long-term ranging patterns of wild gelada
TRANSCRIPT

LONG-TERM RANGING PATTERNS OF WILD GELADA MONKEYS
(THEROPITHECUS GELADA) ON AN INTACT AFRO-ALPINE
GRASSLAND AT GUASSA, ETHIOPIA
____________________________________
A Thesis
Presented to the
Faculty of
California State University, Fullerton
____________________________________
In Partial Fulfillment
of the Requirements for the Degree
Master of Arts
in
Anthropology
____________________________________
By
Cha Moua
Thesis Committee Approval:
Associate Professor Peter J. Fashing, Chair
Associate Professor Nga Nguyen, Department of Anthropology
Associate Professor Elizabeth G. Pillsworth, Department of Anthropology
Fall, 2015

ii
ABSTRACT
Long-term studies of animal ranging ecology are critical to understanding how
animals utilize their habitat across space and time. Although gelada monkeys
(Theropithecus gelada) inhabit an unusual, high altitude habitat that presents unique
ecological challenges, no long-term studies of their ranging behavior have been
conducted. To close this gap, I investigated the daily path length (DPL), annual home
ranges (95%), and annual core areas (50%) of a band of ~220 wild gelada monkeys at
Guassa, Ethiopia, from January 2007 to December 2011 (for total of n = 785 full-day
follows). I estimated annual home ranges and core area using the fixed kernel reference
(FK REF) and smoothed cross-validation (FK SCV) bandwidths, and the minimum
convex polygon (MCP) method. Both annual home range (MCP - 2007: 5.9 km2; 2008:
8.6 km2; 2009: 9.2 km2; 2010: 11.5 km2; 2011: 11.6 km2) and core area increased over
the 5-year study period. The MCP and FK REF generated broadly consistent, though
slightly larger estimates that contained areas in which the geladas were never observed.
All three methods omitted one to 19 sleeping sites from the home range depending on the
year. Thus, neither the MCP nor fixed kernel estimators were more accurate than the
other. Similarly, mean annual DPL (± SE m) increased over the study period (2007:
2,848±57 m; 2008: 3,339±65 m; 2009: 3,272±72 m; 2010: 3,835±80 m; 2011: 4,100±86
m). In general, the geladas showed remarkable variation in daily, monthly, and annual

iii
DPL. I also investigated the effects of movement across uneven topography on DPL, and
I discuss the ecological implications of these findings. I compare the ranging behavior of
geladas at Guassa to (a) geladas at other study sites, (b) to Papio (baboon) species, (c) to
both terrestrial and arboreal primates, and (d) to grazing ungulates. The extensive inter-
annual variability in ranging patterns in this study demonstrates the importance of long-
term monitoring for wild nonhuman primates and its implications for conservation policy.

iv
TABLE OF CONTENTS
ABSTRACT ................................................................................................................... ii
LIST OF TABLES ......................................................................................................... vi
LIST OF FIGURES ....................................................................................................... vii
ACKNOWLEDGMENTS ............................................................................................. viii
Chapter
1. INTRODUCTION ................................................................................................ 1
Research in Animal Ranging Ecology .................................................................. 1
The Importance of Long-Term Ranging Studies ........................................... 4
Gelada Monkeys as a Model System ............................................................. 6
Gelada Monkeys Study Site, Guassa, Ethiopia .............................................. 8
Objectives of the Study .................................................................................. 9
2. METHODS ........................................................................................................... 11
Study site............................................................................................................... 11
The Qero System and its Future .................................................................... 12
Study Subjects ................................................................................................ 13
Data Collection and Analysis ............................................................................... 14
Daily Ranging Data ....................................................................................... 15
Ranging Analysis: Calculation of Daily Path Lengths .................................. 17
Ranging Analysis: Amending Daily Path Lengths to Account for Changes
in Altitude ................................................................................................. 18
Home Range Analysis .......................................................................................... 19
Home Range Estimator: Minimum Convex Polygon .................................... 20
Home Range Estimator: Fixed kernel ............................................................ 23
Autocorrelation: Implications on Home Range Analysis .............................. 27
Statistical Analysis ................................................................................................ 30
3. RESULTS ............................................................................................................. 32
Annual Home Range Estimates: MCP.................................................................. 32
Annual Home Range Estimates: FK REF ...................................................... 33
Annual Home Range Estimates: FK SCV ..................................................... 41

v
Comparison of Annual Home Range Across Methods ......................................... 43
Trends in Annual Home Range ..................................................................... 43
Annual Core Area: Use and Trends ............................................................... 46
Ranging Patterns: Daily, Monthly, and annual trends in DPL ............................. 49
Monthly Mean DPL ....................................................................................... 51
Annual Mean DPL ......................................................................................... 53
4. DISCUSSION ....................................................................................................... 56
Summary of Findings............................................................................................ 56
Evaluation of the MCP Method ..................................................................... 57
Evaluation of the Kernel Estimators .............................................................. 60
Implications and Suggestions for Future Research ........................................ 63
Comparison of Gelada Monkey Ranging Behavior Across Sites ......................... 67
How do the Annual Home Range Estimates of Geladas at Guassa Compare
to Those for Geladas at Other Sites?......................................................... 68
How do Geladas Utilize Their Home Range at Guassa and How Does it
Compare to That of Geladas at Other Sites? ............................................. 70
How do the DPL of Geladas at Guassa Compare to Those of Geladas at
Other Sites? .............................................................................................. 71
Comparison of Gelada Monkey Ranging Behavior Across Taxa......................... 71
Comparison of Gelada Ranging Behavior to Papio Species ......................... 72
Comparison of Gelada Ranging Behavior to Terrestrial Nonhuman Primate
Species ...................................................................................................... 77
Comparison of Gelada Ranging Behavior to Arboreal Nonhuman Primate
Species ...................................................................................................... 83
Comparison of Gelada Ranging Behavior to Terrestrial Ungulate Species .. 89
Implications of Inhabiting in a Topographically Variable Environment on
Calculations of Distance Traveled ..................................................................... 91
Ecological Implications of Movement Across Uneven Topography............. 92
Critiques of the Altitudinal Change Formula ................................................ 96
Conclusions ........................................................................................................... 97
APPENDIX: ADDING ERROR TO USER IDENTIFIED DUPLICATE PAIRS ...... 100
BIBLIOGRAPHY .......................................................................................................... 105

vi
LIST OF TABLES
Table Page
2.1 Results of Autocorrelation Analysis ....................................................................... 30
3.1 Comparison of Annual Home Range Estimates for MCP ...................................... 33
3.2 Core Areas (50%) and Annual Home Ranges (95%) Based on the FK
0.6*REF .................................................................................................................. 34
3.3 Core Areas (50%) and Annual Home Ranges (95%) Based on the FK SCV ......... 41
3.4 Monthly Mean DPL ± SE (m), Number of Full-days, and Range of DPL
for Each and all Years. ............................................................................................ 52
4.1 Comparison of Gelada Monkey Ranging Patterns Across Sites ............................. 69
4.2 DPL, Home Range, and Core Area of Papio Species............................................. 75
4.3 DPL, Home Range, and Core Area of Terrestrial and Arboreal Nonhuman
Primates................................................................................................................... 78
4.4 DPL, Home Range, and Core Area of Terrestrial Ungulate Species ...................... 86

vii
LIST OF FIGURES
Figure Page
2.1 Monthly Mean Rainfall at Guassa, Ethiopia .......................................................... 12
2.2 Monthly Mean Temperature at Guassa, Ehiopia ................................................... 13
2.3 Diagram Showing the Corrected DPL Based on a2 + b2 = c2. ................................ 19
3.1 Comparison of Annual Home Ranges From 2007 to 2008 Using the MCP
Method .................................................................................................................... 35
3.2 Comparison of Annual Home Ranges for 2007 Based on Scaling the FK REF..... 36
3.3 Comparison of Annual Home Ranges for 2008 Based on Scaling the FK REF..... 37
3.4 Comparison of Annual Home Ranges for 2009 Based on Scaling the FK REF..... 38
3.5 Comparison of Annual Home Ranges for 2010 Based on Scaling the FK REF..... 39
3.6 Comparison of Annual Home Ranges for 2011 Based on Scaling the FK REF..... 40
3.7 Comparison of Annual Home Ranges From 2007 to 2011 Using the FK SCV ..... 42
3.8 Cumulative 10-day Home Range Size Calculated Using the MCP Method
(95% Solid and 100% Dotted) .............................................................................. 45
3.9 Cumulative Annual Home Range Estimates Calculated Using the MCP
Method .................................................................................................................. 48
3.10 Comparison of the Relationship Between Time and DPL for Each and all
Range Years ......................................................................................................... 50
3.11 Comparison of Monthly Mean DPL and for all Range Years .............................. 51
3.12 Plot of Monthly Mean DPL on a Continuous Time Scale ................................... 54
3.13 Comparison of Annual Mean DPL + SE (m)........................................................ 55

viii
ACKNOWLEDGMENTS
I owe thanks and am indebted to many people who helped made this thesis
possible.
First and foremost, I would like to thank my life-long partner, the love of my life,
and my only best friend, Judy N. Vang, for her unconditional and unwavering support
and love these last five long and arduous years. Her presence and comfort were
instrumental in keeping me on the right path, and her happiness and health push me to
always do better and attain great things for the betterment of our lives. My daughter,
Julianne Dej Ntshiab Moua, though she is too young to realize, has been a constant bright
spot in my life, uplifting my spirit and resparking my resolve.
Next, I owe thanks to my parents, Vang Moua and Mai Lor, for giving me the
opportunity to receive an education, and in essence, experience the wonders of education
in their place. It is without doubt my parents’ struggles working in the fields to this day
and their individual and collective strengths to stay strong and unrelenting have changed
my siblings’ and my life for the better. I cannot appreciate them enough for all that they
have done for my siblings and me. I would like to thank my grandparents, Wa Lee Moua
and Xiong Thao, who were instrumental in raising my siblings and me during our
childhood years. I only wish they could be here still to share this moment with me.

ix
Life in Fullerton was made easier thanks to my brother, Sher, who was also
working on his Master’s at the time, spent some of his time in my car driving back and
forth between Fullerton and Long Beach just so that I could be closer to my school. I also
would like to thank my younger brother, Tao, and his girlfriend, Jennifer Lee, who
opened their home up to me every time I visited them in San Diego. Furthermore, I thank
my youngest brother, Yen Kong, and my younger sisters, Panglee, Gao Nou, Chamee, for
helping take care of our parents while we were away home for college.
Moreover, I thank Dr. John V.H. Constable, my undergraduate advisor and
mentor at Fresno State who continued to give me life lessons and guidance about
graduate school.
Last but not least, I thank the wonderful people I met during my time here at CSU
Fullerton, from my peers to my professors to the office staff, in particular Tannise
Collymore, and to my Thesis Committee advisors, Drs. Elizabeth Pillsworth, Nga
Nguyen, and Peter J. Fashing. I am especially indebted to Drs. Nguyen and Fashing, two
of the hardest working, devoted, and generous people I know. I will always remember
and be thankful for the patience, support, friendliness, and hospitality they have given me
and my family these past five years. They remained by me and were there for me
whenever I needed them. I cannot thank them enough for giving me the opportunity to
explore and expand my mind and develop as a scientist and a scholar. Thank you.

1
CHAPTER 1
INTRODUCTION
Research in Animal Ranging Ecology
Over the last half century, studies of animal ranging ecology have played an
integral role in expanding our knowledge of the behavior and ecology of numerous
species of animals, from ungulates (pronghorn antelope Antilocapra americana:
Buechner 1950; elk Cervus canadensis: Craighead et al. 1975), to birds (breeding,
feeding, and ranging ecology reviewed in Sutherland et al. 2004 and Wiens 1989), and
land mammals (giraffe Girafa camelopardalis: Dagg and Foster 1976, Leuthold and
Leuthold 1978; leopard Panthera onca: Rabinowitz and Nottingham 1986; African
elephant Loxodonta africana: Sikes 1971) including nonhuman primates (chimpanzees
Pan troglodytes: Boesch and Achermann 2000; L’Hoest’s monkeys Cercopithecus
lhoesti: Kaplin 2001; Bale monkey Chlorocebus djamdjamensis: Mekonnen et al. 2010;
mountain gorilla Gorilla beringei beringei: Vedder 1984; Watts 1998).
Being able to monitor and document an animal’s behavior and ecology over time
can clarify or reveal the role some animals have on the biological integrity of their
ecosystem. African elephants (Loxodonta africana), for example, consume or destroy
woody vegetation, which allows light to penetrate into the forest floor thereby facilitating
light-dependent plant species to establish and diversify (Field 1971; Western 1989).
Further, organisms like bumble bees, birds (Avian spp.), and (arguably) nonhuman

2
primates engage in pollination or seed dispersal, which facilitates reproductive success
and genetic diversity of plant species (Chapman et al. 1994; Dew and Wright 1998;
Howe 1977; Wallace and Trueman 1995). Lastly, predators consume prey to regulate
prey population densities (Berger et al. 2001; Bergerud et al. 1983; Mills 1984).
While these studies demonstrate that animals can play an integral part in the
success and health of their ecosystems, they also show that the relationship between
organisms and their environment is a highly complex and deeply interconnected one.
This suggests that as resources, such as food, water, and shelter, and ecological variables,
such as weather patterns, predation pressure, habitat loss, and group size vary across
space and time, we can expect animals to adjust their behaviors and movements in
accordance to these changes in order to maintain continued acquisition of the resources
required for survival and reproduction.
In response to ecological variability, for example, terrestrial ungulates (Albon and
Langvatn 1992; Lesage et al. 2000; Luccarini et al. 2006; Marra et al. 2005) and
nonhuman primates (Li et al. 2008) have been shown to migrate to or occupy temporary
(or seasonal) home ranges. Furthermore, animals may make minor or major shifts within
or outside their normal home range (Asensio et al. 2012; Donaldson and Echternacht
2005; Edwards et al. 2009; Fashing et al. 2007; Ferguson et al. 1999; Li et al. 2010), or
exploit alternative or fall back resources (Doran-Sheehy et al. 2009; Dunbar 1977; Li and
Rogers 2005; Pavelka et al. 2003) when primary resources become scarce. Alternatively,
large groups may fission into smaller factions to mitigate the effects of within-group
feeding competition while simultaneously reducing travel distance needed to search for
(more) food (Chapman and Chapman 2000; Chapman and Pavelka 2005; Dias and Strier

3
2003; Oluput et al. 1994). Indeed, studies of animal ranging ecology have provided
researchers with a valuable tool for documenting the behavioral responses animals make
in relation to changes in their surrounding environment.
In addition to information about space use and movement patterns, studies of
animal ranging ecology can also provide valuable data or uncover aspects of an animal’s
ranging ecology essential to making informed conservation decisions (e.g., Covert et al.
2008; Hervert et al. 2005; Heymann and Aquino 2010; Hillman 1988; Kaplin 2001;
Mekonnen et al. 2010; Laliberte and Ripple 2004; Rabinowitz and Nottingham 1986).
Recently, Mekonnen et al. (2010) completed the first study on the ranging and feeding
behavior of the Bale monkey (Chlorocebus djamdjamensis). They were able to
document, among other things, the monkey’s immense reliance on bamboo leaves,
despite inhabiting an area where the resource is heavily exploited by the local human
population (Mekonnen et al. 2010). Alternatively, information about an animal’s
behavioral or ranging ecology may be lacking or unclear. In these instances, researchers
may reinvestigate to gather additional data (e.g., Georgii 1980) or reevaluate the existing
literature in order to arrive at more robust conclusions about an animal’s ranging
behavior or habitat use preferences (e.g., Heymann and Aquino 2010). The Peruvian red
uakari monkey (Cacajao calvus ucayalii), for example, was once widely considered to be
apt to flooded-forest habitats. A recent review of all available data on its sightings and
whereabouts by Heymann and Aquino (2010), however, left the authors to reject this
notion and instead conclude that the monkeys show a preference for habitats mixed with
flooded-forest, terra firme (comprised of differently sized vegetation and terrain), or palm
swamps.

4
Indeed, the types of information obtained from studies of animal ranging, such as
the types of resources an animal relies on for survival (or the foods it substitutes when
conditions worsen), its habitat use patterns (over time), or its ecological role in its
environment, not only represent valuable assets concerned individuals need in order to
make informed conservation and management-related decisions, but also demonstrates
the importance of monitoring in animal populations.
The Importance of Long-Term Ranging Studies
Despite the significance of studies of animal ranging ecology, most ranging
studies (in nonhuman primates in particular) have only been carried out over several
months (e.g., Baoping et al. 2009; Doran 1997; Dunbar and Dunbar 1975; Mekonnen et
al. 2010; Zinner et al. 2002) or a single annual cycle (e.g., Albernaz and Magnusson
1999; Barton et al. 1992; Fashing 2001; Hunter 2001; Poulsen et al. 2001; Schreier 2010;
Willems et al. 2009). Though informative, some social activities, such as mating or births
(Carnegie et al. 2011; Janson and Verdolin 2005) and group size (Dias and Strier 2003;
Wieczkowski 2005), and ecological factors, such as food resources (Li and Walker 1986)
and climatic patterns (Haile 2005; Malhi and Wright 2004), may vary seasonally or occur
only during certain years, but not others. Therefore, the ability to acquire data over a long
stretch of time is important because it may lend researchers the opportunity to identify
behavior or movement trends that would otherwise be imperceptible with studies shorter
in duration.
For example, in their investigation of the longitudinal ranging patterns of muriqui
(Brachyteles arachnoides hypoxanthus) at Estação Biolόgica de Caratinga, Minas Gerais,
Brazil, across two temporally distinct study periods 15 years apart, Dias and Strier (2003)

5
reported an increase in the home range size (from 1.68 km2 to 3.09 km2) of their group of
muriquis between the study periods, which they attributed to a concomitant increase in
group size (from 23-27 to 57-63 individuals) over this same period. Similarly,
Wieczkowski (2005), in her re-examination of the ranging behavior of a group of Tana
River mangabeys in Kenya, first studied by Homewood (1976) and then later by Kinnaird
(1990), spanning more than two decades, found that home range size in their group of
mangabeys had increased over time (0.17 km2 to 0.19 km2 to 0.47 km2 in 1974, 1988-
1989, and 2000-2001, respectively). This pattern of increasing range use coincided with,
and was likely explained by the large group sizes found during each study period (36 to
17 to 50 members in 1974, 1988-1989, and 2000-2001, respectively) (Wieczkowski
2005). (In the 1960s, habitat disturbance led to a reduction in available habitat, which is
argued to explain the high density of mangabeys and their smaller range sizes in the 1970
study by Homewood [Wieczkowski 2005].) Lastly, Li et al. (2010), studying the long-
term ranging patterns of the Yunnan snub-nosed monkey (Rhinopithecus bieti) at Samage
Forest in the Baimaxueshan Nature Reserve, Yunnan, China, from 1998 to 2007, found
that annual home range size increased each year until it reached an asymptote after the
seventh year of observation, where it decreased slightly thereafter (7.67 km2 in 1998 to
18.77 km2 in 2004 to 17.14 km2 in 2007). These findings are important because they
capture the adaptive responses animals make as resources and conditions vary across
space and time, and further demonstrate the value of longitudinal monitoring in wild
nonhuman primate populations—and animals in general.

6
Gelada Monkeys as a Model System
Gelada monkeys (Theropithecus gelada) are an ideal model system with which to
employ home range estimators to investigate and uncover how these animals utilize their
home range across space and time. First and foremost, gelada monkeys live in a complex
fission-fusion social system (Kawai et al. 1983). Multiple one-male units can come
together and form a unit called the band which consists of units that are typically seen
ranging together and share a common home range (Kawai et al. 1983; Snyder-Mackler et
al. 2012). Units from different bands sometimes aggregate to form a larger unit called the
herd, or all the individuals seen traveling together at a particular time (Dunbar and
Dunbar 1975; Kawai et al. 1983; Ohsawa 1979; Snyder-Mackler et al. 2012). Not all of
the one-male units belonging to a single band are necessarily present at any given time
(Dunbar 1980; Dunbar and Dunbar 1975; Ohsawa 1979; Snyder-Mackler et al. 2012).
Therefore, herd size can fluctuate considerably across time. Secondly, gelada monkeys
live in an environment characterized by high altitude (range: 1700 – 4200 m: Dunbar
1998), cold temperatures, and rugged and mountainous topography (Ashenafi 2001;
Dunbar and Dunbar 1974, 1975; Fashing et al. 2014; Hunter 2001; Kawai 1979; Mori and
Belay 1990). We can therefore expect the physical constraints imposed by their
environment, coupled with the instability of their herd sizes, to shape the decisions these
monkeys make in terms of movement and habitat selection in both the short- and long-
term.
Despite the wealth of literature on the social system and behavior of gelada
monkeys, relatively little is known about how gelada monkeys utilize their unusual
habitat, especially in regard to home range size and daily movement patterns (e.g., Hunter

7
2001) over an extended and continuous period of time. Prior research has indicated that
gelada monkeys exhibit marked variations in both the distance they travel on a daily basis
and in their use of certain parts of the home range relative to other areas over time (Crook
1966; Dunbar and Dunbar 1974, 1975; Hunter 2001; Kawai 1979), and that such
movement patterns may be related to variations in resource availability and distribution,
band (herd) size, and weather conditions, e.g., fog, rainfall, and hail (Dunbar and Dunbar
1975; Hunter 2001; Iwamoto and Dunbar 1983; Kawai and Iwamoto 1979). Though
intriguing and informative, these findings only describe the ranging behavior of gelada
monkeys over the short-term (i.e., no more than one year of continuous observation:
Hunter 2001), and more pertinent to the objectives of this study, lack detailed
investigations into the home range size and use patterns of geladas in the long-term and
the specific analytical tools used to estimate home range (Dunbar and Dunbar 1974,
1975; Kawai 1979).
The scarcity of reports on the ranging ecology of geladas is alarming given the
number of potential challenges the species faces in the following years, including rising
global temperatures (Dunbar 1998), human encroachment and hunting pressures at
Sankaber, Gich, and Bole (Dunbar 1977; but see Beehner et al. 2008), and the potentially
tenuous status of long-standing traditional conservation bylaws at Guassa, the most
pristine of all established gelada study sites (Ashenafi 2001; Ashanfi and Leader-
Williams 2005). In combination, these challenges threaten the integrity of the remaining
gelada habitat and ultimately their long-term existence. Therefore, the need to quantify
how gelada monkeys utilize their unusual habitat, including their movement patterns and
spatial requirements, is at an all-time high. Critical questions include: (i) How large of an

8
area do gelada monkeys utilize on a year-to-year basis?; (ii) How do gelada monkeys use
their home range and how do their home range use patterns change over time?; (iii) How
far do gelada monkeys travel on a daily basis, how do their daily movements vary month-
to-month and year-to-year, and how does living in an uneven and hilly habitat influence
total daily distance traveled? Obtaining answers to these questions will undoubtedly
expand our knowledge about their short-term and long-term ranging patterns, and more
importantly, provide information essential for making informed conservation-related
decisions (Beehner et al. 2008; Cowlishaw and Dunbar 2000; Dunbar 1998).
Furthermore, the information obtained as a result of these questions can facilitate
comparisons of gelada ranging patterns to other species of nonhuman primates and
ungulates, and possibly help evaluate their utility in hypotheses about human and
nonhuman primate evolution (Jolly 1970; Jablonski 1993; Wrangham 1980; Fashing et al.
2014).
Gelada Monkey Study Site, Guassa, Ethiopia
In December 2005, Nguyen and Fashing (2009) established a new gelada monkey
study site at Guassa, an ecologically intact afro-alpine grassland in the Ethiopian
Highlands (Ashenafi 2001; Ashenafi and Leader-Williams 2005). Before this research
commenced at Guassa, the only sites where gelada monkeys had been studied were at
three more disturbed sites in the northern Ethiopian Highlands—Sankaber and Gich, both
located in the Simen Mountains, and Bole (Crook 1966; Dunbar and Dunbar 1974, 1975;
Kawai 1979)—and at one location, Arsi, in central Ethiopia south of the Rift Valley
(Mori and Belay 1990). Recently, reports by Fashing, Nguyen, and colleagues (Fashing et
al. 2010, 2011, 2014; Lee 2011; Moua et al. 2012; Nguyen and Fashing 2012; Nguyen et

9
al. 2015; Venkataraman et al. 2014, 2015) have offered a glimpse into the behavioral
ecology of a band of ~220 free-ranging gelada monkeys at this new relatively
undisturbed location. For example, geladas at Guassa eat a more varied diet than geladas
at more disturbed sites, incorporating not only graminoids (grasses and sedges), but also
forbs (herbs), invertebrates, and occasionally bird eggs into their diet (Fashing et al.
2010, 2014). Geladas at Guassa also suffer from large parasitic swellings caused by a
tapeworm (Taenia serialis) which represent a significant contributor to mortality in this
population (Nguyen et al. 2015). Ethiopian wolves (Canis simensis) also sometimes form
mixed-species associations with geladas at Guassa, but do not prey on the monkeys.
Wolves appear to be benefit from these associations in that they are more successful at
capturing rodents when among geladas than when they are hunting for rodents solitarily
(Venkataraman et al. 2015). Thus, the geladas at Guassa are clearly an interesting, and in
some ways unique, study population and are particularly ideal subjects for the study of
ecology, given the relatively undisturbed nature of their habitat.
Objectives of the Study
In an effort to fill gaps in our understanding about the ecology of geladas at
Guassa—and as a species—we present data on the ranging patterns of geladas at Guassa,
Ethiopia, studied over a five-year period from January 2007 to December 2011. First and
foremost, the primary objectives of this five-year study were to (a) assess the annual
home range size and core area use; (b) evaluate the accuracy of the minimum convex
polygon (MCP) and fixed kernel techniques for estimating home range size and core
area; (c) test the relationship between sample size and home range size in the MCP
method; (d) discuss the theoretical and practical implications of (b) and (c) for future

10
research; and (e) determine the total distance traveled daily and explore the effects of
living in an environment with uneven topography on estimates of distance traveled. Our
secondary objectives were to compare the ranging behavior of gelada monkeys at Guassa
to (f) gelada monkeys at other study sites where similar data are available; (g) to Papio
spp., terrestrial nonhuman primates (e.g., chimpanzees, patas monkeys, etc.), and both
arboreal frugivorous and folivorous nonhuman primates; and (h) to terrestrial ungulate
species (because of their similar gramnivorous diet to gelada monkeys). Lastly, we
provide a discussion the importance of longitudinal monitoring for conservation and
management purposes and suggestions for future research.
The objectives of this study will afford us the opportunity to evaluate the
effectiveness of the MCP and fixed kernel methods in estimating the home range and
core area use patterns in this band of gelada monkeys, and also provide us with the
valuable information we have been missing about the long-term ranging behavior of
gelada monkeys.

11
CHAPTER 2
METHODS
Study Site
The Guassa study area, ~111 km2 in area (Lat 10 15’ - 10 - 27’ N and Lon 39
45’ - 39 48’ E), is an unusually intact afro-alpine grassland located in the Central
Highlands of Ethiopia (Ashenafi 2001; Ashenafi and Leader-Williams 2005). The study
site rests between 3200-3600 m above sea level on the western border of the Greater Rift
Valley (Ashenafi 2001; Fashing et al. 2010). Guassa’s unique geographic location makes
the study site extremely hilly and mountainous, with steep drop offs of greater than 1 km
along the eastern edge of the study area (Ashenafi 2001). Moreover, Guassa experiences
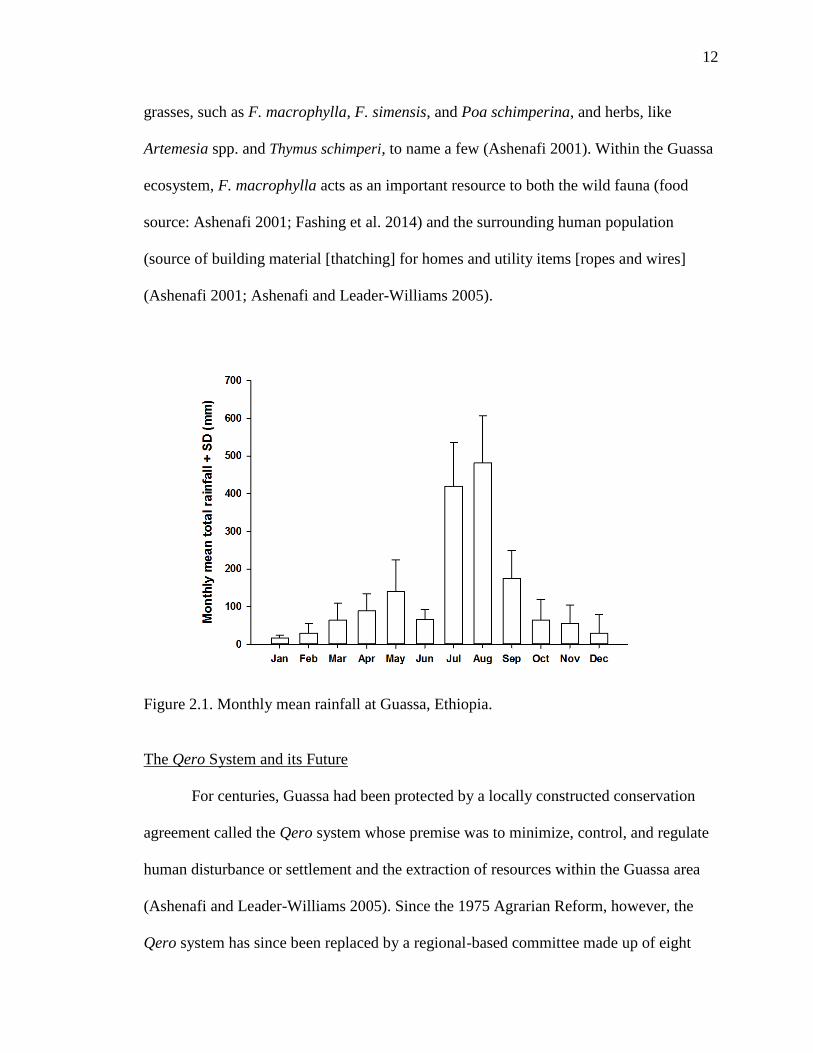
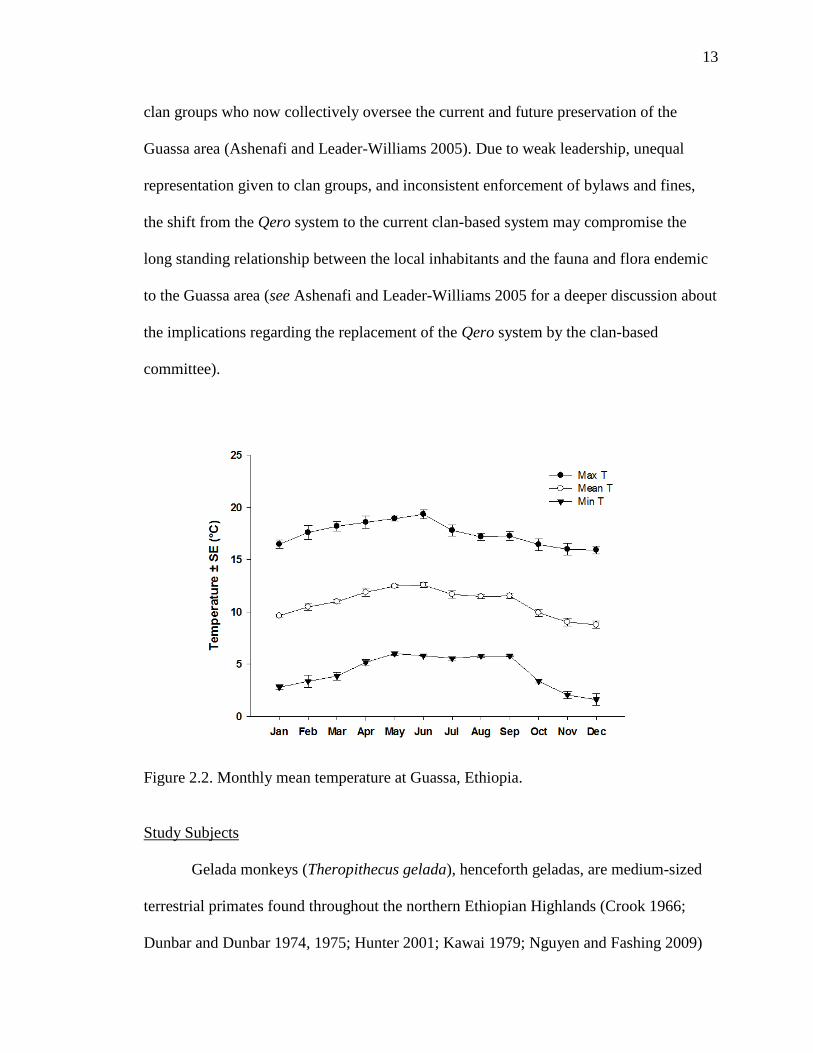
highly seasonal weather patterns. Rainfall occurs throughout the year (range of monthly
mean rainfall: 17 mm to 482 mm), but is mostly concentrated between July and August
(Figure 2.1). Monthly mean maximum temperatures typically range from 16 to 19 C,
whereas monthly mean minimum temperatures generally range from 1 to 6 C; the
overall monthly mean daily temperature ranges from 9 to 12.5 C (Figure 2.2) (Fashing
et al. 2014).
Guassa’s pristine afro-alpine grassland can be categorized into distinctive
vegetation zones depending on the composition of the plants in each area (Ashenafi
2001). The Festuca grassland (also known locally as guassa), the second largest
vegetation zone which covers ~19.9% of Guassa, is composed of various species of
12
grasses, such as F. macrophylla, F. simensis, and Poa schimperina, and herbs, like
Artemesia spp. and Thymus schimperi, to name a few (Ashenafi 2001). Within the Guassa
ecosystem, F. macrophylla acts as an important resource to both the wild fauna (food
source: Ashenafi 2001; Fashing et al. 2014) and the surrounding human population
(source of building material [thatching] for homes and utility items [ropes and wires]
(Ashenafi 2001; Ashenafi and Leader-Williams 2005).
Figure 2.1. Monthly mean rainfall at Guassa, Ethiopia.
The Qero System and its Future
For centuries, Guassa had been protected by a locally constructed conservation
agreement called the Qero system whose premise was to minimize, control, and regulate
human disturbance or settlement and the extraction of resources within the Guassa area
(Ashenafi and Leader-Williams 2005). Since the 1975 Agrarian Reform, however, the
Qero system has since been replaced by a regional-based committee made up of eight
13
clan groups who now collectively oversee the current and future preservation of the
Guassa area (Ashenafi and Leader-Williams 2005). Due to weak leadership, unequal
representation given to clan groups, and inconsistent enforcement of bylaws and fines,
the shift from the Qero system to the current clan-based system may compromise the
long standing relationship between the local inhabitants and the fauna and flora endemic
to the Guassa area (see Ashenafi and Leader-Williams 2005 for a deeper discussion about
the implications regarding the replacement of the Qero system by the clan-based
committee).
Figure 2.2. Monthly mean temperature at Guassa, Ethiopia.
Study Subjects
Gelada monkeys (Theropithecus gelada), henceforth geladas, are medium-sized
terrestrial primates found throughout the northern Ethiopian Highlands (Crook 1966;
Dunbar and Dunbar 1974, 1975; Hunter 2001; Kawai 1979; Nguyen and Fashing 2009)

14
and at one location, Arsi, in central Ethiopia south of the Rift Valley (Mori and Belay
1990; Mori et al. 1999). Geladas sleep alongside cliff edges, but conduct their daily
activities on the plateau above (Dunbar and Dunbar 1974, 1975; Kawai and Iwamoto
1979). Their diet consists mostly of grasses (Crook 1966; Dunbar and Dunbar 1975;
Iwamoto 1979; Fashing et al. 2014), however, geladas have also been observed to
consume herbs, roots, and insects (Dunbar 1977; Iwamoto 1993), especially at Guassa
where these items play an important role in the gelada diet (Fashing et al. 2010, 2014).
The foundation of the gelada monkey multi-level social system is the one-male
unit (OMU), which consists of 1-3 males, 1-9 females, juveniles, and dependent young
(Dunbar 1980; Kawai 1979; Kawai et al. 1983; Nguyen and Fashing 2009, 2012).
Alternatively, males without any alliance to an OMU may group together to form an all-
male unit (Kawai et al. 1983). Multiple OMUs that tend to range within the same
geographic location is called a band (Dunbar 1980; Kawai et al. 1983). A temporary mass
of OMUs or bands without a social or reproductive connection is called a herd (Kawai et
al. 1983). Prior research has indicated that though OMUs (with the occasional cycling of
the alpha male due to male-to-male competition) and bands may remain stable over time,
herds have been shown to be much more fluid and unpredictable in duration and number
(Dunbar and Dunbar 1975; Hunter 2001; Ohsawa 1979; P. Fashing, unpub.data).
Data Collection and Analysis
The data used for this study were collected by members of the Guassa Gelada
Research Project, headed by Peter Fashing and Nga Nguyen, and span a five-year period
from January 2007 to December 2011.

