molecular mechanisms underlying the rewarding effects of cocaine
TRANSCRIPT

47
Ann. N.Y. Acad. Sci. 1025: 47–56 (2004). © 2004 New York Academy of Sciences.doi: 10.1196/annals.1316.006
Molecular Mechanisms Underlying the Rewarding Effects of Cocaine
F. SCOTT HALL,a ICHIRO SORA,b JANA DRGONOVA,a XIAO-FEI LI,a MICHELLE GOEB,a AND GEORGE R. UHLa
aMolecular Neurobiology Branch, NIDA-IRP, NIH/DHHS, Baltimore, Maryland 21224, USAbTohoku University Graduate School of Medicine, Department of Neuroscience,Sendai, Japan
ABSTRACT: The initially surprising observation that cocaine retains its re-warding effects in dopamine transporter (DAT) knockout (KO) mice led ourlaboratory to examine the effects of deletion of other monoaminergic genes oncocaine reward. Our initial approach to this problem was to combine DAT KOmice with serotonin transporter (SERT) KO mice to make combined DAT/SERT KO mice. The combination of these knockouts eliminates cocaine re-ward as assessed in the conditioned place preference (CPP) paradigm. We havealso identified evidence that, in the absence of DAT, there is greater participa-tion in cocaine reward by serotonin (SERT) and norepinephrine (NET) trans-porters. Both NET and SERT blockers (nisoxetine and fluoxetine) producedsignificant CPPs in DAT KO mice, but not in wild-type (WT) mice. The strik-ing elimination of cocaine CPP in combined DAT/SERT KO mice contrastswith effects that we have identified in combined NET/SERT knockout mice,which display increases in cocaine reward, and with recent reports that suggestthat DAT/NET combined KOs retain substantial cocaine CPP. Overall, thesestudies indicate important requirements for several monoaminergic systemgenes to fully explain cocaine reward, in particular those expressed by dopam-ine and serotonin systems.
KEYWORDS: cocaine reward; transporters; dopamine; serotonin; norepinephrine
INTRODUCTION
The rewarding actions of addictive drugs, as assessed in tests such as conditionedplace preference (CPP) or self-administration, could be based on actions at singlesites or the sorts of actions at multiple sites that have been described as “dirty drugs.”The rewarding effects of several drugs can be eliminated by genetic deletion of a sin-gle gene that encodes a single drug target. Morphine place preference is eliminated,for example, in µ opiate receptor gene knockouts.30 As we note in more detail below,we and others have shown that cocaine reward cannot be eliminated by knockout ofany of the three major molecular targets for cocaine action: the plasma membrane
Address for correspondence: F. Scott Hall, Ph.D., Molecular Neurobiology, Box 5180,Baltimore, MD 21224. Voice: 410-550-1589; fax: 410-550-1535.

48 ANNALS NEW YORK ACADEMY OF SCIENCES
transporters for dopamine (DAT), serotonin (SERT), and norepinephrine (NET).32,42
Furthermore, these studies revealed enhanced cocaine place preference after remov-al of SERT or NET.
With this information in hand, we postulated that several gene knockouts werelikely to be required to eliminate the rewarding effects of cocaine, and we were ableto identify the only combined transporter knockouts to date that have eliminated co-caine place preference, knockouts of DAT and SERT, as detailed below.
We were also aware of the possibility that the mechanism by which cocaine re-ward was mediated in either single- or multiple-transporter gene knockout mice thatdisplayed retained cocaine reward was unlikely to be identical to the ways in whichit was mediated in wild-type (WT) animals. The greater reward produced by cocainein some single-transporter knockouts and the evidence for novel rewarding proper-ties acquired by SERT and NET blockers that was absent in WT animals providedevidence for such differences. These lines of evidence are also detailed below.
