have suggested thatthe contribution of several brain areas to movements differs dependingon the internal versus external control of movements (Debaere et al.,2003; Jueptner et al., 1996; Ogawa et al., 2006; Taniwaki et al., 2006).Therefore, to investigate in humans the relationship between handmovement kinematics and brain activity in the context of natural self-initiated voluntary movements, we developed a novel method, cortico-kinematic coherence (CKC), that maps synchronization between MEGoscillations and the kinematics of self-paced voluntary repetitive handmovements monitored by a 3-axis accelerometer (Acc) (Bourguignonet al., 2011). Using this method, we were able to show that cortical ac-tivity is coherent with hand acceleration during natural repetitive handmovements at the hand movement frequency (F0, ~3 Hz) and its !rstharmonic (F1, ~6 Hz). Equivalent current dipole (ECD) modeling inthe time domain using the cross-correlogram pinpointed the hand M1cortex contralateral to hand movement as the main source of thecoherent activity. Nevertheless, this ECD modeling approach did notallow to characterize the neural network involved in the differentcoherent frequencies (F0 and F1). In addition, the existence of a moredistributed coherent neuronal network, as suggested by weaker ipsilat-eral coherent signals in some subjects' cross-correlogram, could not beidenti!ed with the source reconstruction method used. In this paper,we used the DICS approach in combination with a group-level coher-ence analysis to better describe the CKC phenomenon in two types ofhandmovements differing in the amount of tactile synchronous stimu-lations. This study was speci!cally designed 1) to characterize theneuronal network involved in self-initiated fast repetitive hand move-ments, 2) to search for differences in the coherent neuronal networkdepending on the amount of somatosensory inputs, and 3) to assessdifferences in the brain regions involved at coherent frequencies.
Materials and methods
Fig. 1 summarizes the corticokinematic coherence method used inthis study.
Parts of the data used in this study have been analyzed in a previ-ous study using equivalent current dipole modeling in the time do-main (Bourguignon et al., 2011).
Subjects
Ten healthy subjects (range 24–40 yrs; mean age 30.5 yrs; 5males, 5 females) without any history of neuropsychiatric disease ormovement disorders were studied. All subjects were right-handedaccording to Edinburgh handedness inventory (Old!eld, 1971).
The study was approved by the ULB-Hôpital Erasme Ethics Com-mittee. Subjects participated after informed consent. The measure-ments were carried out at the ULB-Hôpital Erasme, Brussels, Belgium.
Experimental paradigm
Subjects were asked to make self-paced "exions–extensions of allright-hand !ngers at about 3 Hz for 3 min in two conditions differingin the amount of tactile somatosensory inputs. In the touch condition(TOUCH), subjects were asked to touch the thumb with the other !n-gers during the "exions, leading to a high degree of tactile input from!ngers. In the non-touch condition (noTOUCH), they were asked toavoid touching the thumb with the other !ngers during the "exionsin order to limit as much as possible the level of tactile information.The order of condition was balanced across subjects. In order to min-imize effects of visual inputs and eye-movements artifacts, subjectswere asked to !xate the gaze at a point in the magnetically shieldedroom to avoid any gaze contact with the moving hand.
Data acquisition
Neuromagnetic signals were recorded with a whole-scalp-covering neuromagnetometer in a magnetically shielded room
Euclidian norm
0 2Time (s)
0 2Time (s)
Acc signals
MEG recording
Signal spaceseparation
Acc recording
Coherence analysis
Acc signal = x MEG signals = y
Pxy( f ) = 1K
Xk ( f )Yk* ( f )
k
!
Cohxy( f ) =Pxy( f )
2
Pxx( f )Pyy( f )
Fig. 1. Summary of the processing steps described in the coherence method. Upper Left.The three measured orthogonal Acc signals are band-pass !ltered and then combinedinto a single Acc signal using the Euclidian norm. Upper Right. Magnetoencephalo-graphic (MEG) signals are !ltered using signal space separation (SSS) method to cor-rect for head movements and subtract external interferences. Bottom. Both signalsare epoched to compute the individual coherence maps, which are thereafter normal-ized to standard MNI space and used to compute a coherence map at the group level.
1685M. Bourguignon et al. / NeuroImage 59 (2012) 1684–1691

Author's personal copy
(Vectorview & MaxshieldTM; Elekta Oy, Helsinki, Finland). Head po-sition inside the MEG helmet was continuously monitored usingfour head tracking coils. The locations of the coils with respect to an-atomical !ducials were determined with an electromagnetic tracker(Fastrak, Polhemus, Colchester, VT, USA). The movements of theright index !nger were monitored time-locked to MEG signals witha 3-axis Acc (ADXL330 iMEMS Accelerometer, Analog Devices, Inc.,Norwood, MA, USA). Time-locked MEG and Acc signals wererecorded using a bandpass of 0.1–330 Hz and a sampling rate of1 kHz. High-resolution 3D-T1 cerebral magnetic resonance images(MRIs) were acquired on a 1.5 Tesla MRI scan (Intera, Philips, TheNetherlands).
Data preprocessing
Continuous MEG data were !rst preprocessed off-line using thesignal space separation method to suppress external interferencesand to correct for head movements (Taulu et al., 2005). To performfrequency and coherence analyses, continuous data were split into2048 ms epochs with 1638-ms epoch overlap, leading to a frequencyresolution of ~0.5 Hz (Bortel and Sovka, 2007). Such frequency reso-lution is typically used in coherence analyses (Baker and Baker,2003; Semmler and Nordstrom, 1998). MEG epochs exceeding 3 pT(magnetometers) or 0.7 pT/cm (gradiometers) were excluded fromfurther analysis to avoid contamination of our data by eye move-ments, muscle activity, and artifacts in the MEG sensors. These stepsled to more than 400 artifact free MEG and Acc epochs for each sub-ject and condition. The acceleration corresponding to each epochwas computed at every sample as the Euclidian norm of the threeband-passed (1–200 Hz) Acc channels.
