nectar biodiversity: a short review
TRANSCRIPT

Nectar biodiversity: a short review
E. Pacini1, M. Nepi
1, and J. L. Vesprini
2
1Department of Environmental Sciences ‘‘G. Sarfatti’’, Siena, Italy2Faculty of Agricultural Sciences, Zavalla, Argentina
Received July 23, 2002; accepted September 18, 2002Published online: June 2, 2003� Springer-Verlag 2003
Abstract. Nectaries differ in many aspects but acommon feature is some kind of advantage for theplant conferred by foraging of consumerswhichmaydefend the plant from predators in the case ofextrafloral nectaries, or be agents of pollination inthe case of floral nectaries. This minireview isconcerned mainly with floral nectaries and examinesthe following characteristics: position in flower;nectary structure; origin of carbohydrates, amino-acids andproteins;manner of exposure of nectar; siteof nectar presentation; volume and production ofnectar in time; sexual expression of flower andnectary morphology; nectar composition and floralsexual expression; variability of nectar composition;fate of nectar; energy cost of nectar production. Thespecies of certain large families, such asBrassicaceae,Lamiaceae and Asteraceae, resemble each other innectary organisation; other families, such as Cucur-bitaceae and Ranunculaceae, have various types oforganisation. A scheme is presented to illustratefactors influencing nectary and nectar biodiversity.
Key words: Floral nectaries, nectar, secretion,carbohydrates.
Introduction
The term nectary does not indicate a welldefined anatomical structure because there arevarious types of nectary with different
anatomical origins and positions. The termhas ecological significance, in that nectaries arethe places where liquid substances involved ininteractions with animals are produced andoffered.
The most ancient plant with nectaries stillliving today is in Pteridophyta. Pteridiumaquilinum, a quasi-cosmopolitan fern in whichnectaries provide rewards for ants that defendthe plants from predators (Heads and Lawton1985 and references therein). These authorsdemonstrated nectaries in fern populations inGreat Britain, but they were not found inpopulations in South Africa (Rashbrook et al.1992). Thus, ecological significance of nectar-ies may vary in relation to environment.
In gymnosperms, nectaries are only foundin certain Chlamydospermae such as Ephedrasp. and Welwitschia, being involved in polli-nation (Bino et al. 1984, Wetschnig andDepisch 1999). Nectaries are more commonin angiosperms, dating back to the LateCretaceous, when they were both floral andextrafloral (Friis and Endress 1990). Theformer are involved in pollination and thelatter in rewards for animals, mainly ants, thatkeep predators away, but in some cases theyare also involved in pollination. Examples ofthis type are the cyathial nectaries of the genus
Plant Syst. Evol. 238: 7–21 (2003)DOI 10.1007/s00606-002-0277-y

Euphorbia and the stipules of leaves in Acacia(Kenrick et al. 1987 and references therein).
Fahn (1952) gives an anatomical andfunctional description of floral nectaries, iden-tifying five types. In a later paper (Fahn 1979),he identifies 12 topographic positions of floraland extrafloral nectaries. Vogel (1997, 1998a,1998b, 1998c) gives an exhaustive descriptionof the type and structure of nectaries in themain angiosperm families. Percival (1961)describes types of nectaries in 889 species ofangiosperms. Nectaries vary in morphology,nectar varies in chemical composition andthese characteristics reflect the type of pollina-tor (Baker and Baker 1983).
There are families of angiosperms in whichall species have nectaries, whereas nectaries areabsent in Orchidaceae wherever pollinationoccurs by deception (Neiland and Wilcock1998). These authors noticed that in thisfamilies reproductive success was statisticallygreater when nectaries were present.
Schmid (1988) published a paper on termi-nology which summarises research on nectariesup to the 1980s. A detailed survey of distribu-tion, types and evolution of monocot nectarieswas given by Smets et al. (2000); data on dicotsis divided among various papers (Schmid 1988and references therein).
The present minireview is mainly con-cerned with floral nectaries. Its aim is toexamine cytological and ecological aspectsrather than anatomical and topographic ones.The latter are well known and are more easilystudied and interpreted.
Observations and discussion
Position of nectaries in flowers
Floral nectaries may be at the same level as thesurface of the organ that bears them, form aprotrusion, or be deeply embedded, as in thecase of septate nectaries (Fahn 1979). If they
Table 1. Relationships between features of nectary and nectar and some environmental parameters. Thepresence of ‘‘X’’ means a correlation between parameters
Pollinatortype
Pollinatorchoices
Pollencharge anddischarge
Predation Bloomingperiod
No. ofovules perovary
– Floral nectaryposition
X X X
– Nectary structure X X X– Nectar carbohydratesource
X X
– Nectar productionrate
X X
– Site and mode ofnectar presentation
X X
– Nectar volume andproduction in time
X X X X X
– Flower sexualexpression andnectary morphology
X X
– Nectar compositionand flower sexualexpression
X X
– Variability of nectarcomposition
X X X
– Fate of nectar X X
8 E. Pacini et al.: Nectar biodiversity

are exposed, as in the case of Hedera helix, thenectar may be collected by many animalswith different pollination efficiencies. Exposednectar also evaporates readily and is notprotected by insect predation. On the otherhand, if the nectar is secreted inside thecorolla, or in a spur, it is protected againstevaporation, especially if the cuticle of thefloral part collecting nectar is thick, but it isnot protected from robbery (Inouye 1983), seealso Table 1.
Nectary structure
The nectary consists of three components: a.epidermis with or without stomata whichnormally mediates nectar release; b. parenchy-ma that produces or stores substances thatbecome dissolved in the nectar; c. the vascularbundle which conveys water or nutrients to theparenchyma. These three parts may varygreatly, depending on the type of nectary.The reasons for this diversity depend largelyon the type of pollinator, the manner in whichpollen is loaded/unloaded and on the numberof ovules per ovary (Table 1).
The epidermis of the nectary may be:
1. flat;1.1 devoid of stomata as in the case of
Helleborus foetidus (Fig. 1);1.2 with stomata as in the case of Cucurbita
pepo (Figs. 1 and 2);2. with secreting hairs as in Cyclanthera pedata
(Figs. 1, 2, 3).
The nectar may be exposed in microdroplets atthe end of hairs or in a continuous layer overthe nectary surface. There is normally a cuticle(Fig. 4) but it may be perforated to enablenectar to go out to exit or the cuticle may havemodified permeability, as in C. pepo in whichnectar uncollected by pollinators is resorbed(Nepi et al. 1996a).