15
Daily Ranging Data
Ranging data were collected on a band of approximately 220 geladas, known as
Steelers band, grouped into 16 OMUs (Nguyen et al. 2015). Fashing and Nguyen first
started habituating Steelers band in December 2005 and, along with field assistants and
student researchers, have continued to monitor the animals’ behaviors and movements on
a near-daily basis since November 2006 (Fashing et al. 2010). Follows started at 0700-
0800 in the morning before the geladas departed their sleeping cliffs and concluded at
1730-1800 in the evening, depending on the geladas’ distance from the camp and weather
conditions. The location of the Steelers band OMU currently followed was recorded
every half-hour with a handheld GPS device (Garmin GPSMAP 62). During instances
when the researchers had to switch to a different OMU of Steelers band during the daily
follow, e.g., to carry out behavior sampling on a different OMU, the researchers selected
the next OMU of Steelers band within five to 10 meters to the OMU being followed
currently as the new follow unit. This was done to minimize the distance between the old
and new OMU and to ensure an accurate depiction of the band’s (or herd’s) movement.
All half-hour readings were recorded with an error of less than 10 meters (m), unless
striving to obtain a reading with an error of less than 10 m placed the researcher in a
precarious situation, e.g., the band was at the edge of a cliff.
Data Analysis: To be considered a valid ranging day, henceforth full-day, each
full-day had to have both a morning and an evening sleeping cliff reading and at least a
1600 (i.e., 4:00 PM) reading. There was no minimum number of half-hour readings so as
long as the aforementioned criteria were met. Based on the criteria above, I identified a
total of n = 785 full-day follows (mean = 157, range = 145 – 168) from January 2007

16
through December 2011, with an average of 20 ± 1.4 (SD) number of readings per day
(range = 14 – 24).
Sometimes the researchers were unable to remain with the herd until an evening
sleeping cliff site was chosen. In these cases, the researchers returned the next morning
before the geladas departed from their morning sleeping cliff and recorded the exact
location of the current sleeping cliff and used this sleeping cliff reading as the evening
sleeping cliff for the previous ranging day. Since the geladas had yet to venture from this
sleeping cliff, the researchers were confident that the gelada monkeys slept on this
sleeping cliff the entire night. Under this circumstance, the researchers assumed the
geladas took the shortest possible route from their last known location the prior day to
their sleeping cliff site that night. It is therefore likely that the animals’ path lengths and
daily path length (for such days) may have been slightly underestimated for some full-
day follows (e.g., Swedell 2006), though this is not expected to present any major
problems to the analysis conducted here. All GPS locations were recorded in Latitude and
Longitude (Lat and Lon), Geographic World Coordinate System WGS 84 and
subsequently uploaded to MapSource® (Garmin 2011) at the end of each month.
Data Analysis: Preparing the Data for ArcMap 10 I used Microsoft Excel 2010 to
organize and prepare the data for ranging analysis. First, I matched each GPS location
data point, identified by its waypoint number (i.e., the unique ID number indicating the
order in which the GPS point was taken) and Lat and Lon coordinate, to their respective
researcher notes. The researcher notes were entered into a palm device (Palm m500) in
the field at the time of each reading, and describe the number and sequence of the
reading, the time and date of the reading, whether the reading was a sleeping cliff or

17
regular half-hour reading, and any relevant information that may be used to assess the
validity of that particular reading. Then, I uploaded the organized Excel documents into
ArcMap 10 (ESRI 2012) under the coordinate system Geographic World Coordinate
System, WGS 84. Thereafter, I changed the Layers Data Frame properties to the
Projected Coordinate System, UTM (i.e., Universal Trans Mercator), WGS 84, Northern
Hemisphere, WGS 84 UTM Zone 37N, the coordinate zone to which Ethiopia belongs.
This series of changes transforms the Lat and Lon decimal degree coordinates into UTM
meter coordinates, making it possible to calculate the distance between consecutive half-
hour readings (for daily path length) and to estimate fixed kernel home ranges. Since the
coordinate transformation is not permanent using this procedure, I exported the data as a
shapefile, and then implemented the addxy command in Geospatial Modeling
Environment 0.7.2 (GME; Beyer 2012) to replace the original Lat and Lon coordinates
with the newly defined UTM coordinates.
Ranging Analysis: Calculation of Daily Path Lengths
I calculated all half-hour path lengths and daily path length (henceforth DPL)
using GME 0.7.2 (Beyer 2012). I define DPL as the sum of all consecutive half-hour
readings belonging to each unique full-day follow.
I identified two approaches in GME that can be used to calculate DPL, and I
utilized both approaches to validate my estimates. The first approach utilizes the
convert.pointstolines and addlength commands. The former command uses a line to
connect all of the consecutive half-hour readings belonging to a unique full-day follow
while the latter then calculates the total distance of that line (in a unit of distance
specified by the user, such as m [meters] in this case); the final value represents the DPL.

18
Alternatively, the second approach utilizes the movement.pathmetrics command. Because
this command calculates the distance of each half-hour reading, I first re-organized all
half-hour readings that belong to the same full-day, and then I obtained the sum of all the
half-hour readings to determine the DPL. Lastly, I compared the DPL estimates produced
via both of these methods and verified that both techniques produced identical estimates
(Moua unpub. data).
Ranging Analysis: Amending Daily Path Lengths to Cccount for Changes in Altitude
Once I confirmed the validity of the DPL estimates, I manually reanalyzed each
half-hour path length reading to account for the influence of changing altitude on distance
traveled. I reasoned that the extremely rugged and mountainous topography of Guassa
will cause the geladas to travel longer distances than traditionally calculated (e.g.,
Sprague 2000). To test the influence of changing altitude on distance traveled in this band
of geladas, I adapted Pythagora’s theorem for the three sides of a right triangle (i.e., a2 +
b2 = c2) (Figure 2.3). Specifically, I assumed that: (i) x1 and x2 denotes the location of
subsequent half-hour readings and a2 represents the distance, in meters, between these
two readings, squared; (ii) line segment x2x3, denoted as b2, represents the change in
altitude, in meters, squared, between half-hour readings x1 and x2; and (iii) lastly, c2 is the
sum of a2 and b2, where after solving for c2, I obtain c, the corrected path length after
taking into account change in altitude. I implemented this formula to calculate the
corrected path length for all half-hour readings. Then, I summed all corrected half-hour
path length readings belonging to each unique full-day follow to obtain the overall
corrected DPL, henceforth referred to as simply DPL (i.e., all reports of DPL henceforth
refer to the corrected estimate described here, unless otherwise stated).

19
Figure 2.3. Diagram showing the corrected DPL based on a2 + b2 = c2.
Home Range Analysis
I estimated annual home ranges using two common techniques: the minimum
convex polygon (MCP) and the fixed kernel (FK). Home ranges are defined and were
calculated based on 95% of the data, or fixes. I also calculated 100% annual home ranges
using the MCP to compare results with the 95% annual home range estimates of the MCP
method. I used the FK method to estimate core area, defined as the 50% density contour,
to identify localities of concentrated activity. Both the 50% and 95% designation for core
area (e.g., Asensio et al. 2011; Donaldson and Echternacht 2005; Fashing et al. 2007;
Loveridge et al. 2009; Rowe and Dalgarn 2010; Wartman et al. 2010; but see Powell
2000) and home range (Laver and Kelly 2008; Powell 2000; Seaman and Powell 1996;
White and Garrott 1990; Worton 1989; but see Bӧrger et al. 2006; Seaman et al. 1999)
are in line with the conventional method of home range analysis, and therefore facilitate
comparisons across studies.
Additionally, ranging data for individual range years were combined into larger
datasets to produce cumulative annual home ranges. For example, the 2007 and 2008
datasets were combined into one dataset and then analyzed to produce a cumulative

20
annual home range for 2007-2008. I repeated this process of adding subsequent datasets
for the remaining range years. In the end, I obtained a total of five datasets, four of which
contained data from subsequent years (i.e., 2007-2008, 2007-2009, 2007-2010, and 2007-
2011, except for 2007). All cumulative annual home ranges were estimated using the
MCP method only.
Lastly, I calculated cumulative home ranges at every 10 full-days. For example,
the first dataset started at full-days 1-10, then full-days 1-20, then full-days 1-30, until
full-days 1-785. As with the cumulative annual home ranges, all cumulative 10-day home
ranges were estimated using the MCP method only.
Home Range Estimator: Minimum Convex Polygon
The MCP (Mohr 1947) is a relatively old method researchers have utilized to
extrapolate home range. The MCP method uses straight lines and convex angles of less
than 180 degrees to connect the outermost points in a distribution of “fixes” (i.e.,
telemetry or geographic location data) to produce a home range in the shape of a polygon
(Anderson 1982a; Mohr 1947). Mechanically and conceptually the MCP is simple to
understand and implement, but the practical applicability of a polygon-shaped home
range has engendered a variety of issues that have severely hampered its long-standing
use as a home range tool (e.g., Borger et al. 2006; Laver and Kelly 2008; Powell 2000). A
commonly cited drawback associated with the MCP method is its tendency to
erroneously include areas the focal subject has never visited or been observed in within
the home range estimate (Andreka et al. 1999; Pebsworth et al. 2012; Powell 2000). This
leads to two additional problems: first, it does not accurately reflect the focal subject’s
movement and home range use patterns, and second, it overestimates the actual extent of

21
the focal subject’s home range (e.g., Andreka et al. 1999; Pebsworth et al. 2012).
Furthermore, outliers or unusual movements, due to their location generally being on the
periphery of the home range, will exacerbate the issues above. This is because the MCP
connects the farthest points together, and since outliers are usually farther away from
more common movements near the center of the home range, areas of space that lie
between adjacent data will be inadvertently included in the home range, inflating the
home range estimate. Moreover, the accuracy of the MCP has been tied to sample size
such that the larger the sample size, the larger (and more accurate) the home range
estimate (Bekoff and Mech 1984; Boyle et al. 2009; Girard et al. 2002; Jennrich and
Turner 1969; Schoener 1981; Seaman and Powell 1996). Lastly, the MCP fails to
produce any meaningful conclusions about trends in the focal subject’s activity inside the
home range. This inability to assess space use patterns within the home range is
significant considering the question of how an animal utilizes its home range is equally, if
not arguably more, important to how large the home range is.
Despite the aforementioned limitations of the MCP method, I constructed MCP
home ranges using both 95% and 100% of the data points in Home Range Tools, version
1.1 for ArcGIS 9.3 (Rodgers et al. 2007). I calculated 95% MCP home ranges using the
“Fixed Mean” default option in HRT. I note sample size where appropriate. The scientific
community has generally chosen to construct MCP home ranges using only a percentage
of the data points, usually 95% (Anderson 1982; Powell 2000; Powell et al. 1997),
because the MC method is highly susceptible to outliers (Andreka et al. 1982; Bekoff and
Mech 1984; Börger et al. 2006; Pebsworth et al. 2012; Powell 2000). However, several
authors (e.g., Kernohan et al. 2001; White and Garrott 1990) have argued there is no

22
biological support for the removal of the top 5% of the data, because it can result in the
loss of valuable data, e.g., removal of sleeping cliff sites from the (95%) home range
estimate (Pebsworth et al. 2012). Rodgers et al. (2007) advise that researchers should use
the “Remove X/Y Duplicates" command to remove all duplicate data points prior to
home range analysis because calculating the distances between duplicate data can result
in a “division by zero” error that can lead to a software crash. I was reluctant to
implement this command to remove duplicate data for several reasons: geladas at Guassa
routinely reuse sleeping cliffs, and they often remain immobile during periods of extreme
weather conditions, such as hail, rainfall, and thick fog (Dunbar 1977; Dunbar and
Dunbar 1975; Hunter 2001; Kawai and Iwamoto 1979; this study). Indeed, both of these
behaviors often result in duplicate or clumping of data points because the geladas are in
the same location for an extended period of time. Third, I was unsure of the ramifications
that removing duplicate data would have on the overall home range, both in terms of the
home range area estimate and possible biological interpretations (e.g., Blundell et al.
2001; de Solla et al. 1999). To determine whether or not removing the duplicate data
would have any effect on the home range estimate, I calculated home ranges with (i.e.,
the original datasets) and without duplicate data points using the “Remove X/Y
Duplicates” command in HRT. I compared the results and found that 95% MCP home
ranges constructed without duplicate points were (1-3%) larger than those constructed
with duplicate points for four of the five years (Moua, unpub. data). Further, ArcGIS 9.3
did not force quit or malfunction when the MCP command was used to calculate annual
home ranges with duplicate points in the dataset. Based on the findings above, I decided
to estimate all home ranges using data from the original datasets.

23
Though the MCP command successfully calculated all the annual and cumulative
annual home ranges from the original datasets, I later discovered that the MCP command
failed to produce any cumulative 10-day home range estimates for the first 100 full-days
(i.e., full-days 1-10, 1-20, 1-30, . . . 1-100) with the original datasets. In some cases,
ArcGIS 9.3 unexpectedly shut down without warning. This experience is indicative of the
software crash Rodgers et al. (2007) warned that can occur because of duplicate data in
the dataset. It appears that the effects of duplicate data on the home range analysis of the
MCP method in HRT is much more pronounced in smaller sample sizes than larger (since
no similar issues occurred with the larger datasets). Since I was unable to calculate 10-
day cumulative home ranges for the first 100 days of the study using the original data, I
re-analyzed all annual, cumulative annual, and cumulative 10-day home range estimates
without any duplicate data (to ensure all estimates of home range were calculated from
the same data using the MCP method). Therefore, all reports of MCP annual home range,
cumulative annual home range, and cumulative 10-day home range estimates have been
derived from the datasets containing no duplicate data. Lastly, as I previously determined
that 95% home ranges calculated without duplicate data were slightly larger than those
calculated from the original data, I acknowledge that the 95% home range estimates
reported in this study could be slightly overestimated, though a negligible difference.
Further, I acknowledge that MCP and fixed kernel annual home ranges will be calculated
using different sample sizes, however, I anticipate the results to be negligible (above).
Home Range Estimator: Fixed Kernel
Unlike the MCP method, kernel estimators can provide information about how an
animal utilizes its home range, in addition to several other features that make kernel

24
estimators currently the most preferred home range tool (Borger et al. 2006; Gitzen et al.
2006; Laver and Kelly 2008; Nilsen et al. 2009; Powell 2000; Seaman and Powell 1996).
Kernel estimators construct a home range, called a kernel or density estimate, based on
the relative density of points in a utilization distribution (UD)—a juxtaposition of fixes
(Silverman 1986; Worton 1989). It accomplishes this placing a fixed or an adaptive
kernel around each data point. A fixed kernel applies a constant smoothing factor (or
bandwidth), h, to the data, whereas an adaptive kernel adjusts its bandwidth relative to
the concentration of points in the regions such that more concentrated areas receive less
smoothing and vice versa (Silverman 1986; Worton 1989). Seaman and Powell (1996)
have demonstrated that the fixed kernel produces home range estimates that more closely
reflect the UD than the adaptive kernel.
Currently, the fixed kernel is the preferred home range estimator due to its ability
to generate density estimates of animal ranging behavior and produce a home range
estimate that may fit the shape of the focal subject’s distribution (as opposed to the MCP
which is confined to a polygon) (Laver and Kelly 2008; Powell 2000; Seaman and Powell
1999; Worton 1989). A density estimate, or density contour, represents the probability
value of the focal subject being in that location relative to other areas in the home range
(Worton 1989). The density estimate is extrapolated to identify areas within the home
range that are of relative importance to the focal subject, such as a core area, information
critical to uncovering how animals use their home range and for conservation-related
purposes. Lastly, kernels, unlike other home range estimators, such as the MCP, grid cell,
and ellipse, are free from issues that constrain the home range to a rigid and fixed shape,
and this characteristic allows kernels to produce a home range estimate that may captures

25
the focal subject’s fluid movements and dynamic home range use patterns (reviewed in
Powell 2000).
Effects of Bandwidth Estimator on Kernel Home Range Estimates Despite
possessing features that are ideal to any home range estimator, the accuracy and
performance of kernel estimators have been shown to be highly dependent on the
bandwidth used to assess the data (Borger et al. 2006; Gitzen et al. 2006; Powell 2000;
Seaman and Powell 1996; Worton 1989). Currently, the least-squares cross validation
bandwidth (LSCV) is the bandwidth of choice (Borger et al. 2006; Gitzen et al. 2006;
Powell 2000; Seaman and Powell 1999); the LSCV bandwidth, however, is limited in
application and prone to errors (Blundell et al. 2001; Gitzen et al. 2006; Horne and
Garton 2006).
Recently, research by Gitzen et al. (2006) and Horne and Garton (2006) found
that bandwidths such as the plug-in and solve-the-equation, and the likelihood cross-
validated bandwidths, respectively, performed similarly or better than the widely
considered LSCV bandwidth under identical experimental conditions. Horne and Garton
(2006) demonstrated, for example, that the likelihood cross-validated bandwidth
generated density contours that were relatively more accurate and indicative of the focal
subject’s home range use patterns than those estimated using the LSCV bandwidth when
sample size was ≤50. (Both the LSCV and likelihood cross-validated bandwidths
produced similar density estimates as sample size increased, indicating that sample size
has a relatively larger impact on density estimates than the choice of smoothing
parameter [Horne and Garton 2006; Seaman et al. 1999].) Indeed, these findings
corroborate the push by many researchers who advocate using multiple bandwidth

26
estimators in an effort to gauge the performance capabilities of each relative to the other
(Börger et al. 2006; Boyle et al. 2009; Powell 2000; Seaman and Powell 1996; White and
Garron 1990; Worton 1989). Based on these suggestions, I implemented FK analysis
using the reference, or ad hoc (REF); the LSCV; plug-in; and smoothed cross-validation
(SCV) bandwidth estimators. I used Home Range Tools (HRT) (Rodgers et al. 2007) to
conduct fixed kernel REF and LSCV home range analyses, whereas I used GME to
calculate fixed kernel LSCV, plug-in, and SCV home ranges (Beyer 2012). (GME does
not possess the REF option, whereas calculating the LSCV in both HRT and GME
provides comparability of results across programs.) Following the recommendation of
many researchers (Gitzen et al. 2006; Pebsworth et al. 2012; Rodgers et al. 2007; Seaman
and Powell 1996; Worton 1989), I multiplied the REF by a fixed proportion (e.g., 0.2,
0.4, 0.6, 0.8, and 1.0), also known as scaling the REF, which may circumvent its
tendency to underestimate or overestimate the home range.
Results of Preliminary Analyses of FK Kernel Estimators I conducted a series of
preliminary analyses to evaluate the performance capabilities of each bandwidth
estimator.
During the preliminary analysis phase, I discovered that HRT was unable to
produce any home range estimates using the LSCV bandwidth estimator, in which the
following error message appeared: “Warning: the LSCV function failed to minimize
between 0.5*HREF and 2.00*HREF. The bandwidth defaulted to HREF.” It appears that
when the LSCV bandwidth fails to reduce the mean integrated square error to an
appreciable level, it reverts to the REF bandwidth (Gitzen et al. 2006; Rodgers et al.
2007). Conversely, I found that GME successfully generated LSCV home ranges. I

27
compared the LSCV home ranges obtained in GME to the 1.0*REF home ranges
calculated in HRT, and found that the density contours of each were remarkably similar
(Moua unpub. data). I suspect GME was also unable to process the LSCV and
automatically reverted to the REF bandwidth, though without notifying the user about the
underlying reasons for the change. Following the unraveling of these findings, I omitted
the LSCV bandwidth estimator from this study altogether.
Both the SCV and plug-in bandwidth estimators produced home range estimates
with highly disconnected and scattered density contours (Moua unpub. data). Given the
similarity in the estimates produced by these two bandwidths, I report the findings for the
SCV bandwidth only. In sum, I report home range estimates for only the FK REF and
SCV bandwidths.
Autocorrelation: Implications on Ranging Analysis
Autocorrelation is defined as the aggregation of (location) data points that are
spaced too close in time that their association is no longer the result of random movement
(Legendre 1983; Swihart and Slade 1985a). It is generally assumed that data are
independent of one another, i.e., not autocorrelated (Legendre 1993; Swihart and Slade
1985b), because data that are autocorrelated may lead researchers to support or reject a
hypothesis without a statistically significant finding (Legendre 1993). The purported
impacts of autocorrelated data on estimates of animal ranging ecology are mixed at best.
Studies have shown, for example, that autocorrelated data generate MCP home ranges
that underestimated and did not correctly portray the focal subject’s space use patterns,
and also reduced the detail and length of travel paths (Swihart and Slade 1985b).
Conversely, numerous studies have demonstrated that eliminating autocorrelation may

28
actually diminish the quality and interpretational power of the findings (e.g., Blundell et
al. 2001; de Solla et al. 1999; Hansteen et al. 1997; Legendre 1993; Otis and White
1990). de Solla et al. (1999) found, for instance, that measurements of movement patterns
of both antler files (Protopiophila litigata) and snapping turtles (Chelydra serpentina)
were negatively affected at the expense of increasing the sampling time interval (to reach
independence of observations), such that a longer sampling interval resulted in a
reduction in the detail of the animal’s whereabouts and thus underestimated total distance
traveled. It appears that deleting data or increasing the time interval between subsequent
readings to reach independence of observations (as suggested by Swihart and Slade
1985a, b) may actually do more harm to the data analysis than intended (de Solla et al.
1999; Legendre 1993), and others have shown that autocorrelated data may actually help
interpret results (Hansteen et al. 1997). For example, in their examination of root vole
(Microtus oeconomus) ranging behavior, Hansteen et al. (1997) found that male root
voles tended to exhibit autocorrelated movement at short sampling intervals (i.e., at 30
and 60 mins). The authors posit this phenomenon may be explained by the animals
having large home ranges but not moving far enough between consecutive time intervals
to reach independence of observations (Hansteen et al. 1997) (an animal with a large
home range needs relatively more time between consecutive time intervals to distance
itself from its previous location if independence of observations is to be met: Schoener
1981). Indeed, these results suggest that, in some cases, autocorrelation may provide
researchers with added analytical and interpretational power about the behavior and
ecology of the focal subject.

29
I have described the disadvantages and advantages of autocorrelation on estimates
of animal ranging parameters, and the possible solutions to remedy autocorrelated data
(e.g., increase time interval or delete data points: Swihart and Slade 1985a, b). However,
I feel that increasing the time interval between consecutive observations or deleting data
until independence of observations is met (Swihart and Slade 1985a,b) would result in the
loss of crucial data and possibly inferential power about the movement patterns of the
geladas at Guassa. Ranging data in this study were collected at regular 30-minute
intervals throughout each full study day to insure a complete record was obtained of
gelada monkey movement patterns at Guassa. Furthermore, geladas are known to remain
immobile or inactive during periods of extreme weather and they frequently re-use
sleeping sites (Dunbar and Dunbar 1975; Hunter 2001; Kawai and Iwamoto 1979; this
study), behaviors that are likely to lead to autocorrelation (clumping of data points).
Indeed, eliminating data from the analysis for the sole purpose of reaching independence
of observations could potentially diminish the quality of the estimates (e.g., de Solla et al.
1999; Hansteen et al. 1997). I feel that this was something I did not want to risk.
Prior to fixed kernel analysis, I subjected the data to both Schoener’s Index (Schoener
1981) and Swihart and Slade’s Index (Swihart and Slade 1985b) to test for serial
autocorrelation with the option provided in HRT. The results (Table 2.1) of the
autocorrelation analysis indicate that the data are autocorrelated. Values of <1.6 or >2.4
for Schoener’s Index or >0.6 for Swihart and Slade’s Index indicate autocorrelation.
Given the discussion on the issue of autocorrelation (above), and in spite of the
autocorrelation test results (below), I opted to analyze the data without amending them to
reach independence of observations.

30
Table 2.1 Results of autocorrelation analysis
Year n Swihart and Slade Schoener’s Index
2007 2,944 0.04 2.18
2008 3,325 0.06 2.60
2009 3,420 0.06 2.33
2010 3,078 0.06 2.40
2011 3,326 0.07 2.45
Statistical Analysis
A goal of this study was to ascertain how DPL varied over the course of the year.
To investigate the relationship between time and DPL, I employed linear regression
analysis. I plotted each full-day follow (identified by its date) and its respective DPL
value on the x and y axis, respectively, of a scatterplot diagram. Once the data were
plotted, I obtained the line of best fit for the relationship using the least-squares method.
The least-squares method draws a straight line through the data such that the line is
exactly the same distance from each data point on the diagram (Salkind 2009). I then
calculated the Pearson product-moment correlation coefficient, r, for the given line of
best-fit. The Pearson correlation coefficient ranges from a value of -1 to 1 and describes
the strength and direction of the relationship (Salkin 2009).
Afterwards, I implemented one-way analysis of variance (ANOVA) to assess
whether the observed variations in the monthly mean DPL and annual mean DPLs were
significantly different within and across years. It is generally assumed in an ANOVA that
each group exhibits a similar degree of variation, also known as the homogeneity of
variances. I used Levene’s test of homogeneity of variances to investigate whether or not
the data supported this assumption. In instances where the homogeneity of variances
assumption was rejected, i.e., Levene’s statistic < 0.05, I re-tested the data using both the

31
Welch and Brown-Forsythe tests, ideal in cases in which the homogeneity of variances
assumption has been rejected (Pallant 2010).
All statistical tests were implemented using SPSS 20 (IBM 2012) and tested with
a significance level of α = 0.05, unless otherwise stated.
All figures were created using SigmaPlot 12.5.

32
CHAPTER 3
RESULTS
Annual Home Range Estimates: MCP
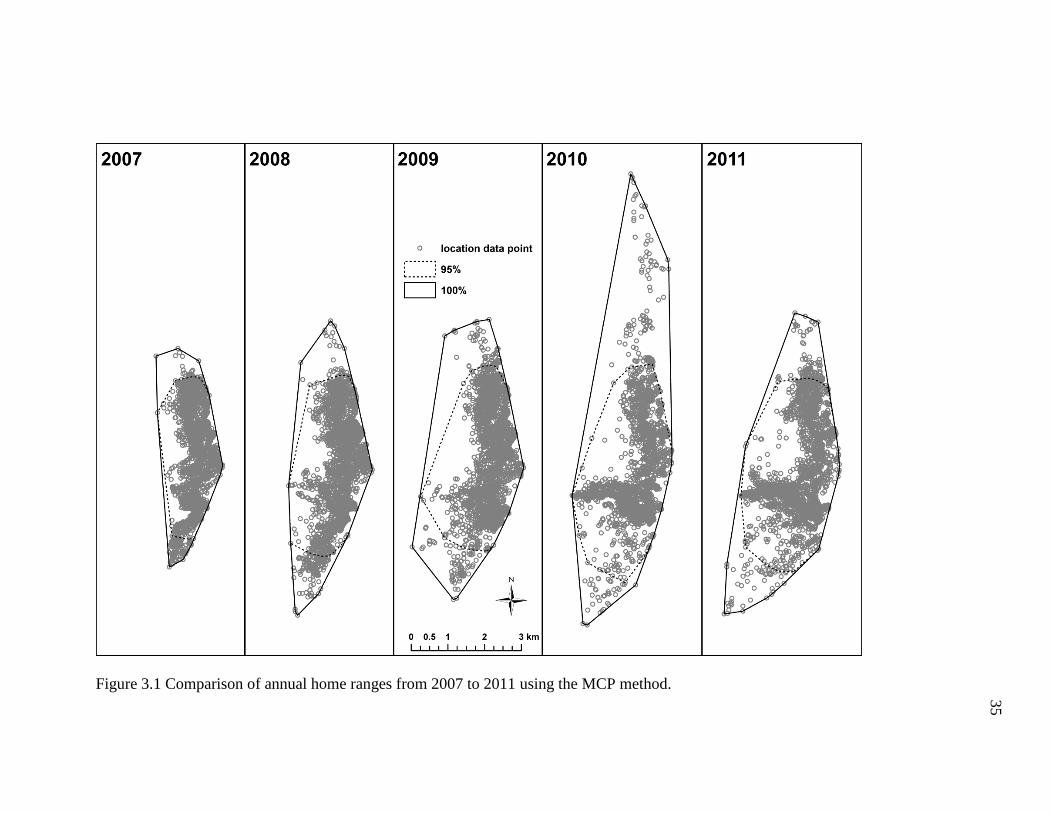
Annual home range (95%) increased in size over the five-year study period
(Figure 3.1), being smallest in 2007 (5.7 km2) and largest in 2011 (11.6 km2). The 95%
and 100% annual home range and the percentage difference between them are shown in
Table 3.1. Overall, this difference in increase in home range area from 2007 to 2011
amounts to a percent of increase of more than 50% over this time period.
All annual home range estimates from 2007 to 2011 contained areas the geladas
were not observed in (Figure 3.1). However, these areas of empty and unused areas were
relatively fewer and reduced in the 95% estimates compared to the 100% estimates,
which resulted in home ranges that were 23.4% (2007) to 48.4% (2010) smaller than their
respective 100% home range estimates (Table 3.1). Despite these findings of reduced
home range size, numerous sleeping sites (2007: 2; 2008: 1; 2009: 10; 2010: 9; 2011: 19),
which are all located along the cliff edges that border the eastern edge of the study area,
were erroneously excluded from the 95% home range estimates.