SINGLE GENE KNOCKOUTS AND COMPENSATORY CHANGES INTHE MAINTENANCE OF COCAINE REWARD
The prototypical psychomotor stimulant cocaine increases locomotor activity, in-duces stereotypical behavior, and is highly reinforcing (for review see Ref. 15). Co-caine also produces aversive effects in humans8,27 and animals (for review see Ref.2). Cocaine blocks monoamine uptake by neuronal plasma membrane transportersfor dopamine (DAT), serotonin (SERT), and norepinephrine (NET). Recent studieshave concentrated on the effects of deletion of these transporters alone, and in com-bination, but cocaine also blocks ligand- and voltage-gated channels, although atlower potencies.16 Considering each of these molecular targets of cocaine is impor-tant for understanding its behavioral sequelae and, in particular, for understandingthe consequences of gene deletions.37
Prior to the use of gene knockout techniques, the rewarding and locomotor stim-ulating effects of cocaine and other drugs of abuse were generally associated withdopamine systems. This hypothesis was based on several lines of evidence from le-sion and pharmacological experiments (see Ref. 40). Consequently, this became theinitial focus of the first transgenic studies of the molecular bases of cocaine rewardwhich were thought to support the role of DAT as the sole mediator of cocaine’s ac-tions.9 This presumption was based on the elimination of the locomotor stimulatingeffects of cocaine in DAT knockout mice and its misinterpretation as “indifference”to cocaine.9 However, studies from our laboratory and others documented that therewarding effects of cocaine were virtually unaltered in each of two separate lines ofDAT knockout mice.25,32
We then proceeded under the hypothesis that the elimination of multiple genesmight be necessary to eliminate the rewarding effects of cocaine. Single gene knock-out of DAT failed to affect cocaine CPP, while deletion of SERT or NET increasedcocaine CPP.32,42 The simplest hypotheses that might account for these results werethat cocaine actions at no one transporter were sufficient to produce full cocaine re-warding effects, and that actions at SERT and NET were at least partially aversive,such that their removal enhanced cocaine reward.37 Such data are consistent with therelatively weak rewarding effects of DAT or DAT/NET blockers.33,35,41
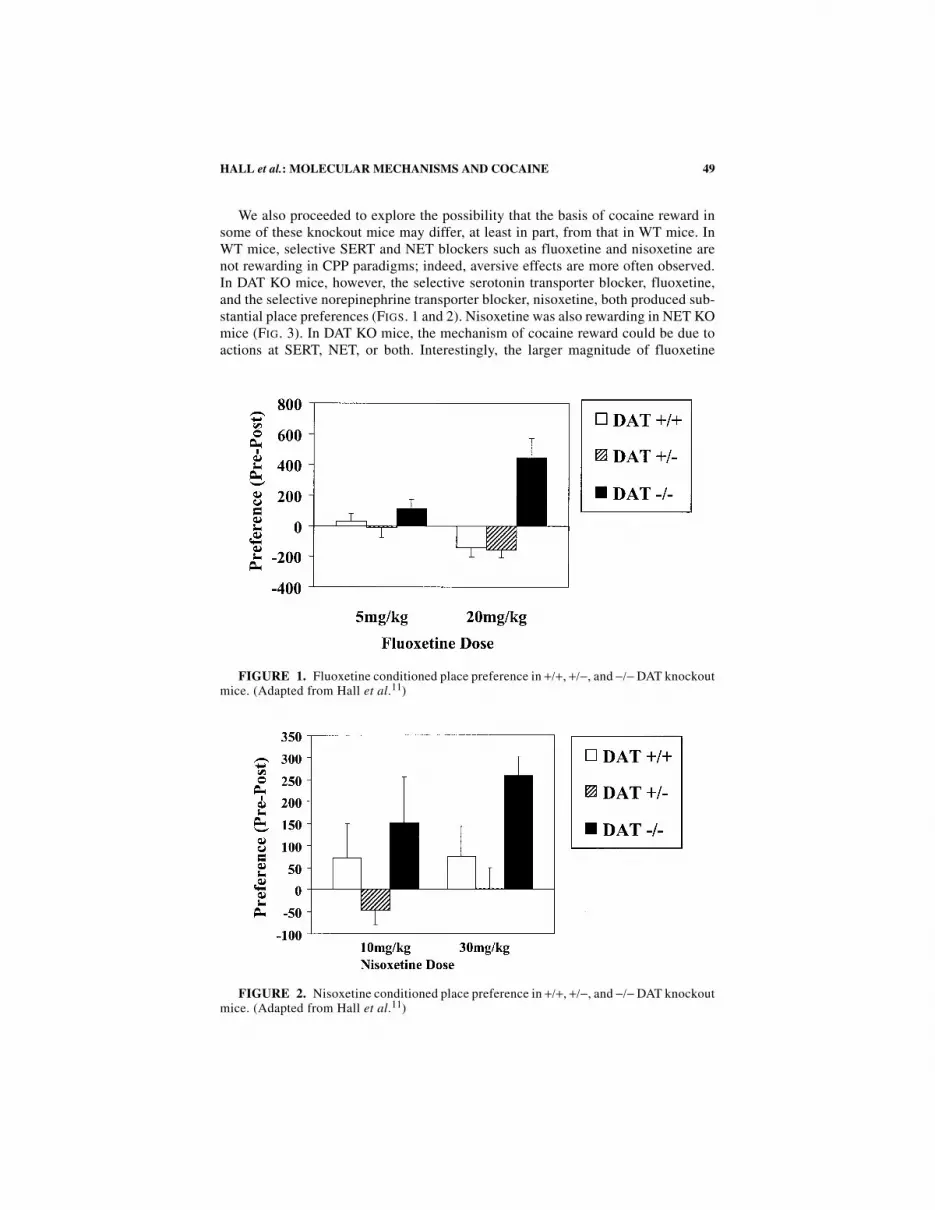
49HALL et al.: MOLECULAR MECHANISMS AND COCAINE
