mycophenolate mofetil and roscovitine decrease cyclin expression and increase p27kip1 expression in...
TRANSCRIPT

Clinical and Experimental Immunology
© 2004 British Society for Immunology,
Clinical and Experimental Immunology
,
139:
225–235
225
doi:10.1111/j.1365-2249.2004.02684.x
et al.
Accepted for publication 25 October 2004
Correspondence: Dr Elisa Menegatti, Diparti-
mento di Medicina e Oncologia Sperimentale,
Università di Torino, Corso Raffaello 30, 10125
Torino, Italy.
E-mail: [email protected]
OR IG INAL ART I C L E
Mycophenolate mofetil and roscovitine decrease cyclin expression and increase p
27
kip
1
expression in anti Thy
1
mesangial proliferative nephritis
M. Chiara,*
†
E. Menegatti,*
†
D. Di Simone,*
†
A. Davit,*
†
D. Bellis,
‡
D. Sferch,
‡
G. De Rosa,
‡
O. Giachino,*
†
L. M. Sena*
†
and D. Roccatello*
†
*Cattedre e Scuola di Specializzazione di Patologia
Clinica, Università di Torino,
†
Centro
Multidisciplinare di Immunopatologia e
Documentazione su Malattie Rare, CMID, Osp.
G. Bosco (ASL4) and
‡
Divisione di Anatomia
Patologica, ASL4, Torino, Italy
Summary
The response of mesangial cells to a phlogistic challenge includes cell prolif-eration and mesangial matrix expansion. Cell proliferation is a highly regu-lated process which includes enhancing factors such as cyclins, cyclindependent kinases, and inhibitory proteins, such as p27
kip1
. The aim of thestudy was to evaluate the effects of Mycophenolate mofetil (MMF), and ros-covitine (R), on the cell cycle regulatory system when administered in theflorid phase of the experimental model of mesangial proliferative nephritisinduced by the anti Thy-1 antigen monoclonal antibody. Three days afternephritis induction, different groups were given MMF and R. Rats treatedwith MMF or R showed a slight decrease in mesangial proliferation andmatrix expansion. Samples of cortical tissue were tested by ‘real time’ RT-PCRin order to study gene expression of cyclins B, D1, D2, D3, E, and the cyclininhibitor p27
kip1
. Localization of mRNA was evaluated by
in situ
hybridiza-tion. Real time RT-PCR analysis showed a significant decrease in cyclins B,D1, D2, and D3 in rats treated with either MMF or R as compared to controls.Both MMF and R treatment induced a significant increase in p27
kip1
mRNAexpression.
In situ
hybridization showed a mesangial-endothelial expressionpattern in glomeruli. The number of labelled cells per glomerulus, the num-ber of positive glomeruli in each examined slide as well as cyclin D2 and D3signal intensity was significantly lower in rats treated with MMF or R as com-pared to controls, whereas MMF or R treatment up-regulated p27
kip1
mRNAexpression. Immunohistochemical evaluation of p27
kip1
aimed to examine theinfluence of MMF or R on protein expression confirmed up-regulation.
Keywords:
mycophenolate mofetil, cyclin, p27
kip1
, mesangial proliferation, antiThy1 model
IIII
ntroduction
Glomerular cell hypertrophy occurs in many forms of acuteand chronic disease and may precede the development ofglomerulosclerosis [1,2]. One of the key features of severalforms of glomerulonephritis is massive proliferation ofglomerular mesangial cells and deposition of mesangialmatrix in response to a phlogistic challenge [1,2]. Cell pro-liferation is dependent on the co-ordinated activation of aseries of cell cycle regulatory genes.
Cyclins and cyclin-dependent kinases (CDKs) representthe main positive regulatory proteins, while CDK-inhibi-tors (CKI) are their negative counterpart. Cyclins and theirpartner CDKs form an active holoenzyme, which is spe-
cific for each phase of the cell cycle. These complexes phos-phorilate substrates which are critical for cell division.Cyclins have a very short half-life (30–60 min) and theirlevels fluctuate in each phase of the cell cycle [3]. They aretranscriptionally and post-translationally regulated [4].Entry into cell cycle from quiescent state is governed by
d
-type cyclins (namely D1, D2, D3). They form an activecomplex with their specific CDKs, that are responsiblefor phosphorilation and functional inactivation of the ret-inoblastoma protein, an event that implies DNA synthesis[5].
Cyclin E is expressed in the late G1 phase and is essential inorder to overcome the G1/S restriction point. Cyclin A ismaximally expressed during the S-phase since it is involved

M. Chiara
et al.
226
© 2004 British Society for Immunology,
Clinical and Experimental Immunology
,
139:
225–235
in DNA-synthesis and G2/M transition. Cyclin B governs celldivision during the M-phase [6].
The cell cycle is also regulated by inhibitory proteins,including the p21 and the p27 family, which inhibit allcyclin-CDKs complexes and by the so-called INK4 family(p15, p16, p18, p19), which in turn specifically inhibits
d
-type cyclin-CDKs complexes [7].
The anti Thy-1 experimental nephritis is a well establishedrat model that is characterized by a marked proliferation ofmesangial cells [8]. This is an acute and self-limiting diseasein which mesangial proliferation represents the reparativeresponse to massive mesangiolysis. Thus, anti Thy-1 nephri-tis is different from human mesangial glomerulonephritis,in which, mesangial hyperplasia is followed by glomerularsclerosis. Nevertheless, this experimental model offers theunique opportunity to evaluate possible interferences ofnovel pharmacological compounds on mesangial prolifera-tion, although no estimation of the effects of these com-pounds in the long term is possible. In this model, the onsetof mesangial cell proliferation is associated with an increasein the number of cyclin D1 and cyclin A-positive mesangialcells, and an increase in expression and activity of CDK2.While levels of p27, the CDK inhibitor, decrease, there is anup-regulation of p21 which is induced in the disease state[9,10]. Nephritic rats given roscovitine, a purine analoguewhich is known to inhibit
in vitro
CDK activity withoutaffecting cyclin or CDK levels [11], were found to have amarked reduction in mesangial cell proliferation and a def-inite amelioration in renal function compared to diseased,untreated animals [12]. Other compounds known to inter-fere with the development of renal changes in this modelinclude retinoids. Retinoids directly modulate growth fac-tors and vasoactive substances (such as endothelin 1) andinterfere with proliferative transcription factors (includingcyclin D
1
) [13].Mycophenolate mofetil (MMF) is a powerful immunosup-
pressive agent used extensively for the prevention of acuteallograft rejection [14]. The immunosuppressive activity ofMMF mainly derives from the inhibition of inosine mono-phosphate dehydrogenase (IMPDH) [15]. Other pharmaco-logical activities of MMF include functional impairment ofadhesion molecules and – with yet unknown mechanisms –inhibition of proliferation of smooth muscle cells in culture[16] and mesangial cells both
in vitro
and
in vivo
[17].In the present study the effects of MMF and roscovitine on
cell cycle regulatory system were investigated in the experi-mental model of anti Thy-1 monoclonal antibody-inducedproliferative nephritis.
