metabolite induction via microorganism co-culture: a potential way to enhance chemical diversity for...
TRANSCRIPT

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/260996012
Metaboliteinductionviamicroorganismco-culture:Apotentialwaytoenhancechemicaldiversityfordrugdiscovery
ARTICLEinBIOTECHNOLOGYADVANCES·NOVEMBER2014
ImpactFactor:9.02·DOI:10.1016/j.biotechadv.2014.03.001·Source:PubMed
CITATIONS
33
READS
775
6AUTHORS,INCLUDING:
SamuelBertrand
UniversityofNantes
71PUBLICATIONS176CITATIONS
SEEPROFILE
NadineBohni
UniversityofZurich
35PUBLICATIONS129CITATIONS
SEEPROFILE
SylvainSchnee
Agroscope
13PUBLICATIONS96CITATIONS
SEEPROFILE
KatiaGindro
Agroscope
94PUBLICATIONS1,065CITATIONS
SEEPROFILE
Availablefrom:SamuelBertrand
Retrievedon:04February2016

Biotechnology Advances xxx (2014) xxx–xxx
JBA-06793; No of Pages 25
Contents lists available at ScienceDirect
Biotechnology Advances
j ourna l homepage: www.e lsev ie r .com/ locate /b iotechadv
Research review paper
Metabolite induction via microorganism co-culture: A potential way toenhance chemical diversity for drug discovery
Samuel Bertrand a,b,1, Nadine Bohni a,1, Sylvain Schnee c, Olivier Schumpp c,Katia Gindro c, Jean-Luc Wolfender a,⁎a School of Pharmaceutical Sciences, EPGL, University of Geneva, University of Lausanne, quai Ernest-Ansermet 30, CH-1211 Geneva 4, Switzerlandb Groupe Mer, Molécules, Santé-EA 2160, Faculté des Sciences pharmaceutiques et biologiques, Université de Nantes, 9 rue Bias, BP 53508, F-44035 Nantes Cedex 01, Francec Mycology and Biotechnology group, Institute for Plant Production Sciences IPS, Agroscope, Route de Duillier 50, P.O. Box 1012, 1260 Nyon, Switzerland
Abbreviations: 1H-NMR, proton nuclear magnetic resodiscriminant analysis; DESI, desorption electrospray ionizaFDA, US Food and Drug Administration; FID, flame ionizatresolution; HTS, high-throughput screening; IMS, imagingspectrometry; MS/MS, tandem mass spectrometry; MSn, mNMR, nuclear magnetic resonance; NP, natural product; NPAIN, pan-assay interference; PCA, principal component aprojected orthogonalized chemical encounter monitoringphism; RI, refractive index; SAHA, suberoylanilide hydroxtotal ion chromatograms; TLC, thin layer chromatographyviolet; ZON, zearalenone.⁎ Corresponding author. Tel.: +41 22 379 33 85; fax: +
E-mail address: [email protected] (J.-L. Wo1 These authors contributed equally to this work.
http://dx.doi.org/10.1016/j.biotechadv.2014.03.0010734-9750/© 2014 Elsevier Inc. All rights reserved.
Please cite this article as: Bertrand S, et al, Mdrug discovery, Biotechnol Adv (2014), http
a b s t r a c t
a r t i c l e i n f oAvailable online xxxx
Keywords:Mixed fermentationCo-cultureNatural productsInterspecies communicationGene cluster activationMicroorganismsMass spectrometryMetabolomicsMicrobiomeAntimicrobials
Microorganisms have a long track record as important sources of novel bioactive natural products, particularly inthe field of drug discovery. While microbes have been shown to biosynthesize a wide array of molecules, recentadvances in genome sequencing have revealed that such organisms have the potential to yield evenmore struc-turally diverse secondary metabolites. Thus, many microbial gene clusters may be silent under standard labora-tory growth conditions. In the last ten years, several methods have been developed to aid in the activation ofthese cryptic biosynthetic pathways. In addition to the techniques that demand prior knowledge of the genomesequences of the studied microorganisms, several genome sequence-independent tools have been developed.One of these approaches is microorganism co-culture, involving the cultivation of two or more microorganismsin the same confined environment. Microorganism co-culture is inspired by the natural microbe communitiesthat are omnipresent in nature. Within these communities, microbes interact through signaling or defensemolecules. Such compounds, produced dynamically, are of potential interest as new leads for drug discovery. Mi-croorganism co-culture can be achieved in either solid or liquid media and has recently been used increasinglyextensively to study natural interactions and discover new bioactive metabolites. Because of the complexity ofmicrobial extracts, advanced analytical methods (e.g., mass spectrometry methods and metabolomics) are keyfor the successful detection and identification of co-culture-induced metabolites.This review focuses on co-culture studies that aim to increase the diversity of metabolites obtained from mi-crobes. The various strategies are summarized with a special emphasis on the multiple methods of performingco-culture experiments. The analytical approaches for studying these interaction phenomena are discussed,and the chemical diversity and biological activity observed among the induced metabolites are described.
© 2014 Elsevier Inc. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 02. Strategies for increasing the chemodiversity of microbes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
2.1. Accessing and altering the genes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
nance spectroscopy; 3D, three-dimensional; ANOVA, analysis of variance; APCI, atmospheric pressure chemical ionization; DA,tion; DI–MS, desorption ionization–MS; DNMT, DNAmethyltransferase; DON, deoxynivalenol; EI–MS, electron ionization–MS;ion detection; GC, gas chromatography; HDAC, histone deacetylase; HPLC, high performance liquid chromatography; HR, high-MS; LDPI, laser desorption postionization;MALDI, matrix-assisted laser desorption ionization;MF,molecular formula;MS,massultistage mass spectrometry;m/z, mass-to-charge ratio; NDF, non-dermatophyte filamentous fungus; NI, negative ionization;RPS, non-ribosomal peptide synthases; OPLS, orthogonal projection to latent structure; OSMAC, one strain-many compounds;nalysis; PCR, polymerase chain reaction; PI, positive ionization; PKS, polyketide synthase; PLS, partial least square; POChEMon,; qRT-PCR, quantitative real-time polymerase chain reaction; QS, quorum sensing; RFLP, restriction fragment length polymor-amic acid; SPME, solid phase microextraction; SUMO, small ubiquitin-related modifier; SUS plot, share and unique plot; TIC,; TRFLP, terminal restriction fragment length polymorphism; UHPLC, ultra high performance liquid chromatography; UV, ultra-
41 22 379 33 99.lfender).
etabolite induction via microorganism co-culture: A potential way to enhance chemical diversity for://dx.doi.org/10.1016/j.biotechadv.2014.03.001

2 S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
2.2. Interfering with transcription . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 02.3. Altering biosynthesis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 02.4. Modifying metabolite structures in extracts . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
3. Genome sequence-independent approaches for inducing chemodiversity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 03.1. Variation of culture conditions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 03.2. Mixed fermentation and solid medium co-culture . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
4. Microbial interactions in nature . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 04.1. Fungal communities . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 04.2. Bacterial communities . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 04.3. Bacterial–fungal communities . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
5. From natural communities to chemodiversity enhancement . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 05.1. Co-cultures on solid substrates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 05.2. Co-cultures in liquid substrates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 05.3. The induction of chemical diversity in microbes via co-culture . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 05.4. Advantages and drawbacks of solid medium co-culture over mixed fermentation . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
6. Monitoring co-culture metabolite induction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 06.1. Targeted and untargeted metabolite profiling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 06.2. Metabolite profiling and fingerprinting methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 06.3. Metabolomics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 06.4. Imaging mass spectrometry . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 06.5. Metabolite identification . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
7. Bioactivity of metabolites induced via co-culture . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 07.1. Type of induced metabolites . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 07.2. Biological activity of the induced metabolites . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 07.3. Chemical novelty among the induced metabolites . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
8. Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Appendix A. Supplementary data . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0References. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
1. Introduction
Natural products (NPs) are important sources of novel bioactivecompounds. Although many industries have ceased or significantly re-duced their NP drug discovery programs, NPs continue to be of interestto pharmaceutical companies (Newman and Cragg, 2012; Rosén et al.,2009). Indeed, nature provides a massive reservoir of organisms thatproduce potentially beneficial compounds to be discovered and ex-plored (bioprospecting) (Zhu et al., 2011). The inventiveness of natureregarding the production of innovative and unusual molecular skeletons(chemodiversity) is unmatched.
Nevertheless, because NPs are obtained from living species—as op-posed to via combinatorial synthesis—there are several aspects thatmake NP drug discoverymore complicated. For example, the organismsfromwhichNP extracts are to be obtainedmust be identified, and ques-tions concerning re-sourcing and intellectual property rights must beaddressed. The production of secondarymetabolites (metabolic expres-sion) in these living species depends on growth or culture conditions,which can be difficult to control. In addition, NP extracts are complexmixtures, which further complicates their assessment for bioactivitybecause the presence of overly toxic or PAIN (pan-assay interference)compounds may mask the effects of the sought-after compounds oractive constituents are present in too small a quantity. NPs or NP ex-tracts that are compatible with high-throughput assays can be obtainedthrough the enrichment or isolation of desirable compounds, but thesetasks can be labor-intensive and difficult. Furthermore, one of themajorchallenges in working with NPs is the rediscovery of previously identi-fiedmolecules. Avoiding rediscovery demands accurate chemical analy-sis of the extracts and detailed databases of all known compounds(dereplication (El-Elimat et al., 2013; Mitova et al., 2008; Nielsenet al., 2011; Wolfender et al., 2010)). In addition, NPs are often com-plex molecules and exhibit numerous stereocenters, making the elu-cidation of their structures challenging. Overall, drug discovery fromNPs is not considered to be highly compatible with high-throughputscreening (HTS) techniques or the time frame allocated for the
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
validation of a hit compound (Lam, 2007) demanded by industrialdrug discovery programs. Thus, in the industry, drug discoveryprograms mainly operate on the basis of NP-like compounds pro-duced through combinatorial synthesis (Feher and Schmidt, 2002;Pascolutti and Quinn, 2013).
However, in recent years, progress has beenmade on several fronts.These include improvements in chromatography techniques, whichallow accelerated isolation of molecules from complex mixtures(Bohni et al., 2013a; Bugni et al., 2008; Eldridge et al., 2002;Wolfender and Queiroz, 2012); in analytical techniques, notably in nu-clear magnetic resonance (NMR) detection, which now requires onlymicrogram amounts of compounds for identification (Dalisay et al.,2009; Gronquist et al., 2005; Hu et al., 2005); and in molecular biology,which provides for alternative routes to generate NPs (Brakhage andSchroeckh, 2011). Hence, these advances should facilitate the compati-bility of NPs with industry drug discovery programs.
Among the established sources of NPs, microorganisms have provento be promising candidates for the production of novel scaffolds as wellas marketable drugs (Cragg and Newman, 2013; Demain, 2014). Oneparticularly well-known example is provided by the β-lactam, penicillin(Fig. 1). Other examples of important leads isolated from microorgan-isms (Fig. 1) include the macrolides used as antibiotics (erythromycin),antifungals (amphotericin B) and immunosuppressive drugs (cyclospor-in) and depsipeptides with antibiotic activity (fusafungin). Furthermore,the cholesterol-lowering agents of the statin class (e.g., lovastatin) werethe fourth bestselling group of pharmaceuticals in 2013 in the US(Drugs.com, 2013) and were the bestselling NP-derived drugs. More-over, microorganisms are also known to produce particularly innovativescaffolds, such as pleurotine (Grandjean and Huls, 1974), enediyne anti-biotics (Smith and Nicolaou, 1996) or diketopiperazines (Guo et al.,2009).
Bacteria and fungi are therefore important sources of drugs and leadcompounds (Berdy, 2012; Cragg and Newman, 2013; Pearce et al.,2009). For example, 33% of all natural product-related antitumordrugs approved by the US Food and Drug Administration (FDA) by
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

HN
ON
O
SH
OHOpenicillin G
Penicillium sp.
O
OHHO
O
O
C2H5
O
O
OH
O
NHO
O OH
OCH3
erythromycin AStreptomyces erythreus
amphotericin BStreptomyces nodosum
O
O OH OH
OH
OH OH
OH
O
OH
O
OH
O
HO
OOH
OHH2N
NN
HN
OO
O
N O
O
HN
O
NH
N
O
N
O
NO
O
N
HO
NH
O
ciclosporin ATolypocladium inflatum
fusafungin (depsipeptide)Fusarium lateritium
NO
O
O
O
N O
O
ON
O
O
O
HO O
HO
O
lovastatinAspergillus terreus
O
O O
O
O
H
H
H
pleurotinePleurotus griseus
dynemicin A (enediyne)Micromonospora chersina
glionitrin A (diketopiperazine)Aspergillus fumigatus
OHNO
OOH
OH
OH
O
O
OH
N
N
O2N
SS
O OH
O
Fig. 1.Microbial natural products. Molecules of pharmaceutical interest or exhibiting particular structural diversity from microbial sources.
3S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
2012 were of microbial origin or were derivatives of microbial metabo-lites (Giddings and Newman, 2013). Compounds from microorganismsand marine organisms constituted the largest group of promising anti-cancer drugs among the compounds evaluated in oncology clinical trialsas of 2004 (Butler, 2005). Compared to other natural resources, mi-crobes are presently themost attractive source of NPs in drug discovery(Lam, 2007), mainly because of their ubiquitous occurrence (Bull andStach, 2007; Gunatilaka, 2006; Wilson and Brimble, 2009), their exten-sive biodiversity and the large chemodiversity that can be foundwithina given species (Akondi and Lakshmi, 2013; Wilson and Brimble, 2009;Zhu et al., 2011). To further broaden the diversity of species to bestudied, methods have been developed to utilize thus far uncultivatablemicroorganisms (Lefevre et al., 2008; Piel, 2011).
Another key feature of these organisms is their ability to be culturedin the laboratory and the fact that the production of NPs can be opti-mized and scaled up as required. For microorganisms, culturing can beperformed in liquid or solid media (Adelin et al., 2011) or in fermenters(Omura, 1992). The culturing conditions are substantially controlled bythe composition of the culture medium (Creek et al., 2013; Kaeberleinet al., 2002).
With the development of molecular biology techniques, microor-ganisms can also be engineered to produce specific compounds. The de-siredmolecules are then obtained through heterologous expression in ahost organism to circumvent culturing problems or to access otherwisesilent gene clusters (Gomez-Escribano and Bibb, 2014; Schneider et al.,2008). Particular efforts were made to develop additional methods forthe activation of silent gene clusters (Brakhage and Schroeckh, 2011;
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
Chiang et al., 2009, 2011; Marmann et al., 2014; Ochi and Hosaka,2012; Scherlach and Hertweck, 2009). A method has been developedto screen bacterial DNA for preserved gene regions that code for differ-ent classes of polyketides, non-ribosomal peptides or diterpenes. Thismethod is complementary to the other existing methods and allowsfor the selection and prioritization of promising producers (Xie et al.,2014). Moreover, methods combining genomics and chemistry havebeen established that allow the simultaneous identification of novelbioactive compounds and their respective mechanisms of action(Ho et al., 2011). All of these advances have led to a favorable situa-tion, in both academia and the pharmaceutical industry, in whichthe use of NP libraries continues “to cataly[z]e innovation in biomed-ical research” (Lam, 2007; Pevarello, 2009; Roemer et al., 2011;Schmitt et al., 2011).
Among these general considerations regarding the usefulness of mi-croorganisms in NP research, a core area of interest is the possibility ofgenerating new scaffolds to discover novel bioactive compounds. In ad-dition to the bioprospecting of species not previously studied, whichcarries a high risk of redundancy, one strategy consists of exploiting bio-synthetic pathways that are not triggered under normal laboratorygrowth conditions in known producers.
There are various methods for inducing chemodiversity in selectedorganisms at different levels, ranging from the genome to the metabo-lome (Fig. 2). In addition, the chemical–ecological relationships thatoccur in microorganism communities (interspecies interactions) canbe exploited. In this respect, the novel approach in which microbesare grown together (co-culture or confrontation experiments) has
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

mutasynthesis
Genome Transcriptome Proteome Metabolome
heterologousexpression
metabolicengineering
co-culture (solid medium co-culture and mixed fermentation)
OSMAC
substrate feeding
semi-synthesis
chemodiversityenhancement
precursor-directed biosynthesis
epigenetic modification
enzymeinhibition
Fig. 2.Methods to influence secondary metabolite biosynthesis in microorganisms. Possible methods for influencing the production of secondary metabolites in microorganisms or aug-menting their chemodiversity. The upper methods (blue and green backgrounds) demand prior knowledge of the genome sequence. Abbreviation: OSMAC, one strain-many compoundsapproach. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
4 S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
received increasing interest related to the potential discovery of newleads and to understand the triggering of specific biosynthetic path-ways, mainly related to defense.
This review assembles various historical examples of microbial co-culture as well as recent applications that highlight the use of suchmethods from the perspective of increasing microbial chemodiversity.A general introduction that summarizes ways of modeling chemo-diversity is provided, with a particular focus on co-culture in solid andliquid media. The different analytical approaches that are used tostudy the microbial metabolome and to highlight novel metabolite in-duction in co-cultures are then described. Finally, the challenges andperspectives arising from this new field of research are discussed.
2. Strategies for increasing the chemodiversity of microbes
Based on the genome sequences of some fungi (e.g., (Galagan et al.,2005)), it is estimated that microorganisms are capable of producingmany more compounds than are observed in the analysis of conven-tional in vitro cultures (Nett et al., 2009). Thus, many biosyntheticgene clusters are presumably silent under standard laboratoryconditions (Wiemann andKeller, 2014). Severalmethods have beende-veloped to activate these cryptic gene clusters or, more generally, toaugment the chemodiversity of which the microorganisms are capable(Table 1). This can be achieved at the genome, transcriptome, proteomeor metabolome levels (Fig. 2), for which the methods are briefly de-scribed below and have been extensively reviewed (e.g.,Cichewicz,2010; Goss et al., 2012; Ochi and Hosaka, 2012; Scherlach andHertweck, 2009; Winter et al., 2011). Section 3 provides further detailson methods that involve varying the culture conditions or co-culturingmultiple organisms to modulate the chemical composition of microbialextracts.
2.1. Accessing and altering the genes
Since the genome sequences of several fungi and bacteria have beenmade available, influencing the biosynthesis of secondary metabolitesthrough the manipulation of genome sequences has become possible(Brakhage and Schroeckh, 2011; Scherlach and Hertweck, 2009;
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
Winter et al., 2011). The methods used for this purpose—summarizedas metabolic engineering—include various methods for gene knockout,promoter exchange and overexpression of transcription factors(Bergmann et al., 2007; Bok et al., 2006). NPs that are generated fromthese modified gene clusters are then obtained through either heterol-ogous expression, where the gene cluster is inserted into a differentmi-croorganism in which culturing is straightforward, i.e., a host organismsuch as Escherichia coli, or homologous expression,where the gene clus-ter is back-inserted into the original microorganism. Alternatively, themicroorganism's genome can be altered to induce greater artificialchemodiversity (Goss et al., 2012) via mutasynthesis. In this technique,the genes responsible for theuptake of the starting substrate of ametab-olite are disabled, and the starting block of choice is fed to the microor-ganism (combinatorial biosynthesis). Thus, an altered NP is generatedusing the starting block provided to the organism. Mutasynthesisdemands substrate flexibility concerning these unnatural substrates.All of these molecular methods require that the biosynthetic gene clus-ters are sequenced and that putative functions are assigned (throughwhole-genome sequencing or via genome mining (Bergmann et al.,2007)). An alternative approach is randomor ultraviolet (UV)mutagen-esis, which has been successfully applied to generate antibacterialcompounds from Aspergillus oryzae (Leonard et al., 2013) and a novelmacrolide from a Sphaeropsidales sp. mutant (Bode et al., 2000a).
Another field that seems promising for increasing and accessing thechemodiversity of microorganisms is metagenomics (Lefevre et al.,2008). Under this approach, DNA from complexmicrobial communities(e.g., soil or gut microbiomes) is extracted and cloned in laboratoryhosts to create a DNA library. The DNA from these metagenomic librar-ies is then purified and fractionated for direct recombinant expressionin a host. The generated clones are either tested directly for the produc-tion of bioactive compounds in a particular assay or themetabolites areextracted and tested through a HTS approach (Lefevre et al., 2008; Piel,2011).
2.2. Interfering with transcription
Inside eukaryotic cells, the genome is packaged into chromatin withhistone proteins, and this structure exerts profound control over gene
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

