low external ph induces hog1‐dependent changes in the organization of the saccharomyces cerevisiae...
TRANSCRIPT

Molecular Microbiology (2001) 39(2), 469±479
Low external pH induces HOG1-dependent changesin the organization of the Saccharomyces cerevisiaecell wall
J. C. Kapteyn,² B. ter Riet, E. Vink, S. Blad,
H. De Nobel, H. Van Den Ende and F. M. Klis*
Fungal Research Group, Swammerdam Institute of Life
Sciences, University of Amsterdam, Kruislaan 318,
1098 SM Amsterdam, The Netherlands.
Summary
Low environmental pH strongly affected the organiz-
ation of the Saccharomyces cerevisiae cell wall,
resulting in rapidly induced resistance to b1,3-
glucanase. At a molecular level, we found that a
considerable amount of Cwp1p became anchored
through a novel type of linkage for glycosylpho-
sphatidylinositol (GPI)-dependent cell wall proteins,
namely an alkali-labile linkage to b1,3-glucan. This
novel type of modification for Cwp1p did not require
the presence of a GPI-derived structure connecting
the protein with b1,6-glucan. In addition, we found
high levels of Cwp1p, which was double-anchored
through both the novel alkali-sensitive bond to b1,3-
glucan and the alkali-resistant GPI-derived linkage to
b1,6-glucan. Further cell wall analyses demonstrated
that Pir2p/Hsp150 and possibly other Pir cell wall
proteins, which were already known to be linked to
the b1,3-glucan framework by an alkali-sensitive
linkage, were also more efficiently retained in the
cell wall at pH 3.5 than at pH 5.5. Consequently, the
alkali-sensitive type of linkage of cell wall proteins to
b1,3-glucan was induced by low pH. The low pH-
induced alterations in yeast cell wall architecture
were demonstrated to be dependent on a functional
HOG1 gene, but not on the Slt2p-mediated MAP
kinase pathway. Consistent with this observation,
DNA microarray studies revealed transcriptional
induction of many known high-osmolarity glycerol
(HOG) pathway-dependent genes, including four cell
wall-related genes, namely CWP1, HOR7, SPI1 and
YGP1.
Introduction
The molecular organization of the cell wall of the baker's
yeast Saccharomyces cerevisiae is relatively well docu-
mented in comparison with that of other fungi (KollaÂr et al.,
1995; 1997; Lipke and Ovalle, 1998; Kapteyn et al., 1999a;
Smits et al., 1999). The S. cerevisiae cell wall consists of
b1,3-glucan, b1,6-glucan, chitin and mannoproteins in
proportions of approximately 55%, 5%, about 2% and
40% of the cell wall dry weight respectively (Orlean, 1997).
The b1,3-glucan chains constitute a three-dimensional
resilient framework that, together with chitin, accounts for
the cell's shape and its physical strength. The b1,3-glucan
molecules have multiple side-branches, which may function
as attachment sites for chitin and b1,6-glucan chains. The
b1,6-glucan molecules interconnect the largest class of
covalently linked cell wall proteins (CWPs), the glycosyl-
phosphatidylinositol (GPI)-CWPs, to the b1,3-glucan
framework (Kapteyn et al., 1996; 1999a; KollaÂr et al.,
1997). The other, less abundant group of covalently linked
CWPs, known collectively as Pir-CWPs, are linked to
b1,3-glucan without an interconnecting b1,6-glucan chain
(Kapteyn et al., 1999b) and can be liberated from the wall
by mild alkali extraction (Mrsa et al., 1997).
The molecular architecture of the cell wall of S.
cerevisiae has a highly dynamic nature. This is illustrated
by yeast mutants with reduced levels of b1,3- and/or b1,6-
glucan, which display a characteristic set of modifications
in their cell wall structure (Kapteyn et al., 1997; 1999b).
First, yeast walls of these glucan-deficient mutants are
enriched in chitin (Kapteyn et al., 1997, 1999b; Popolo
et al., 1997; Dallies et al., 1998; Ram et al., 1998; Ketela
et al., 1999; Garcia-Rodriguez et al., 2000a; Valdivieso et al.,
2000), and the number of linkages between chitin and b1,6-
glucan is augmented (Kapteyn et al., 1997). This augmenta-
tion is particularly evident in b1,3-glucan-deficient mutants.
Secondly, the expression of several GPI-CWPs, Pir-
CWPs and of Fks2p, the alternative catalytic subunit of
b1,3-glucan synthase, also becomes upregulated (Mazur
et al., 1995; Ram et al., 1998; Kapteyn et al., 1999b; De
Nobel et al., 2000; Terashima et al., 2000). For example,
Cwp1p is a GPI-CWP that is consistently induced upon
cell wall weakening (Ram et al., 1998; Kapteyn et al.,
1999a; Terashima et al., 2000). Thirdly, in mutant cells
with low levels of b1,6-glucan, a strong decrease in the
Q 2001 Blackwell Science Ltd
Accepted 18 October, 2000. *For correspondence. E-mail [email protected]; Tel. (131) 20 525 7834; Fax (131) 20 525 7934. ²Presentaddress: Crucell, PO Box 2048, 2301 CA Leiden, The Netherlands.

level of GPI-CWPs is accompanied by a concomitant
increase in the incorporation of Pir-CWPs (Kapteyn et al.,
1999b).
Similar cell wall compensatory responses to those
outlined above have also been demonstrated in yeast cells
challenged with cell wall-destabilizing agents (Roncero and
Duran, 1985; Ketela et al., 1999; De Nobel et al., 2000).
Consequently, these data vividly illustrate the ability of
yeast cells to adapt the architecture of their cell wall as a
protective mechanism against cell wall-weakening condi-
tions. Several lines of evidence indicate that cell wall
weakening is sensored by the Wsc and Mid families of
membrane proteins and relayed via Rho1p to the PKC1-
SLT2 signal transduction pathway, resulting in an
enhanced expression of many cell wall-related genes
(Popolo et al., 1997; Verna et al., 1997; Bickle et al., 1998;
Zhao et al., 1998; Jung and Levin, 1999; Ketela et al.,
1999; Rajavel et al., 1999; Smits et al., 1999; De Nobel
et al., 2000; Martin et al., 2000; Turchini et al., 2000).
Interestingly, evidence is also accumulating that the high-
osmolarity glycerol (HOG) pathway is implicated in
modulating the expression of several cell wall-related
genes in response to hyperosmotic shock (Rep et al.,
2000), and under other cell wall stress situations (Jiang
et al., 1995; Bourdineaud, 2000; Garcia-Rodriguez et al.,
2000b).
Here, we demonstrate that the molecular organization
of the cell wall of S. cerevisiae is strongly affected by
external pH. Cell wall analyses revealed that, at pH 3.5,
yeast cells exhibited high levels of incorporation of Cwp1p
through an alkali-sensitive bond to b1,3-glucan, which
represents a novel type of linkage for GPI-CWPs.
Concomitantly, the level of Cwp1p being double-anchored
through an alkali-sensitive bond to b1,3-glucan and an
alkali-resistant bridge to b1,6-glucan was also increased
under low pH conditions. At pH 3.5, the levels of
incorporation of Pir2p and, possibly, other Pir-CWPs
through an alkali-sensitive linkage to b1,3-glucan were
also higher than at pH 5.5. These low pH-induced
structural changes were demonstrated to be dependent
on a functional HOG1 gene, which is consistent with DNA
microarray studies, revealing transcriptional induction of
many known HOG pathway-dependent genes, including
CWP1, when yeast cells were confronted with a pH shift
from 5.5 to 3.5.
