mesozoic araucariaceae: morphology and systematic relationships
TRANSCRIPT

J. Plant Res. 107 : 493-502, 1994 Journal of Plant Research by The Botanical Society of Japan 1994
JPR Symposium
Mesozoic Araucariaceae : Morphology and Systematic Relationships
Ruth A. S t o c k e y
Department of Biological Sciences, University of Alberta, Edmonton, AB, Canada, T6G 2E9
The Southern Hemisphere conifer family Araucar- iaceae has a very restricted present day distribution, but was more widespread in the past. The genus Araucaria is represented by good fossil material in both hemi- spheres as early as the Jurassic, while Agathis is only known from the Southern Hemisphere beginning in the Cretaceous. Cuticle studies of extant araucarians have enabled accurate comparisons of fossil leaves to living taxa. Araucaria Section Bunya is represented by cones of several types in the Jurassic. In addition to these remains, a suite of araucarian cones showing affinities to several sections of the genus Araucaria have been de- scribed from England, Japan and North America. Evi- dence that fossil araucarian cones may have produced seeds with hypogeal germination is discussed in light of recent work on germination of extant bunya seedlings and the discovery of new fossil shoots from the Jurassic Morrison Formation of Utah.
Key words : Agathis-- Araucaria - - Araucariaceae - - Cuticle - - Germination - - Mesozoic
The Araucariaceae, a family of large, coniferous trees, contains the two extant genera Agathis Salisbury and Araucaria de Jussieu. While the genus Araucaria is confined today to the Southern Hemisphere, Agathis extends into Malaysia across the equator. Florin (1963) believed that the distribution of this genus probably re- flected a migration northwestward in "Plio-Pleistocene times". The genera Araucaria and Agathis today contain about 21 species each.
In 1982, I discussed our current state of knowledge of the fossil record of the family Araucariaceae (Stockey 1982). The present contribution serves to bring the last twelve years of research on the Mesozoic fossil record of this family together with work on the extant species to provide a more up-to-date summary of our knowledge of the Araucariaceae and its evolution.
Materials and Methods
Many of the permineralized fossils described here have been studied using the cellulose acetate peel technique (Joy et al. 1956). Cuticle preparations were made using
the chromium trioxide techniques of Alvin and Boulter (1974) and Stockey and Ko (1986).
Results
Form and organ genera Leaves : Many types of fossils have been referred to as
"araucarian": however, many of these are form genera that have doubtful affinites with the family (Stockey 1982). In particular, the identity of fossil leaves has been difficult. Harris (1979) pointed out the difficulty in assigning genera such as Brachyphyllum and Pagiophyllum to the Araucar- iaceae or other conifer families. Some species of Bra- chyphyllum such as B. mamillare Lindley and Hutton from the Jurassic of Yorkshire belong the the Araucariaceae based on a combination of leaf morphology, cuticular features, their association with araucarian cone scales and their attachment to pollen cones containing Araucar- iacites pollen (Kendall 1949). Recently, Brachyphyllum vulgare (Stopes and Fujii) Jeffrey leaves from the Upper Cretaceous of Hokkaido (Ohana and Kimura 1993) have been found in attachment to immature cones of Araucaria nihongii Stockey, Nishida et Nishida (T. Ohsawa, M. Ni- shida and H. Nishida, pers. commun., 1993). Other Bra- chyphyllum species, however, show affinities to the extinct conifer family, Cheirolepidiaceae (Harris 1979). The same is true for Pagiophyllum species (Harris 1979).
Without the actual attachment of cones to branches bearing leaves with the helically arranged imbricate leaves typical of many araucarians, cuticular remains are essential to determine affinities of leafy twigs. This work was begun by Florin (1931) using the light microscope and has been continued recently using the scanning electron microscope. Two recent studies by Stockey and Ko (1986) and Stockey and Atkinson (1993) have outlined the cuticular micromorphology of all the the extant species in the family.
The genus Agathis is characterized by broad, lan- ceolate to ovate leaves, with a distinct petiole, and usually tapering apices (Fig. 14). They are arranged oppositely on branches, and therefore, without cuticle
494 R.A. Stockey

Mesozoic Araucariaceae 495
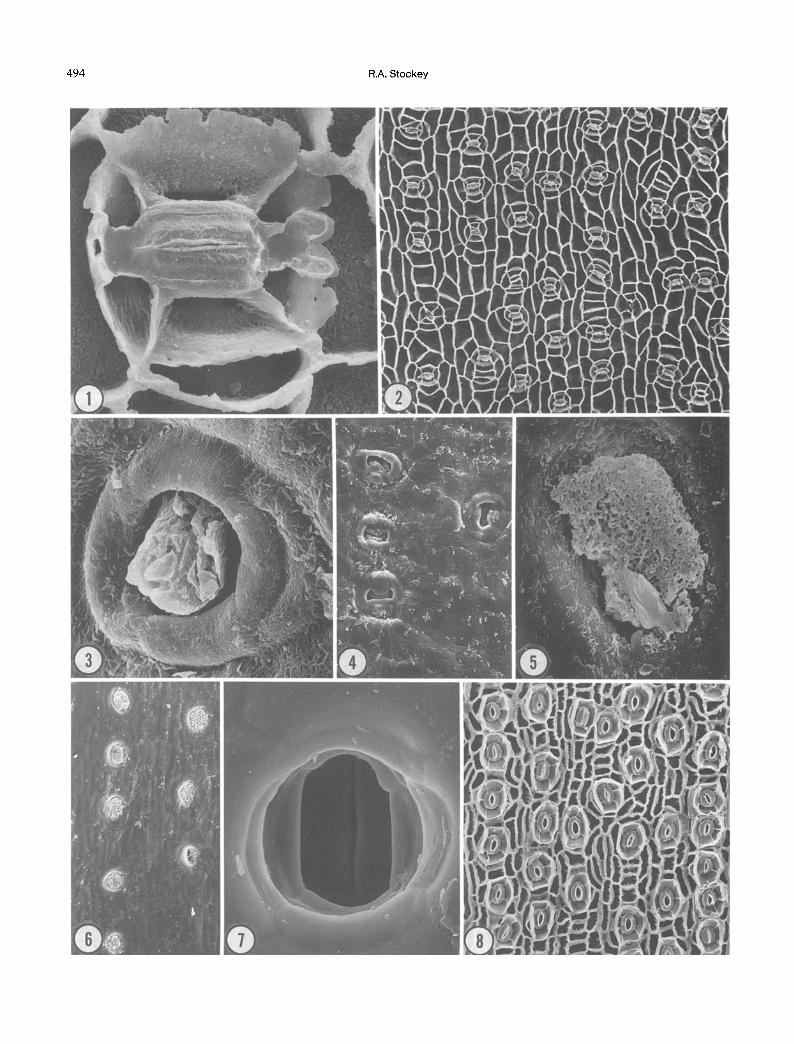
could be easily confused with those of the genus Nageia Gaertner (Podocarpaceae). If isolated or broken, they might also be confused with some of the broad leaved Araucaria species, in particular Araucaria bidwillfi or close- ly related fossil taxa. Cuticle micromorphology, however, distinguishes them from Nageia. Agathis leaf surfaces are undulating and exhibit stomatal plugs and Florin rings (Figs. 3, 4 ; Stockey and Atkinson 1993). Most species are hypostomatic. Stomata are in discontinuous rows and usually oriented perpendicularly or obliquely to the long axis of the leaf (Fig. 2). Four or five subsidiary cells are the common number but from three to nine have been observed (Fig. 1). Bilobed polar extensions of cuticle are characteristic of the genus as are variable-shaped epider- mal cells (Figs. 1, 2).
