mating in the monogamous male: behavioral consequences
TRANSCRIPT

Pergamon
0031-9384(94)00362-9
Physiology & Behavior, Vol. 57, No. 4, pp. 615-627, 1995 Copyright © 1995 Elsevier Science Ltd Printed in the USA. All rights reserved
0031-9384/95 $9.50 + .00
Mating in the Monogamous Male: Behavioral Consequences
T H O M A S R. I N S E L , l S T E P H A N I E P R E S T O N A N D J A M E S T. W I N S L O W
Laboratory o f Neurophysiology, National Institute of Mental Health, Poolesville, MD 20837
R e c e i v e d 11 Apr i l 1994
INSEL, T. R., S. PRESTON AND J. T. WINSLOW. Mating in the monogamous male: Behavioral consequences. PHYSIOL BEHAV 57(4) 615-627, 1995.--In monogamous mammals, males typically show selective affiliation with a single mate, high levels of paternal care, and aggression towards conspecifics to protect mate and offspring. We have previously described how selective aggression and affiliation increase after mating in the male prairie vole, Microtus ochrogaster. The current studies further explored the behavioral changes that follow mating in the male of this species. The first set of experiments tested males on several behavioral measures after 24 h of either mating, social (but not sexual) exposure, or no social contact. After 24 h of mating, but not after the other two conditions, aggression and affiliation (partner preference) increased as previously reported. In addition, mated animals showed increased exploration of the open arms of a plus maze, consistent with decreased fearfulness. There were no group differences in paternal behavior (which was high in all three conditions) or analgesia (assessed by tail flick latency). To determine the minimum amount of mating necessary for the induction of aggression, males were tested in a resident-intruder paradigm after 1,6, or 24 h of mating. Although 1 h of mating was associated with a transient increase in the frequency of threats and attacks, the full spectrum of enduring aggression was observed only in the males given 24 h of mating. In a final experiment, the behavioral consequences of mating were studied in males of the closely related montane vole (Microtus montanus) which does not pair bond. Males of this nonmonogamous vole species did not show increased aggression, partner preference, or alterations in plus maze exploration following 24 h of mating. These results demonstrate the importance of prolonged mating for the induction of pair bonding in the monogamous male and they suggest that increases in aggression and affiliation are associated with decreased fearfulness in pair bonded males.
Vole Pair bonding Plus maze Preference Aggression Reproductive behavior
M O N O G A M O U S social organization is usually characterized by selective affiliation with a partner, high levels of paternal behav- ior, and intense aggression towards strangers for defense of ter- ritory, nest, and mate (6,16). For many monogamous mammals, these behaviors are not present throughout development, but ap- pear rapidly after mating. The prairie vole, Microtus ochrogaster, a rodent which appears monogamous in both field and laboratory studies, has proven especially instructive in this regard (4,11). Prairie voles are gregarious and highly affiliative as long as they remain sexually quiescent in their natal group. Once a female vole is exposed to pheromones from an unrelated male, she be- comes sexually receptive within 24 h and, as a consequence of mating, she develops a marked preference for the partner. This partner preference appears to be enduring (although the female may be induced to mate with a novel male) and thus, may rep- resent the first phase of pair bonding (3,12).
We have previously described how male prairie voles embark on an analogous transition after mating (31). Within 24 h of mat- ing, male prairie voles change from showing diffuse social con- tact to highly selective affiliation with increased aggression to- wards conspecifics other than the mate. These changes are of
interest because the emergence of selective affiliation (partner preference) and selective aggression (mate guarding) are instru- mental to pair bonding in the male prairie vole. Pair bonding is a theoretical construct that involves more than selective aggres- sion and affiliation, but these behaviors may be necessary (if not sufficient) for the development of a monogamous bond.
Although this previous report demonstrated that mating-in- duced aggression was selective and enduring [males continued to be aggressive even after social isolation for 1 wk (31)], several issues remained unresolved. In this study, nonmating males used as controls were males who failed to mate after 24 h of exposure to an estrous female. The lack of aggression in these animals may have resulted from some general difference in this control group (e.g., over-representation of subordinate males) rather than the specific absence of mating. In addition, this study did not define what amount of mating was necessary to observe the increase in aggression, nor did it address other behaviors that may have changed as a consequence of mating. The current studies were designed to resolve these issues. To determine the importance of mating, as opposed to nonspecific factors in the control males, we designed an experiment with multiple control groups. To in-
Requests for reprints should be addressed to Thomas R. Insel, M.D., Office of the Director, Yerkes Regional Primate Research Center, Emory University, Atlanta, GA 30322.
615

616 INSEL, PRESTON AND WINSLOW
vestigate the specificity of the changes in aggression and affili- ation, we examined several behavioral dependent variables in- cluding paternal care, fearfulness, and pain sensitivity. An increase in paternal care, would support the notion of changes postmating subserving pair bonding. Fearfulness and pain sen- sitivity were studied because of reports in other species linking benzodiazepine and opiate systems to social interaction (10,15). To address the time course of the mating effects, we examined the induction of aggression in males with various amounts of mating. Finally, to determine if these behavioral changes subse- quent to mating were characteristic of monogamous social or- ganization or more generally related to microtine rodents, we investigated the consequences of mating in the nonmonogamous montane vole male.
EXPERIMENT 1: BEHAVIORAL CONSEQUENCES OF MATING VS. SOCIAL CONTACT
METHODS
Animals
Prairie voles, originally derived from feral stock captured near Urbana, Illinois, were laboratory-reared at the NIH Animal Cen- ter in Poolesville, MD. Breeding voles were housed in polycar- bonate cages (55 x 31 x 21 cm) on cedar chip bedding with approx. 10 cm of alfalfa cover for burrowing and nest building. Offspring were weaned away from both parents at 22 days of age and housed with same sex littermates until 60-90 days of age when they were used for behavioral experiments. Housing was on a 12:12 light:dark schedule (lights on at 7AM) with food (Purina rabbit chow) and water available ad lib; substrate was cedar chip bedding without alfalfa covering.
Experimental Design
This experiment employed a triadic design based on trios of sexually naive male siblings housed together since weaning at age 22 days. Between 60 and 90 days of age, each sibling was housed separately for 3 days (to minimize potential effects of social status in communal cages) and then assigned randomly to one of three experimental groups: MATED, EXPOSED, and CONTROL. Each male in the MATED group was given a sex- ually receptive female for 24 h. Sexually receptive females had been ovariectomzed at least 2 wk earlier, injected with estradiol benzoate (5.0 ug, SC) for 3 days, and tested with a sexually active male to ensure lordosis prior to use as the stimulus female in the mating condition. Males in the EXPOSED group were given so- cial exposure to a sexually nonreceptive female for 24 h. These stimulus females were also ovariectomized, but not given estra- diol benzoate. The CONTROL group was left undisturbed through the same 24 h period. Eleven trios were studied in this fashion. Males were filmed throughout the 24 h experimental intervention, using time-lapse video recording with a 12:1 re- duction (12 min of real time for each 1 minute of tape) and a low-light adjustment to accomodate filming throughout the dark cycle. This video record provided verification of mating or non- mating in the different experimental conditions and also allowed the detection of group differences in nonmating behaviors, such as side-by-side contact, which might confound the interpretation of our results.
Our original triadic design was complicated by the discovery that 6 of the males in the MATED group were not observed to mate on the time-lapse video tapes. This apparent absence of mating could not be attributed to the E.B.-primed females, as these females were shown to be receptive prior to pairing with the males. Moreover, given the number of bouts of mating usu-
ally seen, it was unlikely that mating occurred but we failed to detect it (although males that mated only once or twice during the dark phase may have been overlooked). These nonmating males were henceforth treated as a fourth experimental group (NON-MATED, n = 6). Three additional males were added to the MATED group (final n = 8) to compensate for the creation of the NON-MATED category. These additional males were tested simultaneously with males from other groups to avoid bi- asing the results.
The behavioral measures were exploration in the plus maze, paternal behavior, aggression in a resident-intruder paradigm, and tail-flick as a measure of analgesia. These behaviors were studied sequentially (midway through the daily light phase) both prior to and immediately following the 24 h experimental con- dition. A fifth measure, partner preference, was found to be in- compatible with the other four behavioral measures in pilot stud- ies because of its length (3 h) and its sensitivity to context (i.e., the four preceding tests appeared to affect the partner preference). Therefore, partner preference was assessed in an entirely differ- ent group of males following either the MATING (n = 9) or the EXPOSED (n = 9) condition.
Elevated Plus Maze Exploration
The elevated plus maze has been widely used to assess fear- fulness or anxiety in both rats and mice (17,22). This test exploits the natural aversion rodents demonstrate to heights and open spaces. Animals spend more time in the exposed or "open" arms of the maze following administration of a clinically anxiolytic compound and less time in the open arms of the maze following anxiogenic drugs or fear-inducing experiences (23). Recently this test has been used to detect strain differences in fearfulness (25).