33
Table 3.1 Comparison of annual home range estimates for MCP.
Year n 95% 100% % DIFF
2007 2,611 5.9 7.7 23.4
2008 3,169 8.6 12.1 29.0
2009 3,071 9.2 14.7 37.4
2010 2,812 11.5 22.3 48.4
2011 3,148 11.6 16.7 30.5
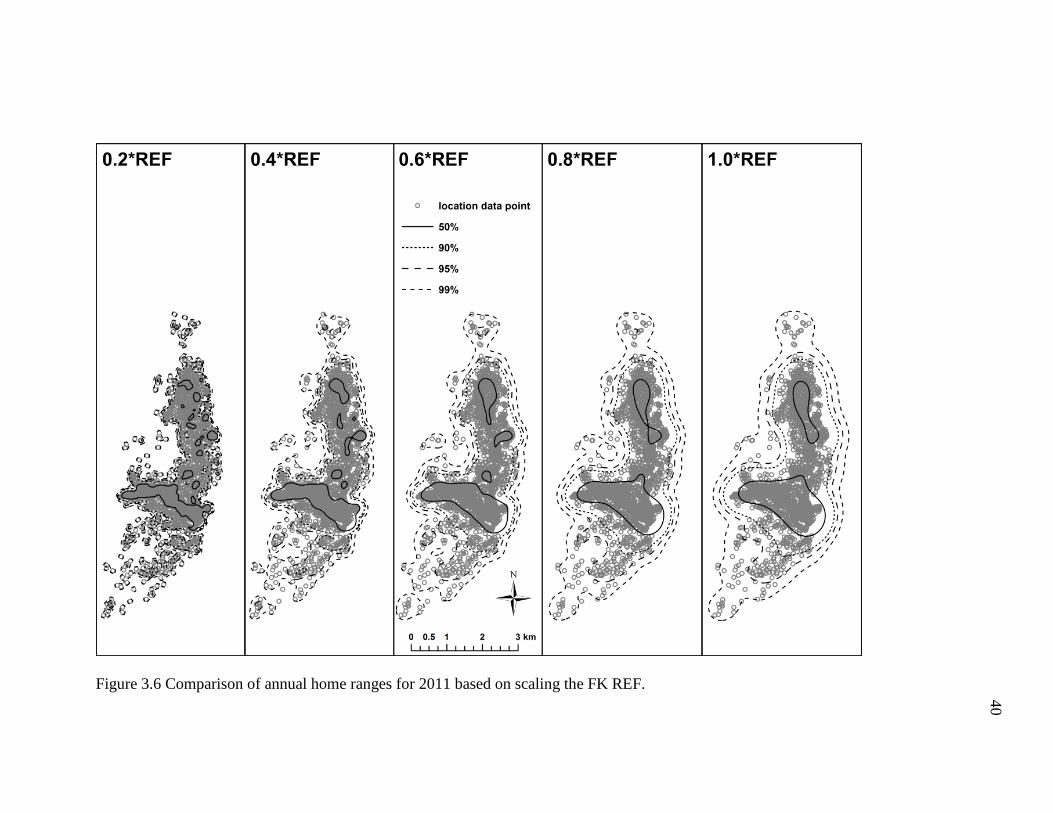
Annual Home Range Estimates: FK REF
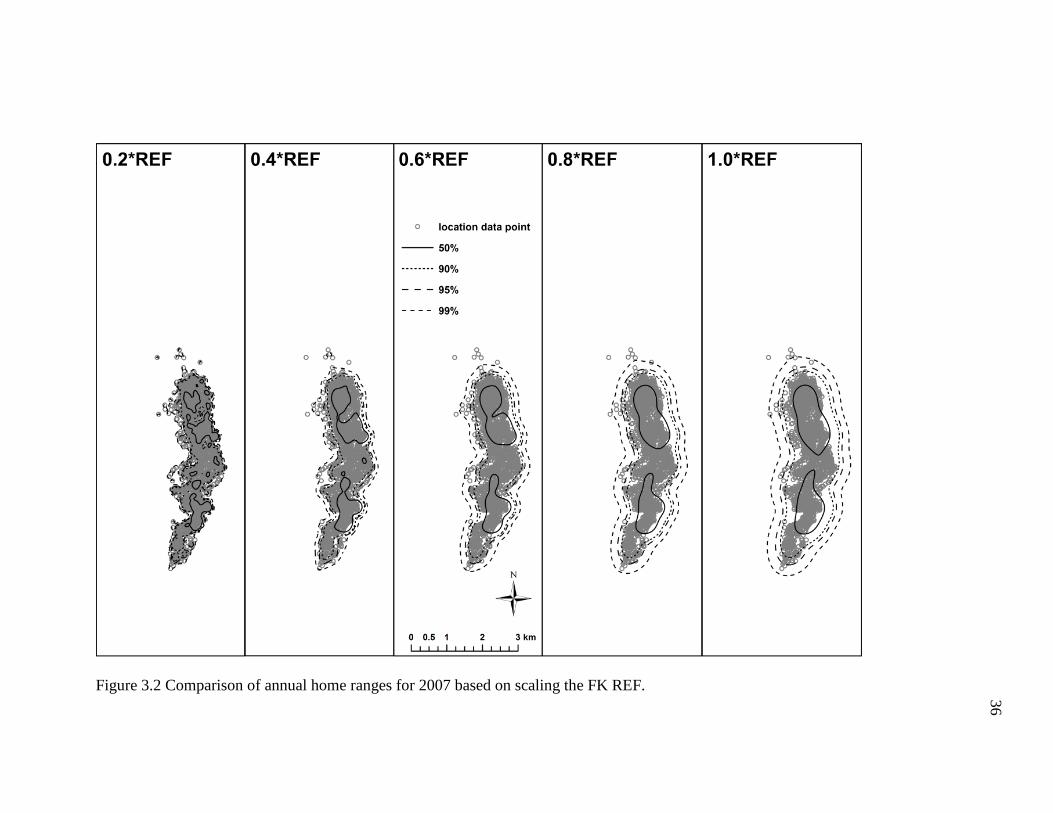
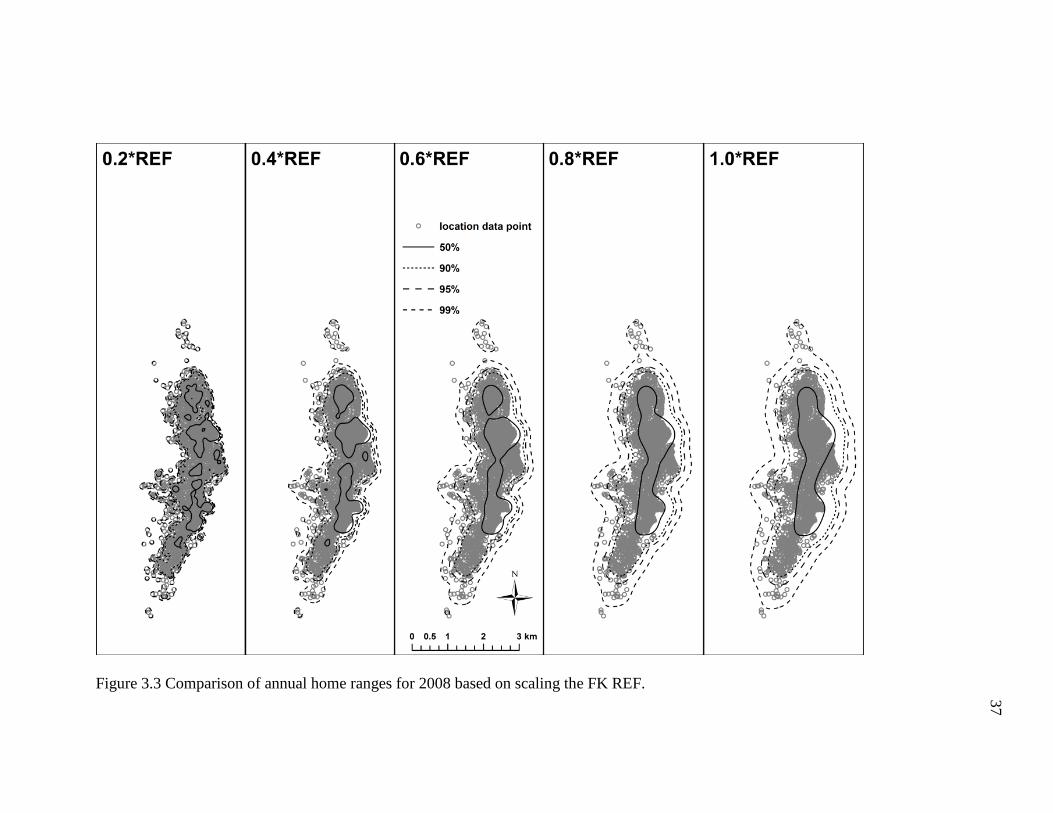
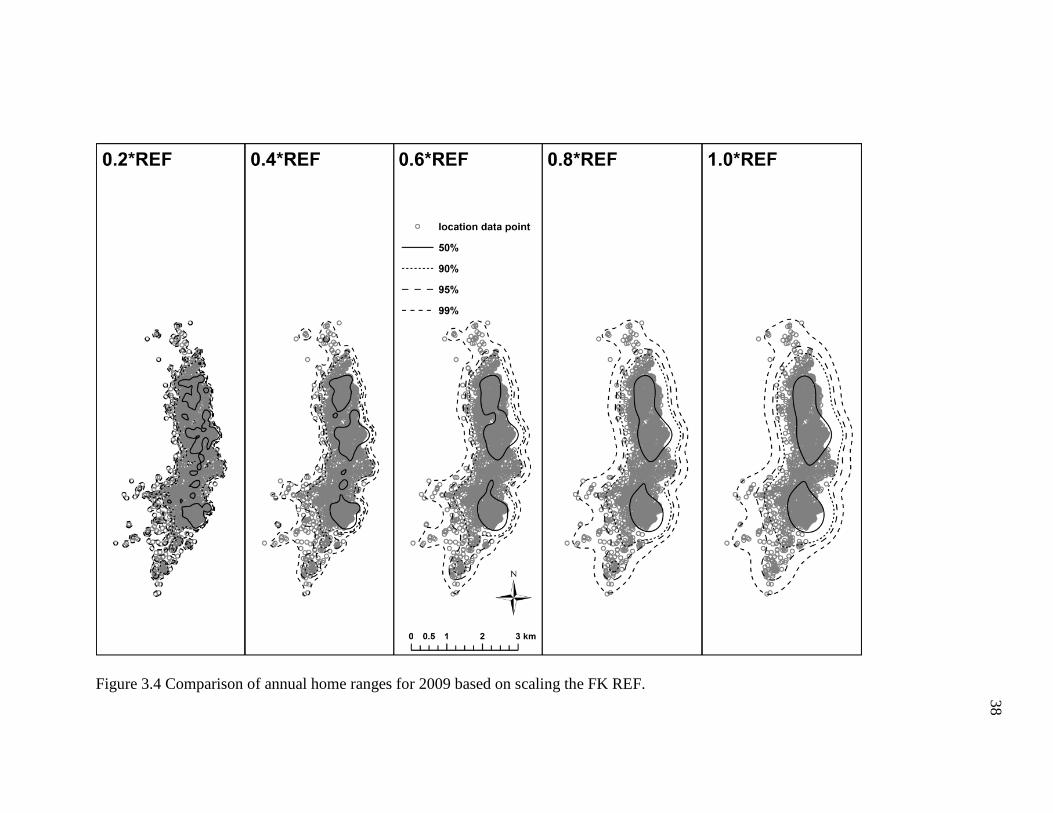
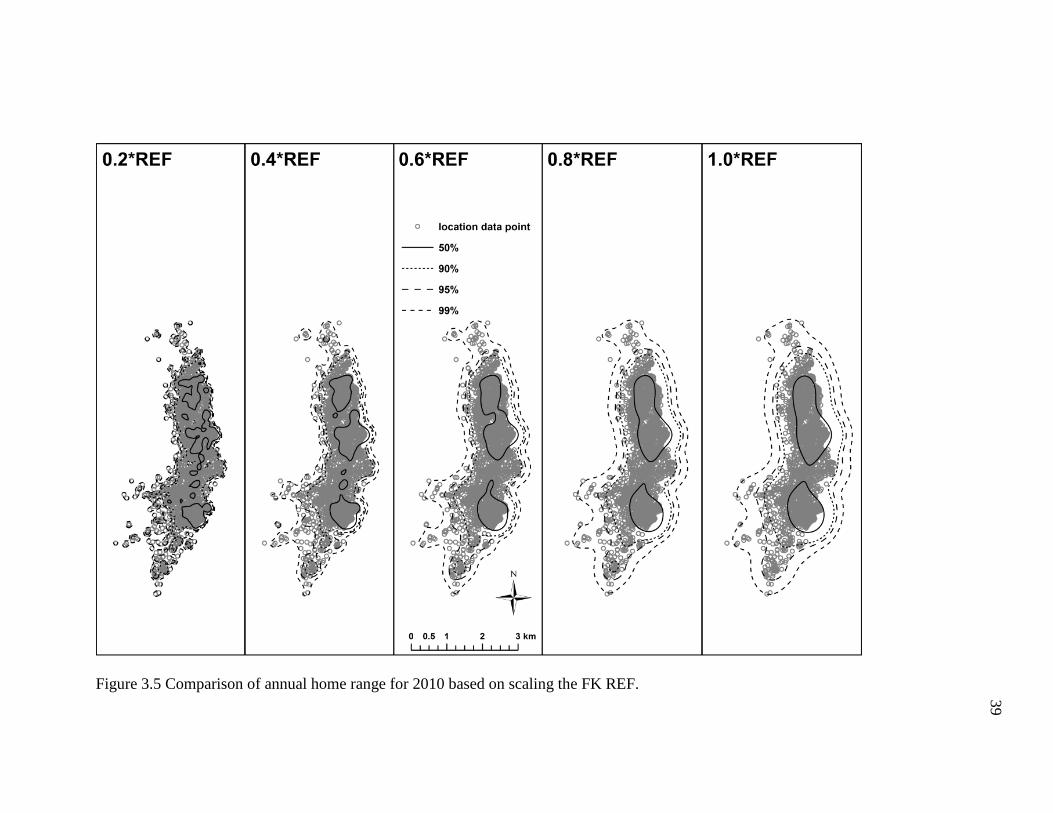
I found that multiplying the FK REF bandwidth by proportions of 0.2 to 1.0
resulted in noticeably dissimilar annual home range estimates (Figures 3.2-3.6; Table
3.2). For example, the smaller proportions of the REF (e.g. ≤0.4) produced density
contours that were fragmentary and shaped irregularly, whereas the higher proportions of
the REF (≥0.6) generated density contours that were relatively smoother and more
continuous. However, the density contours of the former incorporated copious areas the
geladas did not visit (Figure 3.2-3.6), particularly some uninhabitable areas east of the
geladas’ sleeping cliffs. Indeed, these differences in the density contours are depicted in
the area estimates for each annual home range. To briefly elaborate, the 2007 annual
home range for the 0.2*REF, 4.1 km2, was substantially smaller (47%) than the 7.7 km2
estimate obtained for the 1.0*REF (Table 3.2). In general, the number and size of the
empty, unused spaces, and the total area of each home range tended to increase as the
proportion increased (i.e., 0.2 1.0). Conversely, the density contours became more
fragmented and broken as the proportion decreased (i.e., 1.0 0.2). Based on the results
obtained here, it appears that the 0.6*REF produced annual home ranges that had
relatively continuous density contours and incorporated areas the geladas did not visit the
fewest. The 0.8*REF produced much smoother and less irregularly shaped density
34
contours, but the density contours were comparatively wider and thus tended to include
more areas the geladas never visited. As such, I conclude that the 0.6*REF produced
annual home ranges that appear to most accurately reflect the geladas’ ranging behavior.
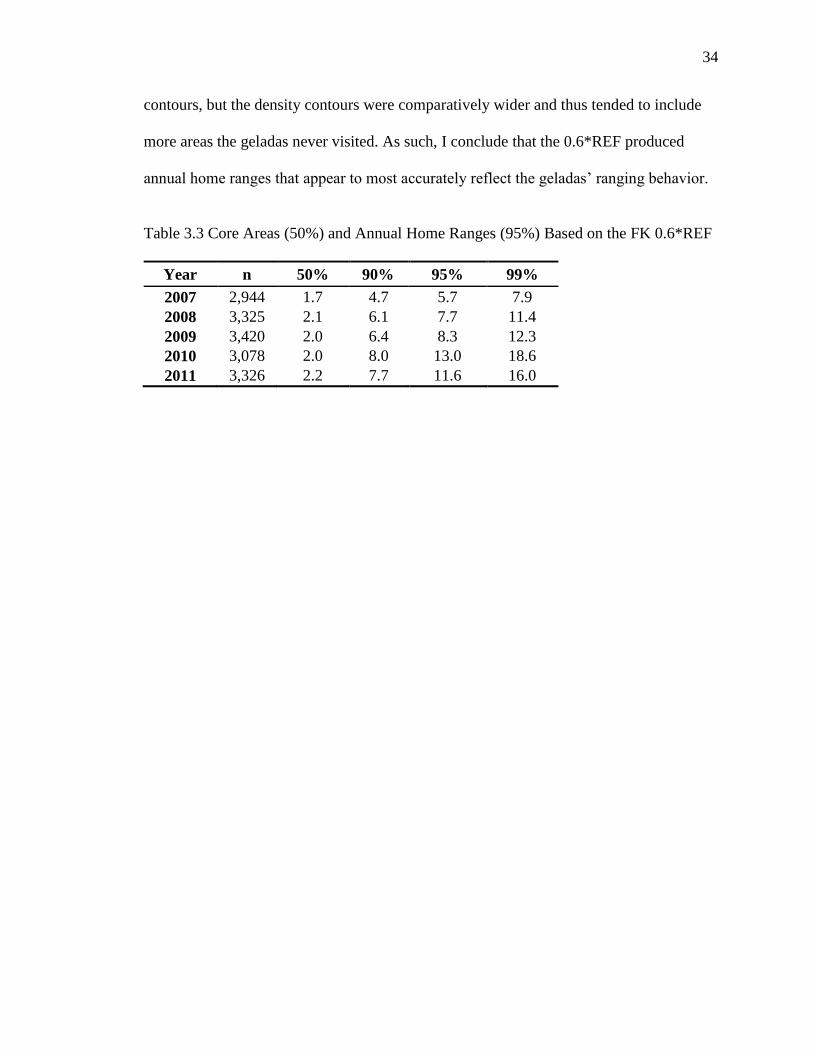
Table 3.3 Core Areas (50%) and Annual Home Ranges (95%) Based on the FK 0.6*REF
Year n 50% 90% 95% 99%
2007 2,944 1.7 4.7 5.7 7.9
2008 3,325 2.1 6.1 7.7 11.4
2009 3,420 2.0 6.4 8.3 12.3
2010 3,078 2.0 8.0 13.0 18.6
2011 3,326 2.2 7.7 11.6 16.0
35
Figure 3.1 Comparison of annual home ranges from 2007 to 2011 using the MCP method.
36
Figure 3.2 Comparison of annual home ranges for 2007 based on scaling the FK REF.
37
Figure 3.3 Comparison of annual home ranges for 2008 based on scaling the FK REF.
38
Figure 3.4 Comparison of annual home ranges for 2009 based on scaling the FK REF.
39
Figure 3.5 Comparison of annual home range for 2010 based on scaling the FK REF.
40
Figure 3.6 Comparison of annual home ranges for 2011 based on scaling the FK REF.
41
Annual Home Range Estimates: FK SCV
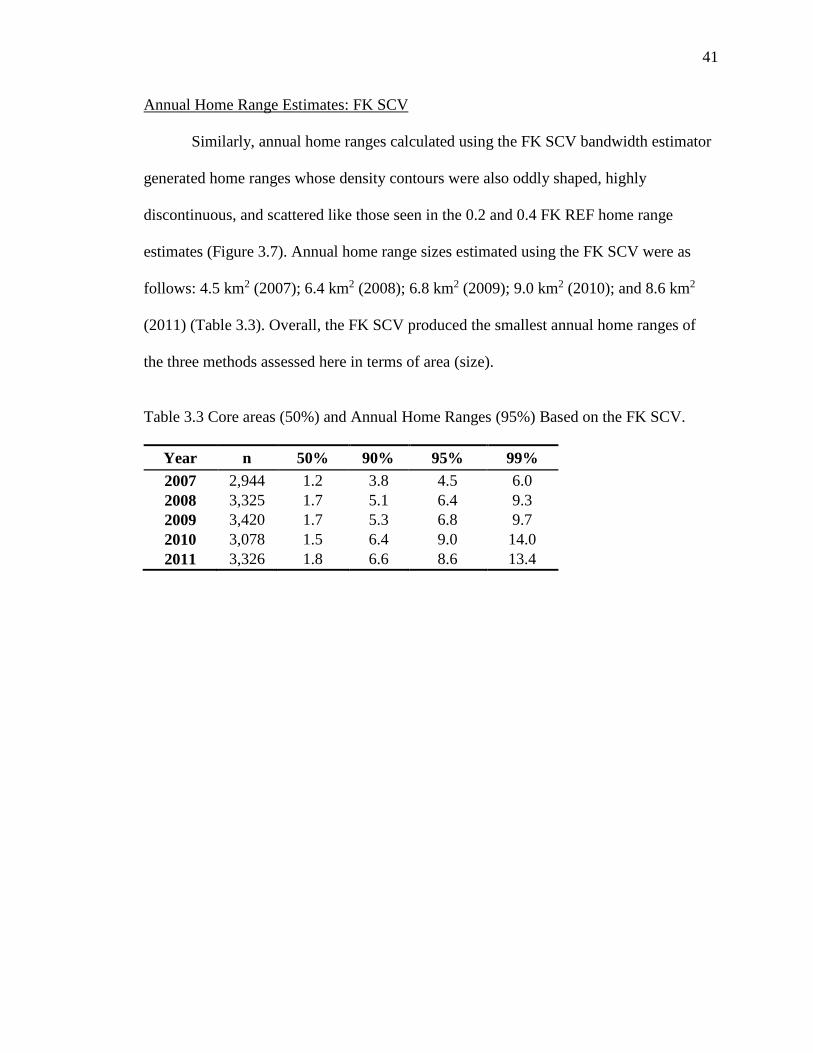
Similarly, annual home ranges calculated using the FK SCV bandwidth estimator
generated home ranges whose density contours were also oddly shaped, highly
discontinuous, and scattered like those seen in the 0.2 and 0.4 FK REF home range
estimates (Figure 3.7). Annual home range sizes estimated using the FK SCV were as
follows: 4.5 km2 (2007); 6.4 km2 (2008); 6.8 km2 (2009); 9.0 km2 (2010); and 8.6 km2
(2011) (Table 3.3). Overall, the FK SCV produced the smallest annual home ranges of
the three methods assessed here in terms of area (size).
Table 3.3 Core areas (50%) and Annual Home Ranges (95%) Based on the FK SCV.
Year n 50% 90% 95% 99%
2007 2,944 1.2 3.8 4.5 6.0
2008 3,325 1.7 5.1 6.4 9.3
2009 3,420 1.7 5.3 6.8 9.7
2010 3,078 1.5 6.4 9.0 14.0
2011 3,326 1.8 6.6 8.6 13.4
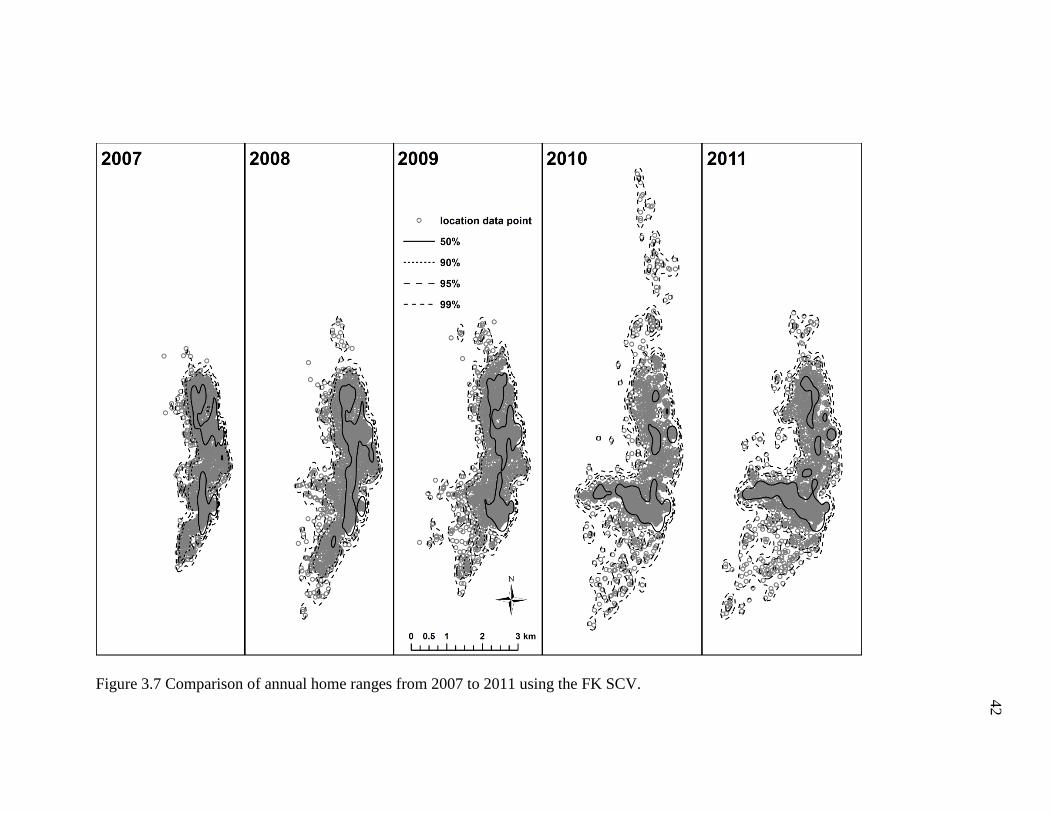
42
Figure 3.7 Comparison of annual home ranges from 2007 to 2011 using the FK SCV.

43
Comparison of Annual Home Ranges Across Methods
Despite the considerable variation in the home range estimates produced by each
method, there is evidence that illustrates some degree of commonality in the home ranges
among the MCP and both fixed kernel REF and SCV methods. To being with, the FK
SCV and some proportions of the REF method (e.g., 0.2 and 0.4) generally produced
small, disconnected, and incongruous density contours that resulted in small range size
estimates. Conversely, the ≥0.6 FK REF generated mostly contiguous and smooth density
contours, particularly at the higher density contours (e.g., >90%). Further, like the MCP
method, the ≥0.6 FK REF tended to incorporate areas never used by the animals, which
consequently resulted in inflated area estimates (Table 3.1 and 3.2). Additionally, each
home range method omitted one to 19 sleeping sites from the 95% estimates. Lastly,
despite the observed differences in the appearance of the annual home ranges estimated
by these methods, the trend of increasing annual home range size over time was evident
across all three home range estimate methods.
Trends in Annual Home Range
In general, annual home range size increased gradually over the five-year study
period: it was smallest in 2007, largest in 2010, and dropped slightly in 2011 (except for
the MCP in 2010 and 2011 where we found the reverse to be true). A closer examination
of the relationship between number of study days and home range size found that growth
in the home range was greatest during the first two to three years of the study period
(study days 460-470), but has since slowed down and appears to have reached an
asymptote after the 2010 and 2011 range years, with an occasional peak in home range

44
(e.g., study days 550-560, 660-670, and 780-785) (Figure 3.8). These findings imply an
underlying relationship between the number of study days and the size of the home range.

45
Figure 3.8 Cumulative 10-day home range size calculated using the MCP method (95% solid and 100% dotted).
Number of study days
0 50 100 150 200 250 300 350 400 450 500 550 600 650 700 750
Ho
me
ra
ng
e s
ize
(k
m2)
0
5
10
15
20
25
30

46
Cumulative Annual Home Range Estimates In general, annual home range size
increased with the addition of ranging data from subsequent years (Table 3.4). Using the
MCP method, the cumulative annual home range for 2007-2011 was slightly smaller
(11.2 km2) than the annual home range for 2011 (11.6 km2). One possible explanation for
this observation is that I calculated 95% MCP home ranges using the “Fixed Mean”
option in HRT, which obtains the mean of all the Lat and Lon coordinate pairs, then
removes the top 5% coordinate pairs from the dataset that are farthest from this mean.
Since the cumulative annual home range for 2007-2011 contained the largest sample size
and therefore most data near the center of the home range, the mean for the 2007-2011
dataset was likely smaller than the mean for the 2011 dataset, which meant that more data
were considered “farther” away and thus removed.
Annual Core Area: Use and Trends
Core area use during the five-year study indicates the geladas concentrated the
majority of their activities in roughly two to five regions within the home range (Figures
3.2-3.7): one to four in the northern and one in the southern region. The geladas’ space
use patterns did not remain static over time. According to both the FK REF and SCV, in
2010 and 2011 the core area in the southern region of the home range underwent a
westward expansion after having remained relatively stable (in size) the three years prior.
Core area size estimates varied considerably among the various proportions of the
FK REF bandwidth (Table 3.2). Roughly, annual core area for the FK REF increased in
size from 2007 to 2008, and depending on the particular proportion examined, it either
increased again or reduced in size until it finally increased to its largest size in 2011.
Similarly, annual core area for the FK SCV increased from 2007 (1.2 km2) to 2008 (1.7

47
km2), where it remained unchanged in 2009, but then it dipped back down in 2010 (1.5
km2), before it increased to its largest (combined) size in 2011 (1.8 km2). Overall, core
area use can be characterized as being smallest in the first year of study and largest in the
last year, though the number and size of the core areas changed year to year as estimated
using the FK SCV method.
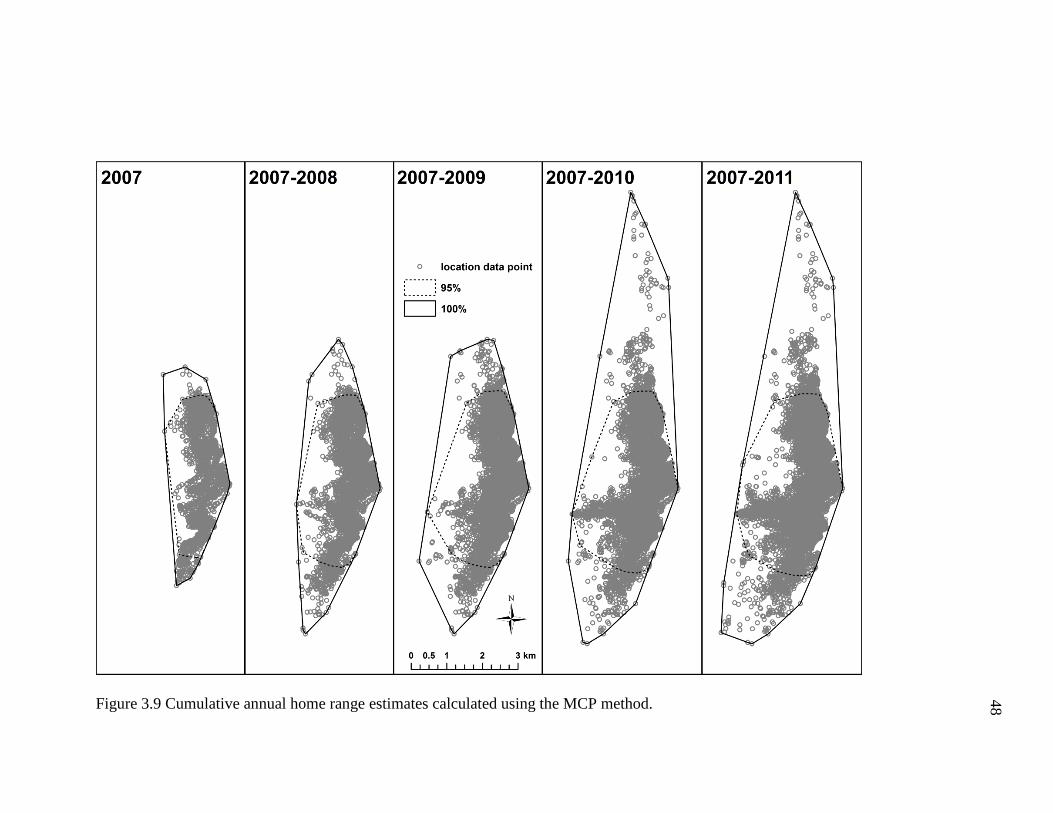
48
Figure 3.9 Cumulative annual home range estimates calculated using the MCP method.