Nageia juvenile foliage, in particular, is similar to Agath- is (Stockey and Atkinson 1993). Leaves possess several veins, Florin rings, and from four to six subsidiary cells (Greenwood 1987, Hill and Pole 1992). Stomatal orienta- tions in Nageia are usually parallel to the long axis of the leaf, (Greenwood 1987, Hill and Pole 1992). Polar exten- sions are delicate, lacking the two cuticular nobs of Agathis. Thus, stomatal orientation and polar extension morphology are distinguishing features.
Cuticle of Araucaria is characterized by a lack of Florin rings or papillae on the outer cuticle surface (Figs. 5-7 ; Stockey and Ko 1986). Like Agathis, leaves of Araucaria have undulating cuticle surfaces and stomatal plugs (Figs. 5,6). Broad-leaved araucarians have discontinuous rows of stomata that are oriented parallel to the long axis of the leaf (Fig. 10), while araucarians with imbricate, rhomboidal leaves usually have oblique orientations. Four or five subsidiaries are common with from three to seven reported (Figs. 8, 9). Epidermal cell outlines are usually elongate and rectangular (Stockey and Ko 1986). Polar extensions often occur but do not show the bilobed structure seen in those of Agathis.
With the characterization of cuticle micromorphology of all of the species in both genera, we can now more meaningfully treat fossil leaves with cuticles in a tax- onomic framework. The oldest known Agathis leaves based on cuticular remains come from the Cretaceous (Albian) Otway Formation at Moonlight Head, Victoria, Australia (Cantrill 1992). These Agathis victoriensis leaves show impressions of Florin rings and a stomatal orientation similar to living Agathis.
Cantrill (1992) further discussed the micromorphology of fossil Araucaria from the Lower Cretaceous of Australia.
Although preservation of leaf cuticles in many instances was fragmentary, cuticular micromorphology was used to characterize several species. Araucaria seorsum Cantrill (1992) cannot be assigned to an extant section of the genus but shows characters of Eutacta, Intermedia and Araucaria (=Columbea). Araucaria lanceolatus Cantrill (1992) shows similarities to Sections Araucaria and Bunya. Araucaria acutifoliatus Cantrill (1992) shows similarities to Section Eutacta, but differences in stomatal distribution. Araucaria falcatus Cantrill and A. carinatus Cantrill, in contrast to many of the other species described by Cantrill (1992), show affinities to one section, Eutacta. The confi- dent placement of leaves into any extant section of the genus, thus, seems difficult with Mesozoic foliage, and is presumably an indication that evolutionary rates were more rapid in the family at this time in the past.
If we compare the Cretaceous reports with those of the Tertiary, we see a different pattern emerging. In a recent study of Australian Tertiary leaves, Hill (1990) reexamined the previously described species of fossil Araucaria and described four new species based on cuticular micromor- phology. Placement of these leaves into sections of the genus is easier than with Mesozoic fossils. In the Sec- tion Eutacta are: Araucaria lignitici Cookson and Duigan (1951), A. readiae Hill and Bigwood (1987), A. planus Hill (1990), A. prominens Hill (1990), and A. uncinatus Hill (1990). Section Araucaria (=Columbea) contains A. bal- combensis Selling (1950) and A. hastiensis Hill and Bigwood (1987). The eighth species, A. fimbriatus Hill (1990) cannot be placed into any of the sections of the genus with certainty suggesting that the genus was more diverse than at present time.
Cuticles of Araucarioides Bigwood and Hill (1985) from the Eocene of Tasmania come from leaves that are broad and elongate similar to Agathis. These leaves, however, are incomplete. Stomatal orientations are also similar to Agathis rather than broad-leaved Araucaria species. Two of the species described by Bigwood and Hill (1985), Araucarioides linearis and A. sinuosa do not, however, show Florin rings. A third species A. annulata, does show what are interpreted as rings that are not prominent. Stockey and Atkinson (1993) suggested that these three species externally resemble Araucaria but are similar to Agathis in stomatal orientation. Epidermal cell shapes are more similar to Araucaria than Agathis.