Results
Low extracellular pH decreases Quantazyme sensitivity of
yeast cells
b1,3-Glucanase sensitivity assays have been shown to be
valuable in monitoring alterations in yeast cell wall
architecture induced by various stress situations (Popolo
et al., 1993; Shimoi et al., 1998; Ovalle et al., 1998; De
Nobel et al., 2000). Here, we used this assay to study the
molecular organization of cell walls isolated from a yeast
wild-type strain cultivated at pH 3.5, pH 4.5, pH 5.5 and
pH 6.5. The pH of the growth medium was shown to have
a dramatic effect on the resistance of intact yeast cells to
Quantazyme, a recombinant b1,3-glucanase (Fig. 1A).
Decreasing the pH of the medium resulted in cells
becoming increasingly resistant (up to almost 100%)
(Fig. 1A). The cells reacted rapidly to lowering the pH of
the medium from 5.5 to 3.5. Within 30 min after the shift,
the cells exhibited a significant increase (< fourfold) in
resistance to Quantazyme (Fig. 1B). This response was
not exhibited by cells that continued to grow at pH 5.5
(data not shown). It is also noted that, during the duration
of all experiments, external pH values were stable.
Hypothetically, the observed resistance to Quantazyme
exhibited by cells grown at low pH might be attributed to
an enhanced number of linkages between b-glucan
molecules and chitin. Alternatively, the external protein
layer of the cell wall, which limits the accessibility of the
internal glucan layer to b1,3-glucanase in intact cells,
might have become less permeable (De Nobel et al.,
1990). The first possibility, however, did not turn out to be
the case, as the differential Quantazyme sensitivity was
displayed only by intact cells and not by isolated SDS-
extracted cell walls, indicating that the external protein
layer was affected at acidic pH. Consistent with this
observation, chitin levels were hardly affected by the
external pH, varying between 1% and 2% of cell wall dry
weight (Table 1). At pH 3.5, the chitin content was even
reduced by about 40% compared with the chitin level of
cells grown at pH 5.5 (Table 1). Unexpectedly in view of
the data from the Quantazyme sensitivity assay, the total
protein content of isolated cell walls was also not greatly
affected, varying between 4% and 5% of cell wall dry
weight (Table 1). However, yeast cells grown at low pH
did tend to have slightly higher levels of CWPs than those
grown at higher pH (Table 1).
Efficiency of Pir2/Hsp150 incorporation in the cell wall is
enhanced at low external pH
The observed lack of large quantitative differences in total
CWP expression by cells grown at differential pH
prompted us to look for pH-dependent qualitative differ-
ences in the expression of CWPs. In S. cerevisiae, two
major classes of CWPs have been identified, namely
the b1,6-glucanase-extractable GPI-CWPs (Fig. 7A) and
the b1,6-glucanase-resistant, b1,3-glucanase- or alkali-
extractable Pir-CWPs (Fig. 7B; Kapteyn et al., 1999a).
Hence, we decided to look for pH-dependent changes in
the relative frequency of cell wall incorporation of these
two classes of CWPs. Interestingly, we observed clear
470 J. C. Kapteyn et al.
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 39, 469±479

pH-induced alterations in the glucanase extractability
of CWPs (Table 1). At pH 6.5, the b1,6-glucanase-
extractable GPI-CWPs and the b1,6-glucanase-resistant
Pir-CWPs accounted for approximately 85% and 14% of
the total CWP content respectively (Table 1). In contrast,
these percentages shifted to 55% and 44%, respectively,
when yeast cells were grown at pH 3.5 (Table 1).
Consequently, these data suggest that the expression
and immobilization of Pir-related CWPs in the cell wall
were induced under conditions of low pH. Accordingly,
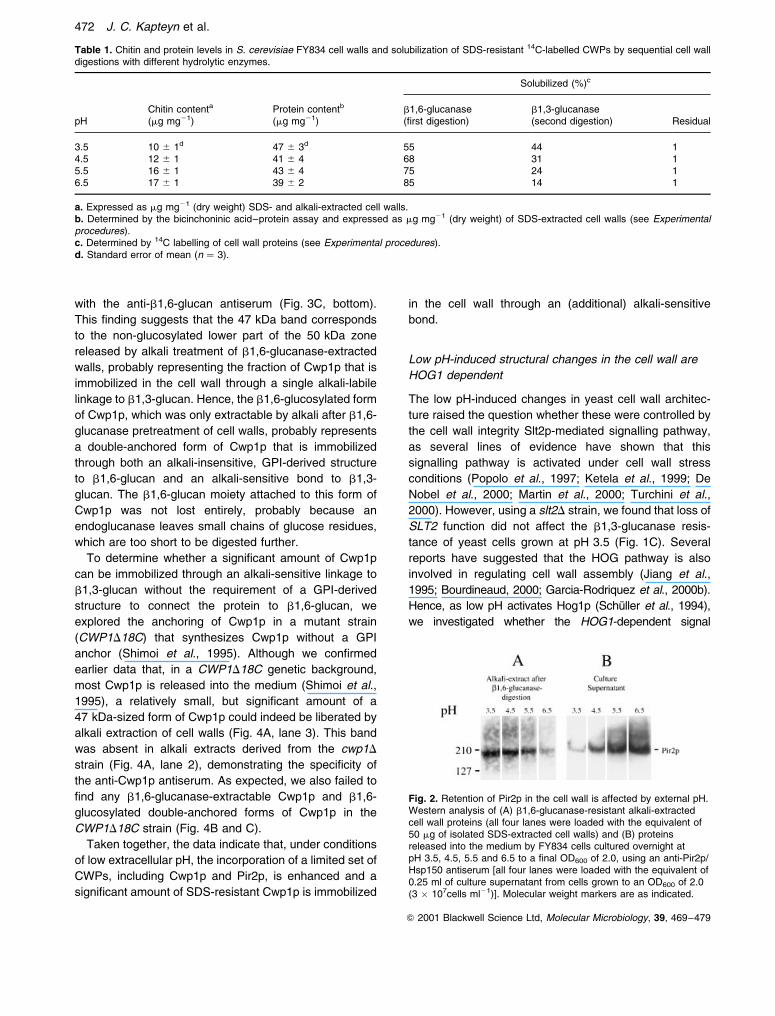
Western analyses demonstrated that the b1,6-glucanase-
resistant, alkali-releasable wall protein fractions from cells
grown at pH 3.5 and pH 4.5 contained higher levels of
Pir2p/Hsp150 than the corresponding ones from cells
grown at higher pH values (Fig. 2A). As expected, Pir2p/
Hsp150 was not solubilized by b1,6-glucanase digestion
(data not shown). We also determined the effect of
external pH on the level of Pir2p released into the
medium. At pH values of 4.5 and higher, increasing
amounts of Pir2p were secreted into the medium
(Fig. 2B). In stark contrast, under low pH conditions,
hardly any Pir2p was found to be released (Fig. 2B).
These findings strongly suggested that the increased level
of Pir2p in the wall of cells grown at low pH is largely the
result of a more efficient incorporation of this protein.