Coherence analyses
The coherence is an extension of Pearson correlation coef!cient tothe frequency domain, which determines the degree of coupling be-tween two signals x(t) and y(t), providing a number between 0 (nolinear dependency) and 1 (perfect linear dependency) for each fre-quency (Halliday et al., 1995). Let Xk(f) and Yk(f) be the Fourier trans-form of the kth segment of x(t) and y(t), by de!ning
Pxy f! " # 1K!kXk f! "Y$
k f! "; !1"
K being the number of averaged epochs, the coherence can bewritten as
Cohxy f! " #Pxy f! "!! !!2
Pxx f! "Pyy f! " : !2"
Considering the linear relationship between acceleration and ve-locity in the frequency domain, coherence analyses were performedusing the acceleration directly available from Acc signals rather thanwith velocity signals derived from the acceleration by time integra-tion (Bourguignon et al., 2011).
Frequencies of interest
To determine the frequencies used for coherent source analyses,namely the frequencies of interest, coherence spectra were computedusing (2) between Acc and MEG signals (i.e. sensor space). Frequen-cies that showed in all subjects signi!cant coupling between Accand MEG signals in sensor space were de!ned as the frequencies ofinterest. To perform coherent source analyses, the (Nb+1!Nb+1)cross-spectral density matrix was computed between all possiblecombinations of MEG and acceleration signals at the frequencies ofinterest.
Coherent source analyses
Individual MRIs were !rst segmented using Freesurfer software(Martinos Center for Biomedical Imaging, Massachusetts, USA).Then, the MEG forward model for two orthogonal tangential cur-rent dipoles placed on a homogeneous 7 mm grid source spacecovering the whole brain was computed using MNE suite (Marti-nos Center for Biomedical Imaging, Massachusetts, USA). Coherencemaps at the frequencies of interest were !nally produced withinthe computed source space using the Dynamic Imaging of CoherentSources (DICS) approach with minimum variance beamformer(Gross et al., 2001). Both planar gradiometers and magnetometerswere used for inverse modeling, by using a constant non-adaptive weighting. Magnetometer noise was !xed at 300 fT andgradiometer noise at 50 fT/cm. On average, this weighting gave aslightly higher weight to gradiometer signals as compared to mag-netometer signals.
Group level analyses
A 12–parameters af!ne transformation from individual MRIs tothe standard Montreal Neurological Institute (MNI) brain was !rstcomputed using the spatial normalization algorithm implementedin Statistical Parametric Mapping (SPM8, Wellcome Department ofCognitive Neurology, London, UK) and then applied to individualMRIs and coherence maps. This procedure generated a normalizedcoherence map in the MNI space for each subject, condition, and fre-quency of interest. To produce coherence maps at the group level foreach condition and frequency of interest, we computed the general-ized f-mean of individual normalized maps, according tof "! " # arctanh
"""""""! "
p# $, namely the Fisher z-transform of the square
root. This procedure transforms the noise on the coherence into anapproximately normally distributed noise (Rosenberg et al., 1989).Therefore, the computed coherence is an unbiased estimation of themean coherence at the group level. Moreover, this averaging proce-dure lessens the relative contribution of subjects characterized byhigh coherence values to the group analysis. Finally, difference of f-transformed TOUCH–noTOUCH group-level maps (difference maps)was computed.
Statistical analyses
Simulated data were used to assess the threshold for statisticalsigni!cance of coherence values in single subject's sensor space andgroup-level coherence maps. This approach overcomes the multiplecomparison issue, which has no straightforward analytical solutionwhen dealing with highly dependent time series.
To assess the statistical signi!cance of our results, for both condi-tions and frequencies of interest separately, we repeated 10,000 timesthe procedure of computing subject-level coherence values in thesensor space, group-level coherence maps, and difference mapsusing real MEG signals and Fourier transform surrogate accelerationsignals (Faes et al., 2004). The Fourier transform surrogate imposespower spectrum to remain the same as in the original signal but re-places the phase of Fourier coef!cients by random numbers in therange [#!,!] in the surrogate signals (Faes et al., 2004). Maximal co-herence and difference values were then extracted for each simula-tion to compute the cumulative density function of the maximalcoherence and difference values occurring due to stochastic matchingbetween acceleration and MEG sensor or source signals. The coher-ence or difference thresholds at pb0.05 corrected for multiple com-parisons were then evaluated as the 0.95 percentile of thecorresponding cumulative density function.
Statistical differences between movement frequencies and maxi-mal coherence levels between TOUCH and noTOUCH conditionswere assessed using Wilcoxon signed rank test at the sensor level.
1686 M. Bourguignon et al. / NeuroImage 59 (2012) 1684–1691

Author's personal copy
Assessment of coherence maps
Local coherence maxima were identi!ed in group-level coherencemaps for each condition and frequencies of interest. Only local coher-ence maxima with values above the statistical threshold were consid-ered. The number of subjects for which normalized coherence mapsdisplayed a local coherence maximum less than 10 mm away fromthe corresponding group-level local coherence maximum was thendetermined.
Results
Acceleration and coherence spectra
All subjects performed the tasks with a movement frequency ofaround 3 Hz. The movement frequency was signi!cantly higher(p=0.0039) in TOUCH (3.61±0.52 Hz; mean±sd) than in noTOUCH(2.73±0.66 Hz). In both, TOUCH and noTOUCH, acceleration powerspectra and coherence spectra showed peaks at F0 and F1 in all sub-jects (see Fig. 2). Coherence values at these frequencies were statisti-cally signi!cant for all subjects and conditions (pb0.05, Table 1). At
F0 (varying with subject and condition), maximal coherence valuesover all sensors were signi!cantly higher in TOUCH than in noTOUCH(pb0.05). The same tendency was observed at F1 but did not reachsigni!cance (p=0.13). In addition, signi!cant coherence peaks wereobserved at higher harmonics in some subjects (see Fig. 2). Only F0and F1 were considered as frequencies of interest for coherent sourceanalyses.