49
Ranging Patterns: Daily, Monthly, and Annual Trends in DPL
The geladas traveled, on average, 3495 ± 1017.1 (SD) m per day (n = 785). The
day of shortest travel occurred on April 24, 2008 when the geladas ranged only 690 m,
while the day of furthest travel happened on November 11, 2011, when the geladas
traveled 7970 m.
Daily path lengths varied considerably within years and across the five-year study
period (Figure 3.10). Despite this wide variation in DPL, a modest, though statistically
significant increasing time trend was evident for three of the five years (2008: r2adj = 0.07;
2009: r2adj = 0.04; and 2011: r2
adj = 0.19; p < 0.001 for all three years), and for all five
years combined (r2adj = 0.20, p < 0.01). Conversely, no discernible pattern in DPL was
evident for the 2007 (r2adj = 0.01; p = 0.16) and 2010 range years (r2
adj = -0.01; p = 0.89).
Despite the significant increases in DPL (for three of the five range years), only some of
the observed variation in DPL (between 4-20%) could be explained by time (i.e., year),
which left a substantially high proportion (between 80-96%) unaccounted. We therefore
presume additional as yet unidentified variables are responsible for explaining the
remaining proportion of variation in DPL.
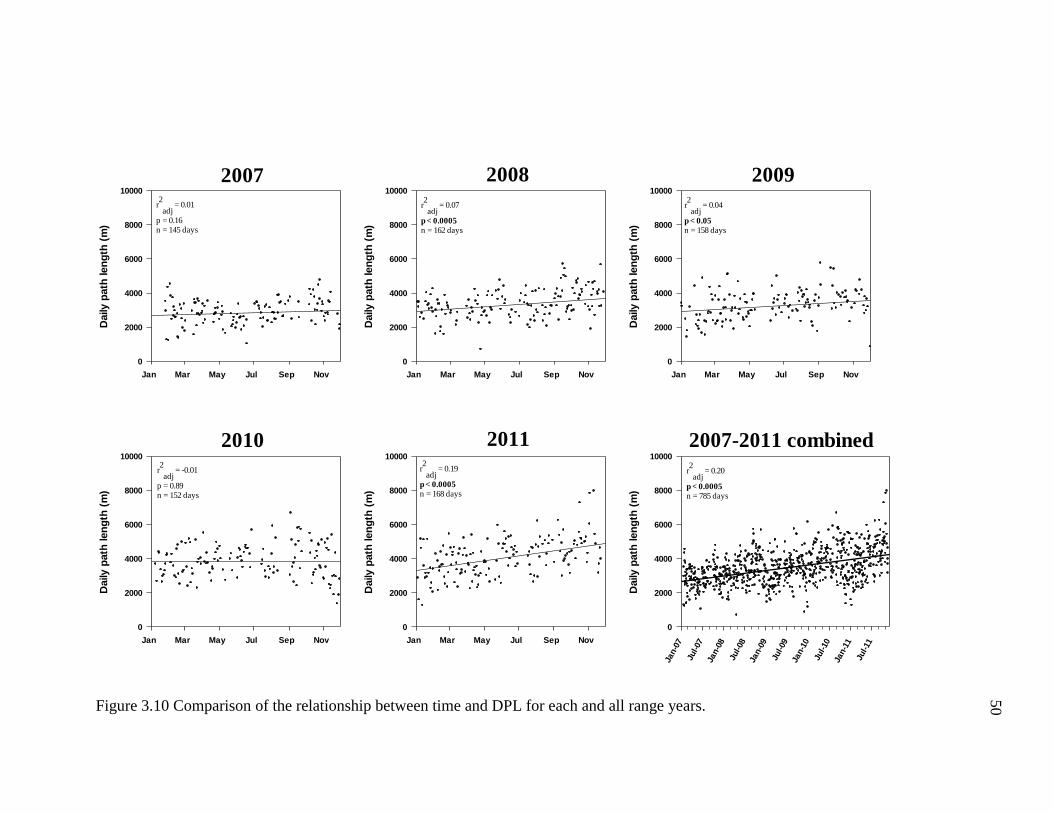
50
Figure 3.10 Comparison of the relationship between time and DPL for each and all range years.
Jan-0
7
Jul-07
Ja
n-0
8
Jul-08
Ja
n-0
9
Jul-09
Ja
n-1
0
Jul-10
Ja
n-1
1
Jul-11
Daily p
ath
len
gth
(m
)0
2000
4000
6000
8000
10000
r2
adj = 0.20
p < 0.0005
2007-2011 combined
Jan Mar May Jul Sep Nov
Daily p
ath
len
gth
(m
)
0
2000
4000
6000
8000
10000
Jan Mar May Jul Sep Nov D
aily p
ath
len
gth
(m
)0
2000
4000
6000
8000
10000
Jan Mar May Jul Sep Nov
Daily p
ath
len
gth
(m
)
0
2000
4000
6000
8000
10000
Jan Mar May Jul Sep Nov
Daily p
ath
len
gth
(m
)
0
2000
4000
6000
8000
10000
Jan Mar May Jul Sep Nov
Daily p
ath
len
gth
(m
)
0
2000
4000
6000
8000
10000
2007 2008 2009
2010 2011
r2
adj = 0.20
p < 0.0005
n = 785 days
r2
adj = 0.19
p < 0.0005
n = 168 days
r2
adj = -0.01
p = 0.89n = 152 days
r2
adj = 0.04
p < 0.05
n = 158 days
r2
adj = 0.07
p < 0.0005
n = 162 days
r2
adj = 0.01
p = 0.16n = 145 days
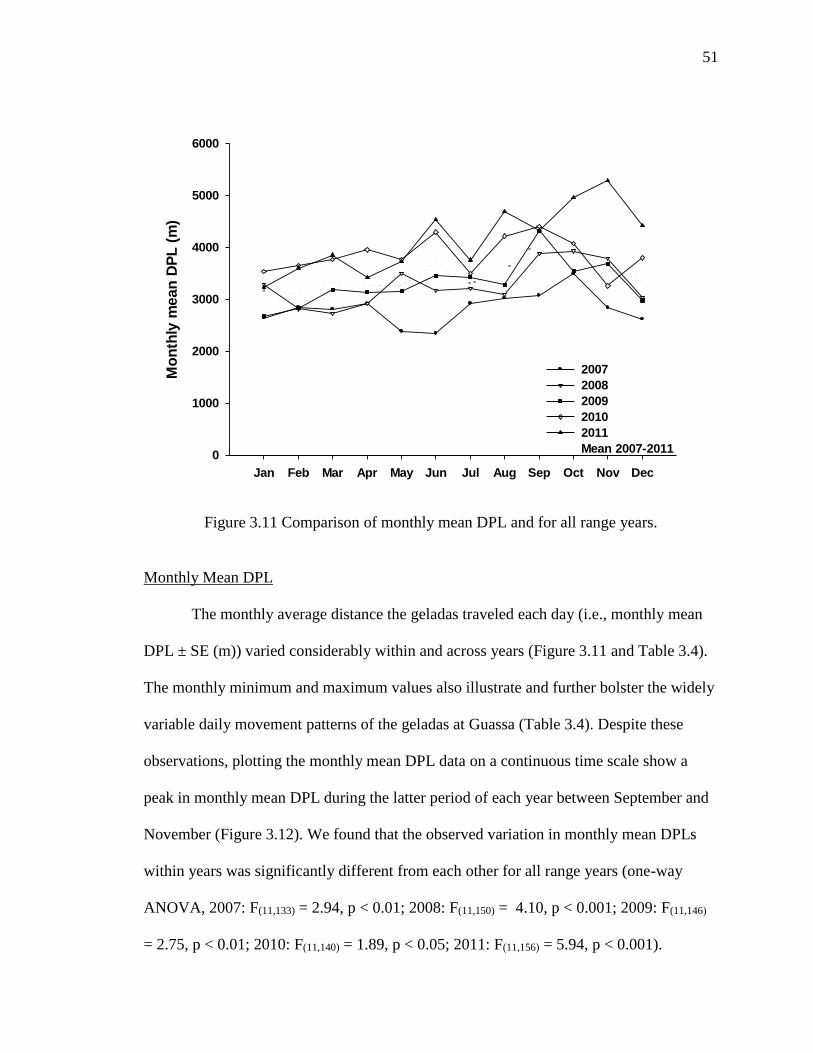
51
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Mo
nth
ly m
ean
DP
L (
m)
0
1000
2000
3000
4000
5000
6000
2007
2008
2009
2010
2011
Mean 2007-2011
Figure 3.11 Comparison of monthly mean DPL and for all range years.
Monthly Mean DPL
The monthly average distance the geladas traveled each day (i.e., monthly mean
DPL ± SE (m)) varied considerably within and across years (Figure 3.11 and Table 3.4).
The monthly minimum and maximum values also illustrate and further bolster the widely
variable daily movement patterns of the geladas at Guassa (Table 3.4). Despite these
observations, plotting the monthly mean DPL data on a continuous time scale show a
peak in monthly mean DPL during the latter period of each year between September and
November (Figure 3.12). We found that the observed variation in monthly mean DPLs
within years was significantly different from each other for all range years (one-way
ANOVA, 2007: F(11,133) = 2.94, p < 0.01; 2008: F(11,150) = 4.10, p < 0.001; 2009: F(11,146)
= 2.75, p < 0.01; 2010: F(11,140) = 1.89, p < 0.05; 2011: F(11,156) = 5.94, p < 0.001).
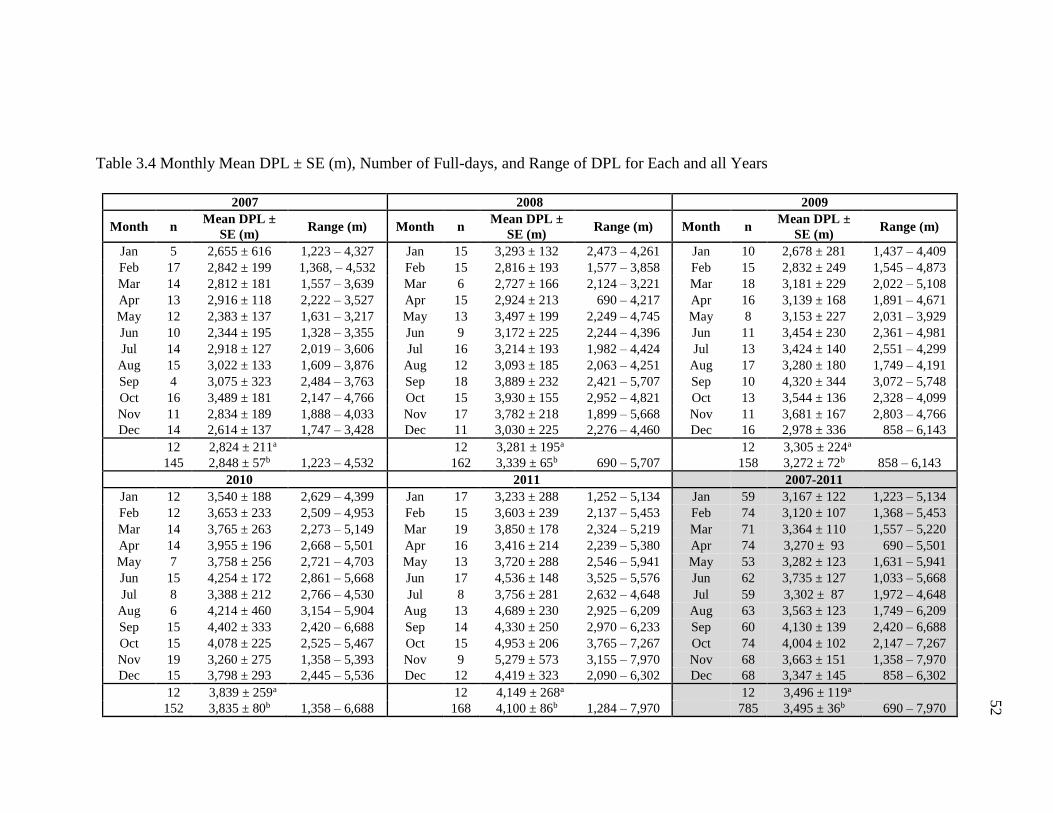
52
Table 3.4 Monthly Mean DPL ± SE (m), Number of Full-days, and Range of DPL for Each and all Years
2007 2008 2009
Month n Mean DPL ±
SE (m) Range (m) Month n
Mean DPL ±
SE (m) Range (m) Month n
Mean DPL ±
SE (m) Range (m)
Jan 5 2,655 ± 616 1,223 – 4,327 Jan 15 3,293 ± 132 2,473 – 4,261 Jan 10 2,678 ± 281 1,437 – 4,409
Feb 17 2,842 ± 199 1,368, – 4,532 Feb 15 2,816 ± 193 1,577 – 3,858 Feb 15 2,832 ± 249 1,545 – 4,873
Mar 14 2,812 ± 181 1,557 – 3,639 Mar 6 2,727 ± 166 2,124 – 3,221 Mar 18 3,181 ± 229 2,022 – 5,108
Apr 13 2,916 ± 118 2,222 – 3,527 Apr 15 2,924 ± 213 690 – 4,217 Apr 16 3,139 ± 168 1,891 – 4,671
May 12 2,383 ± 137 1,631 – 3,217 May 13 3,497 ± 199 2,249 – 4,745 May 8 3,153 ± 227 2,031 – 3,929
Jun 10 2,344 ± 195 1,328 – 3,355 Jun 9 3,172 ± 225 2,244 – 4,396 Jun 11 3,454 ± 230 2,361 – 4,981
Jul 14 2,918 ± 127 2,019 – 3,606 Jul 16 3,214 ± 193 1,982 – 4,424 Jul 13 3,424 ± 140 2,551 – 4,299
Aug 15 3,022 ± 133 1,609 – 3,876 Aug 12 3,093 ± 185 2,063 – 4,251 Aug 17 3,280 ± 180 1,749 – 4,191
Sep 4 3,075 ± 323 2,484 – 3,763 Sep 18 3,889 ± 232 2,421 – 5,707 Sep 10 4,320 ± 344 3,072 – 5,748
Oct 16 3,489 ± 181 2,147 – 4,766 Oct 15 3,930 ± 155 2,952 – 4,821 Oct 13 3,544 ± 136 2,328 – 4,099
Nov 11 2,834 ± 189 1,888 – 4,033 Nov 17 3,782 ± 218 1,899 – 5,668 Nov 11 3,681 ± 167 2,803 – 4,766
Dec 14 2,614 ± 137 1,747 – 3,428 Dec 11 3,030 ± 225 2,276 – 4,460 Dec 16 2,978 ± 336 858 – 6,143
12 2,824 ± 211a 12 3,281 ± 195a 12 3,305 ± 224a
145 2,848 ± 57b 1,223 – 4,532 162 3,339 ± 65b 690 – 5,707 158 3,272 ± 72b 858 – 6,143
2010 2011 2007-2011
Jan 12 3,540 ± 188 2,629 – 4,399 Jan 17 3,233 ± 288 1,252 – 5,134 Jan 59 3,167 ± 122 1,223 – 5,134
Feb 12 3,653 ± 233 2,509 – 4,953 Feb 15 3,603 ± 239 2,137 – 5,453 Feb 74 3,120 ± 107 1,368 – 5,453
Mar 14 3,765 ± 263 2,273 – 5,149 Mar 19 3,850 ± 178 2,324 – 5,219 Mar 71 3,364 ± 110 1,557 – 5,220
Apr 14 3,955 ± 196 2,668 – 5,501 Apr 16 3,416 ± 214 2,239 – 5,380 Apr 74 3,270 ± 93 690 – 5,501
May 7 3,758 ± 256 2,721 – 4,703 May 13 3,720 ± 288 2,546 – 5,941 May 53 3,282 ± 123 1,631 – 5,941
Jun 15 4,254 ± 172 2,861 – 5,668 Jun 17 4,536 ± 148 3,525 – 5,576 Jun 62 3,735 ± 127 1,033 – 5,668
Jul 8 3,388 ± 212 2,766 – 4,530 Jul 8 3,756 ± 281 2,632 – 4,648 Jul 59 3,302 ± 87 1,972 – 4,648
Aug 6 4,214 ± 460 3,154 – 5,904 Aug 13 4,689 ± 230 2,925 – 6,209 Aug 63 3,563 ± 123 1,749 – 6,209
Sep 15 4,402 ± 333 2,420 – 6,688 Sep 14 4,330 ± 250 2,970 – 6,233 Sep 60 4,130 ± 139 2,420 – 6,688
Oct 15 4,078 ± 225 2,525 – 5,467 Oct 15 4,953 ± 206 3,765 – 7,267 Oct 74 4,004 ± 102 2,147 – 7,267
Nov 19 3,260 ± 275 1,358 – 5,393 Nov 9 5,279 ± 573 3,155 – 7,970 Nov 68 3,663 ± 151 1,358 – 7,970
Dec 15 3,798 ± 293 2,445 – 5,536 Dec 12 4,419 ± 323 2,090 – 6,302 Dec 68 3,347 ± 145 858 – 6,302
12 3,839 ± 259a 12 4,149 ± 268a 12 3,496 ± 119a
152 3,835 ± 80b 1,358 – 6,688 168 4,100 ± 86b 1,284 – 7,970 785 3,495 ± 36b 690 – 7,970

53
Annual Mean DPL
Annual mean DPL increased significantly over the course of the five-year period
(Figure 3.13; one way ANOVA: F(4,780) = 44.1, p < 0.001). Results from Scheffe’s post-
hoc test reveal the annual mean DPL for 2007 was significantly different from the annual
mean DPL for 2008-2011 (p < 0.001 for all cases); the annual mean DPL for both 2008
and 2009 were significantly different from 2007 and 2010-2011 (p < 0.01 for all cases);
and the annual mean DPL for both 2010 and 2011 were significantly different from 2007-
2009 (p < 0.001 for all cases) (in Figure 3.6, letters that are shared among years indicate
where years are not significantly different from each other, whereas letters that differ
among years indicate where years are significantly different from each other). Despite the
significant increases in annual mean DPL, approximately only 18% of the variation in
annual mean DPL can be explained by time (i.e., year). We therefore suspect the
remaining 82% of the variation in annual mean DPL to be explained by additional though
currently unidentifiable variables (shaping DPL at the annual time scale).
Effect of Change in Altitude on Calculations of Distance Traveled
We found that accounting for changes in altitude on half-hourly distance traveled
resulted in a 0.15% (8 August 2011) to 12.2% (1 December 2009) increase in DPL (n =
785, mean = 1.67 ± 0.9% (SD)). In terms of actual additional distance traveled, these
percentages amount to an increase in DPL by 8 to 278 m (mean = 53 ± 22.4 m (SD)).
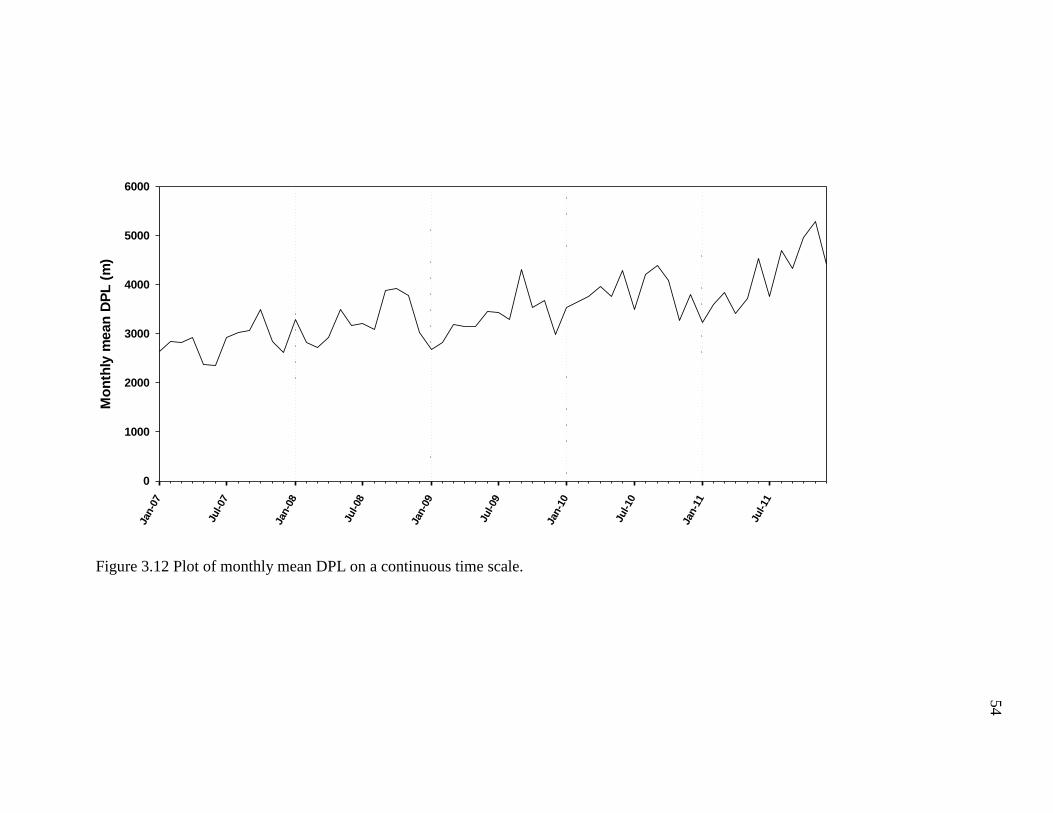
54
Jan-0
7
Jul-07
Jan-0
8
Jul-08
Jan-0
9
Jul-09
Jan-1
0
Jul-10
Jan-1
1
Jul-11
Mo
nth
ly m
ean
DP
L (
m)
0
1000
2000
3000
4000
5000
6000
Figure 3.12 Plot of monthly mean DPL on a continuous time scale.
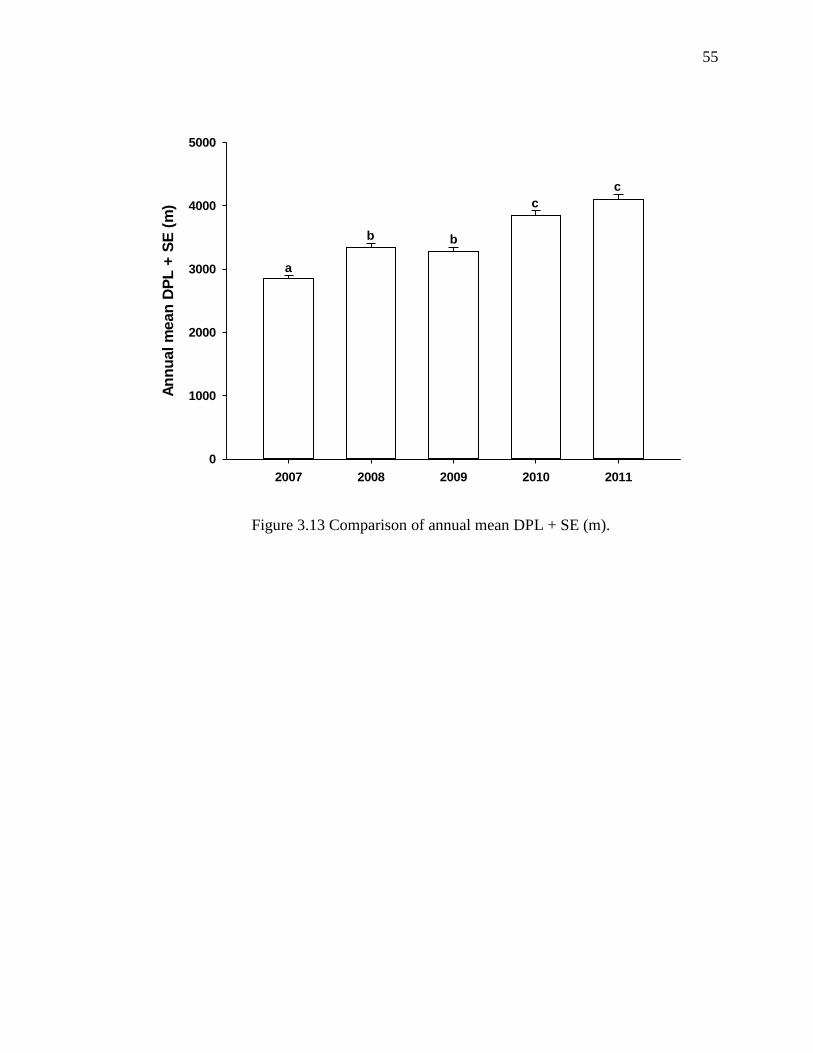
55
2007 2008 2009 2010 2011
An
nu
al m
ean
DP
L +
SE
(m
)
0
1000
2000
3000
4000
5000
a
b b
c
c
Figure 3.13 Comparison of annual mean DPL + SE (m).

56
CHAPTER 4
DISCUSSION
Summary of Findings
Annual home ranges estimated using the MCP method included spacious areas
the geladas never used, which were exacerbated by outliers and ultimately inflated home
range area estimates. Additionally, several sleeping sites were omitted from the 95%
home range estimates.
Like the MCP, both the fixed kernel SCV and REF bandwidths omitted several
sleeping sites from the 95% estimate. Both the SCV and (0.2 and 0.4) REF bandwidths
constructed annual home ranges with discontinuous and broken density contours.
Conversely, the >0.6*REF bandwidth produced continuous density contours, though
these density contours included areas the geladas did not and could not use.
In general, home range estimates estimated with the MPC were larger in area than
those estimated using both the REF and SCV bandwidths.
Gelada monkey annual home range and core area use increased in size from year
to year and were generally larger at the end of the study than at the beginning of the
study, patterns evident across all home range estimation techniques.
Daily path length (DPL), on the other hand, varied considerably within years,
though the total distance the geladas covered on any given day tended to increase from
January through to December. Monthly mean DPLs also varied extensively within and

57
between years, though it can be said that monthly mean DPLs generally increased over
the five-year study period. Overall, annual mean DPL increased significantly between
years, being shortest in 2007 and longest in 2011.
Evaluation of the MCP Method
Annual home ranges calculated using the MCP method were generally larger in
size than those estimated using both fixed kernel methods (Pimley et al. 2005; but see
Boyle et al. 2009), and tended to include areas the geladas never visited (Andreka et al.
1999; Fashing et al. 2007; Pebsworth et al. 2012; Powell 2000), trends that are evident
across the literature. A likely explanation for these observations is that the MCP method
uses straight lines and convex angles to connect the outermost data points to create a
home range whose shape is confined to a rigid and static figure that is unable to capture
the fluid and dynamic movements and space use patterns of animals. All annual home
ranges, for instance, contained a sizeable pocket of empty space in the western region of
the home range, despite the lack of evidence suggesting the geladas were observed in this
area over the five-year study period.
The decision to exclude a small portion of data from home range analysis,
typically the top 5%, has become common practice among studies of animal ranging
ecology (e.g., Andreka et al. 1999; Fashing et al. 2007; Pebsworth et al. 2012; Pimley et
al. 2005; Powell 2000; but see Borger et al. 2006), since this technique has demonstrated
consistently the ability to remove outliers or unusual movements that can have
detrimental impacts on the home range estimate, or at least mitigate their effects. Indeed,
removal of the top 5% from home range analysis reduced the size of the 95% annual
home ranges by 23.4% to 48.4% relative to their respective 100% estimates. The largest

58
reduction occurred between the annual home ranges for the 2010 range year, which saw
the 100% MCP drop from 22.3 km2 to 11.5 km2 in the 95% MCP, a decrease of 48.4%.
Comparisons of the 95% and 100% annual home ranges for all five years show a
decrease in the inclusion of data located on the periphery of the home range, which in
turn reduced the size and prevalence of empty, unused areas within the home range. This
was most evident for the 2010 range year when the geladas made two separate and
uncharacteristically long excursions to the far north of the Guassa study area. The
removal of these extreme data points explains the significant reduction in the home range
area for the 95% annual home range for 2010, and further highlights the susceptibility of
home range estimators such as the MCP to outliers.
Several authors have questioned the decision to exclude data from the home range
analysis (Kernohan et al. 2001; Powell 2000; White and Garrott 1990), and though the
findings reported here appear to show that this strategy of excluding the top 5% is
effective at eliminating unusual data points and producing smaller and relatively more
accurate home ranges, there is evidence that supports the concerns raised by these
authors. Specifically, several sleeping sites, from one to 19 depending on the range year,
were omitted from the 95% annual home range estimates (e.g., Pebsworth et al. 2012).
Recently, Pebsworth et al. (2012), in their investigation of the ranging ecology of chacma
baboons (Papio hamadryas ursinus) at Wildcliff Nature Reserve in Western Cape, South
Africa, reported the loss of two of seven sleeping sites from their 95% MCP estimates. It
is difficult to assess how often biologically important data such as sleeping sites get
removed from the 95% home range estimate, but it appears to be related to the practice of
excluding some portion of the top data from the home range analysis. Presumably, the top

59
5% generally represent the data located on the boundary or fringes of the home range,
movements considered to have relatively little biological significance. The findings here
show, however, that data on the fringes of the home range, such as sleeping sites, can
present critical challenges on the MCP method because such data may not only be
considered an outlier and therefore are more likely to excluded from the home range
analysis, but they can also represent areas essential to the focal subject’s ecology, in this
case sleeping sites (Powell 2000). Indeed, the findings of loss of sleeping sites reported
here and in Pebsworth et al. (2012) question the merit behind the removal of data from
the home range analysis and stress the need for researchers to consider the advantages
and disadvantages of their analytical decisions (e.g., Kernohan et al. 2001; Powell 2000;
White and Garrott 1990).
Home ranges estimated using the MCP method tend to increase as sample size
increases (Bekoff and Mech 1984; Boulander and White 1990; Boyle et al. 2009;
Jennrich and Turner 1969; Schoener 1981). The home ranges estimated at 10-day
intervals—a proxy measure of sample size—demonstrated that the geladas’ home range
increased in size as the number of study days increased, until home range size reached an
asymptote after approximately 570 days of study, a sample size of n = 12,244. Studies
using simulation data (Bekoff and Mech 1984) and telemetry data (Girard et al. 2002)
found 100-200 and 100-300 data points, respectively, could be enough to produce reliable
and accurate home range estimates. The finding that the geladas’ home range continued
to increase well beyond the 200-300 data point threshold suggests that the relationship
between home range size and sample size is much more complex and most likely
involves research-related components, such as sampling regime and analytical methods,

60
and species-specific variables, such as behavior and ecology (Bekoff and Mech 1984;
Boyle et al. 2009). The current study, however, did not categorically attempt to test the
relationship between home range size and sample size in the MCP method (e.g., Boyle et
al. 2009), and thus the conclusions reached here are derived from the analysis of the
home range size at 10-day intervals. Nonetheless, this finding of increasing home range
size with increasing study days is important for two reasons: firstly, it appears to support
the conclusion that home range size is directly correlated with sample size in the MCP
method; and secondly, but more importantly, it demonstrates the value of long-term
studies in discovering and illuminating the nuances in animal ranging behavior attainable
only through extended and continuous research. Lastly, this finding of increasing home
range size with increasing sample days invokes the need to investigate the determinants
of this phenomena in geladas at Guassa, Ethiopia.
Evaluation of the Kernel Estimators
Both fixed kernel estimators produced widely disparate results. The fixed kernel
SCV, for example, generated annual home ranges whose density contours were relatively
more disconnected, fragmented (i.e., islands), and shaped irregularly. As this is the first
study to utilize the fixed kernel SCV bandwidth in a practical setting (see Duong and
Hazelton 2005 for multivariate SCV; see Hall et al. 1992 for univariate SCV for full
descriptions of the bandwidth; see Dobrovidov and Rud’ko 2009 for the SCV bandwidth
applied in statistical and theoretical settings), it is difficult to explain the observed density
contours. Similar findings of fragmented and disconnected density contours have been
reported for the LSCV bandwidth (Amstrup et al. 2004; Blundell et al. 2001; Gitzen et al.
2006), which occur when the LSCV fails to find an appropriate smoothing value for the

61
given dataset (Gitzen et al. 2006; Rodgers et al. 2007; Seaman and Powell 1996). Perhaps
a similar issue may explain the fragmentary contours observed in the SCV estimates,
though additional research is required to confirm whether or not this is the case.
Whereas the MCP incorporated empty, unused areas predominantly in the western
region of home range, the fixed kernel REF, and to a certain extent the fixed kernel SCV,
included inhospitable areas east of the geladas’ sleeping sites that line the eastern border
of the Guassa study site. This finding suggests that data such as sleeping sites that are
located near the periphery of the home range pose an issue to all three home range
estimators as each included areas the animals were never observed in, and thus produced
annual home range values that were overestimates. One possible explanation for these
results is that the REF bandwidth has been reported to produce wide and expansive
density contours—that is, it tends to overestimate the boundaries of the home range—
when used to analyze utilization distributions with more than one center of activity (i.e.,
core area) (Gitzen et al. 2006; Seaman and Powell 1996; Seaman et al. 1999). The
findings reported here suggest that the geladas utilized more than one core area (based on
the 50% density contour), which supports the conclusion that the REF bandwidth is not
an appropriate bandwidth estimator for distributions with multiple centers of activity
(Gitzen et al. 2006; Seaman and Powell 1996; Seaman et al 1999).
Not all annual home ranges estimated using the REF bandwidth included areas the
geladas neither used nor visited. Rather, we found that scaling—i.e., multiplying—the
REF bandwidth by a fixed proportion (i.e., 0.2, 0.4, 0.6, 0.8, and 1.0) (e.g., Gitzen et al.
2006; Pebsworth et al. 2012; Seaman and Powell 1996; Worton 1989) produced annual
home ranges with density contours that varied in degree of continuity and

62
disconnectivity. To determine which proportion produced the most accurate and reliable
home range, Pebsworth et al. (2012) assessed each home range on its ability to include all
sleeping sites and major centers of activity, and the least amount of empty, unused areas.
Based on these criteria, Pebsworth et al. (2012) concluded that a proportion of 0.65 of the
REF generated the most reliable and accurate home range estimate for their study group
of chacma baboons. Using these criteria to assess the results reported here, I conclude
that a proportion of 0.6 of the REF produced a home range that had a combination of
smooth and continuous density contours and density contours that incorporated relatively
smaller degree of areas the geladas were never observed in. Unfortunately, none of the
proportions I analyzed produced an annual home range that contained all sleeping sites
for all five study years while meeting the two criteria above. The findings reported here
and in Pebsworth et al. (2012) demonstrate that the practice of scaling the REF
bandwidth by a fixed proportion affords the user the ability to decide the appropriate
bandwidth value for the given utilization distribution.
Kernel estimators are widely purported to be superior to the MCP method in
home range estimation (Borger et al. 2006; Laver and Kelly 2008), but the findings here
do not definitively support this popular supposition. The results show that both kernel
estimators, like the MCP, incorporated areas the geladas did not utilize and omitted
sleeping sites from the annual home range estimates. In turn, these results demonstrate
the value of utilizing multiple home range tools to estimate home range as results may
not conform to previous findings (Boyle et al. 2009).