Level of b1,6-glucanase-resistant Cwp1p is enhanced at
low extracellular pH
We have shown previously that, in cells grown at pH 5.5,
most Cwp1p is b1,6-glucanase extractable, whereas only
a trace amount of Cwp1p is b1,6-glucanase resistant
(Kapteyn et al., 1999b). b1,6-Glucanase-resistant Cwp1p
has been suggested to be double-anchored through an
alkali-insensitive GPI-derived structure to b1,6-glucan and
through an unclarified alkali-sensitive linkage to b1,3-glucan
(Kapteyn et al., 1999b). Using an anti-Cwp1p antiserum,
we looked for pH-dependent changes in the level of both
b1,6-glucanase-extractable Cwp1p and b1,6-glucanase-
resistant, alkali-extractable Cwp1p. The level of b1,6-
glucanase-extractable Cwp1p was not significantly
affected by external pH (Fig. 3A, top). However, at
pH 3.5 and 4.5, a large increase was seen in the level
of b1,6-glucanase-resistant, alkali-extractable Cwp1p
(Fig. 3B, top). b1,6-Glucanase-resistant Cwp1p migrated
as a relatively broad zone with an average molecular
mass of 50 kDa (Fig. 3B, top). Interestingly, upon strip-
ping and reprobing the blot with an anti-b1,6-glucan
antiserum, the upper, but not the lower, part of this 50 kDa
zone was shown to contain a small b1,6-glucan moiety
(Fig. 3B, bottom). Consequently, these findings indicate
that alkali extraction of b1,6-glucanase-pretreated cell
walls results in the release of a b1,6- and a non-
glucosylated form of Cwp1p, which apparently migrate
very close to each other in SDS±PAGE. As a next step,
we explored whether these two b1,6-glucanase-resistant
forms of Cwp1p could also be released directly by mild
alkali treatment. Alkali extraction of untreated cell walls
resulted in the release of only a relatively discrete 47 kDa-
sized form of Cwp1p (Fig. 3C, row 1) that did not react
Fig. 1. Cells cultured at low pH exhibit Hog1p-dependent resistanceto b1,3-glucanase digestion. b1,3-Glucanase sensitivity of (A)FY834 cells cultivated at various pHs to a final OD600 of 2.0; (B)FY834 cells grown at pH 5.5 to OD600 of 0.3 and subsequentlyincubated at pH 3.5 for various periods of time; and (C) FY834 (X),hog1D (W) and slt2D (B) cells grown at pH 3.5 to a final OD600 of2.0. Cells were washed and digested with a recombinant b1,3-glucanase Quantazyme as described in Experimental procedures.The decrease in OD600 represents cell lysis and was expressed asa percentage of the starting OD600.
Low pH-induced alterations in yeast cell wall architecture 471
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 39, 469±479
with the anti-b1,6-glucan antiserum (Fig. 3C, bottom).
This finding suggests that the 47 kDa band corresponds
to the non-glucosylated lower part of the 50 kDa zone
released by alkali treatment of b1,6-glucanase-extracted
walls, probably representing the fraction of Cwp1p that is
immobilized in the cell wall through a single alkali-labile
linkage to b1,3-glucan. Hence, the b1,6-glucosylated form
of Cwp1p, which was only extractable by alkali after b1,6-
glucanase pretreatment of cell walls, probably represents
a double-anchored form of Cwp1p that is immobilized
through both an alkali-insensitive, GPI-derived structure
to b1,6-glucan and an alkali-sensitive bond to b1,3-
glucan. The b1,6-glucan moiety attached to this form of
Cwp1p was not lost entirely, probably because an
endoglucanase leaves small chains of glucose residues,
which are too short to be digested further.
To determine whether a significant amount of Cwp1p
can be immobilized through an alkali-sensitive linkage to
b1,3-glucan without the requirement of a GPI-derived
structure to connect the protein to b1,6-glucan, we
explored the anchoring of Cwp1p in a mutant strain
(CWP1D18C) that synthesizes Cwp1p without a GPI
anchor (Shimoi et al., 1995). Although we confirmed
earlier data that, in a CWP1D18C genetic background,
most Cwp1p is released into the medium (Shimoi et al.,
1995), a relatively small, but significant amount of a
47 kDa-sized form of Cwp1p could indeed be liberated by
alkali extraction of cell walls (Fig. 4A, lane 3). This band
was absent in alkali extracts derived from the cwp1D
strain (Fig. 4A, lane 2), demonstrating the specificity of
the anti-Cwp1p antiserum. As expected, we also failed to
find any b1,6-glucanase-extractable Cwp1p and b1,6-
glucosylated double-anchored forms of Cwp1p in the
CWP1D18C strain (Fig. 4B and C).
Taken together, the data indicate that, under conditions
of low extracellular pH, the incorporation of a limited set of
CWPs, including Cwp1p and Pir2p, is enhanced and a
significant amount of SDS-resistant Cwp1p is immobilized
in the cell wall through an (additional) alkali-sensitive
bond.
Low pH-induced structural changes in the cell wall are
HOG1 dependent
The low pH-induced changes in yeast cell wall architec-
ture raised the question whether these were controlled by
the cell wall integrity Slt2p-mediated signalling pathway,
as several lines of evidence have shown that this
signalling pathway is activated under cell wall stress
conditions (Popolo et al., 1997; Ketela et al., 1999; De
Nobel et al., 2000; Martin et al., 2000; Turchini et al.,
2000). However, using a slt2D strain, we found that loss of
SLT2 function did not affect the b1,3-glucanase resis-
tance of yeast cells grown at pH 3.5 (Fig. 1C). Several
reports have suggested that the HOG pathway is also
involved in regulating cell wall assembly (Jiang et al.,
1995; Bourdineaud, 2000; Garcia-Rodriquez et al., 2000b).
Hence, as low pH activates Hog1p (SchuÈller et al., 1994),
we investigated whether the HOG1-dependent signal
Table 1. Chitin and protein levels in S. cerevisiae FY834 cell walls and solubilization of SDS-resistant 14C-labelled CWPs by sequential cell walldigestions with different hydrolytic enzymes.
Solubilized (%)c
pHChitin contenta
(mg mg21)Protein contentb
(mg mg21)b1,6-glucanase(first digestion)
b1,3-glucanase(second digestion) Residual
3.5 10 ^ 1d 47 ^ 3d 55 44 14.5 12 ^ 1 41 ^ 4 68 31 15.5 16 ^ 1 43 ^ 4 75 24 16.5 17 ^ 1 39 ^ 2 85 14 1
a. Expressed as mg mg21 (dry weight) SDS- and alkali-extracted cell walls.b. Determined by the bicinchoninic acid±protein assay and expressed as mg mg21 (dry weight) of SDS-extracted cell walls (see Experimentalprocedures).c. Determined by 14C labelling of cell wall proteins (see Experimental procedures).d. Standard error of mean (n � 3).
Fig. 2. Retention of Pir2p in the cell wall is affected by external pH.Western analysis of (A) b1,6-glucanase-resistant alkali-extractedcell wall proteins (all four lanes were loaded with the equivalent of50 mg of isolated SDS-extracted cell walls) and (B) proteinsreleased into the medium by FY834 cells cultured overnight atpH 3.5, 4.5, 5.5 and 6.5 to a final OD600 of 2.0, using an anti-Pir2p/Hsp150 antiserum [all four lanes were loaded with the equivalent of0.25 ml of culture supernatant from cells grown to an OD600 of 2.0(3 � 107cells ml21)]. Molecular weight markers are as indicated.
472 J. C. Kapteyn et al.
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 39, 469±479

transduction cascade was involved in the observed
changes in cell wall organization. We demonstrated that,
under low pH conditions, hog1D cells completely lost the
ability to resist cell lysis by b1,3-glucanase digestion
(Fig. 1C), whereas at pH 5.5, wild-type, slt2D and hog1D
cells were equally sensitive to b1,3-glucanase (data not
shown). Therefore, the HOG pathway is strongly impli-
cated in mediating the structural alterations in the cell wall
in response to low external pH. Notably, like the parental
wild-type strain, hog1D cells did become resistant to b1,3-
glucanase upon entering the stationary phase (data not
shown), confirming that yeast cell wall architecture is also
changed upon diauxic shift (De Nobel et al., 1990), but
that this is HOG1 independent. In comparison with wild-
type cells, the hog1D strain did not exhibit any obvious
growth defect when grown at low pH (data not shown),
indicating that the Hog1p-dependent changes in cell wall
architecture are non-essential under low pH conditions.