We also proceeded to explore the possibility that the basis of cocaine reward insome of these knockout mice may differ, at least in part, from that in WT mice. InWT mice, selective SERT and NET blockers such as fluoxetine and nisoxetine arenot rewarding in CPP paradigms; indeed, aversive effects are more often observed.In DAT KO mice, however, the selective serotonin transporter blocker, fluoxetine,and the selective norepinephrine transporter blocker, nisoxetine, both produced sub-stantial place preferences (FIGS. 1 and 2). Nisoxetine was also rewarding in NET KOmice (FIG. 3). In DAT KO mice, the mechanism of cocaine reward could be due toactions at SERT, NET, or both. Interestingly, the larger magnitude of fluoxetine
FIGURE 1. Fluoxetine conditioned place preference in +/+, +/−, and −/− DAT knockoutmice. (Adapted from Hall et al.11)
FIGURE 2. Nisoxetine conditioned place preference in +/+, +/−, and −/− DAT knockoutmice. (Adapted from Hall et al.11)

50 ANNALS NEW YORK ACADEMY OF SCIENCES
place preference compared to nisoxetine place preference in DAT KO mice is con-sistent with data from combined DAT × SERT KO mice (see below). Further supportfor this novel action of fluoxetine in DAT KO mice comes from in vivo microdialy-sis data, demonstrating that fluoxetine increases extracellular dopamine in DAT KOstriatum but not in WT striatum.28
Other mechanisms are certainly necessary to explain the effects of cocaine in oth-er single transporter knockout mice. The effects of nisoxetine in the CPP paradigmwere unchanged in SERT KO mice, suggesting that the basis of enhanced cocainereward in these mice does not involve alterations in the rewarding valence of NETblockade.
MULTIPLE MONOAMINE TRANSPORTER KNOCKOUTS IDENTIFY GENE–GENE INTERACTIONS IN COCAINE REWARD
The creation of multiple gene knockouts was motivated by the inability of singlegene knockouts to eliminate cocaine reward25,32 and by pharmacological consider-ations that cocaine might produce its rewarding effects through actions at multiplesites of action. Removing multiple genes would eliminate functions that normally re-quire both genes and also alter compensatory adaptations to removal of the first genethat might require the second gene. Because of the strong evidence for serotonergicinvolvement in drug reward, we first combined DAT and SERT KO strains.31 Initialobservations of these mice showed that few baseline behavioral differences wereseen in these mice that were not observed in either of the single knockouts (e.g.,DAT-induced hyperactivity was also observed in DAT/SERT combined knockouts).