Coherent source analyses
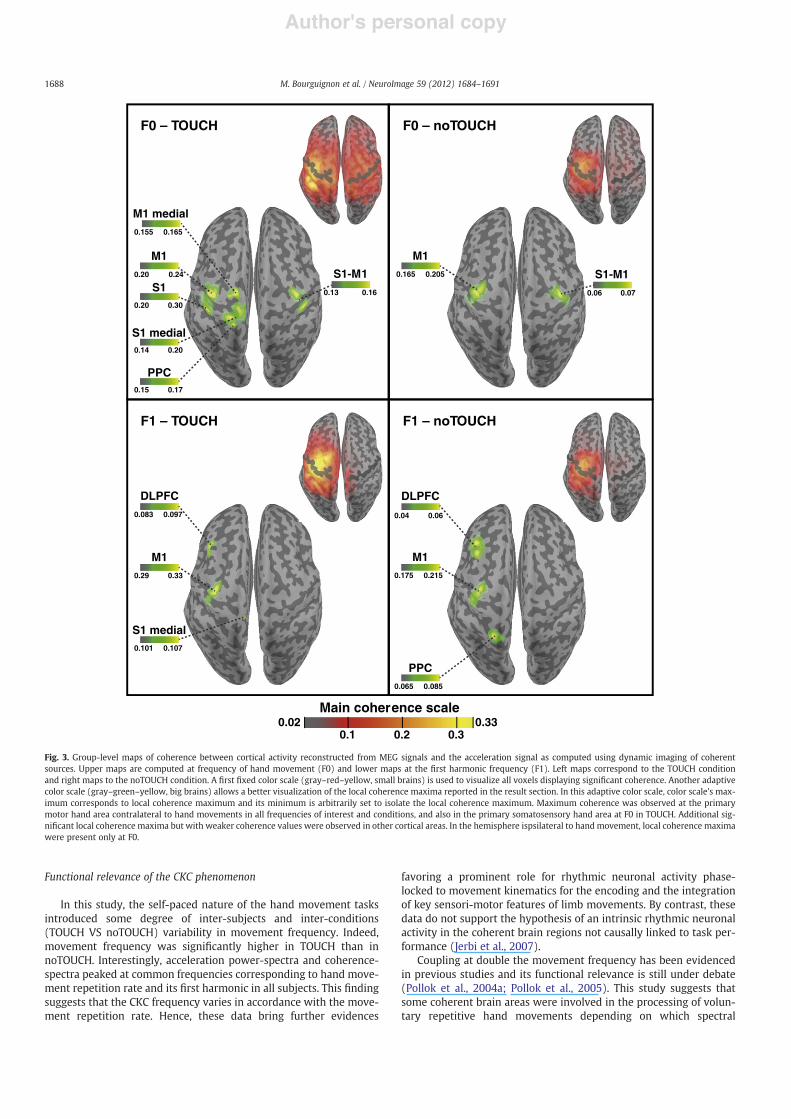
Fig. 3 illustrates the results of coherent source analyses at thegroup level.
The coherence values for the signi!cant coherent cortical areas atthe group level as well as the number of individual subjects in whomthose coherent cortical areas were observed are reported in Table 2.
At the group level, the maximum coherence was observed at M1hand area contralateral to hand movements in all frequencies of in-terest and conditions, except at F0 in TOUCH where S1 hand areawas the most coherent brain area. MNI coordinates of the local coher-ence maximum observed at the left M1 hand area were very closeto each other in all conditions and frequencies of interest, with max-imum distance of 3.6 mm from the mean coordinate [# 40.75# 17.559.5].
Signi!cant local coherence maxima with weaker coherence valueswere observed across conditions or coherent frequencies in othersegregated cortical areas. In the hemisphere contralateral to handmovement, these maxima were located in the dorsolateral prefrontalcortex (DLPFC) at F1 in both conditions, in the medial S1-M1 area atF0 and F1 in TOUCH, and in the PPC at F0 in TOUCH and at F1 innoTOUCH. In the hemisphere ipsilateral to hand movement, local co-herence maxima were present only at F0, at S1-M1 area in bothconditions.
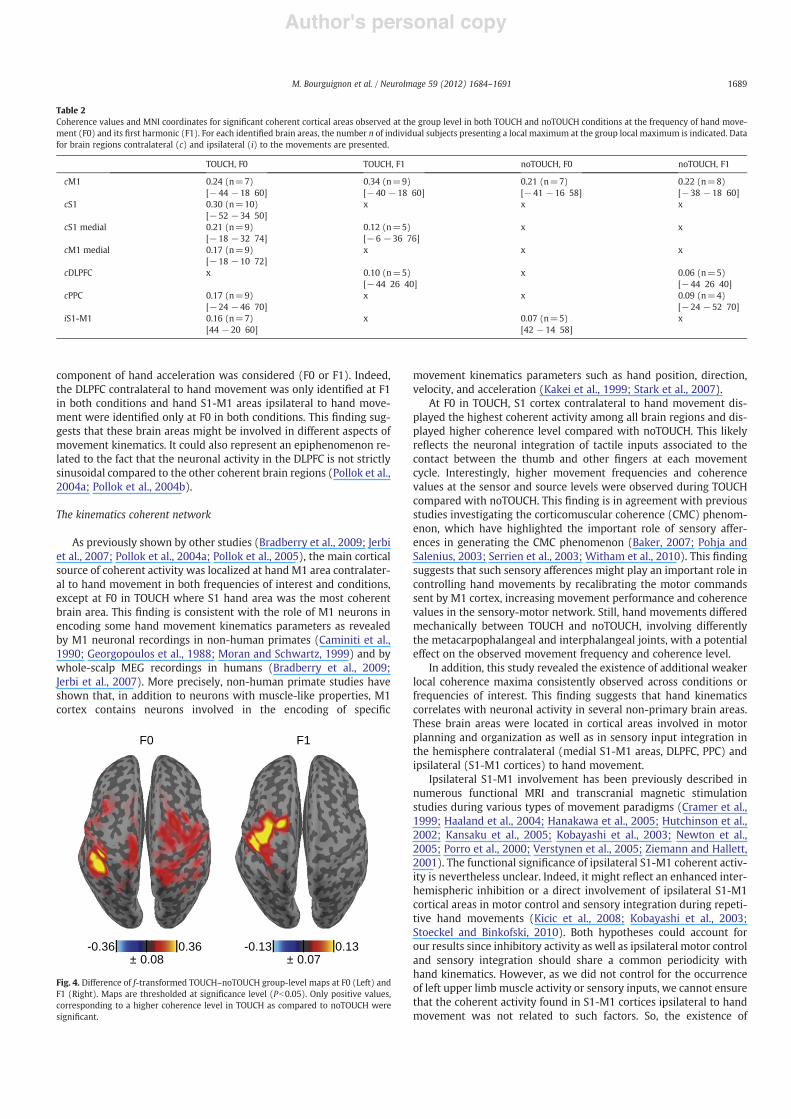
At F0, difference maps showed that coherence values were signif-icantly higher at all local coherence maxima observed in TOUCHcompared with noTOUCH, except at M1 cortex contralateral tohand movement (Fig. 4). At F1, coherence values were signi!cantlyhigher at M1 cortex contralateral to hand movement in TOUCH com-pared with noTOUCH. Difference maps also revealed that coherencevalues were signi!cantly higher in some parieto-frontal regions inTOUCH compared with noTOUCH but no manifest local coherencemaximum was identi!ed in these regions in group level coherencemaps.