The existence of leaves in the Eocene fossil record that combine features of the two genera is intriguing, suggest- ing the presence of additional araucarian genera in the
Fig. 1. Agathis labillardieri juvenile foliage, abaxial cuticle, inner surface showing one stomatal apparatus with four subsidiary cells and bilobed polar extensions. •
Fig. 2. Agathis moorei, inner view, abaxial cuticle, showing stomatal rows. x100. Fig. 3. Agathis microstachya, outer view, abaxial surface showing Florin ring and stomatal plug. • Fig. 4. Agathis orbicula, outer view, abaxial surface showing undulating cuticle surface and Florin rings. X210. Fig. 5. Araucaria bidwillii, outer view, abaxial surface showing stomatal plug. X1,200. Fig. 6. Araucaria bidwillii, outer view, abaxial surface showing undulating cuticle and stomata with plugs lacking Florin rings or
papillae. X 200. Fig. 7. Araucaria hunsteinii, outer view, abaxial surface showing stomatal apparatus with plug removed. • Fig. 8. Araucaria angustifolia, inner view, abaxial surface showing stomatal rows and stomatal orientations nearly parallel to the
long axis of the leaf. •
496 R.A. Stockey

Mesozoic Araucariaceae 497
Mesozoic that have so far been described (Stockey 1990). A second taxon from the Cretaceous, Araucaria otwayen- sis Cantrill (1992) from Australia, is poorly preserved but shows polar extensions similar to those of Agathis, while leaf shaped is similar to species of Araucaria Section Eutacta. In addition to these Australian remains, leaves described recently as Dammarites coriacea (Barale 1992) are known from the Cretaceous of Spain. These strap- shaped leaves have decurrent bases and appear similar to some broad-leaved araucarians. A similar cuticular structure, at the light microscope level, has been observed on the bracts at the base of pollen cones from the same locality containing Araucariacites pollen. The name Dammarites Presl ex Sternberg was originally used for ovulate cones. Some of those described in the genus previously have now been transferred to the Taxodiaceae (Stockey 1982). The original type specimen could be araucarian; but it is a sandstone cast and difficult to interpret. Hlustik (1976) further reports that the original type specimen is not the specimen reported by Knobloch (1973) as the type, causing some confusion in the litera- ture. Undoubtedly, some of the material described as Dammarites albens or D. coriacea is araucarian. Whether the generic name Dammarites should be used for this material is not certain. I would suggest that some of it should be included in the genus Araucaria, but a closer investigation is necessary. Wood: Araucarian wood, usually described under the genus Araucarioxylon Kraus, has been reported from the lower Carboniferous to the Tertiary (Stockey 1982). Iso- lated, there is little one can do with wood of this type. It is difficult to distinguish from cordaite wood, usually treated in the genus Dadoxylon Endlicher. In addition to cordaites, a very similar wood was also borne by glos- sopterids (Taylor and Taylor 1993). In fact, woods of Araucaria and Agathis are so similar to one another that there is no way of distinguishing between the two genera (Greguss 1955, Jane 1970). Thus, the use of isolated twigs or logs to indicate the presence of Araucariaceae or its component genera in the fossil record is questionable. Pollen : The pollen type Araucariacites australis Cookson is widespread during the Jurassic, Cretaceous and Terti- ary in both the Northern and Southern Hemispheres (Hughes 1969). Again, as with wood, it is difficult to distinguish between Araucaria and Agathis based on pollen. Since Araucaria fossils are now known from the Northern Hemisphere it seems reasonable to believe that
most of this pollen belonged to Araucaria rather than to Agathis.
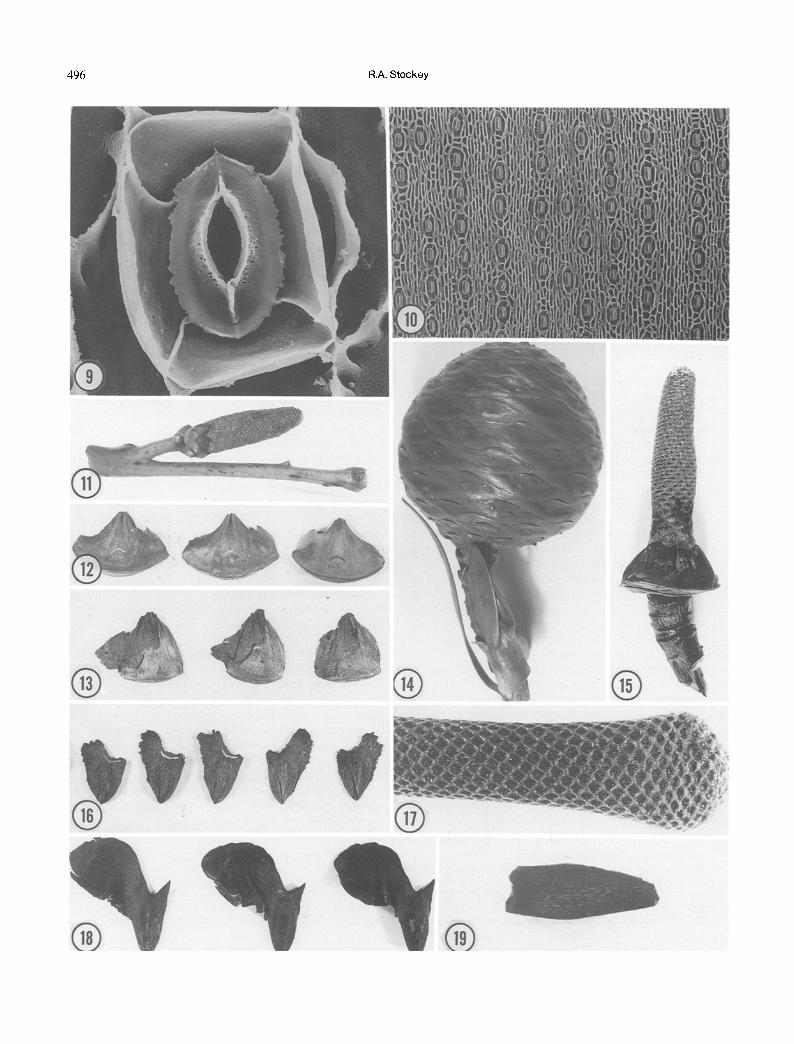
Pollen cones attributed to the Araucariaceae have not been extremely helpful in evolutionary studies (Stockey 1982). The large size of extant cones (Fig. 11) is not reflected in the meager fossil record, except for those cones associated with Dammarites coriaceous (Barale 1992). The small pollen cones of Brachyphyllum mamil- lare are borne terminally on the ends of leafy twigs (Ken- dall 1949) similar to those of Araucaria Section Eutacta (Wilde and Eames 1952). These remains are especially important as cuticles have also linked a cone-scale type with pollen cones and foliage (Kendall 1949, Harris 1979). Combined with other araucarian remains, attached cones can provide much more information about the plants in question. Ovulate cones: The best types of remains to indicate araucarian affinities based on isolated organs are those of ovulate cones. The typical araucarian cone is spherical to ovoid (Fig. 14) and sheds its cone-scale complexes at maturity (Figs. 12, 13, 15). Cones of Agathis show a fused bract and ovuliferous scale (Figs. 12, 14), unlike those of Araucaria which usually show a free ovuliferous scale tip (Eames 1913). The seeds of Agathis show a lateral wing near one side of the chalaza and a pointed projection on the other side of the chalaza (Figs. 13,16,18). These seeds separate from the cone-scale complex and can be dispersed by wind from the trees as cones disaggregate.