Table 1Methods to increase chemodiversity in microorganisms through manipulation or activation of biosynthetic genes.Several facets of the usability of the presented methods are given. The general prerequisites, whether the microorganisms have to be cultivatable (cultivation-dependence) and whetherthe genome-sequence has to be known (genetic information) are indicated. Another aspect is whether themethod permits the activation of silent biosynthetic genes (activation of silentgenes). Then, an estimation is given on whether the method potentially allows to generate larger quantities of compound (upscalability) and whether the method is capable of possiblyyielding novel structures (generation of novel structures). Abbreviations: NP, natural product; OSMAC, one-strain-many-compounds; ⊠, Yes; □, No.
Method Prerequisite Activationofsilent genes
Upscalability Generation ofnovelstructures
Referencesa
Cultivation-dependence
Geneticinformation
Mutasynthesis ⊠ ⊠ ⊠ ⊠ ⊠ Goss et al. (2012), Kirschning and Hahn (2012)Heterologous expression □ ⊠ ⊠ ⊠ ⊠ Gomez-Escribano and Bibb (2014), Scherlach and Hertweck (2009)Metabolic engineering ⊠ ⊠ ⊠ ⊠ ⊠ Kirschning and Hahn (2012); Scherlach and Hertweck (2009)Enzyme inhibition ⊠ ⊠ ⊠ ⊠ ⊠ Bode and Zeeck (2000b), Craney et al. (2012), Scherlach and Hertweck
(2009)Precursor-directedbiosynthesis
⊠ ⊠ □ ⊠ □ Goss et al. (2012)
Substrate feeding ⊠ ⊠ □ ⊠ □ Goss et al. (2012), Kirschning and Hahn (2012)Co-culture, liquidmediumb
⊠ □ ⊠ ⊠ ⊠ Pettit (2009), Scherlach and Hertweck (2009)
Co-culture, solid mediumb ⊠ □ ⊠ □c ⊠ Bertrand et al. (2013a), Yang et al. (2009)OSMAC ⊠ □ ⊠ ⊠ ⊠ Bode et al. (2002), Iwai and Omura (1982), Omura (1992),
Scherlach and Hertweck (2009), Takahashi et al. (2013)Epigenetic modificationd ⊠ □ ⊠ ⊠ ⊠ Bok et al. (2009), Cichewicz (2010), Scherlach and Hertweck (2009)
a Review articles describing the different methods from the viewpoint of NP and drug discovery (unless not available, then, original articles are given).b Involved microorganisms must tolerate same culture condition.c A large-scale solid-state fermenter has been developed recently (Adelin et al., 2011). The applicability to solid-media co-cultures remains to be proven.d Has not been tried on bacteria yet.
publication year1950 1960 1970 1980 1990 2000 2010
num
ber
of p
ublic
atio
ns
0
10
20
30
40
50
60
70
Fig. 3.Number of publications per year in the field of microorganism co-culture. Evolutionof research efforts in the field of microorganism co-culture, as indicated by the number ofpublications per year from 1950 to 2013. Journal articles, reviews, conference proceedingsand book chapters with the following keywords were searched in theWeb of Knowledge(Thomson Reuters): microorganism co-culture, mixed fermentation, coculture andcombined-culture.
5S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
transcription (Felsenfeld and Groudine, 2003). The chromatin structureis closely associated with the epigenetic state of histones, alkaline pro-teins that package and order the DNA into nucleosomes. Epigenetic his-tone modifications include the production of acetylated, methylated orubiquitinated lysines, methylated arginines and phosphorylatedserines. Together with DNA methylation, which also occurs in non-eukaryotes, the modulation of chromatin structure has major effectson the overall expression of genes and thus subsequent metabolitebiosynthesis.
For example, a deficiency in the methylation of specific histonespermits the induction of silent gene clusters in Aspergillus nidulans(Bok et al., 2009). This gives rise to the possibility of using epigeneticmodifiers to induce the transcription of otherwise silent gene clusters(Williams et al., 2008; X.-L. Yang et al., 2013). Such epigenetic manipu-lation can be achieved using small molecules, such as 5-azacytidine,suberoylanilide hydroxamic acid (SAHA) or nicotinamide, that inhibiteither DNA methyltransferase (DNMT) or histone deacetylase (HDAC).This technique has been applied in several fungi (Cichewicz, 2010).
Additionally, global manipulation of transcription factors allows thesimultaneous targeting of the expression of several secondary metabo-lite gene clusters. This approach has been demonstrated in Aspergillusspp., in which the overexpression of laeA, a protein with homology tomethyltransferase, activated or enhanced the production of severalknown bioactive compounds (non-ribosomal peptides, polyketides,terpenes, etc.) (Bok and Keller, 2004).
Because epigenetic modulation affects the transcription ofmany dif-ferent gene clusters, interference at this level can have negative effectson the survival of the targeted microorganism. Indeed, reduced growthhas been observed under the influence of epigeneticmodifiers, and suchan approach can currently be considered untargeted with an unpredict-able likelihood of success (Williams et al., 2008). The diversity ofepigenome-related targets among fungi is large, and not all fungalstrains respond to currently known epigenetic modifiers (Cichewicz,2010).
2.3. Altering biosynthesis
The addition of enzyme inhibitors to the culture medium can blockcertain biosynthetic pathways, thus shifting secondary metabolite bio-synthesis towards the production of other NPs from either silent or
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
poorly expressed gene clusters (Bode and Zeeck, 2000b; Craney et al.,2012). Enzyme inhibition may cause both the elicitation and inhibitionof NP biosynthesis. For example, tricyclazole inhibits the production of1,8-dihydroxynaphthalene in Sphaeropsidales sp. and simultaneouslyinduces the production of sphaerolone and dihydrosphaerolone (Bodeand Zeeck, 2000a; Bode et al., 2000b).
One possible target for secondary metabolite induction is theSUMOylation of proteins (SUMO: small ubiquitin-related modifier),which is involved in transcriptional regulation by post-translationalmodification of the proteins. A study conducted in anA. nidulansmutantincapable of protein SUMOylation showed altered NP formation com-pared to the intact fungus (Szewczyk et al., 2008). Various inhibitorsof SUMOylation have been discovered (Fukuda et al., 2009; Hirohamaet al., 2013; Takemoto et al., 2014), but their utility in the induction ofnovel NPs remains to be proven.
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

6 S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
As discussed previously (see Section 2.1), metabolomic modifica-tions can be performed upstream in biosynthetic pathways by feedingthe microorganisms unnatural substrates (precursor-directed biosyn-thesis (Goss et al., 2012)). This approach requires substrate flexibilityof the enzymes involved, and the corresponding biosynthetic origin ofthe targeted NPmust be known. This method presents a high likelihoodof leading to pharmaceutically relevant compounds because functional-ities leading to drug-like properties can specifically be introduced intothe modified molecules. This approach is useful for optimizing particu-lar scaffolds to increase bioactivity or reduce toxicity.
Overall, the use of enzyme inhibitors/inducers, similar to the use ofepigenetic modifiers, mainly relies on serendipity, except when thecomplete regulation of a gene cluster has previously been described.
2.4. Modifying metabolite structures in extracts
Enhancing the chemodiversity of metabolites is also possiblethrough chemical modification of their structures. Classically, minorskeleton modifications are attained through various methods of deriva-tization, and major alterations can be achieved using semi-synthesis onisolated microbial metabolites (Goss et al., 2012; Kirschning and Hahn,2012). Alternatively, chemical modifications can be performed directlyat the metabolome level (on the crude extract), without prior isolationof individual compounds (chemodiversity enhancement). This strategywas successfully applied to modify β-caryophyllene and flavonoids(Krchňák et al., 2011; López et al., 2007).
Again, the chance of obtaining pharmaceutically relevant com-pounds is increased, as drug-like properties can be specifically intro-duced into the modified molecules to optimize the bioactivity andsafety profile of an NP.
3. Genome sequence-independent approaches forinducing chemodiversity
The production of secondary metabolites by microorganisms isstrongly dependent on environmental factors, such as growth condi-tions and biotic and abiotic stresses (Bode et al., 2002; Iwai andOmura, 1982; Omura, 1992). Thus, the selective variation of the cultureconditions (mainly abiotic) for a given microbe (Bode et al., 2002;Takahashi et al., 2013) and/or stress induction through interactionwith other competing microorganisms in a co-culture (Frey-Klettet al., 2011; Pettit, 2009; Scherlach et al., 2013; Shank and Kolter,2009) represent interesting ways to generate enhanced chemo-diversity. Such approaches can affect various levels of the cellular ma-chinery (genome, transcriptome, proteome or metabolome).
Because genomic information is only available for a limited numberof microorganisms, precluding the use of sophisticatedmolecular biolo-gy techniques, multiple other methods have been developed to over-come the difficulty of NP production under certain culture conditions.These approaches have been inspired by various examples in nature,where microbial communities (the so-called microbiome) are omni-present (see Section 4).
3.1. Variation of culture conditions
Culture conditions affect the metabolite profiles of microorganisms,which has led researchers to perform experiments with different cul-ture media to optimize metabolite production. This method is knownas the OSMAC (one strain-many compounds) approach (Bode et al.,2002), in which culture media and nutrients are systematically variedto simulate different environments and consequently induce the pro-duction of different metabolites. The parameters that are varied includelight (Tisch and Schmoll, 2010), pH, temperature and the oxygen sup-ply. The media are varied in terms of carbon, nitrogen and phosphorussources, inorganic salts or trace metal contents (Iwai and Omura,1982). The culture medium can also be supplemented with other
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
small molecules or heavy metals that function as growth inducers orsuppressors (as detailed above, e.g., for HDAC or enzyme inhibitors) orthrough unknown mechanisms (e.g., when using the organic solventsdimethylsulfoxide and ethanol (Pettit, 2011)).
In some cases, these modifications influence transcription factors(Keller et al., 2005 and references therein; Rigali et al., 2008) and canbe considered a form of epigenetic modulation.
Applications of the OSMAC approach have proven to be successfulfor the production of drug-like compounds and novel metabolites(Bode and Zeeck, 2000a; Bode et al., 2002; Masuma et al., 1986).
3.2. Mixed fermentation and solid medium co-culture
Another way to stimulate the biosynthesis of secondary metabolitesis to challenge a given microorganism with biotic stress, which can beinduced through interaction with other microbial partners.
Indeed, in nature, microorganisms are often found in communities(see Section 4), where they produce secondary metabolites related tothe interaction type involved (sexualization, growth inhibition orstimulation), mainly in the context of defense or nutrient competition.Such situations can be reproduced artificially by culturing two or moremicroorganisms together (De Roy et al., 2013). This strategy has oftenbeen defined as ‘co-culture’ when solid media are used or ‘mixedfermentation’ when liquid media are involved. These interactionsmight either mimic naturally occurring communities (mainly in theframework of chemical ecology studies) or constitute an artificial com-munity for studying NP induction specifically in confrontation zones(mainly to discover new bioactive compounds). All of these aspectswill be discussed in the second part of this review.
The concept of co-culture has generated increasing interest from thescientific community, and as shown in Fig. 3, the number of applicationsof this strategy has increased greatly over the last decade. Although themorphology ofmicroorganismal interactions has been of interest tomi-crobiologists for years, the resurgence of interest in this field, especiallyconcerning biochemical aspects, is most likely related to the technolog-ical advances in the analysis of such complex biological systems (seeSection 6).
The co-culture approach has been applied to study diverse issues,such as i) in the fundamental investigation of natural communities inan agricultural context (the rhizosphere or mycorrhizosphere)(Bonfante and Anca, 2009); ii) to elucidate symbiosis phenomena(e.g., the antibiotic protection of a coral by its symbiont (Moree et al.,2013)); iii) to investigate humanmicrobiome interactions (e.g., positiveinteractions leading to a ‘healthy situation’) (Cui et al., 2013; Ghannoumet al., 2010;Huffnagle andNoverr, 2013; Ridaura et al., 2013; Turnbaughet al., 2006); iv) for the induction of pharmaceutically interesting sec-ondary metabolites (Bertrand et al., 2013d; König et al., 2013;Schroeckh et al., 2009), such as glionitrin A (Park et al., 2009); andv) in targeted applications to improve the production yields of specificfermentation products (e.g., vitamin C synthesis (Du et al., 2012)).
4. Microbial interactions in nature
As mentioned above, microbial interactions are ubiquitous and canbe found in every biocoenosis. Soil (Ross-Davis et al., 2013) and aquaticor terrestrial organisms, such as plants (Gaiero et al., 2013; Hyde andSoytong, 2008; Oldroyd, 2013), animals (Douglas, 2013; Misra, 1998;Ross et al., 2013), fungi (Kiss et al., 2004; Lawrey, 1995) and protists(Wodarz, 2013), generally host a set of microbes sharing specific eco-logical niches. Thesemicroorganisms include the Prokaryotes (bacteria,archaea) and Eukaryotes (protists, fungi) as well as viruses. They growand survive in these environments as long as the nutrient sources aresufficient, either inmutualistic relationships or as antagonists. Classically,studies on the biodiversity of microorganisms in a given environmenthave only been possible in specific cultures using adapted culturemedia. However, the approach is limited to several groups of
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

7S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
microorganisms, including some fungi and bacteria. Some microorgan-isms are impossible to cultivate artificially because of their dependenceon microbe–microbe interactions for development (Beck et al., 2012).Others are difficult to detect due to their lack of competitiveness,which leads to growth inhibition in the presence of another microorgan-ism. The recent development of methods that are totally independent ofclassical cultures, such as polymerase chain reaction (PCR) (Sachse,2004; vanBelkumet al., 1998), restriction fragment lengthpolymorphism(RFLP) (Vaneechoutte, 1996), pyrosequencing (Petrosino et al., 2009)and, more recently, metagenomics (Miller et al., 2013; Ross et al., 2013),metatranscriptomics (McGrath et al., 2010) and other ‘meta'omics’methods (Lepage et al., 2013), represent newways to study complex eco-systems and their interactions (Segata et al., 2013). As a result, fungal andbacterial communities have been discovered in substrates that were pre-viously considered to be nearly devoid of microorganisms, further em-phasizing the importance of studying communities (Delhaes et al., 2012).
These complex fungal or bacterial communities have been definedas ‘microbiomes.’ This term includes all of the microbes (bacteria,archaea, viruses, protists and fungi) living in a particular environmentor host (Methe et al., 2012), and this concept has received strong inter-est, especially in humans (e.g., gut or lung microbiomes) (Huffnagleand Noverr, 2013). In 2010, the concept of a mycobiome emerged(Ghannoum et al., 2010), referring to the fungal biota in a particular en-vironment (fungal microbiome).
The coexistence of several microorganisms that share the sameniche can affect the organisms' growth, adaptation patterns, morpholo-gy and developmental patterns (Rico-Gray, 2001; Sandland et al., 2007),aswell as their ability to synthesize proteins and secondarymetabolites.Thus,microbial communities have amajor influence on their immediateenvironment or host. For example, in soy sauce production, the compo-sition of the microbial community and the dynamic changes it un-dergoes during fermentation have been found to be crucial to thequality of the soy sauce (Wei et al., 2013).
Additionally, the level of microbiome organization (e.g., in biofilms)confers advantages to a community, e.g., by increasing resistance orvirulence (Sandland et al., 2007).
Historically, microbial communities have mainly been considered tostudy antagonistic growth-inhibiting interactions. These interactionshave been widely detailed in wood-inhabiting fungal communities,bacterial soil communities (An et al., 2013; Sullivan et al., 2013) andspecific bacterial–fungal interactions related to the synthesis of antibi-otics (Park et al., 2009). The best-known example of such bacterial–fungal interactions is the accidental discovery of penicillin in an unintend-ed co-culture (contamination) of Staphylococcus spp. with Penicillium sp.in 1928 by Sir Alexander Fleming (Fleming, 1929). Recent research hasdemonstrated that the interaction between each partner is, amongother factors, mediated by the expression of small molecules (Shankand Kolter, 2009).
4.1. Fungal communities
An important phenomenon in triggering the production of second-ary metabolites is the interaction between fungi (Keller et al., 2005)or between fungi and bacteria (Scherlach and Hertweck, 2009). Thecompetition for nutrients and space leads fungi to synthesize enzymesand secondary metabolites to enhance their own growth. Fungal colo-nies develop as multicellular filaments (hyphae) that form aninterconnecting network (mycelium) (Donnelly and Boddy, 2001).Two mycelia in close proximity to each other can interact in differentways, specifically inmutualistic, neutralistic or competitive interactions,and can even switch from one interaction type to another (Rayner,1988). In confined spaces, the fungi are restricted to only one resourceunit; i.e., both nutrients and space are very limited. There are twotypes of competitive mycelial interactions: either one fungus inhibitsother individuals or one fungus uses a nutrient/space resource, whichis consequently not available to another fungus, a process referred to
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
as exploitation competition. These types of antagonistic interactionsoccur either at a distance or upon/after physical contact between indi-vidual hyphae or mycelial networks (Falconer et al., 2008). This impliesthat active recognition of the presence of other mycelia occurs and ini-tiates a combative response. However, the biological mechanismslinked to this recognition are not clear (Peiris et al., 2008). Reactivityof mycelia at a distance would require the release and recognition ofboth water-soluble and volatile diffusible compounds, such as in therecognition of sexual partners with trisporic acids in the fungal orderMucorales (Schachtschabel et al., 2008). However, in some cases,contact between individual hyphae must occur before a response isstimulated, sometimes followed by cell death at one or both mycelialfronts (Ikediugwu and Webster, 1970a, 1970b).
The changes that occur during interactions can include the produc-tion of extracellular secondary metabolites, notably phenolic andquinonic compounds (Griffith et al., 1994). A number of such metabo-lites have been identified during the past decade in interactionsamong the mycelia of wood-decaying fungi. In addition to changes inthe metabolome, it has been shown that production of differentenzymes can be induced during these mycelial interactions. For exam-ple, in wood, the production of enzymes such as phenoloxidases andperoxidases (Score et al., 1997), including laccase (Wei et al., 2010)and manganese-dependent peroxidase (White and Boddy, 1992), aswell as different lignin-degrading enzymes (Chi et al., 2007), can beinduced.
Wood also represents a good context in which to study fungus–fungus interactions in a confined space (Rayner and Todd, 1980). Thishighly complex substrate shelters a dynamic fungal community, includ-ing numerous fungal species occurring in succession (Casieri et al.,2009; Frankland, 1998; Hofstetter et al., 2012). Competitive mycelialinteractions are very important in the overall development of fungalcommunities in wood (Boddy, 2000; Heilmann-Clausen and Boddy,2005).
It has been shown that the secondary metabolites of some wood-decaying fungi can act as total inhibitors or stimulators of the growthof other fungal protagonists (Boddy and Rayner, 1983). In the case ofcompetitive interactions, competing fungi can form zone lines (alsodefined as confrontation or barrage zones) that are morphologically dif-ferent from the pure cultures (Campbell, 1932). The strong coloration ofsuch zones denotes significant metabolic activity, which can potentiallybe exploited to search for novel metabolites (Bohni et al., 2013b; Peiriset al., 2008). A number of such metabolites involved in interactions be-tween the mycelia of wood-decaying fungi (e.g., volatile compounds,such as acetone and 2-methyl-1-butanol, or quinones, such aspodosporin A) have been identified during the past decade (Gloer,1995; Humphris et al., 2001; Shearer, 1995; Wheatley, 2002). However,very little attention has been paid to competition between fungi to pro-duce phytotoxic or antifungal substances. Similarly, few studies discussthe exploitation of such phenomena to detect disease markers. Recently,a studyusing the fungi implicated in esca, an apoplectic disease of grapes,demonstrated the complexity and richness of such zone lines. Transposi-tion of parts of the fungal community onto an artificial medium permit-ted the identification of melleins with fungitoxic, phytotoxic andbactericidal activity (Glauser et al., 2009). Solid medium co-culture hasbeen applied based on the approach detailed in Section 5,which illustrat-ed its potential for the discovery of new compounds with such activities.
Other ecological niches where fungi live in a confined space includehuman finger- and toenails, where opportunistic fungi may interactwith the dermatophytes that are involved in superficial mycoses(Monod et al., 2006; Verrier et al., 2012). The presence of non-dermatophyte filamentous fungi (NDF) and dermatophytes that may actas the etiological agents of onychomycoses (Summerbell et al., 2005)has been recently demonstrated via PCR (Verrier et al., 2012). The processthat governs these fungal interactions is not yet known, but artificial co-culturing of hospital isolates has revealed the induction of quinones andother secondary metabolites (Bertrand et al., 2012; Bohni et al., 2013b).
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

8 S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
Other mycobiomes that are now well studied with respect to theprogression of fungal or fungus-associated human pathologies (Cuiet al., 2013; Verrier et al., 2013) include those of the oral and nasal cav-ities, and the lungs, gut or skin (Ghannoumet al., 2010). Cui et al. (2013)demonstrated that specific mycobiomes are associated with differentdiseases and that these communities contribute to disease through in-teractions within fungi and those between the whole mycobiome andthe host.
4.2. Bacterial communities
Bacteria often organize into multicellular populations, mainly asbiofilms, to colonize an ecological niche (Romling, 2013). One or severalbacterial species interact closely and evolve in communities to exploitlimited resources in a confined environment to ensure species survivaland procure advantages such as access to nutrients, dynamic growthor increased antibiotic resistance (Percival and Bowler, 2004). Chemicalcommunication among the bacterial population appears to be crucial forthe integration of new species and the formation of the so-called ‘climaxcommunity’, where the protagonists co-exist in a balanced and stablemanner as a function of the surroundingmicro-environment. For exam-ple, the plasticity of cyanobacterial communities permits them to sur-vive in a wide range of salinities (Junier et al., 2013), and conversely,environmental modifications evoke dynamic changes in the composi-tion and ratio of bacteria within the communities (Lawrence et al.,2012). In Drosophila melanogaster, it has been shown that the dietplays a crucial role in influencing the bacterial microbiome of the diges-tive tract (Chandler et al., 2011). The microbiome composition evolvesin relation to host physiology or according to modification of the sub-strate over different time scales, as shown, for example, in human ca-davers (Hyde et al., 2013). Similarly, changes in coral physiology dueto environmental factors impact the diversity of the mucus-associatedbacterial communities (Kooperman et al., 2007).
4.3. Bacterial–fungal communities
Mixed populations of bacteria and fungi occur in a wide variety ofecological niches and are conditioned by the access to nutrients andsubsequent colonization of the substrates. The composition of suchcomplex communities is directly linked to specific conditions, such asa particular health or disease state in the case of human microbiomes(Jenkinson and Lamont, 2005).
Rich microbial diversity is also observed in soil. One gram of soil cancontain up to ten billionmicroorganisms, likely including between 1000and 10,000 species of unknown prokaryotes (Rosselló-Mora andAmann, 2001). The interactive relationships and combinations of indi-viduals and communities are therefore essentially infinite. In such com-munities, complex mutualistic interactions exist, including the use offungal hyphae by bacterial cells for protection against soil modificationsor for access to nutrients. Some of these interactions are key factors, asexemplified by the oxalate–carbonate pathway in soil, which implies aspecific interaction of fungi and bacteria (Martin et al., 2012).
All these examples of specific natural microbial communities dem-onstrate the importance of the cohabitation abilities of the implicatedspecies, allowing them to reach an equilibrium in a given ecologicalniche. These microbiomes are governed by complex multidimensionalinteractions, among which chemical communication and secondarymetabolite induction represent promising sources of chemical diversity.These communities are worth being studied in depth, both to under-stand the key regulation phenomena and to assess the bioactivity ofthe identified molecules.
5. From natural communities to chemodiversity enhancement
Numerous natural interactions between microbes can be exploitedto mimic confrontations under artificial conditions. From a biochemical
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
perspective, all of these interaction phenomena are characterized by theinduction of bioactive defense metabolites or toxins, such as phyto-alexins in plants (Ahuja et al., 2012) or mycoalexins in fungi (Glauseret al., 2009). Thus, accurate knowledge of the secondary metabolitecomposition of the organisms entering into the competition is requiredfor an in-depth investigation of these interactions. Sophisticated analyt-ical methods are often required to detect prominent as well as subtlebut significant metabolome variations (detailed in Section 6).
Biotechnology research has attempted to mimic culture conditionsin vitro for selected microbes to generate natural microbiota, producehigh-value compounds and increase the productivity of particularstrains. The transfer of a species from its natural habitat to an artificialenvironment must be carried out via successive improvement steps,based on a good understanding of the growthmechanisms of the targetmicroorganisms. This is important, for example, because the productionof secondarymetabolites is associatedwith the formation of the asexualreproduction organs in fungi (Prosser and Tough, 1991). The coloniza-tion of the medium is dependent on the type of cultured microorgan-ism: filamentous fungi ensure their growth through the extension ofhyphae producing lateral or apical branches, leading to the formationof macroscopic mycelia, whereas bacterial cells attach to a surface andcan organize themselves as a biofilm, embedded in an extracellular ma-trix of polymeric substances (mainly polysaccharides) (Donlan, 2002).
However, microorganismal morphology can be modified in relationto the substrate type and culture conditions. For example, in mostcases, the development of a fungal species on a solidmedium leads to cir-cular colonies with characteristic growthmargins, whereas in liquidme-dium, the same species could develop as plates, crusts or ‘pellets’ ofmycelial aggregations. Therefore, upscaling microbial cultures for thebiosynthesis of secondary metabolites is strongly dependent on thetype of substrate involved and on several qualitative biological parame-ters, such as the carbohydrate and nitrogen sources and macro- and mi-croelement availability, as well as engineering constraints (temperature,hygrometry, light, oxygen supply) (Falconer et al., 2008). Complex mul-tidimensional interactions cannot be studied directly because the contri-butions from the different individuals are difficult to isolate. One strategyconsists of studying the interaction between two representative partnersunder controlled conditions. For this purpose,microbes can be either cul-tivated on solid or in liquid media, but the culture conditions must becompatible for both partners.
5.1. Co-cultures on solid substrates
Fungi are well adapted to develop on solid media because they usehyphal growth to colonize unexplored regions containing nutrients(Prosser and Tough, 1991). The growing fungal tip is the center of in-tense metabolic activity, mainly to ensure hyphal extension.
Morphologically, the interaction of two fungi can lead to four major‘interaction types’ (distance inhibition, zone lines, contact inhibitionand overgrowth), as defined through the observation of numerous co-culture experiments performed on solid agar plates (Fig. 4) (Bertrandet al., 2013c). This type of behavior can also be observed in natural hab-itats, such as the colored zone lines in wood (Boddy, 2000).
Historically, the first in vitro co-cultures of fungi were performed onsolid media. This culturing format has permitted the study of morpho-genetic and metabolic changes that occur at the mycelial front(Sonnenbichler et al., 1989) as well as interaction patterns (Bertrandet al., 2013c, 2013d; Boddy, 2000; Woodward and Boddy, 2008).Many additional studies have been accomplished based on the confron-tation of fungi, bacteria and protists with each other (Cheng et al., 2013;Martin et al., 2012; Ola et al., 2013; Schoeman et al., 1996;Sonnenbichler et al., 1989), including studies of fungus–bacterium in-teractions (Mela et al., 2011) or fungus–fungus interactions (Melaet al., 2011; Wald et al., 2004; Woodward and Boddy, 2008; Yuenet al., 1999). To investigate the metabolic processes related to mycelialinteraction and their relevance for industrial purposes, sets of various
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