Parenchyma specialised for nectar secretionmay be mono- or multi-layered. Its volumeis proportional to the quantity of nectarproduced and hence the type and number ofpollinator visits to the nectary (Table 1).
The vascular bundle which supplies theparenchyma may consist of the followingelements, depending on secretory mode: a.phloem only, e.g. floral nectaries of Brassicanapus (Davis et al. 1986), Vicia faba (Daviset al. 1988), Digitalis purpurea (Gaffal et al.1998) and Cyclanthera pedata (Fig. 3); b. phlo-em and xylem: as in extrafloral nectaries ofVicia faba (Davis et al. 1988) and otherexamples in Fahn (2000).
From a systematic viewpoint, there arefamilies, such as the Brassicaceae, Lamiaceaeand Asteraceae, in which the nectaries aresimilar throughout the family (for Asteraceaesee Mani and Saravanan 1999). On the otherhand, at least two different types exist in theCucurbitaceae, with nectar being secreted intwo different ways but presented in only oneway (Fig. 1). In Cucurbita pepo, the nectarcomes out of stomata (Figs. 1, 2) and is exposedin a continuous liquid layer. InCyclanthera, theepidermis has multicellular hairs and the nec-tar, produced in the hair tip, builds up andforms a continuous layer (Fig. 1).
Origin of nectar carbohydrates
According to Fahn (2000), the origin ofsecreted nectar is the phloem sap. Directly orphotosynthesis in the parenchyma. This meansindividual and seasonal variations, especiallyin plants that bloom over long periods(Table 1). The parenchyma with chlorophyllmay belong to the nectary itself, as in Cyclan-thera pedata (Figs. 1, 5a, b), certain Ranuncul-aceae such as Helleborus (Vesprini et al. 1999),and Brassicaceae, or it may be in other parts ofthe flower or plant, as in Cucurbita pepo (Nepiet al. 1996b).
In Cucurbita and Passiflora (Table 2),products of photosynthesis by parts of flowersother than nectaries or by other parts of theplant, accumulate in the nectar-producingparenchyma as starch in amyloplasts, begin-ning several days before anthesis. Amyloplastsin the nectar-producing parenchyma aregenerally almost devoid of stroma becausethey are fully packed with starch (Nepi et al.
E. Pacini et al.: Nectar biodiversity 9

2
Nectarsecretion
APAP
AP
CP
CP
Beforesecretion
Dur
ing
secr
etio
n
Cucurbita pepo Cyclanthera pedata Helleborus foetidus
Nectarexposure
Nectarconsumption
Eventualnectarresorption
PP
PPPP
CP CP
AP
APAP
CP
CP1223
45678
CP
CP
CP
CP
1
2
3456
7
8
1
2
345
678
Fig. 1. Scheme of formation, exposure and presentation of nectar in three species differing in aspects such asorigin of sugar component of nectar, presentation and reabsorption. Nectar is shown as dots. Amyloplasts(AP), proplastids (PP), chloroplasts (CP). In Cucurbita pepo nectar is derived from hydrolysis of starch in theparenchyma. Nectar is exposed as a continuous layer on the epidermis and is resorbed if not consumed. InCyclanthera pedata it is produced by photosynthesis by the parenchyma, secreted through pluricellulartrichomes and presented as a continuous layer on the epidermis. If not collected, it is not resorbed. InHelleborusfoetidus the nectary surface is disposed vertically, the sugar component is produced by the parenchyma but thenectar is exposed gradually by epidermal cells which empty completely and die. After daily secretion, theepidermis and the nectary reorganise. The nectar flows along the nectary and that not collected falls to theground
Fig. 2. Cucurbita pepo PAS stained nectary producing nectar (arrows). There is a monolayered epidermis (E )with stomata (s) and nectar producing parenchyma (P) containing starch. Nectar accumulates in the stomatacavities (arrow) and on the surface of the nectary (arrowheads). Nectar is PAS positive, viscous and notremoved during fixation and dehydration. Bar¼ 50 lm. Fig. 3. Cyclanthera pedata PAS stained nectary. Thenectary is of the trichome type consisting of a monolayered epidermis (E ) with secretory capitate trichomes andparenchyma with chloroplasts, not evident in the picture, with vascular bundles (vb) and phloem. The trichomeshave a peduncle consisting of two cells and an apex consisting of 6–7 cells. Bar¼ 50 lm. Fig. 4. Cyclantherapedata auramine O stained nectary. A cuticle (arrow) is more evident above the trichome than above the otherepidermal cells. Bar¼ 75 lm. Fig. 5a and b. Hand section of Cyclanthera pedata nectary observed in brightfield (5a) and UV light (5b); fluorescence is from chloroplasts in parenchyma cells. Trichomes do not containchloroplasts and do not fluoresce. Bar¼ 200 lm
c
10 E. Pacini et al.: Nectar biodiversity
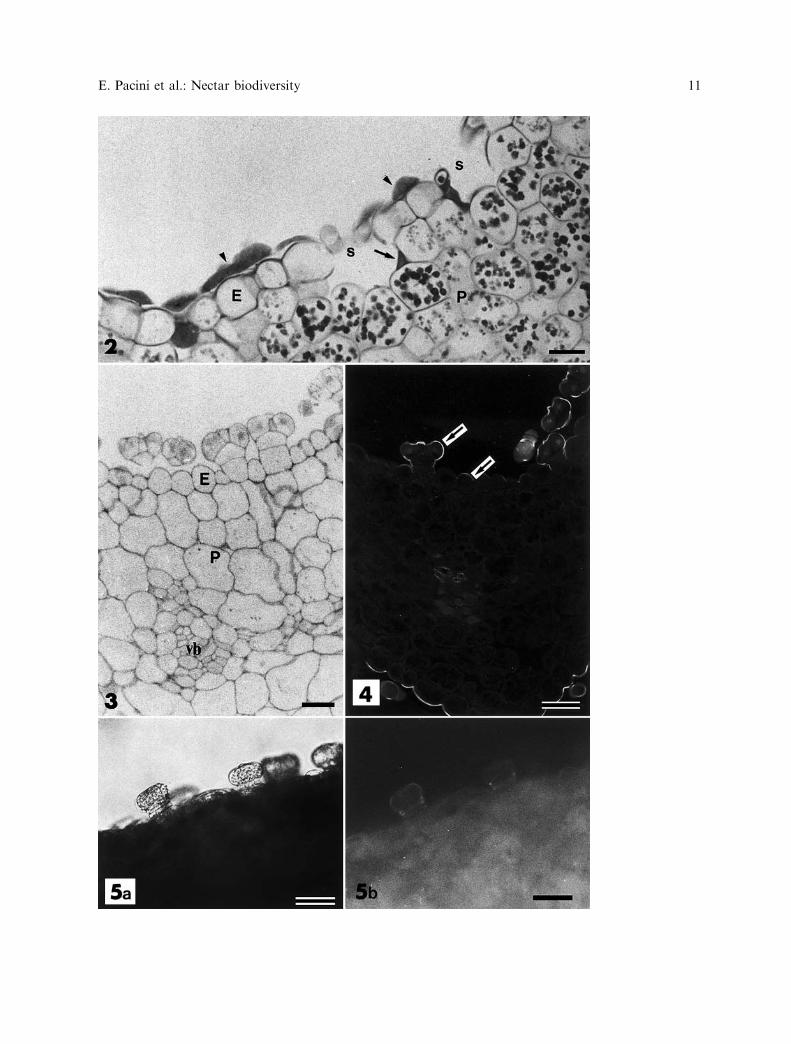
E. Pacini et al.: Nectar biodiversity 11

1996b). Moreover, there are many starchgranules per amyloplast; this increases starchsurface area, facilitating and speeding hydro-lysis at the time of nectar production andsecretion. The starch is hydrolysed before thestart of secretion (Figs. 1, 2) as seen inCucurbita (Nepi et al. 1996a, b), Passiflorasp. (Durkee et al. 1981), Hibiscus rosa-sinensis(Sawidis 1998), Rosmarinus officinalis (Zer andFahn 1992), Pisum sativum (Razem and Davis1999) and the orchid Limodorum abortivum(Figueiredo and Pais 1992). Nectar productionmay vary in quantity (Table 2); when it is veryabundant amyloplasts may even accumulate inepidermal cells (Nepi et al. 1996b). The abun-dance of starch in parenchyma cells of Cucur-bita pepo and Passiflora biflora (Nepi et al.1996b, Durkee 1982, Durkee et al. 1981) hasbeen linked to the high rate of production ofnectar with a high sugar concentration.