So far, cones of Agathis have not been demonstrated in the Mesozoic fossil record. Cone scales associated with the foliage type Podozamites lanceolatus Lindley et Hutton (=Agathis jurassica White) from the Jurassic Talbragar Fish Bed Flora in New South Wales are preser- ved only as impressions (White 1981). Stockey (1982) suggested that these and other similar scales (Fig. 28) show close similarities to those of Araucaria Section Eutacta rather than the genus Agathis.
The fossil record of Araucaria cones is much more complete than that of Agathis. Cones usually have been described under the genera Araucarites or Araucaria. The genus Araucaria is today divided into four sections : Araucaria ( =Columbea), Eutacta, Intermedia, and Bunya (Wilde and Eames 1952). These sections are distin- guished on characters of leaf morphology, attachment of pollen and ovulate cones, cone-scale morphology, vascu- lar system of the cone-scale complex, type of seedling germination and seedling morphology.
Fig. 9. Araucaria angustifolia, inner view, abaxial surface showing cuticle on stomatal apparatus with four subsidiary cells. X1,000.
Fig. 10. Araucaria bidwillii, inner view, abaxial surface showing stomatal rows and regular stomatal orientation. • Fig. 11. Fig. 12. Fig. 13. Fig. 14. Fig. 15. Fig. 16. Fig. 17. Fig. 18. Fig. 19.
Agathis pollen cone. X1. Agathis lanceolata cone scale complexes. • 0.9. Agathis lanceolata cone scale complexes with seeds on surfaces. • Agathis lanceolata ovulate cone on branch with leaves. X0.8. Agathis cone axis with scales removed. • Agathis lanceolata winged seeds removed from cone scales. • Araucaria hunsteinfi cone axis with scales removed. X2. Agathis moorei winged seeds removed from cone scales. • Fossil araucarian cone axis from the Jurassic Morrison Formation. Xl.
498 R.A. Stockey

Mesozoic Araucariaceae 499
The section Intermedia from New Guinea (White 1947) contains from one to three species. Most workers feel that these are closely related to the Eutacta species whose center of diversity is in New Caledonia. Fossil leaves similar to those of Section Intermedia have been reported from the Upper Cretaceous of New Zealand (Bose 1975). However, no associated cone remains have been reported.
The section Eutacta is the most widespread today in the South Pacific including Australia, New Caledonia and adjacent islands, Norfolk Island, and New Guinea (Florin 1963). The fossils described by Kendall (1947,1949) under Araucarites phill ipsfi cones and cone-scales and Brachyphyllum mamillare leaves and pollen cones are probably the remains of Araucaria Section Eutacta. In addition, Araucaria cutchensis (Feistmantel) Pant and Srivastava (1968) from the Lower Cretaceous of India can probably be included in this group (Stockey 1982).
The section Araucaria (=Columbea Endlicher emend. Wilde and Eames 1952), found in South America today, is represented by leaf remains, A. nathorstii Dusen (Menen- dez and Caccavari 1966) from the Tertiary of Rio Negro. These leaves show close similarities to extant A. araucana (Molina) K. Koch (=A. imbricata Pavon) based on cuticle. Until recently this group was not represented by cone fossils in the Mesozoic.
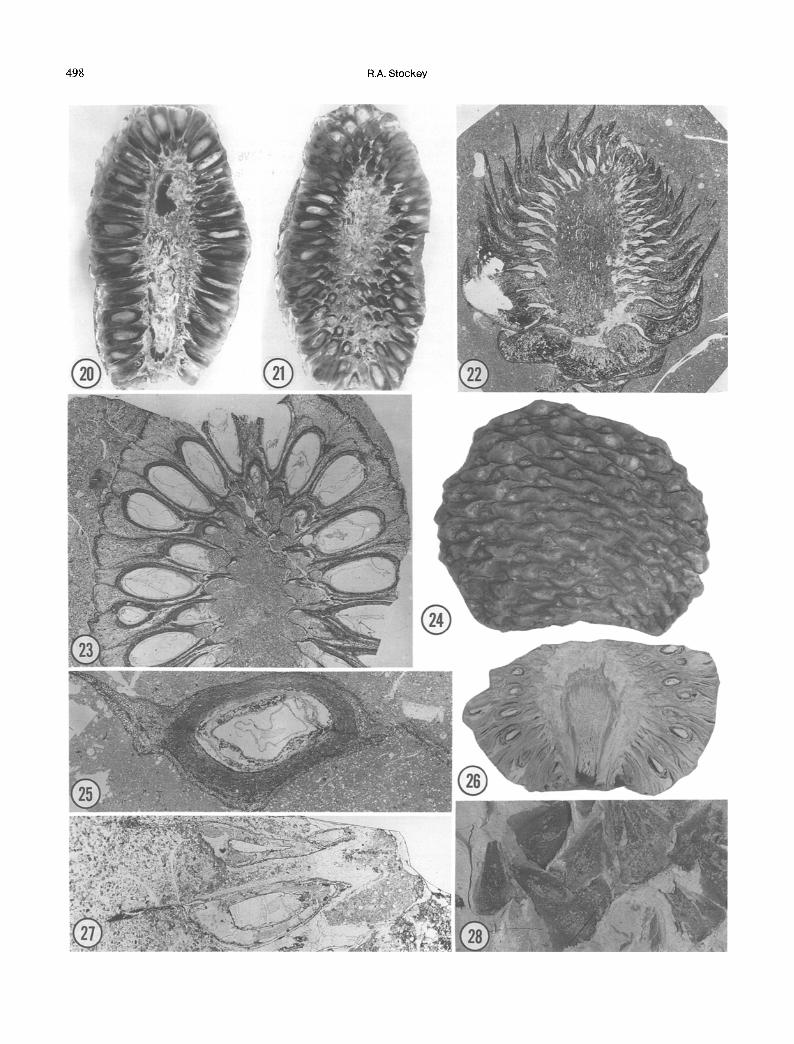
Recently, two cones have been described from the Upper Cretaceous of Hokkaido, Japan and Saghalien, Russia that combine features of several sections of the genus Araucaria. Araucaria nipponensis (Stockey et al. 1994) cones show similarities to cones of Section Araucar- ia (Figs. 23, 25). The very woody nature of the cone- scale complexes a n d a "ligular sulcus" (space between the bract and ovuliferous scale) that is small to absent in these cones are similar to those seen in A. angustifolia (Bertolini) O. Kuntze. Other cone characters such as the lack of a deciduous laminar tip on the bract and the dicotyledonous embryos are similar to Intermedia or Eutacta species (Stockey et al. 1994, Rouane and Woltz 1979, 1980, Woltz 1986).