The maze consisted of two opposing open arms and two op- posing closed arms, each 67 cm long and 5.5 cm wide. The open arms had a 1 cm lip to prevent the vole from falling; the closed arms had a 15 cm dark Plexiglas wall. The maze was 40 cm above the table surface. Pairs of photobeam detectors were placed 4cm down each ann to record entries and exits. Following a 30 min adaptation to the test room, each vole was placed in the neutral square at the cross of the maze, pointing towards an open arm. The following measures were automatically recorded during a 5 min test: number of entries into and duration within open arms, number of entries into and duration within closed arms, time in neutral area. Testing was done under red light conditions (1 x 60W, 3 feet above the center of the maze). The maze was cleaned with a dilute soap solution between each test.
Paternal Behavior
Paternal behavior was assessed in the male's home cage by adding a conspecific 1-5 day old pup. The pup was placed on the opposite side of the cage and the male's behavior was vid- eotaped for the subsequent 10 min. The principal measures were latency to make contact and duration of contact with the pup. If the pup was attacked, the test was stopped immediately. Each pup was used only once.
Analgesia
Analgesia was assessed with a simple tail-flick assay, a method that was originally used to detect changes in pain sen- sitivity (7) and has been used more recently to demonstrate an- algesia following social stress (19) or gonadal steroid treatment (18). The vole was held around the chest and gently lowered until his tail was suspended in water at 56°C. A timer was started with a foot pedal at the time of tail submersion and stopped at the

MATING IN THE MONOGAMOUS MALE 617
moment of tail withdrawal. If no tail movement was detected after 10 s, the test was terminated and a score of 10 was recorded. Each test consisted of three trials, with the median latency used for data analysis.
Resident-Intruder Paradigm
The resident-intruder paradigm has been used previously to study aggression (20). An intruder male was placed in the home cage of the experimental animal for 6 min. Each male was tested at baseline and then tested with a different intruder for subsequent tests. For groups with female exposure, the female was removed prior to adding the intruder male. Intruders were conspecific males housed with 4 -5 other males and previously defeated in a social encounter. Behaviors were recorded on videotape and subsequently scored using a computer-assisted data acquisition system by a rater blind to group assignment and trained to a criterion of greater than 95% reliability. Aggressive behaviors were operationally defined as previously described (24) with re- ported means indicating frequency over the 6 min test. Offensive aggression consisted of attack-bites, lunge threats, and lateral threats. Defensive aggression consisted of the defensive upright posture and escape leaps. In addition, several mutually exclusive activity measures, including rearing (front paws off the floor of the cage), walking (ambulation), sniffing (snout raised), auto- grooming (self-licking or scratching), and digging (scratching bedding with front paws) were assessed for duration.
Partner Preference
The method of assessing partner preference in prairie voles has been previously described (30). Briefly, males received either 24 h of exposure to a sexually receptive female (MATED, n = 9) or a sexually nonreceptive female (EXPOSED, n = 9). Mating was verified by time-lapse video (at least 2 episodes of mating were required in the first 6 h). At the end of 24 h, males were placed into a 3-chamber apparatus in which the test animal chooses between a neutral cage, a cage with a familiar female ("partner"), or a cage with a novel female ("stranger"). The females were tethered to restrain them in their cages, but the male had free access to all three chambers. The activity of the male was recorded for 3 h (using time-lapse recording) with the fol- lowing measures scored from the tapes: entries into and duration within each chamber, time in side-by-side contact with partner or stranger female, and frequency of mating. Previous studies have demonstrated that a 3 h preference test yields results com- parable to tests of 6 or 24 h duration (30).
A critical aspect of this test is the successful matching of stimulus females. Females were matched prior to being placed with a male-- that is, for mated males, both the partner and the stranger were sexually receptive prior to the 24 h of mating and, for nonmated males, neither female had received steroid priming. For both con- ditions, it is possible that the 24 h of mating or cohabitation alters the stimulus properties of the female and thus confound the study of male preference. To role out this possibility, each female was used for two partner preference tests, such that the partner female in one test would be the stranger female in the other.
Data Analysis
Except for the partner preference test, each of the behavioral measures was assessed twice in each individual. Differences were evaluated with a repeated measures ANOVA, with group (MATED vs. EXPOSED vs. CONTROL vs. NON-MATED) as a main factor and time (PRE vs. POST) as the repeated measure. In the presence of a significant main effect or interaction (ie. p
< .05), group differences were further assessed in the "pre" and "post" conditions using a single factor ANOVA with a posthoc Sheff6 test (again, p < .05 adopted as signficant).
For the partner preference test, a preference was defined as a significant difference (p < .05) between time in contact with the partner and time in contact with the stranger by a paired t-test. Between group differences were further assessed by comparing time in side-by-side contact and time in each of the three cages by Student's t-test. Variance is represented by SEM throughout.
RESULTS
Mating
During the 24 h of cohabitation with the female, few differ- ences were apparent between the MATED and the EXPOSED groups. Males that mated always mounted females within 3 h (mean latency = 38.4 _ 16.8 min.) and continued to mount every 40-60 min throughout the 24 h (mean number of mating bouts in the first 6 h = 8.1 _+ 0.7). There was however, no group difference in time spent in side-by-side contact or grooming. Pairs that mated spent 258.5 _ 9.8 rain in contact whereas pairs in the EXPOSED group spent 253.4 _ 16.7 min in contact during the first 6 h (maximum possible = 360 min.). During the same interval, MATED males spent 68.6 _+ 6.9 min. while EXPOSED males spent 59.6 __+ 6.8 min. grooming.
Resident-Intruder Paradigm
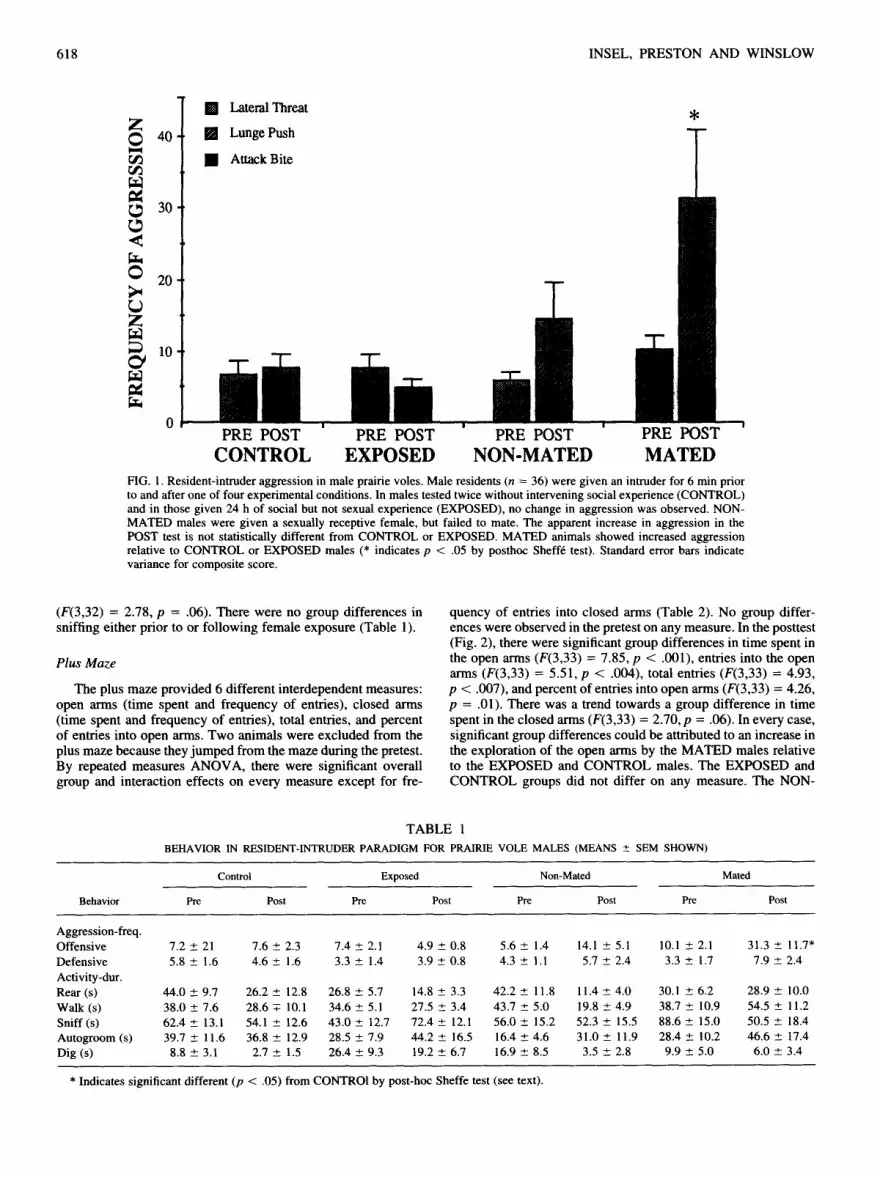
Males (n = 36) in the resident-intruder paradigm were as- sessed on several measures of activity and aggression. There were no group differences in any measure of aggression prior to female exposure. Across all groups, offensive aggression was relatively low and manifested mostly as lateral threats (Fig. l).Attack bites were particularly rare, occurring only once across all pretests. As predicted, the repeated measures analysis revealed a significant group effect (F(3,32) --- 4.912, p = .006) and a significant inter- action term (F(3,3) = 3.60, p = .024). These effects could be largely attributed to changes in the second test (F(3,35) = 4.947, p < .006) when the MATED group showed higher levels of ag- gression than either the EXPOSED or CONTROL males (Fig. 1). There were no other group differences detected by Sheff6 test. The problematic NON-MATED group was not significantly different from either the MATED or the CONTROL males.