Materials and methods
Experimental design
Wistar (Harlan Nossan – Italy) male rats of 200–220 g bodyweight were maintained in standard conditions of light
and dark (light times 0800h
-
2000h), and temperature(21
±
1
∞
C) and given food (standard rat chow with 25% ofprotein) and water ad libitum. The animals were weighedboth before and after each treatment and during the sacrificeand no significant differencies between treated and controlgroup were observed. In each experimental condition, bloodand 24 hour urines were collected and immediately analysedfor determination of proteinuria, and haematuria (SelfStick – IPR Italy) and creatinine clearance.
Treated rats were given a single dose of 0·25 ml/100 gbody weight of anti-Thy-1 monoclonal antibody (OX 7,CEDARLANE, Ontario, Canada), through an intravenousinjection in the tail, according to the Thy-1 model of mesan-gial proliferative glomerulonephritis [18]. Seven groups offive rats each were studied. Six groups of 5 rats receivedmonoclonal antibody. Two groups, which were sacrificedone day and eight days after the induction of nephritis,respectively, served as positive controls (CD1
+
and CD8
+
).From day 3 to day 7 a group (R) received roscovitine (AlexisCo., Lanser, Switzerland), a purine analogue, at the dose of2·12 mg/kg of body weight in 500
m
l of DMSO/die by intra-peritoneum injection, a group (MMF) received
per os
myco-phenolate mofetil (a gift from Roche, Basel, Switzerland) atthe dose of 40 mg/kg of body weight in 400
m
l of bufferedsaline/die. Rats given only the vehicle DMSO (group D)(500
m
l/die) or buffered saline (group F) (400
m
l/die) servedas controls. On days 1, 3 and 7 the animals were separatedand placed in metabolic cages for 24 h for urine collection.
At day 8 all the animals were sacrificed as were healthyrats, given intravenously a sterile saline solution (NaCl 0·9%)(negative control C-).
Both kidneys were withdrawn, weighed and carefullyexamined. The cortical tissue of one kidney was separatedfrom the medulla, and the two portions were immediatelyfrozen in liquid nitrogen and preserved at
-
80
∞
C, for molec-ular biology analyses. The contra lateral kidney tissue wasfixed in buffered formalin at 4
∞
C and processed for histolog-ical analysis and immunohistochemistry.
Histological analysis
Thick sections of all organs were fixed in cold (4
∞
C) 10%buffered formalin for 24 h, and then embedded in paraffinusing routine techniques. An average of 10, 2–3 mm thick,sections were prepared for microscopic analysis. Observa-tions were performed in double blind by two pathologists(G.D and D.B).
Haematoxylyn-eosine, periodic acid-Schiff reagent, andMasson’s staining were used. About 50 glomeruli per sec-tions were examined and assessed for total cellularity, type ofcellular proliferation, detection of inflammatory infiltrates,presence and percentage of apoptotic cells, glomerular base-ment membrane thickness, mesangial expansion, disruptionof the urinary space and Bowman capsule changes.

MMF and cyclins in the anti Thy-1 model
© 2004 British Society for Immunology,
Clinical and Experimental Immunology
,
139:
225–235
227
Morphometric evaluation was performed employing thesemi quantitative method described in Tisher [19], i.e. num-ber of nuclei in the mesangial area per glomerulus.
Gene expression study
RT-real time PCR
Cortical tissue and medulla were processed separately formRNA extraction by RNAcqueous System (Ambion). TotalRNA was treated with DNAse and quantified by fluorimetricdetection (RiboGreen RNA quantification kit, MolecularProbe). Two nanograms of total RNA were reverse tran-scribed into cDNA using random examers as primers.
Relative gene expression quantification of cyclin A, B, D1,D2, D3 and E, and cyclin inhibitor p27 gene expression wasassessed by means of real time PCR (ABI Prism 7700,Applied Biosystems, Foster City, CA, USA) usingSYBR®Green as the detection method.
Primer sequences (Table 1) were chosen by using PrimerExpress software (Applied Biosystems), and optimization ofamplification reaction was assured by dissociation curveanalysis. Data were normalized using glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and were analysed bythe comparative Ct method. In all experiments, only tripli-cates with a standard deviation of the Ct value
<
0·20 wereaccepted.
In situ
hybridization
Cyclin D2, D3 and p27 cyclin kinase inhibitor specificmRNAs were detected by using digoxigenin-labelled ribo-probes that were synthesized by using specific PCR productscloned into pCR®4-TOPO plasmid (TOPO-TA Cloning kit,Invitrogen) as template. In positive clones the insert waschecked by direct sequencing.
Riboprobes were labelled with dig-dUTP (DIG labellingmix, Roche) by an in vitro RNA transcription using T3 or T7RNA polymerases, depending on insert position.
Localization of mRNA was determined on 3 micrometerthick sections of formalin-fixed and paraffin embedded tis-sue. The sections were rehydrated in a graded series of alco-hol and blocked in 10% FCS.