FIGURE 3. Fluoxetine conditioned place preference in +/+, +/−, and −/− NET knockoutmice. (Adapted from Hall et al.11)

51HALL et al.: MOLECULAR MECHANISMS AND COCAINE
Nevertheless, cocaine CPP was completely eliminated in these mice (FIG. 4). It wasnot necessary to eliminate both copies of the SERT gene to produce these effects;neither DAT −/− SERT +/− nor DAT −/− SERT −/− mice exhibited rewarding ef-fects of cocaine. Conversely, eliminating both copies of DAT was necessary. DAT+/− SERT −/− mice exhibited normal rewarding effects of cocaine. These compari-sons are consistent with a necessary role for SERT and a greater overall role for DATto produce normal cocaine reward.
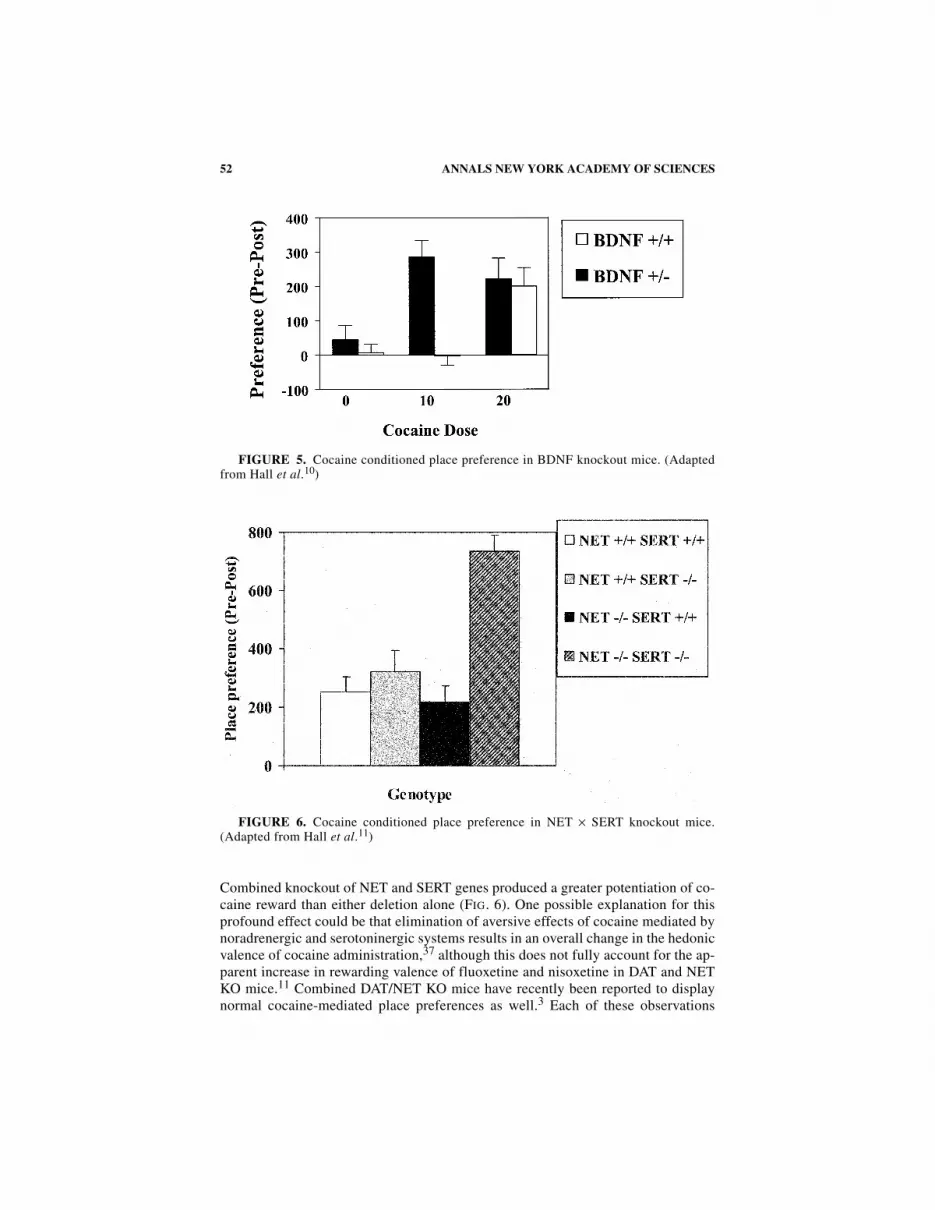
BDNF is a trophic factor that can influence both serotonergic and dopaminergicsystems.4,7,13,14,17,29,43 For this reason, we examined BDNF heterozygous knockoutmice in the same place preference paradigm used in our other studies. BDNF +/− KOmice had reduced cocaine CPP compared to WT mice,10 with a shift to the left in thedose-effect curve (FIG. 5). Initial assessments of dopamine cell counts and mRNAexpression levels for DAT, NET, SERT, and the 5-HT1A, 5-HT1B, 5-HT2A, and 5-HT2C receptors have failed to identify biochemical correlates of this altered cocainereward in monoaminergic systems (Drgonova, Hall, and Uhl, unpublished results).Thus, behavioral effects of single-copy BDNF gene deletion could arise from otheralterations in monoaminergic function that have yet to be detected or changes inother brain systems.