63
Implications and Suggestions for Future Research
Studies of animal ranging ecology are imperative for conservation-related
purposes, and the analytical techniques and tools researchers utilize to investigate and
understand aspects of animal movement and home range use patterns are equally critical
for developing sound and effective conservation plans. The results reported here have
wide implications for future research. I therefore propose the following recommendations
based on the observations reported here.
For the choice of home range estimators, I strongly agree the MCP should be still
utilized as a home range estimator despite its widely documented drawbacks and the
growing consensus against its continued use (Borger et al. 2006; Gitzen et al. 2006; Laver
and Kelly 2008). I contend that using the MCP to estimate home range affords
researchers the invaluable ability of making comparisons between methods (Boyle et al.
2009; Pimley et al. 2005; Pebsworth et al. 2012) and within the same study or across
studies and taxa (Biebouw et al. 2009; Fashing et al. 2007; Grueter et al. 2009; Robbins
and McNeilage 2003; Strier 2003; Wartman et al. 2010; Wieczkowski 2005). Moreover, I
recommend using the fixed kernel method to supplement the MCP method (Borger et al.
2006; Boyle et al. 2009; Gitzen et al. 2006; Laver and Kelly 2008; Powell 2000; Seaman
and Powell 1999). The choice of bandwidth will likely vary depending on the nature of
the data being analyzed (Gitzen et al. 2006; Powell 2000), though I recommend using the
LSCV and REF bandwidth estimators. The LSCV, despite its drawbacks (Blundell et al.
2001; Boyle et al. 2009; Gitzen et al. 2006; Pebsworth et al. 2012), is the bandwidth of
choice among researchers (e.g., Powell 2000) and its use is worth attempting. The REF
bandwidth should be implemented with the scaling option at consistent intervals of 0.05,

64
or an interval the researcher finds suitable for the given dataset (e.g., 0.1). Though the
results reported here regarding the home range estimating power of the SCV bandwidth is
inconclusive at best, additional research is needed to ascertain whether or not the SCV
bandwidth will have a place as a reliable home range estimator. Indeed, this sentiment
applies to many bandwidth estimators that have seen little application in home range
studies, e.g., the plug-in methods and cross-validation bandwidths (Beyer 2012; Gitzen et
al. 2006), and thus I recommend researchers test various bandwidth estimators to assess
its efficacy as a home range estimator.
On the topic of data analysis, the decision to subject the entire dataset (i.e., 100%)
or a majority of the data (e.g., 90% or 95%) to home range analysis should be driven
largely by the research question(s) being asked and the behavioral ecology of the focal
subject. As I have demonstrated, in focal subjects, like the geladas at Guassa, where the
periphery of the home range is the location of critical data essential to understanding the
behavioral ecology of the species, the researcher must select a data analysis procedure
that is line with the research question but will simultaneously produce results that are
reliable and accurate. Nonetheless, I strongly advise researchers to conduct home range
analysis using both 95% and 100% of the dataset, since doing so will allow for
comparisons across techniques and the assurance of making informed and sound
decisions supported by quantitative analysis and field notes.
Additional components to consider include sample size and sampling regime.
With regard to sample size, the number of data points needed to produce a reliable and
accurate home range estimate is contingent upon a variety of factors, such as home range
estimator (e.g., Anderson 1982; Bekoff and Mech 1984; Boyle et al. 2009; Seaman and

65
Powell 1999), behavior and ecology of the focal subject (see Bekoff and Mech 1984 for
examples; Girard et al. 2002), and body size (mass) (Clutton-Brock and Harvey 1977).
Research has shown, for example, that the MCP method can underestimate (Girard et al.
2002) and overestimate home range at small sample sizes (Glessener and Britt 2004),
whereas the fixed kernel method can produce a home range that provides a general, but
accurate account of the focal subject’s space use patterns with a small sample size
(sample size of 10 in Borger et al. 2006; sample size of 30-50 in Seaman and Powell
1999; but see Boyle et al. 2009). Whether or not the focal subject lives independently or
in a group can possibly have an effect on the number of data points required to reproduce
an accurate depiction of its home range. Group living may invoke greater instances of
intra-specific competition for resources, which in turn will force the group to make
additional or further movements in search of food (Chapman and Chapman 2000; Dunbar
and Dunbar 1975; Dunbar and Iwamoto 1983; Fashing et al. 2007). Similarly, diet, such
as fruigivory, insectivory, and gummivory, has been shown to influence home range size
(Clutton-Brock and Harvey 1977; Isbell 1998). In light of these variables, the ability to
find the “optimal” sample size may be difficult given the number of factors that influence
the relationship between sample size and home range estimation. I therefore recommend
a sample size that falls within the means of the research goal(s), taking into account each
of the variables described above, including the likelihood and practicality of gathering
such data and the associated costs (i.e., time and money). Finally, it may be more
strategic to gather a plethora of data at the onset and then sub-sample or condense the
data later whenever needs change.

66
The sampling regime, defined as the time interval between subsequent data
points, the length of the study, and which locations should be recorded, is arguably
influenced by the same factors that affect sample size as discussed above. For instance, a
short time between subsequent data points, such as 15 to 30 minutes, may be ideal for
focal subjects that cover long distances in short periods of time. Conversely, a longer
time between subsequent data points of one hour or greater may be more appropriate for
focal subjects that remain immobile in a general area over an extended period of time or
do not travel far enough between sampling intervals (e.g., Hansteen et al. 1997). If the
time interval between subsequent data points is too short, this may lead to data being
autocorrelated, which has been argued to undermine the integrity of the results (Swihart
and Slade 1985a,b; but see Fieberg 2007; de Solla et al. 1999). Depending on the focal
subject, some locations may be more critical than others, e.g., sleeping sites, water holes,
territorial zones, etc., and thus their acquisition should take precedence over other
location data, particularly in the event that time or cost may prevent ample data
acquisition. Where sufficient time and funding are available, researchers beginning
detailed studies of animal ranging ecology should plan to undertake several years of
continuous observation as this will result in (a) more complete estimations of overall
home range size and (b) the identification of any inter-annual differences in movement
and home range use patterns.
In sum, the recommendations described above provide the researcher with the
greatest degree of comparability of results across studies and confidence in research
method supported by extensive research (e.g., Borger et al. 2006; Pimley et al. 2009;
Pebsworth et al. 2012; Powell 2000).

67
Comparison of Gelada Monkey Ranging Behavior Across Sites
One purpose of this study was to compare the DPL, home range size, core area,
and furthest distance traveled from a sleeping cliff site (edge) for geladas at Guassa,
Ethiopia, to those at Sankaber, Gich, and Bole (no ranging data exist for geladas
inhabiting the study area at Arsi, Ethiopia: Mori and Belay 1990; Mori et al. 1999). Both
Sankaber and Gich are located within 15 km of each other in the Simien Mountains
National Park in central Ethiopia (Dunbar and Dunbar 1975; Iwamoto and Dunbar 1983;
Kawai 1979). They are approximately ~305 km and ~320 km northwest of Guassa,
respectively. The study site of Bole, on the other hand, lies ~ 100-200 (south)west of
Guassa, near Addis Ababa, the capital of Ethiopia (Dunbar and Dunbar 1974; Iwamoto
and Dunbar 1983).
One obstacle to comparing home range estimates across sites or species (here and
below) concerns the types of method(s) used to estimate the home range. For example, I
estimated home range size using both the MCP and fixed kernel methods, whereas
Hunter (2001) utilized the grid-cell method with 200 x 200 m quadrats (Dunbar and
Dunbar 1975 and Kawai 1979 did not specify which methods they used). Because each
method employs different assumptions or criteria to construct a home range, the use of
different techniques makes it problematic to make direct and robust comparisons about
space use patterns. In addition to using different methods to calculate home range size,
the time scale in which the ranging data are collected is another obstacle to consider.
Home ranges estimated from data gathered over only a few weeks or several months, for
example, are not the same as home ranges estimated using data obtained from an entire
year or more, because each estimate reflects the habitat and space use patterns of the

68
animal during different times of the year and under (presumably) different environmental
conditions.
How do the Annual Home Range Estimates of Geladas at Guassa Compare to Those for
gGeladas at Other Sites?
Mean home range size of the geladas at Guassa during the five-year period was
7.06 km2 (fk SCV), 9.12 km2 (fk LSCV), and 9.28 km2 (MCP). The five-year mean home
range estimates reported for the fk LSCV and MCP, but not the fk SCV, methods are
similar to the home range size reported for geladas at Sankaber, 9.28 km2 (Hunter 2011),
studied over a one-year period. Meanwhile, mean home ranges of both the geladas at
Guassa (this study) and at Sankaber (Hunter 2011) are much larger than those reported
for geladas at Bole and Gich (Dunbar 1977; Dunbar and Dunbar 1975; Iwamoto and
Dunbar 1983; Kawai and Iwamoto 1979).
A comparison of annual home ranges, on the other hand, indicates that though
annual home range in the geladas at Guassa started out smaller (from 2007-2009), it
eventually eclipsed (in 2010 and 2011) the estimate reported by Hunter (2001). This
indicates that the range use patterns of geladas at Guassa exhibit extreme inter-annual
variability, and demonstrates the value of long-term monitoring of nonhuman primate
ranging patterns, particularly for gelada populations.
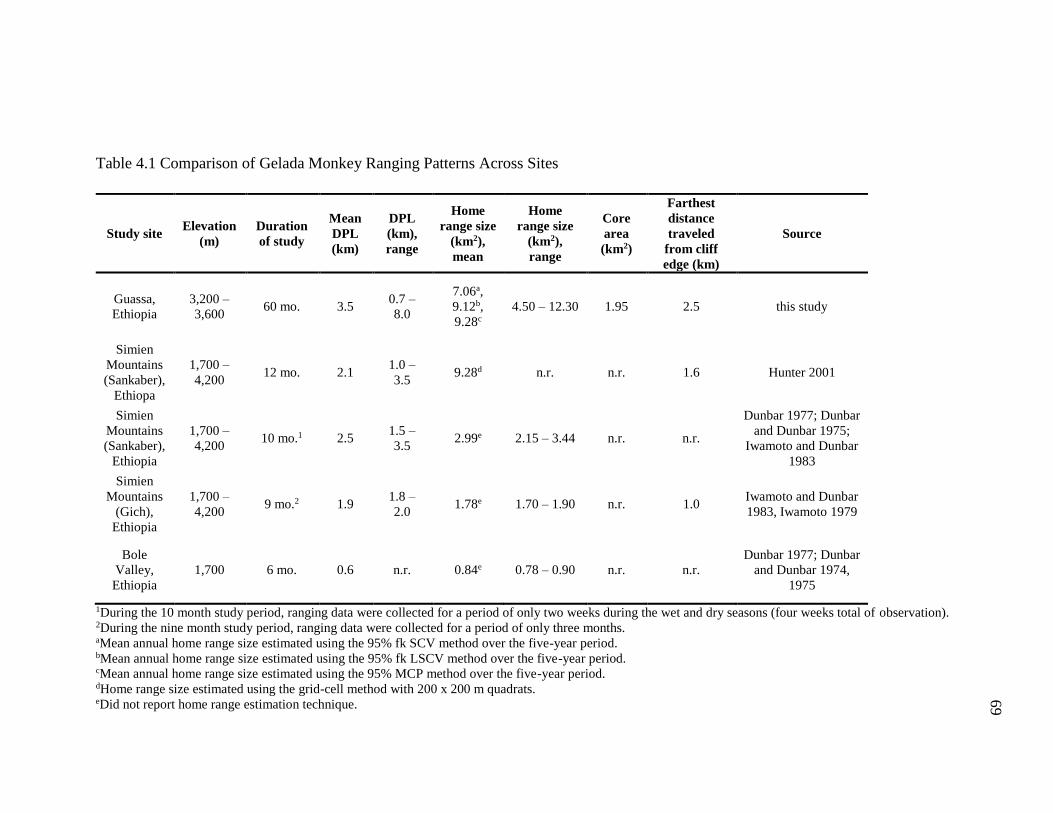
69
Table 4.1 Comparison of Gelada Monkey Ranging Patterns Across Sites
Study site Elevation
(m)
Duration
of study
Mean
DPL
(km)
DPL
(km),
range
Home
range size
(km2),
mean
Home
range size
(km2),
range
Core
area
(km2)
Farthest
distance
traveled
from cliff
edge (km)
Source
Guassa,
Ethiopia
3,200 –
3,600 60 mo. 3.5
0.7 –
8.0
7.06a,
9.12b,
9.28c
4.50 – 12.30 1.95 2.5 this study
Simien
Mountains
(Sankaber),
Ethiopa
1,700 –
4,200 12 mo. 2.1
1.0 –
3.5 9.28d n.r. n.r. 1.6 Hunter 2001
Simien
Mountains
(Sankaber),
Ethiopia
1,700 –
4,200 10 mo.1 2.5
1.5 –
3.5 2.99e 2.15 – 3.44 n.r. n.r.
Dunbar 1977; Dunbar
and Dunbar 1975;
Iwamoto and Dunbar
1983
Simien
Mountains
(Gich),
Ethiopia
1,700 –
4,200 9 mo.2 1.9
1.8 –
2.0 1.78e 1.70 – 1.90 n.r. 1.0
Iwamoto and Dunbar
1983, Iwamoto 1979
Bole
Valley,
Ethiopia
1,700 6 mo. 0.6 n.r. 0.84e 0.78 – 0.90 n.r. n.r.
Dunbar 1977; Dunbar
and Dunbar 1974,
1975
1During the 10 month study period, ranging data were collected for a period of only two weeks during the wet and dry seasons (four weeks total of observation). 2During the nine month study period, ranging data were collected for a period of only three months. aMean annual home range size estimated using the 95% fk SCV method over the five-year period. bMean annual home range size estimated using the 95% fk LSCV method over the five-year period. cMean annual home range size estimated using the 95% MCP method over the five-year period. dHome range size estimated using the grid-cell method with 200 x 200 m quadrats. eDid not report home range estimation technique.

70
How do Geladas Utilize Their Home Range at Guassa and How Does It Compare to That
of Geladas at Other Sites?
Geladas, like many other species of non-human primates, (e.g., spider monkeys:
Asensio et al. 2011; chimpanzees: Basabose 2005; sifakas: Gerber et al. 2011; mountain
gorillas: Watts 1998), do not utilize their home range in a uniform manner, and instead
exhibit preferential use of some areas relative to others (Dunbar 1977; Dunbar and
Dunbar 1975; Hunter 2001; Kawai and Iwamoto 1979; Ohsawa 1979; this study). The
Guassa area consists of a blend of numerous habitat types interspersed within its
boundaries (Ashenafi 2001; Fashing et al. 2014). It is conceivable that the distribution of
the main food source of geladas, green grass blades (Dunbar and Dunbar 1975; Hunter
2001; Iwamoto 1979; Fashing et al. 2014), within these patchily distributed habitats is
likely to result in differential space use patterns (core areas) over time, because animals
are expected to situate themselves in areas where they can maximize energy acquisition
while simultaneously minimizing energy expenditure (Stephen and Krebs 1986). Indeed,
as Dunbar and others (Dunbar 1977; Dunbar and Dunbar 1975; Hunter 2001; Kawai
1979) have indicated in their shorter duration studies of gelada monkey ranging ecology,
the space use patterns of geladas appear to be related to the spatial and temporal
availability and distribution of resources and weather conditions, e.g., thick fog or
rainfall. The fact that the geladas at Guassa are exhibiting preferential space use, as
indicated by the core areas, lends credence to the hypothesis that the animals may be
selecting and concentrating most of their activities in areas of relatively higher resource
availability. Future research should seek to obtain detailed data on resource availability
and distribution over time to further investigate this hypothesis.

71
How do the DPL of Geladas at Guassa Compare to Those of Geladas at Other Ses?
Geladas at Guassa travel, on average, substantially further per day than those at
Sankaber, Gich, and Bole (Dunbar 1977; Dunbar and Dunbar 1974, 1975; Hunter 2001;
Iwamoto and Dunbar 1983; Kawai and Iwamoto1979). Furthermore, the longest DPL
observed at Guassa was 8.0 km, more than twice the longest distance (3.5 km) recorded
for geladas at Sankaber (Hunter 2001) and four times the distance (2.0 km) for geladas at
Gich (Kawai and Iwamoto 1979). No minimum or maximum DPL estimates were
reported for geladas at Bole.
Comparison of Ranging Behavior Across Taxa
A secondary objective of this study was to compare the DPL, annual home range,
and core area of geladas to the ranging behavior reported for (various) species of
nonhuman primates. I separated my analysis into four categories: Papio spp.
(phylogenetic relationship) in Table 4.2; terrestrial nonhuman primates (mode of
locomotion); arboreal frugivores; and arboreal folivores (mode of locomotion and dietary
profile), all in Table 4.3. Dividing the numerous species of nonhuman primates in this
manner helped facilitate direct comparisons among the different ecological groups to
which nonhuman primates make up (e.g., Clutton-Brock and Harvey 1977). Ideally, I
would have liked to base the comparisons on all species of nonhuman primates for which
ranging data are available, because this would provide us with a (near) complete and
thorough analysis of nonhuman primate ranging behavior (please refer to Campbell et al.
2011 and the various chapters in this text for a complete list of ranging studies on
nonhuman primates). However, I decided to only include data from nonhuman primate
populations studied over a period of one year or greater, because the data are more likely

72
to depict the animals’ ranging patterns over the course of (at least) an entire annual cycle
(which also corresponds with the data presented in our study). Lastly, as I have discussed
above, making comparisons across species is problematic due to the differential use of
methods to estimate home range in each study. Therefore, I focus my attention only on
the value(s) reported and do not make any assumptions beyond that.
Comparison of Gelada Ranging Behavior to Papio Species
Baboons (Papio spp.), travel, on average, further per day and occupy home ranges
larger in size than geladas (see Table 4.2). If I compare the ranging behavior of gelada
monkeys to individual species of Papio, however, different relationships emerge.
Geladas, particularly the Guassa geladas (mean DPL = 3.5 km), exhibit a mean
DPL comparable to, though slightly shorter than chacma baboons (P. cyncephalus
ursinus) studied at the Drakensberg Mountains, Natal Province, South Africa (4.1 km:
Henzi et al. 1992; Whiten et al. 1987) and at the Suikerbosrand Nature Reserve,
Transvaal, South Africa (4.1 km: Anderson 1981, 1982). No study populations of
geladas, however, travel as far per day as those chacma baboons at Tshipise, Transvaal,
South Africa (8.5 km: Stoltz and Saayman 1970). Despite their fairly similar DPLs, home
range size in chacma baboons (17.2 – 24.6 km2) are several times larger than those
reported for geladas (0.84 – 12.3 km2).
The mean DPL reported for yellow (P. cynacephalus) (5.6 km: Barton et al.
1992), olive (P. anubis) (5.0 km: Bronikowski and Altmann 1996), and hamadyras (P.
hamadryas) (7.5 – 13.2 km) baboons are longer than those observed for all studied
groups of geladas. Similarly, home range sizes for olive (4.1 – 43.8 km2) and hamadyras

73
(28 – 30 km2) baboons are (considerably) larger than those reported for geladas in
general.
What can explain the observed differences and similarities in the ranging behavior
of geladas and Papio spp.? Research has indicated that variations in group (herd) size,
temperature, food availability and distribution, access to water, weather patterns, and
proximity or access to sleeping cliff sites have been suggested to influence both the
movement and space use patterns among Papio (Barton et al. 1992; Bronikowski and
Altmann 1996; Henzi et al. 1992; Kunz and Lisenmair 2008; Stoltz and Saayman 1970;
Schreier 2010; Smuts 1985; Swedell 2006, 2011; Whiten et al. 1987) and geladas
(Dunbar and Dunbar 1975; Hunter 2001; Kawai 1979). Here, I elaborate on two of these
factors and their effects on ranging behavior in baboons and geladas: access to sleeping
sites and waterholes.
Whiten et al. (1987) have postulated that access to multiple sleeping cliff sites and
the differential use of sleeping cliff sites on a nightly basis may explain the shorter day
ranges observed in their population of chacma baboons at the Drakensberg Mountains,
Natal Province, South Africa, than compared to other species of baboons (e.g.,
hamadryas and olive baboons: Kunz and Lisenmair 2008; Sigg and Stolba 1981; Smuts
1985; Swedell 2006). Having access to multiple (and suitable) sleeping sites reduces the
amount of travel an animal has to invest in when searching for a place to sleep (Whiten et
al. 1987). Like chacma baboons, geladas have access to multiple sleeping sites and also
tend to use different sleeping sites on a nightly basis, though repeated use of a sleeping
cliff site on consecutive days has been observed many times in this band of geladas as
well (Moua unpubl. data). Conversely, hamadryas baboons regularly use the same

74
sleeping cliff site for numerous days on end (Sigg and Stolba 1981; Swedell 2006), and
coupled with the lack of potential sleeping cliff sites (at Filoha: Swedell 2006; and
Comoé National Park, Ivory Coast: Kunz and Linsenmair 2008), this may explain the
longer day ranges in these animals relative to chacma baboons and geladas.
In addition to the impact access to sleeping sites can have on the movement and
space use patterns, access to water has also been considered a factor in determining how
far baboons travel on a daily basis, which areas of their habitat are used, and the size of
the home range (Barton et al. 1992; Hamilton et al. 1976; Kunz and Linsenair 2008;
Smuts 1985; Stoltz and Saayman 1970; Swedell 2011). For example, chacma baboons
inhabiting the study site at Tshipise, Transvaal, South Africa, traveled long distances to
waterholes widely dispersed throughout their home range (Stoltz and Saayman 1970).
Furthermore, limited access to water can result in larger home ranges (olive baboons:
Kunz and Linsenmair 2008), because the animals would have to incorporate a home
range of relatively larger area to compensate for the lack of available waterholes. In
geladas, such as those at Gich (Kawai and Iwamoto 1979), the high availability of water
sources within the home range facilitates minimal movement to and from waterholes;
however, this implies that where access to water is (becomes) limited, it is conceivable
geladas, like baboons, are likely to increase movement or expand their home range in
search for additional sources of water. Future research should aim to obtain more
information about the geographic distribution of waterholes within the home range of the
geladas at Guassa, and record the behavior of the Guassa geladas in relation to drinking
water to better understand the relationship between access to water and movement and
space use patterns.
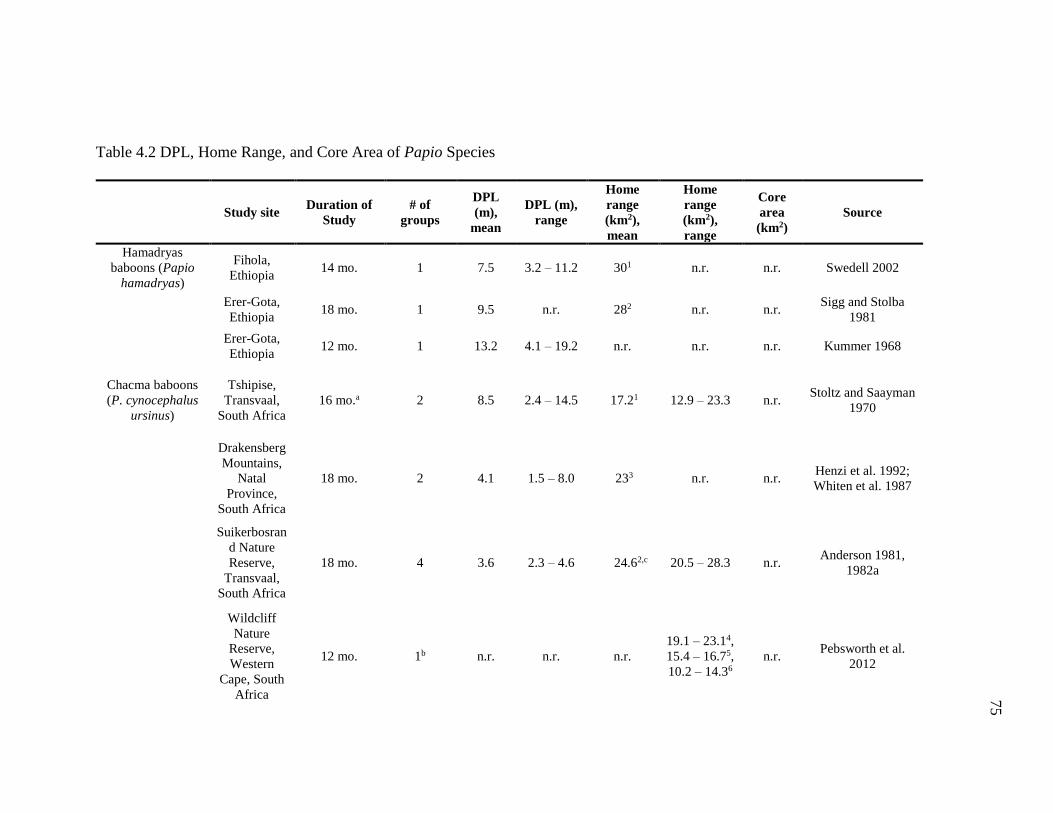
75
Table 4.2 DPL, Home Range, and Core Area of Papio Species
Study site Duration of
Study
# of
groups
DPL
(m),
mean
DPL (m),
range
Home
range
(km2),
mean
Home
range
(km2),
range
Core
area
(km2)
Source
Hamadryas
baboons (Papio
hamadryas)
Fihola,
Ethiopia 14 mo. 1 7.5 3.2 – 11.2 301 n.r. n.r. Swedell 2002
Erer-Gota,
Ethiopia 18 mo. 1 9.5 n.r. 282 n.r. n.r.
Sigg and Stolba
1981
Erer-Gota,
Ethiopia 12 mo. 1 13.2 4.1 – 19.2 n.r. n.r. n.r. Kummer 1968
Chacma baboons
(P. cynocephalus
ursinus)
Tshipise,
Transvaal,
South Africa
16 mo.a 2 8.5 2.4 – 14.5 17.21 12.9 – 23.3 n.r. Stoltz and Saayman
1970
Drakensberg
Mountains,
Natal
Province,
South Africa
18 mo. 2 4.1 1.5 – 8.0 233 n.r. n.r. Henzi et al. 1992;
Whiten et al. 1987
Suikerbosran
d Nature
Reserve,
Transvaal,
South Africa
18 mo. 4 3.6 2.3 – 4.6 24.62,c 20.5 – 28.3 n.r. Anderson 1981,
1982a
Wildcliff
Nature
Reserve,
Western
Cape, South
Africa
12 mo. 1b n.r. n.r. n.r.
19.1 – 23.14,
15.4 – 16.75,
10.2 – 14.36
n.r. Pebsworth et al.
2012
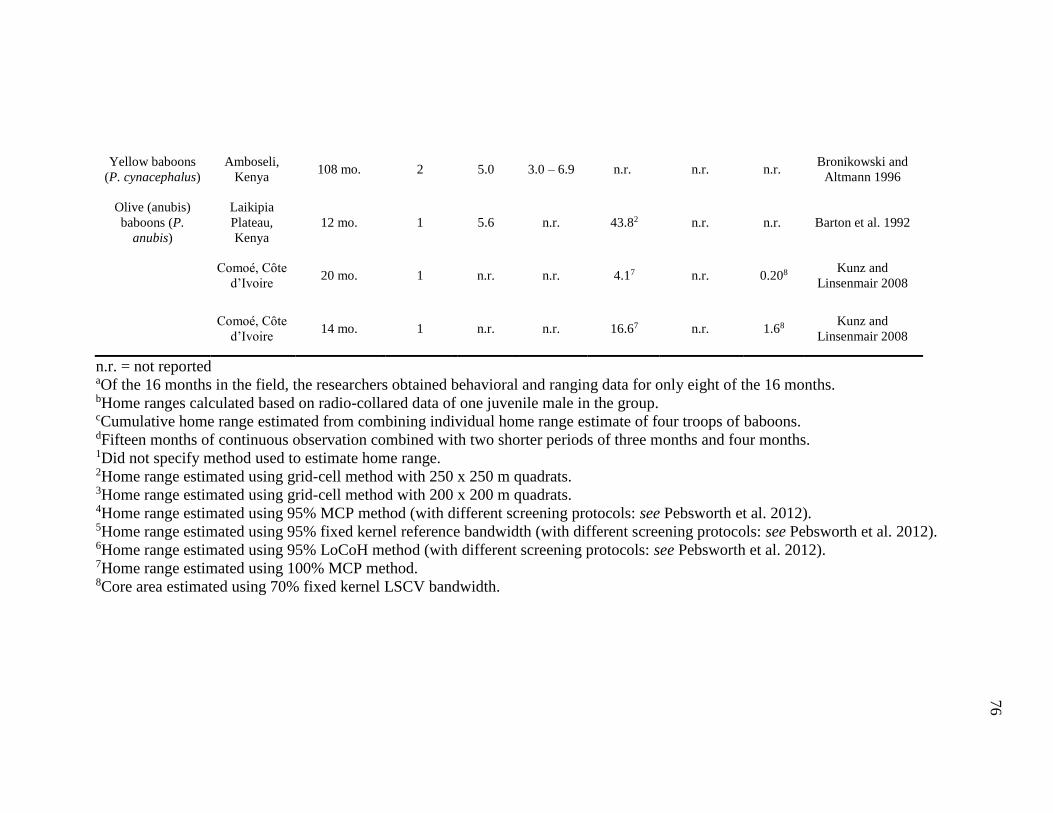
76
Yellow baboons
(P. cynacephalus)
Amboseli,
Kenya 108 mo. 2 5.0 3.0 – 6.9 n.r. n.r. n.r.
Bronikowski and
Altmann 1996
Olive (anubis)
baboons (P.
anubis)
Laikipia
Plateau,
Kenya
12 mo. 1 5.6 n.r. 43.82 n.r. n.r. Barton et al. 1992
Comoé, Cȏte
d’Ivoire 20 mo. 1 n.r. n.r. 4.17 n.r. 0.208 Kunz and
Linsenmair 2008
Comoé, Cȏte
d’Ivoire 14 mo. 1 n.r. n.r. 16.67 n.r. 1.68 Kunz and
Linsenmair 2008
n.r. = not reported aOf the 16 months in the field, the researchers obtained behavioral and ranging data for only eight of the 16 months. bHome ranges calculated based on radio-collared data of one juvenile male in the group. cCumulative home range estimated from combining individual home range estimate of four troops of baboons. dFifteen months of continuous observation combined with two shorter periods of three months and four months. 1Did not specify method used to estimate home range. 2Home range estimated using grid-cell method with 250 x 250 m quadrats. 3Home range estimated using grid-cell method with 200 x 200 m quadrats. 4Home range estimated using 95% MCP method (with different screening protocols: see Pebsworth et al. 2012). 5Home range estimated using 95% fixed kernel reference bandwidth (with different screening protocols: see Pebsworth et al. 2012). 6Home range estimated using 95% LoCoH method (with different screening protocols: see Pebsworth et al. 2012). 7Home range estimated using 100% MCP method. 8Core area estimated using 70% fixed kernel LSCV bandwidth.