Next, we explored the possibility that the observed
b1,3-glucanase sensitivity of hog1D cells at pH 3.5 is
related to a decreased ability to enhance the levels of
Cwp1p and Pir2p in the cell wall. Consistent with the
increased b1,3-glucanase sensitivity of the hog1D strain
at pH 3.5, the levels of both the b1,6-glucanase- and
alkali-extractable forms of Cwp1p were markedly reduced
(Fig. 5). In contrast, at all external pH values tested, the
level of Pir2p was not or hardly at all affected by the loss
of HOG1 function (data not shown). Consequently, unlike
Cwp1p, the increase in Pir2p in the cell wall at acidic pH
does not rely on the HOG1 gene functions.
Genomic analysis of gene expression induced by low
environmental pH
Several lines of evidence suggested that the regulation of
low pH-induced alterations in the cell wall is determined at
the transcriptional level. First, we perceived that the
kinetics of acquisition of resistance to b1,3-glucanase
were rapid; within 30 min after pH shift from 5.5 to 3.5,
cells were already markedly resistant to b1,3-glucanase
digestion (Fig. 1B). Secondly, Northern analyses revealed
that transcription of CWP1 mRNA was rapidly induced by
about threefold, already peaking at < 30 min after shifting
from pH 5.5 to 3.5 (data not shown). To identify other
transcriptional changes in response to acidification of the
medium, we performed genomic microarray experiments.
Based on the transcriptional kinetics of CWP1 (data
not shown) and the kinetics of Quantazyme resistance
displayed by S. cerevisiae upon pH shift (Fig. 1B), these
microarray experiments were conducted with RNA iso-
lated from yeast cells at two time points, namely 30 and
120 min after the pH shift. We found that the mRNA level
of a total of 36 genes, including four cell wall-related ones,
was on average in three hybridization experiments at least
1.4-fold higher when cells were grown for 30 and/or
120 min after shifting from pH 5.5 to 3.5. As expected
from our Northern analyses, CWP1 was one of the low
pH-induced genes and displayed a significantly higher
level of transcriptional accumulation after 30 min than
after 2 h of growth at pH 3.5 (Fig. 6; Table 2). Transcrip-
tion levels of the three other cell wall-related genes, SPI1,
HOR7 and YGP1, were higher after growth for 2 h than
after growth for 30 min (Table 2). SPI1 encodes a
stationary phase-induced GPI-CWP of unknown function,
which has recently been found also to be induced under
heat shock and osmotic stress (Puig and PerezOrtin,
Fig. 3. Augmented retention of Cwp1p in thecell wall through an alkali-sensitive linkage tob1,3-glucan in response to low external pH.Western analysis of (A) b1,6-glucanase-extracted cell wall proteins; (B) b1,6-glucanase-resistant, alkali-extracted cell wallproteins; and (C) cell wall proteins extracteddirectly by alkali treatment of FY834 cellsgrown at different extracellular pHs. All laneswere loaded with the equivalent of 50 mg ofisolated SDS-extracted cell walls. External pHvalues, molecular weight markers andantisera used are as indicated.
Fig. 4. Retention of Cwp1p through an alkali-sensitive linkage tob1,3-glucan does not require a GPI anchor addition sequence.Western analysis of (A) alkali-extracted cell wall proteins, (B) b1,6-glucanase-extracted cell wall proteins and (C) b1,6-glucanase-resistant, alkali-extracted cell wall proteins of YHP499 (wild type),YHS201 (cwp1D) and YHS211 (CWP1D18C) cells grown at pH 3.5using anti-Cwp1p antiserum. All lanes were loaded with theequivalent of 50 mg of isolated SDS-extracted cell walls. Molecularweight markers are as indicated.
Low pH-induced alterations in yeast cell wall architecture 473
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 39, 469±479

2000; Rep et al., 2000). HOR7 also codes for a putative
GPI-CWP of unknown function, which is reported to be
involved in the hyperosmolarity response (Hirayama et al.,
1995) and to be transiently repressed in response to a-
factor mating pheromone (Seidl and Tanner, 1997). YGP1
encodes a highly glycosylated extracellular protein of
unknown function produced in response to nutrient
depletion (Destruelle et al., 1994) and secreted into the
medium by regenerating spheroplasts (Pardo et al.,
1999). Notably, transcription of PIR2 was only slightly
(1.2-fold) induced, supporting the data (Fig. 2) that
suggest that the low pH-dependent increased incorpora-
tion of Pir2p in the wall results from increased incor-
poration efficiency rather than a massively enhanced
expression of the protein. The transcription of PIR1 and
PIR3 was induced to a similar fold to that found for PIR2
(data not shown).
The other groups of genes that were induced by at least
1.4-fold are related to carbohydrate metabolism [GCY1,
GPD1, GPP2/HOR2, GRE3, HXK1, HXT6, SOL4, TPS1
(YBR126C), TSL1 (YML100W)], redox metabolism
(ALD2, ALD3, MCR1, YER053C YNL134C), control of
gene expression (CUP1), nuclear pore transport (GSP2),
protection against various stress conditions (CTT1,
DDR2, GRE2, HSP12, HSP42, YDL204W, YDR453C,
YML128C) or encode proteins of unknown function
(YDR070C, YDR533C, YGL037C, YGR043C, YHR056C,
YHR087W, YIL169C). As expected from the observed
Hog1p-dependent changes in the molecular architecture
upon shifting from pH 5.5 to 3.5 (Fig. 1), most low pH-
induced genes (29 out of 36), including the four cell wall-
related genes (Table 2), have previously also been reported
to be induced after osmotic shock with 0.7 M NaCl (Rep
et al., 2000). Of these genes, 23 genes have previously
also been shown to be dependent on HOG1 and/or
MNS2/4 (Costanzo et al., 2000; Rep et al., 2000).
Consistent with these observations, most of the low pH-
induced genes contain several transcriptional stress
response elements (STREs) that have been demon-
strated to mediate gene transcriptional induction by heat
shock, nitrogen starvation, oxidative stress (Marchler
et al., 1993), hyperosmotic stress and, interestingly, also
by low external pH (SchuÈ ller et al., 1994).
Transcription of only three genes (YCL075W,
YEL071W, HXT2) was at least 1.4-fold higher at pH 5.5
than at pH 3.5. The first two open reading frames (ORFs)
are genes with unknown function, and HXT2 is involved
in hexose transport across the plasma membrane
(Costanzo et al., 2000).
Discussion
In this paper, we demonstrate that S. cerevisiae alters its
molecular cell wall organization when confronted with
changes in environmental pH. At acidic pHs, much higher
levels of Pir2p and, possibly, other Pir-CWPs were found
in the cell wall (Table 1, Fig. 3), concomitantly with the
acquisition of resistance to cell lysis by b1,3-glucanase
digestion (Fig. 1). The higher level of Pir2p found in the
cell wall at acidic pHs resulted from more efficient
incorporation rather than an increased expression of
Pir2p, as hardly any Pir2p was secreted into the medium
at pH 3.5 in contrast to the situation at pH 5.5 (Fig. 3).
Moreover, microarray studies showed that the mRNA
level of PIR2 was only slightly upregulated at 30 min after
pH shift (data not shown). Consistent with the observed
high level of Pir proteins in the yeast cell wall at acidic pH,
growth at pH 3.5 was reduced dramatically in comparison
with the situation at pH 5.5, when yeast cells were deleted
in PIR1, PIR2 and PIR3 (data not shown). The question of
why Pir2p and possibly other Pir proteins are more
efficiently incorporated in the wall at acidic pHs remains
unclarified, but it might be that more b1,3-glucan acceptor
sites are available for these proteins at pH 3.5. Alterna-
tively, possible enzyme activities required for actually
connecting the Pir proteins to the b1,3-glucan network
may also be stimulated by low pH.