As can be seen in Fig. 1, not only the quantity but the quality of the aggression changed after mating. For instance, attack-bites were more frequent in the second test, although almost exclu- sively in the MATED males. NON-MATED males, intermediate between the other groups, showed little change in lateral threats or attack-bites, but a modest increase in the frequency of lunge pushes. It should also be noted that among the MATED males, the expression of aggression was highly variable (Table 1).
There were neither group nor time effects for defensive ag- gression (composite measure of defensive upright posture and escape responses). The frequency of these behaviors was gen- erally low across all conditions (Table 1), with no evidence for a change in the pattern of the components of defensive responses following mating.
Activity measures also showed few experimental differences. There were no overall group differences in duration of rearing, walking, sniffing, grooming, or digging. Significant time effects were apparent for rearing and digging, both of which decreased overall in the second test. Interactions were notable for walking (F(3,3) = 4.50, p < .010) and sniffing (F(3,3) = 3.41, p < .029). In the posttest, MATED males showed a trend towards increased duration of walking relative to males in any of the other groups
618 INSEL, P R E S T O N A N D W l N S L O W
Z 40
30 r~
20
Z
10
• Lateral Threat
• Lunge Push 7"-
| • Attack Bite
| !
PRE POST PRE POST PRE POST PRE POST
CONTROL EXPOSED NON-MATED MATED FIG. 1. Resident-intruder aggression in male prairie voles. Male residents (n = 36) were given an intruder for 6 min prior to and after one of four experimental conditions. In males tested twice without intervening social experience (CONTROL) and in those given 24 h of social but not sexual experience (EXPOSED), no change in aggression was observed. NON- MATED males were given a sexually receptive female, but failed to mate. The apparent increase in aggression in the POST test is not statistically different from CONTROL or EXPOSED. MATED animals showed increased aggression relative to CONTROL or EXPOSED males (* indicates p < .05 by posthoc Sheff6 test). Standard error bars indicate variance for composite score.
(F(3,32) = 2.78, p = .06). There were no group d i f fe rences in sniff ing ei ther pr ior to or fo l lowing female exposure (Table 1).
Plus Maze
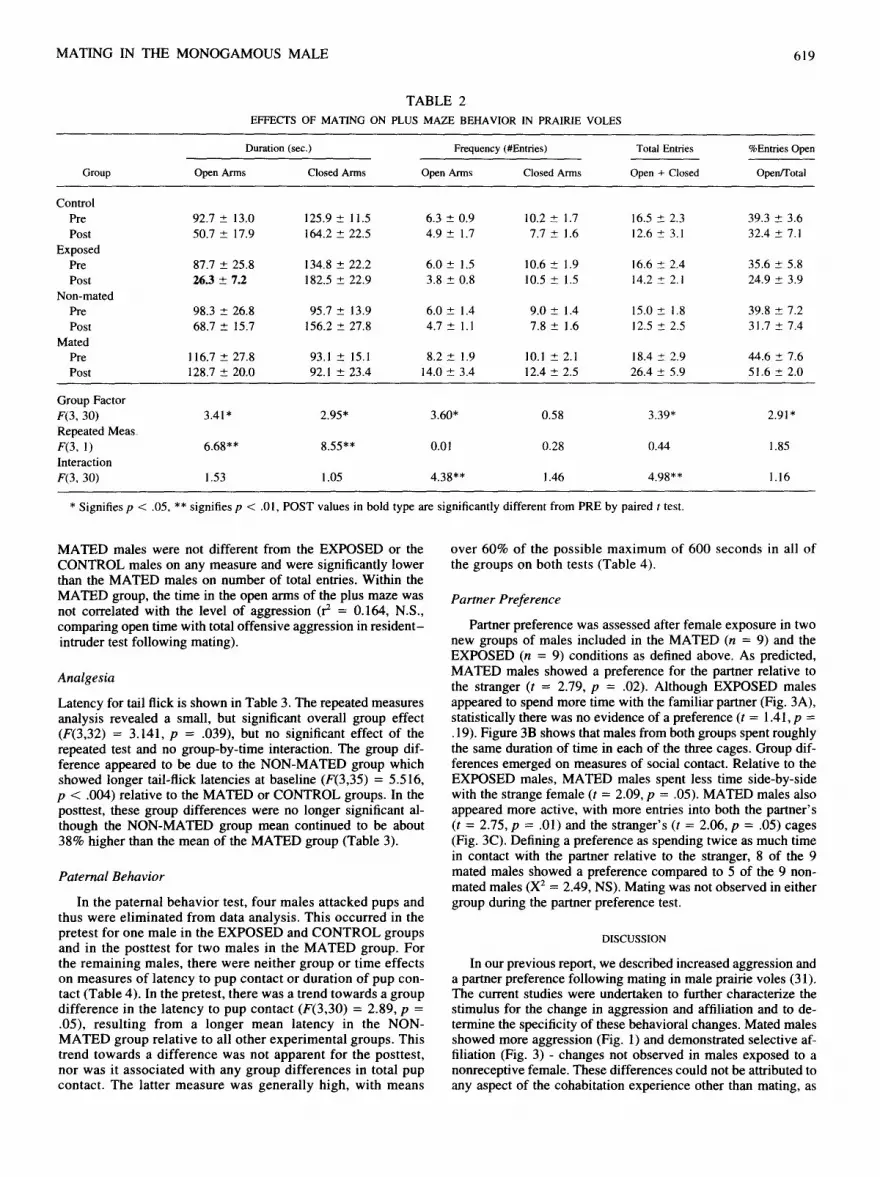
The plus maze p rov ided 6 d i f ferent in te rdependen t measures : open arms ( t ime spent and f r equency o f entries) , c losed arms ( t ime spent and f requency o f entries), total entr ies, and percent o f entr ies into open arms. T w o animals were exc luded f rom the plus maze because they j u m p e d f rom the m a z e during the pretest . By repeated measures A N O V A , there were significant overal l group and in teract ion effects on every measure excep t for fre-
quency o f entr ies into c losed arms (Table 2). N o group differ- ences were obse rved in the pretes t on any measure . In the post tes t (Fig. 2), there were significant g roup d i f ferences in t ime spent in the open arms (F(3,33) = 7.85, p < .001), entr ies into the open arms (F(3,33) = 5.51, p < .004), total entr ies (F(3,33) = 4.93, p < .007), and percent o f entr ies into open arms (F(3,33) = 4.26, p = .01). There was a t rend towards a group d i f ference in t ime spent in the c losed arms (F(3,33) = 2.70, p = .06). In every case, s ignificant group d i f ferences could be at tr ibuted to an increase in the explora t ion o f the open arms by the M A T E D males relative to the E X P O S E D and C O N T R O L males . The E X P O S E D and C O N T R O L groups d id not d i f fer on any measure . The N O N -
T A B L E 1
BEHAVIOR IN RESIDENT-INTRUDER PARADIGM FOR PRAIRIE VOLE MALES (MEANS +__ SEM SHOWN)
Control Exposed Non-Mated Mated
Behavior Pre Post Pre Post Pre Post Pre Post
Aggression-freq. Offensive 7.2 +__ 21 Defensive 5.8 ± 1.6 Activity-dur. Rear (s) 44.0 ± 9.7 Walk (s) 38.0 ± 7.6 Sniff (s) 62.4 ± 13.1 Autogroom (s) 39.7 ___ l 1.6 Dig (s) 8.8 ± 3.1
7.6 ± 2.3 7.4 ± 2.1 4.9 ± 0.8 5.6 ± 1.4 14.1 ± 5.1 10.1 ± 2.1 31.3 +_ 11.7" 4.6 ± 1.6 3.3 ± 1.4 3.9 +_ 0.8 4.3 ± 1.1 5.7 +_ 2.4 3.3 ± 1.7 7.9 ± 2.4
26.2 ± 12.8 26.8 ___ 5.7 14.8 + 3.3 42.2 ± 11.8 11.4 ± 4.0 30.1 ± 6.2 28.9 ± 10.0 28.6 ¥ 10.1 34.6 ± 5.1 27.5 ± 3.4 43.7 ± 5.0 19.8 ___ 4.9 38.7 +_ 10.9 54.5 +_ 11.2 54.1 ± 12.6 43.0 ± 12.7 72.4 ± 12.1 56.0 ___ 15.2 52.3 ± 15.5 88.6 ___ 15.0 50.5 ± 18.4 36.8 ± 12.9 28.5 ± 7.9 44.2 ± 16.5 16.4 ± 4.6 31.0 ± 11.9 28.4 ± 10.2 46.6 ± 17.4
2.7 ± 1.5 26.4 + 9.3 19.2 ± 6.7 16.9 ± 8.5 3.5 ___ 2.8 9.9 ___ 5.0 6.0 ± 3.4
* Indicates significant different (p < .05) from CONTROl by post-hoc Sheffe test (see text).