77
Comparison of Gelada Ranging Behavior to Terrestrial Nonhuman Primate Species
A comparison of the DPL and home range of geladas to various terrestrial
primates, such as patas monkeys (Erythrocebus patas pyrrhonotus), vervet monkeys
(Cercopithecus aethiops), gorillas (Gorilla beringei beringei), and chimpanzees (Pan
troglodytes), show geladas are somewhere intermediate among those terrestrial primates
(Table 4.3). Geladas, particularly the Guassa geladas, and patas monkeys, for example,
exhibit similar day journey lengths (600 – 3,495 m and 3,830 – 4,220 m, respectively),
however, patas monkeys live in much larger home ranges (28.5 km2: Chism and Rowell
1988; Isbell 1998; Isbell et al. 1999) than do geladas (0.78 km2 – 9.28 km2). Vervet
monkeys (Isbell et al. 1999), on the other hand, do not occupy home ranges (0.15 – 1.15
km2) as large as those reported for geladas. In comparison to specific chimpanzee
populations, geladas, particularly those at Guassa and Sankaber, utilize home ranges
similar in area to the chimpanzees studied at Kahuzi (mean: 7.55 km2; total area: 12.81
km2: Basabose 2005), half the area compared to the chimpanzees at Taї National Park (27
km2: Boesch and Boesch 1989), and significantly smaller than the chimpanzees at Mt.
Assirik, Senegal (278 – 333 km2: Baldwin et al. 1982). Lastly, in comparison to mountain
gorillas, geladas (at Guassa and Sankaber) occupy home ranges similar in size to the
group studied by Vedder (1984: 8.56 km2) and Watts (1998: 8.1 km2), but much smaller
than the group studied recently by Robbins and McNeilage (2003: mean = 27.7 km2,
range = 21.1 – 40.1 km2, using the MCP method).
Overall, our findings with regards to the relationship between DPL and home
range and feeding ecology in nonhuman primates are mostly consistent with conclusions
reached earlier by the influential work of Clutton-Brock and Harvey (1977). Despite
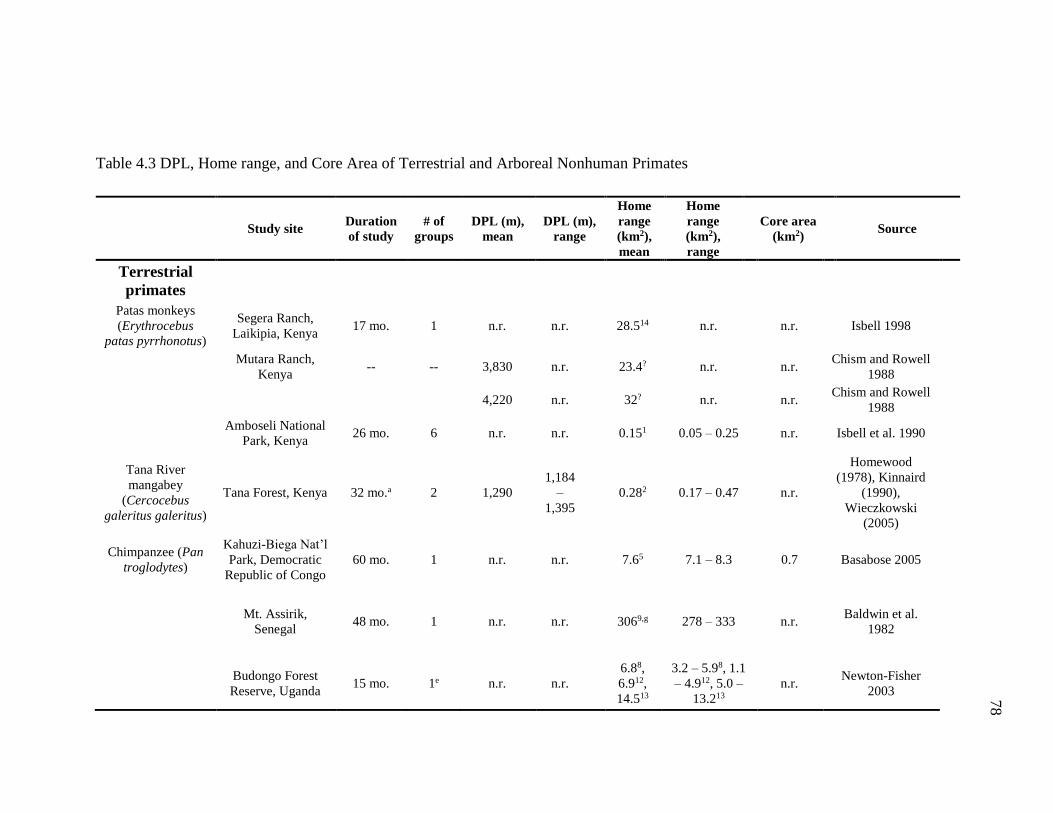
78
Table 4.3 DPL, Home range, and Core Area of Terrestrial and Arboreal Nonhuman Primates
Study site Duration
of study
# of
groups
DPL (m),
mean
DPL (m),
range
Home
range
(km2),
mean
Home
range
(km2),
range
Core area
(km2) Source
Terrestrial
primates
Patas monkeys
(Erythrocebus
patas pyrrhonotus)
Segera Ranch,
Laikipia, Kenya 17 mo. 1 n.r. n.r. 28.514 n.r. n.r. Isbell 1998
Mutara Ranch,
Kenya -- -- 3,830 n.r. 23.4? n.r. n.r.
Chism and Rowell
1988
4,220 n.r. 32? n.r. n.r. Chism and Rowell
1988
Amboseli National
Park, Kenya 26 mo. 6 n.r. n.r. 0.151 0.05 – 0.25 n.r. Isbell et al. 1990
Tana River
mangabey
(Cercocebus
galeritus galeritus)
Tana Forest, Kenya 32 mo.a 2 1,290
1,184
–
1,395
0.282 0.17 – 0.47 n.r.
Homewood
(1978), Kinnaird
(1990),
Wieczkowski
(2005)
Chimpanzee (Pan
troglodytes)
Kahuzi-Biega Nat’l
Park, Democratic
Republic of Congo
60 mo. 1 n.r. n.r. 7.65 7.1 – 8.3 0.7 Basabose 2005
Mt. Assirik,
Senegal 48 mo. 1 n.r. n.r. 3069,g 278 – 333 n.r.
Baldwin et al.
1982
Budongo Forest
Reserve, Uganda 15 mo. 1e n.r. n.r.
6.88,
6.912,
14.513
3.2 – 5.98, 1.1
– 4.912, 5.0 –
13.213
n.r. Newton-Fisher
2003

79
Mountain gorilla
(Gorilla beringei
beringei)
Bwindi
Impenetrable
National Park,
Uganda
36 mo. 1 n.r. n.r. 206, 289 16.3 – 286,
21.1 – 40.19 9.5b-c Robbins and
McNeilage 2003
Bwindi
Impenetrable
National Park,
Rwanda
84 mo. 3 – 5 n.r. n.r. 8.15 3.1 – 15.6 2.9 Watts 1998
Bwindi
Impenetrable
National Park,
Rwanda
18 mo. 1 n.r. n.r. 8.565 n.r. n.r. Vedder 1984
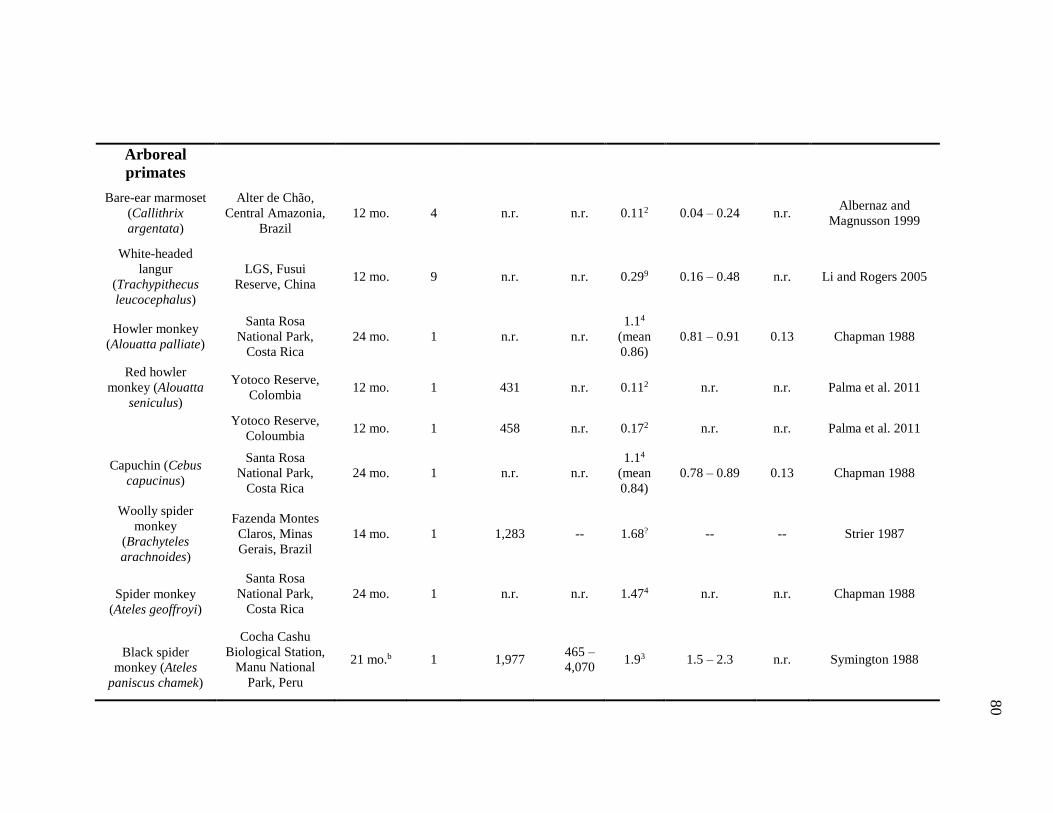
80
Arboreal
primates
Bare-ear marmoset
(Callithrix
argentata)
Alter de Chão,
Central Amazonia,
Brazil
12 mo. 4 n.r. n.r. 0.112 0.04 – 0.24 n.r. Albernaz and
Magnusson 1999
White-headed
langur
(Trachypithecus
leucocephalus)
LGS, Fusui
Reserve, China 12 mo. 9 n.r. n.r. 0.299 0.16 – 0.48 n.r. Li and Rogers 2005
Howler monkey
(Alouatta palliate)
Santa Rosa
National Park,
Costa Rica
24 mo. 1 n.r. n.r.
1.14
(mean
0.86)
0.81 – 0.91 0.13 Chapman 1988
Red howler
monkey (Alouatta
seniculus)
Yotoco Reserve,
Colombia 12 mo. 1 431 n.r. 0.112 n.r. n.r. Palma et al. 2011
Yotoco Reserve,
Coloumbia 12 mo. 1 458 n.r. 0.172 n.r. n.r. Palma et al. 2011
Capuchin (Cebus
capucinus)
Santa Rosa
National Park,
Costa Rica
24 mo. 1 n.r. n.r.
1.14
(mean
0.84)
0.78 – 0.89 0.13 Chapman 1988
Woolly spider
monkey
(Brachyteles
arachnoides)
Fazenda Montes
Claros, Minas
Gerais, Brazil
14 mo. 1 1,283 -- 1.68? -- -- Strier 1987
Spider monkey
(Ateles geoffroyi)
Santa Rosa
National Park,
Costa Rica
24 mo. 1 n.r. n.r. 1.474 n.r. n.r. Chapman 1988
Black spider
monkey (Ateles
paniscus chamek)
Cocha Cashu
Biological Station,
Manu National
Park, Peru
21 mo.b 1 1,977 465 –
4,070 1.93 1.5 – 2.3 n.r. Symington 1988
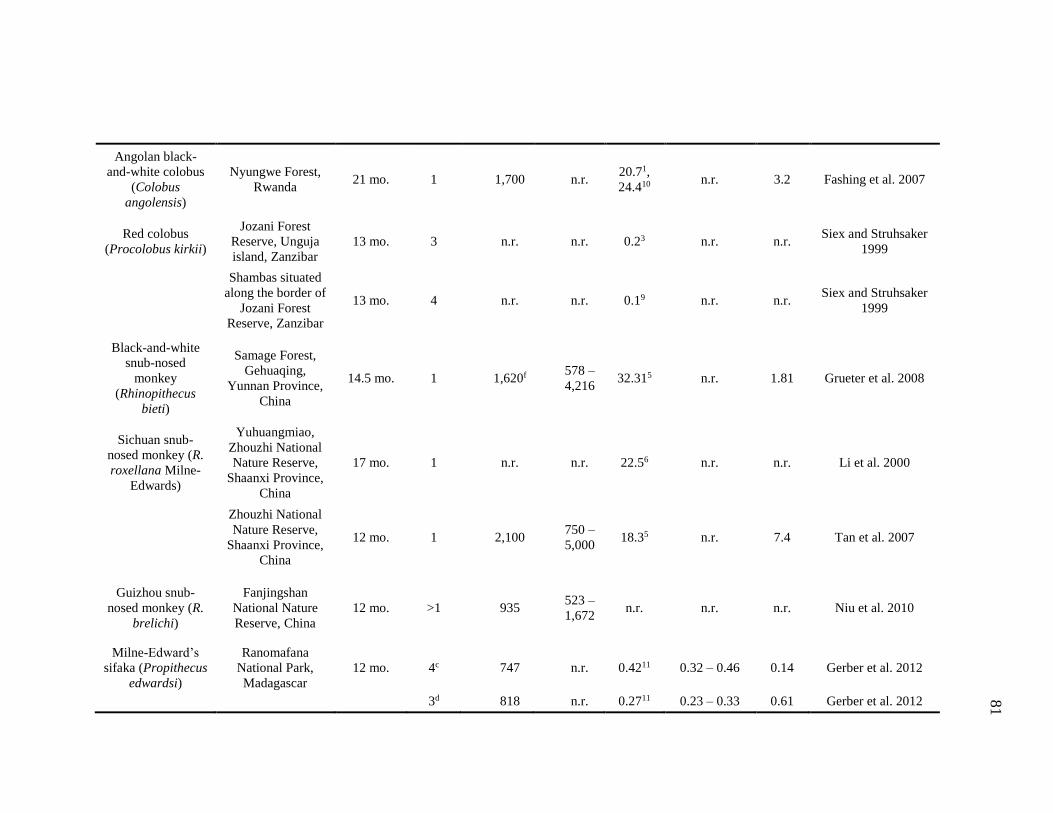
81
Angolan black-
and-white colobus
(Colobus
angolensis)
Nyungwe Forest,
Rwanda 21 mo. 1 1,700 n.r.
20.71,
24.410 n.r. 3.2 Fashing et al. 2007
Red colobus
(Procolobus kirkii)
Jozani Forest
Reserve, Unguja
island, Zanzibar
13 mo. 3 n.r. n.r. 0.23 n.r. n.r. Siex and Struhsaker
1999
Shambas situated
along the border of
Jozani Forest
Reserve, Zanzibar
13 mo. 4 n.r. n.r. 0.19 n.r. n.r. Siex and Struhsaker
1999
Black-and-white
snub-nosed
monkey
(Rhinopithecus
bieti)
Samage Forest,
Gehuaqing,
Yunnan Province,
China
14.5 mo. 1 1,620f 578 –
4,216 32.315 n.r. 1.81 Grueter et al. 2008
Sichuan snub-
nosed monkey (R.
roxellana Milne-
Edwards)
Yuhuangmiao,
Zhouzhi National
Nature Reserve,
Shaanxi Province,
China
17 mo. 1 n.r. n.r. 22.56 n.r. n.r. Li et al. 2000
Zhouzhi National
Nature Reserve,
Shaanxi Province,
China
12 mo. 1 2,100 750 –
5,000 18.35 n.r. 7.4 Tan et al. 2007
Guizhou snub-
nosed monkey (R.
brelichi)
Fanjingshan
National Nature
Reserve, China
12 mo. >1 935 523 –
1,672 n.r. n.r. n.r. Niu et al. 2010
Milne-Edward’s
sifaka (Propithecus
edwardsi)
Ranomafana
National Park,
Madagascar
12 mo. 4c 747 n.r. 0.4211 0.32 – 0.46 0.14 Gerber et al. 2012
3d 818 n.r. 0.2711 0.23 – 0.33 0.61 Gerber et al. 2012

82
n.r. = not reported aCombined study durations of Homewood (1978) (7 mo.), Kinnaird (1990) (15 mo.), and Wieczkowski (2005) (12 mo.) because same
groups were studied across all three study periods. bStudied over a four-year period from June 1982 to June 1986. cA total of 9 adults were sampled from 4 different groups inhabiting the ‘logged site’. dA total of 6 adults were sampled from 3 different groups inhabiting the ‘unlogged site’. eHome ranges estimated for male chimpanzees. Values in ‘HOME RANGE SIZE (km2), MEAN’ represent a composite home range
estimated using ranging data for all males, whereas values in ‘HOME RANGE SIZE (km2), RANGE’ represent the variation in the
home range size of individual male chimpanzees. fFull data for ranging were only available for the month of September, and thus DPL for entire study period was extrapolated based on
data in this month only, however, Grueter et al. (2008) indicate September ranging was representative of the group’s movement
patterns throughout the entire year. gUsed the minimum convex polygon method to estimate initial home range (did not specify percentage used), but also used nest sites
and density of chimpanzees to extrapolate home range size (Baldwin et al. 1982). 1Home range estimated using the grid-cell method with 33 x 33 m quadrats. 2Home range estimated using the grid-cell method with 50 x 50 m quadrats. 3Home range estimated using the grid-cell method with 100 x 100 m quadrats. 4Home range estimated using the grid-cell method with 120 x 120 m quadrats. 5Home range estimated using the grid-cell method with 250 x 250 m quadrats. 6Home range estimated using the grid-cell method with 500 x 500 m quadrats. 7Home range estimated using 95% minimum convex polygon method. 8Home range estimated using 100% minimum convex polygon method. 9Home range estimated using minimum convex polygon method (did not specify percentage used). Li and Rogers (2005) used the
MCP method, but selectively removed unused or inhabitable areas, e.g., flat land between hills. 10Home range estimated using the 95% fixed kernel with least squares cross-validation bandwidth. 11Home range estimated using the 95% fixed kernel with root-n bandwidth. 12Home range estimated using the 99% fixed kernel with least squares cross-validation bandwidth. 13Home range estimated using the 99% adaptive kernel with least squares cross-validation bandwidth. 14Did not specify home range method used. ?Method of home range technique unknown.

83
these general conclusions about the relationship between ranging behavior and feeding
ecology in nonhuman primates, it is intriguing to see, for example, why geladas and patas
monkeys exhibit such disparate ranging behaviors given that they are both terrestrial and
live in relatively large group sizes. One possible explanation concerns the (primary) food
item(s) that makes up each animal’s respective dietary profile. It is considered, for
example, that the consumption of fruits or invertebrates would result in longer DPLs and
larger home ranges due to the wide spatial and temporal variability of these resources
(Clutton-Brock and Harvey 1977). Though patas monkeys primarily consume insects and
gum (Chism and Rowell 1988; Isbell 1998), a study by Isbell (1998) found that access to
waterholes, and not invertebrates or gums, was the main contributing factor for the large
home range size reported in her study group of patas monkeys.
Comparison of Gelada Ranging Behavior to Arboreal Nonhuman Primate Species
In comparison to both arboreal frugivores and folivores, with the exception of the
Angolan black-and-white colobus (Colobus angolensis ruwenzorii) group studied by
Fashing et al. (2007) at Nyungwe Forest, Rwanda, geladas appear to exhibit day journey
lengths and home ranges that are generally longer and larger, respectively, than their
arboreal fruit and non-fruit eating counterparts (see Table 4.3). For example, both fruit
and non-fruit eating arboreal primates, such as muriquis (Brachyteles arachnoides
hypoxanthus) (Dias and Strier 2003), red colobus (Procolobus badius), black-and-white
colobus (Colobus guereza) (Chapman and Pavelka 2005), spider monkeys (Ateles
geofroyi) (Chapman 1990), and Javan gibbons (Hyobates moloch) (Kim et al. 2011), to
name a few, all occupy home ranges similar to (e.g., Gich and Bole) or smaller (e.g.,
Sankaber and Guassa) in size than geladas. Some species of arboreal nonhuman primates,

84
however, do appear to exhibit mean DPLs similar to or longer than the populations of
geladas at Gich, Bole, and Sankaber, but not those at Guassa: black spider monkey
(Ateles geoffroyi, 1,977 m: Chapman 1988; Angolan black-and-white colobus (Colobus
angolensis, 1,700 m: Fashing et al. 2007); black-and-white snub-nosed monkey
(Rhinopithecus bieti, 1,620 m: Grueter et al. 2008; and the Sichuan snub-nosed monkeys
(R. roxellana Milne-Edwards, 2,100 m: Tan et al. 2007).
Furthermore, as mentioned above, in their investigation of the ranging behavior of
Angolan black-and-white colobus (Colobus angolensis ruwenzorii) at Nyungwe, Fashing
et al. (2007) found these monkeys occupied a home range size of 20.7 to 24.4 km2
(estimated using the 95% fixed kernel with LSCV and MCP methods, respectively),
considerably larger than the home range reported for any other species of colobus
(Fashing et al. 2007) and numerous species of nonhuman primates reported here,
including geladas (see Tables 4.1-4.3). Despite being primarily leaf consumers and living
in a habitat shown to have high resource availability, the authors suggest that the
monkeys’ uncharacteristically large group size (> 300 individuals) may be creating a
situation in which even the abundant supply of resources is insufficient to support a group
size of that magnitude (Fashing et al. 2007). Therefore, to compensate for such a large
group size, the study group invested more time in moving and feeding at the expense of
resting time, while simultaneously increasing their DPL and home range (Fashing et al.
2007). Dunbar and Dunbar (1975) and Iwamoto and Dunbar (1983) have indicated gelada
monkeys travel further when there are more animals present in the herd. A similar
argument has been made for various species of nonhuman primates (Barton et al. 1992;

85
Chapman and Pavelka 2005; Schreier 2010). Future research should investigate the
relationship between herd size and DPL and home range in this band of gelada monkeys.
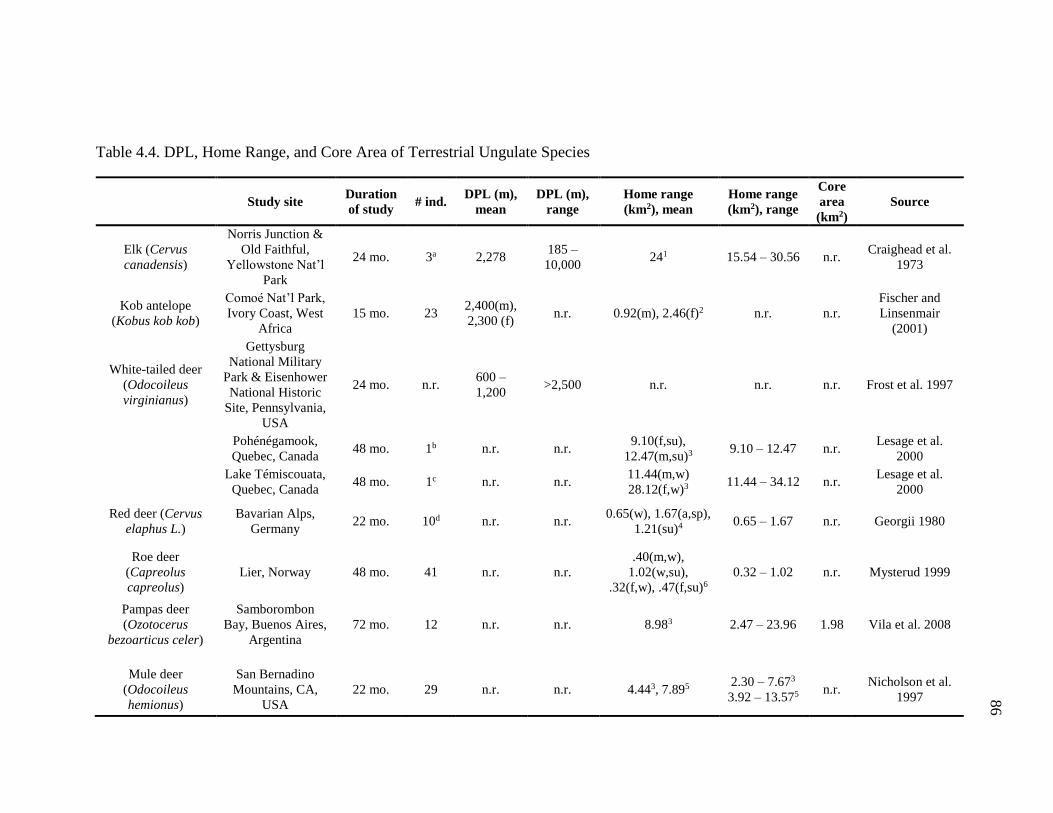
86
Table 4.4. DPL, Home Range, and Core Area of Terrestrial Ungulate Species
Study site Duration
of study # ind.
DPL (m),
mean
DPL (m),
range
Home range
(km2), mean
Home range
(km2), range
Core
area
(km2)
Source
Elk (Cervus
canadensis)
Norris Junction &
Old Faithful,
Yellowstone Nat’l
Park
24 mo. 3a 2,278 185 –
10,000 241 15.54 – 30.56 n.r.
Craighead et al.
1973
Kob antelope
(Kobus kob kob)
Comoé Nat’l Park,
Ivory Coast, West
Africa
15 mo. 23 2,400(m),
2,300 (f) n.r. 0.92(m), 2.46(f)2 n.r. n.r.
Fischer and
Linsenmair
(2001)
White-tailed deer
(Odocoileus
virginianus)
Gettysburg
National Military
Park & Eisenhower
National Historic
Site, Pennsylvania,
USA
24 mo. n.r. 600 –
1,200 >2,500 n.r. n.r. n.r. Frost et al. 1997
Pohénégamook,
Quebec, Canada 48 mo. 1b n.r. n.r.
9.10(f,su),
12.47(m,su)3 9.10 – 12.47 n.r. Lesage et al.
2000
Lake Témiscouata,
Quebec, Canada 48 mo. 1c n.r. n.r.
11.44(m,w)
28.12(f,w)3 11.44 – 34.12 n.r. Lesage et al.
2000
Red deer (Cervus
elaphus L.)
Bavarian Alps,
Germany 22 mo. 10d n.r. n.r.
0.65(w), 1.67(a,sp),
1.21(su)4 0.65 – 1.67 n.r. Georgii 1980
Roe deer
(Capreolus
capreolus)
Lier, Norway 48 mo. 41 n.r. n.r.
.40(m,w),
1.02(w,su),
.32(f,w), .47(f,su)6
0.32 – 1.02 n.r. Mysterud 1999
Pampas deer
(Ozotocerus
bezoarticus celer)
Samborombon
Bay, Buenos Aires,
Argentina
72 mo. 12 n.r. n.r. 8.983 2.47 – 23.96 1.98 Vila et al. 2008
Mule deer
(Odocoileus
hemionus)
San Bernadino
Mountains, CA,
USA
22 mo. 29 n.r. n.r. 4.443, 7.895 2.30 – 7.673
3.92 – 13.575 n.r.
Nicholson et al.
1997
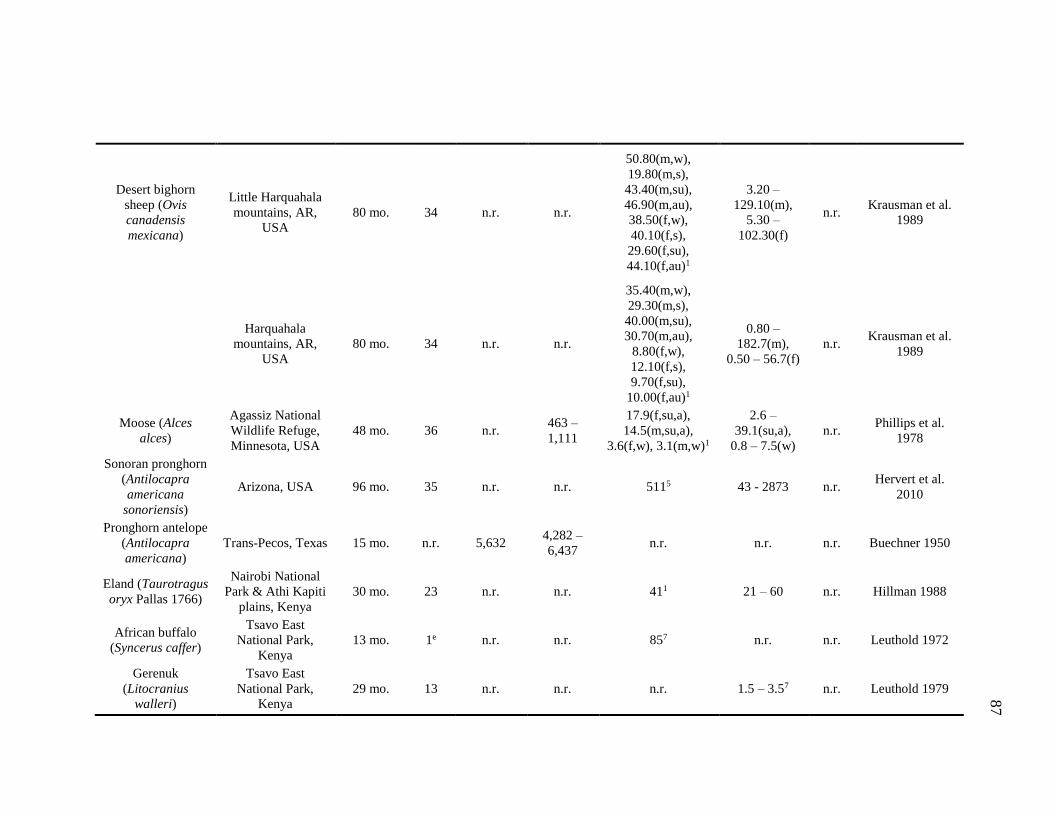
87
Desert bighorn
sheep (Ovis
canadensis
mexicana)
Little Harquahala
mountains, AR,
USA
80 mo. 34 n.r. n.r.
50.80(m,w),
19.80(m,s),
43.40(m,su),
46.90(m,au),
38.50(f,w),
40.10(f,s),
29.60(f,su),
44.10(f,au)1
3.20 –
129.10(m),
5.30 –
102.30(f)
n.r. Krausman et al.
1989
Harquahala
mountains, AR,
USA
80 mo. 34 n.r. n.r.
35.40(m,w),
29.30(m,s),
40.00(m,su),
30.70(m,au),
8.80(f,w),
12.10(f,s),
9.70(f,su),
10.00(f,au)1
0.80 –
182.7(m),
0.50 – 56.7(f)
n.r. Krausman et al.
1989
Moose (Alces
alces)
Agassiz National
Wildlife Refuge,
Minnesota, USA
48 mo. 36 n.r. 463 –
1,111
17.9(f,su,a),
14.5(m,su,a),
3.6(f,w), 3.1(m,w)1
2.6 –
39.1(su,a),
0.8 – 7.5(w)
n.r. Phillips et al.
1978
Sonoran pronghorn
(Antilocapra
americana
sonoriensis)
Arizona, USA 96 mo. 35 n.r. n.r. 5115 43 - 2873 n.r. Hervert et al.
2010
Pronghorn antelope
(Antilocapra
americana)
Trans-Pecos, Texas 15 mo. n.r. 5,632 4,282 –
6,437 n.r. n.r. n.r. Buechner 1950
Eland (Taurotragus
oryx Pallas 1766)
Nairobi National
Park & Athi Kapiti
plains, Kenya
30 mo. 23 n.r. n.r. 411 21 – 60 n.r. Hillman 1988
African buffalo
(Syncerus caffer)
Tsavo East
National Park,
Kenya
13 mo. 1e n.r. n.r. 857 n.r. n.r. Leuthold 1972
Gerenuk
(Litocranius
walleri)
Tsavo East
National Park,
Kenya
29 mo. 13 n.r. n.r. n.r. 1.5 – 3.57 n.r. Leuthold 1979

88
n.r. = not reported
w = winter home range, su = summer home range, sp = spring home range, a = autumn home range
m = adult male, f = adult female aData collected on and presented for cow (female) elk only. bData collected on white-tailed deer from high-density site (Lesage et al. 2000). cData collected on white-tailed deer from low-density site (Lesage et al. 2000). dData collected on and presented for (female) red deer hind only. eData collected on herd of African buffalo. 1Home range estimated with MCP method (did not specify percentage used). 2Home range estimated with 90% MCP method. 3Home range estimated with 95% MCP method. 4Home range estimated with grid-cell method with 200 x 200 m quadrats. 5Home range estimated with 95% adaptive kernel. 6Home range estimated with 90% kernel method (did not specify whether fixed or adaptive or bandwidth used). 7Did not specify method used to estimate home range.