The acquired resistance of low pH-grown cells to lysis
by b1,3-glucanase digestion can probably also be
Fig. 6. A downshift in the external pH shift from 5.5 to 3.5 rapidlyinduces the expression of CWP1 in S. cerevisiae. Northern blotanalysis of CWP1 expression in wild-type cells of S. cerevisiae at0.5 h and 2 h after pH shift. Hybridization was performed with 32P-labelled CWP1 and ACT1 RNA probes.
Fig. 5. Incorporation of Cwp1p in the cell wall at low pH isdependent on a functional HOG1 gene. Western analysis of (A)b1,6-glucanase-extracted cell wall proteins, (B) b1,6-glucanase-resistant, alkali-extracted cell wall proteins and (C) cell wall proteinsextracted directly by alkali treatment of FY834 and hog1D cell wallsusing anti-Cwp1p antiserum. Strains were grown at pH 3.5. Alllanes were loaded with the equivalent of 50 mg of isolated SDS-extracted cell walls. Molecular weight markers are as indicated.
474 J. C. Kapteyn et al.
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 39, 469±479

attributed to the observed rapid induction of expression of
CWP1 (Table 2) and the low pH-induced changes in the
relative frequencies of methods of Cwp1p anchoring
(Fig. 3). At extracellular pH values of 5.5 and higher,
most Cwp1p was extractable by b1,6-glucanase digestion
only (Fig. 3). However, under more acidic conditions, an
increasing amount of Cwp1p was immobilized in the wall,
like the Pir-CWPs, through an alkali-labile linkage to b1,3-
glucan (Figs 3 and 7C), which represents a novel type of
post-translational modification for GPI-CWPs. In addition,
at pH 3.5 and 4.5, an increased amount of Cwp1p was
found to be double-anchored through an alkali-resistant
linkage to b1,6-glucan and this novel alkali-sensitive bond
to b1,3-glucan (Figs 3 and 7D). Deletion of the C-terminal
18 amino acids did not lead to disappearance of the alkali-
extractable forms of Cwp1p (Fig. 4), excluding the
involvement of the GPI anchor in the alkali-sensitive
incorporation of Cwp1p. We are currently investigating
whether the alkali-sensitive method of anchoring of
Cwp1p is a common incorporation mechanism for other
GPI-CWPs.
Notably, chitin deposition did not seem to play a role in
the observed b1,3-glucanase resistance of yeast cells
induced by low extracellular pH (Table 1). At pH 3.5, cell
wall chitin levels were even lower than in cells grown at
higher pH (Table 1), which might be attributed to an
increase in chitinase activity at acidic pH values, as has
been reported before (Cabib et al., 1989).
The observed alterations in cell wall architecture
induced by a pH shift from 5.5 to 3.5 were demonstrated
to be dependent on HOG1, but not on SLT2 (Figs 1B and
5). This was supported by DNA microarray experiments,
indicating that many genes that have previously been
shown to be inducible by hyperosmotic shock and to
depend on HOG1 and MNS2/4 (Rep et al., 2000) were
also induced by low pH. In addition, Hog1p has been
shown to be activated by low external pH (SchuÈ ller et al.,
1994). Consistent with the findings above, genes that are
known to be upregulated when Slt2 activity is enhanced
(Jung and Levin, 2000) were not found among those
induced by low external pH with the exception of CWP1.
Finally, the HOG1-dependent changes in the cell wall of
S. cerevisiae were not essential, as the hog1D strain did
not show any growth defect under low pH conditions (data
not shown). However, the observed low pH-induced
glucanase resistance may be vital under natural condi-
tions when yeast is growing on rotting fruits, which
represents an acidic environment with many harmful cell
wall hydrolytic enzymes. Adaptation to ambient environ-
mental pH may also be important for human fungal
pathogens, such as for example Candida albicans, that
are faced with pHs in the range of 4.5 in vaginal infections
and pHs near neutrality in systemic infections. In support
of this idea, the expression of genes that are essential for
the survival of C. albicans within a particular host niche
has been shown to be regulated by the pH of the host
niche (De Bernardis et al., 1998). This raises the question
whether the cell wall architecture of C. albicans, which
has recently been shown to resemble closely that of
S. cerevisiae (Kapteyn et al., 2000), is also changed in
response to differences in environmental pH.
Experimental procedures
Yeast strains and growth media
The yeast strains (Table 3) were grown at 308C in syntheticcomplete (SC) medium containing 0.17% (w/v) yeast nitrogen
Fig. 7. Proposed cell wall modules in S. cerevisiae. Module A isbased on data from Kapteyn et al. (1996) and KollaÂr et al. (1997).Module B is based on data by Mrsa et al. (1997) and Kapteyn et al.(1999b). Modules C and D are based on data in this paper. Notethat Pir-CWPs are linked to b1,3-glucan through an as yetuncharacterized linkage that is highly sensitive to alkali. For thesake of clarity, the linkages between chitin and b-glucans are notshown.
Table 2. Cell wall-related genes induced in S. cerevisiae FY834 by a pH shift from 5.5 to 3.5.
Gene name
Fold induction after Induced byhyperosmoticshock Description of gene product30 min 120 min
CWP1 1.5 1.2 1 Cell wall stress protein with GPI anchorHOR7 1.3 1.5 1 Cell wall protein with GPI anchor, involved in responsiveness to hyperosmolaritySPI1 1.5 1.6 1 Cell wall protein with GPI anchor related to Sed1pYGP1 1.4 1.6 1 Secretory glycoprotein, induced by nutrient depletion, related to Sps100p
The transcript levels of the genes presented were induced by at least 1.4-fold during 30 and/or 120 min after shifting the external pH from 5.5 to3.5. Information about the transcriptional induction of genes by hyperosmotic shock and the description of the gene product was retrieved from theYPD protein database (Costanzo et al., 2000). Note that the procedure used leads to a systematic underestimation of the fold induction (seeExperimental procedures).
Low pH-induced alterations in yeast cell wall architecture 475
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 39, 469±479

base without amino acids and ammonium sulphate (Difco),2% (w/v) glucose, 0.5% (w/v) casein hydrolysate, 0.5% (w/v)ammonium sulphate, 0.25% (w/v) succinate, 0.01% (w/v)uracil, 0.01 (w/v) tryptophan and 0.01% (w/v) adeninesulphate at pH 3.5, pH 4.5, pH 5.5 or pH 6.5.
RNA isolation and Northern hybridization
To study the transcriptional kinetics of CWP1 in response topH shift, RNA was isolated using the FastRNA Red kit (BIO101). RNA (1.0±2.0 mg per lane) was separated on aformaldehyde±formamide-containing agarose gel, trans-ferred to a Hybond-XL membrane (Amersham) by capillaryblotting with 10 � SSC and cross-linked to this membrane byUV radiation and baking for 2 h at 808C. Northern hybridiza-tions were performed in MicroHyb solution (ResearchGenetics) at 428C using 32P-labelled CWP1 gene fragments.The blot was washed with 2 � SSC21.0% SDS at 428C.Transcript levels of CWP1 were normalized to those of ACT1.