The observation that combined gene deletion of DAT and SERT eliminates co-caine reward contrasts with the effects of other combined transporter knockouts.11
FIGURE 4. Cocaine conditioned place preference in SERT × DAT double knockoutmice. (Adapted from Sora et al.31)
52 ANNALS NEW YORK ACADEMY OF SCIENCES
Combined knockout of NET and SERT genes produced a greater potentiation of co-caine reward than either deletion alone (FIG. 6). One possible explanation for thisprofound effect could be that elimination of aversive effects of cocaine mediated bynoradrenergic and serotoninergic systems results in an overall change in the hedonicvalence of cocaine administration,37 although this does not fully account for the ap-parent increase in rewarding valence of fluoxetine and nisoxetine in DAT and NETKO mice.11 Combined DAT/NET KO mice have recently been reported to displaynormal cocaine-mediated place preferences as well.3 Each of these observations
FIGURE 5. Cocaine conditioned place preference in BDNF knockout mice. (Adaptedfrom Hall et al.10)
FIGURE 6. Cocaine conditioned place preference in NET × SERT knockout mice.(Adapted from Hall et al.11)

53HALL et al.: MOLECULAR MECHANISMS AND COCAINE
demonstrates the specificity of the effects of combined DAT/SERT knockouts: theablation of cocaine CPP identified in these mice is found in no other single- ordouble-transporter knockout mice.
The role for SERT in cocaine reward identified in this double knockout is alsoconsistent with the hedonic consequences of pharmacologic, genetic, and neurotoxicmanipulations of brain serotonin systems.5,6,12,18–22,24,26,34,36 Because manipula-tion of serotonergic systems can alter both rewarding and aversive effects of cocaine,elimination of cocaine reward in DAT × SERT KO mice could involve a selectedsubset of serotonin system components.
One approach to dissecting these subcomponents of the serotonin system thatmay be involved in cocaine reward is to determine whether the effects of DAT/SERTknockout on cocaine reward can also be produced by combining DAT knockout withknockout of serotonin receptor subtypes, and/or combining SERT knockout withknockout of dopamine receptor subtypes. We have combined deletion of DAT withdeletion of the serotonin 1B receptor (5-HT1B), deletion of DAT with deletion of theserotonin 1A receptor (5-HT1A), and deletion of SERT with deletion of the dopam-ine D2 receptor (DRD2). None of these combined knockouts has produced dimin-ished cocaine CPP similar to that observed in DAT/SERT knockout mice (Hall andUhl, in preparation). Interestingly, however, the combined DAT KO/5HT1B receptorheterozygote mice display restoration of cocaine induced locomotion, which is lostin single DAT KO mice, and near-reversal of DAT hyperactivity (Hall and Uhl, inpreparation).