Fig. 4.Morphological interactions observed among two fungi in a Petri dish. Colonymorphologies of fungal co-cultures showing the four major described ‘interaction types’: distance in-hibition, zone lines, contact inhibition and overgrowth, according to Bertrand et al. (2013c).
2
16
3 3
17
6
14 4
1 1
5
0
5
10
15
20
Nu
mb
er o
f re
po
rted
anal
ytic
al t
ech
niq
ues
TLC
HPLC-U
V
HPLC-R
I
GC-FID
LC-M
S
GC-MS
DI-MS
Nano-
DESI-MS
MALD
I-MS
LDPI-M
SNM
R
Not re
porte
d
Fig. 5. Analytical techniques that permit monitoring of microorganism co-culture. Preva-lence of analytical techniques used to monitor chemical changes in microbial co-culturegiven in Supplementary Table 1. Abbreviations: TLC, thin layer chromatography; HPLC-UV; high performance liquid chromatography-ultra violet; HPLC-RI, HPLC-refractiveindex; GC-FID; gas chromatography-flame ionization detection; LC–MS, liquid chroma-tography–mass spectrometry; DI–MS, direct ionization–MS; DESI–MS, direct electrosprayionization–MS; MALDI–IMS, matrix-assisted laser desorption-ionization-imaging MS;LDPI–MS, laser desorption postionization–MS; NMR, nuclear magnetic resonance.
9S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
fungi could be selected. The relevant fungi can be chosen on the basis oftaxonomic criteria, ecological data (sharing an ecological niche) (Billset al., 2013), or growth rate behavior, or they can be selected randomly.Because more than 5 million fungal species could exist (Bass andRichards, 2011; Blackwell, 2011), there are incredible opportunitiesfor developing an almost infinite number of interactions, which couldlead to bioactive NPs. This is also true for other microorganisms, forwhich the biosynthetic potential of such groups has been greatly under-explored. Conducting species confrontations on solidmedia permits themorphology of the species involved in the interactions to be distin-guished, and thus, the area of ‘chemical warfare’ between partners,wheremetabolite induction phenomena are likely to occur, can be accu-rately localized. However, solid medium cultures are usually performedat the Petri dish scale, and only limited amounts of metabolites can beextracted from such culturing conditions (Bertrand et al., 2013d). Thisrepresents a drawback when specific metabolites need to be isolatedfor de novo identification or bioactivity studies.
5.2. Co-cultures in liquid substrates
The co-culture of different species ofmicroorganisms in liquid nutri-ent media is referred to as mixed fermentation, as observed in the nat-ural processes employed in wine fermentation. For instance, Capeceet al. (2013)were able to obtain 160 isolates of Saccharomyces cerevisiaeassociatedwith Sangiovese grapes, and the interactions of six represen-tative strains were studied to understand the natural community thatwas important during wine making. In liquid media, it is not possibleto exploit the modalities of the interactive behavior between microor-ganisms, but the induction of NPs can be monitored.
Fermentation is a well-established method to produce secondarymetabolites from pure strains. Interesting metabolites that have tradi-tionally been extracted from the fruiting bodies of medicinal mush-rooms with high production costs are now obtained in liquid culture,which is an efficientway to produce increased amounts of thesemetab-olites (Tang et al., 2007). However, each parameter involved in cultiva-tion within the bioreactor must be adapted to the developmentalpattern and physiology of the fungus. This is important for the optimiza-tion of the rheological properties and metabolic activity during the de-velopment of the biomass (Znidarsic and Pavko, 2001). Bacterial co-cultures are readily performed in liquid medium, and growth is easilymonitored via turbidimetric methods.
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
Mixed fermentation systems have been developed among variousmicroorganisms, including bacterium–fungus (Mendes et al., 2013; Ohet al., 2005, 2007; Rateb et al., 2013), bacterium–bacterium (Kumaret al., 2013), bacterium–protist (Verhoeven et al., 2010), archaea–fungus(Cheng et al., 2013) and fungus–fungus (Zhu and Lin, 2006) co-cultures,involving both budding and filamentous forms. As reported by Pettit(2009), mixed fermentation is an effective way to increase NP libraries.
5.3. The induction of chemical diversity in microbes via co-culture
Several groups have shown that co-culturing can activate silent geneclusters, but the molecular mechanisms by which this is achieved oftenremain unknown. In fact, microbes can produce compounds that func-tion as transcriptional regulators and epigenetic modifiers. In onestudy, a co-culture of Aspergillus fumigatus with a bacterium was dem-onstrated to activate the same silent pathways as the addition of an epi-genetic modulator (König et al., 2013).
Co-cultivation of microbes can also result in genemutation and sub-sequent expression of otherwise silent gene clusters (Charusanti et al.,2012) or even the exchange of whole gene fragments (horizontal gene
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001
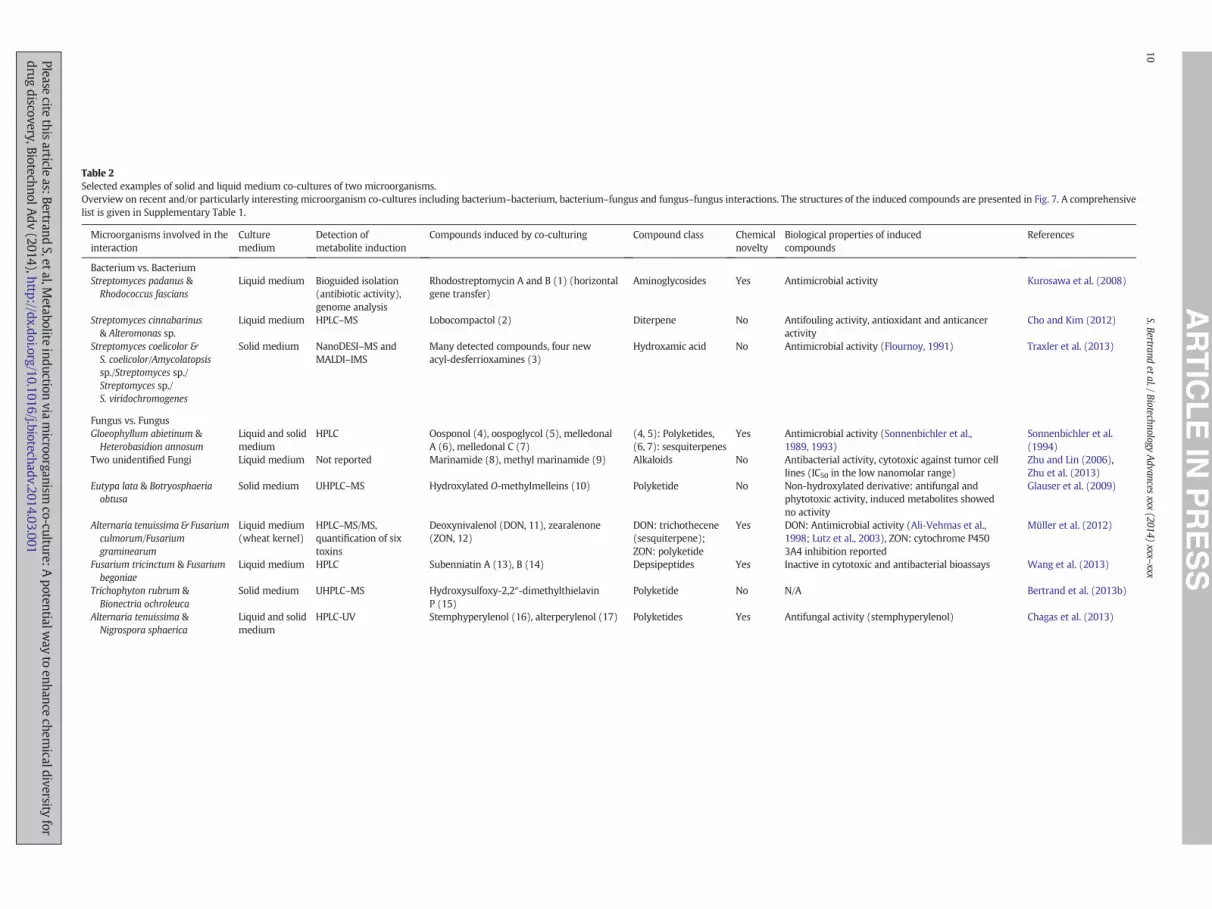
Table 2Selected examples of solid and liquid medium co-cultures of two microorganisms.Overview on recent and/or particularly interesting microorganism co-cultures including bacterium–bacterium, bacterium–fungus and fungus–fungus interactions. The structures of the induced compounds are presented in Fig. 7. A comprehensivelist is given in Supplementary Table 1.
Microorganisms involved in theinteraction
Culturemedium
Detection ofmetabolite induction
Compounds induced by co-culturing Compound class Chemicalnovelty
Biological properties of inducedcompounds
References
Bacterium vs. BacteriumStreptomyces padanus &Rhodococcus fascians
Liquid medium Bioguided isolation(antibiotic activity),genome analysis
Rhodostreptomycin A and B (1) (horizontalgene transfer)
Aminoglycosides Yes Antimicrobial activity Kurosawa et al. (2008)
Streptomyces cinnabarinus& Alteromonas sp.
Liquid medium HPLC–MS Lobocompactol (2) Diterpene No Antifouling activity, antioxidant and anticanceractivity
Cho and Kim (2012)
Streptomyces coelicolor &S. coelicolor/Amycolatopsissp./Streptomyces sp./Streptomyces sp./S. viridochromogenes
Solid medium NanoDESI–MS andMALDI–IMS
Many detected compounds, four newacyl-desferrioxamines (3)
Hydroxamic acid No Antimicrobial activity (Flournoy, 1991) Traxler et al. (2013)
Fungus vs. FungusGloeophyllum abietinum &Heterobasidion annosum
Liquid and solidmedium
HPLC Oosponol (4), oospoglycol (5), melledonalA (6), melledonal C (7)
(4, 5): Polyketides,(6, 7): sesquiterpenes
Yes Antimicrobial activity (Sonnenbichler et al.,1989, 1993)
Sonnenbichler et al.(1994)
Two unidentified Fungi Liquid medium Not reported Marinamide (8), methyl marinamide (9) Alkaloids No Antibacterial activity, cytotoxic against tumor celllines (IC50 in the low nanomolar range)
Zhu and Lin (2006),Zhu et al. (2013)
Eutypa lata & Botryosphaeriaobtusa
Solid medium UHPLC–MS Hydroxylated O-methylmelleins (10) Polyketide No Non-hydroxylated derivative: antifungal andphytotoxic activity, induced metabolites showedno activity
Glauser et al. (2009)
Alternaria tenuissima & Fusariumculmorum/Fusariumgraminearum
Liquid medium(wheat kernel)
HPLC–MS/MS,quantification of sixtoxins
Deoxynivalenol (DON, 11), zearalenone(ZON, 12)
DON: trichothecene(sesquiterpene);ZON: polyketide
Yes DON: Antimicrobial activity (Ali-Vehmas et al.,1998; Lutz et al., 2003), ZON: cytochrome P4503A4 inhibition reported
Müller et al. (2012)
Fusarium tricinctum & Fusariumbegoniae
Liquid medium HPLC Subenniatin A (13), B (14) Depsipeptides Yes Inactive in cytotoxic and antibacterial bioassays Wang et al. (2013)
Trichophyton rubrum &Bionectria ochroleuca
Solid medium UHPLC–MS Hydroxysulfoxy-2,2″-dimethylthielavinP (15)
Polyketide No N/A Bertrand et al. (2013b)
Alternaria tenuissima &Nigrospora sphaerica
Liquid and solidmedium
HPLC-UV Stemphyperylenol (16), alterperylenol (17) Polyketides Yes Antifungal activity (stemphyperylenol) Chagas et al. (2013)
10S.Bertrand
etal./BiotechnologyAdvances
xxx(2014)
xxx–xxx
Pleasecite
thisarticle
as:BertrandS,etal,M
etaboliteinduction
viamicroorganism
co-culture:Apotentialw
ayto
enhancechem
icaldiversityfor
drugdiscovery,BiotechnolA
dv(2014),http://dx.doi.org/10.1016/j.biotechadv.2014.03.001

Fungus vs. BacteriumFusarium oxysporum &Serratia sp./Achromobacter sp.
Liquid medium SPME-GC–MS Isocaryophyllene (18),α-humulene (19), cyclocaryophyllan-4-ol (20)
Sesquiterpenes Yes Suppression of virulence in F. oxysporum,antimicrobial activity of isocaryophyllene
Minerdi et al. (2009)
Aspergillus fumigatus &Sphingomonas sp.
Liquid medium HPLC Glionitrin A (21) Diketopiperazinealkaloids
No Antimicrobial activity and cytotoxicity Park et al. (2009)
Aspergillus nidulans &Escherichia coli
Liquid medium qRT-PCR of thepolyketide gene, HPLC
Orsellinic acid (22), lecanoric acid (23),polyketides F-9775A (24) and F-9775B (25)
Polyketides Yes (22): Antimicrobial activity (Gomes et al., 2003),(23): inhibits ATP synthesis and electron transfer,antimicrobial activity (Gomes et al., 2003;Ranković and Mišić, 2008),(24–25): antiosteoporosis activity
Schroeckh et al. (2009)
Aspergillus fumigatus &Streptomyces peucetius
Liquid medium HPLC–MS Fumiformamide (26), N,N′-((1Z,3Z)-1,4-bis(4-methoxyphenyl)β-1,3-diene-2,3-diyl)diformamide (27), together with two knownN-formyl derivatives and the xanthocillinanalogue BU-4704
Alkaloids Yes Cytotoxic activity Zuck et al. 2011
Aspergillus fumigatus &Streptomyces rapamycinicus
Liquid medium Full genomemicroarray,HPLC-UV
Fumicyclines A (28), B (29) Meroterpenoids No Moderate activity against S. rapamycinicus, responsiblegene cluster found
König et al. (2013)
Fusarium tricinctum & Bacillussubtilis
Solid medium HPLC-UV Induction of 13 compounds, three novelcompounds macrocarpon C (30), 2-(carboxymethylamino)benzoic acid (31), (−)-citreoisocoumarinol (32)
Polyketides No No bioactivity of de novo-induced compounds,antimicrobial activity of some of the upregulatedcompounds
Ola et al. (2013)
Aspergillus fumigatus &Streptomyces bullii
Liquid medium HPLC–MS 11-O-Methylpseurotin A2 (33), and severaldiketopiperazine alkaloids and other knowncompounds
(33): Lactam(pyrrolidone);diketopiperazinealkaloids
No Trypanocidal and leishmanicidal activity andtoxicity
Rateb et al. (2013)
OthersPhytophthora nicotianae &Phytophthora nicotianae
Liquid medium Counting of oospores,HPLC–MS
Mating hormones α1 (34)and α2 (35)
Diterpenoids No The hormones induce sexual reproduction in theopposite mating type
Jee et al. (2002), Ojikaet al. (2011)
11S.Bertrand
etal./BiotechnologyAdvances
xxx(2014)
xxx–xxx
Pleasecite
thisarticle
as:BertrandS,etal,M
etaboliteinduction
viamicroorganism
co-culture:Apotentialw
ayto
enhancechem
icaldiversityfor
drugdiscovery,BiotechnolA
dv(2014),http://dx.doi.org/10.1016/j.biotechadv.2014.03.001

12 S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
transfer), which can result in the production of previously undetectedchemical structures (1) (Kurosawa et al., 2008).
Microbial co-cultures have also been used to explore themodulationof quorum sensing (QS) (Kalia, 2013) in the search for new antibioticsbecause QS modifiers might function as this type of drug. Indeed, QSmolecules play effective regulatory roles within bacterial populationsby adapting global growth to the level of nutrient availability. QSmight also be of interest in other aspects of drug discovery; for instance,a QS lactone produced by Pseudomonas aeruginosa was shown to haltcell proliferation and induce cell apoptosis in human breast cancercells (Li et al., 2004).
5.4. Advantages and drawbacks of solid medium co-culture over mixedfermentation
Microbial co-culture could represent a challenging approach for in-ducing chemodiversity in microbes compared to epigenetic modulationor OSMAC, mainly because of reproducibility issues (Williams et al.,2008). Nevertheless, there is evidence that in certain cases, the activa-tion of silent pathways requires the physical presence of a second mi-crobe (cell–cell interaction) and that metabolites alone (e.g., heat-killed cells, cell-free supernatants or extracts) are not always sufficientto induce the production of secondary metabolites. König et al. (2013)showed that A. fumigatus produced fumicyclines (28, 29) only duringmixed fermentation with Streptomyces rapamycinicus and not in thepresence of the bacterial metabolites. In this particular case, the novelmetabolite could be obtained through overexpression of the corre-sponding polyketide synthase (PKS) gene or the addition of the HDACinhibitor SAHA. Similar results were observed in A. nidulans followingdirect physical contact with specific actinomycete strains. The physicalcontact caused the activation of gene clusters and resulted in the pro-duction of orsellinic acid (Schroeckh et al., 2009), which was not pro-duced during pure-strain culture. In selected conditions, comparablemolecular processes might be involved in co-culture as in epigeneticmodification. This is supported by a study involving the co-culture ofthe plant pathogens Eutypa lata and Botryosphaeria obtusa, which dem-onstrated the upregulation of O-methylmellein (Glauser et al., 2009), acompound that was upregulated in the plant pathogen Stagonosporanodorum through the addition of the epigenetic modifiers SAHA andnicotinamide (X.-L. Yang et al., 2013).
The generation of secondary metabolites is also linked to the nutri-tional environment, and it has been shown that in some cases, antibioticproduction and sporulation are increased in nutritionally poor media(Rigali et al., 2008). Similarly, in a co-culture experiment, the numberof induced metabolites was found to be greater when a smaller volumeof growth medium was used (Bertrand et al., 2012). Furthermore, solidmedium cultures have been found to yield a substantially greater num-ber of metabolites compared with liquid medium cultures (Zahn et al.,2001). Solid medium co-culturing constitutes an easy, flexible andlow-cost format in which to screen NP production. However, thelarge-scale production of co-cultures on solid media remains a rathercomplicated process. Several techniques have been developed to enablethe upscaling of solid medium co-cultures (Adelin et al., 2011; Bohniet al., 2013b; Glauser et al., 2009). Such approaches may generate suffi-cient material to allow the isolation of induced microbial metabolites inadequate amount for in-depth bioactivity studies. However, the use ofpure strains and mixed fermentation remains critical if upscaling to in-dustrial production is needed.
6. Monitoring co-culture metabolite induction
As discussed above, the co-culture ofmicroorganisms leads to variousmorphological interaction patterns, andmodificationsmight occur at dif-ferent levels (from genome tometabolome, Fig. 2). It was clearly demon-strated that various silent genes are expressed in microbial strains thatare confronted in co-cultures. This phenomenon highlights the potential
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
for co-culture of microorganisms to induce secondary metabolite pro-duction, possibly leading to the discovery of new chemical entities.
To highlight significant changes in the microbial metabolome com-position in either solidmedium co-cultures ormixed fermentation, var-ious analytical strategies, including simple targeted quantification,differential metabolite profiling and metabolomics or mass spectrome-try imaging, must be applied.
In some cases, the observed metabolite induction is striking and canbe easily monitored using simple methods, such as thin layer chroma-tography (TLC) or high performance liquid chromatography (HPLC). Inother cases, when no significant modifications are observed, sensitivemetabolomics approaches involving advanced data mining are neces-sary (Nguyen et al., 2012). Alternatively, the induction of metabolitescan be highly localized to the zone of confrontation, forwhichmolecularimaging approaches are well adapted.
The various analytical techniques reported for studying the induc-tion of microbial metabolites (Table 2, Supplementary Table 1) areshown in Fig. 5, together with their frequency of use to date. Somemethods only allow the detection of particular metabolites, whereasothers also provide structural information to allow their identificationin databases (mass spectrometry (MS), tandem mass spectrometry(MS/MS), UV) or de novo identification (NMR). The various analyticalstrategies applied will be discussed in the following sections.
6.1. Targeted and untargeted metabolite profiling
Targeted analyses (quantification of particular compounds, mainlytoxins) were first used to demonstrate induction phenomena duringmicrobial growth. This was achieved through simple differential com-parison via HPLC-UV (Jee et al., 2002; Losada et al., 2009; Ojika et al.,2011; Sonnenbichler et al., 1993; Zhang et al., 2008), gas chromatogra-phy–flame ionization detection (GC–FID) (Do Nascimento et al., 2013;Losada et al., 2009; Thierry et al., 2005) or HPLC–MS (Müller et al.,2012; Soliman and Raizada, 2013), or through GC–MS (Thierry et al.,2005). In addition, bio-guided fractionation procedures have beenemployed to isolate particular compounds from microbial co-culturebroths when a particular induction of bioactivity was observed (Cuetoet al., 2001; Nielsen et al., 1998; Watanabe et al., 1982a, 1990). Thishas generally led to the identification of the induced metabolites re-sponsible for that particular activity. In a number of microbial co-culture experiments, over-production of pigments in the culture medi-um or at the confrontation zone has been observed (Angell et al., 2006;Bertrand et al., 2013d; Glauser et al., 2009; Schürch et al., 2010; Shinet al., 1998; Tsao et al., 1985). In these cases, analytical methods basedon color detection (UV/VIS), TLC (Peiris et al., 2008) or HPLC-UV(König et al., 2013; Nielsen et al., 1998; Onaka et al., 2011) wereemployed to demonstrate the induction and subsequent identificationof the pigments.
6.2. Metabolite profiling and fingerprinting methods
In addition to these targeted analyses, untargeted metabolite profil-ing approaches have recently been applied. These generic methods aimto be as comprehensive as possible and provide data that can either beused for simple differential analyses or for qualitative comparisons. Al-ternatively, the fingerprints obtained in various biological replicatescan be employed in metabolomic analyses following the application ofthe chemometric analysis.
The induction of primary microbial metabolites has been assessedthrough proton nuclear magnetic resonance (1H-NMR) (Cheng et al.,2013), HPLC-refractive index (RI) detection (Hickert et al., 2013;Oliveira et al., 2012) or GC–MS (Du et al., 2012; Paul et al., 2012; Peiriset al., 2008; Zhou et al., 2011). For secondary metabolites, simple TLC(Tokimoto et al., 1987) or HPLC-UV has been successfully employed(Chagas et al., 2013; Nonaka et al., 2011; Ola et al., 2013; Park et al.,2009; Schroeckh et al., 2009; Shanahan et al., 1992; Tokimoto et al.,
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