Depending on the origin of the nectar, thenectary parenchyma may be of two kinds, itmay have chlorophyll or it may be reserveparenchyma for starch storage (Figs. 1, 6). Inthe former, starch is produced directly byphotosynthesis and occurs in small quantities,stored for only a few hours, to be hydrolysedat night; photosynthesis may proceed for manydays, producing small quantities of starch andnectar every day.
There would seem no reason why bothtypes of parenchyma should not occur simul-taneously or in succession, as reported inSanango racemosum (Gesneriaceae) (Maldona-do and Otegui 1997). In this species the nectar-producing parenchyma also contains starch.According to these authors, the starch does notcontribute to nectar production but is, never-theless, gone by the end of anthesis.
Such a process would be regulated byplastid conversion processes, as amyloplastsare normally a final stage of plastid differen-tiation (Wheatley 1977).
Once nectar secretion has ended, the fate ofthe two types of parenchyma is again different.Photosynthesizing parenchyma may continueto function for a while, with the productsgoing to other activities, such as growth of the
ovary. Finally, the nectary degenerates. Stor-age parenchyma just seems to degenerate as inCucurbita (Nepi et al. 1996b).
InHelleborus bocconei andH. foetidus bothof which have both types of parenchyma,the two parenchymas have the same modeof secretion but the origin of the sugar compo-nent of nectar is slightly different. In H. bocc-onei, the photosynthesizing parenchyma seemsto produce all the nectar, whereas inH. foetidusthere must be a contribution from photosyn-thesis by other parts of the flower and/or plant.H. bocconei has an intense green and exposednectary and its cells are packed with chlorop-lasts; the nectar, predominantly sucrose (meansucrose concentration 275.18 mg/ml), is pro-duced when the flower is exposed. InH. foetidusthe nectary is always protected within theflower, it is pale green and the nectar, whichis sucrose dominant (607.81 mg/ml), is pro-duced before the flower opens. These charactersand the different mean sucrose contents suggestthat some of the nectar originates elsewhere(Vesprini et al. 1999 and our unpublished data).
The two modes of nectar carbohydrateproduction may serve to produce nectar con-sumed by pollinators at different times of day(Fig. 6). If sugars are derived from photosyn-thesis in the parenchyma itself, they may beproduced over a long period, even weeks as inthe floral nectaries of Eucalyptus (Davis 1997)and Helleborus (Vesprini et al. 1999), and in allextrafloral nectaries (Table 2). However,because production only occurs by day, thenectar must be ready at dusk if pollination isnocturnal. To have nectar ready at sunset,starch stored in chloroplasts during the daywould have to be quickly hydrolysed at dusk.That is unlike what happens in leaves, wherehydrolysis is slower and continues throughoutthe night. Photosynthesizing nectaries haveintermittent nectar production because nectarcan be produced only during the day. Howeverif material for nectar sugar comes fromhydrolysis of starch stored in the parenchyma,nectar may be produced at any time of day,even before sunrise. By this method it is alsopossible to producemuch nectar in a short time.
12 E. Pacini et al.: Nectar biodiversity
The difference between the two modes of nectarcarbohydrate formationare compared inFig. 6.
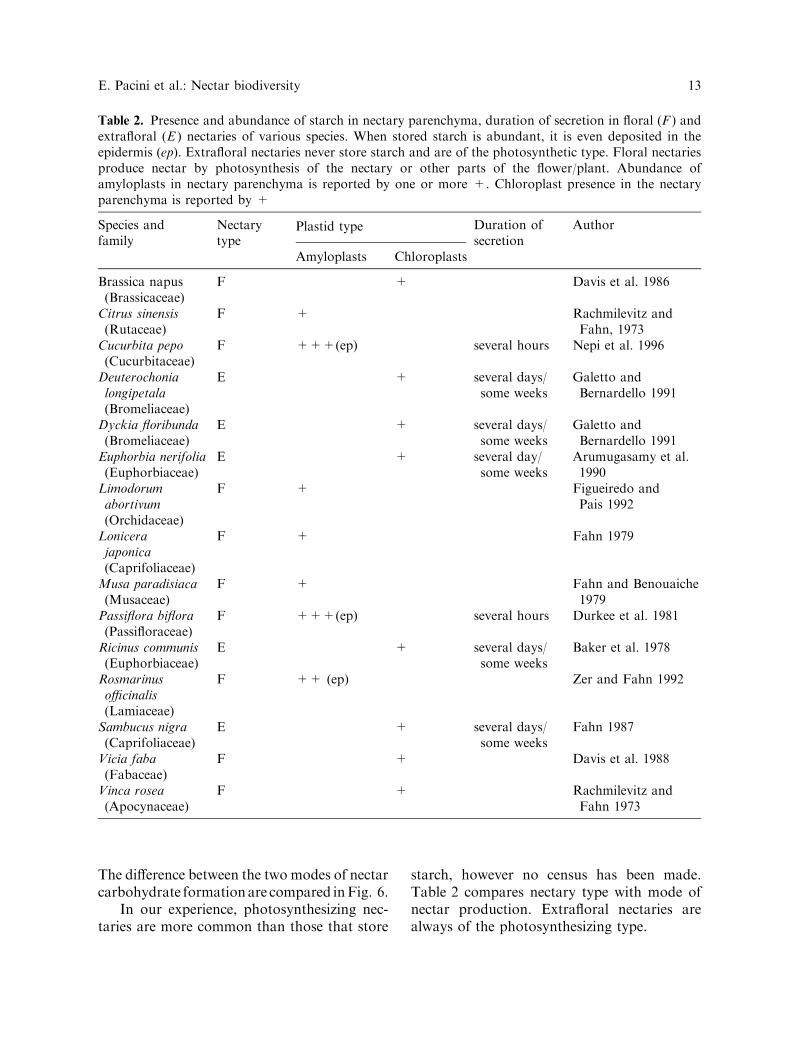
In our experience, photosynthesizing nec-taries are more common than those that store
starch, however no census has been made.Table 2 compares nectary type with mode ofnectar production. Extrafloral nectaries arealways of the photosynthesizing type.