Microarray filter membrane hybridization
33P-labelled cDNA was prepared by hybridizing RNA (0.5±1.0 mg) prepared as described above with 2 ng of oligo(dT10220). After heat denaturation for 10 min at 708C andchilling on ice, the resulting hybrid was used as a templatefor reverse transcriptase (Superscript II; Life Technologies)to synthesize a 33P-labelled first-strand cDNA probeaccording to the Genefilters Microarray protocol (ResearchGenetics). The probe was purified over a Sephadex G-50column. Two separate microarray filters (Research Genet-ics) containing more than 98% of all yeast ORFs weresimultaneously but separately hybridized with two probes(made from RNA isolated from cells grown at pH 3.5 andpH 5.5 respectively) according to the manufacturer's proto-col. The filters were exposed for 5 days to a storagephosphor screen, and the data were collected and analysedusing STORM systems (Molecular Dynamics) and PATHWAYS
software (Research Genetics). After signal detection,analysis and stripping, each of the filters was rehybridizedwith freshly prepared probe, eventually resulting in threesuccessive hybridizations with the two probes. We observedthat hybridization profiles of different probes on the samefilter were very similar to those that were obtained bysimultaneous hybridization of these probes on separatefilters. Transcription levels are only shown for genes thatgave on average an induction of more than 1.4-fold in threehybridization experiments. It is emphasized here that theextent of transcriptional induction of genes determined by
genomic microarray analysis as described here is generallytwo- to threefold lower than that assessed by performingNorthern blot experiments (Ter Linde et al., 1999; Rep et al.,2000). We found this also to be the case for CWP1expression (data not shown). Hence, transcriptional aug-mentations by more than the seemingly low cut-off of 1.4correspond to genes whose actual transcription is enhancedby at least two- to threefold.
Isolation of cell wall proteins
Cells were harvested, washed and homogenized asdescribed previously (Montijn et al., 1994; Kapteyn et al.,1995). Subsequently, cell walls were isolated and washedextensively with 1 M NaCl. Isolated cell walls were boiledtwice in the presence of SDS, EDTA and b-mercaptoethanol(Kapteyn et al., 1996) to solubilize the non-covalently linkedCWPs and to remove any contaminants derived from thecytosol and/or plasma membrane (Mrsa et al., 1997; Kliset al., 1998). SDS-extracted cell walls were washed andtreated with recombinant Trichoderma harzianum endo-b1,6-glucanase (Bom et al., 1998) (0.8 U g21 wet weight of cellwalls) to isolate the GPI-CWPs (Kapteyn et al., 1996; 1999b).Completeness of the glucanase treatment was ensuredby performing a second b1,6-glucanase digestion. Hardlyany proteins were released upon a second b1,6-glucanasedigestion (J. C. Kapteyn, unpublished data). Each enzymaticdigestion was stopped by adding SDS at a final concentrationof 0.4% (w/v) and heating for 3 min at 1008C. Subsequently,b1,6-glucanase-treated cell walls were treated with Quanta-zyme, a recombinant b1,3-glucanase (Quantum Biotechnol-ogies), or ice-cold 30 mM NaOH for 17 h to release the b1,6-glucanase-resistant Pir-CWPs and trace amounts of Cwp1p,as described by Kapteyn et al. (1999b).
The efficacy of various hydrolytic enzymes to solubilizeCWPs was studied by extracting isolated walls from cells thathad been radiolabelled with [14C]-protein hydrolysate accord-ing to Kapteyn et al. (1996). Labelled cell walls were isolated,SDS extracted, rinsed and digested with hydrolytic enzymesas described above. Protein contents of the different fractionswere calculated as percentages of total label incorporatedinto SDS-extracted walls.
Analysis of cell wall proteins
CWPs were separated by electrophoresis using linear 2.2±20% polyacrylamide gels and electrophoretically transferredonto an Immobilon polyvinylidene difluoride (PVDF) mem-brane (Millipore) (Montijn et al., 1994). Mannosylated CWPs
Table 3. Yeast strains used in this study.
Strain Genotype Source or reference
FY834 MATa , leu2D1, ura3-52, his3D300, lys2D202, trp1D63 Winston et al. (1995)EV71 slt2D::HIS3 in FY834 E. VinkEV118 hog1D::URA3 in FY834 E. VinkYPH499 MATa, ura3-52, lys2-801, ade2-101, trp1D63, his3D200, leu2-D1 Sikorski and Hieter (1989)YHS201 cwp1D::URA3 in YPH499 Shimoi et al. (1995)YHS211 cwp1D18C in YPH499 Shimoi et al. (1995)
476 J. C. Kapteyn et al.
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 39, 469±479

were visualized by probing the membranes with peroxidase-labelled Concanavalin A (ConA; 1 mg ml21) in phosphate-buffered saline (PBS), containing 3% (w/v) bovine serumalbumin (BSA), 2.5 mM CaCl2 and 2.5 mM MnCl2 (Klis et al.,1998). Western immunoblot analyses were performed withpolyclonal antisera directed against b1,6-glucan (Montijnet al., 1994), Cwp1p (Shimoi et al., 1995) and Pir2p/Hsp150(Russo et al., 1992), as described before (Kapteyn et al.,1997; 1999b; Klis et al., 1998). As for the antisera directedagainst Cwp1p and Pir2p/Hsp150, before the blocking step,the membranes were treated for 30 min with 50 mM periodicacid and 100 mM sodium acetate (pH 4.5) to enhance thespecificity of the antisera. Binding of rabbit antibodies wasassessed with goat anti-rabbit IgG peroxidase (Pierce) at adilution of 1:10 000 in PBS25% (w/v) milk powder. The blotswere visualized with ECL Western blotting detection reagents(Amersham) according to the manufacturer's instructions.
Determination of protein content in cell walls
Isolated cell walls (about 4 mg dry weight) were extractedtwice in SDS extraction buffer for 5 min at 1008C, rinsed fivetimes with water and heated in 1 N NaOH (200 ml) at 1008Cfor 10 min. Subsequently, the alkaline extract was neutral-ized by adding 1 N HCl (200 ml) and, upon centrifugation(5 min at 14 000 g), the protein concentration in the super-natant was determined using the BCA protein assay reagents(Pierce) with BSA as reference.
Determination of chitin content in cell walls
SDS-extracted cell walls (about 4 mg dry weight) werehydrolysed in 6 N HCl (1 ml) at 1008C for 17 h. Uponevaporation at 658C, the samples were taken up in water(1 ml), and 100 ml of 1.5 N Na2CO3 in 4% acetylacetone wasadded to 100 ml of sample. The mixture was incubated at1008C for 20 min, and 0.7 ml of 96% ethanol was added.Subsequently, 100 ml of 2.7% (w/v) p-dimethylaminobenz-aldehyde in 50% (v/v) HCl±ethanol was added and, after 1 hincubation at room temperature, the absorbance at 520 nmwas measured and compared with a standard curve of0±30 mg of glucosamine (Popolo et al., 1997).
Quantazyme sensitivity assay
Quantazyme is a recombinant b1,3-glucanase (QuantumBiotechnologies). The Quantazyme sensitivity assay tomonitor structural changes in the yeast cell wall wasconducted as described by De Nobel et al. (2000). Briefly,yeast cells were resuspended in 40 mM b-mercaptoethanol,50 mM Tris-HCl, pH 7.5, to a concentration of 1.5 � 107
cells ml21 (OD600 of 1.0). After incubation for 1 h, Quanta-zyme was added to a final concentration of 0.1 U ml21, andcell lysis was determined by following the decrease inturbidity at 600 nm (OD600) over time. Similarly, sensitivityof isolated cell walls to degradation by Quantazyme wasmeasured by determining the decrease in turbidity of cell wallsuspensions with an initial OD600 of 1.0 over time. Quanta-zyme sensitivity was expressed as a percentage of the OD600
before the addition of the glucanase.
Acknowledgements
We thank Jose Ibeas (Purdue University, West Lafayette,USA) for providing the pirD strains, Hitoshi Shimoi (NationalInstitute of Brewing, Higashihiroshima-shi, Japan) for strainsYPH499, YHS201, YHS211 and the anti-Cwp1p antiserum.We are grateful to Marja Makarow (University of Helsinki,Helsinki, Finland) for sharing the anti-Pir2p/Hsp150, andIsaac Bom (Unilever Research Laboratories, Vlaardingen,The Netherlands) for donating samples of purified endo-b1,6-glucanase. Robbert Damveld is thanked for his help inmaking photo images, and Neta Dean (State University ofNew York, NY, USA) for critical reading of the manuscript.This research was supported financially by the NetherlandsTechnology Foundation (STW) and was co-ordinated by theEarth Life Sciences Foundation (ALW).