13S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
1987). However, for such generic fingerprinting, directMS or hyphenat-ed MS methods are much more powerful because of their sensitivityand the resolving power of such detectors (Wolfender, 2009;Wolfender et al., 2013). Furthermore, MS and especially high-resolution MS (HRMS) can provide key structural information for theidentification of the induced metabolites (see Section 6.5).
Furthermore, direct analyseswithminimal or no sample preparationhave been efficiently applied, including direct ionization–MS (DI–MS)(Degenkolb et al., 2002), desorption electrospray ionization–MS(nano-DESI–MS) (J. Watrous et al., 2013; Traxler et al., 2013; Watrouset al., 2012), matrix-assisted laser desorption ionization–MS (MALDI–MS) (Moree et al., 2012, 2013; Traxler et al., 2013; Yang et al., 2009)and laser desorption postionization–MS (LDPI–MS) (Bhardwaj et al.,2013). Total MS spectra of microbial extracts or particular spots fromsolid medium cultures can be generated using all of these approaches.
DI–MSprovides spectra that reflect themetabolite compositionwith-in an extract through simple infusion in an electrospray source. MALDI–MS (O'Connor, 2007) and LDPI–MS (Hanley and Zimmermann, 2009)are desorption methods that allow the ionization of metabolites directlyfrom fungal or bacterial strains on solid medium. Using such methods,rastering over an agar plate provides a good method for recording thetotalMS spectra at definedpositions, and the compilation of such data fa-cilitates MS imaging of a particular microbial co-culture zone (seeSection 6.4). Nano-DESI allows the sampling of amicrobial colony direct-ly from a Petri dish, without sample preparation (J. Watrous et al., 2013;Traxler et al., 2013). This apparatus performs nanoliter-scale liquid ex-traction of the biological surface, and the extract is directly infused intoa mass spectrometer. Compared to desorption techniques such asMALDI, which efficiently analyze metabolites of high molecular weightas well as peptides and proteins, nano-DESI-based approaches involvean electrospray ionization process and therefore can also be efficientlyused to analyze small molecules (Roach et al., 2010). All of these tech-niques cause the direct ionization of the samples butmay suffer from im-portant matrix effects due to the large variety of compounds that areionized and detected simultaneously.
Metabolite profiling strategies using hyphenated MS techniquessuch as LC–MS (Bertrand et al., 2013c, 2013d; Cho and Kim, 2012;Combès et al., 2012; Glauser et al., 2009; Jonkers et al., 2012; Kossugaet al., 2013; Oh et al., 2005, 2007; Park et al., 2009; Rateb et al., 2013;Rodriguez Estrada et al., 2011; Wang et al., 2013; Zuck et al., 2011) orGC–MS (Minerdi et al., 2009; Peiris et al., 2008; Thierry et al., 2005)have often been employed. In such methods, MS spectra are acquiredfor microbial metabolites that are first separated through chromatogra-phy. Such approaches generally exhibit a lower throughput than directMS methods and require more tedious sample preparation. However,they suffer less from ion suppression effects, which are likely to occurin MS. In addition, they provide a good means of separating isobaricstructures, which are often found in NP extracts.
Gas chromatography generally generates HR profiles, andmolecularidentification is easily achieved through database searches based onelectron ionization–MS (EI–MS) spectra and retention indices (Peiriset al., 2008) when reference metabolites are present in the database.In comparison, the resolution of liquid chromatography is generallylower (Giddings, 1967), but when combined with an HR mass spec-trometer using soft ionization methods, such as ESI, this method pro-vides a good separation of metabolome constituents in both thechromatographic and MS dimensions.
The resolution or throughput of LC can be greatly improved throughultra high performance liquid chromatography (UHPLC) using sub-2 μmsilica beads (Eugster et al., 2012) compared to standard HPLC. Using thistype of UHPLC-HRMS approach, very rapid fingerprinting can beachieved over a gradient of 2–5 min (Eugster et al., 2011; Fekete andGuillarme, 2013; Ito et al., 2011). Alternatively, very high resolutioncan be obtained using longer columns and gradient times, where peakcapacities exceeding 1000 can be achieved (Eugster et al., 2012). FastUHPLC–MS is commonly employed to profile large numbers of
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
biological replicates, which are usually necessary for metabolomicsstudies (Want et al., 2010; Wolfender et al., 2013). In LC–MS, HRMSspectra recorded with state-of-the-art instruments provide mass accu-racies below 5 ppm. When combined with heuristic filtering, this levelof accuracy usually enables the molecular formulas of the metabolitesto be confirmed with good confidence (Kind and Fiehn, 2007). This isoften sufficient for peak annotation when the metabolites of interesthave been previously reported, and further information can be generat-ed through MS/MS (detailed in Section 6.5).
6.3. Metabolomics
Once microbial strains have been chemically analyzed using any ofthe methods or combinations of the methods described above, the col-lected data must be analyzed. In co-culture experiments, the goal is tohighlight secondary metabolites that are either produced de novo orare up- or downregulated upon interspecies competition. A simple com-parison of chromatographic tracesmay be sufficient to identify the com-pounds that are strongly induced. This approach is suitable for theanalysis of chromatograms obtained via HPLC-UV (Chagas et al., 2013;Losada et al., 2009; Nonaka et al., 2011; Ola et al., 2013; Schroeckhet al., 2009; Wang et al., 2013), TLC (Peiris et al., 2008) and GC-FID(Do Nascimento et al., 2013; Losada et al., 2009; Thierry et al., 2005).In most cases, this methodology allows the selection of compounds de-tected in the co-culture but not detected in cultures of the pure strains,referred to as de novo-induced metabolites.
MS detection generates large datasets, and in addition to the majormetabolites, minor constituents can be sensitively and selectively de-tected. Similar to the other fingerprinting methods, the induction of astrong ion can be directly observed through the comparison of totalMS spectra or LC–MS chromatograms. This is possible when importantfold changes in the intensities (sufficiently high induction rates (Choand Kim, 2012)) are recorded for well-defined ions or for LC peaksthat are well resolved in the total ion chromatograms (TIC) in LC–MS.This approach has been applied to the data generated by various tech-niques, such as DI–MS (Degenkolb et al., 2002) or LC–MS (Glauseret al., 2009; Kossuga et al., 2013; Oh et al., 2005, 2007; Rateb et al.,2013; Wang et al., 2013; Zuck et al., 2011).
A better overview of all of themetabolomic changes that occur in co-culture experiments can be obtained if fingerprinting data, especiallythose obtained through MS, are evaluated with newly developedchemometric tools, such as those used for metabolomics studies(Wolfender et al., 2013).
For this purpose, an automated datamining proceduremust be usedto analyze the large MS datasets generated. A first step is to filter thedata (blank subtraction, denoising) and convert them into a datamatrix(Fig. 6A) through alignment of the detected features generated viaspectral binning, in the case of direct MS analysis (Kirwan et al.,2013), or through automatic peak detection (peak picking), in the caseof the hyphenated techniques (Gürdeniz et al., 2012). Various currentlywell-established software packages, including some freeware, can beused to perform this step (Georgios et al., 2012; Wolfender et al.,2013). In a subsequent step, the data matrix is analyzed using statisticaltools, including either supervised or non-supervisedmethods, or elabo-ratemachine-learning algorithms to highlight non-obvious information(Georgios et al., 2012; Hur et al., 2013; Saccenti et al., 2013). This ulti-mately provides a way to visualize the differentiation of the co-culturereplicates from those of the pure strains (score plots) and, hence, high-lights the corresponding regulated metabolites (loading plot, and theirinterrelation in a so-called biplot).
One potential datamining approach consists of automated searchingfor de novo-induced metabolites (Fig. 6A I.). In this case, only features(the detected metabolites, corresponding to the mass-to-charge ratio(m/z) at a particular retention time) that are detected in the co-culture extract and that are not found in the pure-strain cultures areconsidered. Simple scripts can be employed to compare the large
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

A) Differential metabolomic strategy B) Imaging-MS strategy
LC-MS analysis
Peak picking
Data analysis to highlight induction
I. De novo induction detection
stra
in 1 #1
#2#3#4#5#6#1#2#3#4#5#6#1#2#3#4#5#6
stra
in 2
co-c
ultu
re
peak area peak area peak area
II. PCA & PLS approach
III. POChEMonapproach
PCA
PLSCompetition PC 1
Com
petit
ion
PC
2
Mixingmodel
Competition model
MS analysis
Metabolite induction pattern
m/z
Inte
nsity
m/z
Inte
nsity
Imaging treatment
Data analysis
RT
m/ z
RT
m/ z
RT
m/z
PC
2
PC1
F2
F1
m/z
Inte
nsity m/z
Inte
nsity
de
m/z
Inte
nsity m/z
Inte
nsity a
m/z
Inte
nsity
m/z
Inte
nsity
c
m/z
Inte
nsity
b
m/z
Inte
nsity f
ion intensity
Mix
ing
PC
2
Mixing PC 1
Metabolite induction pattern
det
ecte
d io
ns
a b c
d e f
Fig. 6.Differentmetabolomics approaches employed to studymetabolite induction duringmicroorganism co-culture, illustrated for the case of solid medium co-culture. The red and bluecolors in the figure correspond to the cultures of the two pure strains, whereas the purple color indicates the co-culture, inwhich the information from the twomicroorganisms cannot beseparated. (A) represents the metabolomics approach (exemplified by the treatment of LC–MS data); the extracts from cultures of the pure strains and the co-culture are first analyzedthrough LC–MS. The generated raw data are transformed into a data matrix through automatic peak picking. The data are then explored to highlight the induced metabolites. The firstapproach (I.) consists of specific searches for de novo induction via searching peak-by-peak features that are only detected in the co-culture. The second approach, represented in figure(II.), demonstrates a multivariate data analysis, in which the data are first presented through a PCA that indicates metabolite induction. In a second step, (O)PLS-DA analysis leads to se-lection of the inducedmetabolites. The third strategy (III.) is the POChEMon approach, which strategically constructs two complementarymodels, a ‘mixingmodel’ between the twopure-strain culture data and another ‘competitionmodel’ that specifically describes the information on features that are unique to the co-culture data. Finally, typical induction patterns selectedthrough the three different datamining approaches are presented. (B) represents the imagingMS approach. The surface of an agar platewhere bothmicroorganisms are grown together issequentially rastered to acquire anMS spectrum for every location. Images are generated for each ion detected (for example, ions a–f) to highlight their location in the co-culture and, thus,to select thosemetabolites present in the confrontation zoneof the co-culture (as represented by the induction patterngenerated through imagingMS). Abbreviations: LC–MS, liquid chro-matography–mass spectrometry; PCA, principal component analysis; PLS, partial least square; OPLS, orthogonal projection to latent structure; DA, discriminant analysis; POChEMon,projected orthogonalized chemical encounter monitoring. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
14 S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
Please cite this article as: Bertrand S, et al, Metabolite induction via microorganism co-culture: A potential way to enhance chemical diversity fordrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotechadv.2014.03.001

15S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
datasets generated throughMS detection and, thus, allow the detectionof the induced minor compounds (Bertrand et al., 2013c, 2013d). Thesescripts systematically compare the ‘detected’ status (peak height over acertain threshold) of every feature identified, selecting those that areuniquely ‘detected’ in the co-culture replicates. The highlighted featurescorrespond to highly reproducible inducedmetabolites. De novo induc-tion stricto sensu is not always detected in co-cultures, but other impor-tant metabolomic changes correspond to the up- or downregulation ofsome metabolites (Cho and Kim, 2012). To demonstrate all of thetypes of microbial metabolomic changes that may occur due to fungalconfrontation and to take into consideration the dynamic aspectsrelated to fungal growth, more elaborate chemometric tools should beused.
A second data mining approach involves the application of conven-tional statistical tools to replicates of pure strains and co-cultures, ascarried out in metabolomic analyses (Fig. 6A II.). For microbial co-culture experiments, various methods of mining the data have beenemployed, including principal component analysis (PCA) (Bertrandet al., 2013c; Bhardwaj et al., 2013; Du et al., 2012; Peiris et al., 2008;Zhou et al., 2011), analysis of variance (ANOVA) (Combès et al., 2012;Jonkers et al., 2012; Müller et al., 2012; Rodriguez Estrada et al., 2011),partial least squares regression coupled with discriminant analysis(PLS-DA) (Bertrand et al., 2013c), orthogonal projections to latentstructure-DA (OPLS-DA) (Bohni et al., 2013b; Jansen et al., 2013,2014) and similar approaches (Paul et al., 2012).
From a data mining perspective, the two states ‘control’ and‘stressed’ are compared in traditional metabolomic analyses. However,the ‘induced’ state observed in the co-cultures should be comparedwith the two pure strains simultaneously, and none of the conventionalmethods accomplishes this.
As a first exploratory step, it is often informative to explore thedata through an unsupervised approach, such as PCA (Jolliffe,2002). PCA displays the relationships among the sample distribution,whichmay reveal groupings, trends or outliers (Boccard et al., 2010).In co-culture experiments, three well-differentiated clusters areoften observed, confirming the differences in metabolite composi-tion between the pure strains and the compared co-culture.When the co-culture cluster is located away from the barycenter ofthe two pure-strain clusters, it most likely indicates the existenceof metabolic modifications that are not related to the mixing of themetabolomes of the two pure strains and suggests the presence ofnew metabolites that are specifically induced by the interaction(Fig. 6A II.). One approach for selecting the induced biomarkers con-sists of comparing the co-culture data with the pure-strain datausing univariate approaches (Hur et al., 2013; Vinaixa et al., 2012).In this case, the biomarkers of interest are selected through theexploration of each feature detected individually. The selection isbased on the fold change in the peak area between the co-cultureand the two pure-strain cultures and therefore requires an evalua-tion of statistical significance based on Student's t-test (with onlythe most intense pure-strain peak area being considered). Incomparison with the previously described de novo detectionapproach, this strategy focuses more on fold-change information,rather than the ‘detected’ status. Therefore, upregulated ions arealso highlighted. A second approach consists of the construction oftwo (O)PLS-DA models through multivariate statistical analysis(comparison of the co-culture with each pure-strain culture). Eachmodel permits the classification of features according to its capacityto separate two groups (the co-culture and one of the pure-straincultures). Thus, features can be selected that are highly specific tothe co-culture compared to a single pure-strain culture. Finally,shared and unique (SUS) plots are presented, providing a two-dimensional representation of the various features (Wiklund et al.,2008). Both dimensions correspond to the relevance of the bio-markers in the two designed models. This approach may help tohighlight the induced biomarkers in both models.
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
However, these last approaches are not well adapted for studyingmixed microbial cultures (Jansen et al., 2013, 2014). PCA describes themetabolome variation among all samples but does not highlight co-culture-induced modifications specifically. Similarly, (O)PLS-DAconstructs models that are highly focused on comparison of the co-culture with the individual pure-strain cultures but does not considerthe co-culture as a mixture of the two microbial species and thereforedoes not describe the metabolic variation expected to occur in the co-culture experiment in an interpretable form. None of the previously de-scribed approaches takes advantage of the particular biological setupprovided by the co-culture experiment, and therefore, their biologicalrelevance is limited. Thus, these conventional statistical tools mightnot always highlight critical co-culture-specific information. In addition,the biological variations observed during microbial growth (Williamset al., 2008) complicate these tasks. Thus, there is a need for innovativedatamining approaches to highlight significant upregulation phenome-nawhen extracts obtained from solidmedium co-cultures ormixed fer-mentation are analyzed.
To address this particular need, a novel approachwas recently devel-oped, referred to as projected orthogonalized chemical encounter mon-itoring (POChEMon, Fig. 6A III.) (Jansen et al., 2013, 2014). This datamining strategy is based on the intrinsic structure of the co-cultureexperiment. Initially, the co-culture data are mathematically re-constructed as a mixture of all pure-strain cultures in which metabolicvariation can be explored. The first step is the creation of a mixedmodel from the two pure-strain metabolomes that represents the co-culture experiments. The position of the co-culture replicates is alreadyinformative: if they are positioned closer to one of the two pure strains,it indicates that this particular strain is themajormetabolite producer inthe particular co-culture. The residual information in the co-culture rep-licates is then likely associated with the interspecies competition, spe-cifically when shared by multiple replicates. From this information, asecond model is built. This latter model, referred to as the competitionmodel, explores these residuals through PCA. All of the co-culture ex-periments are compared, and upregulated features are highlightedusing a biplot (Gower et al., 2011). This approach has been found toprovide information about generally induced metabolites and aboutmetabolites induced only in specific replicates (Jansen et al., 2013,2014). Because specific replicated behaviors are taken into account,clear induction phenomena found only in a few replicates can still behighlighted. This approach might be superior in accounting for thehigh variability that is found in such complex systems. This approachcan also highlight features that are downregulated in co-cultures andtherefore help to delineate interspecies crosstalk in microbial modelsrelated to naturally co-habiting microorganisms.
All of these approaches provide a list of biomarkers with a particularm/z or anm/z at a particular retention time together with its specific in-tensity across all replicates. For key features, it is always important toverify the modifications in the raw MS data. Often, several featuresmay be related to one specific metabolite (due to the detection of ad-ducts), and relationships between features (correlated features) mustbe established to further validate the induction of a specific metabolite.Typical differences in metabolite behavior highlighted among the purestrains and co-cultures using these different data mining approachesare shown in Fig. 6A. The de novo induction detection approach enablesthe selection of de novo-induced metabolites corresponding to highlyreproducible and highly induced compounds that are not detected inpure-strain cultures. The classical statistical approach (e.g., using PCAfollowed by (O)PLS) highlights highly reproducible upregulatedmetab-olites. Among the induced compounds that are identified, de novo-inducedmetabolites are generally highlighted unlessmore significantlyupregulated features are present in the data. Finally, the recently de-scribed POChEMon approach also indicates induced metabolites withhigh specific reproducibility in certain co-culture replicates. Thus, allof these approaches are complementary for the selection of compoundsof interest in natural product drug development.
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