Discussion
Using natural hand movement paradigms and non-invasive MEGrecordings, this study shows that rhythmic neuronal activity withinan extensive cortical sensori-motor network is coupled with hand ac-celeration during repetitive self-initiated voluntary hand movements.This study highlights that the main source of the coherent activity atthe movement frequency is located in M1 or S1 hand cortical areacontralateral to hand movement depending on the amount of tactilesomatosensory inputs (TOUCH or noTOUCH). It also demonstratesthat coherence values are signi!cantly higher when the movementtask is associated with a high level of somatosensory inputs.
Subject 1
0
1
0 10 20 30
Acc
Coh
Subject 5
0
1
0 10 20 30
Acc
Coh
Subject 2
0
1
0 10 20 30
Acc
Coh
Subject 6
0
1
0 10 20 30
Acc
Coh
Subject 9
0
1
0 10 20 30
Acc
Coh
Subject 3
0
1
0 10 20 30
Acc
Coh
Subject 7
0
1
0 10 20 30
Acc
Coh
Subject 10
0
1
0 10 20 30
Acc
Coh
Subject 4
0
1
0 10 20 30
Acc
Coh
Subject 8
0
1
0 10 20 30
Acc
Coh
TOUCH noTOUCH
Frequency (Hz)Frequency (Hz)
Fig. 2. Acceleration power spectra (upper graph) and MEG–acceleration coherencespectra (lower graph) for all subjects. The coherence was computed between a singleMEG channel located above the left rolandic hand region and the hand acceleration sig-nal both when the !ngers were touching each other (TOUCH, solid lines) or not(noTOUCH, dashed lines). The correspondence between frequencies maximizing bothcoherence and acceleration power spectra can be noted. Frequencies maximizing thecoherence are higher during TOUCH than noTOUCH.
Table 1Mean and range of maximum coherence values at the sensor level in TOUCH andnoTOUCH conditions for the fundamental frequency (F0) and the !rst harmonic fre-quency (F1).
F0 F1
TOUCH noTOUCH TOUCH noTOUCH
Mean 0.55 0.31 0.48 0.29Min 0.23 0.12 0.19 0.07Max 0.78 0.83 0.75 0.80
1687M. Bourguignon et al. / NeuroImage 59 (2012) 1684–1691
Author's personal copy
Functional relevance of the CKC phenomenon
In this study, the self-paced nature of the hand movement tasksintroduced some degree of inter-subjects and inter-conditions(TOUCH VS noTOUCH) variability in movement frequency. Indeed,movement frequency was signi!cantly higher in TOUCH than innoTOUCH. Interestingly, acceleration power-spectra and coherence-spectra peaked at common frequencies corresponding to hand move-ment repetition rate and its !rst harmonic in all subjects. This !ndingsuggests that the CKC frequency varies in accordance with the move-ment repetition rate. Hence, these data bring further evidences
favoring a prominent role for rhythmic neuronal activity phase-locked to movement kinematics for the encoding and the integrationof key sensori-motor features of limb movements. By contrast, thesedata do not support the hypothesis of an intrinsic rhythmic neuronalactivity in the coherent brain regions not causally linked to task per-formance (Jerbi et al., 2007).
Coupling at double the movement frequency has been evidencedin previous studies and its functional relevance is still under debate(Pollok et al., 2004a; Pollok et al., 2005). This study suggests thatsome coherent brain areas were involved in the processing of volun-tary repetitive hand movements depending on which spectral
S1-M1
0.02 0.33
0.13 0.16
M1
DLPFC
0.065 0.085
0.175 0.215
0.04 0.06
PPC
M10.165 0.205 S1-M1
0.06 0.07
F0 – TOUCH F0 – noTOUCH
F1 – TOUCH F1 – noTOUCH
M1
S1
M1 medial
S1 medial
PPC
M1
S1 medial
DLPFC
0.15 0.17
0.14 0.20
0.20 0.30
0.20 0.24
0.155 0.165
0.101 0.107
0.29 0.33
0.083 0.097
Main coherence scale
0.1 0.2 0.3
Fig. 3. Group-level maps of coherence between cortical activity reconstructed from MEG signals and the acceleration signal as computed using dynamic imaging of coherentsources. Upper maps are computed at frequency of hand movement (F0) and lower maps at the !rst harmonic frequency (F1). Left maps correspond to the TOUCH conditionand right maps to the noTOUCH condition. A !rst !xed color scale (gray–red–yellow, small brains) is used to visualize all voxels displaying signi!cant coherence. Another adaptivecolor scale (gray–green–yellow, big brains) allows a better visualization of the local coherence maxima reported in the result section. In this adaptive color scale, color scale's max-imum corresponds to local coherence maximum and its minimum is arbitrarily set to isolate the local coherence maximum. Maximum coherence was observed at the primarymotor hand area contralateral to hand movements in all frequencies of interest and conditions, and also in the primary somatosensory hand area at F0 in TOUCH. Additional sig-ni!cant local coherence maxima but with weaker coherence values were observed in other cortical areas. In the hemisphere ispsilateral to handmovement, local coherence maximawere present only at F0.
1688 M. Bourguignon et al. / NeuroImage 59 (2012) 1684–1691
Author's personal copy
component of hand acceleration was considered (F0 or F1). Indeed,the DLPFC contralateral to hand movement was only identi!ed at F1in both conditions and hand S1-M1 areas ipsilateral to hand move-ment were identi!ed only at F0 in both conditions. This !nding sug-gests that these brain areas might be involved in different aspects ofmovement kinematics. It could also represent an epiphenomenon re-lated to the fact that the neuronal activity in the DLPFC is not strictlysinusoidal compared to the other coherent brain regions (Pollok et al.,2004a; Pollok et al., 2004b).