References
Bickle, M., Delley, P.A., Schmidt, A., and Hall, M.N. (1998)Cell wall integrity modulates RHO1 activity via theexchange factor ROM2. EMBO J 17: 2235±2245.
Bom, I.J., Dielbandhoesing, S.K., Harvey, K.N., Oomes,S.J.C.M., Klis, F.M., and Brul, S. (1998) A new tool forstudying the molecular architecture of the fungal cell wall:one-step purification of recombinant Trichoderma b-(1,6)-glucanase expressed in Pichia pastoris. Biochim BiophysActa 1425: 419±424.
Bourdineaud, J.-P. (2000) At acidic pH, the diminishedhypoxic expression of the SRP1/TIR1 yeast gene dependson the GPA2-cAMP and HOG pathways. Res Microbiol151: 43±52.
Cabib, E., Sburlati, A., Bowers, B., and Silverman, S.J.(1989) Chitin synthase 1, an auxiliary enzyme for chitinsynthesis in Saccharomyces cerevisiae. J Cell Biol 108:1665±1672.
Costanzo, M.C., Hogan, J.D., Cusick, M.E., Davis, B.P.,Fancher, A.M., Hodges, P.E., et al. (2000) The Yeastproteome database (YPD) and Caenorhabditis elegansproteome database (WormPD): comprehensive resourcesfor the organization and comparison of model organismprotein information. Nucleic Acids Res 28: 73±76.
Dallies, N., FrancËois, J., and Paquet, V. (1998) A new methodfor quantitative determination of polysaccharides in theyeast cell wall. Application to the cell wall defective mutantsof Saccharomyces cerevisiae. Yeast 14: 1297±1306.
De Bernardis, F., MuÈhlschlegel, F.A., Cassone, A., andFonzi, W.A. (1998) The pH of the host niche controls geneexpression in and virulence of Candida albicans. InfectImmun 66: 3317±3325.
De Nobel, H., Klis, F.M., Priem, J., Munnik, T., and Van DenEnde, H. (1990) The glucanase-soluble mannoproteinslimit cell wall porosity in Saccharomyces cerevisiae. Yeast6: 491±499.
De Nobel, H., Ruiz, C., Martin, H., Morris, W., Brul, S.,Molina, M., et al. (2000) Cell wall perturbation in yeastresults in dual phosphorylation of the Slt2/Mpk1 MAPkinase and in an Slt2-mediated increase in FKS2-lacZexpression, glucanase resistance and thermotolerance.Microbiology 146: 2121±2131.
Destruelle, M., Holzer, H., and Klionsky, D.J. (1994)
Low pH-induced alterations in yeast cell wall architecture 477
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 39, 469±479

Identification and characterization of a novel yeast gene:the YGP1 gene product is a highly glycosylated secretedprotein that is synthesized in response to nutrient limitation.Mol Cell Biol 14: 2740±2754.
Garcia-Rodriguez, L.J., Trilla, J.A., Castro, C., Valdivieso,M.H., Duran, A., and Roncero, C. (2000a) Characterizationof the chitin biosynthesis process as a compensatorymechanism in the fks1 mutant of Saccharomyces cerevi-siae. FEBS Lett 478: 84±88.
Garcia-Rodriguez, L.J., Duran, A., and Roncero, C. (2000b)Calcofluor antifungal action depends on chitin and afunctional high-osmolarity glycerol response (HOG) path-way: evidence for a physiological role of the Saccharo-myces cerevisiae HOG pathway under non-inducingconditions. J Bacteriol 182: 2428±2437.
Hirayama, T., Maeda, T., Saito, H., and Shinozaki, K. (1995)Cloning and characterization of seven cDNAs for hyper-osmolarity-responsive (HOR) genes of Saccharomycescerevisiae. Mol Gen Genet 249: 127±138.
Jiang, B., Ram, A.F.J., Sheraton, J., Klis, F.M., and Bussey,H. (1995) Regulation of cell wall b-glucan assembly: PTC1negatively affects PBS2 action in a pathway that includesmodulation of EXG1 transcription. Mol Gen Genet 248:260±269.
Jung, U.S., and Levin, D.E. (1999) Genome-wide analysis ofgene expression regulated by the yeast cell wall integritysignalling pathway. Mol Microbiol 34: 1049±1057.
Kapteyn, J.C., Montijn, R.C., Dijkgraaf, G.J.P., Van DenEnde, H., and Klis, F.M. (1995) Covalent association ofb1,3-glucan with b1,6±glucosylated mannoproteins in cellwalls of Candida albicans. J Bacteriol 177: 3788±3792.
Kapteyn, J.C., Montijn, R.C., Vink, E., De La Cruz, J., Llobell,A., Douwes, J.E., et al. (1996) Retention of Saccharo-myces cerevisiae cell wall proteins through a phosphodie-ster-linked b1,3-/b1,6±glucan heteropolymer. Glycobiology6: 337±345.
Kapteyn, J.C., Ram, A.F.J., Groos, E.M., KollaÂr, R., Montijn,R.C., Van Den Ende, H., et al. (1997) Altered extent ofcross-linking of b1,6-glucosylated mannoproteins to chitinin Saccharomyces cerevisiae mutants with reduced cellwall b1,3-glucan content. J Bacteriol 179: 6279±6284.
Kapteyn, J.C., Van Den Ende, H., and Klis, F.M. (1999a) Thecontribution of cell wall proteins to the organization of theyeast cell wall. Biochim Biophys Acta 1426: 373±383.
Kapteyn, J.C., Van Egmond, P., Sievi, E., Van Den Ende, H.,Makarow, M., and Klis, F.M. (1999b) The contribution ofthe O-glycosylated protein Pir2p/Hsp150 to the construc-tion of the yeast cell wall in wild-type cells and b1,6-glucan-deficient mutants. Mol Microbiol 31: 1835±1844.
Kapteyn, J.C., Hoyer, L.L., Hecht, J.E., MuÈ ller, W.H., Andel,A., Verkleij, A.J., et al. (2000) The cell wall architecture ofCandida albicans wild-type cells and cell wall-defectivemutants. Mol Microbiol 35: 601±611.
Ketela, T., Green, R., and Bussey, H. (1999) Saccharomycescerevisiae Mid2p is a potential cell wall stress sensorand upstream activator of the PKC1±MPK1 cell integritypathway. J Bacteriol 181: 3330±3340.
Klis, F.M., Ram, A.F.J., Montijn, R.C., Kapteyn, J.C., Caro,L.H.P., Vossen, J.H., et al. (1998) Posttranslationalmodifications of secretory proteins. Methods Microbiol 26:223±238.
KollaÂr, R., PetraÂkovaÂ, E., Ashwell, G., Robbins, P.W., andCabib, E. (1995) Architecture of the yeast cell wall: thelinkage between chitin and b1,3-glucan. J Biol Chem 270:1170±1178.
KollaÂr, R., Reinhold, B.B., PetraÂkovaÂ, E., Yeh, H.J.C.,Ashwell, G., Drgonova , J., et al. (1997) Architecture ofthe yeast cell wall: b1,6-glucan interconnects mannopro-teins, b1,3-glucan, and chitin. J Biol Chem 272: 17762±17775.
Lipke, P.N., and Ovalle, R. (1998) Cell wall architecture inyeast: new structure and new challenges. J Bacteriol 180:3735±3740.