16 S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
6.4. Imaging mass spectrometry
The comparison of extract profiles from pure-strain cultures and co-cultures allows the globalmetabolic changes taking place in solid medi-um co-cultures or in mixed fermentation to be highlighted. The induc-tion of metabolites in co-cultures is usually localized at the zone ofconfrontation in solid media, and methods allowing precise spatial lo-calization are particularly useful to highlight the induction in the specif-ic zones of interaction and to understand which partner is producing aparticular metabolite (Yang et al., 2009). Recently developed imagingMS (IMS) technologies provide an efficient method to analyze thespatial distribution of molecules within solid agar media (Shih et al.,2014). Various technologies have been used to explore the spatial distri-bution of metabolites via MS in co-cultures or in solid media, includingMALDI and nano-DESI (Moree et al., 2012;Watrous et al., 2012). The se-quential rastering across a surface while acquiring a mass spectrumfrom every location is necessary for IMS. From the obtained dataset,multiple images can be extracted that represent the spatial distributionof each ion detected with high spatial resolution, depending on the ras-ter step size (Simmons et al., 2008). The analysis of these images(throughmapping) allows the presence of some secondary metabolitesto be visualized as a function of their location in the solid culture medi-um (Fig. 6B). The major limitation of this method lies in its spatial reso-lution, which is generally approximately 10 μm, although the mostrecent advances allow IMS with a resolution of up to 1 μm to be per-formed (Römpp and Spengler, 2013). However, most of the availablemethods exhibit a sufficient spatial resolution to localize induction inthe confrontation zone between microorganisms.
The choice of ionizationmethodhas a strong influence on the type ofmetabolites detected. MALDI is better suited for large metabolites,peptides or proteins, while nano-DESI reveals smaller secondarymetabolites.
The spatial distribution ofmetabolites in interactingmicroorganismshas been studied only using MALDI and nano-DESI (J. Watrous et al.,2013; Moree et al., 2013; Traxler et al., 2013; Watrous et al., 2012;Yang et al., 2009). In these analyses, themetabolites of interest were se-lected based on their spatial distribution, specifically, the metaboliteslocated in the interaction zone where the two microorganisms are incontact. This approach provides interesting insight into microbial inter-actionmechanisms andmay also help delineate interspecies crosstalk inmicrobial models. Three-dimensional (3D) visualization of the metabo-lite distribution was recently achieved via 3D IMS (J.D. Watrous et al.,2013). This technique takes advantage of traditional technology in serialcross-sections of microbial colonies grown on agar. The reconstructed3D images of selectedmetabolites allow their spatial distributionswith-in the microorganisms to be visualized.
Such methods are powerful but may suffer from ion suppression. Acomplete survey of all induction phenomena in the interactingmicroor-ganisms most likely requires a combination of imaging and metabolo-mics approaches.
6.5. Metabolite identification
In most studies addressing microbial co-cultures (N60% see Table 2,Supplementary Table 1), the induction of previously isolated com-pounds has been reported. This information can be important if theco-culture is studiedwithin the context of an ecological niche. However,if the goal is to generate new chemical diversity through the activationof silent biosynthetic pathways, it is important to identify the inducedmetabolites correctly. This can be achieved through the dereplicationof MS and MS/MS data (El-Elimat et al., 2013; Nielsen et al., 2011;Wolfender et al., 2013) prior to purifying the highlighted metabolites.
The identification of the highlighted metabolites based only on theMSdata represents themost challenging step (El-Elimat et al., 2013). In-deed, MS identification requires comparison with databases, and inmany cases, information for the majority of microbial metabolites is
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
not accessible. As a result, only putative assignments based on molecu-lar formulae (MF) and microbial chemotaxonomy can be made. Fur-thermore, even when the appropriate databases are available, becausechemical novelty is expected in the study of induced secondary micro-bial metabolites, de novo structure elucidation is required, which mustinvolve NMR.
When GC–MS is employed for profiling, the identification workflowis partially based on the comparison of the EI–MS spectrawith referencecompounds andwith database searches based onMS spectra and reten-tion indexes (Du et al., 2012; Kind et al., 2009; Minerdi et al., 2009; Paulet al., 2012; Peiris et al., 2008; Thierry et al., 2005; Zhou et al., 2011).When no clear identification is achieved, the evaluation of compoundsis much more difficult. In such cases, preparative GC may be used forcompound purification for further identification (Zuo et al., 2013). Theuse of soft ionization techniques, such as chemical ionization,field emis-sion and field desorption, can be employed in GC; these methods areless informative but may be employed to confirm themolecular weightof the metabolites of interest.
When DI–MS, nano-DESI–MS and LC–MS are used, the metabo-lites are ionized through soft ionization techniques, mostly involvingESI. However, atmospheric pressure chemical ionization (APCI) canalso be employed. Such methods mainly produce molecular ion spe-cies that appear in the form of adducts, such as [M+H]+, [M+Na]+,[M + H + CH3CN]+ if acetonitrile is used as solvent, [M + H–H2O]+
in positive ion mode or [M–H]−, [M + HCO2]−, or [M–H + CO2]− innegative ion mode (Kind et al., 2009; Nielsen et al., 2011). Further-more, dimers, which complicate interpretation, may also be formedin the ion source. Therefore, prior to any dereplication, the correctmolecular weight should be defined through adduct recognition(Kuhl et al., 2011). In addition, comparison of the different ionizationmodes (positive or negative ionization (PI, NI)) may help to unam-biguously reveal molecular ions (Funari et al., 2012). The presenceof these different ions complicates the dereplication process, but itis important to note that during the data mining step, several m/zfeatures can be associated with a single induced metabolite. Thismust also be taken into account when analyzing the loading resultsfrom metabolomic datasets.
To improve identification, HRMS detectors are now generallyemployed. Using such instruments, the MF can be determined basedon an accurate mass, along with a high spectral accuracy of particularmolecular ions. However, the unambiguous determination of the MF isstill difficult, even when mass accuracies below 1 ppm are obtained,particularly for compounds with a high molecular weight (N500 Da)(Kind and Fiehn, 2006). To reduce the number of possible MF, differentheuristic filters can be applied (Kind and Fiehn, 2007): i) restrictions onthe number of elements; ii) LEWIS and SENIOR chemical rules; iii) iso-topic patterns; iv) hydrogen/carbon ratios; v) the elemental ratios of ni-trogen, oxygen, phosphorus and sulfur versus carbon; vi) elementalratio probabilities; and vii) in the case of GC–MS data, the presence oftrimethylsilylated compounds. When high-resolution multistage MS(HRMSn) spectra can be recorded, these spectra can be employed to re-duce the number of MF based on the determination of fragment MF(Konishi et al., 2006). To speed up this time-consuming process, suchworkflows may be automated (Brown et al., 2011).
Based on the putative MF, the identification of metabolites isachieved using various NP databases. This process usually leads to mul-tiple putative identities for each highlighted feature. To further reducethe number of possible structures and improve the accuracy of peak an-notation, other filters based on retention time prediction (Creek et al.,2011; Stanstrup et al., 2013) and microbial phylogeny (Bertrand et al.,2012) can be applied. When applied to co-cultures, these approacheshave resulted in several putative identifications (Bertrand et al., 2012,2013c, 2013d), with the differences in hit rates being greatly reducedin poorly studiedmicroorganisms. A recent systematic study addressingvarious fungal co-culture experiments demonstrated that co-culture-induced metabolites display a lower database hit rate than the
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

O
O
O
NHO
O
OH
O
NH
HO
HO
HO HO
HOHN
NH2
NH
HN
NH2
NH
rhodostreptomycin A and B (1)
* OHO
OHlobocompactol (2)
R1 N NH
NHN
O
OH
O
O
OH
O
N
O
OH
NH2
R1: alkyl, acyl-desferrioxamines (3)
O
OH O
R2HO
R2: =O, oosponol (4)R2: -OH, oospoglycol (5)
HO
HHO
O
OH
O
O
HO
OR3, R4: H, melledonal A (6)R3: CH3, R4: Cl, melledonal C (7)
NH
O O
O
NH
R5: H, marinamide (8)R5: CH3, methylmarinamide (9)
O
OO
H R6 R7
R6, R7: H, OH, hydroxylated O-methylmelleins (10)deoxynivalenol (11)
OH
HO
O O
O
zearalenone (12)
R4
R3
R5
HON
ON
OH
O
O
O
R8O
R8: CH3, subenniatin A (13)R8: C2H5, subenniatin B (14)
O
O
O
O
O
OHO
OH
O
OHO3S
hydroxysulfoxy-2,2’’-dimethylthielavin P (15)
O OH
HOH H
OH
OOH
stemphyperylenol (16)OH
OHO
HOHHO
Oalterperylenol (17)
isocaryophyllene (18)α-humulene (19)
OH
cyclocaryophyllan-4-ol (20)
glionitrin A (21)
HO
OH
O
OHorsellinic acid (22)
O
O
HO OH
OH
OH
O
lecanoric acid (23)
O
O
OHO
O
HO
O
O
O
O
O
HO
O
OHF-9775A (24)F-9775B (25)
O
HO
OH OH O
OH O
OH
HO
OH OH O
OH
OH O
fumicyclines A (28) fumicyclines B (29)
O
NH
O
OMe
OH
O
O
OH
OMe
11-O-methylpseurotin A2 (33)
O
OOH
HO
O
OOH
HO
OHOH
(-)-citreoisocoumarinol (32)
COOH
NH
COOH
macrocarpon C (30)
2-(carboxymethylamino)benzoic acid (31)
HOHO
O
OH
HOHO
OH
alpha1 (34)
alpha2 (35)
NH
HN
O
OO
O
R9
R9: CH3, R10: SO3H, fumiformamide (26)R9: CH3, R10: CH3, N,N’-((1Z,3Z)-1,4-bis(4-methoxyphenyl)β-1,3-diene-2,3-diyl)diformamide (27)
R10
O
O
HOHO
H H
OHO
N
N
O2N
SS
O OH
O
Fig. 7. Selected natural products induced by microorganism co-culture. The numbers of the molecules correspond to the numbers given in Table 2.
17S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
constitutively produced compounds (Bertrand et al., 2013c). This inter-esting result might indicate that induced metabolites exhibit a greaterchance of being novel compared to constitutive microbial metabolites.Itmight also be that the detectedmetabolites correspond to constitutivemetabolites that are biotransformed during the interaction (analogues)corresponding to unreported NPs. In any case, since dereplication ofsuch compounds cannot be performed based on MS only, complete denovo structure identification by NMR would be needed to further con-firm the metabolites' identity.
As a complementary approach, MS/MS spectra networking has beensuccessfully applied for microbial compound identification (J.Y. Yang
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
et al., 2013a). This strategy is based on searching for similarities inMS/MS spectra among various microbial metabolites. In some cases,this procedure has led to the successful identification of a large numberof structurally analogous metabolites based on similar fragmentationpatterns (J. Watrous et al., 2013; Moree et al., 2013). For example, alarge series of desferrioxamines (3) was identified in co-cultures inwhich Streptomyces coelicolorwere combined with various other bacte-ria (Traxler et al., 2013).
As co-culture-induced metabolites are usually unsuccessfullydereplicated, the purification and subsequent de novo identification ofthese metabolites through NMR is the only possible means of achieving
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

18 S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
unambiguous identification (Bucar et al., 2013). For this purpose, classi-cal isolation strategies based onmultiple fractionation steps have large-ly been employed (Angell et al., 2006; Chagas et al., 2013; Cueto et al.,2001; Glauser et al., 2009; König et al., 2013; Kossuga et al., 2013;Kurosawa et al., 2008; Li et al., 2011; Oh et al., 2005, 2007; Rateb et al.,2013; Soman et al., 1999; Tsao et al., 1985; Watanabe et al., 1982a;Zhu and Lin, 2006; Zuck et al., 2011). Recently, LC–MS-targeted isolationbased on software-driven protocol optimization was used to improvethe purification procedures and rapidly isolate the highlighted com-pounds (Bertrand et al., 2013d; Glauser et al., 2008). This approachtakes advantage of the possibility of simulating metabolite elution(Heinisch et al., 1990) to optimize a purification protocol. This targetedstrategy allows a reduction in the number of purification steps requiredto isolate a pure compound from a crude extract. Unfortunately, the pu-rification of metabolites can sometimes be very challenging due to thesmall amount of material available and the small amount of thehighlighted compound produced (Bertrand et al., 2013d). In addition,because biomarker detection is based on MS, the intensity of thepeaks is not directly correlated with the amounts present, and an in-tense ionmay represent an extremely small amount of a very efficientlyionizedmetabolite (Wolfender, 2009). Thus, despite the high sensitivityof the most recent NMR instruments, the intrinsic differences in sensi-tivity between MS and NMR may lead to unsuccessful identification ofsome metabolites (Bertrand et al., 2013d). To solve such problems,large-scale co-culture experiments should be undertaken in both liquidand solidmedia to improve isolation efficiency (Bohni et al., 2013b). Theproduction of metabolites in solid media is now also possible at largerscales due to recently developed agar-supported solid-state fermentersthat mimic what occurs in Petri dishes over an area of two square me-ters (Adelin et al., 2011).
To address the incompatibilities between metabolomic approaches,which require a large number of replicates, and the identification ofthemetabolites following purification, a novel strategywas recently de-veloped (Bertrand et al., 2012). In this approach, the co-culture profilesobtained from 2-cm Petri dishes were comparedwith those obtained in15-cm dishes. Metabolomic analysis was performed on the smallerdishes after four days of growth. Using this method, most of the denovo-induced metabolites (16/18) were detected in the extract obtain-ed in the 15-cm Petri dishes after three weeks of growth. This findingclearly demonstrates that solid medium co-culture can be scaled upwithout loss of metabolome diversity.
7. Bioactivity of metabolites induced via co-culture
Among the 58 examples of co-culture studies that focused on sec-ondary metabolite induction summarized in Supplementary Table 1(Supplementary information, selected examples are shown inTable 2), the identified biomarkers were found to cover almost all clas-ses of microbial metabolites. Some of the induced compounds were ob-served to show numerous activities. Naturally, many antimicrobialswere identified because this type of activity is expected to arise frommicrobe–microbe interactions as a direct defense mechanism.
7.1. Type of induced metabolites
Among the classes of compounds observed in co-culture experi-ments (selected examples shown in Fig. 7), polyketides have oftenbeen reported. This result might be attributed to the high abundanceof polyketide-producing organisms aswell as the great structural diver-sity of this class of compounds (Hertweck, 2009).
The activation of silent PKS gene clusters through co-culturing hasbeen demonstrated. This activation resulted in the production of alarge variety of induced polyketides in various strains of Aspergillus sp.(e.g., A. nidulans (Schroeckh et al., 2009) and in A. fumigatus (Königet al., 2013)). In A. nidulans, the induced compounds were shown tobe 2,4-dihydroxybenzoic acid derivatives and dimers of these
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
compounds (22–25). A similar compound, consisting of a sulfonated tri-mer of 2,4-dihydroxybenzoic acid (15), was also identified in a co-culture of Bionectria ochroleuca and Trichophyton rubrum. WhenA. fumigatus was co-cultured with a bacterium, other polyketides(fumicyclines A (28) and B (29)) were identified. Furthermore, upregu-lation of the polyketide zearalenone (ZON), among other knownmyco-toxins,was observed in co-cultures of Fusarium culmorum and Alternariatenuissima (Müller et al., 2012). In addition, a large variety ofpolyketideswith special structural features has been reported, includingaromatic monocycles (Cueto et al., 2001; Müller et al., 2012; Nonakaet al., 2011; Notz et al., 2002; Ola et al., 2013), linear skeletons(Kossuga et al., 2013; Tokimoto et al., 1987; Watanabe et al., 1982a,1990) and multiple aromatic fused cycles (Chagas et al., 2013; Glauseret al., 2009; Li et al., 2011; Oh et al., 2005; Ola et al., 2013; Onakaet al., 2011; Peiris et al., 2008; Sonnenbichler et al., 1994; Zhang et al.,2008).
The production of peptides through the activation of non-ribosomalpeptide synthase (NRPS) gene clusters (Schwarzer et al., 2003) has alsobeen widely reported in the literature. Various types of peptides havebeen found, such as depsipeptides (13–14) (Oh et al., 2007; Wanget al., 2013), iturins (Moree et al., 2013) and lipoaminopeptides(Degenkolb et al., 2002). NRPS can also be involved in the productionof hydroxamic acid compounds (e.g., 3) such as siderophores (Kadiand Challis, 2009), and such compounds have been reported to be in-duced through co-culture (Jonkers et al., 2012; Moree et al., 2012;Traxler et al., 2013). These molecules are involved in the scavenging ofiron, which is important for cell growth, from the environment.
Some of thesemetabolites may be generated throughmixed biosyn-thetic pathways. This is the case, for example, for the cytochalasins ob-served to be induced during the co-culture of two Aspergillus spp.(Losada et al., 2009) as a result of the activation of hybrid NRPS-PKSgene clusters (Fujii et al., 2013). Another example is the tetramic acid-derived 11-O-methylpseurotin A2 (33) that was induced in a co-culture of A. fumigatus with Streptomyces bullii (Rateb et al., 2013).PKS-NRPS gene clusters have been shown to be required for the biosyn-thesis of the pseurotin compound class (Maiya et al., 2007).
The induction of terpenes via activation of terpene synthase(Smanski et al., 2012) has also been reported. This class includes com-pounds such as sesquiterpenes (11, 18–20) observed in mixed fermen-tations involving Fusarium spp. (Minerdi et al., 2009; Müller et al.,2012), diterpenes (2, 34, 35) produced in various settings (Cho andKim, 2012; Jee et al., 2002; Soliman and Raizada, 2013) and triterpenesoccurring in a mixed fermentation of Aspergillus sp. with Streptomycessp. (Rateb et al., 2013). Other mixed biosynthetic pathways based onterpene synthases leading to meroterpenoides have been reported aswell (Geris and Simpson, 2009; König et al., 2013). Furthermore, variousalkaloids were shown to be induced through co-culture. The reportedalkaloids mostly corresponded to cyclic aromatic amines such asprocyanines (monomers or dimers) (Angell et al., 2006; Moree et al.,2012), marinamides (8, 9) (Zhu and Lin, 2006; Zhu et al., 2013),pyrazines (Zhu et al., 2009), pyrrols (Onaka et al., 2011; Straight et al.,2007; Tsao et al., 1985) and picolinic acid derivatives (Jonkers et al.,2012). An unusual example is the N-formyl alkaloids (26, 27) producedin a co-culture of A. fumigatuswith S. peucetius (Zuck et al., 2011). Addi-tionally, the production of free fatty acids (Combès et al., 2012; Thierryet al., 2005) and glycosides (Kurosawa et al., 2008) has been reported infew cases.
Overall, no clear trend has yet been demonstrated regardingwheth-er particular compound classes are more prone to be induced throughmicrobial co-culture. This may be related to the small number of poten-tial microbe interactions studied to date and to the relatively low diver-sity of the microorganism species involved. Aspergillus spp. andStreptomyces spp. have mainly been studied in this context. Neverthe-less, in comparison with strategies based on gene activation foraccessing novel molecules, co-culture appears to activate specificgenes, but depending on the interaction type, metabolites resulting
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

19S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
from various unrelated pathways may be induced. This contrasts withthe molecular biological approaches, in which the PKS and NRPS geneclusters have been found to be the predominant pathways activated(Brakhage and Schroeckh, 2011). The limited knowledge of other geneclusters may explain why only a few studies using molecular biologicalapproaches have observed the activation of other gene cluster families.Therefore, co-culture can be considered an untargeted method of geneactivation that allows the production of very structurally different sec-ondary metabolites.
7.2. Biological activity of the induced metabolites
During co-culture on solid media, clear long-distance growth inhibi-tion is observed in some cases (Bertrand et al., 2013d; Chagas et al.,2013; Combès et al., 2012; de los Santos-Villalobos et al., 2012; Nielsenet al., 1998, 1999; Schürch et al., 2010;Watanabe et al., 1982b;Wolfenderand Queiroz, 2012; Zuck et al., 2011). This observation is expected to belinked to the production of antimicrobial compounds as a defense mech-anismby one of the twomicroorganisms being studied. The activity of theisolated metabolites induced by the co-culture has been evaluated insome instances, but this has not always led to the successful identificationof the antimicrobial compounds (Combès et al., 2012; Zuck et al., 2011). Arecent study that examined a large number of co-cultures indicated thatonly 5% of more than 600 co-cultures tested demonstrated long-distance growth inhibition and that such interactions are not linked to aparticularly striking pattern of metabolite induction compared to othermorphological patterns (Bertrand et al., 2013d).
In general, various studies have observed an increase in the antimi-crobial activity of extracts from co-cultures in comparison to those fromcorresponding pure-strain cultures (Losada et al., 2009). Severaltargeted studies on known antimicrobial compounds have demonstrat-ed that they can be induced by co-culturing (Müller et al., 2012;Sonnenbichler et al., 1994). For example, co-culturing Gloeophyllumabietinum with Heterobasidion annosum induced the production of theknown antibiotics oosponol (4), oospoglycol (5), melledonal A (6) andmelledonal C (7) (Sonnenbichler et al., 1994). This clearly highlightsthe potential for co-culture to induce antimicrobial compounds usefulfor drug discovery. Furthermore, various untargeted differential metab-olite profiling strategies and metabolomic approaches have beenemployed to identify promising molecules, which have, in some cases,led to the identification of antimicrobial compounds (see Supplementa-ry Table 1). In a co-culture of Pseudomonas aeruginosa with Aspergillusfumigatus, IMS revealed local induction of iturins, which are moleculesthat exert antifungal activities, in the confrontation zone (Moree et al.,2013). In addition, the production of iron (III)-scavenging metabolites(siderophores) in the confrontation zones between P. aeruginosa andA. fumigatus and between Streptomyces coelicolor and Amycolatopsis sp.(Moree et al., 2012; Traxler et al., 2013) may highlight survival compe-tition between the twomicroorganisms. This competition for ferric ionsmay lead to growth inhibition of the microorganisms that do not pro-duce these metabolites.
In addition to antimicrobial activity, induced metabolites also showdiverse other biological activities. Various compounds exhibit cytotoxicactivity against different tumor cell lines as well as antiparasitic activity(against malaria, trypanosoma and leishmania, see Table 2, Supplemen-tary Table 1). For example, production of the anticancer agent taxol byParaconiothyrium sp. was induced in presence of Alternaria sp. orPhomopsis sp., all three of which are endophytes of Taxus trees(Soliman and Raizada, 2013). Some hydroxamic acid siderophores(Moree et al., 2012; Traxler et al., 2013) have been reported to possessa wide range of biological activities (Bertrand et al., 2013a; Flournoy,1991). In addition to compounds presenting activities of therapeutic in-terest, induced metabolites with antifouling activity (2) (Cho and Kim,2012) have also been reported.
To further evaluate the potential of induced metabolites as leads forvarious drug targets, co-culture extracts can be subjected to biological
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
HPLC profiling (Potterat and Hamburger, 2013) for the rapid localiza-tion of bioactive metabolites in these microbial extracts. HPLCmicrofractionation is a valuable method that is compatible with thesmall amounts of extracts that are usually obtained from solid mediumco-cultures (milligram amounts). These bioassays can be performed at-line, and this format is compatible with the evaluation of compoundscollected in the microgram range (typically, tenths of micrograms).NMR information complementary to that provided by MS can nowbe obtained for such minute amounts of material using highly sensitivemicroNMR methods (Dalisay and Molinski, 2009) to improvedereplication results, and this information can also be used to quantifythe amounts collected for further assessment of the potency of microbi-al metabolites (Bohni et al., 2013a) in various assays. Regarding antimi-crobial activity, bioassays can be performed at-line using bioautography(Favre-Godal et al., 2013) or microassays (Bertrand et al., 2013b) or on-line, coupled directly to a separation device, such as LC (Potterat andHamburger, 2013). Such micro-isolation procedures are well adaptedto the amounts of material that are usually obtained from microbialmetabolomics studies (e.g., by pooling replicates) andmay lead to effec-tive, very early identification of inducedmetabolites with high bioactivepotential.
7.3. Chemical novelty among the induced metabolites
Among the compounds reported to be induced during microbial co-culture, approximately half of the identified molecules represent newnatural products (see Table 2, Supplementary Table 1). This is a highproportion and indicates that new structures are likely to be foundwhen induced compounds are specifically targeted. This situation iseven more striking for metabolites that are induced de novo and aretherefore not constitutively produced by any of the studied strains.
Dereplication based on MS data is a mandatory step to evaluate thepossible novelty of induced metabolites (Bertrand et al., 2012, 2013d;Roemer et al., 2011). The failure of peak annotation based onmolecularformula assignment and chemotaxonomy cross-searches in databasesof microbial metabolites indicates a high probability that a newmetab-olite has been detected. However, the success of dereplication is highlydependent on whether in-depth secondary metabolite analysis hasbeen previously carried out on the studied microorganisms. Important-ly, as noted above, induced metabolites exhibit lower hit rates indatabases in comparison with constitutively expressed microbial me-tabolites, even in poorly studied strains. This demonstrates the great in-terest in co-culturing to induce interesting, chemodiverse metabolites.A general trend observed among the studied co-cultures is upregulationof the production of a series of analogues that share the same skeleton(Glauser et al., 2009; Moree et al., 2012; Nonaka et al., 2011; Oh et al.,2005; Rateb et al., 2013; Traxler et al., 2013; Zuck et al., 2011).
So far, microbial co-culturing has not yielded many examples of in-duced molecules possessing novel skeletons but instead has resultedin the identification of derivatives of knownmicrobialmetabolites. Sim-ilarly, the bioactivity potency of these compounds has not been found tobe significantly different from other constitutive metabolites. However,several findings, such as the discovery of marinamides (8, 9), whichhave nanomolar anti-tumor activity (Zhu et al., 2013), and glionitrin A(21), which shows submicromolar cytotoxic activity towards theDU145 human cancer cell line (Park et al., 2009), indicate that this ap-proach still has great potential to yield not only derivatives of knowncompounds but also novel skeletons or new lead structures.
8. Conclusion
Microbial genomic data produced in the last decade have revealedan incredible diversity ofmetabolic pathways forwhich nofinal producthas yet been identified. It is now widely accepted that microorganismsharbor massive potential for the biosynthesis of compounds of highchemical diversity. However, many of these pathways are cryptic, and
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