CONCLUSIONS
The accumulating evidence from gene knockout studies of single and multiplegene knockout mice produces a complex picture of interactive gene effects. Thiscomplex picture fits with recent polygenic descriptions of genetic influences on hu-man addiction.38 Our current understanding of the mechanisms of cocaine’s reward-ing effects begins with evidence that no single transporter site alone is bothnecessary and sufficient to produce cocaine reward. This understanding continueswith the realization that cocaine must be working as a “dirty drug” based on the re-quirement that both the dopamine and serotonin transporters must be ablated to elim-inate cocaine’s rewarding effects. Current understanding also postulates thatmultiple genetic mechanisms contribute to aspects of cocaine seeking, includinggenes that regulate both rewarding and aversive consequences of cocaine adminis-tration. Finally, a growing body of evidence indicates that deletion of other genes canincrease cocaine CPP.
Attempts to put the molecular mechanisms of cocaine reward into the context ofspecific anatomical circuitry and biochemical relationships also need to be viewedin the context of learning and memory aspects of drug-seeking behavior, drug con-sumption, and even normal behavior in CPP testing that provides the bulk of the be-havioral underpinnings for the results discussed here. Although drug seeking hascomponents of motivation, sometimes described by the term “craving” (for reviewsee Ref. 1), drug-related stimuli can also evolve to control behavior through roles assecondary reinforcers (see Ref. 23). Many well-learned drug-associated responses(see Ref. 39) can be best described as habits23 and may not involve strong motiva-

54 ANNALS NEW YORK ACADEMY OF SCIENCES
tional components. CPP, although referred to as an index of “reward,” involves seek-ing the environment where the drug was previously experienced. The knockouteffects summarized in this review thus access a portion of the behavioral systems in-volved in mnemonic, appetitive, and consummatory components of addiction.
Evidence of adaptations to gene deletions is accumulating in single transportergene knockout mice, including those that are likely to contribute to the retained orincreased cocaine responses found in some of these strains. Adaptations to chronicgene deletion do provide limitations to interpretation of knockout data in light of thequestion, “What does this gene product normally do?”; nonetheless, knockouts pro-vide substantial insight into the possible mechanisms that can underlie the rewardingeffects of cocaine in animals and, by inference, in humans. Multiple gene knockoutstudies have now revealed additive, subtractive, and interactive effects of gene vari-ants. New phenotypes can emerge in some strains, such as the fluoxetine and nisox-etine reward in DAT and NET KO mice. This use of combined knockout mice toexamine interactions is improving our understanding of addiction genetics and pro-vides insight into the polygenic determination of drug addiction phenotypes in waysthat are not possible with other methods.
ACKNOWLEDGMENTS
The authors are grateful to the many co-investigators who have contributed to thiswork and for the financial support from the National Insitute on Drug Abuse Intra-mural Research Program, as well as to the organizers of the meeting from which thispaper derives.
REFERENCES
1. ALTMAN, J., B.J. EVERITT, S. GLAUTIER, et al. 1996. The biological, social and clinicalbases of drug addiction: commentary and debate. Psychopharmacology (Berl.) 125:285–345.
2. BLANCHARD, D.C. & R.J. BLANCHARD. 1999. Cocaine potentiates defensive behaviorsrelated to fear and anxiety. Neurosci. Biobehav. Rev. 23: 981–991.
3. DYKSTRA, L.A., L.M. BOHN, R.M. RODRIGUIZ, et al. 2003. Rewarding properties ofcocaine in dopamine/norepinephrine transporter knockout mice A.S.P.E.T. (Abstr.).
4. EATON, M.J., J.K. STALEY, M.Y. GLOBUS & S.R. WHITTEMORE. 1995. Developmentalregulation of early serotonergic neuronal differentiation: the role of brain-derivedneurotrophic factor and membrane depolarization. Dev. Biol. 170: 169–182.
5. FLETCHER, P.J. & K.M. KORTH. 1999. Activation of 5-HT1B receptors in the nucleusaccumbens reduces amphetamine-induced enhancement of responding for condi-tioned reward. Psychopharmacology (Berl.) 142: 165–174.
6. FLETCHER, P.J., M. TAMPAKERAS & J.S. YEOMANS. 1995. Median raphe injections of 8-OH-DPAT lower frequency thresholds for lateral hypothalamic self-stimulation.Pharmacol. Biochem. Behav. 52: 65–71.
7. GALTER, D. & K. UNSICKER. 2000. Brain-derived neurotrophic factor and trkB areessential for cAMP-mediated induction of the serotonergic neuronal phenotype. J.Neurosci. Res. 61: 295–301.