Table 2. Presence and abundance of starch in nectary parenchyma, duration of secretion in floral (F ) andextrafloral (E ) nectaries of various species. When stored starch is abundant, it is even deposited in theepidermis (ep). Extrafloral nectaries never store starch and are of the photosynthetic type. Floral nectariesproduce nectar by photosynthesis of the nectary or other parts of the flower/plant. Abundance ofamyloplasts in nectary parenchyma is reported by one or more +. Chloroplast presence in the nectaryparenchyma is reported by +
Species andfamily
Nectarytype
Plastid type Duration ofsecretion
Author
Amyloplasts Chloroplasts
Brassica napus(Brassicaceae)
F + Davis et al. 1986
Citrus sinensis(Rutaceae)
F + Rachmilevitz andFahn, 1973
Cucurbita pepo(Cucurbitaceae)
F +++(ep) several hours Nepi et al. 1996
Deuterochonialongipetala(Bromeliaceae)
E + several days/some weeks
Galetto andBernardello 1991
Dyckia floribunda(Bromeliaceae)
E + several days/some weeks
Galetto andBernardello 1991
Euphorbia nerifolia(Euphorbiaceae)
E + several day/some weeks
Arumugasamy et al.1990
Limodorumabortivum(Orchidaceae)
F + Figueiredo andPais 1992
Lonicerajaponica(Caprifoliaceae)
F + Fahn 1979
Musa paradisiaca(Musaceae)
F + Fahn and Benouaiche1979
Passiflora biflora(Passifloraceae)
F +++(ep) several hours Durkee et al. 1981
Ricinus communis(Euphorbiaceae)
E + several days/some weeks
Baker et al. 1978
Rosmarinusofficinalis(Lamiaceae)
F ++ (ep) Zer and Fahn 1992
Sambucus nigra(Caprifoliaceae)
E + several days/some weeks
Fahn 1987
Vicia faba(Fabaceae)
F + Davis et al. 1988
Vinca rosea(Apocynaceae)
F + Rachmilevitz andFahn 1973
E. Pacini et al.: Nectar biodiversity 13

Origin of the amino acid and proteincomponents of nectar
There has been even less research on the originof the amino acid and protein components ofnectar than on the carbohydrate components.It is likely that the site of production of aminoacids in nectar is not always the same and thatit varies in relation to nectary structure. Thismakes it interesting to discover the relationbetween nectar composition and nectary struc-ture.
A useful case is that of Helleborus, in whichthere is some certainty as to where all or mostof the amino acid component is produced. Theepidermal cells of the nectary of Helleborus dieas they produce nectar (Vesprini et al. 1999).Sugars derived from the underlying photosyn-thesizing parenchyma pass through the epider-mal layer and proteins from the degenerating
cytoplasm enrich the nectar with proteins andamino acids (Fig. 1). The emptying of epider-mal cells as they secretes causes changes inposition of all epidermal cells (Fig. 1); thosethat have secreted nectar are displaced anddeformed by the growth of those about tosecrete.
Manner of exposure of nectar
The manner in which nectar is produced andexposed is summarised in Kronestedt-Ro-bards and Robards (1991). Here, we includetheir data together with that of more recentstudies on nectar exposure. Nectar mayappear on the surface of the nectary in fourways.
1. It comes out of modified stomata whichhave lost the capacity to open and close,
of the nectary nectar is produced for a fewhours or several days
• low rate of secretion• the nectar produced is proportionalto mass and volume of nectary• the pollinator requires little nectar• the nectary stops secreting nectarbut photosynthesis continues and itsproducts may be used for otherpurposes
Examples: Brassicaceae,Lamiaceae,Fabaceaesome Scrophulariaceae
Chloroplasts
of other parts ofthe flower orleaves/stem
products of photosynthesisare stored in the nectary
parenchyma
starch is hydrolyzed andnectar is available at any
time of day
• high rate of secretion• large amounts of nectar can beproduced in a short time• pollinators consume large amountsof nectar• the nectary degenerates
Examples:Cucurbita peposome Malvaceae
Fig. 6. Scheme showing the two possible origins of the carbohydrate part of nectar and eco-physiologicalconsequences
14 E. Pacini et al.: Nectar biodiversity

as in Vicia (Davis and Gunning 1992),Cucurbita (Nepi et al. 1996b) (Figs. 1, 2)and Linaria (Nepi et al., this volume).
2. It crosses the plasma membrane then thecell wall and accumulates in a pocketbetween the cell wall and cuticle. The cuticlebreaks under the nectar pressure. Thisoccurs in Anacardium occidentale (Wunna-chit et al. 1992).
3. The cell wall has ingrowths like transfercells that facilitate and speed secretion as inLonicera japonica (Fahn 1979). This mode isfound mainly in pedunculate nectaries suchas those of Abutilon (Kronestedt et al.1986).
4. The nectar flows out through the externalperiphery of the epidermis which has acuticle, as in Limodorum abortivum spur(Figuereido and Pais 1992).
5. The epidermal cells gradually die, producingnectar for a given period (Fig. 1) as inHelleborus (Vesprini et al. 1999).