20 S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
strategies to activate them are needed to explore the full chemical di-versity of such organisms.
Accessingmetabolites that are not expressed under standard growthconditions is challenging, but such resources are now attainable due tothe recent progress made in the fields of biology and analytical chemis-try as well as data mining and the possibility of applying these ap-proaches in combination.
These developments encompass the stimulation of endogenous me-tabolite production through the so-called ‘cryptic pathways’ via varioustreatments or combining superior genetic engineering approaches withsynthetic chemistry to generate chemical polymorphism. In this con-text, the various recent papers described in this review have shownthat microorganisms can efficiently activate the de novo production orupregulation of a wide range of chemical agents to compete withother microorganisms for nutrient or space when they are co-culturedin artificial conditions. Despite some promising findings, further workneeds to be carried out to verify whether microorganism co-culturepreferentially yields new molecular skeletons or derivatives of knowncompounds.
The investigation of induction phenomena in interactingmicrobes isextremely challenging due to the complex nature and chemical diversi-ty of their metabolomes. Furthermore, microbial metabolomes in co-cultures generate chemical variability that should be accuratelyassessed to highlight the compounds of interest resulting from suchinteractions. This requires sophisticated and sensitive analytical ap-proaches,mainly based on high-resolutionmass spectrometry for eitherextract profiling or direct MS imaging of the co-cultures. Relevant andreproducible biomarkers of microbial interactions are progressivelybeing identified through advanced data mining methods that are capa-ble of tracking subtle induction phenomena. As shown in this review,various approaches havemade this task possible, and innovative strate-gies to efficiently perform such analyses are now at our disposal.
However, particular efforts should be made to identify biomarkersand characterize their bioactivity, even when an efficient dereplicationprotocol based on high-resolution MS or MS/MS is available. Neverthe-less, the accuracy of structural assignment using such methods isstrongly dependent on the availability of databases or standards for mi-crobial metabolites. Depending on the strains examined, the quality ofpeak annotation and biomarker identification may vary greatly.Furthermore, the hit rate in NP databases is low for de novo-inducedmetabolites, indicating a potential to yield novel structures. To achievethe identification of induced metabolites, strategies involving LC–MS-targeted microfractionation and de novo structure assignment basedon sensitive NMR approaches are therefore mandatory. Some of theseapproaches have been illustrated here and provide appropriatemethods to assess the biological activity of the isolated compounds,which is critical to the process of understanding the relevance of the ob-served induction phenomena.
The study of complex microbial interactions has become feasiblethrough combinations of the most recent biotechnological and analyti-cal methods. This promising approach constitutes a strategic methodto highlight novel structures and, possibly, new skeletons with relevantbiological activity and potentially new modes of action in the field ofdrug discovery. These investigations are keys to understanding thechemical events that govern interactions between microorganisms indefined ecological niches from a fundamental perspective. Consideringthe exponential increase in the number of papers on this topic thathave been published in the last several years, it is safe to conclude thatthe use of this approach will expand rapidly and yield important andfascinating discoveries.
Acknowledgments
This work was supported by the Swiss National Science FoundationSinergia Grant CRSII3_127187 and Grant CR23I3_143733/1, whichwere awarded to J.-L.W. and K.G.
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
We gratefully acknowledge the first nine Grands Crus de Bordeaux(Bordeaux, France) for their financial support for the salary of Dr.Sylvain Schnee: château Ausone, château Cheval blanc, château Haut-Brion, château Lafitte Rothschild, château Latour, château Margaux,château Mouton Rothschild, château Petrus and château d'Yquem. Wealso thank Pierre-Marie Allard for helpful discussion on epigeneticmod-ification and Sarah Berndt, Nurhuda Manshoor, Jeroen J. Jansen andMariam Mnatsakanyan for revising the manuscript.
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.biotechadv.2014.03.001.
References
Adelin E, Slimani N, Cortial S, Schmitz-Alfonso I, Ouazzani J. Platotex: an innovative andfully automated device for cell growth scale-up of agar-supported solid-state fermen-tation. J Ind Microbiol Biotechnol 2011;38:299–305.
Ahuja I, Kissen R, Bones AM. Phytoalexins in defense against pathogens. Trends Plant Sci2012;17:73–90.
Akondi KB, Lakshmi VV. Emerging trends in genomic approaches for microbialbioprospecting. OMICS 2013;17:61–70.
Ali-Vehmas T, Rizzo A, Westermarck T, Atroshi F. Measurement of antibacterial activitiesof T-2 toxin, deoxynivalenol, ochratoxin A, aflatoxin B1 and fumonisin B1 usingmicrotitration tray-based turbidimetric techniques. J Vet Med Ser A 1998;45:453–8.
An S, Couteau C, Luo F, Neveu J, DuBow MS. Bacterial diversity of surface sand samplesfrom the Gobi and Taklamaken deserts. Microb Ecol 2013;66:850–60.
Angell S, Bench BJ, Williams H, Watanabe CMH. Pyocyanin isolated from a marine micro-bial population: synergistic production between two distinct bacterial species andmode of action. Chem Biol 2006;13:1349–59.
Bass D, Richards TA. Three reasons to re-evaluate fungal diversity on Earth and in theocean. Fungal Biol Rev 2011;25:159–64.
Beck JM, Young VB, Huffnagle GB. The microbiome of the lung. Transl Res 2012;160:258–66.
Berdy J. Thoughts and facts about antibiotics: where we are now and where we are head-ing. J Antibiot 2012;65:385–95.
Bergmann S, Schumann J, Scherlach K, Lange C, Brakhage AA, Hertweck C. Genomics-driven discovery of PKS-NRPS hybrid metabolites from Aspergillus nidulans. NatChem Biol 2007;3:213–7.
Bertrand S, Azzollini A, Schumpp O, Bohni N, Monod M, Gindro K, et al. Fungal co-cultureas a new source of bioactive induced metabolites: a MS-based metabolomic study.Planta Med 2012;78:CL7.
Bertrand S, Hélesbeux J-J, Larcher G, Duval O. Hydroxamate, a key pharmacophoreexhibiting a wide range of biological activities. Mini Rev Med Chem 2013a;13:1311–26.
Bertrand S, Petit C, Marcourt L, Ho R, Gindro K, MonodM, et al. HPLC profiling with at-linemicrodilution assay for the early identification of antifungal compounds in plantsfrom French Polynesia. Phytochem Anal 2013b;25:106–12.
Bertrand S, Schumpp O, Bohni N, Bujard A, Azzollini A, Monod M, et al. Detection of me-tabolite induction in fungal co-cultures on solid media by high-throughput differen-tial ultra-high pressure liquid chromatography-time-of-flight mass spectrometryfingerprinting. J Chromatogr A 2013c;1292:219–28.
Bertrand S, Schumpp O, Bohni N, Monod M, Gindro K, Wolfender J-L. De novo productionof metabolites by fungal co-culture of Trichophyton rubrum and Bionectria ochroleuca.J Nat Prod 2013d;76:1157–65.
Bhardwaj C, Cui Y, Hofstetter T, Liu SY, Bernstein HC, Carlson RP, et al. Differentiation ofmicrobial species and strains in coculture biofilms by multivariate analysis of laserdesorption postionization mass spectra. Analyst 2013;138:6844–51.
Bills GF, Gloer JB, An Z. Coprophilous fungi: antibiotic discovery and functions in anunderexplored arena of microbial defensive mutualism. Curr Opin Microbiol 2013;16:549–65.
Blackwell M. The fungi: 1, 2, 3 … 5.1 million species? Am J Bot 2011;98:426–38.Boccard J, Veuthey J-L, Rudaz S. Knowledge discovery in metabolomics: an overview of
MS data handling. J Sep Sci 2010;33:290–304.Boddy L. Interspecific combative interactions between wood-decaying basidiomycetes.
FEMS Microbiol Ecol 2000;31:185–94.Boddy L, Rayner ADM. Ecological roles of basidiomycetes forming decay communities in
attached oak branches. New Phytol 1983;93:77–88.Bode HB, Zeeck A. Sphaerolone and dihydrosphaerolone, two bisnaphthyl-pigments from
the fungus Sphaeropsidales sp. F-24'707. Phytochemistry 2000a;54:597–601.Bode HB, Zeeck A. UV mutagenesis and enzyme inhibitors as tools to elucidate the late
biosynthesis of the spirobisnaphthalenes. Phytochemistry 2000b;55:311–6.Bode HB, Walker M, Zeeck A. Structure and biosynthesis of mutolide, a novel macrolide
from a UV mutant of the fungus F-24'707. Eur J Org Chem 2000a;2000:1451–6.Bode HB, Wegner B, Zeeck A. Biosynthesis of cladospirone bisepoxide, a member of the
spirobisnaphthalene family. J Antibiot 2000b;53:153–7.Bode HB, Bethe B, Höfs R, Zeeck A. Big effects from small changes: possible ways to
explore nature's chemical diversity. Chembiochem 2002;3:619–27.Bohni N, Cordero-Maldonado ML, Maes J, Siverio-Mota D, Marcourt L, Munck S, et al. In-
tegration of microfractionation, qNMR and zebrafish screening for the in vivo
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

21S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
bioassay-guided isolation and quantitative bioactivity analysis of natural products.PLoS One 2013a;8:e64006.
Bohni N, Schumpp O, Schnee S, Bertrand S, Gindro K, Wolfender J-L. Targeted isolation ofinduced and bioactive metabolites from fungal co-cultures. Planta Med 2013b;79:SL41.
Bok JW, Keller NP. LaeA, a regulator of secondary metabolism in Aspergillus spp. EukaryotCell 2004;3:527–35.
Bok JW, Hoffmeister D, Maggio-Hall LA, Murillo R, Glasner JD, Keller NP. Genomic miningfor Aspergillus natural products. Chem Biol 2006;13:31–7.
Bok JW, Chiang Y-M, Szewczyk E, Reyes-Dominguez Y, Davidson AD, Sanchez JF, et al.Chromatin-level regulation of biosynthetic gene clusters. Nat Chem Biol 2009;5:462–4.
Bonfante P, Anca I-A. Plants, mycorrhizal fungi, and bacteria: a network of interactions.Annu Rev Microbiol 2009;63:363–83.
Brakhage AA, Schroeckh V. Fungal secondary metabolites—strategies to activate silentgene clusters. Fungal Genet Biol 2011;48:15–22.
Brown M, Wedge DC, Goodacre R, Kell DB, Baker PN, Kenny LC, et al. Automatedworkflows for accurate mass-based putative metabolite identification in LC/MS-derived metabolomic datasets. Bioinformatics 2011;27:1108–12.
Bucar F, Wube A, Schmid M. Natural product isolation—how to get from biological mate-rial to pure compounds. Nat Prod Rep 2013;30:525–45.
Bugni TS, Richards B, Bhoite L, Cimbora D, Harper MK, Ireland CM. Marine natural productlibraries for high-throughput screening and rapid drug discovery. J Nat Prod 2008;71:1095–8.
Bull AT, Stach JEM. Marine actinobacteria: new opportunities for natural product searchand discovery. Trends Microbiol 2007;15:491–9.
Butler MS. Natural products to drugs: natural product derived compounds in clinicaltrials. Nat Prod Rep 2005;22:162–95.
Campbell WG. The chemistry of the white rots of wood. III. The effect on wood substanceof Ganoderma applanatum (Pers.) Pat., Fomes fomen-tarius (Linn.) Fr., Polyporusadustus (Willd.) Fr., Pleurotus ostreatus (Jacq.) Fr., Armillaria mellea (Vahl.) Fr.,Trametes pini (Brot.) Fr., and Polystictus abietinus (Dicks.) Fr. Biochem J 1932;26:1829–38.
Capece A, Siesto G, Romaniello R, Lagreca VM, Pietrafesa R, Calabretti A, et al. Assessmentof competition in wine fermentation among wild Saccharomyces cerevisiae strainsisolated from Sangiovese grapes in Tuscany region. LWT Food Sci Technol 2013;54:485–92.
Casieri L, Hofstetter V, Viret O, Gindro K. Fungal communities living in the wood ofdifferent cultivars of young Vitis vinifera plants. Phytopathol Mediterr 2009;48:73–83.
Chagas F, Dias L, Pupo M. A mixed culture of endophytic fungi increases production ofantifungal polyketides. J Chem Ecol 2013;39:1335–42.
Chandler JA, Lang JM, Bhatnagar S, Eisen JA, Kopp A. Bacterial communities of diverseDrosophila Species: ecological context of a host-microbe model system. PLoS Genet2011;7:e1002272.
Charusanti P, Fong NL, Nagarajan H, Pereira AR, Li HJ, Abate EA, et al. Exploiting adaptivelaboratory evolution of Streptomyces clavuligerus for antibiotic discovery and over-production. PLoS One 2012;7:e33727.
Cheng YF, Jin W, Mao SY, Zhu W-Y. Production of citrate by anaerobic fungi in the pres-ence of co-culture methanogens as revealed by 1H NMR spectrometry. AsianAustralas J Anim Sci 2013;26:1416–23.
Chi Y, Hatakka A, Maijala P. Can co-culturing of two white-rot fungi increase lignin deg-radation and the production of lignin-degrading enzymes? Int Biodeterior Biodegrad2007;59:32–9.
Chiang Y-M, Lee K-H, Sanchez JF, Keller NP, Wang CCC. Unlocking fungal cryptic naturalproducts. Nat Prod Commun 2009;4:1505–10.
Chiang Y-M, Chang S-L, Oakley BR, Wang CCC. Recent advances in awakening silentbiosynthetic gene clusters and linking orphan clusters to natural products inmicroor-ganisms. Curr Opin Chem Biol 2011;15:137–43.
Cho JY, Kim MS. Induction of antifouling diterpene production by Streptomycescinnabarinus PK209 in co-culture withmarine-derived Alteromonas sp. KNS-16. BiosciBiotechnol Biochem 2012;76:1849–54.
Cichewicz RH. Epigenome manipulation as a pathway to new natural product scaffoldsand their congeners. Nat Prod Rep 2010;27:11–22.
Combès A, Ndoye I, Bance C, Bruzaud J, Djediat C, Dupont J, et al. Chemical communicationbetween the endophytic fungus Paraconiothyrium variabile and the phytopathogenFusarium oxysporum. PLoS One 2012;7:e47313.
Cragg GM, Newman DJ. Natural products: a continuing source of novel drug leads.Biochim Biophys Acta Gen Subj 2013;1830:3670–95.
Craney A, Ozimok C, Pimentel-Elardo Sheila M, Capretta A, Nodwell Justin R. Chemicalperturbation of secondary metabolism demonstrates important links to primarymetabolism. Chem Biol 2012;19:1020–7.
Creek DJ, Jankevics A, Breitling R, Watson DG, Barrett MP, Burgess KEV. Toward globalmetabolomics analysis with hydrophilic interaction liquid chromatography–massspectrometry: improved metabolite identification by retention time prediction.Anal Chem 2011;83:8703–10.
Creek DJ, Nijagal B, Kim D-H, Rojas F, Matthews KR, Barrett MP. Metabolomics guides ra-tional development of a simplified cell culture medium for drug screening againstTrypanosoma brucei. Antimicrob Agents Chemother 2013;57:2768–79.
Cueto M, Jensen PR, Kauffman C, Fenical W, Lobkovsky E, Clardy J. Pestalone, a new anti-biotic produced by a marine fungus in response to bacterial challenge. J Nat Prod2001;64:1444–6.
Cui L, Morris A, Ghedin E. The human mycobiome in health and disease. Genome Med2013;5:63.
Dalisay DS, Molinski TF. NMR quantitation of natural products at the nanomole scale. J NatProd 2009;72:739–44.
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
Dalisay DS, Rogers EW, Edison AS, Molinski TF. Structure elucidation at the nanomolescale. 1. Trisoxazole macrolides and thiazole-containing cyclic peptides from thenudibranch Hexabranchus sanguineus. J Nat Prod 2009;72:732–8.
de los Santos-Villalobos S, Barrera-Galicia G, Miranda-Salcedo M, Peña-Cabriales J.Burkholderia cepacia XXVI siderophore with biocontrol capacity against Colletotrichumgloeosporioides. World J Microbiol Biotechnol 2012;28:2615–23.
De Roy K, Marzorati M, Van den Abbeele P, Van de Wiele T, Boon N. Synthetic microbialecosystems: an exciting tool to understand and apply microbial communities. Envi-ron Microbiol 2013. http://dx.doi.org/10.1111/462-2920.12343. [Epub Dec 16, 2013].
Degenkolb T, Heinze S, Schlegel B, Strobel G, Gräfe U. Formation of newlipoaminopeptides, acremostatins A, B, and C, by co-cultivation of Acremoniumsp. Tbp-5 and Mycogone rosea DSM 12973. Biosci Biotechnol Biochem 2002;66:883–6.
Delhaes L, Monchy S, Frealle E, Hubans C, Salleron J, Leroy S, et al. The airway microbiotain cystic fibrosis: a complex fungal and bacterial community-implications fortherapeutic management. PLoS One 2012;7:e36313.
Demain AL. Importance of microbial natural products and the need to revitalize theirdiscovery. J Ind Microbiol Biotechnol 2014;41:185–201.
Do Nascimento M, MdlA Dublan, Ortiz-Marquez JCF, Curatti L. High lipid productivity ofan Ankistrodesmus–Rhizobium artificial consortium. Bioresour Technol 2013;146:400–7.
Donlan RM. Biofilms: microbial life on surfaces. Emerg Infect Dis 2002;8:881–90.Donnelly DP, Boddy L. Mycelial dynamics during interactions between Stropharia caerulea
and other cord-forming, saprotrophic basidiomycetes. New Phytol 2001;151:691–704.
Douglas AE. Microbial brokers of insect–plant interactions revisited. J Chem Ecol 2013;39:952–61.
Drugs.com [Internet]. U.S. pharmaceutical sales—2013 [accessed Nov 1, 2013]. Availablefrom: http://www.drugs.com/stats/top100/2013/sales.
Du J, Zhou J, Xue J, Song H, Yuan Y. Metabolomic profiling elucidates community dynam-ics of the Ketogulonicigenium vulgare–Bacillus megaterium consortium. Metabolomics2012;8:960–73.
Eldridge GR, Vervoort HC, Lee CM, Cremin PA, Williams CT, Hart SM, et al. High-throughput method for the production and analysis of large natural product librariesfor drug discovery. Anal Chem 2002;74:3963–71.
El-Elimat T, Figueroa M, Ehrmann BM, Cech NB, Pearce CJ, Oberlies NH. High-resolutionMS, MS/MS, and UV database of fungal secondary metabolites as a dereplicationprotocol for bioactive natural products. J Nat Prod 2013;76:1709–16.
Eugster PJ, Guillarme D, Rudaz S, Veuthey J-L, Carrupt P-A, Wolfender J-L. Ultra high pres-sure liquid chromatography for crude plant extract profiling. J AOAC Int 2011;94:51–70.
Eugster PJ, Biass D, Guillarme D, Favreau P, Stöcklin R, Wolfender J-L. Peak capacity opti-misation for high resolution peptide profiling in complex mixtures by liquid chroma-tography coupled to time-of-flight mass spectrometry: application to the Conusconsors cone snail venom. J Chromatogr A 2012;1259:187–99.
Falconer RE, Bown JL, White NA, Crawford JW. Modelling interactions in fungi. J R SocInterface 2008;5:603–15.
Favre-Godal Q, Queiroz EF, Wolfender J-L. Latest developments in assessing antifungalactivity using TLC-bioautography: a review. J AOAC Int 2013;96:1175–88.
FeherM, Schmidt JM. Property distributions: differences between drugs, natural products,and molecules from combinatorial chemistry. J Chem Inf Comput Sci 2002;43:218–27.
Fekete S, Guillarme D. Possibilities of new generation columns packed with 1.3 μm core-shell particles in gradient elution mode. J Chromatogr A 2013;1320:86–95.
Felsenfeld G, Groudine M. Controlling the double helix. Nature 2003;421:448–53.Fleming A. On the antibacterial action of cultures of a penicillium, with special reference
to their use in the isolation of B. influenzae. Br J Exp Pathol 1929;10:226–36.Flournoy DJ. In vitro antimicrobial properties of deferoxamine mesylate. Eur J Clin
Microbiol Infect Dis 1991;10:597–8.Frankland JC. Fungal succession—unravelling the unpredictable. Mycol Res 1998;102:
1–15.Frey-Klett P, Burlinson P, Deveau A, Barret M, Tarkka M, Sarniguet A. Bacterial–fungal in-
teractions: hyphens between agricultural, clinical, environmental, and food microbi-ologists. Microbiol Mol Biol Rev 2011;75:583–609.
Fujii R, Minami A, Gomi K, Oikawa H. Biosynthetic assembly of cytochalasin backbone.Tetrahedron Lett 2013;54:2999–3002.
Fukuda I, Ito A, Hirai G, Nishimura S, Kawasaki H, Saitoh H, et al. Ginkgolic acid inhibitsprotein SUMOylation by blocking formation of the E1-SUMO intermediate. ChemBiol 2009;16:133–40.
Funari CS, Eugster PJ, Martel S, Carrupt P-A,Wolfender J-L, Silva DHS. High resolution ultrahigh pressure liquid chromatography–time-of-flight mass spectrometrydereplication strategy for the metabolite profiling of Brazilian Lippia species. JChromatogr A 2012;1259:167–78.
Gaiero JR, McCall CA, Thompson KA, Day NJ, Best AS, Dunfield KE. Inside the rootmicrobiome: bacterial root endophytes and plant growth promotion. Am J Bot2013;100:1738–50.
Galagan JE, Calvo SE, Cuomo C, Ma L-J, Wortman JR, Batzoglou S, et al. Sequencing ofAspergillus nidulans and comparative analysis with A. fumigatus and A. oryzae. Nature2005;438:1105–15.
Georgios AT, Helen GG, Elizabeth JW, Ian DW. Liquid chromatography–mass spectrome-try based global metabolite profiling: a review. Anal Chim Acta 2012;711:7–16.
Geris R, Simpson TJ. Meroterpenoids produced by fungi. Nat Prod Rep 2009;26:1063–94.
GhannoumMA, Jurevic RJ, Mukherjee PK, Cui F, Sikaroodi M, Naqvi A, et al. Characteriza-tion of the oral fungal microbiome (mycobiome) in healthy individuals. PLoS Pathog2010;6:e1000713.
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