Tissue sections were permeabilized by treatment in PBS-glycine 0·1
m
buffer, PBS-TritonX 100 0·3%, and proteinaseK (1
m
g/ml), incubated in prehybridization buffer (40% for-mamide, 50% dextran sulphate, 10
¥
Denhart’s, 0·1
m
DTT,0·5 ng/ml yeast t-RNA) for 3 h and then incubated overnightat 42
∞
C with 100 ng of labelled probe (2 ng/
m
l in 50
m
l/sec-tion). Sections were washed in 5
¥
SSC buffer at 37
∞
C for 1 h,in 0·2
¥
SSC at 37
∞
C for 1 h and in 0·2
¥
SSC at room tem-perature for 5 min. Probe signals were then visualized byimmunoreactions using an antidigoxigenin antibodylabelled with alkaline phosphatase and using NBT/BCIP aschromogen. Labelling was independently evaluated in ablind fashion in controls and in Thy-1 glomerulonephritisrats by three investigators (EM, DB and GD). Each rat (atleast 4 per treatment) was individually assessed. Fifty glom-eruli for 2 cross section were evaluated by counting thenumber of nuclei that stained positive. Results representedthe mean of the independent evaluation of the threeresearchers.
Immunohistochemistry
p27 expression was analysed by immunohistochemistry on3
m
m sections of formalin-fixed and paraffin embedded tis-sue. The sections were rehydrated in a graded series of alco-hol and blocked in 10% horse serum. The endogenousperoxidases were inactivated by treating slices with an 0·3%hydrogen peroxide/methanol solution. Antigen unmaskingwas performed by 3 passages of sodium citrate buffer(10 mmol/l pH 6·0) dipped slices in a microwave oven. Sec-tions were then incubated overnight at 4
∞
C with polyclonalantibody to p27 (C-19) (1 : 500 Santa Cruz Biotechnology,Santa Cruz, CA, USA), and goat anti rabbit IgG conjugatedwith horseradish peroxidase (HRP, BIO-RAD, Hercules, CA,USA), was used as the secondary antibody (1 : 200) for30 min. Sections were incubated for 30 min with avidin-biotin complex and developed with diaminobenzidine(DAB) as chromogen. p27 expression was graded semi quan-titatively as in ‘
in situ
hybridization’ experiments.
Statistical analyses
Results are expressed as mean
±
SD. Statistical analysis wasperformed by using commercial software (Stat View, AbacusConcepts Inc, Berkeley, CA, USA). Comparisons of meanswere carried out by the Student’s
t
-test. For each experimenta
P
-value
<
0·05 was considered statistically significant.
Table 1.
Primer sequence used for real time PCR.
GAPDH FW 5
¢
-CCTGATGCAAACACCCAACTT-3
¢
REV 5
¢
-TCTGTGGGGCCTTGAAGAATATAG-3
¢
CB FW 5
¢
-CTCTCCAAGCCCAATGGAAA-3
¢
REV 5
¢
-TTTCTCCGATGTGATTCTTGCA-3
¢
CD1 FW 5
¢
-CCCACGATTTCATCGAACACT-3
¢
REV 5
¢
-GATCATCCGCAAACATGCA-3
¢
CD2 FW 5
¢
-AAGGACATCCAGCCGTACATG-3
¢
REV 5
¢
-TGTGAGGGACAGAAGTGTGAAGA-3
¢
CD3 FW 5
¢
-TGGATGCTGGAGGTGTGTGA-3
¢
REV 5
¢
-AAAGGCGCAACTGCAGCTT-3
¢
CE FW 5
¢
-TGGATGTTGGCTGCTTAGAATTT-3
¢
REV 5
¢
-TTTCCTCGTTGGAGTTGATGC-3
¢
P27 KIP1 FW 5
¢
-CGGCTGGGTTAGCGGAG-3
¢
REV 5
¢
-GATTCTTCTTCGCAAAACAAAAGG-3
¢
M. Chiara
et al.
228
© 2004 British Society for Immunology,
Clinical and Experimental Immunology
,
139:
225–235
Results
Histological analysis
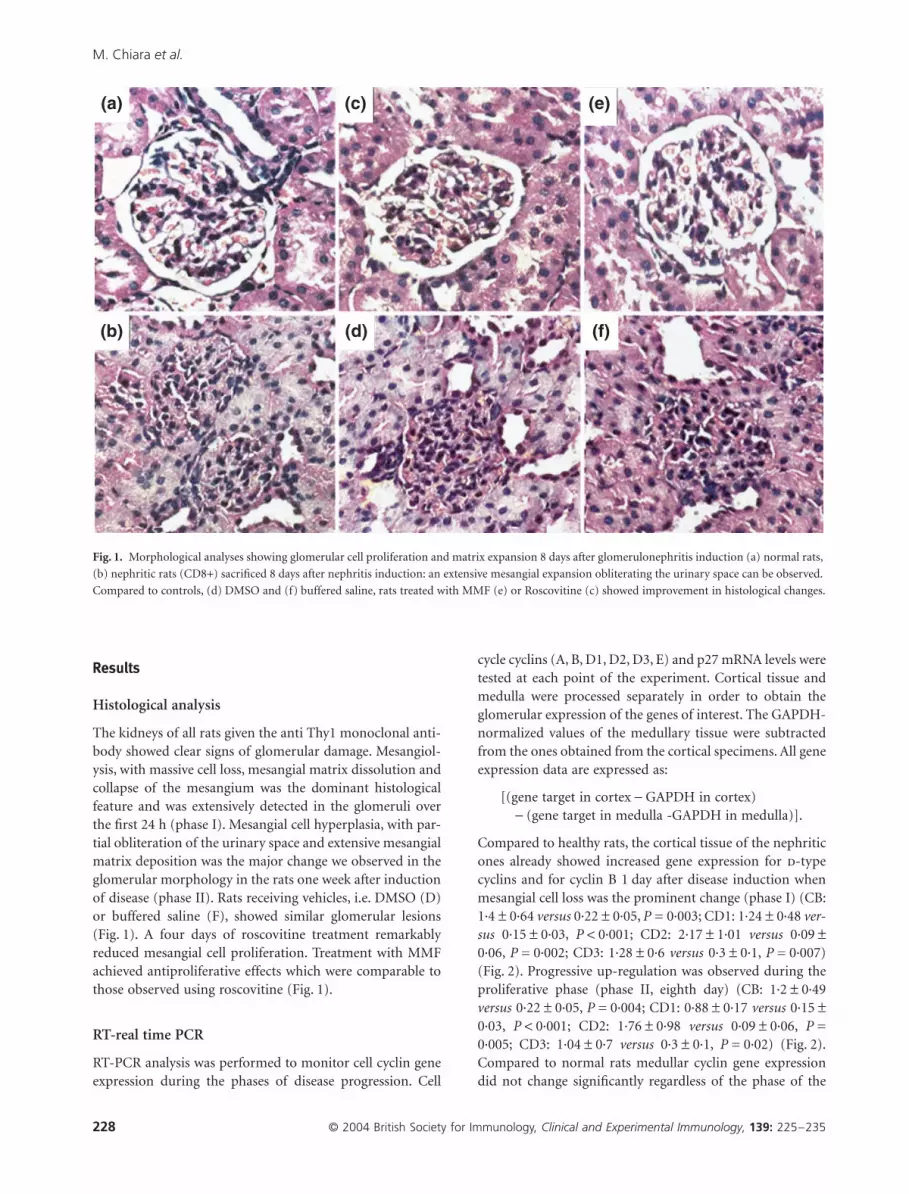
The kidneys of all rats given the anti Thy1 monoclonal anti-body showed clear signs of glomerular damage. Mesangiol-ysis, with massive cell loss, mesangial matrix dissolution andcollapse of the mesangium was the dominant histologicalfeature and was extensively detected in the glomeruli overthe first 24 h (phase I). Mesangial cell hyperplasia, with par-tial obliteration of the urinary space and extensive mesangialmatrix deposition was the major change we observed in theglomerular morphology in the rats one week after inductionof disease (phase II). Rats receiving vehicles, i.e. DMSO (D)or buffered saline (F), showed similar glomerular lesions(Fig. 1). A four days of roscovitine treatment remarkablyreduced mesangial cell proliferation. Treatment with MMFachieved antiproliferative effects which were comparable tothose observed using roscovitine (Fig. 1).