Cones of Araucaria nihongfi (Stockey et al. 1992), like those of A. nipponensis, show characters of several sections of the genus Araucaria (Fig. 22). General cone structure with woody winged bracts and small, papery- thin ovuliferous scales is most similar to cones of Section Eutacta. The presence of a deciduous laminar tip on the bract has been reported in Section Araucaria, in A. angus- tifolia (Pilger 1926).
The embryo structure in A. nihongii is the best preser- ved in all the fossil cones so far described. Two cotyle- dons per embryo, as in this species, also have been reported in Section Intermedia (A. klinkii) Section Eutacta (A. heterophylla), Section Araucaria (A. araucana) and Section Bunya (A. bidwillfi). However, the germination of A. nihongii seeds was probably epigeal, unlike Sections Araucaria or Bunya. The vascularization of the cotyle- dons is similar to A. klinkfi Lauterbach as described by Rouane and Woltz (1980) or Agathis species. Embryos probably lacked a cotyledonary tube as in Section Bunya. The separation of the cotyledons and the anatomy of the cotyledonary node are similar to Eutacta species as described by Woltz (1986). The positions of resin canals in the hypocotyl of the embryo are similar to Intermedia species (Rouane and Woltz 1980).
The section Bunya, that contains only the extant A. bidwil l i i Hooker from Australia, appears to have been more widespread in the past (Stockey 1982). Included in this group are the cones of Araucaria mirabilis (Spegazzini) Windhausen (Figs. 24, 26) from the Jurassic Cerro Cua- drado petrified forest of Argentina (Wieland 1935, Calder 1953, Stockey 1975, t978). Like those of A. bidwill i i, these cones show a double vascular system to the cone- scale complex at its origin in the axis stele. Winged bracts and ovuliferous scale tips that are vascularized and free from the bract for about half their length are similar to the Australian species (Wilde and Eames 1948). There is a "vascular plexus" at the seed chalaza (Stockey 1975) as in A. bidwillf i (Wilde and Eames 1948). Embryos have two cotyledons.
Other fossil cones that have been assigned to the Section Bunya in the past include A. sphaerocarpa Car- ruthers (1866) from the Middle Jurassic Inferior Oolite Formation (Stockey 1980a) and A. brownfi Stockey (1980b) from the Jurassic of Dorset (Fig. 27) both from the U.K. Evidence now available from several sources indicates that some of the "Bunya" fossils should be reevaluated (Stockey et al. 1992). Araucaria brownfi, in particular, should probably be removed from the Section Bunya (Stockey et al. 1992). These cones show characters in common with cones such as those of A. nihongfi from the Cretaceous of Japan.
New cone material has been discovered in the Jurassic Morrison Formation of Utah (Figs. 20, 21). Cones definite- ly represent the genus Araucaria and are the first per- mineralized cone remains from North America. Cone shape and size are similar to A. mirabilis from the Cerro
Fig. 20. Permineralized Araucaria cone from the Jurassic Morrison Formation. X1.2. Fig. 21. Counterpart to cone in Fig. 20. Xl.2. Fig. 22. Immature Araucaria nihongii cone from the Upper Cretaceous Upper Yezo Group, Hokkaido, Japan. 91059 #24. Fig. 23. Araucaria nipponensis cone longitudinal section. 921803 A side #51. X2.5. Fig. 24. Araucaria mirabilis from the Cerro Cuadrado petrified forest of Patagonia. x2. Fig. 25. Araucaria nipponensis cone scale in cross section showing winged bracts. 840203 #33 X12. Fig. 26. Araucaria mirabilis from the Cerro Cuadrado petrified forest of Patagonia. X1.2. Fig. 27. Araucaria brownii longitudinal section of cone-scale complex. BMNH V59205 B #6. X5. Fig. 28. Araucarites sp. cone scales from the Jurassic Bex Hill Claypit, Walloon Coal Measures, N.S.W., Australia. X1.
X3.5.

500 R.A. Stockey
Cuadrado, however, ovuliferous scales are small and unvascularized. Typical araucarian seed integuments with a zig-zag pattern of branched sclereids are present. The wavy nucellar apices and mouth-shaped micropyles typical of the genus (Eames 1913) are also present. This new material currently under investigation by Stockey and Tidwell appears to show characters of several sections of the genus Araucaria.