The kinematics coherent network
As previously shown by other studies (Bradberry et al., 2009; Jerbiet al., 2007; Pollok et al., 2004a; Pollok et al., 2005), the main corticalsource of coherent activity was localized at hand M1 area contralater-al to hand movement in both frequencies of interest and conditions,except at F0 in TOUCH where S1 hand area was the most coherentbrain area. This !nding is consistent with the role of M1 neurons inencoding some hand movement kinematics parameters as revealedby M1 neuronal recordings in non-human primates (Caminiti et al.,1990; Georgopoulos et al., 1988; Moran and Schwartz, 1999) and bywhole-scalp MEG recordings in humans (Bradberry et al., 2009;Jerbi et al., 2007). More precisely, non-human primate studies haveshown that, in addition to neurons with muscle-like properties, M1cortex contains neurons involved in the encoding of speci!c
movement kinematics parameters such as hand position, direction,velocity, and acceleration (Kakei et al., 1999; Stark et al., 2007).
At F0 in TOUCH, S1 cortex contralateral to hand movement dis-played the highest coherent activity among all brain regions and dis-played higher coherence level compared with noTOUCH. This likelyre"ects the neuronal integration of tactile inputs associated to thecontact between the thumb and other !ngers at each movementcycle. Interestingly, higher movement frequencies and coherencevalues at the sensor and source levels were observed during TOUCHcompared with noTOUCH. This !nding is in agreement with previousstudies investigating the corticomuscular coherence (CMC) phenom-enon, which have highlighted the important role of sensory affer-ences in generating the CMC phenomenon (Baker, 2007; Pohja andSalenius, 2003; Serrien et al., 2003; Witham et al., 2010). This !ndingsuggests that such sensory afferences might play an important role incontrolling hand movements by recalibrating the motor commandssent by M1 cortex, increasing movement performance and coherencevalues in the sensory-motor network. Still, hand movements differedmechanically between TOUCH and noTOUCH, involving differentlythe metacarpophalangeal and interphalangeal joints, with a potentialeffect on the observed movement frequency and coherence level.
In addition, this study revealed the existence of additional weakerlocal coherence maxima consistently observed across conditions orfrequencies of interest. This !nding suggests that hand kinematicscorrelates with neuronal activity in several non-primary brain areas.These brain areas were located in cortical areas involved in motorplanning and organization as well as in sensory input integration inthe hemisphere contralateral (medial S1-M1 areas, DLPFC, PPC) andipsilateral (S1-M1 cortices) to hand movement.
Ipsilateral S1-M1 involvement has been previously described innumerous functional MRI and transcranial magnetic stimulationstudies during various types of movement paradigms (Cramer et al.,1999; Haaland et al., 2004; Hanakawa et al., 2005; Hutchinson et al.,2002; Kansaku et al., 2005; Kobayashi et al., 2003; Newton et al.,2005; Porro et al., 2000; Verstynen et al., 2005; Ziemann and Hallett,2001). The functional signi!cance of ipsilateral S1-M1 coherent activ-ity is nevertheless unclear. Indeed, it might re"ect an enhanced inter-hemispheric inhibition or a direct involvement of ipsilateral S1-M1cortical areas in motor control and sensory integration during repeti-tive hand movements (Kicic et al., 2008; Kobayashi et al., 2003;Stoeckel and Binkofski, 2010). Both hypotheses could account forour results since inhibitory activity as well as ipsilateral motor controland sensory integration should share a common periodicity withhand kinematics. However, as we did not control for the occurrenceof left upper limb muscle activity or sensory inputs, we cannot ensurethat the coherent activity found in S1-M1 cortices ipsilateral to handmovement was not related to such factors. So, the existence of
Table 2Coherence values and MNI coordinates for signi!cant coherent cortical areas observed at the group level in both TOUCH and noTOUCH conditions at the frequency of hand move-ment (F0) and its !rst harmonic (F1). For each identi!ed brain areas, the number n of individual subjects presenting a local maximum at the group local maximum is indicated. Datafor brain regions contralateral (c) and ipsilateral (i) to the movements are presented.
TOUCH, F0 TOUCH, F1 noTOUCH, F0 noTOUCH, F1
cM1 0.24 (n=7) 0.34 (n=9) 0.21 (n=7) 0.22 (n=8)[#44 #18 60] [#40 #18 60] [#41 #16 58] [#38 #18 60]
cS1 0.30 (n=10) x x x[#52 #34 50]
cS1 medial 0.21 (n=9) 0.12 (n=5) x x[#18 #32 74] [#6 #36 76]
cM1 medial 0.17 (n=9) x x x[#18 #10 72]
cDLPFC x 0.10 (n=5) x 0.06 (n=5)[#44 26 40] [#44 26 40]
cPPC 0.17 (n=9) x x 0.09 (n=4)[#24 #46 70] [#24 #52 70]
iS1-M1 0.16 (n=7) x 0.07 (n=5) x[44 #20 60] [42 #14 58]
± 0.080.36-0.36
± 0.070.13-0.13
F0 F1
Fig. 4. Difference of f-transformed TOUCH–noTOUCH group-level maps at F0 (Left) andF1 (Right). Maps are thresholded at signi!cance level (Pb0.05). Only positive values,corresponding to a higher coherence level in TOUCH as compared to noTOUCH weresigni!cant.
1689M. Bourguignon et al. / NeuroImage 59 (2012) 1684–1691

Author's personal copy
ipsilateral S1-M1 coherent activity in the context of self-paced repet-itive movements should be investigated in further studies controllingfor these aspects.
The DLPFC is thought to integrate multiple instructions to gener-ate an action plan; it is a major control center of goal-directed behav-iors (Abe and Hanakawa, 2009). In addition, this brain region isactivated during self-initiated tasks, in which decisions are requiredabout the timing of movements (Jenkins et al., 2000; Karch et al.,2009). The DLPFC coherent activity might therefore re"ect the in-volvement of this brain region in the self-initiation of repetitivehand/!ngers movements.