RT-real time PCR
RT-PCR analysis was performed to monitor cell cyclin geneexpression during the phases of disease progression. Cell
cycle cyclins (A, B, D1, D2, D3, E) and p27 mRNA levels weretested at each point of the experiment. Cortical tissue andmedulla were processed separately in order to obtain theglomerular expression of the genes of interest. The GAPDH-normalized values of the medullary tissue were subtractedfrom the ones obtained from the cortical specimens. All geneexpression data are expressed as:
[(gene target in cortex
-
GAPDH in cortex)
-
(gene target in medulla -GAPDH in medulla)].
Compared to healthy rats, the cortical tissue of the nephriticones already showed increased gene expression for
d
-typecyclins and for cyclin B 1 day after disease induction whenmesangial cell loss was the prominent change (phase I) (CB:1·4
±
0·64
versus
0·22
±
0·05,
P
= 0·003; CD1: 1·24
±
0·48
ver-sus
0·15 ± 0·03, P < 0·001; CD2: 2·17 ± 1·01 versus 0·09 ±0·06, P = 0·002; CD3: 1·28 ± 0·6 versus 0·3 ± 0·1, P = 0·007)(Fig. 2). Progressive up-regulation was observed during theproliferative phase (phase II, eighth day) (CB: 1·2 ± 0·49versus 0·22 ± 0·05, P = 0·004; CD1: 0·88 ± 0·17 versus 0·15 ±0·03, P < 0·001; CD2: 1·76 ± 0·98 versus 0·09 ± 0·06, P =0·005; CD3: 1·04 ± 0·7 versus 0·3 ± 0·1, P = 0·02) (Fig. 2).Compared to normal rats medullar cyclin gene expressiondid not change significantly regardless of the phase of the
Fig. 1. Morphological analyses showing glomerular cell proliferation and matrix expansion 8 days after glomerulonephritis induction (a) normal rats,
(b) nephritic rats (CD8+) sacrificed 8 days after nephritis induction: an extensive mesangial expansion obliterating the urinary space can be observed.
Compared to controls, (d) DMSO and (f) buffered saline, rats treated with MMF (e) or Roscovitine (c) showed improvement in histological changes.
(c) (e)(a)
(b) (d) (f)

MMF and cyclins in the anti Thy-1 model
© 2004 British Society for Immunology, Clinical and Experimental Immunology, 139: 225–235 229
experiment (data not shown). Compared to controls nochanges in the 1-day (phase I) gene expression of p27, theCDKs inhibitor, were detectable in the cortical tissue. How-ever, p27 expression decreased remarkably during mesangialcell proliferation (phase II) (CD1+ versus C-: 1·49 ± 0·45 ver-sus 1·6 ± 0·45, P = NS; CD8+ versus C-: 0·6 ± 0·21 versus1·6 ± 0·45, P = 0·002)(Fig. 2).
The glomerular expression of cyclin D1, cyclin D2, cyclinD3 and mainly, cyclin B increased significantly during theearly phase of the disease (phase I) as compared to quiescentglomeruli, while glomerular p27 mRNA levels, which areusually highly expressed in the mature kidney, decreasedduring phase II of sustained mesangial cell proliferation.
As compared to rats given DMSO and buffered saline,animals given roscovitine and MMF during the peak ofmesangial cell proliferation (phase II) showed a remarkable
reduction in glomerular d-type cyclins and cyclin B mRNAlevels, which was coupled with an increase in p27 expression(Fig. 3). Cyclin expression in medullar tissue did not vary inof the any experimental conditions (i.e. roscovitine, MMFand vehicle administration). (data not shown).