In the same formation, what appear to be isolated araucarian cone axes have also been found (Fig. 19). Similar appearing fossil cone axes were illustrated by Wieland (1935) and Stockey (1978). The small circular scars on these axes are the former positions of cone- scale complexes (Figs. 15, 17). Since both Araucaria and Agathis shed their scales at maturity, investigation of the morphology and affinities of these fossil remains and associated foliage wi l l be the focus of future work. Seedlings: The description of carrot-l ike structures as fossil "seedlings" from the Cerro Cuadrado petrified forest (Stockey and Taylor 1978) associated with cones of Araucaria mirabilis has recently been called to question. Some fossil remains from Cerro Cuadrado are definitely seedlings (Stockey and Taylor 1978, figs. 15, 19). Others were thought likely to be seedlings (Stockey and Taylor 1978). While others were questionable seedlings. These latter two categories contain structures that vary from turbinate to top-shaped and up to 6 cm in length, and many appear very similar to seedlings of A. bidwillii and A. angustifolia with swollen hypocotyls and hypogeal germi- nation (Stockey 1982). Very similar structures, but with much better preservation of internal tissues, were also described by Stockey et al. (1992) from the Upper Cretaceous of Hokkaido, Japan. These structures, too, were associated with araucarian cone remains. How- ever, the internal anatomy of these corm-like structures showed that at least three types were present in Japanese deposits based on differences in the pith structure and the presence of resin canals in the primary and secondary xylem. Since xylem resin canals do not occur in the Araucariaceae, at least some of these structures were thought to have been produced by other conifers as well. Similar woody corm-like structures were observed in living seedlings of A. bidwillii with root apex damage (Stockey et al. 1990).
In a studies of the morphology of Araucaria bidwillfi seedlings, Burrows et al. (1992) and Burrows and Stockey (1994) showed that the internal tissue structure of typical bunya pine seedlings was very different from that seen in the fossils from the Cerro Cuadrado or the Upper Cretaceous of Hokkaido. Concentric rings of vascular tissue near the apical end of these seedlings, even in abnormally developed seedlings such as those illustrated by Stockey et al. (1990), were not present in the fossil "seedlings". This left considerable doubt as to the iden- tity of the corm-like structures in both Argentine and Japanese deposits.
Burrows et al. (1993) reviewed the literature on an- giosperm and gymnosperm taxa with hypogeal
(cryptogeal) germination. These taxa have cotyledons that remain inside the seeds and that transfer food reserves to swollen hypocotyls that are then used as storage organs. All of the extant taxa that show this type of germination also have large seeds with copious amounts of stored food reserves. Seeds from cones of Araucaria mirabilis from the Cerro Cuadrado (Stockey 1975) or A. nihongfi (Stockey et al. 1992) and A. nipponen- sis (Stockey et al. 1994) from the Upper Cretaceous of Hokkaido and Saghalien are not large enough to have the starch reserves to produce such large seedling-l ike struc- tures. Contrary to my earlier opinions, I now believe that it is probable that all of the fossil Araucaria species described so far had epigeal germination based on seed size (Burrows et al. 1992, Clarkson and Clifford 1987). This combined with an appreciation of the variability of araucarian embryo structure from the work of Rouane and Woltz (1979, 1980) and Woltz (1986), leads me to suggest that hypogeal germination was a relatively recent phe- nomenon in the Araucariaceae.
The other line of evidence suggesting that the corm- like structures are not seedlings comes from the Jurassic of North America. Tidwell and Medlyn (1992) described a startling set of specimens on which the "seedling"-Iike structures were found in attachment to shoot systems. These authors interpreted the structures as short shoots of extinct conifers. Two types are present in the Mor- rison Formation of Utah, Wyoming and Colorado. It is interesting to note that these short shoot systems occur .in the same formation with the araucarian cone remains.
In addition to the cones of Araucaria in the Morrison Formation, however, several other cone types are also found. Cones of a similar appearance to Pararaucaria patagonica Wieland, which co-occurs with Araucaria mirabilis at the Jurassic Cerro Cuadrado localities, are also present in some Morrison Formation localities, per- haps suggesting a possible affinity for the shoots. Parar- aucaria cones show many similarities to those of the Taxodiaceae and Pinaceae ; and Stockey (1977) suggest- ed that it probably should be in its own extinct family and not associated with the Araucariaceae in anyway. The shoot systems of Pararaucaria from Argentina are unknown at this time. In addition, Pararaucaria is not known to occur in the Upper Cretaceous of Hokkaido. The Mor- rison Formation material is currently under investigation.
Obviously, the story with respect to these shoot sys- tems is not a simple one. There may, in fact, be several types of remains represented. We have to take into account several conifer groups in the Hokkaido deposits as evidenced by the number and variety of seed cones, leaves and conifer wood types described so far (see Nishida 1991, for summary). This number is growing every year.
Conclusions
While we have learned a great deal about the fossil record of the Araucariaceae and morphology and anatomy

Mesozoic Araucariaceae 501
of living taxa in the last 12 years, we have still a long way to go in our understanding of the origins of araucarians and of the two extant genera. Many questions have been answered, but many more generated by our investi- gations. Most exciting to me is the recent find of per- mineralized cone material from North America. This fossil record has been virtually inaccessible because of preservation type and the kinds of "araucarian" plant organs that are present in many Mesozoic localities ; for example, the Triassic Petrified Forest from the Chinle Formation of Arizona, and its rafted Araucarioxylon logs. It is hoped that future concentration of effort into wel l- preserved araucarian remains from the Jurassic Morrison Formation of Utah, Wyoming and Colorado, in particular, will add significantly to the current database.
My earlier opinions of the fossil record of the Section Bunya of the genus Araucaria have been considerably modified, largely due to the character combinations dis- played in the excellently preserved Upper Cretaceous remains from Hokkaido and Saghalien. As more of these taxa are reconstructed as whole plants, it is hoped that a phylogenetic analysis of this group will be forthcoming. Current research on extant araucarians will continue add to the existing database.