Marchler, G., SchuÈ ller, C., Adam, G., and Ruis, H. (1993) ASaccharomyces cerevisiae UAS element controlled byprotein kinase A activates transcription in response to avariety of stress conditions. EMBO J 12: 1997±2003.
Martin, H., Rodriguez-Pachon, J.M., Ruiz, C., Nombela, C.,and Molina, M. (2000) Regulatory mechanisms for modula-tion of signaling through the cell integrity Slt2-mediatedpathway in Saccharomyces cerevisiae. J Biol Chem 275:1511±1519.
Mazur, P., Morin, N., Baginsky, W., el-Sherbeini, M., Clemas,J.A., Nielsen, J.B., and Foor, F. (1995) Differentialexpression and function of two homologous subunits ofyeast 1,3-beta-D-glucan synthase. Mol Cell Biol 15: 5671±5681.
Montijn, R.C., Van Rinsum, J., Van Schagen, F.A., and Klis,F.M. (1994) Glucomannoproteins in the cell wall ofSaccharomyces cerevisiae contain a novel type of carbo-hydrate side chain. J Biol Chem 269: 19338±19342.
Mrsa, V., Seidl, T., Gentzsch, M., and Tanner, W. (1997)Specific labelling of cell wall proteins by biotinylation.Identification of four covalently linked O-mannosylatedproteins of Saccharomyces cerevisiae. Yeast 13: 1145±1154.
Orlean, P. (1997) Biogenesis of yeast wall and surfacecomponents. In Molecular and Cellular Biology of the YeastSaccharomyces, Vol. 3, Cell Cycle and Cell Biology.Pringle, J.R., Broach, J.R., and Jones, E.W. (eds). ColdSpring Harbor, NY: Cold Spring Harbor Laboratory Press,pp. 229±362.
Ovalle, R., Lim, S.T., Holder, B., Jue, C.K., Wood Moore, C.,and Lipke, P.N. (1998) A spheroplast rate assay fordetermination of cell wall integrity in yeast. Yeast 14:1159±1166.
Pardo, M., Monteoliva, L., Pla, J., Sanchez, M., Gil, C., andNombela, C. (1999) Two-dimensional analysis of proteinssecreted by Saccharomyces cerevisiae regeneratingspheroplasts: a novel approach to study the cell wall.Yeast 15: 459±472.
Popolo, L., Vai, M., Gatti, E., Porello, S., Bonfante, P.,Balestrini, R., et al. (1993) Physiological analysis ofmutants indicates involvement of the Saccharomycescerevisiae glycosylphosphatidylinositol-anchored proteingp115 in morphogenesis and cell separation. J Bacteriol175: 1879±1885.
Popolo, L., Gilardelli, D., Bonfante, P., and Vai, M. (1997)Increase in chitin as an essential response to defects inassembly of cell wall polymers in the ggp1D mutant ofSaccharomyces cerevisiae. J Bacteriol 179: 463±469.
Puig, S., and PerezOrtin, J.E. (2000) Stress response and
478 J. C. Kapteyn et al.
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 39, 469±479

expression patterns in wine fermentations of yeast genesinduced at the diauxic shift. Yeast 16: 139±148.
Rajavel, M., Philip, B., Buehrer, B.M., Errede, B., and Levin,D.E. (1999) Mid2 is a putative sensor for cell integritysignalling in Saccharomyces cerevisiae. Mol Cell Biol 19:3969±3976.
Ram, A.F.J., Kapteyn, J.C., Montijn, R.C., Caro, L.H.P.,Douwes, J.E., Baginsky, W., et al. (1998) Loss of theplasma membrane-bound protein Gas1p in Saccharo-myces cerevisiae results in the release of b1,3-glucaninto the medium and induces a compensation mechanismto ensure cell wall integrity. J Bacteriol 180: 1418±1424.
Rep, M., Krantz, M., Thevelein, J.M., and Hohmann, S.(2000) The transcriptional response of Saccharomycescerevisiae to osmotic shock. J Biol Chem 275: 8290±8300.
Roncero, C., and Duran, A. (1985) Effect of calcofluor whiteand Congo red on fungal wall morphogenesis: in vivoactivation of chitin polymerization. J Bacteriol 170: 1950±1954.
Russo, P., Kalkinen, N., Sareneva, H., Paakkola, J., andMakarow, M. (1992) A heat shock gene from Sacchar-omyces cerevisiae encoding a secretory protein. Proc NatlAcad Sci USA 89: 3671±3675.
SchuÈ ller, G., Brewster, J.L., Alexander, M.R., Gustin, M.C.,and Ruis, H. (1994) The HOG pathway controls osmoticregulation of transcription via the stress response element(STRE) of the Saccharomyces cerevisiae CTT1 gene.EMBO J 13: 4382±4389.
Seidl, J., and Tanner, W. (1997) Characterization of two newgenes down-regulated by a-factor. Yeast 13: 809±817.
Shimoi, H., Iimura, Y., and Obata, T. (1995) Molecularcloning of CWP1: a gene encoding a Saccharomycescerevisiae cell wall protein solubilized with Rarobacterfaecitabus protease I. J Biochem 118: 302±311.
Shimoi, H., Kitagaki, H., Ohmori, H., Iimura, Y., and Ito, K.(1998) Sed1p is a major cell wall protein of Saccharomycescerevisiae in the stationary phase and is involved in lyticenzyme resistance. J Bacteriol 180: 3381±3387.
Sikorski, R.S., and Hieter, P. (1989) a system of shuttlevectors and yeast host strains designed for efficientmanipulation of DNA in Saccharomyces cerevisiae. Genet-ics 122: 19±27.
Smits, G.J., Kapteyn, J.C., Van Den Ende, H., and Klis, F.M.(1999) Cell wall dynamics in yeast. Curr Opin Microbiol 2:348±352.
Terashima, H., Yabuki, N., Arisawa, M., Hamada, K., andKitada, K. (2000) Up-regulation of genes encoding glyco-sylphosphatidylinositol (GPI)-attached proteins in responseto cell wall damage caused by disruption of FKS1 inSaccharomyces cerevisiae. Mol Gen Genet 264: 64±74.
Ter Linde, J.J.M., Liang, H., Davis, R.W., Steensma, H.Y.,Van Dijken, J.P., and Pronk, J.T. (1999) Genome-widetranscriptional analysis of aerobic and anaerobic chemo-stat cultures of Saccharomyces cerevisiae. J Bacteriol 181:7409±7413.
Turchini, A., Ferrario, L., and Popolo, L. (2000) Increase ofexternal osmolarity reduces morphogenetic defects andaccumulation of chitin in a gas1 mutant of Saccharomycescerevisiae. J Bacteriol 182: 1167±1171.
Valdivieso, M.H., Ferrario, L., Vai, M., Duran, A., and Popolo,L. (2000) Chitin synthesis in a gas1 mutant of Sacchar-omyces cerevisiae. J Bacteriol 182: 4752±4757.
Verna, J., Lodder, A., Lee, K., Vagts, A., and Ballester, R.(1997) A family of genes required for maintenance of cellwall integrity and for the stress response in Saccharo-myces cerevisiae. Proc Natl Acad Sci USA 94: 13804±13809.
Winston, F., Dollard, C., and Ricupero, S.L. (1995) Con-struction of a set of convenient Saccharomyces cerevisiaestrains that are isogenic to S288C. Yeast 11: 53±55.
Zhao, C., Jung, U.S., Garrett-Engele, P., Roe, T., Cyert,M.S., and Levin, D.E. (1998) Temperature-induced expres-sion of yeast FKS2 is under the dual control of proteinkinase C and calcineurin. Mol Cell Biol 18: 1013±1022.
Low pH-induced alterations in yeast cell wall architecture 479
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 39, 469±479