In situ hybridization
Light microscopy observation revealed a positive pattern inthe mesangium and in endothelial cells. Normal glomeruli
Fig. 2. Cyclin and p27 gene expression in cortical renal tissue of
nephritic rats treated with the anti Thy 1 monoclonal antibody.Com-
pared to normal rats, the cortical tissue of nephritic animals already
showed increased gene expression for D-type cyclins and for cyclin B
one day after disease induction (phase I, CD1+ group). This up-regu-
lation was maintained throughout the proliferative phase (until the
eighth day, phase II, CD8+ rats). Gene expression of the CDK inhibitor
p27, which was found to be intensively positive in CD1+ nephritic rats
as well as in normal ones, considerably decreased during mesangial cell
proliferation (phase II, CD8+ rats). All gene expression data are
expressed as [(gene target in cortex – GAPDH in cortex) – (gene target
in medulla -GAPDH in medulla)]. Levels of significance refer to CD1+
and CD8+ compared to C-: ***P < 0·001; **P < 0·01; *P < 0·05.
2·5
2
1·5
1
0·5
0[(C
t gen
e ta
rget
-Ct G
AP
DH
) co
rtte
x –
(Ct g
ene
targ
et-G
AP
DH
) m
edul
la]
CB CD1 CD2 CD3 CE p27 p21
**** **
**
***
***
*
* **
**
** **
C–
CD1+
CD8+
CB
CD1
CD2
CD3
CE
p27
p21
C–
0·22 ± 0·05
0·15 ± 0·05
0·09 ± 0·06
0·3 ± 0·1
0·29 ± 0·09
1·6 ± 0·45
0·04 ± 0·007
1·4 ± 0·64
1·24 ± 0·48
2·17 ± 1·01
1·28 ± 0·6
1·04 ± 0·214
1·49 ± 0·46
1 ± 0·27
1·2 ± 0·49
0·88 ± 0·17
1·76 ± 0·98
1·04 ± 0·70
0·87 ± 0·195
0·6 ± 0·21
1 ± 0·31
CD1+ CD8+
Fig. 3. Cyclin and p27 gene expression in cortical renal tissue of
nephritic rats treated with mycophenolate mofetil and roscovitine.
(a) Plot of gene expression data of the rats treated with roscovitine (R)
and roscovitine vehicle (DMSO, D). (b) Plot of gene expression data of
the rats treated with MMF and MMF vehicle (buffered saline, F). All
gene expression data are expressed as [(gene target in cortex – GAPDH
in cortex) – (gene target in medulla -GAPDH in medulla)]. Levels of
significance refer to R compared to D: ∞∞P < 0·01; ∞P < 0·05; MMF com-
pared to F: §§§P < 0·001; §P < 0·05.
1·81·61·41·2
10·80·60·40·2
0CB CD1 CD2 CD3 CE p27
[(C
t gen
e ta
rget
-Ct G
AP
DH
) co
rtex
–(C
t gen
e ta
rget
-GA
PD
H)
med
ulla
]
RDo
o
o
oo
CBR D
0·60 ± 0·08 0·93 ± 0·010·88 ± 0·39 1·02 ± 0·361·54 ± 0·14 1·99 ± 0·140·77 ± 0·01 0·96 ± 0·080·94 ± 0·47 1·20 ± 0·321·09 ± 0·01 0·91 ± 0·09
CD1CD2CD3CEP27
2·5
2
1·5
1
0·5
0CB CD1 CD2 CD3 CE p27
[(C
t gen
e ta
rget
-Ct G
AP
DH
) co
rtex
–(C
t gen
e ta
rget
-GA
PD
H)
med
ulla
]
MMFF§ §§§ §
§
CBMMF F
0·92 ± 0·12 1·06 ± 0·070·99 ± 0·17 1·17 ± 0·060·87 ± 0·12 2·10 ± 0·050·92 ± 0·07 1·08 ± 0·011·19 ± 0·28 1·14 ± 0·271·02 ± 0·01 0·96 ± 0·03
CD1CD2CD3CEP27
(a)
(b)

M. Chiara et al.
230 © 2004 British Society for Immunology, Clinical and Experimental Immunology, 139: 225–235
presented a cyclin D weak positive pattern, absence of signalfor cyclin B and a diffuse positive pattern for p27 mRNA.
As compared to DMSO, roscovitine induced a decrease inthe number group D cyclin positive glomeruli (D2 and D310% versus 90%, P < 0·001) (Fig. 4), and in the number ofpositive cells per glomerulus’s (D2: 4·7 ± 1 versus 45·4 ± 9·6,P < 0·001; D3: 7·15 ± 1·9 versus 25 ± 6·5, P < 0·001), as wellas, with a decrease in signal intensity as determined by dou-ble blind microscopy observation (CD2: 1+ to 0; CD3: 2+ to±) (Fig. 5).
Compared to vehicle, mycophenolate mofetil induced adecrease in the cyclin D2 signal (2+ to ±), a decrease in thenumber of positive cells per glomerulus (3·8 ± 1·5 versus23 ± 8·2, P < 0·001) and a slight reduction in the number ofpositive glomeruli (70% versus 95%) (Fig. 6). In MMFtreated rats, cyclin D3 was expressed in 20% of the glomer-uli and 4·8 ± 1·6 cells/glomerulus versus 90% of glomeruliand 33 ± 7·9 cells/glomerulus (P < 0·001) in controls (Figs 6and 7).
Cyclin inhibitor p27 gene expression was found to be sig-nificantly higher in roscovitine and MMF-treated rats ascompared to controls, with a definite increase in both thenumber of positive glomeruli (R 90% versus DMSO 26%;MMF 100% versus F 20%) (Figs 4–7) and number of positivecells per glomerulus (R 32·6 ± 11·75 versus DMSO 11·2 ± 3·2
P < 0·001; MMF 27·5 ± 7·58 versus F 1–2 cells, P < 0·001)(Figs 4 and 6).
Immunohistochemistry
Cyclin inhibitor p27kip1 is post-transcriptionally regulated bychanges in protein translation and degradation [20]. p27kip1
protein levels are critical for cell proliferation [21]. Thiscyclin inhibitor is widely expressed in quiescent cells [21].
In untreated rats (CD1-), p27kip1 was found to be diffuselyexpressed both in mesangial and in epithelial cells. CD1+rats, that were sacrificed 1 day after glomerulonephritisinduction, presented a decrease in labelled glomeruli (62%versus 100%).
p27kip1 expression was completely abrogated in nephriticrats (CD8+) whose tissue sections showed clear glomerularhypercellularity with a reduction in urinary space.