I would like to thank the following people for assistance in specimen loans and the acquistion of extant material: Dr. Harufumi Nishida, International Budo University, Chiba, Japan; Prof. Makoto Nishida, Setagaya, Japan; Mr. Takeshi Ohsawa, Laboratory of Phylogenetic Botany, Chiba University, Japan; Dr. Kazuhiko Uemura, National Science Museum, Tokyo; Dr. W.D. Tidwell, Department of Botany, Brigham Young University; Drs. Gordon McPherson and Peter Raven, Missouri Botantical Garden ; Dr. P. Morat, Musee National d'Histoire Naturelle (Botani- que-Phanerogamie) Paris, Organisation Recherche Scientifique d'Outre Mer (ORSTOM), Noumea, New Caledonia; Eaux et Forets, Noumea, N.C. ; Dr. Philippe Woltz, Laboratoire de Morphogenese Vegetale, Universite d'Aix, Centre St. J6rbme, Marseille III, France; Dr. Garth Nikles, Mr. Robert Newton, Mr. Alan Harvey, Dr. Richard Gould, Dr. Russell Haines, Department of Forestry, Bris- bane and Gympie, QId. ; Dr. B.P.M. Hyland, Atherton, QId. ; Dr. H. J&hnichen, Palaeontologisches Museum der Humboldt-Universitat zu Berlin ; Field Museum of Natural History, Chicago; Dr. Britta Lundblad, Naturhistoriska Riksmuseet, Stockholm ; Gray Herbarium and the Arnold Arboretum, Harvard University ; Dr. John Silba, New York Botanical Garden. I thank Dr. G.W. Rothwell, Ohio Uni- versity for the use of laboratory facilities and George Braybrook, Helen Ko, lan Atkinson and Brenda Frevel, University of Alberta for laboratory assistance. Support- ed in part by NSERC grants A6908 and ICR145941 to RAS.
References
Alvin, K.L. and Boulter, M.C. 1974. A controlled method of comparative study of taxodiaceous cuticles. J.
Linn. Soc. (Bot.) 69 : 277-286. Barale, G. 1992. De nouveaux restes fossiles attribues
aux Araucariacees dan les calcaires lithographiques du Cr6tace inferieur du Montsec (province de Lerida, Espagne). Rev. Palaeobot. Palynol. 75: 53-64.
Bigwood, A.J. and Hill, R.S. 1985. Tertiary araucarian macrofossils from Tasmania. Aust. J. Bot. 33 : 645- 656.
Bose, M.N. 1975. Araucaria haastii Ettingshausen from Shag Point, New Zealand. Palaeobotanist 22: 76- 80.
Burrows, G.E., Boag, T.S., and Stockey, R.A. 1992. A morphological investigation of the unusual cryptogeal germination strategy of bunya pine (Araucaria bidwillii)- an Australian rain forest conifer. Int. J. Plant Sci. 153 : 503-512.
Burrows, G.E. and Stockey, R.A. 1994. The develop- mental anatomy of cryptogeal germination in bunya pine (Araucaria bidwillii). Int. J. Plant Sci. 155 : 519- 537.
Cookson, I.C. and Duigan, S.L. 1951. Tertiary Araucar- iaceae from southeastern Australia, with notes on living species. Aust. J. Sci. Res. Ser. B. 4 : 415-449.
Calder, M.G. 1953. A coniferous petrified forest in Patagonia. Bull. Br. Mus. (Nat. Hist.) Geol. 2: 197- 137.
Cantrill, D.J. 1992. Araucarian foliage from the Lower Cretaceous of southern Victoria, Australia. Int. J. Plant Sci. 153 : 622-645.
Carruthers, W. 1866. On araucarian cones from the Secondaw Beds of Britain. Geol. Mag. 3 : 249-252.
Clarkson, J.R. and Clifford, H.T. 1987. Germination of Jedda multicau/is J.R. Clarkson (Thymelaeaceae) : an example of cryptogeal germination in the Australian flora. Aust. J. Bot. 35: 715-720.
Eames, A.J. 1913. The morphology of Agathis austra/is. Ann. Bot. 27: 1-38.
Florin, R. 1931. Untersuchungen zur Stammesgeschichte der Coniferales und Cordaitales. K. Svensk. Vetens- kapakad. Handl. Ser. 5. 10 : 1-588.
Florin, R. 1963. The distribution of conifer and taxad genera in time and space. Acta Horti Bergiani 20: 121-312.
Greenwood, D.R. 1987. Early Tertiary Podocarpaceae: megafossils from the Eocene Anglesea Locality, Victoria, Australia. Aust. J. Bot. 35: 111-133.
Greguss, P. 1955. Identification of Living Gymnosperms on the Basis of Xylotomy. Akad. Kiado, Budapest.
Harris, T.M. 1979. The Yorkshire Jurassic Flora. V. Coniferales. Br. Mus. (Nat. Hist.) 803: 1-166.
Hill, R.S. 1990. Araucaria (Araucariaceae) species from Australian Tertiary sediments-a micromorphological study. Aust. Syst. Bot. 3: 203-220.
Hill, R.S. and Bigwood, A.J. 1987. Tertiary gymnosperms from Tasmania : Araucariaceae. Alcheringa 11 : 325-335.
Hill, R.S. and Pole, M.S. 1992. Leaf and shoot morphol- ogy of extant Afrocarpus, Nageia and Retrophy/lum (Podocarpaceae) species, and species with similar leaf arrangement from Tertiary sediments in Aus- tralasia. Aust. Syst. Bot. 5: 337-358.

502, R.A. Stockey
Hlustik, A. 1976. Contribuion to the systematic and leaf anatomy of the genus Dammarites Presl in Sternberg. Sbornik Narodniho Muzea V Praze. Rada B. 30: 49-70.
Hughes, N.F. 1969. Jurassic and Early Cretaceous pollen and spores, In R.H. Tschudy and R.A. Scott, eds., Aspects of Palynology. Wiley, New York. pp. 311- 329.
Jane, F.W. 1970. The Structure of Wood. 2nd ed. Adam and Charles Black, London.
Joy, K.V., Willis, A.J. "and Lacey, W.S. 1956. A rapid cellulose peel technique in Paleobotany. Ann. Bet. N.S. 20 : 635-637.
Kendall, M.W. 1947. Some conifers from the Jurassic of England. Ann. Mag. Nat. Hist. Ser. 12 : 583-594.