M A T I N G IN THE M O N O G A M O U S MALE 619
TABLE 2
EFFECTS OF MATING ON PLUS MAZE BEHAVIOR IN PRAIRIE VOLES
Duration (sec.) Frequency (#Entries) Total Entries %Entries Open
Group Open Arms Closed Arms Open Arms Closed Arms Open + Closed Open/Total
Control Pre 92.7 ± 13.0 125.9 ± 11.5 6.3 ± 0.9 10.2 ± 1.7 16.5 ± 2.3 39.3 ± 3.6 Post 50.7 ± 17.9 164.2 ± 22.5 4.9 ± 1.7 7.7 ± 1.6 12.6 ± 3.1 32.4 + 7.1
Exposed Pre 87.7 ± 25.8 134.8 ± 22.2 6.0 ± 1.5 10.6 ± 1.9 16.6 ± 2.4 35.6 _ 5.8 Post 26.3 ± 7.2 182.5 ± 22.9 3.8 ± 0.8 10.5 ± 1.5 14.2 ± 2.1 24.9 ± 3.9
Non-mated Pre 98.3 ± 26.8 95.7 ± 13.9 6.0 ± 1.4 9.0 ± 1.4 15.0 ± 1.8 39.8 ± 7.2 Post 68.7 ___ 15.7 156.2 ± 27.8 4.7 ± 1.1 7.8 + 1.6 12.5 ± 2.5 31.7 _+ 7.4
Mated Pre 116.7 ± 27.8 93.1 ± 15.1 8.2 _+ 1.9 10.1 ± 2.1 18.4 _+ 2.9 44.6 ± 7.6 Post 128.7 ± 20.0 92.1 ± 23.4 14.0 ± 3.4 12.4 ± 2.5 26.4 ± 5.9 51.6 z 2.0
Group Factor F(3, 30) 3.41 * 2.95* 3.60* 0.58 3.39* 2.91 * Repeated Meas F(3, 1) 6.68** 8.55** 0.01 0.28 0.44 1.85 Interaction F(3, 30) 1,53 1.05 4.38** 1.46 4.98** 1.16
* Signifies p < .05, ** signifies p < .01, POST values in bold type are significantly different from PRE by paired t test.
M A T E D males were not different from the EXPOSED or the CONTROL males on any measure and were significantly lower than the M A T E D males on number of total entries. Within the M A T E D group, the t ime in the open arms of the plus maze was not correlated with the level of aggression (r z = 0.164, N.S., comparing open time with total offensive aggression in r e s iden t - intruder test following mating).
Analgesia
Latency for tail flick is shown in Table 3. The repeated measures analysis revealed a small, but significant overall group effect (F(3,32) = 3.141, p = .039), but no significant effect of the repeated test and no group-by-t ime interaction. The group dif- ference appeared to be due to the N O N - M A T E D group which showed longer tail-flick latencies at baseline (F(3,35) = 5.516, p < .004) relative to the M A T E D or CONTROL groups. In the posttest, these group differences were no longer significant al- though the N O N - M A T E D group mean continued to be about 38% higher than the mean of the MATED group (Table 3).
Paternal Behavior
In the pa terna l behav io r test, four males at tacked pups and thus were e l imina ted f rom data analysis . This occurred in the pretest for one male in the E X P O S E D and C O N T R O L groups and in the post tes t for two males in the M A T E D group. For the r emain ing males, there were ne i ther group or t ime effects on measures of la tency to pup contac t or durat ion of pup con- tact (Table 4). In the pretest , there was a t rend towards a group di f ference in the la tency to pup contac t (F(3,30) = 2.89, p = .05), resul t ing f rom a longer mean la tency in the NON- M A T E D group re la t ive to all o ther exper imenta l groups. This t rend towards a d i f fe rence was not apparen t for the posttest , nor was it associa ted wi th any group d i f ferences in total pup contact . The lat ter measure was genera l ly high, with means
over 60% of the possible m a x i m u m of 600 seconds in all of the groups on both tests (Table 4).
Partner Preference
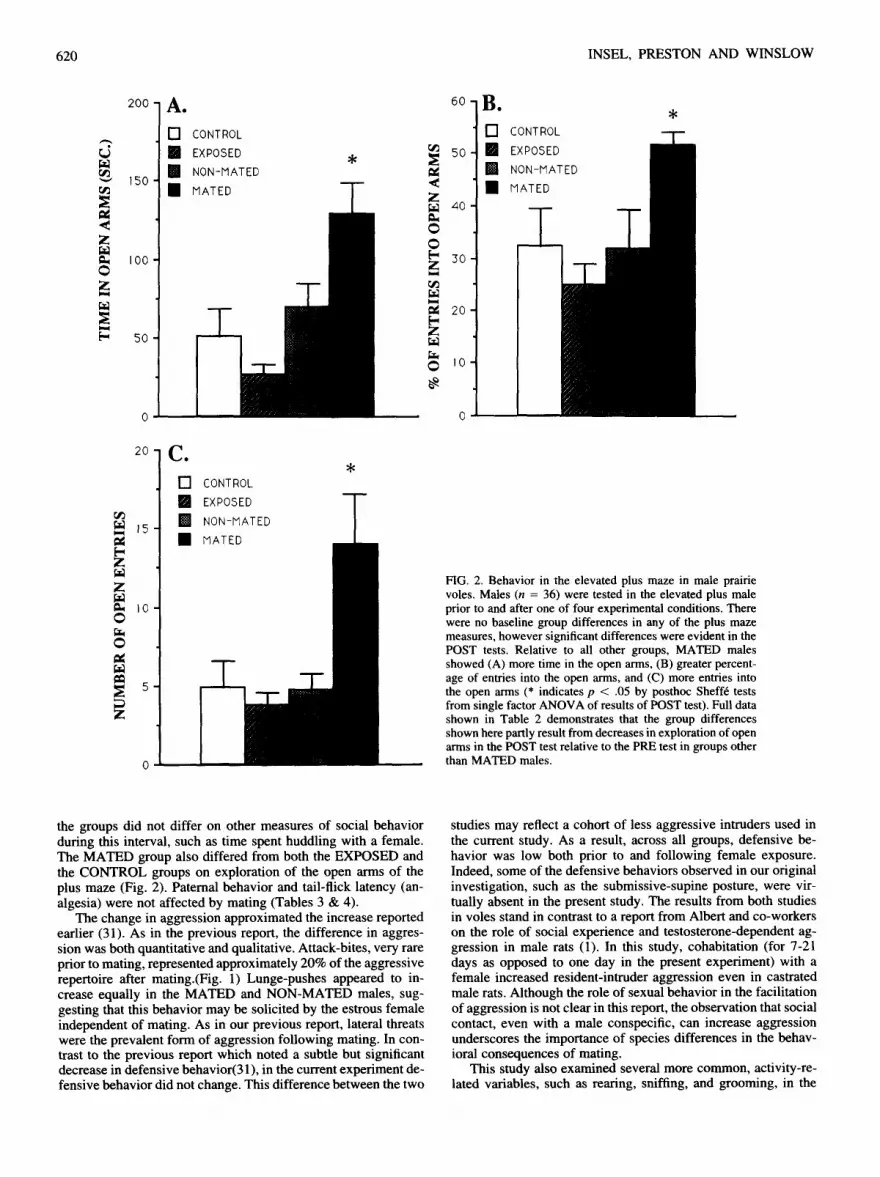
Partner preference was assessed after female exposure in two new groups of males included in the M A T E D (n = 9) and the EXPOSED (n = 9) conditions as defined above. As predicted, MATED males showed a preference for the partner relative to the stranger (t = 2.79, p = .02). Although EXPOSED males appeared to spend more time with the familiar partner (Fig. 3A), statistically there was no evidence of a preference (t = 1.41, p = • 19). Figure 3B shows that males from both groups spent roughly the same duration of t ime in each of the three cages. Group dif- ferences emerged on measures of social contact. Relative to the EXPOSED males, MATED males spent less time side-by-side with the strange female (t = 2.09, p = .05). MATED males also appeared more active, with more entries into both the partner 's (t = 2.75, p = .01) and the s tranger 's (t = 2.06, p = .05) cages (Fig. 3C). Defining a preference as spending twice as much time in contact with the partner relative to the stranger, 8 of the 9 mated males showed a preference compared to 5 of the 9 non- mated males (X 2 = 2.49, NS). Mating was not observed in either group during the partner preference test.