Clearly the way nectar is exposed andpresented determines the type of nectar col-lector and robbers (Table 1). Case 1 is perhapsthe most common. All parenchyma and epi-dermal cells produce and secrete nectar at thesame time. In cases 2, 3 and 4, parenchymacells operate simultaneously and secretoryepidermal cells are synchronous in the pro-duction/exposure of nectar. In case 5, thesecretory epidermal cells are asynchronous(Fig. 1).
Site and mode of nectar presentationto pollinators
Just as pollen has primary and secondarypresentation (Faegri and van der Pijl 1979), wepropose the same for nectar. Primary presen-tation is exemplified when nectar is exposed topollinators on: a. the surface of the nectaryitself, like pollen presented in the anther; b. oncontinuous surfaces; c. or on the surface ofcapitate nectaries (Fig. 1). Presentation issecondary if the nectar flows from the nectaryand collects in another part of the flower, as in
spurs or similar structures the task of whichseems to be to hold the nectar until it iscollected. Obviously the position, length andaccessibility of the spur determine the type ofcollector and any predators. The spur may bethe nectary itself if it produces nectar as inLimodorum abortivum (Figueiredo and Pais1992) or it may merely be where nectaraccumulates if the nectary is in another partof the flower, as in Linaria (Nepi et al., thisvolume) and the spur is a recipient. InAsclepias syriaca the nectary is in the stigmaticchamber and the nectar flows from there innectar reservoirs (cuculli) (Kevan et al. 1989).
There is an intermediate case in verticallydisposed bell-shaped or tubular flowers andnectaries. In some Aloe species the nectary is atthe base of the corolla and the nectar flowsalong the corolla tube; in Helleborus, thenectary performs photosynthesis and is tubular(Vesprini et al. 1999); the nectar flows alongthe nectary itself (Fig. 1).
Volume and production of nectar in time
The volume and production of nectar in timeare influenced by environmental parameters(Table 1). The quantity of nectar produced bya nectary varies enormously from fractions ofa microlitre (difficult to detect) to thousandsof microlitres (Opler 1983). Abundant pro-duction of nectar may be related to numerouspollinator visits (Waddington 1983), especial-ly in species with many ovules/ovary, or tolarge pollinators. The larger the pollinator,the more the nectar consumed per visit, thelarger the body surface for collecting anddepositing pollen. Repeated visits create suc-cessive waves of pollen tubes in the style(Nepi and Pacini 2001), reducing male andfemale competition and enabling scalar fertil-isation of ovules in the ovary, especially inspecies with many ovules (Nepi and Pacini2001).
The nectar may be produced for a brieftime (only a few minutes or hours) or formany days. When much nectar is produced ina short time, it normally comes from hydro-
E. Pacini et al.: Nectar biodiversity 15

lysis of starch stored in the parenchyma(Fig. 6).
Sexual expression of flowersand nectary morphology
In the case of unisexual flowers, nectaries maybe on both flowers or on only one, as in Osyrisalba (Aronne et al. 1993) where they are onmale flowers and pollinators are induced tovisit female flowers by deception (Table 1). Asimilar situation is found in Ecballium elateri-um where nectaries of male flowers producenectar and those of female flowers do notsecretemeasurableamounts(FahnandShimony2001). In Cucurbita pepo, flowers of both sexeshave nectaries but they are dimorphic in posi-tion and form, causing different patterns ofnectar collection which optimises pollen collec-tion and deposition (Nepi and Pacini 1993).Moreover, the nectar ofmale and female flowersdiffers in quantity and composition inCucurbitapepo and Cocos nucifera (Kevan 1993; Nepi etal. 1996a, 2001). The nectaries of male andfemale flowers are also different in Crotonsarcopetalus (Freitas et al. 2001) and femaleflowers have two types. Septate nectaries ofEuterpe precatoria are only slightly different(Kuchmeister et al. 1997).
Sexual expression of flowersand nectar composition
Nectar composition may differ in male andfemale flowers of monoecious and dioeciousspecies. That produced by female flowers ofCucurbita pepo, Euterpe precatoria and Rhushirta is more abundant and more concentrat-ed than that of male flowers (Greco et al.1996, Kuchmeister et al. 1997, Nepi et al.2001). On the other hand, in the three speciesof the genus Silene, Witt et al. (1999) foundmore concentrated nectar in male flowers. InCucurbita pepo, it has been observed that thedifference in quality and quantity of nectar inthe two flowers is reflected by the fact thatfemale flowers receive more visits (Table 2)(Nepi and Pacini 1993; Nepi et al. 1996a, b).
In hermaphrodite flowers, nectar compositionmay vary in relation to the sexual stage of theflower. For example in Alstroemeria aureaand Echium vulgare, nectar produced in themale phase is more abundant and concentrat-ed (Klinkhamer and de Jong 1990, Aizen andBasilio 1998), whereas in Polyascias sambuci-folia that produced in the female phase ismore concentrated (Gillespie and Henwood1994).
Variability of nectar composition
Besides programmed variability related tosexual expression of the flower, there are othertypes of variability that depend on the envi-ronment and the plant itself (Table 1).
There is variability in flowers of the sameplant, or of flowers of individual plants ofpopulation of the same species, or betweendifferent populations of the same species.These different types of variability probablyat least partly reflect different photosyntheticactivity of the plants or of various parts of aplant, especially in the case of trees. Becausenectar is derived directly or indirectly fromphotosynthesis, its composition may varywidely in relation to exposure of the flower,or parts of the plant carrying flowers, to light,and consequently in relation to temperature,and availability of water. This explains whytemperature can affect nectar secretion andcomposition (Jacobsen and Kistjansson 1994).Freeman and Head (1990) demonstrated that apositive correlation exists between increase intemperature, obviously within limits, and in-crease in nectar concentration. If nectar pro-duction and composition are influenced bytemperature, it follows that nectar qualitywould change in time in species with longblooming times, such as Capparis spinosa(Petanidou et al. 1996) as well as with the ageof the plant, as in Silene dioica (Hemborg1998). Pleasants and Chaplin (1983) demon-strated in Asclepias quadrifolia a positivecorrelation between nectar production androot weight, and a negative one between nectarproduction and inflorescence size.