89
Comparison of Gelada Ranging Behavior to Terrestrial Ungulate Species
Geladas are unique among non-human primates in that they are primarily grass
consumers (Crook 1966; Dunbar and Dunbar 1975; Iwamoto 1979; Nguyen and Fashing
2012). Therefore, the graminivorous diet and terrestrial nature of geladas make them an
intriguing species to compare to grazing terrestrial ungulate species in terms of ranging
ecology (e.g., Iwamoto 1979). Earlier work by Iwamoto (1979) described briefly the
similarities and differences in the feeding behavior between geladas and ungulates,
however, his discussion focused primarily on how a diet consisting mainly of grasses
could (continue to) sustain a large population of geladas. To my knowledge, this is the
first study to compare the ranging behavior of gelada monkeys to that of terrestrial
ungulate species (Table 4.4).
Based on the data in this analysis, geladas appear to occupy home ranges similar
to (Bole and Gich) or larger (Sankaber and Guassa) than those reported for various
species of deer, e.g., red deer (Cervus elaphus), roe deer (Capreolus capreolus), white-
tailed deer (Odocoileus virginianus), and rusa deer (Rusa timorensis), and other species
of ungulates, such as the gerenuk (Litocranius walleri) and Kob antelope (Kobus kob
kob). Conversely, ungulates such as the Sonoran pronghorn (Antilocapra americana
sonoriensis), elk (Cervus elaphus), wild eland (Taurotragus oryx Pallas 1766), moose
(Alces alces), gerenuk (Litocranius walleri), and the African buffalo (Syncerus caffer),
occupy home ranges several times larger than those reported for geladas across all study
sites.
Few data are available on the daily movement distances of ungulates. Where such
data do exist for ungulates, e.g., white-tailed deer, kob antelope, and pronghorn antelope,

90
geladas, depending on the site, exhibit day journey lengths that are similar to (Bole and
Gich), longer (Sankaber and Guassa), or shorter (all sites) than those reported for these
species of ungulates (see Table 4.4).
Reports of home range for ungulates are generally derived from observations
made by radio-tracking of individuals (e.g., Lesage et al. 2000; Mysterud 1999; Tufto et
al. 1996). Due to the disparate behavioral ecology of male (e.g., territorial) and female
(e.g., movement in relation to parturition) ungulates and the impact of seasonal patterns
on space use patterns (e.g., migration between winter and summer ranges) (Luccarini et
al. 2006), home ranges are further—and appropriately—categorized by sex (e.g., Hillman
1988; Lesage et al. 2000) or season, such as summer vs. winter ranges, pre-parturition vs.
post-parturition, etc., (Anderson et al. 2005; Girard et al. 2002; Lesage et al. 2000), and in
some cases, based on the movements of entire groups (herd) (Hervert et al. 2010;
Leuthold 1972; Luccarini et al. 2006). Furthermore, where the annual or total home range
estimate was not reported (e.g., Anderson et al. 2005; Lesage et al. 2000), space use
patterns were described using seasonal home ranges. In some cases, home range sizes for
juveniles or younglings were also reported (e.g., Hillman 1988; Lesage et al. 2000)—
however, I did not include these estimates in the analysis. The use of seasonal home
ranges or home ranges based on behavioral or reproductive events (when an annual home
range was not reported) often made it difficult to compare to the home ranges estimated
for the geladas. Despite the difficulties with the home range estimates I encountered
above, the use of the 95% MCP and fixed kernel (LSCV bandwidth in many cases)
methods in studies of ungulate ranging ecology facilitated direct comparisons of home
range estimates in those studies to this study (but not with estimates reached by Dunbar

91
and others due to methodological differences). In some cases, ungulate researchers (e.g.,
Mysterud 1999; Tufto et al. 1996) estimated kernel home ranges using the 90% density
contour; however, I did not consider the 5% difference to have a major impact on
comparisons (Börger et al. 2006; Seaman and Powell 1996).
Implications of Inhabiting in a Topographically Variable Environment on Calculations
of Distance Traveled
One goal of this study was to assess how living in a topographically variable
environment would affect estimates of distance traveled. I have shown mathematically
that movement across an altitudinal gradient can result in longer DPLs by up to 8% (a
mean increase of up to 3% was found for monthly mean DPL). This finding corroborates
an earlier conclusion reached by Sprague (2000) who found similar, though higher
increases (mean: 9.5%; range: 2.5 – 21.5%) in the DPL of the Yaku monkey (Macaca
fuscata yakui) at Kirishima-Yaku National Park, Japan, after he accounted for changes in
altitude. Prior reports by Whiten et al. (1987) and Hunter (2001), and more recently Niu
et al. (2010), also investigated the effects of topography on distance traveled in chacma
baboons (P. ursinus), geladas (at Sankaber), and Guizhou snub-nosed monkeys
(Rhinopithecus brelichi), respectively; however, these authors did not specify the amount
of percentage difference they found between the original and corrected DPLs, which
makes it difficult to make any meaningful comparisons. Furthermore, another factor
limiting comparability of results is the manner in which each study assessed the impact of
change in altitude on DPL. This study and Whiten et al.’s (1987), for instance, utilized
Pythagora’s theorem of the three sides of a right triangle to account for the net change in
elevation between consecutive readings, though in Whiten et al.’s (1987) study, vertical
travel was calculated based on the number of 50 m contour lines the animals passed

92
through (whereas we found the exact change in altitude by subtracting the altitude
reading between consecutive path lengths: see Methods). Alternatively, Sprague (2000)
and Niu et al. (2010) used the mean slope angle of their respective study sites to examine
the influence of topography on movement. Lastly, Hunter (2001) combined the distance
the geladas traveled horizontally and the distance they traveled vertically (e.g., Whiten et
al. 1987) to determine the observed difference in altitude for individual day journey
lengths. The degree of difference between the original and corrected DPL readings
reported in each study, therefore, may simply be a by-product of the differences inherent
in each method. Despite the different methods employed in each study, the results are
clear: topography can have a small, but noticeable effect on estimates of distance traveled
and should therefore be accounted for whenever possible.
Ecological Implications of Movement Across Uneven Topography
Nonhuman primates, such as geladas (Ashenafi 2001; Dunbar 1998), some
populations of chacma baboons (Henzi et al. 1992; Whiten et al. 1987), Japanese
macaques (Wada and Ichiki 1989), black-crested gibbons (Fan and Jiang 2010), and
snub-nosed monkeys (Kirkpatrick et al. 1999; Li et al. 2008; Long et al. 1994) live in
high altitude, montane environments characterized by cold temperatures, strongly
seasonal weather patterns, and wide temporal variability of resources (Ashenafi 2001;
Hanya et al. 2003; Li and Walker 1986; Nguyen and Fashing 2012; Whiten et al. 1987).
In spite of the documented effects of temperature and climate patterns on energy
expenditure and thermoregulation (Bronikowski and Altmann 1996; Fan and Jiang 2010;
Henzi et al. 1992; Hill 2006; Iwamoto and Dunbar 1983; Stelzner 1988; Wada et al.
2007; Yang 2003), these animals may traverse across altitudinal gradients of more than

93
30 m on a daily basis to satisfy their basic biological needs, e.g., acquire food, return to
sleeping sites, etc. (Dunbar and Dunbar 1975; Fan and Jiang 2010; Hanya et al. 2003;
Kawai and Iwamoto 1979; Niu et al. 2010; Whiten et al. 1987; Yang 2003; Moua unpubl.
data). Furthermore, not only does upslope movement result in longer (than expected)
DPLs (Sprague 2000; this study), but an animal moving across uneven topography
expends relatively more energy (related to locomotion) than it would if moving across
horizontal or downslope landscape (researched in small ruminants and grazing mammals:
Dailey and Hobbs 1989; Lachica and Aguilera 2000; Lachica et al. 1997; reviewed by
Lachica and Aguilera 2005). It is therefore conceivable that the cumulative effects of
upslope travel, longer travel distances, and thermoregulatory responses in relation to
existing temperature and weather conditions can be expected to impose (considerable)
energetic constraints on an animal’s overall energy expenditure and behavioral ecology.
In their investigation of the altitudinal ranging patterns of Guizhou snub-nosed
monkeys at Fanjinshan National Nature Reserve, China, Niu et al. (2010) uncovered that
the monkeys would travel from lower to higher elevations to feed and then return to
lower elevations to sleep at night. This type of ‘oscillatory’ (Niu et al. 2010: 241)
behavior in movement is similar to the ranging behavior reported for geladas (Dunbar
and Dunbar 1975; Kawai and Iwamoto 1979; Moua unpubl. data) and possibly chacma
baboons (Henzi et al. 1992; Stoltz and Saayman 1970), where the animals sleep alongside
cliff edges down on the slopes below but conduct the rest of their daily activities on
higher elevation above the sleeping sites. Fittingly, Niu et al. (2010) have questioned why
nonhuman primates instead simply adopt a routine where the animals ‘sleep where they
eat’ (Niu et al. 2010: 241) to minimize traveling up and down slopes and therefore

94
conserve energy. Niu et al. (2010) have suggested that such altitudinal movement patterns
in the Guizhou snub-nosed monkey mark a trade-off between acquiring resources (in the
higher elevations where food is more abundant) and avoiding predators (in the lower
elevations where the animals can hide in the dense foliage and where snowfall and
temperatures are thwarted by the canopy cover). Perhaps this trade-off of increased
energy costs related to movement across an altitudinal gradient and reduced predation
risk may also explain the similar up and down movement patterns and use of sleeping
cliffs in geladas (and maybe chacma baboons).
Though the trade-off between resource acquisition and anti-predatory defense
may potentially explain why nonhuman primates such as geladas and Guizhou snub-
nosed monkeys exhibit such energy costly movement patterns over the course of a day,
the trade-off explanation does not explain the energetic costs the monkeys experience at
the time of their of upslope travels, or the impact of current environmental conditions on
their energy budget (i.e., thermoregulatory responses). For example, black-crested
gibbons studied at Mt. Wuliang, central Yunnan, China, tended to range at lower
elevations in the morning when it was cold but moved to higher elevations in the
afternoon when temperatures got warmer (Fan and Jiang 2010). Alternatively, during
snow storms, for example, Japanese macaques (Macaca fuscata) in the Shiga Heights,
Japan, huddled closely to one another in order to conserve energy and create heat (Wada
et al. 2007). Lastly, laboratory experiments using rats showed that the animals increased
energy intake and metabolic activity (e.g., become more active or mobile) during low
temperatures in order to provide the body with (additional) energy to create heat to
reduce hypothermia (Brobeck 1948). These findings indicate that animals may adopt or

95
develop a variety of responses to conserve energy or maintain optimal body temperature
under energetically costly situations. It is in this regard that I discuss the ecological
implications of the ‘shuffle’ technique in gelada monkeys (Wrangham 1980).
In his study of gelada monkey ranging behavior, Wrangham (1980) noted that
geladas often shuffle while feeding and that shuffling accounted for between 14-30% of
actual distance traveled per day (cited in Iwamoto and Dunbar 1983). During a ‘shuffle’,
geladas slide their hind legs back and forth with the legs maintaining continuous contact
with the ground (Hunter 2001). It therefore appears that geladas should ‘shuffle’ in
situations where they only wish to move a short distance, e.g., within a food patch,
whereas quadruped locomotion should be used in circumstances where long(er) distance
movement is required or where speed is a factor (e.g., running away from a predator). If
this assumption is correct, it implies that a ‘shuffle’ would be energetically more efficient
than quadruped locomotion, otherwise it would be impractical from an evolutionary and
energetic perspective to ‘shuffle’ when moving on all four limbs covers more ground for
the same amount of energy used. Therefore, I hypothesize that the ‘shuffle’ technique in
geladas could have evolved as a more (energetically) efficient form of short movement
bout in an environment characterized by cold temperatures and low nutrient quality
resources. However, further more detailed data on the movement behavior of geladas,
such as when geladas shuffle and when they use quadruped location and the ecological
context in which each behavior was observed, are required in order to fully test this
hypothesis.

96
Critiques of Altitudinal Change Formula
There are a couple shortcomings associated with the altitudinal change formula
applied here that warrant mention. First, the exact geographic location of the herd was
recorded only once every half-hour. Since it is conceivable the geladas could have
traveled multiple times upslope or downslope between subsequent half-hour readings, I
cannot know for certain the exact change in elevation that occurred between subsequent
readings, and it is therefore possible that the estimates of half-hour path lengths (and
ultimately DPL) may have been underestimated. Moreover, the monkeys could have
moved exactly the same distance upslope and downslope between consecutive half-hour
readings, thereby resulting in a net change in altitude of 0 m when in fact movement
across an altitudinal cline had occurred. In fact, the mean net change in altitude between
the very first and last readings (the difference in elevation between the morning and
evening sleeping site locations, respectively) on the same full-day was only +5 m, but I
was able to demonstrate the high degree of movement across uneven topography in our
study group of geladas (Moua unpub. data). The finding that the mean net change in
elevation on a daily basis was +5 m suggests it may be important to record the animals’
whereabouts in a time frame capable of capturing their movement across uneven
landscape, because of the possibility that such (small) changes in elevation may be lost in
consecutive readings with longer time intervals. Despite the simplicity of the formula, the
corrected path length values obtained as a result of altitudinal effects nonetheless provide
an estimate that reflects the influence of change in altitude on distance traveled. The
findings presented here suggest that the conventional method of calculating DPL may be
unsuitable for study sites where the topography is heterogeneous or when a percentage of

97
the animals’ movement involves upslope or downslope travel, because of the likelihood
that path lengths and DPL will be underestimated (Sprague 2000). Thus, I advise that
future studies of primates and other terrestrial animals that range over rugged terrain
consider the influence of elevation and topographic variation when calculating movement
parameters.
Conclusions
The continual expansion of the annual home range reported in this study has
considerable implications for the conservation and management of the geladas and their
natural habitat at Guassa (and for geladas elsewhere), and further demonstrates the
importance of long-term monitoring in wild animal populations. The increasing trend in
annual home range suggests that one or two years of observation would not have
provided sufficient data to make an accurate conclusion about the geographic extent of
the geladas’ range at Guassa. Moreover, this pattern also indicates that the home range
estimates reported for the gelada populations at Sankaber, Gich, and Bole (Dunbar and
Dunbar 1974, 1975; Hunter 2001; Kawai and Iwamoto 1979), which reflect data gathered
from a few weeks to a single annual cycle, may not be entirely representative of the
animals’ space use patterns over a longer period of time (i.e., it is possible that home
range size may be larger than what was reported at these sites had the animals been
observed for a longer period of time).
Furthermore, most, if not all, of the expansions in the annual range over the five-
year period have occurred primarily in the southern and western regions of the Guassa
area. Though the geladas are understandably unable to expand any further east because of
the cliff edges that are situated along this side of their range, movement into the northern

98
region of the range, though plausible, has been rare (except for two unusually far
excursions in May and August of 2010). Why the geladas seldom range in this region
remains unknown, though a likely explanation may have to do with the local inhabitants
who reside in this area. During both trips to this region in 2010, for example, researchers
were unable to remain with the geladas because it was part of a different administrative
unit in which the study team did not have permission to work. Therefore, it is presently
unknown the type of relationship that exists between the locals in this region and the
geladas. Similarly, recent studies by Li et al. (2008, 2010) on the ranging ecology of the
Yunnan snub-nosed monkeys (R. bieti) and the black-and-white snub nosed monkeys (R.
bieti) at Samage Forest, Baimaxue-shan National Nature Reserve, Yunnan, China, found
evidence to suggest that human encroachment and disturbance into the surrounding
habitat may be limiting the monkeys’ ability to expand their home range and completely
avoiding ranging in areas disturbed by humans.
For geladas and animals in general, the restriction of range expansion or space use
related to human activity may present a serious (and compounding) issue for the integrity
of the species and their natural habitat going into the future. Computer simulations
developed by Dunbar (1998), for example, demonstrate the potential implications of
rising world temperatures on the distribution and viability of grasslands throughout the
geladas’ home ranges in the northern Ethiopian Highlands. Human encroachment and
agricultural cultivation have already penetrated other gelada monkey study sites (e.g.,
Sankaber, Gich, and Bole: Dunbar and Dunbar 1974, 1975; Hunter 2001; Kawai 1979),
while Guassa remains relatively intact ecologically in large part because of the (Qero)
conservation system put in place there hundreds of years ago (Ashenafi 2001; but see

99
Ashenafi and Leader-Williams 2005 for the implications of the recent change in
management regime for the Guassa area). Should rising world temperatures result in the
demise of the gelada monkeys’ grassland ecosystem as Dunbar (1998) projected, and
coupled with continuing human encroachment, the monkeys can be expected to be
pushed to higher elevations with far reduced available habitat (Dunbar 1998). I advise
researchers to continue monitoring the ranging behavior and habitat quality of gelada
monkeys at these study sites and at other locations where such observations are possible.
Doing so should provide valuable data about the monkeys’ space use patterns (over an
extended period) in an environment that is likely to change due to global climate change
and human activity.
In sum, I advise that ongoing and future studies of animal ranging ecology
attempt to invest several years of continuous observation for the highest possibility of
acquiring sufficient data about the space use patterns of animals in relation to ecological
variability across space and time. The information obtained from these studies can prove
crucial in helping us make informed conservation and management-related decisions.

100
APPENDIX A
ADDING ERROR TO USER-IDENTIFIED DUPLICATE PAIRS
Using a kernel bandwidth, such as the least-squares cross validation (LSCV), to
analyze a utilization distribution with duplicate data or data that clump can cause it to fail
(Beyer pers. comm.; Gitzen et al. 2006; Tufto et al. 1996). Some authors suggest (e.g.,
Beyer pers. comm.; Rogers et al. 2007) that adding error to duplicate or clumped data can
resolve bandwidth issues associated with duplicate or clumped data, however, little is
known about the ramifications of adding error to duplicate data (Rogers et al. 2007). In
accordance with the recommendations of several researchers, I added error to duplicate
coordinate pairs I identified using the procedure below to address the issues associated
with duplicate data on kernel home range analysis.
I used Microsoft 2010 to organize, sort, identify, and add error all duplicate data. I
outline this procedure below.
Step 1: Identifying Duplicates in the Lat Coordinates First, I sorted the Lat
coordinates into ascending order. (Ensure the corresponding Lon coordinate stays with its
corresponding Lat coordinate.) Next, I implemented Equation 1 to identify all Lat
coordinates that are a duplicate. Equation 1 compares a specified Lat coordinate to the
Lat coordinate in the cell directly above and below it and expresses a value to indicate if a
duplicate does or does not exist.
= 𝐼𝐹(𝑂𝑅(𝐸[𝑥] = 𝐸[𝑥 − 1], 𝐸[𝑥] = 𝐸[𝑥 + 1]), 𝑎, 𝑏) Equation 1

101
, where E[x] is the cell number of the specified Lat coordinate; E[x-1] represents the cell
number of the Lat coordinate immediately before (above) the specified Lat coordinate;
E[x+1] represents the cell number of the Lat coordinate immediately after (below) the
specified Lat coordinate; a and b denote a value (≥0) (chosen by the user) to indicate
whether the “if_or” statement is true or false, respectively (Figure 1).
Figure 1. Hypothetical example showing how Equation 1 is being applied.
Upon the completion of Equation 1, I immediately copied the output values in the
“Lat duplicate?” column (Column C, Figure 1) and (re)pasted the “Values” over the
originals, essentially eliminating the formula and leaving only the value in the cell. (This
is critical because it ensures that each Lat coordinate retains its proper identification of
“1” or “0” even after they get rearranged in Step 2.)
Step 2: Identifying Duplicates in the Lon Coordinates. Similarly, I used Equation
1 to identify duplicates in the Lon coordinates (Figure 2). After the duplicates for the Lon
coordinates were identified, I copied the output values in the “Lon duplicate?” column
(Column E, Figure 2) and (re)pasted the “Values” over the originals.
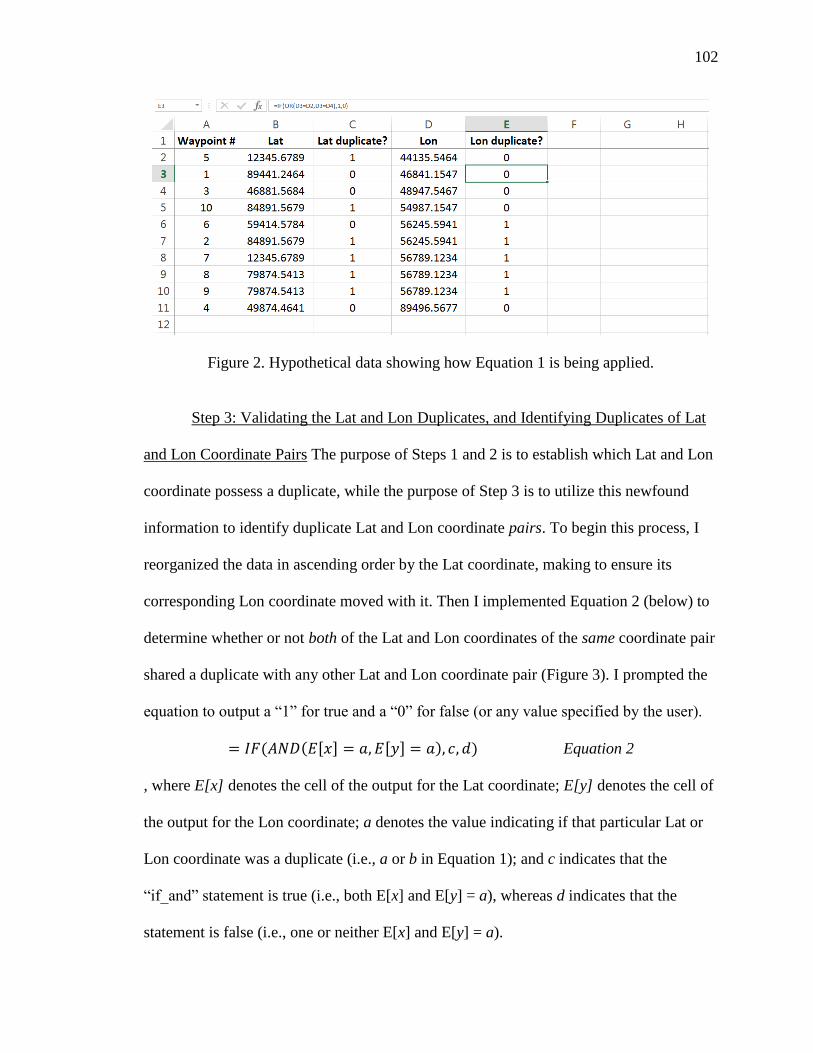
102
Figure 2. Hypothetical data showing how Equation 1 is being applied.
Step 3: Validating the Lat and Lon Duplicates, and Identifying Duplicates of Lat
and Lon Coordinate Pairs The purpose of Steps 1 and 2 is to establish which Lat and Lon
coordinate possess a duplicate, while the purpose of Step 3 is to utilize this newfound
information to identify duplicate Lat and Lon coordinate pairs. To begin this process, I
reorganized the data in ascending order by the Lat coordinate, making to ensure its
corresponding Lon coordinate moved with it. Then I implemented Equation 2 (below) to
determine whether or not both of the Lat and Lon coordinates of the same coordinate pair
shared a duplicate with any other Lat and Lon coordinate pair (Figure 3). I prompted the
equation to output a “1” for true and a “0” for false (or any value specified by the user).
= 𝐼𝐹(𝐴𝑁𝐷(𝐸[𝑥] = 𝑎, 𝐸[𝑦] = 𝑎), 𝑐, 𝑑) Equation 2
, where E[x] denotes the cell of the output for the Lat coordinate; E[y] denotes the cell of
the output for the Lon coordinate; a denotes the value indicating if that particular Lat or
Lon coordinate was a duplicate (i.e., a or b in Equation 1); and c indicates that the
“if_and” statement is true (i.e., both E[x] and E[y] = a), whereas d indicates that the
statement is false (i.e., one or neither E[x] and E[y] = a).

103
It is important to organize the data in ascending order relative to the Lat
coordinate, not by the Waypoint #, as this will facilitate identifying duplicates.
Furthermore, at this juncture the data of greatest import are those in the “Lat+Lon
duplicate?” column with the “1,” which indicate that the Lat and Lon coordinates (may)
share a duplicate with another coordinate pair. The word “may” is used here because,
according to Figure 3, even though both the Lat and Lon coordinates in Waypoint #7
share duplicates, Waypoint #7 is the only one of its kind; in other words, even though the
Lat coordinate of Waypoint #7 shares a duplicate with the Lat coordinate of Waypoint
#5, the corresponding Lon coordinate of each is different, which means the coordinate
pairs of Waypoints #7 and #5 do not share any duplicates with any other coordinate pair.
Waypoints #2 and #10 paint a similar situation.
It is critical to be fully aware of instances such as this one because failure to pay
careful attention may result in accidentally, and needlessly, adding error when there is no
need. Indeed, this means that the user will need to use the information in the “Lat+Lon
duplicate?” column to manually identify duplicate coordinate pairs and add error to any
and all such duplicate pairs (Step 4).
Figure 3. Hypothetical data showing how Equation 2 is being applied.

104
Step 4: Adding Error to User-Identified Duplicate Coordinate Pairs I added error
in increments of two meters) to both the Lat and Lon coordinates of each duplicate
coordinate pair, beginning with the second duplicate coordinate pair. (I always left one
duplicate coordinate pair in its original state.) To elaborate, if a Lat and Lon coordinate
pair had a total of three duplicates, I (i) left one of the three duplicates in its original state;
(ii) then added two m of error to the Lat and Lon coordinate of the second of three
duplicates; and (iii) lastly added four m of error to the Lat and Lon coordinate of the
remaining duplicate pair. (Similarly, I added six m of error would be added to the fourth
duplicate pair, eight m of error would added to the fifth duplicate pair, etc.) I continued
this process of adding error to each of the remaining duplicate coordinate pairs until error
had been added to all user-identified duplicate coordinate pairs. The most random error
added this way was 20 m (most likely to a sleeping cliff coordinate value as the geladas
regularly re-used sleeping cliff sites throughout the five-year study). The new coordinate
pairs will then replace the original coordinate pairs.
Step 5: Re-analysis of data for (inadvertent) duplicate data It is quite conceivable
that coordinate data may be inadvertently duplicated during the error adding phase of
Step 4. This is plausible because the user, by adding error in the form of a distance,
changes the makeup of each coordinate pair, creating a new Lat and Lon that may in turn
share the same value as another Lat and Lon coordinate pair. To ensure this phenomenon
of creating inadvertent duplicate data does not happen, I repeated Steps 1 through 3 to
identify any duplicate data, and where there were duplicate pairs, I added error to them. I
continued this process (of rearranging and adding random error) until there were no
longer any duplicate pairs in the dataset.

105
BIBLIOGRAPHY
Albernaz A, and Magnusson W. 1999. Home-range size of the bare-ear marmoset
(Callithrix argentata) at Alter do Chão, Central Amazonia, Brazil. International
Journal of Primatology 20(5):665-677.
Albon SD, and Langvatn R. 1992. Plant phenology and the benefits of migration in a
temperate ungulate. Oikos 65(3):502-513.
Amstrup SC, McDonald TL, and Durner GM. 2004. Using satellite radiotelemetry data to
delineate and manage wildlife populations. Wildlife Society Bulletin
32(3)(Autumn 2004):661-679.
Anderson C. 1981. Intertroop relations of chacma baboon (Papio ursinus). International
Journal of Primatology 2(4):285-310.
Anderson CM. 1982. Baboons below the tropic of capricorn. Journal of Human Evolution
11(3):205-217.
Anderson D. 1982b. The home range: A new nonparametric estimation technique.
Ecology 63(1):103-112.
Anderson DP, Forester JD, Turner MG, Frair JL, Merrill EH, Fortin D, Mao JS, and
Boyce MS. 2005. Factors influencing female home range sizes in elk (Cervus
elaphus) in North American landscapes. Landscape Ecology 20(3):257-271.
Andreka G, Linn I, Perrine M, and Maddock A. 1999. Range use by the wild dog in the
Hluhluwe-Umfolozi Park, South Africa. South African Journal of Wildlife
Research 29(1):1-9.
Asensio N, Schaffner CM, and Aureli F. 2012. Variability in core areas of spider
monkeys (Ateles geoffroyi) in a tropical dry forest in Costa Rica. Primates
53(2):147-156.
Ashenafi ZT. 2001. Common Property Resource Management of an Afro-Alpine Habitat
Supporting a Population of the Critically Endangered Ethiopian Wolf (Canis
simensis). Ph.D. Thesis. University of Kent, Canterbury, UK.

106
Ashenafi ZT, and Leader-Williams N. 2005. Indigenous common property resource
management in the Central Highlands of Ethiopia. Human Ecology 33(4):539-
563.
Baldwin PJ, McGrew WC, and Tutin CEG. 1982. Wide-ranging chimpanzees at Mt.
Assirik, Senegal. International Journal of Primatology 3(4):367-385.
Baoping R, Ming L, Yongcheng L, and Fuwen W. 2009. Influence of day length, ambient
temperature, and seasonality on daily travel distance in the Yunnan snub-nosed
monkey at Jinsichang, Yunnan, China. American Journal of Primatology
71(3):233-241.
Barton R, Whiten A, Strum S, Byrne R, and Simpson A. 1992. Habitat use and resource
availability in baboons. Animal Behaviour 43(5):831-844.
Basabose AK. 2005. Ranging patterns of chimpanzees in a montane forest of Kahuzi,
Democratic Republic of Congo. International Journal of Primatology 26(1):33-54.
Beehner J, Berhanu G, and McCann C. 2008. Population estimate for geladas
(Theropithecus gelada) living in and around the Simien Mountains National Park,
Ethiopia. SINET 30:149-154.
Bekoff M, and Mech LD. 1984. Simulation analyses of space use: Home range estimates,
variability, and sample size. Behavior Research Methods, Instruments, &
Computers 16(1):32-37.
Berger J, Stacey PB, Bellis L, and Johnson MP. 2001. A mammalian predator-prey
imbalance: Grizzly bear and wolf extinction affect Avian neotropical migrants.
Ecological Applications 11(4):947-960.
Bergerud A, Wyett W, and Snider B. 1983. The role of wolf predation in limiting a
moose population. The Journal of Wildlife Management:977-988.
Beyer HL. 2012a. Geospatial Modelling Environment. 0.7.2.0 ed.
Beyer HL. 2012b. Geospatial Modelling Environment Help PDF.
Biebouw K. 2009. Home range size and use in Allocebus trichotis in Analamazaotra
special reserve, central eastern Madagascar. International Journal of Primatology
30(2):367-386.
Blundell G, Maier J, and Debevec E. 2001. Linear home ranges: effects of smoothing,
sample size, and autocorrelation on kernel estimates. Ecological Monographs
71(3):469-489.