22 S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
Giddings JC. Maximum number of components resolvable by gel filtration and otherelution chromatographic methods. Anal Chem 1967;39:1027–8.
Giddings L-A, Newman D. Microbial natural products: molecular blueprints for antitumordrugs. J Ind Microbiol Biotechnol 2013;40:1181–210.
Glauser G, Guillarme D, Grata E, Boccard J, Thiocone A, Carrupt P-A, et al. Optimized liquidchromatography–mass spectrometry approach for the isolation of minor stress bio-markers in plant extracts and their identification by capillary nuclear magnetic reso-nance. J Chromatogr A 2008;1180:90–8.
Glauser G, Gindro K, Fringeli J, De Joffrey J-P, Rudaz S, Wolfender J-L. Differentialanalysis of mycoalexins in confrontation zones of grapevine fungal pathogensby ultrahigh pressure liquid chromatography/time-of-flight mass spectrome-try and capillary nuclear magnetic resonance. J Agric Food Chem 2009;57:1127–34.
Gloer JB. The chemistry of fungal antagonism and defense. Can J Bot 1995;73:S1265–74.Gomes AT, Smania Júnior A, Seidel C, EdFA Smania, Honda NK, Roese FM, et al. Antibacte-
rial activity of orsellinates. Braz J Microbiol 2003;34:194–6.Gomez-Escribano JP, Bibb MJ. Heterologous expression of natural product biosynthetic
gene clusters in Streptomyces coelicolor: from genome mining to manipulation of bio-synthetic pathways. J Ind Microbiol Biotechnol 2014;41:425–31.
Goss RJM, Shankar S, Fayad AA. The generation of “unNatural” products: synthetic biologymeets synthetic chemistry. Nat Prod Rep 2012;29:870–89.
Gower JC, Lubbe S, Le Roux N. Understanding biplots. Chichester: Wiley; 2011.Grandjean J, Huls R. Structure de la pleurotine: une benzoquinone extraite de Pleurotus
griseus. Tetrahedron Lett 1974;15:1893–5.Griffith GS, Rayner ADM, Wildman HG. Extracellular metabolites and mycelial morpho-
genesis of Hypholoma fasciculare and Phlebia radiata (Hymenomycetes). NovaHedwigia 1994;59:311–29.
Gronquist M, Meinwald J, Eisner T, Schroeder FC. Exploring uncharted terrain in nature'sstructure space using capillary NMR spectroscopy: 13 Steroids from 50 fireflies. J AmChem Soc 2005;127:10810–1.
Gunatilaka AAL. Natural products from plant-associated microorganisms: distribution,structural diversity, bioactivity, and implications of their occurrence. J Nat Prod2006;69:509–26.
Guo H, Sun B, Gao H, Chen X, Liu S, Yao X, et al. Diketopiperazines from the Cordyceps-colonizing fungus Epicoccum nigrum. J Nat Prod 2009;72:2115–9.
Gürdeniz G, Kristensen M, Skov T, Dragsted LO. The effect of LC–MS data preprocessingmethods on the selection of plasma biomarkers in fed vs. fasted rats. Metabolites2012;2:77–99.
Hanley L, Zimmermann R. Light and molecular ions: the emergence of vacuumUV single-photon ionization in MS. Anal Chem 2009;81:4174–82.
Heilmann-Clausen J, Boddy L. Inhibition and stimulation effects in communities of wooddecay fungi: exudates from colonized wood influence growth by other species.Microb Ecol 2005;49:399–406.
Heinisch S, Rocca JL, Kolosky M. Computerized optimization of the mobile phasecomposition in gradient elution reversed-phase HPLC. Chromatographia 1990;29:482–8.
Hertweck C. The biosynthetic logic of polyketide diversity. Angew Chem Int Ed 2009;48:4688–716.
Hickert LR, PBd Souza-Cruz, Rosa CA, Ayub MAZ. Simultaneous saccharification andco-fermentation of un-detoxified rice hull hydrolysate by Saccharomyces cerevisiaeICV D254 and Spathaspora arborariae NRRL Y-48658 for the production of ethanoland xylitol. Bioresour Technol 2013;143C:112–6.
HirohamaM, Kumar A, Fukuda I, Matsuoka S, Igarashi Y, Saitoh H, et al. Spectomycin B1 asa novel SUMOylation inhibitor that directly binds to SUMO E2. ACS Chem Biol 2013;8:2635–42.
Ho CH, Piotrowski J, Dixon SJ, Baryshnikova A, CostanzoM, Boone C. Combining functionalgenomics and chemical biology to identify targets of bioactive compounds. Curr OpinChem Biol 2011;15:66–78.
Hofstetter V, Buyck B, Croll D, Viret O, Couloux A, Gindro K. What if esca disease of grape-vine were not a fungal disease? Fungal Divers 2012;54:51–67.
Hu J-F, Garo E, Yoo H-D, Cremin PA, Zeng L, Goering MG, et al. Application of capillary-scale NMR for the structure determination of phytochemicals. Phytochem Anal2005;16:127–33.
Huffnagle GB, Noverr MC. The emerging world of the fungal microbiome. TrendsMicrobiol 2013;21:334–41.
Humphris SN, Wheatley RE, Bruce A. The effects of specific volatile organic compoundsproduced by Trichoderma spp. on the growth of wood decay basidiomycetes.Holzforschung 2001;55:233–7.
Hur M, Campbell AA, Almeida-de-Macedo M, Li L, Ransom N, Jose A, et al. A globalapproach to analysis and interpretation of metabolic data for plant natural productdiscovery. Nat Prod Rep 2013;30:565–83.
Hyde KD, Soytong K. The fungal endophyte dilemma. Fungal Divers 2008;33:163–73.Hyde ER, Haarmann DP, Lynne AM, Bücheli SR, Petrosino JF. The living dead: bacterial
community structure of a cadaver at the onset and end of the blaot stage of decom-position. PLoS One 2013;8:e77733.
Ikediugwu FEO,Webster J. Antagonism between Coprinus heptemerus and other coproph-ilous fungi. Trans Br Mycol Soc 1970a;54:181–204.
Ikediugwu FEO, Webster J. Hyphal interference in a range of coprophilous fungi. Trans BrMycol Soc 1970b;54:205–10.
Ito T, Odake T, Katoh H, Yamaguchi Y, Aoki M. High-throughput profiling of microbial ex-tracts. J Nat Prod 2011;74:983–8.
Iwai S, Omura S. Culture conditions for screening of new antibiotics. J Antibiot 1982;35:123–41.
Jansen JJ, Blanchet I, Buydens LMC, van Dam NM, Bertrand S, Wolfender J-L. ProjectedOrthogonalized Chemical Encounter Monitoring (POChEMon). Dutch Computers inChemistry symposium, 5th July 2013. Amsterdam, Netherlands; 2013.
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
Jansen JJ, Blanchet L, Buydens LMC, Bertrand S, Wolfender J-L. Projected OrthogonalizedChemical Encounter Monitoring (POChEMon) for fungal interactions during co-culture. 2014:Unpublished results.
Jee H-J, Tang C-S, Ko W-H. Characterization of phytochemicals stimulatory to sexual re-production in Phytophthora cactorum and P. parasitica. Bot Bull Acad Sin 2002;43:203–10.
Jenkinson HF, Lamont RJ. Oral microbial communities in sickness and in health. TrendsMicrobiol 2005;13:589–95.
Jolliffe IT. Principal component analysis. New York: Springer; 2002.Jonkers W, Rodriguez Estrada A, Lee K, Breakspear A, May G, Kistler H. Metabolome and
transcriptome of the interaction between Ustilago maydis and Fusarium verticillioidesin vitro. Appl Environ Microbiol 2012;78:3656–67.
Junier P, Kim O-S, Imhoff JF, Witzel K-P, Hadas O. Effect of salinity on cyanobacterial com-munity composition along a transect from Fuliya spring into the water of LakeKinneret, Israel. Fundam Appl Limnol 2013;182:99–107.
Kadi N, Challis GL. Chapter 17 Siderophore biosynthesis: a substrate specificity assay fornonribosomal peptide synthetase‐independent siderophore synthetases involvingtrapping of acyl‐adenylate intermediates with hydroxylamine. In: HopwoodDA, editor. Methods in enzymology. San Diego: Academic Press; 2009. p. 431–57.
Kaeberlein T, Lewis K, Epstein SS. Isolating “uncultivable”microorganisms in pure culturein a simulated natural environment. Science 2002;296:1127–9.
Kalia VC. Quorum sensing inhibitors: an overview. Biotechnol Adv 2013;31:224–45.Keller NP, Turner G, Bennett JW. Fungal secondary metabolism—from biochemistry to
genomics. Nat Rev Microbiol 2005;3:937–47.Kind T, Fiehn O. Metabolomic database annotations via query of elemental compositions:
mass accuracy is insufficient even at less than 1 ppm. BMC Bioinformatics 2006;7:234.
Kind T, Fiehn O. Seven golden rules for heuristic filtering of molecular formulas obtainedby accurate mass spectrometry. BMC Bioinformatics 2007;8:105.
Kind T, Wohlgemuth G, Lee DY, Lu Y, Palazoglu M, Shahbaz S, et al. FiehnLib: mass spec-tral and retention index libraries for metabolomics based on quadrupole and time-of-flight gas chromatography/mass spectrometry. Anal Chem 2009;81:10038–48.
Kirschning A, Hahn F. Merging chemical synthesis and biosynthesis: a new chapter in thetotal synthesis of natural products and natural product libraries. Angew Chem Int Ed2012;51:4012–22.
Kirwan JA, Broadhurst DI, Davidson RL, ViantMR. Characterising and correcting batch var-iation in an automated direct infusion mass spectrometry (DIMS) metabolomicsworkflow. Anal Bioanal Chem 2013;405:5147–57.
Kiss L, Russell JC, Szentivanyi O, Xu X, Jeffries P. Biology and biocontrol potential ofAmpelomyces mycoparasites, natural antagonists of powderymildew fungi. BiocontrolSci Technol 2004;14:635–51.
König CC, Scherlach K, Schroeckh V, Horn F, Nietzsche S, Brakhage AA, et al. Bacterium in-duces cryptic meroterpenoid pathway in the pathogenic fungus Aspergillus fumigatus.Chembiochem 2013;14:938–42.
Konishi Y, Kiyota T, Draghici C, Gao J-M, Yeboah F, Acoca S, et al. Molecular formulaanalysis by an MS/MS/MS technique to expedite dereplication of natural products.Anal Chem 2006;79:1187–97.
Kooperman N, Ben-Dov E, Kramarsky-Winter E, Barak Z, Kushmaro A. Coral mucus-associated bacterial communities from natural and aquarium environments. FEMSMicrobiol Lett 2007;276:106–13.
Kossuga MH, Ferreira AG, Sette LD, Berlinck RGS. Two polyketides from a co-culture oftwo marine-derived fungal strains. Nat Prod Commun 2013;8:721–4.
Krchňák V, Zajíček J, Miller PA, Miller MJ. Selective molecular sequestration with concur-rent natural product functionalization and derivatization: from crude natural productextracts to a single natural product derivative in one step. J Org Chem 2011;76:10249–53.
Kuhl C, Tautenhahn R, Böttcher C, Larson TR, Neumann S. CAMERA: an integrated strategyfor compound spectra extraction and annotation of liquid chromatography/massspectrometry data sets. Anal Chem 2011;84:283–9.
Kumar RS, Moorthy IMG, Baskar R. Modeling and optimization of glutamic acid produc-tion usingmixed culture of Corynebacterium glutamicumNCIM2168 and Pseudomonasreptilivora NCIM2598. Prep Biochem Biotechnol 2013;43:668–81.
Kurosawa K, Ghiviriga I, Sambandan TG, Lessard PA, Barbara JE, Rha C, et al.Rhodostreptomycins, antibiotics biosynthesized following horizontal gene transferfrom Streptomyces padanus to Rhodococcus fascians. J Am Chem Soc 2008;130:1126–7.
Lam KS. New aspects of natural products in drug discovery. Trends Microbiol 2007;15:279–89.
Lawrence D, Fiegna F, Behrends V, Bundy JG, Phillimore AB, Bell T, et al. Species inter-actions alter evolutionary responses to a novel environment. PLoS Biol 2012;10:1–11.
Lawrey JD. The chemical ecology of lichen mycoparasites—a review. Can J Bot 1995;73:S603–8.
Lefevre F, Robe P, Jarrin C, Ginolhac A, Zago C, Auriol D, et al. Drugs from hidden bugs:their discovery via untapped resources. Res Microbiol 2008;159:153–61.
Leonard CA, Brown SD, Hayman JR. Random mutagenesis of the Aspergillus oryzaegenome results in fungal antibacterial activity. Int J Microbiol 2013;2013:901697.
Lepage P, Leclerc MC, Joossens M, Mondot S, Blottiere HM, Raes J, et al. A metagenomicinsight into our gut's microbiome. Gut 2013;62:146–58.
Li L, Hooi D, Chhabra SR, Pritchard D, Shaw PE. Bacterial N-acylhomoserine lactone-induced apoptosis in breast carcinoma cells correlated with down-modulation ofSTAT3. Oncogene 2004;23:4894–902.
Li C, Zhang J, Shao C, DingW, She Z, Lin Y. A new xanthone derivative from the co-culturebroth of two marine fungi (strain No. E33 and K38). Chem Nat Compd 2011;47:382–4.
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

23S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
López SN, Ramallo IA, Sierra MG, Zacchino SA, Furlan RLE. Chemically engineered extractsas an alternative source of bioactive natural product-like compounds. Proc Natl AcadSci U S A 2007;104:441–4.
Losada L, Ajayi O, Frisvad JC, Yu J, NiermanWC. Effect of competition on the production andactivity of secondary metabolites in Aspergillus species. Med Mycol 2009;47:S88–96.
Lutz MP, Feichtinger G, Défago G, Duffy B. Mycotoxigenic Fusarium and deoxynivalenolproduction repress chitinase gene expression in the biocontrol agent Trichodermaatroviride P1. Appl Environ Microbiol 2003;69:3077–84.
Maiya S, Grundmann A, Li X, Li S-M, Turner G. Identification of a hybrid PKS/NRPS re-quired for pseurotin A biosynthesis in the human pathogen Aspergillus fumigatus.Chembiochem 2007;8:1736–43.
Marmann A, Aly A, Lin W, Wang B, Proksch P. Co-cultivation—a powerful emerging toolfor enhancing the chemical diversity of microorganisms. Mar Drugs 2014;12:1043–65.
Martin G, Guggiari M, Bravo D, Zopfi J, Cailleau G, Aragno M, et al. Fungi, bacteria and soilpH: the oxalate–carbonate pathway as a model for metabolic interaction. EnvironMicrobiol 2012;14:2960–70.
Masuma R, Tanaka Y, Tanaka H, Omura S. Production of nanaomycin and other antibioticsby phosphate-depressed fermentation using phosphate-trapping agents. J Antibiot1986;39:1557–64.
McGrath KC, Mondav R, Sintrajaya R, Slattery B, Schmidt S, Schenk PM. Development ofan environmental functional gene microarray for soil microbial communities. ApplEnviron Microbiol 2010;76:7161–70.
Mela F, Fritsche K, de Boer W, van Veen JA, de Graaff LH, van den Berg M, et al. Dual tran-scriptional profiling of a bacterial/fungal confrontation: Collimonas fungivorans versusAspergillus niger. ISME J 2011;5:1494–504.
Mendes R, Garbeva P, Raaijmakers JM. The rhizosphere microbiome: significance of plantbeneficial, plant pathogenic, and human pathogenic microorganisms. FEMSMicrobiolRev 2013;37:634–63.
Methe BA, Nelson KE, Pop M, Creasy HH, Giglio MG, Huttenhower C, et al. A frameworkfor human microbiome research. Nature 2012;486:215–21.
Miller RR, Montoya V, Gardy JL, Patrick DM, Tang P. Metagenomics for pathogen detectionin public health. Genome Med 2013;5:81.
Minerdi D, Bossi S, Gullino ML, Garibaldi A. Volatile organic compounds: a potential directlong-distance mechanism for antagonistic action of Fusarium oxysporum strain MSA35. Environ Microbiol 2009;11:844–54.
Misra JK. Trichomycetes—fungi associated with arthropods: review and world literature.Symbiosis 1998;24:179–219.
Mitova MI, Murphy AC, Lang G, Blunt JW, Cole ALJ, Ellis G, et al. Evolving trends in thedereplication of natural product extracts. 2. The isolation of chrysaibol, an antibioticpeptaibol from a New Zealand sample of the mycoparasitic fungus Sepedoniumchrysospermum. J Nat Prod 2008;71:1600–3.
Monod M, Bontems O, Zaugg C, Lechenne B, Fratti M, Panizzon R. Fast and reliable PCR/sequencing/RFLP assay for identification of fungi in onychomycoses. J Med Microbiol2006;55:1211–6.
Moree WJ, Phelan VV, Wu C-H, Bandeira N, Cornett DS, Duggan BM, et al. Interkingdommetabolic transformations captured by microbial imaging mass spectrometry. ProcNatl Acad Sci U S A 2012;109:13811–6.
Moree WJ, Yang JY, Zhao X, Liu W-T, Aparicio M, Atencio L, et al. Imaging mass spectrom-etry of a coral microbe interaction with fungi. J Chem Ecol 2013;39:1045–54.
Müller MEH, Steier I, Köppen R, Siegel D, Proske M, Korn U, et al. Co-cultivation of phyto-pathogenic Fusarium and Alternaria strains affects fungal growth and mycotoxin pro-duction. J Appl Microbiol 2012;113:874–87.
Nett M, Ikeda H,Moore BS. Genomic basis for natural product biosynthetic diversity in theactinomycetes. Nat Prod Rep 2009;26:1362–84.
Newman DJ, Cragg GM. Natural products as sources of new drugs over the 30 years from1981 to 2010. J Nat Prod 2012;75:311–35.
Nguyen Q-T, Merlo ME, Medema MH, Jankevics A, Breitling R, Takano E. Metabolomicsmethods for the synthetic biology of secondary metabolism. FEBS Lett 2012;586:2177–83.
Nielsen MN, Sørensen J, Fels J, Pedersen HC. Secondary metabolite- and endochitinase-dependent antagonism toward plant-pathogenic microfungi of Pseudomonas fluorescensisolates from sugar beet rhizosphere. Appl Environ Microbiol 1998;64:3563–9.
Nielsen TH, Christophersen C, Anthoni U, Sørensen J. Viscosinamide, a new cyclicdepsipeptide with surfactant and antifungal properties produced by Pseudomonasfluorescens DR54. J Appl Microbiol 1999;87:80–90.
Nielsen KF, Månsson M, Rank C, Frisvad JC, Larsen TO. Dereplication of microbial naturalproducts by LC-DAD-TOFMS. J Nat Prod 2011;74:2338–48.
Nonaka K, Abe T, Iwatsuki M, Mori M, Yamamoto T, Shiomi K, et al. Enhancement ofmetabolites productivity of Penicillium pinophilum FKI-5653, by co-culture withTrichoderma harzianum FKI-5655. J Antibiot 2011;64:769–74.
Notz R, Maurhofer M, Dubach H, Haas D, Defago G. Fusaric acid-producing strains ofFusarium oxysporum alter 2,4-diacetylphloroglucinol biosynthetic gene expressionin Pseudomonas fluorescens CHA0 in vitro and in the rhizosphere of wheat. Appl Envi-ron Microbiol 2002;68:2229–35.
Ochi K, Hosaka T. New strategies for drug discovery: activation of silent or weaklyexpressed microbial gene clusters. Appl Microbiol Biotechnol 2012;97:87–98.
O'Connor PB. MALDI mass spectrometry instrumentation. In: Hillenkamp F, Peter-Katalinic J, editors. MALDI-MS. Weinheim: Wiley-VCH Verlag GmbH & Co. KGaA;2007. p. 29–82.
Oh D-C, Jensen PR, Kauffman CA, Fenical W. Libertellenones A–D: induction of cytotoxicditerpenoid biosynthesis by marine microbial competition. Bioorg Med Chem 2005;13:5267–73.
Oh D-C, Kauffman CA, Jensen PR, Fenical W. Induced production of emericellamides A andB from the marine-derived fungus Emericella sp. in competing co-culture. J Nat Prod2007;70:515–20.
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
Ojika M, Molli SD, Kanazawa H, Yajima A, Toda K, Nukada T, et al. The secondPhytophthora mating hormone defines interspecies biosynthetic crosstalk. NatChem Biol 2011;7:591–3.
Ola ARB, Thomy D, Lai D, Brötz-Oesterhelt H, Proksch P. Inducing secondary metaboliteproduction by the endophytic fungus Fusarium tricinctum through coculture withBacillus subtilis. J Nat Prod 2013;76:2094–9.
Oldroyd GED. Speak, friend, and enter: signalling systems that promote beneficial symbi-otic associations in plants. Nat Rev Microbiol 2013;11:252–63.
Oliveira RPdS, Perego P, de Oliveira MN, Converti A. Growth, organic acids profile andsugar metabolism of Bifidobacterium lactis in co-culture with Streptococcusthermophilus: the inulin effect. Food Res Int 2012;48:21–7.
Omura S. The search for bioactive compounds frommicroorganisms. New York: Springer;1992.
Onaka H, Mori Y, Igarashi Y, Furumai T. Mycolic acid-containing bacteria induce natural-product biosynthesis in Streptomyces species. Appl EnvironMicrobiol 2011;77:400–6.
Park HB, KwonHC, Lee C-H, Yang HO. Glionitrin A, an antibiotic–antitumormetabolite de-rived from competitive interaction between abandoned mine microbes. J Nat Prod2009;72:248–52.
Pascolutti M, Quinn RJ. Natural products as lead structures: chemical transformations tocreate lead-like libraries. Drug Discov Today 2013;19:215–21.
Paul C, Mausz MA, Pohnert G. A co-culturing/metabolomics approach to investigatechemically mediated interactions of planktonic organisms reveals influence of bacte-ria on diatom metabolism. Metabolomics 2012;9:349–59.
Pearce C, Eckard P, Gruen-Wollny I, Hansske FG. Chapter 7 Microorganisms: their role inthe discovery and development ofmedicines. Natural product chemistry for drug dis-covery. Cambridge: The Royal Society of Chemistry; 2009. p. 215–41.
Peiris D, DunnW, Brown M, Kell D, Roy I, Hedger J. Metabolite profiles of interacting my-celial fronts differ for pairings of the wood decay basidiomycete fungus, Stereumhirsutum with its competitors Coprinus micaceus and Coprinus disseminatus. Metabo-lomics 2008;4:52–62.
Percival SL, Bowler PG. Biofilms and their potential role in wound healing. Wounds 2004;16:234–40.
Petrosino JF, Highlander S, Luna RA, Gibbs RA, Versalovic J. Metagenomic pyrosequencingand microbial identification. Clin Chem 2009;55:856–66.
Pettit R. Mixed fermentation for natural product drug discovery. Appl MicrobiolBiotechnol 2009;83:19–25.
Pettit RK. Small-molecule elicitation of microbial secondary metabolites. MicrobBiotechnol 2011;4:471–8.
Pevarello P. Recent drug approvals from the US FDA and EMEA: what the future holds.Future Med Chem 2009;1:35–48.
Piel J. Approaches to capturing and designing biologically active small molecules pro-duced by uncultured microbes. Annu Rev Microbiol 2011;65:431–53.
Potterat O, Hamburger M. Concepts and technologies for tracking bioactive compounds innatural product extracts: generation of libraries, and hyphenation of analytical pro-cesses with bioassays. Nat Prod Rep 2013;30:546–64.
Prosser JI, Tough AJ. Growth mechanisms and growth-kinetics of filamentous microor-ganisms. Crit Rev Biotechnol 1991;10:253–74.
Ranković B, Mišić M. The antimicrobial activity of the lichen substances of the lichensCladonia furcata, Ochrolechia androgyna, Parmelia caperata and Parmelia conspresa.Biotechnol Biotechnol Equip 2008;22:1013–6.
Rateb ME, Hallyburton I, HoussenW, Bull A, Goodfellow M, Santhanam R, et al. Inductionof diverse secondarymetabolites in Aspergillus fumigatus by microbial co-culture. RSCAdv 2013;3:14444–50.
Rayner A. Life in a collective—lessons from the fungi. New Sci 1988;120:49–53.Rayner ADM, Todd NK. Population and community structure and dynamics of fungi in
decaying wood. In: Woolhouse HW, editor. Advances in botanical research. London:Academic Press; 1980. p. 333–420.
Rico-Gray V. Interspecific interaction. Chichester: John Wiley & Sons Ltd.; 2001.Ridaura VK, Faith JJ, Rey FE, Cheng J, Duncan AE, Kau AL, et al. Gut microbiota from twins
discordant for obesity modulate metabolism in mice. Science 2013;341:6150.Rigali S, Titgemeyer F, Barends S, Mulder S, Thomae AW, Hopwood DA, et al. Feast or fam-
ine: the global regulator DasR links nutrient stress to antibiotic production by Strep-tomyces. EMBO Rep 2008;9:670–5.
Roach PJ, Laskin J, Laskin A. Nanospray desorption electrospray ionization: an ambientmethod for liquid-extraction surface sampling in mass spectrometry. Analyst 2010;135:2233–6.
Rodriguez Estrada AE, Hegeman A, May G. In vitro interactions between Fusariumverticillioides and Ustilago maydis through real-time PCR andmetabolic profiling. Fun-gal Genet Biol 2011;48:874–85.
Roemer T, Xu D, Singh Sheo B, Parish Craig A, Harris G, Wang H, et al. Confronting thechallenges of natural product-based antifungal discovery. Chem Biol 2011;18:148–64.
Romling U. Microbiology: bacterial communities as capitalist economies. Nature 2013;497:321–2.
Römpp A, Spengler B.Mass spectrometry imagingwith high resolution inmass and space.Histochem Cell Biol 2013;139:759–83.
Rosén J, Gottfries J, Muresan S, Backlund A, Oprea TI. Novel chemical space exploration vianatural products. J Med Chem 2009;52:1953–62.
Ross EM, Moate PJ, Marett LC, Cocks BG, Hayes BJ. Metagenomic predictions: frommicrobiome to complex health and environmental phenotypes in humans and cattle.PLoS One 2013;8:e73056.
Ross-Davis AL, Stewart JE, Shaw JD, Kim MS, Klopfenstein NB. Metagenomic approachesfor surveying forest soil microbial communities on permanent plots. Phytopathology2013;103:123.
Rosselló-Mora R, Amann R. The species concept for procaryotes. FEMS Microbiol Rev2001;25:39–67.
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