DISCUSSION
In our previous report, we described increased aggression and a partner preference following mating in male prairie voles (31). The current studies were undertaken to further characterize the stimulus for the change in aggression and affiliation and to de- termine the specificity of these behavioral changes. Mated males showed more aggression (Fig. 1) and demonstrated selective af- filiation (Fig. 3) - changes not observed in males exposed to a nonreceptive female• These differences could not be attributed to any aspect of the cohabitat ion experience other than mating, as
620 INSEL, PRESTON AND WINSLOW
,(
Z
IO0 0 Z W,,W
W,w
[ - 5O
200 _.~.
[]
150
20
{n [#.l 15 ~ i (
Z
Z
0
0
CONTROL
EXPOSED
NON-MATED
Ce [ ] CONTROL • EXPOSED • NON-MATED
• MATED
Z
0 0 [ - z
60
50
~.0
30
20
I0
Be
m E R . R ~
FIG. 2. Behavior in the elevated plus maze in male prairie voles. Males (n = 36) were tested in the elevated plus male prior to and after one of four experimental conditions. There were no baseline group differences in any of the plus maze measures, however significant differences were evident in the POST tests. Relative to all other groups, MATED males showed (A) more time in the open arms, (B) greater percent- age of entries into the open arms, and (C) more entries into the open arms (* indicates p < .05 by posthoc Shelf6 tests from single factor ANOVA of results of POST test). Full data shown in Table 2 demonstrates that the group differences shown here partly result from decreases in exploration of open arms in the POST test relative to the PRE test in groups other than MATED males.
the groups did not differ on other measures of social behavior during this interval, such as time spent huddling with a female. The MATED group also differed from both the EXPOSED and the CONTROL groups on exploration of the open arms of the plus maze (Fig. 2). Paternal behavior and tall-flick latency (an- algesia) were not affected by mating (Tables 3 & 4).
The change in aggression approximated the increase reported earlier (31). As in the previous report, the difference in aggres- sion was both quantitative and qualitative. Attack-bites, very rare prior to mating, represented approximately 20% of the aggressive repertoire after mating.(Fig. 1) Lunge-pushes appeared to in- crease equally in the MATED and NON-MATED males, sug- gesting that this behavior may be solicited by the estrous female independent of mating. As in our previous report, lateral threats were the prevalent form of aggression following mating. In con- trast to the previous report which noted a subtle but significant decrease in defensive behavior(31), in the current experiment de- fensive behavior did not change. This difference between the two
studies may reflect a cohort of less aggressive intruders used in the current study. As a result, across all groups, defensive be- havior was low both prior to and following female exposure. Indeed, some of the defensive behaviors observed in our original investigation, such as the submissive-supine posture, were vir- tually absent in the present study. The results from both studies in voles stand in contrast to a report from Albert and co-workers on the role of social experience and testosterone-dependent ag- gression in male rats (1). In this study, cohabitation (for 7-21 days as opposed to one day in the present experiment) with a female increased resident-intruder aggression even in castrated male rats. Although the role of sexual behavior in the facilitation of aggression is not clear in this report, the observation that social contact, even with a male conspecific, can increase aggression underscores the importance of species differences in the behav- ioral consequences of mating.
This study also examined several more common, activity-re- lated variables, such as rearing, sniffing, and grooming, in the

MATING IN THE M O N O G A M O U S MALE 621
TABLE 3
TAIL FLICK LATENCY FOR PRAIRIE VOLE MALES (MEANS _+ SEM SHOWN)
Group Pre Post
Control 2.1 ± 0.1 2.3 ___ 0.3 Exposed 2.2 ± 0.2 1.8 __+ 0.3 Non-mated 3.0 ± 0.4* 2.6 ± 0.4 Mated 1.5 ± 0.2 1.8 _.+ 0.4
Values represent means ± sem of median values from 3 trials in each animal.
Repeated measures ANOVA finds significant Group factor F(3, 32) = 3.14, p = 0.04).
Repeated measure (i.e., PRE vs. POST) and interaction terms not sig- nificant.
Single factor ANOVA finds significant difference in PRE values F(3, 35) = 5.52, p = .004).
* Signifies p < .05 by post-boc analysis between NON-MATED and MATED PRE values.
resident-intruder paradigm. There were no striking group effects observed on any of these measures nor was there any significant difference after mating, although the variance was high on some measures and it is possible that subtle differences in activity were overlooked. One other result from the resident-intruder test de- serves note. The increase in aggression in the MATED group (Table 1 & Fig. 1) was highly variable. Although overall there was a three-fold increase in offensive aggression, three of the eight males that mated showed little change (ie. increase less than 5 episodes of threats or bites per 6 minute test). It remains unclear why some males increase several-fold while others show little change in aggression after mating. This variability could not be attributed to baseline aggression or to mating behavior.
The MATED group also differed from both the EXPOSED and the CONTROL groups on exploration of the open arms of the plus maze. As group differences were not observed for entries into the closed arms, the increased frequency of entries into the open arms probably did not reflect simply increased activity. The specific increase in number of entries and time spent in the open arms of the plus maze were consistent with the notion that mating decreases fearfulness. As similar changes were not observed in any of the other experimental groups, this apparent decrease in fearfulness cannot be attributed to social contact or repeated test- ing. The results strongly suggest that this change is a conse- quence of the mating experience.
The failure to detect differences in paternal behavior is not altogether surprising given the short interval from mating or co- habitation to testing. In other rodent species, copulation has been reported to either reduce infanticide or promote paternal care, but these effects are usually delayed by several days (9,13,29). Bam- shad et al. have previously described increased paternal behavior in male prairie voles tested following various durations of female exposure (2). In this study with prairie voles, as with previous studies with mice, increases in paternal behavior were not ob- served after 3 days of cohabitation, but were significant by 13 days and appeared maximal at 21 days of cohabitation. Although these males were not monitored for mating, the females' subse- quent dates for parturition suggested that mating occurred in the first 3 days of cohabitation. Thus, in our study which looked at the earliest behavioral changes following mating, the failure to detect an increase in paternal behavior may reflect the longer time course for this behavior to be affected. It should also be consid- ered that the high levels of paternal care at baseline may have
obscured an increase following mating in our study (ie. ceiling effect).
As in the previous report, mating was associated with an in- crease in partner preference. This effect was more modest than expected. Indeed, there was no difference between MATED and EXPOSED males in the time spent in side-by-side contact with the partner (Fig. 3). The most consistent difference was, surpris- ingly, the reduced time spent in side-by-side contact with the stranger, suggesting that the MATED males were avoiding the stranger rather than attracted to the partner. This apparent stranger avoidance might simply reflect neophobia after mating, but the results from the plus maze test would argue against in- creased fearfulness in the MATED group. Moreover, the groups spent approximately equal time in the neutral cage, so that the decreased time with the stranger was not a reflection of gener- alized avoidance of social contact. The avoidance of the stranger in the MATED group could reflect a difference in the stimulus animal in this condition. In contrast to the EXPOSED condition, in the MATED condition the unfamiliar female had recently mated with another male. Although we know of no literature that suggests that a prairie vole male will avoid another male 's mate, females develop a preference of their own and may reject ad- vances from an unfamiliar male (30). Female-rejection seems a reasonable explanation for the apparent "avoidance" of the MATED males, however studies of prairie vole females suggest that 24 h of exposure without mating is sufficient for the induc- tion of a partner preference(30). Thus, female rejection should be equivalent in both the MATED and the EXPOSED groups and may not explain the observed difference in contact with the unfamiliar female. Finally, the MATED males appeared more active in the partner preference test, entering both Partner and Stranger cages more often than the EXPOSED males. Because of this activation, the ratio of time spent with partner vs. time spent with stranger may be a better reflection of the preference rather than absolute durations. These ratios for MATED males (6.6) and EXPOSED males (1.9) approximate the values previ- ously reported for female prairie voles tested under similar con- ditions after 6 h of mating or 6 h of male exposure, respectively (30).