Both Roscovitine and Mycophenolate mofetil treatmentinduced a significant up-regulation of p27kip1 expression whencompared to controls (60% versus 0% P < 0·001) (Fig. 8).
Renal function and urinary parameters
The anti Thy-1 glomerulonephritis caused a decrease in cre-atinine clearance: 0·99 ± 0·31 and 1·07 ± 0·28 in saline and
Fig. 4. In situ hybridization: quantification of
labelling after roscovitine treatment. (a) plot of
the number of labelled cells per glomerulus (b)
plot of the percentage of labelled glomeruli per
section.Labelling was evaluated in at least 4 rats
per treatment, in 2 sections per rat, and in at least
50 glomeruli per section. R, roscovitine;
D, DMSO (roscovitine vehicle). Levels of signifi-
cance refer to R compared to D: ***P < 0·001.
50
(a)
(b)
45
40
35
30
25
20
15
10
5
0D2 D3 p27
Mea
n ce
lls/g
lom
erul
a
90%
90%
90%
10%
10%
26%P27
CD3
CD2
******
***
RD
RD
*** P < 0·001

MMF and cyclins in the anti Thy-1 model
© 2004 British Society for Immunology, Clinical and Experimental Immunology, 139: 225–235 231
Fig. 5. In situ hybridization in roscovitine treated rats. Decrease in signal intensity for group D cyclins and increase in p27 gene expression in roscovitine
(R) treated rats compared to controls receiving DMSO (D).
Fig. 6. In situ hybridization: quantification of
labelling after mycophenolate mofetil treatment.
(a) plot of the number of labelled cells per glom-
erulus. (b) plot of the percentage of labelled
glomeruli per section.Labelling was evaluated
in at least 4 rats per treatment, in 2 sections
per rat, and in at least 50 glomeruli
per section.MMF = mycophenolate
mofetilF = buffered saline (MMF vehicle) Levels
of significance refer to MMF compared to F:
***P < 0·001.
35
(a)
(b)
30
25
20
15
10
5
0D2 D3 p27
Mea
n ce
lls/g
lom
erul
a
100%
90%
95%
20%
70%
20%P27
CD3
CD2
******
***
MMFF
MMFF
*** P < 0·001

M. Chiara et al.
232 © 2004 British Society for Immunology, Clinical and Experimental Immunology, 139: 225–235
Fig. 7. In situ hybridization in mycophenolate mofetil treated rats. Decrease in signal intensity for group D cyclins and increase in p27 gene expression
in MMF-treated rats (MMF) compared to controls receiving buffered saline (F).
Fig. 8. p27 expression. p27kip1 in untreated rats (CD1-) was found to be diffusely expressed both in mesangial and in epithelial cells, as per examination
by immunohistochemical techniques, while CD1+ rats presented a decrease in labelled glomeruli (not shown). p27kip1 expression was completely
abrogated in nephritic rats (CD8+), where tissue sections showed clear glomerular hypercellularity with a reduction in urinary space. Compared to
controls (DMSO, D and saline, F) both Roscovitine (R) and Mycophenolate mofetil MMF treatment induced a significant up-regulation of p27kip1
expression.

MMF and cyclins in the anti Thy-1 model
© 2004 British Society for Immunology, Clinical and Experimental Immunology, 139: 225–235 233
DMSO-treated rats, and 1·03 ± 0·17 ml/min in untreated(CD8+) animals with anti Thy-1 nephritis versus 1·85 ±0·37 ml/min in non-nephritic rats (P < 0·002). In rats givenMMF or roscovitine renal function was preserved with val-ues of creatinine clearance of 1·56 ± 0·32 and 1·63 ± 0·24 ml/min, respectively, P < 0·007 compared to untreated nephriticrats, P = n.s. as compared to non nephritic rats.
Significant differences were found between roscovitine orMMF-treated rats and vehicle-treated diseased animals asregard to the main urinary markers: proteinuria: 32 ± 8 mg/day in roscovitine-treated versus 55 ± 10 mg/day in DMSOtreated-rats, P < 0·02; 35 ± 11 mg/day in MMF-treated versus52 ± 7 mg/day in saline-treated rats, P < 0·05; nephritic rats(CD8+) had a urinary excretion of 60 ± 16 mg/day, P < 0·02as compared to roscovitine and MMF-treated rats. Haema-turia was 0·1 ± 0·04 mg/dl in nephritic (CD8+) rats,0·07 ± 0·02 in DMSO-treated and 0·04 ± 0·01 in saline-treated nephritic rats, but absent in non-nephritic animalsand in roscovitine- and MMF-treated rats.
Discussion
The present study shows that the acute phase of the anti Thy1 experimental model of mesangial proliferative glomerulo-nephritis, which is histologically defined by mesangialhypercellularity and deposition of mesangial matrix, is char-acterized by an increase in cyclin D and B and a decrease inp27kip1 gene expression. Roscovitine and MMF treatmentsduring the florid phase of mesangial cell proliferation causesa similar decrease in cyclin gene expression (both B and D-groups), and a parallel increase in levels of cell cycle inhibitorproteins.
More specifically, a definite increase in cyclin B, D1,D2and D3 gene expression in the cortical portion is seen at day1 after glomerulonephritis induction. Due to the absence ofinfiltrating granulocytes and monocytes, cyclin increase islikely related to intrinsic glomerular cell changes. Cyclin up-regulation starts at the same time as of mesangiolysis andpersists over the entire proliferative phase up to the 8th day.Conversely, p27 mRNA levels show a progressive decreasethat reaches its negative peak eight days after glomerulone-phritis induction.
The role that cell cycle proteins play in renal cell prolifer-ation has been extensively described [22]. Shankland et al.[9,22] showed that mesangial cell proliferation is associatedwith late S-G2 phase cyclin up-regulation, and inverselyrelated to the decrease in p27kip1. The natural resolution ofmesangial cell proliferation in the Thy 1 model ofexperimental nephritis is paralleled by a decrease incyclin expression, with normalization of p27kip1 and p21levels [9].