8. GAWIN, F.H. 1991. Cocaine addiction: psychology and neurophysiology. Science 251:1580–1586.
9. GIROS, B., M. JABER, S.R. JONES, et al. 1996. Hyperlocomotion and indifference tococaine and amphetamine in mice lacking the dopamine transporter. Nature 379:606–612.

55HALL et al.: MOLECULAR MECHANISMS AND COCAINE
10. HALL, F.S., J. DRGONOVA, M. GOEB & G. UHL. 2003. Reduced behavioral effects ofcocaine in heterozygous brain-derived neurotrophic factor (BDNF) knockout mice.Neuropsychopharmacology 28: 1485–1490.
11. HALL, F.S., X.F. LI, I. SORA, et al. 2002. Cocaine mechanisms: Enhanced cocaine, flu-oxetine and nisoxetine place preferences following monoamine transporter deletions.Neuroscience 115: 153–161.
12. HARRISON, A.A., L.H. PARSONS, G.F. KOOB & A. MARKOU. 1999. RU 24969, a 5-HT1A/1B agonist, elevates brain stimulation reward thresholds: an effect reversed byGR 127935, a 5-HT1B/1D antagonist. Psychopharmacology (Berl.) 141: 242–250.
13. HYMAN, C., M. HOFER, Y.A. BARDE, et al. 1991. BDNF is a neurotrophic factor fordopaminergic neurons of the substantia nigra. Nature 350: 230–232.
14. HYMAN, C., M. JUHASZ, C. JACKSON, et al. 1994. Overlapping and distinct actions ofthe neurotrophins BDNF, NT-3, and NT-4/5 on cultured dopaminergic and GABA-ergic neurons of the ventral mesencephalon. J. Neurosci. 14: 335–347.
15. JOHANSON, C.E. & M.W. FISCHMAN. 1989. The pharmacology of cocaine related to itsabuse. Pharmacol. Rev. 41: 3–52.
16. KUHAR, M.J., M.C. RITZ & J.W. BOJA. 1991. The dopamine hypothesis of the reinforc-ing properties of cocaine. Trends Neurosci. 14: 299–302.
17. MAMOUNAS, L.A., M.E. BLUE, J.A. SIUCIAK & C.A. ALTAR. 1995. Brain-derived neu-rotrophic factor promotes the survival and sprouting of serotonergic axons in ratbrain. J. Neurosci. 15: 7929–7939.
18. MILIARESSIS, E. 1977. Serotonergic basis of reward in median raphe of the rat. Pharma-col. Biochem. Behav. 7: 177–180.
19. NOMIKOS, G.G. & C. SPYRAKI. 1988. Effects of ritanserin on the rewarding propertiesof d-amphetamine, morphine and diazepam revealed by conditioned place preferencein rats. Pharmacol. Biochem. Behav. 30: 853–858.
20. PAPP, M. & P. WILLNER. 1991. 8-OH-DPAT-induced place preference and place aver-sion: effects of PCPA and dopamine antagonists. Psychopharmacology 103: 99–102.
21. POSCHEL, B.P. 1974. Proceedings: role of norepinephrine, dopamine, and serotonin inintracranial reward. Psychopharmacol. Bull. 10: 46–47.
22. REDGRAVE, P. 1978. Modulation of intracranial self-stimulation behaviour by local per-fusions of dopamine, noradrenaline and serotonin within the caudate nucleus andnucleus accumbens. Brain Res. 155: 277–295.
23. ROBBINS, T.W., M. CADOR, J.R. TAYLOR & B.J. EVERITT. 1989. Limbic-striatal interac-tions in reward-related processes. Neurosci. Biobehav. Rev. 13: 155–162.
24. ROBERTS, D.C. & G.F. KOOB. 1982. Disruption of cocaine self-administration follow-ing 6-hydroxydopamine lesions of the ventral tegmental area in rats. Pharmacol. Bio-chem. Behav. 17: 901–904.