The most troublesome part of this study was the group of males that failed to mate. These nonmating males have been noted in previous investigations both from our lab (31) and from other investigators working with prairie voles (Carter, personal comm.). The discovery that these males had higher tail-flick la- tencies at baseline might suggest that they were subordinate sib- lings that continued to behave as "defeated males" even after three days of individual housing (19). In the baseline resident- intruder test, their offensive aggression appeared reduced relative to the MATED males (5.6 __. 1.4 vs. 10.1 __+ 2.1 for NON- MATED and MATED, respectively), but there were no differ-
TABLE 4
PATERNAL BEHAVIOR 1N PRAIRIE VOLES (MEANS ~ SEM SHOWN)*
Pup Contact-Latency Pup Contact-Duration
Group Pre Post Pre Post
Control 15.5 ± 2.7 7.5 ± 1.2 382.3 _+ 86.4 377.5 ± 53.5 Exposed 20.1 _+ 4.7 74.7 ± 45.6 379.9 ± 53.5 387.5 ± 57.1 Non-mated 41.7 ± 18.5 90.4 + 83.6 458.0 ± 38.0 397.2 ± 96.1 Mated 10.1 _+ 2.9 13.0 ± 3.2 466.7 ± 70.7 439.1 ± 36.4
* Group means are not significantly different by ANOVA.

622 INSEL, PRESTON AND W I N S L O W
z
8 A .
7
6
5
4
3
2
1
O
40
T
• MATED I ~ E X P O S E D
PARTNER SXS STRANGER SXS
TIME SIDE-BY-SIDE
~o B .
z
PARTNER NEUTRAL STRANGER
TIME IN E A C H OF 3 C A G E S
¢~
ee t,-
PARTNER ENTRIES STRANGER ENTRIES
ENTRIES INTO EACH CAGE
FIG. 3. Partner preference in male prairie voles. Two groups of male prairie voles were compared after 24 h of mating (MATED, n = 9) or social exposure (EXPOSED, n = 9). In a 3 h test, males were given a choice of the familiar partner (partner) or a novel female (stranger) using a three- way test apparatus with restricted access for the females and unlimited access for the male. As shown in (A), MATED males spent less time with the stranger relative to time with the partner (# indicates p < .05 by paired t-test). MATED and EXPOSED males differed in time with the stranger (* indicates group difference, p < .05, by Student' s t-test) but not time with the partner. (B) There were no group differ- ences in time spent each of the three cages--indicating that the preference in (A) is a social preference not a place pref- erence. (C) MATED males were more active than EX- POSED males as demonstrated by increased numbers of en- tries into both partners and strangers cages.
ences at baseline between these groups on several other behav- ioral measures including plus maze exploration (baseline t ime in the open arms = 116.7 ___ 27.7 vs. 98.3 +_ "26.8 for MATED and NON-MATED, respectively). It remains unclear why these males failed to mate, al though a blockade of reproductive status would certainly be consistent with subordinate behavior. In other stud- ies, we have found that most will begin mating with more exten- sive female exposure - also consistent with a subordinate phe- notype.(Insel, unpublished data) But the most troublesome aspect of this group is the implication that both the EXPOSED and the CONTROL groups were comprised of two popu l a t i ons - - one of which was comparable to the M A T E D group and one comparable to the N O N - M A T E D group. Indeed, means for the EXPOSED and CONTROL groups on several baseline measures (tail-flick, resident-intruder aggression, plus maze activity) were interme- diate between M A T E D and N O N - M A T E D groups. For this rea- son, we believe that subsequent experiments will need to use males that are preselected and matched for their ability to mate.
E X P E R I M E N T 2: TIME COURSE OF M A T I N G EFFECTS ON AGGRESSION
Experiment 1 demonstrated that the critical stimulus for in- ducing aggression was mating and not social contact. To deter-
mine the min imum amount of mating sufficient to induce ag- gression, males were compared after variable intervals with sexually receptive females.
M E T H O D
Animals
This experiment involved 32 male prairie voles housed under the conditions described in Experiment 1.
Procedure
Males were prescreened for their ability to mate and then as- signed randomly to each of the following 4 groups: Control, Mated-1 h, Mated-6 h, Mated 24-h. Each of the Mated groups were exposed to an estrous female (in the male ' s home cage) and observed for mating (latency ~-- 1 h). From the the first bout of mating, males were given 1, 6, or 24 h with the same female. At the end of the assigned interval, females were removed. Control males did not receive female exposure. Tests were scheduled to try to minimize potential t ime of day effects (i.e., Mated-6 h. males were begun earlier in the day so that t ime of testing would

MATING IN THE MONOGAMOUS MALE 623
60
Z .¢ 50 r~
40 L~ .<
3O
;;,,.,
Z 20 r,,d
r.,d == 10
TIME FROM MATING:
E3mE [ ] ORB. [ ] 24HR.
1-
N O N - M A T I N G M A T I N G - 1 HR. MATING-6 HR. MA TI N G .24 HR.
(9) (9) (5) (9)
FIG. 4. Time course for mating induction of aggression in male prairie voles. Sexually inexperienced males spent either 1, 6, or 24 h with a sexually receptive female. All males were observed to mate. Aggression was scored as a composite of lateral threat, lunge push, and attack bites. Relative to aggression frequency during a 6 minute baseline resident-intruder test, aggression increased immediately after 1 h of mating but was not sustained. With 24 h of mating, aggression continued to be increased for at least 24 h (* signifies difference from PRE score, p < .05, by ANOVA.
approximate the time used for the Control and Mated-24 h groups).
Males were tested in the resident-intruder paradigm prior to female exposure (PRE), at the completion of the mating interval (0 h), and 24 h later (24 h). Control males were observed in the resident-intruder test three times, 24 h apart. Aggression was as- sessed as described above except that only the composite score of offensive aggression (attack-bite, lunge-push, lateral threat) was scored.
Data Analysis
Frequency of aggression was compared across groups at each time point with a factorial ANOVA. Differences from Control were defined as p < .05 by posthoc Sheff6 test.
RESULTS
Groups did not differ at baseline (PRE) in frequency of ag- gression. Immediately after mating (0 h), increased aggression (relative to Control) was observed in the Mated-1 h and Mated- 24 h groups (Fig. 4). This increase was only sustained in the Mated-24 h group which continued to show high frequencies of aggressive behavior at the 24 h timepoint (48 h after the onset of mating). At the analogous time point in the Mated-1 h group, aggression frequency had dropped to approximate the level ob- served in the PRE test. The Mated-6 h group showed an inter- mediate increase that was not statistically different from either the Control or the Mated-24 h group.
DISCUSSION
As expected from Experiment 1 and our previous report (31), 24 h of sexual experience was associated with a significant and
sustained increase in aggression. It also appears that there is an early increase in aggression, observed after 1 h, that is not sus- tained in the absence of continued mating. This may reflect ac- tivation of a process that requires additional mating to become sustained or there may be two quite different behavioral pro- cesses inducing aggression--an early process with the onset of mating and a later process (after 6 h) that is more enduring.
In our previous report, we described the induction of aggres- sion in male prairie voles with the chronic, continuous central administration of arginine-vasopressin (AVP). In that study, at least 12 h of continuous administration of AVP was required to induce aggression. Although we do not know precisely how much mating experience is necessary for males to demonstrate aggression, the results from the present experiment suggest that the critical period is between 6 and 24 h. It should also be noted that these results do not address the critical duration of mating during this 24 h interval. Because the 1 h and 6 h groups were separated from females to prevent additional mating, we cannot attribute the enduring aggression to persistent mating as opposed to continued social contact.