16 E. Pacini et al.: Nectar biodiversity

Fate of nectar and nectary
Nectar produced by a nectary may be: a. stolenby animals that do not pollinate; b. collectedwholly or partly by animals that pollinate;c. left for lack of pollinators (Table 1). Nectarnot consumed may fall out of the flower ornectary, especially if they are tubular and hangdown (Fig. 1), to the benefit of soil microor-ganisms. In some cases it is resorbed by theplant. The best known examples of nectarresorption are documented in Cucurbita, Lina-ria (Nepi et al. 2001, Nepi et al. this volume),Grevillea (Nicolson 1995) and Eucalyptus(Davis 1997) which differ in aspects such asdifferent origin of nectar carbohydrates, dura-tion of anthesis, site of nectar exposure, and soforth. The corolla closes at the end of anthesisin Cucurbita and in Linaria it is always closedby the labium. In Grevillea and Eucalyptus,nectar droplets remain unprotected duringresorption (Nicolson 1995).
When nectar secretion ceases, the fate ofthe nectary tissue may depends on its positionin the flower and on the origin of the carbo-hydrate part of the nectar. In fact when nectarsugars derive from starch hydrolysis, the nec-tary falls or degenerates because it can not playany other function.
When nectar sugars derive from the pho-tosynthetic activity of the nectary, it may fall,as in the case of Brassicaceae, or persist. Itpersists when the nectary is a part of the ovaryeither under (Linaria vulgaris, Ericaceae etc.)or on the ovary (Hedera helix, Brassicaceae,some Aloe species). In this case the photosyn-thetic activity of the nectary may continue andthe product may contribute to fruit develop-ment. This is only a reasonable hypothesis andresearches are necessary to confirm it.
Energy cost of nectar
Histologically, nectaries consist of secretorytissue and nectar is exposed to rewardseeking pollinators or animals that defendthe plant. Nectar is therefore an investmentthat may be costly in energy terms. The cost
of nectaries can be up to 37% of dailyphotosynthesis (Southwick 1984, Pyke 1991).This may be why plants that produce abun-dant nectar resorb what is left over (Nicolson1995, Nepi et al. 2001, Nepi et al. thisvolume).
There is also another case of nectar saving.Cleistogamy is a temporary phenomenon inmany plants which have flowers that do notopen and automatically self-pollinate. Cleisto-gamic flowers are always smaller than thosecapable of cross pollination (Lord 1981). Theadvantage of cleistogamy is that it producesseeds with less energy investment, albeit withreduced variability. Some species subject totemporary cleistogamy are anemophilous,though there are many entomophilous (Lord1981), some of which have nectaries. At least insome species of the genus Viola (V. canina,V. hirta and V. odorata) cleistogamic flowersare devoid of nectaries (our unpublished data).However, we do not know to what extent thisfeature occurs among species manifesting cleis-togamy.
The lack of nectaries in cleistogamic flow-ers of the genus Viola is an example ofcoordination of plant activity towards energysaving. The case of Pittosporum tobira isdifferent: this plant is male sterile meaningthat the anthers do not produce pollen, but it isapomictic because fruits ripen, dispersing seedswhich germinate. Despite this, the plant hasnectaries and bees visit the flowers and collectnectar (Ricciardelli D’Arbore and PersanoOddo 1978), to no apparent benefit of theplant.
Future research
Existing knowledge on nectaries can be definedas abundant but not integrated or rationalised.Cytological aspects have been related rarely tophysiological and ecological aspects. Apartfrom reviews, papers presenting the results ofresearch usually regard a single species, whichmeans comparisons of different types of nec-tary is rare. Certain cytological and ecologicalaspects have never been studied (e.g. the
E. Pacini et al.: Nectar biodiversity 17

presence of cuticle and the nectariferousparenchyma type) and because laboratoryand field study techniques are not standar-dised, certain data cannot be comparedbetween species.
Apart from the fundamental contributionof Bonnier (1879) and Vogel (1997, 1998a, b,c), there is no systematic view of the variouscytological and ecological aspects of nectariesin angiosperm families.
In our opinion, there are still gaps in ourknowledge of the origin of the polysaccharidecomponent of nectar. Only a few reportsdemonstrate an effect of environmental andexperimental conditions on nectar compositionand abundance (Corbet et al. 1979, Wyatt et al.1992). Vickery and Sutherland (1994) analysedthe volume and concentration of nectar inspontaneous and cultivated plants and foundthat concentration changes little whereas vol-ume changes considerably, especially in green-house plants.
There have been various studies on nectarcomposition, mostly on soluble monosaccha-rides and oligosaccharides, but not on thepresence of PAS-positive polysaccharides thatsurvive fixation, like those observed in Cucur-bita pepo (Fig. 2). No explanation has beenoffered for why nectar, a sugar solution, is notremoved by fixation and dehydration and alsoshows PAS positivity indicative of polysaccha-rides. All this suggests that nectar containsunknown dissolved polysaccharides.
Little work has been done into differencesbetween nectaries and nectar composition inplants with wide distributions, for example theecological studies on Pteridium aquilinum(Heads and Lawton 1985, Raschbrook et al.1992).
Although it is fairly clear where the sugarcomponent of nectar comes from, the origin ofthe amino acid-protein and other componentshas received less attention. Research on thistopic would be useful.
It would also be useful to know if ourobservations on the lack of nectaries in cleis-togamic flowers of Viola are a feature of allcleistogamic plants with nectaries. Other
aspects that deserve attention are the relationsbetween nectary types and breeding systemse.g. dioecy, monoecy, autogamy, xenogamyand parthenocarpy.
References
Aizen M. A., Basilio A. (1998) Sex differentialnectar secretion in protandrous Alstremeriaaurea (Alstremeriaceae); is production alteredby pollen removal and receipt? Amer. J. Bot. 85:245–252.
Aronne G., Wilcock C. C., Pizzolongo P. (1993)Pollination biology and sexual differentiation ofOsyris alba (Santalaceae) in the Mediterraneanregion. Plant Syst. Evol. 188: 1–16.
Arumugasamy K., Subramanian R. B., Inamdar J.A. (1990) Cyathial nectaries of Euphorbia nerii-folia L.: ultrastructure and secretion. Phytomor-phology. 40: 281–288.
Baker H. G., Baker I. (1983) Floral nectar sugarconstituents in relation to pollinator type. In:Jones C. E., Little R. J. (eds.) Handbook ofexperimental pollination biology. Science andAcademic Editions, New York, pp. 117–141.
Baker D. A., Hall J. L., Thorpe J. R. (1978) Astudy of extrafloral nectaries of Ricinus commu-nis. New Phytol. 81: 129–137.
Bino R. J., Devente N., Meeuse A. D. J. (1984)Entomophily in the dioecious gymnosperm Ep-hedra aphylla Forsk. (= E. Alte C. A. Mey.),with some notes on E. campylopoda C. A. Mey.II Pollination droplets, nectaries, and nectarialsecretion in Ephedra. Proceedings of theKoninklijke Nederlandse Akademie vanWetenschappen, Series C. 87: 15–24.