24 S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
Saccenti E, Hoefsloot HJ, Smilde A,Westerhuis J, Hendriks MWB. Reflections on univariateand multivariate analysis of metabolomics data. Metabolomics 2013. http://dx.doi.org/10.1007/s11306-013-0598-6. [Epub Oct 26, 2013].
Sachse K. Specificity and performance of PCR detection assays for microbial pathogens.Mol Biotechnol 2004;26:61–79.
Sandland GJ, Rodgers JK, Minchella DJ. Interspecific antagonism and virulence in hosts ex-posed to two parasite species. J Invertebr Pathol 2007;96:43–7.
Schachtschabel D, David A, Menzel K-D, Schimek C,Wöstemeyer J, BolandW. Cooperativebiosynthesis of trisporoids by the (+) and (−) mating types of the zygomyceteBlakeslea trispora. Chembiochem 2008;9:3004–12.
Scherlach K, Hertweck C. Triggering cryptic natural product biosynthesis in microorgan-isms. Org Biomol Chem 2009;7:1753–60.
Scherlach K, Graupner K, Hertweck C.Molecular bacterial–fungal interactionswith impacton the environment, food and medicine. Annu Rev Microbiol 2013;67:375–97.
Schmitt EK, Moore CM, Krastel P, Petersen F. Natural products as catalysts for innovation:a pharmaceutical industry perspective. Curr Opin Chem Biol 2011;15:497–504.
Schneider P, Misiek M, Hoffmeister D. In vivo and in vitro production options for fungalsecondary metabolites. Mol Pharm 2008;5:234–42.
Schoeman MW, Webber JF, Dickinson DJ. The effect of diffusible metabolites ofTrichoderma harzianum on in vitro interactions between basidiomycete isolates attwo different temperature regimes. Mycol Res 1996;100:1454–8.
Schroeckh V, Scherlach K, Nützmann H-W, Shelest E, Schmidt-Heck W, Schuemann J,et al. Intimate bacterial–fungal interaction triggers biosynthesis of archetypalpolyketides in Aspergillus nidulans. Proc Natl Acad Sci U S A 2009;106:14558–63.
Schürch S, Gindro K, Schumpp O, MonodM, Verrier J, Bohni N, et al. Chemical warfare be-tween fungi: an arsenal of bioactive molecules. Agrarforschung Schweiz 2010;1:442–5.
Schwarzer D, Finking R, Marahiel MA. Nonribosomal peptides: from genes to products.Nat Prod Rep 2003;20:275–87.
Score AJ, Palfreyman JW, White NA. Extracellular phenoloxidase and peroxidase enzymeproduction during interspecific fungal interactions. Int Biodeterior Biodegrad 1997;39:225–33.
Segata N, Boernigen D, Tickle TL, Morgan XC, Garrett WS, Huttenhower C. Computationalmeta'omics for microbial community studies. Mol Syst Biol 2013;9:666.
Shanahan P, O'Sullivan DJ, Simpson P, Glennon JD, O'Gara F. Isolation of 2,4-diacetylphloroglucinol from a fluorescent Pseudomonad and investigation of physiolog-ical parameters influencing its production. Appl Environ Microbiol 1992;58:353–8.
Shank EA, Kolter R. New developments in microbial interspecies signaling. Curr OpinMicrobiol 2009;12:205–14.
Shearer CA. Fungal competition. Can J Bot 1995;73:S1259–64.Shih C-J, Chen P-Y, Liaw C-C, Lai Y-M, Yang Y-L. Bringing microbial interactions to light
using imaging mass spectrometry. Nat Prod Rep 2014. http://dx.doi.org/10.1039/c3np70091g. [EPub Jan 22, 2014].
Shin CS, Kim HJ, Kim MJ, Ju JY. Morphological change and enhanced pigment productionof Monascus when cocultured with Saccharomyces cerevisiae or Aspergillus oryzae.Biotechnol Bioeng 1998;59:576–81.
Simmons TL, Coates RC, Clark BR, Engene N, Gonzalez D, Esquenazi E, et al. Biosyntheticorigin of natural products isolated from marine microorganism–invertebrate assem-blages. Proc Natl Acad Sci U S A 2008;105:4587–94.
Smanski MJ, Peterson RM, Huang S-X, Shen B. Bacterial diterpene synthases: new oppor-tunities for mechanistic enzymology and engineered biosynthesis. Curr Opin ChemBiol 2012;16:132–41.
Smith AL, Nicolaou KC. The enediyne antibiotics. J Med Chem 1996;39:2103–17.Soliman SSM, Raizada MN. Interactions between co-habitating fungi elicit synthesis of
taxol from an endophytic fungus in host Taxus plants. Front Microbiol 2013;4:3.Soman AG, Gloer JB, Wicklow DT. Antifungal and antibacterial metabolites from a
sclerotium-colonizing isolate of Mortierella vinacea. J Nat Prod 1999;62:386–8.Sonnenbichler J, Bliestle IM, Peipp H, Holdenrieder O. Secondary fungal metabolites and
their biological activities, I. Isolation of antibiotic compounds from cultures ofHeterobasidion annosum synthesized in the presence of antagonistic fungi or hostplant cells. Biol Chem Hoppe Seyler 1989;370:1295–304.
Sonnenbichler J, Peipp H, Dietrich J. Secondary fungal metabolites and their biologicalactivities, III. Further metabolites from dual cultures of the antagonistic basidiomy-cetes Heterobasidion annosum and Gloeophyllum abietinum. Biol Chem Hoppe Seyler1993;374:467.
Sonnenbichler J, Dietrich J, Peipp H. Secondary fungal metabolites and their biologicalactivities, V. Investigations concerning the induction of the biosynthesis of toxicsecondary metabolites in basidiomycetes. Biol Chem Hoppe Seyler 1994;375:71–9.
Stanstrup J, Gerlich M, Dragsted LO, Neumann S. Metabolite profiling and beyond:approaches for the rapid processing and annotation of human blood serum massspectrometry data. Anal Bioanal Chem 2013;405:5037–48.
Straight PD, Fischbach MA, Walsh CT, Rudner DZ, Kolter R. A singular enzymaticmegacomplex from Bacillus subtilis. Proc Natl Acad Sci U S A 2007;104:305–10.
Sullivan TS, McBride MB, Thies JE. Soil bacterial and archaeal community composition re-flects high spatial heterogeneity of pH, bioavailable Zn, and Cu in a metalliferous peatsoil. Soil Biol Biochem 2013;66:102–9.
Summerbell RC, Cooper E, Bunn U, Jamieson F, Gupta AK. Onychomycosis: a critical study oftechniques and criteria for confirming the etiologic significance of nondermatophytes.Med Mycol 2005;43:39–59.
Szewczyk E, Chiang Y-M, Oakley CE, Davidson AD, Wang CCC, Oakley BR. Identificationand characterization of the asperthecin gene cluster of Aspergillus nidulans. ApplEnviron Microbiol 2008;74:7607–12.
Takahashi J, Teles A, Almeida Pinto Bracarense A, Gomes D. Classical and epigeneticapproaches to metabolite diversification in filamentous fungi. Phytochem Rev2013:1–17.
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
Takemoto M, Kawamura Y, HirohamaM, Yamaguchi Y, Handa H, Saitoh H, et al. Inhibitionof protein SUMOylation by davidiin, an ellagitannin from Davidia involucrata. JAntibiot 2014. http://dx.doi.org/10.1038/ja.2013.142. [EPub Jan 15, 2014].
Tang Y-J, Zhu L-W, Li H-M, Li D-S. Submerged culture of mushrooms in bioreactors—challenges, current state-of-the-art, and future prospects. Food Technol Biotechnol2007;45:221–9.
Thierry A, Maillard M-B, Richoux R, Kerjean J-R, Lortal S. Propionibacterium freudenreichiistrains quantitatively affect production of volatile compounds in Swiss cheese. Lait2005;85:57–74.
Tisch D, Schmoll M. Light regulation of metabolic pathways in fungi. Appl MicrobiolBiotechnol 2010;85:1259–77.
Tokimoto K, Fujita T, Takeda Y, Takaishi Y. Increased or induced formation of antifungalsubstances in cultures of Lentinus edodes by the attack of Trichoderma spp. Proc JpnAcad Ser B 1987;63:277–80.
Traxler MF, Watrous JD, Alexandrov T, Dorrestein PC, Kolter R. Interspecies interactionsstimulate diversification of the Streptomyces coelicolor secreted metabolome. mBio2013;4:e00459-13.
Tsao S-W, Rudd BAM, He X-G, Chang C-J, Floss HG. Identification of a red pigment fromStreptomyces coelicolor A3(2) as a mixture of prodigiosin derivatives. J Antibiot1985;38:128–31.
Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature2006;444:1027–131.
van Belkum A, Hermans PWM, Licciardello L, Stefani S, Grubb W, van Leeuwen W, et al.Polymerase chain reaction-mediated typing of microorganisms: tracking dissemina-tion of genes and genomes. Electrophoresis 1998;19:602–7.
Vaneechoutte M. DNA fingerprinting techniques for microorganisms—a proposal forclassification and nomenclature. Mol Biotechnol 1996;6:115–42.
Verhoeven AB, Durham-Colleran MW, Pierson T, Boswell WT, van Hoek ML. Francisellaphilomiragia biofilm formation and interaction with the aquatic protist Acanthamoebacastellanii. Biol Bull 2010;219:178–88.
Verrier J, Pronina M, Peter C, Bontems O, Fratti M, Salamin K, et al. Identification of infec-tious agents in onychomycoses by polymerase chain reaction-terminal restrictionfragment length polymorphism. J Clin Microbiol 2012;50:553–61.
Verrier J, Krähenbühl L, Bontems O, Fratti M, Salamin K, Monod M. Dermatophyte identi-fication in skin and hair samples using a simple and reliable nested polymerase chainreaction assay. Br J Dermatol 2013;168:295–301.
Vinaixa M, Samino S, Saez I, Duran J, Guinovart JJ, Yanes O. A guideline to univariate sta-tistical analysis for LC/MS-based untargeted metabolomics-derived data. Metabolites2012;2:775–95.
Wald P, Pitkanen S, Boddy L. Interspecific interactions between the rare tooth fungiCreolophus cirrhatus, Hericium erinaceus and H. coralloides and other wood decayspecies in agar and wood. Mycol Res 2004;108:1447–57.
Wang J-p, Lin W, Wray V, Lai D, Proksch P. Induced production of depsipeptides by co-culturing Fusarium tricinctum and Fusarium begoniae. Tetrahedron Lett 2013;54:2492–6.
Want EJ, Wilson ID, Gika H, Theodoridis G, Plumb RS, Shockcor J, et al. Global metabolicprofiling procedures for urine using UPLC-MS. Nat Protoc 2010;5:1005–18.
Watanabe T, Izaki K, Takahashi H. New polyenic antibiotics active against gram-positiveand -negative bacteria. I. Isolation and purification of antibiotics produced byGluconobacter sp. W-315. J Antibiot 1982a;35:1141–7.
Watanabe T, Izaki K, Takahashi H. New polyenic antibiotics active against Gram-positiveand -negative bacteria. II. Screening of antibiotic producers and taxonomical proper-ties of Gluconobacter sp. W-315. J Antibiot 1982b;35:1148–55.
Watanabe T, Sugiyama T, Takahashi M, Shima J, Yamashita K, Izaki K, et al. The structureof enacyloxin II, a novel linear polyenic antibiotic produced by Gluconobacter sp.W-315. Agric Biol Chem 1990;54:259–61.
Watrous J, Roach P, Alexandrov T, Heath BS, Yang JY, Kersten RD, et al. Mass spectral mo-lecular networking of living microbial colonies. Proc Natl Acad Sci U S A 2012;109:E1743–52.
Watrous J, Roach P, Heath B, Alexandrov T, Laskin J, Dorrestein PC. Metabolic profiling di-rectly from the Petri dish using nanospray desorption electrospray ionization imagingmass spectrometry. Anal Chem 2013a;85:10385–91.
Watrous JD, Phelan VV, Hsu C-C, Moree WJ, Duggan BM, Alexandrov T, et al. Microbialmetabolic exchange in 3D. ISME J 2013b;7:770–80.
Wei F, Hong Y, Liu J, Yuan J, FangW, Peng H, et al. Gongronella sp. induces overproductionof laccase in Panus rudis. J Basic Microbiol 2010;50:98–103.
Wei Q, Wang H, Chen Z, Lv Z, Xie Y, Lu F. Profiling of dynamic changes in the microbialcommunity during the soy sauce fermentation process. Appl Microbiol Biotechnol2013;97:9111–9.
Wheatley RE. The consequence of volatile organic compound mediated bacterial andfungal interactions. Antonie Van Leeuwenhoek 2002;81:357–64.
White NA, Boddy L. Extracellular enzyme localization during interspecific fungal interac-tions. FEMS Microbiol Lett 1992;98:75–9.
Wiemann P, Keller NP. Strategies for mining fungal natural products. J Ind MicrobiolBiotechnol 2014;41:301–13.
Wiklund S, Johansson E, Sjostrom L, Mellerowicz EJ, Edlund U, Shockcor JP, et al. Vi-sualization of GC/TOF–MS-based metabolomics data for identification of bio-chemically interesting compounds using OPLS class models. Anal Chem 2008;80:115–22.
Williams RB, Henrikson JC, Hoover AR, Lee AE, Cichewicz RH. Epigenetic remodeling ofthe fungal secondary metabolome. Org Biomol Chem 2008;6:1895–7.
Wilson ZE, Brimble MA. Molecules derived from the extremes of life. Nat Prod Rep 2009;26:44–71.
Winter JM, Behnken S, Hertweck C. Genomics-inspired discovery of natural products.Curr Opin Chem Biol 2011;15:22–31.
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001

25S. Bertrand et al. / Biotechnology Advances xxx (2014) xxx–xxx
Wodarz D. Evolutionary dynamics of giant viruses and their virophages. Ecol Evol 2013;3:2103–15.
Wolfender J-L. HPLC in natural product analysis: the detection issue. Planta Med 2009;75:719–34.
Wolfender J-L, Queiroz EF. New approaches for studying the chemical diversity of naturalresources and the bioactivity of their constituents. Chimia 2012;66:324–9.
Wolfender J-L, Marti G, Ferreira Queiroz E. Advances in techniques for profiling crude ex-tracts and for the rapid identification of natural products: dereplication, quality con-trol and metabolomics. Curr Org Chem 2010;14:1808–32.
Wolfender J-L, Rudaz S, Hae Choi Y, Kyong Kim H. Plant metabolomics: from holistic datato relevant biomarkers. Curr Med Chem 2013;20:1056–90.
Woodward S, Boddy L. Chapter 7 Interactions between saprotrophic fungi. In: Boddy L,Frankland JC, Van West P, editors. Ecology of saprotrophic basidiomycetes. London:Academic Press; 2008. p. 125–41.
Xie P, Ma M, Rateb ME, Shaaban KA, Yu Z, Huang S-X, et al. Biosynthetic potential-basedstrain prioritization for natural product discovery: a showcase for diterpenoid-producing Actinomycetes. J Nat Prod 2014;77:377–87.
Yang Y-L, Xu Y, Straight P, Dorrestein PC. Translating metabolic exchange with imagingmass spectrometry. Nat Chem Biol 2009;5:885–7.
Yang JY, Sanchez LM, Rath CM, Liu X, Boudreau PD, Bruns N, et al. Molecular networkingas a dereplication strategy. J Nat Prod 2013a;76:1686–99.
Yang X-L, Awakawa T, Wakimoto T, Abe I. Induced biosyntheses of a novel butyrophe-none and two aromatic polyketides in the plant pathogen Stagonospora nodorum.Nat Prod Bioprospect 2013b;3:141–4.
Yuen TK, Hyde KD, Hodgkiss IJ. Interspecific interactions among tropical and subtropicalfreshwater fungi. Microb Ecol 1999;37:257–62.
Zahn JA, Higgs RE, Hilton MD. Use of direct-infusion electrospray mass spectrometry toguide empirical development of improved conditions for expression of secondarymetabolites from actinomycetes. Appl Environ Microbiol 2001;67:377–86.
Please cite this article as: Bertrand S, et al, Metabolite induction via microodrug discovery, Biotechnol Adv (2014), http://dx.doi.org/10.1016/j.biotec
Zhang C-L, Zheng B-Q, Lao J-P, Mao L-J, Chen S-Y, Kubicek C, et al. Clavatol and patulinformation as the antagonistic principle of Aspergillus clavatonanicus, an endophyticfungus of Taxus mairei. Appl Microbiol Biotechnol 2008;78:833–40.
Zhou J, Ma Q, Yi H, Wang L, Song H, Yuan Y-J. Metabolome profiling reveals metaboliccooperation between Bacillus megaterium and Ketogulonigenium vulgare duringinduced swarm motility. Appl Environ Microbiol 2011;77:7023–30.
Zhu F, Lin Y. Marinamide, a novel alkaloid and its methyl ester produced by the applica-tion of mixed fermentation technique to two mangrove endophytic fungi from theSouth China Sea. Chin Sci Bull 2006;51:1426–30.
Zhu F, Hu G, Chen X, Yuan Y, Huang M, Sun H, et al. Studies on the secondary metabolitesproduced by mixed cultures of two epiphytic fungi isolated from a fruit of mangroveAvicennia marina. Zhongshan Daxue Xuebao Ziran Kexueban 2009;48:79–82.
Zhu F, Qin C, Tao L, Liu X, Shi Z, Ma X, et al. Clustered patterns of species origins of nature-derived drugs and clues for future bioprospecting. Proc Natl Acad Sci U S A 2011;108:12943–8.
Zhu F, Chen GY, Wu JS, Pan JH. Structure revision and cytotoxic activity of marinamideand its methyl ester, novel alkaloids produced by co-cultures of two marine-derived mangrove endophytic fungi. Nat Prod Res 2013;27:1960–4.
Znidarsic P, Pavko A. The morphology of filamentous fungi in submerged cultivations as abioprocess parameter. Food Technol Biotechnol 2001;39:237–52.
Zuck KM, Shipley S, Newman DJ. Induced production of N-formyl alkaloids from Aspergil-lus fumigatus by co-culture with Streptomyces peucetius. J Nat Prod 2011;74:1653–7.
Zuo H-L, Yang F-Q, HuangW-H, Xia Z-N. Preparative gas chromatography and its applica-tions. J Chromatogr Sci 2013;51:704–15.
rganism co-culture: A potential way to enhance chemical diversity forhadv.2014.03.001