The 27-kDa protein, p27kip1 (p27) is widely expressed innonproliferating renal and nonrenal cells [23–25]. Unlikep21, p27 expression is post-transcriptionally regulated bychanges in protein translation and degradation through the
ubiquitin proteolytic pathway [26,27]. The protein is alsopost-translationally modified by phosphorilation [28]. p27regulates growth arrest in response to TGFb, rapamycin, andcAMP [24]. This protein is critical in regulating renal cell dif-ferentiation, apoptosis, proliferation, and hypertrophy [29].
In the present study, high protein and mRNA levels of p27were found in untreated animals. Induction of the glomer-ular changes by the anti Thy1 MoAb promoted a slowdecrease in gene and protein expression, which was almostcompletely abrogated in glomeruli after 8 days of disease.
We examined the factors of the cell-cycle regulation sys-tem that can possibly be affected by treatment with antipro-liferative drugs, namely mycophenolate mofetil (MMF) androscovitine, in this experimental model of glomerulonephri-tis. MMF is a powerful immunosuppressor that is used inorgan transplantation [14,15] and that has been successfullyintroduced in the treatment of primary and secondaryhuman glomerulonephritis [30–33]. MMF is an inhibitor ofthe inosin monophosphate dehydrogenase (IMPDH) whichis responsible for the de novo synthesis of guanosine [14,15].The effects of Mycophenolic acid on cell cycle progressionhave been studied in activated normal human T lymphocytes[34]. The expression of cyclin D3, which is a major compo-nent of the cyclin-dependent kinase activity required for pRbphosphorylation, was found to be completely abrogated,whereas the expression of cyclin D2 was attenuated. In addi-tion, MMF prevented the elimination of p27 by sustainingretention of high levels of protein.
These findings support the notion that, apart from itsimmunosuppressive effects, the antiproliferative activities ofMMF on mesangial cells [17] can be, at least in part, medi-ated via intervention on the cell-cycle system. Alternatively,MMF could act via anti-inflammatory pathways, whichresult from the influence on the inducible form of nitricoxide synthase and the generation of peroxynitrite [16].More fundamental examination of the involved mechanismswas beyond the scope of this study.
Roscovitine is a purine analogue known to inhibit CDKcomplexes by interfering with the ATP-binding pocketlocated in the cleft between the small and large lobe of thekinase [11,35,36]. It acts preferentially on CDK2, CDK1 andCDK5 [11]. Roscovitine treatment inhibits the proliferationof rat mesangial cells in vitro [12] and significantly reducesthe magnitude of mesangial cell proliferation in the Thy1model of proliferative glomerulonephritis, thus improvingrenal function and reducing matrix production [12].
In the present study rats sacrificed in the florid phase ofnephritis induction which were treated with MMF and Ros-covitine since day 3 showed, compared to controls, a signif-icant decrease in cyclin gene expression mainly in cyclin B,D2 and D3.
In conclusion, definite interferences by MMF and rosco-vitine cyclin and p27 regulation systems have been demon-strated in this model of renal injury. Both these compoundspromoted resolution of mesangial proliferation and

M. Chiara et al.
234 © 2004 British Society for Immunology, Clinical and Experimental Immunology, 139: 225–235
influenced functional and urinary parameters, such as crea-tinine clearance and proteinuria and haematuria. No signs oftoxicity were observed with any of the drugs used in thisshort-term study. Experimental mesangial proliferativeglomerulonephritis induced by antibody to Thy-1 is anextensively studied model of mesangial cell proliferation inresponse to immune injury. Binding of the Thy1 antibody tomesangial cells causes mesangiolysis [37] which is followedby mesangial proliferation. Although marked and excessive,this PDGF and bFGF-dependent mesangial cell proliferation[38] represents the reparative phase of the Thy1 experimen-tal nephritis. In the present as in other reports [13], bluntingthis phase, which is protective in the animal model, served asa tool to examine the ability of chemical compounds inreducing mesangial proliferation (that in humans is oftenfollowed by glomerular sclerosis), and inhibiting cyclinactivity, which is essential for cell cycle machinery.
Our findings collectively support the rationale of theemploy of MMF in mesangial proliferative glomerulone-phritis and confirm roscovitine to be an interesting novelcandidate for the treatment of glomerular disease.
Acknowledgements
The Authors thank Dr Valery Frances Perricone for help inreviewing the manuscript.
References
1 Ooi BS, Cohen DJ, Veis JH. Biology of the mesangial cell in glom-
erulonephritis- role of cytokines. Proc Soc Exp Biol Medical 1996;
213:230–7.
2 Couser WG. Pathogenesis of glomerular damage in glomerulone-
phritis. Nephrol Dial Transplant 1998; 13:10–5.
3 Pines J. Cyclin and cyclin-dependent kinases: a biochemical view.
Biochem J 1995; 308:697–711.
4 Sherr CJ. Mammalian G1 cyclins. Cell 1993; 73:1059–65.
5 Weiberg RA. The retinoblastoma protein and cell cycle control.
Cell 1995; 81:323–30.
6 Norbury C, Nurse P. Animal cell cycles and their control. Annu
Rev Biochem 1992; 61:441–71.
7 Yeudall WA, Zhang H, Beach D. Cyclin kinase inhibitors add a new
dimension to cell cycle control. Oral Oncol Eur Cancer 1995;
13:291–8.
8 Nakayama H, Oite T, Kawachi H, Marioka T, Kobayashi H, Orikasa
M, Arakawa M, Shimizu F. Comparative nephritogenicity of two
monoclonal antibodies that recognize different epitopes of rat Thy-
1 molecules. Nephron 1998; 78:453–63.
9 Shankland SJ, Hugo C, Coats SR et al. Changes in cell-cycle protein
expression during experimental mesangial proliferative glomeru-
lonephritis. Kidney Int 1996; 50:1230–9.
10 Shankland SJ. Cell-cycle control and renal disease. Kidney Int 1997;
52:294–308.