Bonnier G. (1879) Les Nectaires. Ann. Sci. Nat.Bot. 8: 1–212.
Corbet S. A., Willmer P. G., Beament J. W. L.,Unwin D. M., Prys-Jones O. E. (1979) Post-secretory determinants of sugar concentration innectar. Plant Cell Environ. 2: 293–308.
Davis A. R. (1997) Influence of floral visitation onnectar-sugar composition and nectary surfacechanges in Eucalyptus. Apidologie 28: 27–42.
Davis A. R., Gunning B. E. S. (1992) The modifiedstomata of the floral nectary of Vicia faba L.1. Development, anatomy and ultrastructure.Protoplasma 166: 134–152.
Davis A. R., Peterson R. L., Shuel R. W. (1986)Anatomy and vasculature of the floral nectaries
18 E. Pacini et al.: Nectar biodiversity

of Brassica napus (Brassicaceae) Can. J. Bot. 64:2508–2516.
Davis A. R., Peterson R. L., Shuel R. W. (1988)Vasculature and ultrastructure of the floral andstipular nectaries of Vicia faba (Leguminosae)Can. J. Bot. 66: 1435–1448.
Durkee L. T., Gaal D. J., Reisner W. H. (1981)The floral and extrafloral nectaries of Passiflora.I. The floral nectary. Amer. J. Bot. 68: 453–462.
Durkee L. T. (1982) The floral and extra-floralnectaries of Passiflora. II. The extra-floral nec-tary. Amer. J. Bot. 69 (9): 1420–1428.
Faegri K., van der Pijl L. (1979) The principles ofpollination ecology. Pergamon Press, Oxford.
Fahn A. (1952) On the structure of floral nectaries.Bot. Gaz. 113: 464–470.
Fahn A. (1979) Secretory tissues in plants.Academic Press, London.
Fahn A. (1987) The extrafloral nectaries ofSambucus. Ann. Bot. 60: 299–308.
Fahn A. (2000) Structure and function of secretorycells. Advanc. Bot. Res. 31: 37–75.
Fahn A., Benouaiche P. (1979) Ultrastructure,development and secretion in the nectary ofbanana flowers. Ann. Bot. 44: 85–93.
Fahn A., Shimony C. (2001) Nectary structure andultrastructure of unisexual flowers of Ecballiumelaterium (L.) A. Rich. Cucurbitaceae and theirpresumptive pollinators. Ann. Bot. 87: 27–33.
Figueiredo A. C., Pais M. S. (1992) Ultrastructuralaspects of the nectary spur in Limodorum abort-ivum (L.) Sw. (Orchidaceae). Ann. Bot. 70: 325–331.
Freitas L., Bernardello G., Galetto L., PaoliA. A. S. (2001) Nectaries and reproductivebiology of Croton sarcopetalus (Euphorbiaceae).Bot. J. Linn. Soc. 136: 267–277.
Freeman C. E., Head K. C. (1990) Temperatureand sucrose composition of floral nectar inIpomopsis longiflora under field conditions.Southwest. Nat. 35: 423–426.
Friis E. M., Endress P. K. (1990) Origin andevolution of angiosperm flowers. Advan. Bot.Res. 17: 99–62.
Gaffal K. P., Heimler W., El-Gammal S. (1998)The floral nectary of Digitalis purpurea L.,structure and nectar secretion. Ann. Bot. 81:251–262.
Galetto L., Bernardello L. (1992) Nectar secretionpattern and removal effects in six ArgentineanPitcairnioideae. Bot. Acta 105: 292–299.
Gillespie L. H., Henwood M. J. (1994) Temporalchanges of floral nectar-sugar composition inPolyscias sambucifolia (Sieb. ex DC.) Harms(Araliaceae). Ann. Bot. 74: 227–231.
Greco C. F., Holland D., Kevan P. G. (1996)Foraging behaviour of honey bees (Apis mellif-era L.) on staghorn sumac [Rhus hirta Sudworth(ex-Typhina L.)]: differences and dioecy. Can.Entomol. 128: 355–366.
Heads P. A., Lawton J. H. (1985) Bracken, antsand extrafloral nectaries. III. How insect herbi-vores avoid ant predation. Ecol. Ecomol. 10:29–42.
Hemborg M. (1998) Seasonal dynamics in repro-duction of first-year females and males in Silenedioica. Int. J. Plant Sci. 159: 958–967.
Inouye D. W. (1983) The ecology of nectarrobbing. In: Bentley B., Elias T. (eds.) Thebiology of nectaries. Columbia University Press,New York, pp. 153–173.
Jacobsen H. B., Kistjansson K. (1994) Influence oftemperature and floret age on nectar secretion inTrifolium repens L. Ann. Bot. 74: 327–334.
Kenrick J., Bernardt P., Marginson R., BeresfordG., Knox R. B., Baker I., Baker H. G. (1987)Pollination-related characteristics in the mimo-soid legume Acacia terminalis (Leguminosae).Plant Syst. Evol. 157: 49–62.
Kevan P. G. (1993) Wind or insects: pollination ofcoconut (Cocos nucifera) in the Maldive Islands.In: Connor L. J., Rinderer T., Sylvester H. A.,Wongsiri S. (eds.) Asian apiculture. WicwasPress, Cheshire (Connecticut), pp. 372–377.
Kevan P.G., EisikowitchD., Rathwell B. (1989) Therole of nectar in the germination of pollenin Asclepias syriaca L. Bot. Gaz. 150: 266–270.
Klinkhamer P. G. L., de Jong T. J. (1990) Effect ofplant size, plant density and sex differentialnectar reward on pollinator visitation in theprotandrous Echium vulgare (Boraginaceae).Oikos 57: 399–405.
Kronestedt E., Robards A. W., Stark M., Olesen P.(1986) Development of trichomes in the Abutilonnectary gland. Nord. J. Bot. 6: 627–639.
Kronestedt-Robards E., Robards A. W. (1991)Exocytosis in gland cells. In: Hawes C. R.,Coleman J. O. D., Evans D. E. (eds.) Endocy-tosis, exocytosis and vescicle traffic in plants.Cambridge University Press, Cambridge.
Kuchmeister H., Silberbauer-Gottsberger, Gotts-berger G. (1997) Flowering, pollination,
E. Pacini et al.: Nectar biodiversity 19

nectar standing crop, and nectaries of Euterpeprecatoria (Arecaceae) an Amazonian rain forestpalm. Plant Syst. Evol. 206: 71–97.