107
Boesch C, and Achermann HB. 2000. The chimpanzees of the Taï forest: behavioural
ecology and evolution: New York, Oxford University Press, USA.
Boesch C, and Boesch H. 1989. Hunting behavior of wild chimpanzees in the Tai
National Park. American journal of physical anthropology 78(4):547-573.
Börger L, Franconi N, De Michele G, Gantz A, Meschi F, Manica A, Lovari S, and
Coulson T. 2006. Effects of sampling regime on the mean and variance of home
range size estimates. The Journal of Animal Ecology 75(6):1393-1405.
Boulanger J, and White G. 1990. A comparison of home-range estimators using Monte
Carlo simulation. The Journal of Wildlife Management 54(2):310-315.
Bowman A, and Azzalini A. 1997. Applied Smoothing Techniques for Data Analysis:
The Kernel Approach with S-Plus Illustrations. Oxford: Clarendon Press.
Boyle SA, Lourenço WC, Da Silva LR, and Smith AT. 2009. Home range estimates vary
with sample size and methods. Folia Primatologica 80(1):33-42.
Brobeck JR. 1948. Food intake as a mechanism of temperature regulation. The Yale
Journal of Biology and Medicine 20(6):545.
Bronikowski AM, and Altmann J. 1996. Foraging in a variable environment: weather
patterns and the behavioral ecology of baboons. Behavioral Ecology and
Sociobiology 39(1):11-25.
Buechner HK. 1950. Life history, ecology, and range use of the pronghorn antelope in
Trans-Pecos Texas. American Midland Naturalist: 257-354.
Burgman M, and Fox J. 2003. Bias in species range estimates from minimum convex
polygons: implications for conservation and options for improved planning.
Animal Conservation 6:19-28.
Carnegie S, Fedigan L, and Melin A. 2011. Reproductive seasonality in female capuchins
(Cebus capucinus) in Santa Rosa (Area de Conservación Guanacaste), Costa Rica.
International Journal of Primatology 32(5):1076-1090.
Chapman CA. 1988. Patterns of foraging and range use by three species of neotropical
primates. Primates 29(2):177-194.
Chapman CA, and Chapman L. 2000. Determinants of group size in primates: the
importance of travel costs. In: Boinski S, and Garber P, editors. On the Move:
How and Why Animals Travel in Groups. Chicago and London: The University
of Chicago Press. p 24-41.

108
Chapman CA, and Pavelka MM. 2005. Group size in folivorous primates: ecological
constraints and the possible influence of social factors. Primates 46(1):1-9.
Chism J, and Rowell T. 1988. The natural history of patas monkeys. In: Gautier-Hion A,
Bourliére F, Gautier J, and Kingdon J, editors. Primate Radiation: Evolutionary
Biology of the African Guenons. Cambridge: Cambridge University Press. p 412-
438.
Clutton-Brock TH, and Harvey PH. 1977. Primate ecology and social organization.
Journal of Zoology 183(1):1-39.
Cohen J. 1988. Statistical power analysis for the behavioral sciences. Hillsdale, New
Jersey. Lawrence Erlbaum Associates, Publishers.
Covert H, Quyet L, and Wright B. 2008. On the brink of extinction: Research for the
conservation on the Tonkin snub-nosed monkey (Rhinopithecus avunculus). In:
Fleagle J, and Gilbert C, editors. Ewyln Simons: A Search for Origins: Springer-
Verlag New York. p 409-427.
Cowlishaw G, and Dunbar RI. 2000. Primate Conservation Biology: University of
Chicago Press. Chicago, USA.
Craighead JJ, Craighead FC, Ruff RL, and O'Gara BW. 1973. Home ranges and activity
patterns of nonmigratory elk of the Madison drainage herd as determined by
biotelemetry. Wildlife Monographs (33):3-50.
Crook JH. 1966. Gelada baboon herd structure and movement: a comparative report.
Symp Zool Soc Lond. p 237-258.
Crook JH, and Aldrich-Blake P. 1968. Ecological and behavioural contrasts between
sympatric ground dwelling primates in Ethiopia. Folia Primatologica 8(3-4):192-
227.
Dagg AI, and Foster JB. 1976. The giraffe: its biology, behavior, and ecology: Van
Nostrand Reinhold Company. New York.
de Solla SR, Bonduriansky R, and Brooks RJ. 1999. Eliminating autocorrelation reduces
biological relevance of home range estimates. Journal of Animal Ecology
68(2):221-234.
Dew JL, and Wright P. 1998. Frugivory and seed dispersal by four species of primates in
Madagascar's eastern rain forest. Biotropica 30(3):425-437.
Dias L, and Strier K. 2003. Effects of group size on ranging patterns in Brachyteles
arachnoides hypoxanthus. International Journal of Primatology 24(2):209-221.

109
Dobrovidov AV, and Ruds’ko IM. 2010. Bandwidth selection in nonparametric estimator
of density derivative by smoothed cross-validation method. Autom Remote
Control 71(2):209-224.
Donaldson BM, and Echternacht AC. 2005. Aquatic habitat use relative to home range
and seasonal movement of eastern box turtles (Terrapene carolina carolina:
Emydidae) in eastern Tennessee. Journal of Herpetology 39(2):278-284.
Doran D. 1997. Influence of Seasonality on Activity Patterns, Feeding Behavior,
Ranging, and Grouping Patterns in Taï Chimpanzees. International Journal of
Primatology 18(2):183-206.
Doran-Sheehy D, Mongo P, Lodwick J, and Conklin-Brittain NL. 2009. Male and female
western gorilla diet: Preferred foods, use of fallback resources, and implications
for ape versus old world monkey foraging strategies. American Journal Physical
Anthropology 140(4):727-738.
Dunbar R. 1977. Feeding ecology of gelada baboons: a preliminary report. In: Clutton-
Brock T, editor. Primate Ecology: Studies of feeding and ranging behaviour in
lemurs, monkeys and apes. London, New York, San Francisco: Academic Press. p
251-273.
Dunbar R. 1980. Demographic and life history variables of a population of gelada
baboons (Theropithecus gelada). The Journal of animal ecology:485-506.
Dunbar R, and Dunbar P. 1974. Ecological relations and niche separation between
sympatric terrestrial primates in Ethiopia. Folia Primatologica 21(1):36-60.
Dunbar RIM. 1998. Impact of global warming on the distribution and survival of the
gelada baboon: a modelling approach. Global Change Biology 4(3):293-304.
Dunbar RIM, and Dunbar P. 1975. Social Dynamics of Gelada Baboons: S. Karger.
Duong T, and Hazelton ML. 2005. Cross-validation bandwidth matrices for multivariate
kernel density estimation. Scandinavian Journal of Statistics 32(3):485-506.
Edwards MA, Nagy JA, and Derocher AE. 2009. Low site fidelity and home range drift
in a wide-ranging, large Arctic omnivore. Animal Behaviour 77(1):23-28.
ESRI. 2011. ArcGIS Desktop: Release 10.1. Redlands, CA: Environmental Systems
Research Insititute.
Fashing P. 2001. Activity and ranging patterns of guerezas in the Kakamega Forest:
Intergroup variation and implications for intragroup feeding competition.
International Journal of Primatology 22(4):549-577.

110
Fashing P, Nguyen N, and Fashing N. 2010. Behavior of geladas and other endemic
wildlife during a desert locust outbreak at Guassa, Ethiopia: ecological and
conservation implications. Primates 51(3):193-197.
Fashing PJ, Mulindahabi F, Gakima J-B, Masozera M, Mununura I, Plumptre AJ, and
Nguyen N. 2007. Activity and ranging patterns of Colobus angolensis ruwenzorii
in Nyungwe Forest, Rwanda: possible costs of large group size. International
Journal of Primatology 28(3):529-550.
Fashing PJ, Nguyen N, Barry TS, Goodale CB, Burke RJ, Jones SCZ, Kerby JT, Lee LM,
Nurmi NO, and Venkataraman VV. 2011. Death among geladas (Theropithecus
gelada): a broader perspective on mummified infants and primate thanatology.
American Journal of Primatology 73(5):405-409.
Fashing PJ, Nguyen N, Venkataraman VV, and Kerby JT. 2014. Gelada feeding ecology
in an intact ecosystem at Guassa, Ethiopia: variability over time and implications
for theropith and hominin dietary evolution. American journal of physical
anthropology 155(1):1-16.
Ferguson SH, Taylor MK, Born EW, Rosing-Asvid A, and Messier F. 1999.
Determinants of home range size for polar bears (Ursus maritimus). Ecology
Letters 2(5):311-318.
Fieberg J. 2007. Kernel density estimators of home range: smoothing and the
autocorrelation red herring. Ecology 88(4):1059-1066.
Field CR. 1971. Elephant ecology in the Queen Elizabeth National Park, Uganda. African
Journal of Ecology 9(1):99-123.
Fischer F, and Linsenmair KE. 2001. Spatial and temporal habitat use of kob antelopes
(Kobus kob kob, Erxleben 1777) in the Comoé National Park, Ivory Coast as
revealed by radio tracking. African Journal of Ecology 39(3):249-256.
Frost HC, Storm GL, Batcheller MJ, and Lovallo MJ. 1997. White-tailed deer
management at Gettysburg National Military Park and Eisenhower National
Historic Site. Wildlife Society Bulletin:462-469.
Garber P. 1980. Locomotor behavior and feeding ecology of the Panamanian tamarin
(Saguinus oedipus geoffroyi, Callitrichidae, Primates). International Journal of
Primatology 1(2):185-201.
Georgii B. 1980. Home range patterns of female red deer (Cervus elaphus L.) in the Alps.
Oecologia 47(2):278-285.

111
Gerber BD, Arrigo-Nelson S, Karpanty SM, Kotschwar M, and Wright PC. 2012. Spatial
ecology of the endangered Milne-Edwards’ sifaka (Propithecus edwardsi): Do
logging and season affect home range and daily ranging patterns? International
Journal of Primatology: 1-17.
Girard I, Ouellet J-P, Courtois R, Dussault C, and Breton L. 2002. Effects of sampling
effort based on GPS telemetry on home-range size estimations. The Journal of
Wildlife Management 66(4):1290-1300.
Gitzen RA, Millspaugh JJ, and Kernohan BJ. 2006. Bandwidth selection for fixed-kernel
analysis of animal utilization distributions. Journal of Wildlife Management
70(5):1334-1344.
Glessner KD, and Britt A. 2005. Population density and home range size of Indri indri in
a protected low altitude rain forest. International Journal of Primatology
26(4):855-872.
Gravetter FJ, and Wallnau LB. 2008. Essentials of statistics for the behavioral sciences:
Wadsworth Publishing Company. Belmont, CA, USA.
Grueter C, Li D, Ren B, and Wei F. 2009. Choice of analytical method can have dramatic
effects on primate home range estimates. Primates 50(1):81-84.
Grueter CC, Li D, van Schaik CP, Ren B, Long Y, and Wei F. 2008. Ranging of
Rhinopithecus bieti in the Samage Forest, China. I. Characteristics of range use.
International Journal of Primatology 29(5):1121-1145.
Haile M. 2005. Weather patterns, food security and humanitarian response in sub-
Saharan Africa. Philosophical transactions of the Royal Society of London Series
B, Biological sciences 360:2169-2182.
Hall P, Marron JS, and Park B. 1992. Smoothed cross-validation. Probability Theory and
Related Fields 92(1):1-20.
Hansteen TL, Andreassen HP, and Ims RA. 1997. Effects of spatiotemporal scale on
autocorrelation and home range estimators. The Journal of Wildlife Management:
280-290.
Hanya G, Noma N, and Agetsuma N. 2003. Altitudinal and seasonal variations in the diet
of Japanese macaques in Yakushima. Primates 44(1):51-59.
Hemson G, Johnson P, South A, Kenward R, Ripley R, and McDonald D. 2005. Are
kernels the mustard? Data from global positioning system (GPS) collars suggests
problems for kernel home-range analyses with least-squares cross-validation.
Journal of Animal Ecology 74(3):455-463.

112
Henzi SP, Byrne RW, and Whiten A. 1992. Patterns of movement by baboons in the
Drakensberg mountains: Primary responses to the environment. International
Journal of Primatology 13(6):601-629.
Hervert JJ, Bright JL, Henry RS, Piest LA, and Brown MT. 2005. Home-range and
habitat-use patterns of Sonoran pronghorn in Arizona. Wildlife Society Bulletin
33(1):8-15.
Heymann EW, and Aquino R. 2010. Peruvian red uakaris (Cacajao calvus ucayalii) are
not flooded-forest specialists. International Journal Primatology 31(5):751-758.
Hillman J. 1988. Home range and movement of the common eland (Taurotragus oryx
Pallas 1766) in Kenya. African Journal of Ecology 26(2):135-148.
Homewood KM. 1978. Feeding strategy of the Tana mangabey (Cercocebus galeritus
galeritus) (Mammalia: Primates). Journal of Zoology 186(3):375-391.
Horne JS, and Garton EO. 2006. Likelihood cross-validation versus least squares cross-
validation for choosing the smoothing parameter in kernel home-range analysis.
Journal of Wildlife Management 70(3):641-648.
Hunter CP. 2001. Ecological Determinants of Gelada Ranging Patterns (Theropithecus
gelada). PhD Thesis. The British Library: University of Liverpool. 208 p.
IBM Corp. Released 2012. IBM SPSS Statistics for Windows, Version 20.0. Armonk,
NY: IBM Corp.
Isbell LA. 1998. Diet for a small primate: Insectivory and gummivory in the (large) patas
monkey (Erythrocebus patas pyrrhonotus). American Journal of Primatology
45(4):381-398.
Isbell LA, Cheney DL, and Seyfarth RM. 1990. Costs and benefits of home range shifts
among vervet monkeys (Cercopithecus aethiops) in Amboseli National Park,
Kenya. Behavioral Ecology and Sociobiology 27(5):351-358.
Iwamoto T. 1979. Feeding Ecology. In: Kawai M, editor. Ecological and Sociological
Studies of Gelada Baboons: S. Karger. p 279-330.
Iwamoto T. 1993. The ecology of Theropithecus gelada. In: Jablonski NG, editor.
Theropithecus: The Rise and Fall of a Primate Genus. Great Britain: Cambridge
University Press. p 441-452.
Iwamoto T, and Dunbar RIM. 1983. Thermoregulation, habitat quality and the
behavioural ecology of gelada baboons. Journal of Animal Ecology 52(2):357-
366.

113
Jablonski NG. 1993. Theropithecus: the rise and fall of a primate genus: Cambridge
University Press.
Janson C, and Verdolin J. 2005. Seasonality of primate births in relation to climate. In:
Brockman D, and van Schaik C, editors. Seasonality in Primates: Studies of
Living and Extinct Human and Non-human Primate. New York: Cambridge
University Press. p 307-350.
Jennrich R, and Turner F. 1969. Measurement of non-circular home range. Journal of
Theoretical Biology 22(2):227-237.
Jolly CJ. 1970. The seed-eaters: a new model of hominid differentiation based on a
baboon analogy. [Indianapolis]: [Bobbs-Merrill].
Kaplin BA. 2001. Ranging behavior of two species of guenons (Cercopithecus lhoesti
and C. mitis doggetti) in the Nyungwe Forest Reserve, Rwanda. International
Journal of Primatology 22(4):521-548.
Kawai M, editor. 1979. Ecological and Sociological Studies of Gelada Baboons: S.
Karger.
Kawai M, Ohsawa H, Mori U, and Dunbar R. 1983. Social organization of gelada
baboons: Social units and definitions. Primates 24(1):13-24.
Kernohan B, Gitzen R, and Millspaugh J. 2001. Analysis of animal space use and
movements. In: Millspaugh J, and Marzluff J, editors. Radio tracking and animal
populations. San Diego: Academic Press. p 125-166.
Kinnaird MF. 1990. Behavioral and demographic responses to habitat change by the Tana
River crested mangabey (Cercocebus galeritus galeritus). Ph.D. Dissertation.
Gainesville, FL: University of Florida.
Krausman PR, Leopold BD, Seegmiller RF, and Torres SG. 1989. Relationships between
desert bighorn sheep and habitat in western Arizona. Wildlife Monographs
(102):3-66.
Kummer H. 1968. Social Organization of Hamadryas Baboons: A Field Study. Chicago:
University of Chicago.
Kunz BK, and Linsenmair KE. 2008. The disregarded West: diet and behavioural
ecology of olive baboons in the Ivory Coast. Folia Primatologica 79(1):31-51.
Lachica M, and Aguilera J. 2000. Estimation of the energy costs of locomotion in the
Iberian pig (Sus mediterraneus). British Journal of Nutrition 83(01):35-41.

114
Lachica M, and Aguilera J. 2005. Energy expenditure of walk in grassland for small
ruminants. Small Ruminant Research 59(2):105-121.
Lachica M, Prieto C & Aguilera JF (1997) The energy cost of walking on the level and
on negative and positive slopes in the Granadina goat (Capra hircus). British
Journal of Nutrition 77, 73–81.
Laliberte AS, and Ripple WJ. 2004. Range contractions of North American carnivores
and ungulates. BioScience 54(2):123-138.
Laver PN, and Kelly MJ. 2008. A critical review of home range studies. Journal of
Wildlife Management 72(1):290-298.
Lee, LM. 2011. Parturition among gelada monkeys (Theropithecus gelada) at Guassa,
Ethiopia [M.A. thesis]. Fullerton: California State University Fullerton.
Legendre P. 1983. Spatial autocorrelation: Trouble or new paradigm? Ecology
74(6):1659-1673.
Lesage L, Crête M, Huot J, Dumont A, and Ouellet J-P. 2000. Seasonal home range size
and philopatry in two northern white-tailed deer populations. Canadian Journal of
Zoology 78(11):1930-1940.
Lesage L, Crête M, Huot J, Dumont A, and Ouellet J-P. 2000. Seasonal home range size
and philopatry in two northern white-tailed deer populations. Canadian Journal of
Zoology 78(11):1930-1940.
Leuthold W. 1972. Home range, movements and food of a buffalo herd in Tsavo National
Park. African Journal of Ecology 10(3):237-243.
Leuthold W. 1978. On the ecology of the gerenuk Litocranius walleri. Journal of Animal
Ecology 47(2):561-580.
Leuthold B, and Leuthold W. 1978. Ecology of the giraffe in Tsavo East National Park,
Kenya. African Journal of Ecology 16(1):1-20.
Li B, Chen C, Ji W, and Ren B. 2000. Seasonal home range changes of the Sichuan snub-
nosed monkey (Rhinopithecus roxellana) in the Qinling Mountains of China.
Folia Primatologica 71(6):375-386.
Li D, Grueter CC, Ren B, Long Y, Li M, Peng Z, and Wei F. 2008. Ranging of
Rhinopithecus bieti in the Samage Forest, China. II. Use of land cover types and
altitudes. International Journal of Primatology 29(5):1147-1173.
Li D, Ren B, Li B, and Li M. 2010. Range expansion as a response to increasing group
size in the Yunnan snub-nosed monkey. Folia Primatologica 81(6):315-329.

115
Li X, and Walker D. 1986. The plant geography of Yunnan Province, southwest China.
Journal of Biogeography:367-397.
Li Z, and Rogers ME. 2005. Habitat quality and range use of white-headed langurs in
Fusui, China. Folia Primatologica 76(4):185-195.
Loveridge AJ, Valeix M, Davidson Z, Murindagomo F, Fritz H, and Macdonald DW.
2009. Changes in home range size of African lions in relation to pride size and
prey biomass in a semi-arid savanna. Ecography 32(6):953-962.
Luccarini S, Mauri L, Ciuti S, Lamberti P, and Apollonio M. 2006. Red deer (Cervus
elaphus) spatial use in the Italian Alps: home range patterns, seasonal migrations,
and effects of snow and winter feeding. Ethology Ecology & Evolution
18(2):127-145.
Mahli Y, and Wright J. 2004. Spatial patterns and recent trends in the climate of tropical
rainforest regions. Philosophical transactions of the Royal Society of London
Series B, Biological sciences 359(1443):311-329.
Marra PP, Francis CM, Mulvihill RS, and Moore FR. 2005. The Influence of climate on
the timing and rate of spring bird migration. Oecologia 142(2):307-315.
Mekonnen A, Bekele A, Fashing PJ, Hemson G, and Atickem A. 2010. Diet, activity
patterns, and ranging ecology of the Bale monkey (Chlorocebus djamdjamensis)
in Odobullu Forest, Ethiopia. International Journal of Primatology 31(3):339-362.
Mitchell MS, and Powell RA. 2007. Optimal use of resources structures home ranges and
spatial distribution of black bears. Animal Behaviour 74(2):219-230.
Mori A, and Belay G. 1990. The distribution of baboon species and a new population of
gelada baboons along the Wabi-Shebeli river, Ethiopia. Primates 31(4):495-508.
Mori A, Iwamoto T, Mori U, and Bekele A. 1999. Sociological and demographic
characteristics of a recently found Arsi gelada population in Ethiopia. Primates
40(2):365-381.
Moua C, Fashing PJ, and Nguyen N. 2012. Inter-annual variability in the ranging ecology
of wild geladas (Theropithecus gelada) at Guassa, Ethiopia, from 2007-2011
[Abstract]. American Society of Primatology, 2012.
Mysterud A. 1999. Seasonal migration pattern and home range of roe deer (Capreolus
capreolus) in an altitudinal gradient in southern Norway. Journal of Zoology
247(4):479-486.
Nicholson MC, Bowyer RT, and Kie JG. 1997. Habitat selection and survival of mule
deer: Tradeoffs associated with migration. Journal of Mammalogy 78(2):483-504.

116
Newton‐Fisher NE. 2003. The home range of the Sonso community of chimpanzees from
the Budongo Forest, Uganda. African Journal of Ecology 41(2):150-156.
Nguyen N, and Fashing PJ. 2009. Guassa gelada research project: A guide to working on
the Guassa gelada monkey project for prospective field research assistants.
Unpublished manuscript.
Nguyen N, and Fashing PJ. 2012. Birth seasonality in relation to rainfall and food in wild
geladas (Theropithecus gelada) at Guassa, Ethiopia: Variation within and between
one-male units. American Journal Physical Anthropology 147:223-223.
Nguyen N, Fashing PJ, Boyd DA, Barry TS, Burke RJ, Goodale CB, Jones SC, Kerby JT,
Kellogg BS, and Lee LM. 2015. Fitness impacts of tapeworm parasitism on wild
gelada monkeys at Guassa, Ethiopia. American journal of primatology 77(5):579-
594.
Nilsen EB, Pedersen S, and Linnell JD. 2008. Can minimum convex polygon home
ranges be used to draw biologically meaningful conclusions? Ecological Research
23(3):635-639.
Niu K, Tan CL, and Yang Y. 2010. Altitudinal movements of Guizhou snub-nosed
monkeys (Rhinopithecus brelichi) in Fanjingshan National Nature Reserve,
China: implications for conservation management of a flagship species. Folia
Primatologica 81(4):233-244.
Ohsawa H. 1979. Herd Dynamics. In: Kawai M, editor. Ecological and Sociological
Studies of Gelada Baboons: S. Karger. p 48-80.
Olupot W, Chapman CA, Brown CH, and Waser PM. 1994. Mangabey (Cercocebus
albigena) population density, group size, and ranging: A twenty‐year comparison.
American Journal of Primatology 32(3):197-205.
Otis DL, and White GC. 1999. Autocorrelation of location estimates and the analysis of
radiotracking data. The Journal of Wildlife Management:1039-1044.
Pallant J. 2010. SPSS survival manual: A step by step guide to data analysis using SPSS:
Open University Press.
Palma AC, Vélez A, Gómez‐Posada C, López H, Zárate DA, and Stevenson PR. 2011.
Use of space, activity patterns, and foraging behavior of red howler monkeys
(Alouatta seniculus) in an Andean forest fragment in Colombia. American journal
of primatology 73(10):1062-1071.

117
Pavelka MM, Brusselers O, Nowak D, and Behie A. 2003. Population reduction and
social disorganization in Alouatta pigra following a hurricane. International
Journal of Primatology 24(5):1037-1055.
Pebsworth PA, Morgan HR, and Huffman MA. 2012. Evaluating home range techniques:
use of Global Positioning System (GPS) collar data from chacma baboons.
Primates 53(4):345-355.
Pimley ER, Bearder SK, and Dixson AF. 2005. Home range analysis of Perodicticus
potto edwardsi and Sciurocheirus cameronensis. International Journal of
Primatology 26(1):191-206.
Phillips R, Berg W, and Siniff D. 1973. Moose movement patterns and range use in
northwestern Minnesota. The Journal of Wildlife Management:266-278.
Poulsen JR, Clark CJ, and Smith TB. 2001. Seasonal variation in the feeding ecology of
the grey-cheeked mangabey (Lophocebus albigena) in Cameroon. American
Journal of Primatology 54(2):91-105.
Powell R. 2000. Animal Home Ranges and Territories and Home Range Estimators. In:
Boitani L, and Fuller T, editors. Research Techniques in Animal Ecology:
Controversies and Consequences. New York: Columbia University Press. p 65-
110.
Rabinowitz AR, and Nottingham BG, Jr. 1986. Ecology and behaviour of the Jaguar
(Panthera onca) in Belize, Central America. Journal of Zoology 210(1):149-159.
Robbins MM, and McNeilage A. 2003. Home range and frugivory patterns of mountain
gorillas in Bwindi Impenetrable National Park, Uganda. International Journal of
Primatology 24(3):467-491.
Rodgers AR, Carr A, Beyer H, Smith L, and Kie J. 2007. HRT: home range tools for
ArcGIS. Ontario Ministry of Natural Resources, Centre for Northern Forest
Ecosystem Research, Thunder Bay, Ontario, Canada.
Rowe J, and Dalgarn S. 2010. Home range size and daily movements of midland painted
turtles (Chrysemys picta marginata) in relation to body size, sex, and weather
patterns. Herpetological Conservation and Biology 5(3):461-473.
Rueness EK, Asmyhr MG, Sillero-Zubiri C, Macdonald DW, Bekele A, Atickem A, and
Stenseth NC. 2011. The cryptic African wolf: Canis aureus lupaster is not a
golden jackal and is not endemic to Egypt. PLoS ONE 6(1):e16385.
Salkind NJ. 2009. Statistics for people who (think they) hate statistics: EXCEL 2007
Edition: Sage Publications, Incorporated. Los Angeles, CA, USA.

118
Schaller GB. 2000. The Mountain Gorilla: Ecology and Behavior. Chicago: University of
Chicago Press. 450 p.
Schoener T. 1981. An empirically based estimate of home range. Theoretical Population
Biology 20:281-325.
Schreier AL. 2010. Feeding ecology, food availability and ranging patterns of wild
hamadryas baboons at Filoha. Folia Primatologica 81(3):129-145.
Silverman, BW. 1986. Density Estimation for Statistics and Data Analysis. London:
Chapman and Hall.
Seaman D, and Powell R. 1996. An evaluation of the accuracy of kernel density
estimators for home range analysis. Ecology 77(7):2075-2085.
Seaman DE, Millspaugh JJ, Kernohan BJ, Brundige GC, Raedeke KJ, and Gitzen RA.
1999. Effects of sample size on kernel home range estimates. The Journal of
Wildlife Management: 739-747.
Siex KS, and Struhsaker TT. 1999. Ecology of the Zanzibar red colobus monkey:
Demographic variability and habitat stability. International Journal of
Primatology 20(2):163-192.
Sigg H, and Stolba A. 1981. Home range and daily march in a hamadryas baboon troop.
Folia Primatologica 36(1-2):40-75.
Sikes SK. 1971. The Natural History of the African Elephant. London: Weidenfeld &
Nicolson. 397 p.
Smuts BB. 1985. Sex and friendship in baboons: New York, Aldine Publishing Co.
Sprague D. 2000. Topographic effects on spatial data at a Japanese macaque study site.
American Journal of Primatology 52(3):143-147.
Stephen DW, and Krebs JR. 1986. Foraging Theory. USA: Princeton University Press.
Stelzner JK. 1988. Thermal effects on movement patterns of yellow baboons. Primates
29(1):91-105.
Stoltz L, and Saayman G. 1970. Ecology and behaviour of baboons in the northern
Transvaal. Annals of the Transvaal Museum 26(99):143.
Strier K. 1987. Ranging behavior of woolly spider monkeys, or muriquis, Brachyteles
arachnoides. International Journal of Primatology 8(6):575-591.

119
Sutherland WJ, Newton I, and Green R. 2004. Bird Ecology and Conservation: A
Handbook of Techniques. Oxford, New York: Oxford University Press.
Swedell L. 2006. Strategies of Sex and Survival in Hamadryas Baboons: Through a
Female Lens. Upper Saddle River, New Jersey: Pearson Prentice Hall.
Swedell L. 2011. African Papionins: Diversity of social organization and ecological
flexibility. In: Campbell C, Fuentes A, MacKinnon K, and Panger M, editors.
Primates in Perspective. New York and Oxford: Oxford University Press. p 241-
277.
Swihart RK, and Slade NA. 1985a. Influence of sampling interval on estimates of home-
range size. The Journal of Wildlife Management:1019-1025.
Swihart RK, and Slade NA. 1985b. Testing for independence of observations in animal
movements. Ecology:1176-1184.
Symington MM. 1988. Demography, ranging patterns, and activity budgets of black
spider monkeys (Ateles paniscus chamek) in the Manu National Park, Peru.
American Journal of Primatology 15(1):45-67.
Snyder-Mackler N, Beehner J, and Bergman T. 2012. Defining Higher Levels in the
Multilevel Societies of Geladas (Theropithecus gelada). International Journal of
Primatology 33(5):1054-1068.
Tan CL, Guo S, and Li B. 2007. Population structure and ranging patterns of
Rhinopithecus roxellana in Zhouzhi National Nature Reserve, Shaanxi, China.
International Journal of Primatology 28(3):577-591.
Tufto J, Andersen R, and Linnell J. 1996. Habitat use and ecological correlates of home
range size in a small cervid: the roe deer. Journal of Animal Ecology:715-724.
Vedder AL. 1984. Movement patterns of a group of free‐ranging mountain gorillas
(Gorilla gorilla beringei) and their relation to food availability. American Journal
of Primatology 7(2):73-88.
Venkataraman VV, Glowacka H, Fritz J, Clauss M, Seyoum C, Nguyen N, and Fashing
PJ. 2014. Effects of dietary fracture toughness and dental wear on chewing
efficiency in geladas (Theropithecus gelada). American journal of physical
anthropology 155(1):17-32.
Venkataraman VV, Kerby JT, Nguyen N, Ashenafi ZT, and Fashing PJ. 2015. Solitary
Ethiopian wolves increase predation success on rodents when among grazing
gelada monkey herds. Journal of Mammalogy 96(1):129-137.

120
Vila AR, Beade MS, and Barrios Lamunière D. 2008. Home range and habitat selection
of pampas deer. Journal of Zoology 276(1):95-102.
Wada K, Tokida E, and Ogawa H. 2007. The influence of snowfall, temperature and
social relationships on sleeping clusters of Japanese monkeys during winter in
Shiga Heights. Primates 48(2):130-139.
Wallace HM, and Trueman SJ. 1995. Dispersal of Eucalyptus torelliana seeds by the
resin-collecting stingless bee, Trigona carbonaria. Oecologia 104(1):12-16.
Wartmann FM, Purves RS, and van Schaik CP. 2010. Modelling ranging behaviour of
female orang-utans: a case study in Tuanan, Central Kalimantan, Indonesia.
Primates 51(2):119-130.
Watts D. 1998. Long-Term Habitat Use by Mountain Gorillas (Gorilla gorilla beringei).
1. Consistency, Variation, and Home Range Size and Stability. International
Journal of Primatology 19(4):651-680.
Western D. 1989. The Ecological Role of Elephants in Africa. In: Gakahu C, Western O,
Martin E, Goode B, and Vigne L, editors. Pachyderm: A Publication of the
African Elephant and Rhino Specialist Group: Wildlife Conservation
International. p 42-45.
White G, and Garrott R. 1990. Analysis of wildlife radio-tracking data. University of
Michigan: Academic Press.
Whiten A, Byrne RW, and Henzi SP. 1987. The behavioral ecology of mountain
baboons. International Journal of Primatology 8(4):367-388.
Wieczkowski J. 2005. Examination of increased annual range of a Tana mangabey
(Cercocebus galeritus) group. American Journal Physical Anthropology
128(2):381-388.
Wiens JA. 1989. The Ecology of Bird Communities. Cambridge, UK: Cambridge
University Press.
Willems EP, Barton RA, and Hill RA. 2009. Remotely sensed productivity, regional
home range selection, and local range use by an omnivorous primate. Behavioral
Ecology 20(5):985-992.
Worton BJ. 1989. Kernel methods for estimating the utilization distribution in home-
range studies. Ecology 70(1):164-168.
Wrangham R. 1980. Bipedal locomotion as a feeding adaptation in gelada baboons, and
its implications for hominid evolution. Journal of Human Evolution 9(4):329-331.

121
Wrangham RW, Chapman CA, and Chapman LJ. 1994. Seed dispersal by forest
chimpanzees in Uganda. Journal of Tropical Ecology 10(03):355-368.
Zinner D, Peláez F, and Torkler F. 2002. Distribution and habitat of grivet monkeys
(Cercopithecus aethiops aethiops) in eastern and central Eritrea. African Journal
of Ecology 40(2):151-158.