Signi!cant coupling was observed at F0 and F1 in TOUCH in medi-al S1-M1 areas contralateral to hand movement. According to S1 andM1 classical somatotopy, coherent activity in the medial part of S1-M1 area should re"ect motor control and somatosensory integrationfrom the proximal upper limb extremity. However, this medial partof the precentral gyrus is involved in some aspects of !ngers move-ments as reported by some studies challenging the classic view ofPen!eld's somatotopic organization of S1-M1 area (Geyer et al.,2000; Schieber, 2001). Importantly, these studies suggested that con-trol of each body part is mediated by an extensive territory overlap-ping with the territories controlling other body parts. In particular,electrical stimulation of some medial M1 cortical areas has beenshown to induce either !ngers extension or jerking of the wholearm (Schieber, 2001). The absence of coherent activity in medial S1-M1 areas contralateral to hand movement in the noTOUCH conditionmight suggest that coherence in these brain regions relies on tactileinputs, but this aspect requires further investigations.
The PPC is known to be involved in sensori-motor integration, informing plans for movements, and in monitoring the goal as well asthe dynamic state of the trajectory during movement (Buneo and An-dersen, 2006; Mulliken et al., 2008; Wolpert et al., 1998). The highercoherence level observed in PPC during TOUCH compared withnoTOUCH is in line with this sensori-motor integration hypothesis.The movement monitoring performed by the PPC is supposed to beinvolved in continuous movement control and error detection (Mulli-ken et al., 2008). Its involvement in the CKC phenomenon is in linewith previous studies in non-human primates, which showed thatduring a copy task by joystick manipulation, neuronal ensemble re-cordings in PPC enabled hand speed prediction (Averbeck et al.,2005). Task-related coupling in this cortical area was also identi!edin humans using hand speed or movement frequency and MEG corti-cal signals (Jerbi et al., 2007; Pollok et al., 2004a; Pollok et al., 2005).These data suggest that the CKC phenomenon involves some degreeof somatosensory input integration to organize and control self-paced repetitive hand movements.
Previous studies investigating the coupling between hand speedor movement frequency and MEG signals have shown the existenceof signi!cant coupling in the supplementary motor cortex (SMC) aswell as in the cerebellum ipsilateral to hand movement (Jerbi et al.,2007; Pollok et al., 2004a; Pollok et al., 2005). In this study, local co-herence maxima were not observed in these brain regions in grouplevel coherence maps, even though difference maps indirectly pointsto possible SMC and premotor cortex involvement in CKC phenome-na. The absence of discernible local coherent maxima in the SMC ingroup level coherence maps is challenging since this brain region isknown to be involved in self-initiated movements (Nachev et al.,2008). Apart from resolution issues inherent to the method, it couldbe related to the movement type since SMC mainly contributes tothe control of action sequences rather than !xed movements asused in this study (Deiber et al., 1999; Nachev et al., 2008). The ab-sence of coupling in the cerebellum could be explained by the factthat this structure is mainly involved in externally triggered com-pared to self-initiated movements (Debaere et al., 2003; Jueptner etal., 1996; Ogawa et al., 2006; Taniwaki et al., 2006). Some studies in-deed suggest that the cerebellum uses sensory information to control
voluntary movement (Jueptner et al., 1996). In addition, some datasuggest that the cerebellum is mainly involved in the control of dis-crete compared to continuous movements (Spencer et al., 2007;Spencer et al., 2003).
Conclusion
This study demonstrates that neuronal activity within a largesensori-motor cortical network is coherent with hand kinematicsduring natural self-paced repetitive hand movements. The mainsource of the coherent activity at the movement frequency is locatedin M1 or S1 hand cortical area contralateral to hand movementdepending on the amount of tactile inputs. Coherence values are sig-ni!cantly higher when the movement task is associated with a highlevel of tactile inputs, highlighting the key role of somatosensoryafferences in maintaining a high level of synchronization betweenhand acceleration and the neuronal activity in coherent brain areas.Other weaker local coherence maxima were observed at corticalareas involved in motor planning and organization as well as in sen-sory inputs integration in the hemispheres contralateral and ipsilateralto hand movement.
Acknowledgment
Mathieu Bourguignon bene!ts of a research grant from the FRIA(FRS-FNRS, Belgium). Xavier De Tiège is Clinicien-Chercheur Spécia-liste at the FRS-FNRS, Belgium. This work was supported by a “BrainsBack to Brussels” grant to VJ from the Institut d'Encouragement de laRecherche Scienti!que et de l'Innovation de Bruxelles (Brussels, Bel-gium), the ERC Advanced Grant #232946, the Fonds de la RechercheScienti!que (FRS-FNRS, Belgium, Research Convention 3.4611.08)and the Academy of Finland (National Centers of Excellence Program2006–2011). We thank Helge Kainulainen and Ronny Schreiber at theBrain Research Unit (Aalto University School of Science, Espoo, Fin-land) for technical support. We thank Pr. Riitta Hari for substantialcomments on the manuscript and study.
References
Abe, M., Hanakawa, T., 2009. Functional coupling underlying motor and cognitive func-tions of the dorsal premotor cortex. Behav. Brain Res. 198, 13–23.
Ashe, J., Georgopoulos, A.P., 1994. Movement parameters and neural activity in motorcortex and area 5. Cereb. Cortex 4, 590–600.
Averbeck, B.B., Chafee, M.V., Crowe, D.A., Georgopoulos, A.P., 2005. Parietal representa-tion of hand velocity in a copy task. J. Neurophysiol. 93, 508–518.
Baker, S.N., 2007. Oscillatory interactions between sensorimotor cortex and the periph-ery. Curr. Opin. Neurobiol. 17, 649–655.
Baker, M.R., Baker, S.N., 2003. The effect of diazepam on motor cortical oscillations andcorticomuscular coherence studied in man. J. Physiol. 546, 931–942.
Bortel, R., Sovka, P., 2007. Approximation of statistical distribution of magnitudesquared coherence estimated with segment overlapping. Signal Process. 87,1100–1117.
Bourguignon, M., De Tiège, X., Op de Beeck, M., Pirotte, B., Van Bogaert, P., Goldman, S.,Hari, R., Jousmäki, V., 2011. Functional motor-cortex mapping using corticokine-matic coherence. NeuroImage 55, 1475–1479.
Bradberry, T.J., Rong, F., Contreras-Vidal, J.L., 2009. Decoding center-out hand velocityfrom MEG signals during visuomotor adaptation. NeuroImage 47, 1691–1700.
Buneo, C.A., Andersen, R.A., 2006. The posterior parietal cortex: sensorimotor interfacefor the planning and online control of visually guided movements. Neuropsycholo-gia 44, 2594–2606.
Caminiti, R., Johnson, P.B., Urbano, A., 1990. Making arm movements within differentparts of space: dynamic aspects in the primate motor cortex. J. Neurosci. 10,2039–2058.