EXPERIMENT 3: BEHAVIORAL CONSEQUENCES OF MATING IN A NON-MONOGAMOUS VOLE
The rationale for studying aggression and affiliation in the prairie vole is that these behaviors, which can be operationally defined with the resident-intruder and partner preference tests, measure processes that are instrumental to pair bonding. Exper- iment 1 demonstrated the importance of mating in the induction of both the aggression and the affiliation. Experiment 2 demon- strated that the induction of aggression requires several hours of mating. Neither experiment has tested whether these behavioral changes are related to pair bonding. To address this question, we
624 INSEL, PRESTON AND WINSLOW
14
Z o 12
.~ 8
0
Z
4
2
[ ] Box
[ ] Lunge Push "-'1-- • Attack Bite /
0 PRE POST PRE POST
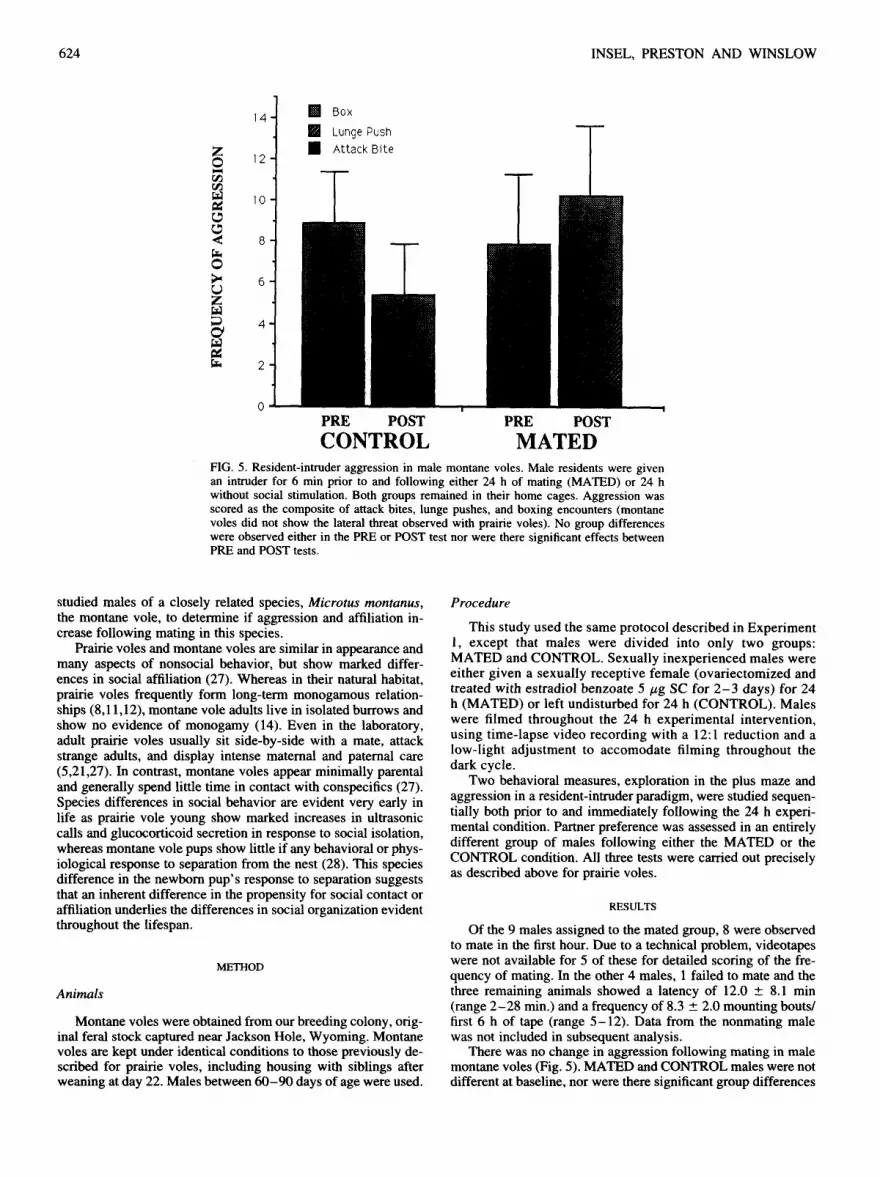
CONTROL MATED FIG. 5. Resident-intruder aggression in male montane voles. Male residents were given an intruder for 6 rain prior to and following either 24 h of mating (MATED) or 24 h without social stimulation. Both groups remained in their home cages. Aggression was scored as the composite of attack bites, lunge pushes, and boxing encounters (montane voles did not show the lateral threat observed with prairie voles). No group differences were observed either in the PRE or POST test nor were there significant effects between PRE and POST tests.
studied males of a closely related species, Microtus montanus, the montane vole, to determine if aggression and affiliation in- crease following mating in this species.
Prairie voles and montane voles are similar in appearance and many aspects of nonsocial behavior, but show marked differ- ences in social affiliation (27). Whereas in their natural habitat, prairie voles frequently form long-term monogamous relation- ships (8,11,12), montane vole adults live in isolated burrows and show no evidence of monogamy (14). Even in the laboratory, adult prairie voles usually sit side-by-side with a mate, attack strange adults, and display intense maternal and paternal care (5,21,27). In contrast, montane voles appear minimally parental and generally spend little time in contact with conspecifics (27). Species differences in social behavior are evident very early in life as prairie vole young show marked increases in ultrasonic calls and glucocorticoid secretion in response to social isolation, whereas montane vole pups show little if any behavioral or phys- iological response to separation from the nest (28). This species difference in the newborn pup's response to separation suggests that an inherent difference in the propensity for social contact or affiliation underlies the differences in social organization evident throughout the lifespan.
METHOD
Animals
Montane voles were obtained from our breeding colony, orig- inal feral stock captured near Jackson Hole, Wyoming. Montane voles are kept under identical conditions to those previously de- scribed for prairie voles, including housing with siblings after weaning at day 22. Males between 60-90 days of age were used.
Procedure
This study used the same protocol described in Experiment l, except that males were divided into only two groups: MATED and CONTROL. Sexually inexperienced males were either given a sexually receptive female (ovariectomized and treated with estradiol benzoate 5 #g SC for 2 - 3 days) for 24 h (MATED) or left undisturbed for 24 h (CONTROL). Males were filmed throughout the 24 h experimental intervention, using time-lapse video recording with a 12"l reduction and a low-light adjustment to accomodate filming throughout the dark cycle.
Two behavioral measures, exploration in the plus maze and aggression in a resident-intruder paradigm, were studied sequen- tially both prior to and immediately following the 24 h experi- mental condition. Partner preference was assessed in an entirely different group of males following either the MATED or the CONTROL condition. All three tests were carried out precisely as described above for prairie voles.
RESULTS
Of the 9 males assigned to the mated group, 8 were observed to mate in the first hour. Due to a technical problem, videotapes were not available for 5 of these for detailed scoring of the fre- quency of mating. In the other 4 males, 1 failed to mate and the three remaining animals showed a latency of 12.0 + 8.1 min (range 2-28 min.) and a frequency of 8.3 _ 2.0 mounting bouts/ first 6 h of tape (range 5-12). Data from the nonmating male was not included in subsequent analysis.
There was no change in aggression following mating in male montane voles (Fig. 5). MATED and CONTROL males were not different at baseline, nor were there significant group differences

M A T I N G IN T H E M O N O G A M O U S M A L E 625
T A B L E 5
EFFECTS OF MATING ON PLUS MAZE BEHAVIOR IN MONTANE VOLES
Control Mated Group T i m e Interaction
Measure Pre Post Pre Post F(I, 26) F(I, 26) F(1, 26)
Open (Time) 76.2 ± 18.7 33.3 ± 9.7 76.9 _+ 16.6 60.9 _+ 8.7 1.014 4.389* 0.916 Open (Frequency) 6.4 ± 1.2 5.0 ± 1.3 8.9 _+ 1.7 8.4 ± 0.7 5.165" 0.567 0.131 Closed (Time) 149.8 ± 20.8 196.3 _+ 12.8 148.2 _ 14.1 162.0 ___ 13.0 1.362 3.862 1.135 Closed (Frequency) 10.4 ± 2.0 14.0 _+ 3.4 14.5 _ 2.5 16.7 _+ 1.6 1.965 1.431 0.074 Center (Time) 74.0 ± 13.7 70.4 ± 9.1 74.9 _+ 9.1 77.1 ___ 8.5 0.137 0.004 0.081 Total Entries 16.9 _ 2.3 19.0 ± 3.7 23.4 _ 3.4 25.1 ± 2.0 4.674* 0.443 0.004 % Entries Open 38.6 ± 5.2 28.3 ± 5.3 39.8 _ 6.5 33.8 ± 2.5 0.432 2.508 0.173
Group represents comparison of Control to Mated; Time represents comparison of PRE to Post; *signifies p < .05. Note: Within group comparison of Pre and Post (paired t test) shows no differences for Mated, but trend of decreased open time and increased
closed time for controls.
6 ] A. [] MONTANE 5, T • PRAIRIE
• , ~ • °
7
T
10 B ,
8 T
PARTNER SXS STRANGER SXS
TIME SIDE-BY-SIDE
60
, '- 5O
4o
PARTNER ENTRIES STRANGER ENTRIES
ENTRIES INTO E A C H CAGE
PARTNER NEUTRAL STRANGER
T I M E IN EACH OF 3 CAGES
FIG. 6. Partner preference in male montane voles. Male montane voles (n = 9) were tested for a partner preference after 24 h of mating. There are no differences in (A) the time spent with partner vs. time spent with stranger, (B) time in the three cages, or (C) number of entries into partner's vs. stranger's cage. The absence of a preference contrasts with results with male prairie voles. Values for mated prairie voles from Fig. 3 are shown here for comparison.

626 INSEL, PRESTON AND WINSLOW
in the posttest or significant within-group differences comparing aggression scores in the pre and post tests.
Group differences (i.e., differences between MATED and CONTROL males) were not observed in the plus maze (Table 5). In the CONTROL males, there was a decrease in time spent in the open arms of the plus maze in the post test relative to the pre test (Paired t = 2.37, P = .05). This difference was not ob- served in the MATED males.
Partner preference was not observed in MATED montane voles. As can be seen in Fig. 6, montane vole males tended to spend more time in the neutral chamber (relative to prairie voles), but there was no evidence for a preference for either the partner or the stranger.