11 Meijer L, Borgne A, Mulner O et al. Biochemical and cellular effects
of roscovitine, a potent and selective inhibitor of the cyclin-
dependent kinases cdc2, cdk2, cdk5. Eur J Biochem 1997; 243:527–
36.
12 Pippin JW, Qu Q, Meijer L, Shankland SJ. Direct in vivo inhibition
of the nuclear cell-cycle cascade in experimenta1 mesangial prolif-
erative glomerulonephritis with roscovitine, a novel cyclin-
dependent kinase antagonist. J Clin Invest 1997; 100:2512–20.
13 Lehrke I, Schaier M, Schade K et al. Retinoid specific-agonist alle-
viate experimental glomerulonephritis. Am J Physiol Renal Physiol
2002; 282:F741–51.
14 Allison AC, Eugui EM. Purine metabolism and immunosuppres-
sive effects of mycophenolate mofetil (MMF). Clin Transplant
1996; 10:77–84.
15 Allison AC, Eugui EM. Mycophenolate mofetil and its mechanism
of action. Immunopharmacology 2000; 47:85–118.
16 Srinivas TR, Kaplan B, Meier-Kriesche HV. Mycophenolate
mofetil-organ transplantation. Expert Opin Pharmacother 2003;
4:2325–45.
17 Ziswiler R, Steinmann-Niggli K, Kappeler A, Daniel C, Marti HP.
Mycophenolic acid: a new approach to the therapy of experimental
mesangial proliferative glomerulonephritis. J Am Soc Nephrol
1998; 9:2055–66.
18 Shimizu A, Kitamura H, Masuda Y, Ishizaki M, Sugisaki Y, Yama-
narka N. Apoptosis in the repair process of experimental prolifer-
ative glomerulonephritis. Kidney Int 1995; 47:114–21.
19 Tisher CC, Brenner BM. Renal Pathology With Clinical and Func-
tional Correlations. Philadelphia: Lippincott, Williams & Wilkins,
1989.
20 Hengst L, Reed SI. Translational control of p27kip1 accumulation
during the cell cycle. Science 1996; 271:1861–4.
21 Hara T, Kamura T, Nakayama K, Oshikawa K, Hatakeyama S. Deg-
radation of p27 (kip1) at the G(0)-G(1) transition mediated by a
Skp2-independent ubiquitination pathway. J Biol Chem 2001;
276:48937–43.
22 Shankland SJ, Wolf G. Cell cycle regulatory proteins in renal dis-
ease. role in hypertrophy, proliferation, and apoptosis. Am J Phys-
iol 2000; 278:F515–F529.
23 Coombs HL, Shankland SJ, Setzer SV, Hudkins KL, Alpers CE.
Expression of the cyclin kinase inhibitor, p27kip1, in developing and
mature human kidney. Kidney Int 1998; 53:892–6.
24 Nourse J, Firpo E, Flanagan WM et al. Interleukin-2-mediated
elimination of the p27kip1 cyclin-dependent kinase inhibitor pre-
vented by rapamycin. Nature 1994; 372:570–3.
25 Shankland SJ, Pippin J, Flanagan M et al. Mesangial cell prolifera-
tion mediated by PDGF and bFGF is determined by levels of the
cyclin kinase inhibitor p27kip1. Kidney Int 1997; 51:1088–99.
26 Nguyen H, Gitig DM, Koff A. Cell-free degradation of p27
(kip1), a G1 cyclin-dependent kinase inhibitor, is dependent on
CDK2 activity and the proteaseome. Mol Cell Biol 1999;
19:1190–201.
27 Toyoshima H, Hunter T. p27, a novel inhibitor of G1 cyclin-Cdk
protein kinase activity, is related to p21. Cell 1994; 78:67–74.
28 Sheaff RJ, Groudine M, Gordon M, Roberts JM, Clurman BE.
Cyclin E-CDK2 is a regulator of p27 (kip1). Genes Dev 1997;
11:1464–78.
29 Shankland SJ. The growing role for the cyclin kinase inhibitor
p27kip1 in renal disease. Kidney Int 1998; 54:2241–2.
30 Chan TM, Li FK, Tang CSO et al. Efficacy of mycophenolate
mofetil in patients with diffuse proliferative lupus nephritis. N
Engl J Med 2000; 343:1156–62.
31 Polenakovich M, Grcevsa L, Dzikova S. Mycophenolate mofetil in
treatment of idiophatic stages III–IV membranous nephropathy.
Nephrol Dial Transplant 2003; 18:1233–4.

MMF and cyclins in the anti Thy-1 model
© 2004 British Society for Immunology, Clinical and Experimental Immunology, 139: 225–235 235
32 Zatz R, Noronha IL, Fujihara CK. Experimental and clinical ratio-
nale for use of MMF in nontrasplant progressive nephropathies.
Am J Physiol 2002; 283:F1167–75.
33 Badid C, Desmouliere A, Laville M. Mycophenolate mofetil. Impli-
cations for the treatment of glomerular disease. Nephrol Dial Trab-
splant 2001; 16:1752–6.
34 Lalibertè J, Yee A, Xiong Y, Mitchell BS. Effects of guanine nucle-
otide depletion on cell cycle progression in human T lymphocytes.
Blood 1998; 91:2896–904.
35 de Arzavedo WF Jr, Leclerc S, Meijer L, Havlicek L, Strnad M, Kim
S-H. Inhibition of cyclin dependent kinases by purine analogues:
crystal structure of human cdk2 complexed with roscovitine. Eur J
Biochem 1997; 243:518–26.
36 Gray N, Detivaud L, Doerig C, Meijer L. ATP-site directed inhibi-
tors of cyclin-dependent kinases. Curr Med Chem 1999; 6:859–75.
37 Yamamoto T, Wilson CB. Quantitative and qualitative studies of
antibody-induced mesangial cell damage in the rat. Kidney Int
1987; 32:514–25.
38 Johnson RJ, Raines EW, Floege J, Yoshimura A, Pritzl P, Alpers C,
Ross R. Inhibition of mesangial cell proliferation and matrix
expansion in glomerulonephritis in the rat by antibody to platelet-
derived growth factor. J Exp Med 1992; 175:1413–6.