Kendall, M.W. 1949. A Jurassic member of the Araucar- iaceae. Ann. Bet. N.S. 13: 151-161.
Knobloch, E. 1973. Nomenklatorische und taxonomis- che Bermerkungen zur Gattung Dammarites Presl in Sternberg. Ber. Dtsch. Gesell. geol. Wiss. A. Geol. Palaont. Berlin. 1 : 967-973.
Menendez, C.A. and Caccavari, M.A. 1966. Estructura epidermica de Araucaria nathorsti Dus. det Terciaro de Pico Quemado, Rio Negro. Ameghiniana 4 : 195- 199.
Nishida, H. 1991. Diversity and significance of Late Cretaceous permineralized plant remains from Hok- kaido, Japan. Bot. Mag. Tokyo. 104 : 253-273.
Ohana, T. and Kimura, T. 1993. Permineralized Brachy- phyl/um leafy branches from the Upper Yezo Group (Coniacian- Santonian), Hokkaido, Japan. Bull. Nat. Sci. Mus. Ser. C. (Geoh and Paleontol.) 19 : 41-64.
Pant, D.D. and Srivastava, G.K. 1968. On the cuticular structure of Araucaria (Araucarites) cutchensis (Feist- mantel) comb. nov. from the Jabalpur Series, India. J. Linn. Soc. (Bot.) 61 : 201-206.
Pilger, R. 1926. Araucariaceae. In A. Engler and K. Prantl, eds., Die nat0rlichen Pflanzenfamilien. 13. Gymnospermae. Engelmann, Leipzig, pp. 249-266.
Rouane, M.L. and Woltz, P. 1979. Apport de I'~tude des plantules pour la taxonomie et I'evolution des Araucariacees. Bull. Soc. Bet. Fr. 126, Actual. Bet. 3 : 67-76.
Rouane, M.L. and Woltz, P. 1980. Interet des plantules pour la systematique et I'evolution des Araucar- iacees. Bull. Soc. Hist. Nat. Toulouse. 116 : 120-136.
Stockey, R.A. 1975. Seeds and embryos of Araucaria mirabilis. Amer. J. Bet. 62 : 856-868.
Stockey, R.A. 1977. Reproductive biology of the Cerro Cuadrado (Jurassic) con i fe rs : Pararaucaria patagonica. Amer. J. Bet. 64 : 733-744.
Stockey, R.A. 1978. Reproductive biology of Cerro Cua- drado fossil conifers: Ontogeny and reproductive strategies in Araucaria mirabilis (Spegazzini) Wind- hausen. Palaeontographica B. 166: 1-15.
Stockey, R.A. 1980a. Anatomy and morphology of Araucaria sphaerocarpa Carruthers from the Jurassic Inferior Oolite of Bruton, Somerset. Bet. Gaz. 141: 116-124.
Stockey, R.A. 1980b. Jurassic araucarian cone from southern England. Palaeontology 23 : 657-666.
Stockey, R.A. 982. The Araucariaceae : an evolutionary perspective. Rev. Palaeobot. Palynol. 37: 133-154.
Stoekey, R.A. 1990. Antarctic and Gondwana conifers. In T.N. Taylor and E.L. Taylor, eds., Antarctic Paleobiology: Its Role in the Recontruction of Gon- dwana. Springer-Verlag, New York, pp. 179-191.
Stockey, R.A. and Atkinson, I.J. 1993. Cuticle micromor- phology of Agathis Salisbury. Int. J. Plant Sci. 154 : 187-225.
Stockey, R.A. and Ko, H. 1986. Cuticle micromorphology of Araucaria de Jussieu. Bet. Gaz. 147 : 508-548.
Stockey, R.A. and Taylor, T.N. 1978. On the structure and evolutionary relationships of the Cerro Cuadrado fossil conifer seedlings. J. Linn. Soc. London (Bet.) 76 : 161-176.
Stockey, R.A., Nishida, M. and Nishida, H. 1990. Struc- ture and diversity of the woody conifer seedling-like structures from the Upper Cretaceous of Hokkaido, Japan. Bet. Gaz. 151: 252-262.
Stockey, R.A., Nishida, H. and Nishida, M. 1992. Upper Cretaceous araucarian cones from Hokkaido : Araucaria nihongii sp. nov. Rev. Palaeobot. Palynol. 72 : 27-40.
Stockey, R.A., Nishida, M. and Nishida, H. 1994. Upper Cretaceous araucarian cones from Hokkaido and Saghalien:Araucaria nipponensis sp. nov. Int. J. Plant Sci. 155 : 800-809.
Taylor, T.N. and Taylor, E.L. 1993. The Biology and Evolution of Fossil Plants. Prentice Hall, Englewood Cliffs, New Jersey..
Tidwell, W.D. and Medlyn, D.A. 1992. Short shoots from the Upper Jurassic Morrison Formation, Utah, Wyom- ing and Colorado, USA. Rev. Palaeobot. Palynol. 71 : 219-238.
White, C.T. 1947. Notes on two species of Araucaria in New Guinea and a proposed new section of the genus. J. Arnold Arbor. 28: 259-260.
White, M.E. 1981. Revision of the Talbragar Fish Bed Flora (Jurassic) of New South Wales. Rec. Aust. Mus. 33 : 695-721.
Wieland, G.W. 1935. The Cerro Cuadrado Petrified For- est. Carnegie Inst. Wash. Publ. 449: 1-183.
Wilde, M.H. and Earnes, A.J. 1948. The ovule and 'seed' of Araucaria bidwil/i with discussion of the taxonomy of the genus. I. Morphology. Ann. Bot. N.S. 12 : 311- 326.
Wilde, M.H. and Eames, A.J. 1952. The ovule and 'seed' of Araucaria bidwi//i with discussion of the taxonomy of the genus. II. Taxonomy. Ann. Bot. N.S. 16 : 27- 47.
Woltz, P. 1986. Les Podocarpus (s.l.) origines et evolu- tion. I~tude des plantules; comparaison avec les families de confees de I'hemisphere sud. Thesis. Univ. Aix-Marseil le III (St-Jerome).
(Received February 1, 1994 : Accepted October 18, 1994)