Carmena, J.M., Lebedev, M.A., Crist, R.E., O'Doherty, J.E., Santucci, D.M., Dimitrov, D.F.,Patil, P.G., Henriquez, C.S., Nicolelis, M.A., 2003. Learning to control a brain-machine interface for reaching and grasping by primates. PLoS Biol. 1, E42.
Coltz, J.D., Johnson, M.T., Ebner, T.J., 1999. Cerebellar Purkinje cell simple spike dis-charge encodes movement velocity in primates during visuomotor arm tracking.J. Neurosci. 19, 1782–1803.
Cramer, S.C., Finklestein, S.P., Schaechter, J.D., Bush, G., Rosen, B.R., 1999. Activation ofdistinct motor cortex regions during ipsilateral and contralateral !nger move-ments. J. Neurophysiol. 81, 383–387.
Debaere, F., Wenderoth, N., Sunaert, S., Van Hecke, P., Swinnen, S.P., 2003. Internal vsexternal generation of movements: differential neural pathways involved in
1690 M. Bourguignon et al. / NeuroImage 59 (2012) 1684–1691

Author's personal copy
bimanual coordination performed in the presence or absence of augmented visualfeedback. NeuroImage 19, 764–776.
Deiber, M.P., Honda, M., Ibanez, V., Sadato, N., Hallett, M., 1999. Mesial motor areas inself-initiated versus externally triggered movements examined with fMRI: effect ofmovement type and rate. J. Neurophysiol. 81, 3065–3077.
Ebner, T.J., Hendrix, C.M., Pasalar, S., 2009. Past, present, and emerging principles in theneural encoding of movement. Adv. Exp. Med. Biol. 629, 127–137.
Faes, L., Pinna, G.D., Porta, A., Maestri, R., Nollo, G., 2004. Surrogate data analysis forassessing the signi!cance of the coherence function. IEEE Trans. Biomed. Eng. 51,1156–1166.
Georgopoulos, A.P., Kettner, R.E., Schwartz, A.B., 1988. Primate motor cortex and freearm movements to visual targets in three-dimensional space. II. Coding of the di-rection of movement by a neuronal population. J. Neurosci. 8, 2928–2937.
Geyer, S., Matelli, M., Luppino, G., Zilles, K., 2000. Functional neuroanatomy of the pri-mate isocortical motor system. Anat. Embryol. (Berl) 202, 443–474.
Gross, J., Kujala, J., Hämäläinen, M., Timmermann, L., Schnitzler, A., Salmelin, R., 2001.Dynamic imaging of coherent sources: studying neural interactions in the humanbrain. Proc. Natl. Acad. Sci. U. S. A. 98, 694–699.
Haaland, K.Y., Elsinger, C.L., Mayer, A.R., Durgerian, S., Rao, S.M., 2004. Motor sequencecomplexity and performing hand produce differential patterns of hemispheric lat-eralization. J. Cogn. Neurosci. 16, 621–636.
Halliday, D.M., Rosenberg, J.R., Amjad, A.M., Breeze, P., Conway, B.A., Farmer, S.F., 1995.A framework for the analysis of mixed time series/point process data–theory andapplication to the study of physiological tremor, single motor unit dischargesand electromyograms. Prog. Biophys. Mol. Biol. 64, 237–278.
Hanakawa, T., Parikh, S., Bruno, M.K., Hallett, M., 2005. Finger and face representationsin the ipsilateral precentral motor areas in humans. J. Neurophysiol. 93,2950–2958.
Hutchinson, S., Kobayashi, M., Horkan, C.M., Pascual-Leone, A., Alexander, M.P.,Schlaug, G., 2002. Age-related differences in movement representation. Neuro-Image 17, 1720–1728.
Jenkins, I.H., Jahanshahi, M., Jueptner, M., Passingham, R.E., Brooks, D.J., 2000. Self-ini-tiated versus externally triggered movements. II. The effect of movement predict-ability on regional cerebral blood "ow. Brain 123 (Pt 6), 1216–1228.
Jerbi, K., Lachaux, J.P., N'Diaye, K., Pantazis, D., Leahy, R.M., Garnero, L., Baillet, S., 2007.Coherent neural representation of hand speed in humans revealed by MEG imag-ing. Proc. Natl. Acad. Sci. U. S. A. 104, 7676–7681.
Jueptner, M., Jenkins, I.H., Brooks, D.J., Frackowiak, R.S., Passingham, R.E., 1996. Thesensory guidance of movement: a comparison of the cerebellum and basal ganglia.Exp. Brain Res. 112, 462–474.
Kakei, S., Hoffman, D.S., Strick, P.L., 1999. Muscle and movement representations in theprimary motor cortex. Science 285, 2136–2139.
Kansaku, K., Muraki, S., Umeyama, S., Nishimori, Y., Kochiyama, T., Yamane, S.,Kitazawa, S., 2005. Cortical activity in multiple motor areas during sequential!nger movements: an application of independent component analysis. NeuroImage28, 669–681.
Karch, S., Mulert, C., Thalmeier, T., Lutz, J., Leicht, G., Meindl, T., Moller, H.J., Jager, L.,Pogarell, O., 2009. The free choice whether or not to respond after stimulus presen-tation. Hum. Brain Mapp. 30, 2971–2985.
Kelso, J.A., Fuchs, A., Lancaster, R., Holroyd, T., Cheyne, D., Weinberg, H., 1998. Dynamiccortical activity in the human brain reveals motor equivalence. Nature 392,814–818.
Kicic, D., Lioumis, P., Ilmoniemi, R.J., Nikulin, V.V., 2008. Bilateral changes in excitability ofsensorimotor cortices during unilateral movement: combined electroencephalo-graphic and transcranial magnetic stimulation study. Neuroscience 152, 1119–1129.
Kobayashi, M., Hutchinson, S., Schlaug, G., Pascual-Leone, A., 2003. Ipsilateral motorcortex activation on functional magnetic resonance imaging during unilateralhand movements is related to interhemispheric interactions. NeuroImage 20,2259–2270.
Lewis, S.M., Jerde, T.A., Tzagarakis, C., Georgopoulos, M.A., Tsekos, N., Amirikian, B., Kim,S.G., Ugurbil, K., Georgopoulos, A.P., 2003. Cerebellar activation during copyinggeometrical shapes. J. Neurophysiol. 90, 3874–3887.