Lord E. (1981) Cleistogamy: a tool for the study offloral morphogenesis, function and evolution.Bot. Rev. 47: 421–449.
Maldonado S., Otegui M. ( 1997) Secretory tissuesof the flower of Sanango racemosum (Gesneria-ceae). I. Light microscopy. Acta Bot Neerl. 46:413–420.
Mani M. S., Saravanan J. M. (1999) Pollinationecology and evolution in Compositae (Astera-ceae). Science Publishers, USA.
Neiland M. R., Wilcock C. C. (1998) Fruit set,nectar reward, and rarity in the Orchidaceae.Am. J. Bot. 85: 1657–1671.
Nepi M., Pacini E. (1993) Pollination, pollenviability and pistil receptivity in Cucurbita pepo.Ann. Bot. 72: 526–536.
Nepi M., Pacini E. (2001) Effect of pistil age onpollen tube growth, fruit development and seedset in Cucurbita pepo L. Acta Soc. Bot. Pol. 70:165–172.
Nepi M., Pacini E., Willemse M. T. M. (1996a)Nectary biology of Cucurbita pepo: ecophysio-logical aspects. Acta Bot. Neerl. 45: 41–54.
Nepi M., Ciampolini E., Pacini E. (1996b)Development and ultrastructure of Cucurbitapepo nectaries of male flowers. Ann. Bot. 78: 95–104.
Nepi M., Guarnieri M., Pacini E. (2001) Nectarsecretion, reabsorption and sugar composition inmale and female flowers of Cucurbita pepo. Int.J. Plant Sci. 162: 353–358.
Nicolson S. W. (1995) Direct demonstration ofnectar reabsorption in the flowers of Grevillearobusta (Proteaceae). Funct. Ecol. 9: 584–588.
Opler P. A. (1983) Nectar production in a tropicalecosystem. In: Bentley B., Elias T. (eds.) Thebiology of nectaries. Columbia University Press,New York, pp. 30–77.
Percival M. (1961) Types of nectar in Angiosperms.New Phytol. 60: 235–281.
Pleasants J. M., Chaplin S. J. (1983) Nectar pro-duction rates of Asclepias quadrifolia: causes andconsequences of individual variation. Oecologia59: 232–238.
Pyke G. H. (1991) What does it cost a plant toproduce floral nectar? Nature 350: 58–59.
Rachmilevitz T., Fahn A. (1973) Ultrastructure ofnectaries of Vinca rosea L., Vinca major L. and
Citrus sinensis Osbeck cv. Valencia and itsrelation to the mechanism of nectar secretion.Ann. Bot. 37: 1–9.
Rashbrook V. K., Compton S. G., Lawton J. H.(1992) Ant-herbivore interactions: reason for theabsence of benefits to a fern with foliar nectaries.Ecology 73: 2167–2174.
Razem F. A., Davis A. R. (1999) Anatomical andultrastructural changes of floral nectary of Pisumsativum L. during flower development. Proto-plasma 206: 57–72.
Ricciardelli D’Arbore G., Persano Oddo L. (1978)Flora Apistica Italiana. Istituto Sperimentale perla Zoologia Agraria, Firenze.
Sawidis T. (1998) The subglandular tissue of Hibis-cus rosa-sinensis nectaries. Flora 193: 327–335.
Schmid R. (1988) Reproductive versus extra-repro-ductive nectaries: histological perspective andterminological recommendations. Bot. Rev. 54:179-252.
Smets E. F., Ronse Decraene L-P, Caris P., RudallP. J. (2000) Floral nectaries in Monocotyledons:distribution and evolution. In: Wilson K. L.,Morrison K. L. (eds.) Monocots: systematicsand evolution. CSIRO, Melbourne, pp. 230–240.
Southwick E. E. (1984) Photosynthate allocation tofloral nectar: a neglected energy investment.Ecology 65: 1775–1779.
Vesprini J. L., Nepi M., Pacini E. (1999) Nectarystructure, nectar secretion patterns and nectarcomposition in two Helleborus species. PlantBiol. 1: 560–568.
Vickery R. K., Sutherland S. D. (1994) Varianceand replenishment of nectar in wild and green-house populations of Mimulus. Great BasinNaturalist 54: 212–227.
Vogel S. (1997) Remarkable nectaries: structure,ecology, organophyletic perspectives. I Substitu-tive nectaries. Flora 192: 305–333.
Vogel S. (1998a) Remarkable nectaries: structure,ecology, organophyletic perspectives. II. Necta-rioles. Flora 193: 1–29.
Vogel S. (1998b) Remarkable nectaries: structure,ecology, organophyletic perspectives. III. Nectarducts. Flora 193: 113–131.
Vogel S. (1998c) Remarkable nectaries: structure,ecology, organophyletic perspectives. IV. Mis-cellaneous cases. Flora 193: 225–248.
Wheatley J. M. (1977) Variations in the basicpathway of chloroplast development. NewPhytol. 78: 407–420.
20 E. Pacini et al.: Nectar biodiversity

Waddington K. D. (1983) Floral visitationsequences by bees: models and experiments. In:Jones C. E., Little R. J. (eds.) Handbook ofexperimental pollination biology. Scientific andAcademic Editions, New York, pp. 461–473.
Wetschnig W., Depisch B. (1999) Pollination biol-ogy of Welwitschia mirabilis Hook. F. (Welwit-schiaceae, Gnetopsida) Phyton (Horn, Austria)39: 167–188.
Wyatt R., Broyles S. B., Derda G. S. (1992)Environmental influences on nectar productionin milkweeds (Asclepia syriaca and A. exaltata).Amer. J. Bot. 79: 636–642.
Witt T., Jurgens A., Geyer R., Gottsberger G.(1999) Nectar dynamics and sugar compositionin flowers of Silene and Saponaria species(Caryophyllaceae). Plant Biolol. 1: 334–345.
Wunnachit W., Jenner C. F., Sedgley M. (1992)Floral and extrafloral nectar production inAnacardium occidentale L. (Anacardiaceae): anandromonoecious species. Int. J. Plant Sci. 153:413–420.
Zer H., Fahn A. (1992) Floral nectaries ofRosmarinus officinalis L.: structure, ultra-structure and nectar secretion. Ann. Bot. 70:391–397.
Addresses of the authors: E. Pacini (e-mail:[email protected]), M. Nepi, Department of Envi-ronmental Sciences ‘‘G. Sarfatti’’, Via P. A. Matt-ioli 4, I-53100 Siena, Italy. J. L. Versprini, Facultyof Agricultural Sciences, UNR CC14 2123 Zavalla,Argentina.
E. Pacini et al.: Nectar biodiversity 21