25. ROCHA, B.A., F. FUMAGALLI, R.R. GAINETDINOV, et al. 1998. Cocaine self-administration indopamine-transporter knockout mice. Nat. Neurosci. 1: 132–137
26. ROCHA, B.A., K. SCEARCE-LEVIE, J.J. LUCAS, et al. 1998. Increased vulnerability tococaine in mice lacking the serotonin-1B receptor. Nature 393: 175–178.
27. SATEL, S.L., S.M. SOUTHWICK & F.H. GAWIN. 1991. Clinical features of cocaineinduced paranoia. NIDA Res. Monogr. 105: 371.
28. SHEN, H.-W., Y. HAGINO, H. KOBAYASHI, et al. 2004. Associations between extracellu-lar monoamines and cocaine reward in mice lacking dopamine and/or serotonintransporters. Neuropsychopharmacology, advance on-line publication, June 30; doi:10.1038/sj. upp. 1300476.
29. SON, J.H., H.S. CHUN, T.H. JOH, et al. 1999. Neuroprotection and neuronal differentia-tion studies using substantia nigra dopaminergic cells derived from transgenic mouseembryos. J. Neurosci. 19: 10–20.
30. SORA, I., G. ELMER, M. FUNADA, et al. 2001. Mu opiate receptor gene dose effects ondifferent morphine actions: evidence for differential in vivo mu receptor reserve.Neuropsychopharmacology 25: 41–54.
31. SORA, I., F.S. HALL, A.M. ANDREWS, et al. 2001. Molecular mechanisms of cocainereward: combined dopamine and serotonin transporter knockouts eliminate cocaineplace preference. Proc. Natl. Acad. Sci. USA 98: 5300–5305.

56 ANNALS NEW YORK ACADEMY OF SCIENCES
32. SORA, I., C. WICHEMS, N. TAKAHASHI, et al. 1998. Cocaine reward models: conditionedplace preference can be established in dopamine- and in serotonin-transporter knock-out mice. Proc. Natl. Acad. Sci. USA 95: 7699–7704.
33. STAFFORD, D., M.G. LESAGE, K.C. RICE & J.R. GLOWA. 2001. A comparison ofcocaine, GBR 12909, and phentermine self-administration by rhesus monkeys on aprogressive-ratio schedule. Drug Alcohol Depend. 62: 41–47.
34. TELLA, S.R. 1995. Effects of monoamine reuptake inhibitors on cocaine self-administration in rats. Pharmacol. Biochem. Behav. 51: 687–692.
35. TELLA, S.R., B. LADENHEIM, A.M. ANDREWS, et al. 1996. Differential reinforcingeffects of cocaine and GBR-12909: biochemical evidence for divergent neuroadap-tive changes in the mesolimbic dopaminergic system. J. Neurosci. 16: 7416–7427.
36. TRAN-NGUYEN, L.T., D.A. BAKER, K.A. GROTE, et al. 1999. Serotonin depletion attenu-ates cocaine-seeking behavior in rats. Psychopharmacology (Berl.) 146: 60–66.
37. UHL, G.R., F.S. HALL & I. SORA. 2002. Cocaine, reward, movement and monoaminetransporters. Mol. Psychiatry 7: 21–26.
38. UHL, G.R., Q.R. LIU & D. NAIMAN. 2002. Substance abuse vulnerability loci: converg-ing genome scanning data. Trends Genet. 18: 420–425.
39. WHITE, N.M. & R.J. MCDONALD. 2002. Multiple parallel memory systems in the brainof the rat. Neurobiol. Learn. Mem. 77: 125–184.
40. WISE, R.A. & M.A. BOZARTH. 1987. A psychomotor stimulant theory of addiction.Psychol. Rev. 94: 469–492.
41. WOOLVERTON, W.L., G.S. HECHT, G.E. AGOSTON, et al. 2001. Further studies of thereinforcing effects of benztropine analogs in rhesus monkeys. Psychopharmacology154: 375–382.
42. XU, F., R.R. GAINETDINOV & W.C. WETSEL. 2000. Mice lacking the norepinephrinetransporter are supersensitive to psychostimulants. Nat. Neurosci. 3: 465–471.
43. ZHOU, J. & L. IACOVITTI. 2000. Mechanisms governing the differentiation of a seroton-ergic phenotype in culture. Brain Res. 877: 37–46.