DISCUSSION
Montane vole males resembled prairie vole males in the gen- eral pattern of mating behavior (ie. latency and frequency of mounts), but failed to show the behavioral changes in aggression or plus maze exploration after mating observed in the prairie voles in Experiments 1 and 2. The apparent absence of an effect of mating could be obscured by a large effect of repeated testing in the montane voles, but this seems an unlikely explanation as the CONTROL males who failed to receive mating experience but were tested twice also showed no change in aggression and only a small albeit significant change in plus maze behavior.
It can be difficult to compare species when different behaviors are involved. As previously described (24), montane voles and prairie voles show different patterns of aggression (boxing in montane voles vs. lateral threats in prairie voles). In addition, their patterns of behavior in the partner preference test are dif- ferent at baseline, with the less social montane vole males spend- ing more time in the neutral chamber. Nevertheless, the within- species comparisons should not be affected by these baseline differences. If mating increased aggressive and affiliative behav- iors in both species, then these behavioral changes could not be attributed to pair-bonding. The failure to find these behavioral changes following mating in the montane vole, which does not
pair bond, is consistent with the hypothesis that both measures are involved in a process that occurs in prairie voles and not montane voles. These data are largely consistent with an earlier report by Shapiro and Dewsbury comparing the effects of mating on partner preferences in female prairie and montane voles (26). Moreover, the absence of a change in plus maze behavior fol- lowing mating in the montane vole underscores the importance of a reduction in fearfulness in this complex of behaviors puta- tively related to pair bonding in the prairie vole.
CONCLUSIONS
Taken together, these studies support the importance of mat- ing for pair bonding in the male prairie vole. Experiment 1 dem- onstrated that mating (but not nonsexual social contact) was as- sociated with an increase in aggression, the formation of a partner preference, and decreased fearfulness. Experiment 2 showed that more than 6 h of mating was necessary for the increase in ag- gression. Experiment 3 implicated the mating-induced changes in the prairie vole in the process of pair bonding as none of these changes were observed following mating in the closely related montane vole which does not form pair bonds. Williams et al.(30) have previously shown that mating facilitates the partner pref- erence observed in female prairie voles. The results with male prairie voles provide additional evidence for the critical role of mating in the formation of social attachments in this species. At the same time, the reader should note that some of the behaviors observed in prairie voles (e.g. patemal care, aggression) are not consistently present in other monogamous mammals. The role of mating in other monogamous species will need to be empirically demonstrated.
Our laboratory has been focused on the neural changes that subserve social attachments. The current studies are important in this effort because they suggest that studying the physiological consequences of prolonged mating in this species may provide the key to understanding pair bonding. Current investigations are examining the effects of prolonged mating on regional gene ex- pression in the brain of the male prairie vole.
REFERENCES
1. Albert, D.; Dyson, E.; Walsh, M.; Petrovic, D. Cohabitation with a female activates testosterone-dependent social aggression in male rats independently of changes in serum testosterone concentration. Physiol. Behav. 44:735-740; 1988.
2. Bamshad, M.; Novak, M.; De Vries, G. Cohabitation alters vaso- pressin and paternal behavior in prairie voles, Microtus ochrogaster. Physiol. Behav. 56(4):751-758; 1994.
3. Carter, C.; Getz, L. Monogamy and the prairie vole. Sci. Am. 268:100-106; 1993.
4. Carter, C. S.; Getz, L. L.; Cohen-Parsons, M. Relationships between social organization and behavioral endocrinology in a monogamous mammal. In: Rosenblatt, J. S.; Beer, C.; Busnel, M. C.; Slater, P. J. B., eds. Advances in the study of behavior. New York: Aca- demic Press, lnc; 1986.
5. Carter, C. S.; Williams, J. R.; Witt, D. M. The biology of social bonding in a monogamous mammal, ln: Balthazart, J., eds. Hor- mones, brain and behavior in vertebrates 2: Behavioral activation in males and females-social interactions and reproductive endocrinol- ogy. Basel, A G: S Karger; 1990:154-164.
6. Clutton-Brock, T. H. Mammalian mating systems. Proc. R. Soc. Lond. 236:339-372; 1989.
7. D'Amour, F.; Smith, D. A method for determining loss of pain sen- sation. J. Pharmacol. Exp. Ther. 72:74-79; 1941.
8. Dewsbury, D. A. An exercise in the prediction of monogamy in the field from laboratory data on 42 species of muroid rodents. Biologist 63:138-162; 1981.
9. Elwood, R.; Ostermeyer, M. Does copulation inhibit infanticide in rodents? Anim. Behav. 32:293-205; 1984.
10. File, S. The use of social interaction as amethod for detecting the anxiolytic activity of chlordiazepoxide-like drugs. J. Neurosci. Methods 2:219-238; 1980.
1 I. Getz, L. L.; Carter, C. S.; Gavish, L. The mating system of the prairie vole Microtus ochrogaster: Field and laboratory evidence for pair bonding. Behav. Ecol. Sociobiol. 8:189-194; 1981.
12. Getz, L. L.; Hofman, J. E. Social organization in free living prairie voles, Microtus ochrogaster. 1986.
13. Gubernick, D.; Schneider, K.; Jeannotte, L. Individual differences in the mechanisms underlying the onset and maintenance of paternal behavior and the inhibition of infanticide in the monogamous bi- parental California mouse, Peromyscus californicus. Behav. Ecol. Sociobiol. (in press).
14. Jannett, F. J. Social dynamics in the montane vole Microtus mon- tanus as a paradigm. Biologist 62:3-19; 1980.
15. Keverne, E. B.; Martensz, N.; Tuite, B. Beta-endorphin concentra- tions in CSF of monkeys are influenced by grooming relationships. Psychoneuroendo 14:155-161; 1989.
16. Kleiman, D. G. Monogamy in mammals. Q. Rev. Biol. 52:39-69; 1977. 17. Lister, R. The use of a plus-maze to measure anxiety in the mouse.
Psychopharmacology (Berlin) 92:180-185; 1987. 18. Martinez-Gomez, M.; Cruz, Y.; Salas, M.; Hudson, R.; Pacheco, P.
Assessing pain threshold in the rat: Changes with estrus and time of day. Physiol. Behav. 55:651-657; 1994.

M A T I N G IN THE M O N O G A M O U S MALE 627
19. Miczek, K; Thompson, M.; Shuster, L. Opioid-like analgesia in de- feated mice. Science 215:1520-1522; 1982.
20. Miczek, K. A.; Winslow, J, T. Psychopharmacologic research on ag- gressive behavior. In: Greenshaw, A. J.; Dourish, C. T., eds. Experi- mental psychopharmacology. Clifton, N J: Humana Press; 1987:27-113.
21. Oliveras, D.; Novak, M. A comparison of paternal behaviour in the meadow vole Microtus pennsylvanicus, the pine vole M. pinetorum and the prairie vole M. ochrogaster. Anim. Behav. 34:519-526; 1986.
22. Pellow, S.; Chopin, P.; File, S.; Briley, M. Validation of open: Closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci. Methods 14:149-167; 1985.
23. Pellow, S.; File, S. Anxiolytic and anxiogenic drug effects in ex- ploratory activity in an elevated plus-maze: A novel measure of anx- iety in the rat. Pharmacol. Biochem. Behav. 24:525-529; 1986.
24. Pierce, J. D. J.; Pellis, V. C.; Dewsbury, D. A.; Pellis, S. M. Targets and tactics of agonistic and precopulatory behavior in montane and prairie voles. Agg. Beh. 17:337-349; 1991.
25. Rodgers, R.; Cole, J. Influence of social isolation, gender, strain, and prior novelty on plus-maze behaviour in mice. Physiol. Behav. 4:729-736: 1993.
26. Shapiro, L.; Austin, D.; Ward, S.; Dewsbury, D. Familiarity and female mate choice in two species of voles (Microtus ochrogaster and Microtus montanus). Anim. Behav. 34:90-97; 1986.
27. Shapiro, L. E.; Dewsbury, D. A. Differences in affiliative behavior, pair bonding, and vaginal cytology in two species of vole. J. Comp. Psychol. 104:268-274; 1990.
28. Shapiro, L. E.; Insel, T. R. Infant's response to social separation reflects adult differences in affiliative behavior: A comparative de- velopmental study in prairie and montane voles. Dev. Psychobiol. 23:375-394; 1990.
29. vom Saal, F. Time-contingent change in infanticide and parental behavior induced by ejaculation in male mice. Physiol. Behav. 34:7- 15; 1985.
30. Williams, J. R.; Catania, K. C.; Carter, C. S. Development of partner preference in female prairie voles (Microtus ochrogas- ter): Role of social and sexual experience. Horm. Behav. 26:339- 349; 1992.
31. Winslow, J. T.; Hastings, N.; Carter, C. S.; Harbaugh, C. R.; Insel, T. R. A role for central vasopressin in pair bonding in monogamous prairie voles. Nature 365:545-548; 1993.