Mehring, C., Rickert, J., Vaadia, E., Cardosa de Oliveira, S., Aertsen, A., Rotter, S., 2003.Inference of hand movements from local !eld potentials in monkey motor cortex.Nat. Neurosci. 6, 1253–1254.
Moran, D.W., Schwartz, A.B., 1999. Motor cortical representation of speed and directionduring reaching. J. Neurophysiol. 82, 2676–2692.
Mulliken, G.H., Musallam, S., Andersen, R.A., 2008. Forward estimation of movementstate in posterior parietal cortex. Proc. Natl. Acad. Sci. U. S. A. 105, 8170–8177.
Nachev, P., Kennard, C., Husain, M., 2008. Functional role of the supplementary andpre-supplementary motor areas. Nat. Rev. Neurosci. 9, 856–869.
Newton, J.M., Sunderland, A., Gowland, P.A., 2005. fMRI signal decreases in ipsilateralprimary motor cortex during unilateral hand movements are related to durationand side of movement. NeuroImage 24, 1080–1087.
Ogawa, K., Inui, T., Sugio, T., 2006. Separating brain regions involved in internally guid-ed and visual feedback control of moving effectors: an event-related fMRI study.NeuroImage 32, 1760–1770.
Old!eld, R.C., 1971. The assessment and analysis of handedness: the Edinburgh inven-tory. Neuropsychologia 9, 97–113.
O'Suilleabhain, P.E., Lagerlund, T.D., Matsumoto, J.Y., 1999. Cortical potentials at thefrequency of absolute wrist velocity become phase-locked during slow sinusoidaltracking movements. Exp. Brain Res. 126, 529–535.
Pohja, M., Salenius, S., 2003. Modulation of cortex-muscle oscillatory interaction byischaemia-induced deafferentation. Neuroreport 14, 321–324.
Pollok, B., Gross, J., Dirks, M., Timmermann, L., Schnitzler, A., 2004a. The cerebral oscil-latory network of voluntary tremor. J. Physiol. 554, 871–878.
Pollok, B., Muller, K., Aschersleben, G., Schnitzler, A., Prinz, W., 2004b. The role of theprimary somatosensory cortex in an auditorily paced !nger tapping task. Exp.Brain Res. 156, 111–117.
Pollok, B., Gross, J., Muller, K., Aschersleben, G., Schnitzler, A., 2005. The cerebral oscil-latory network associated with auditorily paced !nger movements. NeuroImage24, 646–655.
Porro, C.A., Cettolo, V., Francescato, M.P., Baraldi, P., 2000. Ipsilateral involvement ofprimary motor cortex during motor imagery. Eur. J. Neurosci. 12, 3059–3063.
Reina, G.A., Moran, D.W., Schwartz, A.B., 2001. On the relationship between joint angu-lar velocity and motor cortical discharge during reaching. J. Neurophysiol. 85,2576–2589.
Rosenberg, J.R., Amjad, A.M., Breeze, P., Brillinger, D.R., Halliday, D.M., 1989. The Fourierapproach to the identi!cation of functional coupling between neuronal spiketrains. Prog. Biophys. Mol. Biol. 53, 1–31.
Schieber, M.H., 2001. Constraints on somatotopic organization in the primary motorcortex. J. Neurophysiol. 86, 2125–2143.
Semmler, J.G., Nordstrom, M.A., 1998. Motor unit discharge and force tremor in skill-and strength-trained individuals. Exp. Brain Res. 119, 27–38.
Serrien, D.J., Fisher, R.J., Brown, P., 2003. Transient increases of synchronized neural ac-tivity during movement preparation: in"uence of cognitive constraints. Exp. BrainRes. 153, 27–34.
Spencer, R.M., Zelaznik, H.N., Diedrichsen, J., Ivry, R.B., 2003. Disrupted timing of dis-continuous but not continuous movements by cerebellar lesions. Science 300,1437–1439.
Spencer, R.M., Verstynen, T., Brett, M., Ivry, R., 2007. Cerebellar activation during dis-crete and not continuous timed movements: an fMRI study. NeuroImage 36,378–387.
Stark, E., Drori, R., Asher, I., Ben-Shaul, Y., Abeles, M., 2007. Distinct movement param-eters are represented by different neurons in the motor cortex. Eur. J. Neurosci. 26,1055–1066.
Stoeckel, M.C., Binkofski, F., 2010. The role of ipsilateral primary motor cortex in move-ment control and recovery from brain damage. Exp. Neurol. 221, 13–17.
Taniwaki, T., Okayama, A., Yoshiura, T., Togao, O., Nakamura, Y., Yamasaki, T., Ogata, K.,Shigeto, H., Ohyagi, Y., Kira, J., Tobimatsu, S., 2006. Functional network of the basalganglia and cerebellar motor loops in vivo: different activation patterns betweenself-initiated and externally triggered movements. NeuroImage 31, 745–753.
Taulu, S., Simola, J., Kajola, M., 2005. Applications of the signal space separation meth-od. Signal Process., IEEE Trans. 53, 3359–3372.
Turner, R.S., Grafton, S.T., Votaw, J.R., Delong, M.R., Hoffman, J.M., 1998. Motor subcir-cuits mediating the control of movement velocity: a PET study. J. Neurophysiol.80, 2162–2176.
Verstynen, T., Diedrichsen, J., Albert, N., Aparicio, P., Ivry, R.B., 2005. Ipsilateral motorcortex activity during unimanual hand movements relates to task complexity. J.Neurophysiol. 93, 1209–1222.
Witham, C.L., Wang, M., Baker, S.N., 2010. Corticomuscular coherence between motorcortex, somatosensory areas and forearm muscles in the monkey. frontiers in sys-tems neuroscience 4 article 38, 14.
Wolpert, D.M., Goodbody, S.J., Husain, M., 1998. Maintaining internal representations:the role of the human superior parietal lobe. Nat. Neurosci. 1, 529–533.
Ziemann, U., Hallett, M., 2001. Hemispheric asymmetry of ipsilateral motor cortex ac-tivation during unimanual motor tasks: further evidence for motor dominance.Clin. Neurophysiol. 112, 107–113.
1691M. Bourguignon et al. / NeuroImage 59 (2012) 1684–1691