littoral benthos of the saline crater lakes of the basin of oriental, mexico
TRANSCRIPT

IntemationalJoumalofSaltLakeResearch 7: 87-108,1998.@ 1998 Kluwer Academic Publishers. Printed in the Netherlands.
Littoral benthos of the saline crater lakes of the basin
of Oriental, Mexico
JAVIER ALCOCER1,*, ELVA ESCOBAR2, ALFONSO LUGO1 andLAURA PERALTA 1
1 Limnology Lab" Environmental Conservation and Improvement Project, UIICSE, UNAM
Campus Iztacala, Av. de los Barrios s/n, 'Los Reyes Iztacala, Tlalnepantla, Edo. de Mexico
54090, MEXICO; 2Benthic Ecology Lab., Institute of Marine Sciences and Limnology,
UNAM. A.P. 70-405, Ciudad Universitaria 045/0, D.F., MEXICO
(* author for correspondence; E-mail: jalcocer@ servidol:unam.mx)
Abstract. Two saline crater lakes in the basin of Oriental, Puebla- Tlaxcala- Veracruz, wereinvestigated for littoral benthic macroinvertebrates. Fifty taxa were identified with the oligo-chaetes, amphipods, chironornids and leeches the dorninant organisms. These four taxa madeup to 99 per cent in both number and biomass. Limnodrilus hoffmeisteri, Hyalella azteca,Tanypus (Apelopia) sp. and Stictochironomus sp. were the most abundant organisms. Unlikeother saline lakes which have a littoral benthos dorninated by chironornids, Alchichica andAtexcac were dorninated by oligochaetes (70-73 per cent). The gastropod, Physa sp., wasfound up to a salinity of 8 9 L -I; in other studies, it has been found in lower salinities. L.hoffmeisteri is also a typical inhabitant of freshwater lakes, particularly of deep waters. It wasdorninant in the shallow, saline waters of the two lakes studied. Salinity did not affect speciesrichness. Alchichica, the most saline of the six crater lakes of Puebla (salinity, 7.4 9 L -I ),
had 30 per cent more species than the freshwater lakes, and double the species number ofAtexcac. It seems the main factor controlling species richness and the density and biomassof organisms in Alchichica and Atexcac is the presence of aquatic vegetation. It does this byincreasing habitat heterogeneity and providing food and protection against predators.
Key words: benthos, crater lakes, littoral, macroinvertebrates, Mexico, Puebla, saline lakes
Introduction
The basin of Oriental is located in the south-easternmost portion of the Mexi-can Plateau (18°5615111-19°43125'1 N, 97°0711011-98°03104'1 W, 2,312 m.a.s.l.at the lowest part of the basin). In the center of this endorheic basin, thereare s ix crater lakes, four in Los Llanos de San Juan (in the northern portion)and two in Los Llanos de San Andrés (in the southern portion). Alchichicaand Atexcac are saline (i.e., salinity ~ 3 9 L -1); the other four (La Preciosa,Quechulac, Aljojuca and Tecuitlapa) are fresh. A semi-arid climate charac-

88
Figure 1. Aerial photograph of the Llanos de San Juan, basin of Oriental, Mexico, whereAlchichica and Atexcac saline crater lakes are located. The other two crater lakes (La Preciosaand Quechulac) are fresh.
t~rizes the Llanos de San Juan where Alchichica and Atexcac are located(Figure 1 ), with an annual precipitation regime of < 400 mm, an annualevaporation of 500-600 mm, and a mean annual temperature of 12.9 °C
(9.3-15.4 °C) (García, 1988).Alchichica and Atexcac were fonned in the Quaternary by phreatic explo-
sions (Gasca, 1981) and, like the other lakes of the Neovolcanic axis, aresubject to a process of natural desiccation (senescence and a drier climate,Heine, 1973) and human desiccation (e.g. water over-extraction and drainagefor agricultural use). This process of desiccation has already been reported byAlcocer and Escobar.(1990), Alcocer and Williarns (1993) and Alcocer et al.
(1998).Alchichica and Atexcac are deepJakes (ZM = 64 m, meandepth =c38.6 m;
ZM = 39.1, mean depth = 25.7m,respectively), witha surface areaof 1;81and 0.29 km2, a maximum Jength of 1,730 and 780m, a shoreline length of

89
5.06 and 2.18 km, with volumes of 69.92 and 6.15 m3 x 106 of saline water,respectively (Arredondo-Figueroa et al., 1983).
According to Vilaclara et a1. (1993), Alchichica and Atexcac are dom-inated by sodium and magnesium with sma11er amounts of potassium andcalcium, and chloride and bicarbonate with sma11er amounts of sulphate andcarbonate. Both lakes are saline (7.2::1:: 0.1 9 L -I and 6.0::1:: 0.0 9 L -I) and
alkaline (pH = 9.0::1:: 0.1 and 8.4::1:: 0.2) (Alcocer et al., 1993a).Few studies of the benthos of tropical Mexican athalassohaline lakes
have been undertaken (e.g. Alcocer et al., 1993a,b, 1997). This study aimedto describe the abundance and composition of the littoral benthic fauna oftwo Mexican tropical athalassohaline lakes and the environmental factorsinvolved in its temporal and spatial variation.
Material and methods
Based on littoral area characteristics (e.g. vegetation cover and type, sedimenttexture, wave protection, etc.), five sampling zones (habitats) were establishedin Alchichica (ALC1-ALC5) and one in Atexcac (ATE1). Alchichica had aheterogeneous littora1 area, but Atexcac, with steep cliff-like slopes, had alimited or no littoral zone. A seasonal sampling program was designed. Ateach station, triplicate bottom samples were taken with an Ekman dredge of225cm2 gape, and a height of 15cm. Particle size was determined according tothe Wentworth classification. Loss on ignition was determined after one hourat 550°C (organic matter content), and after one hour at 950°C (carbonatecontent). Aquatic vegetation cover was estimated visually. pH, dissolved oxy-gen (mg L -I and per cent saturation), temperature, and salinity (calculated
from K25 measurements) were determined in situ with a calibrated HydrolabDatasonde 3/Surveyor 3 multiparameter water quality datalogger and loggingsystem. Macroinvertebrates were sieved through a 0.59 mm mesh, sorted, pre-served in 70 per cent ethanol, identified, and oven-dried at 60 oC to constantweight. Loss on ignition (roughly equivalent to biomass) was determinedafter one hour at 550 oC for ash-free dried weight (APHA et al., 1985; Weber,
1973).
Results and discussion
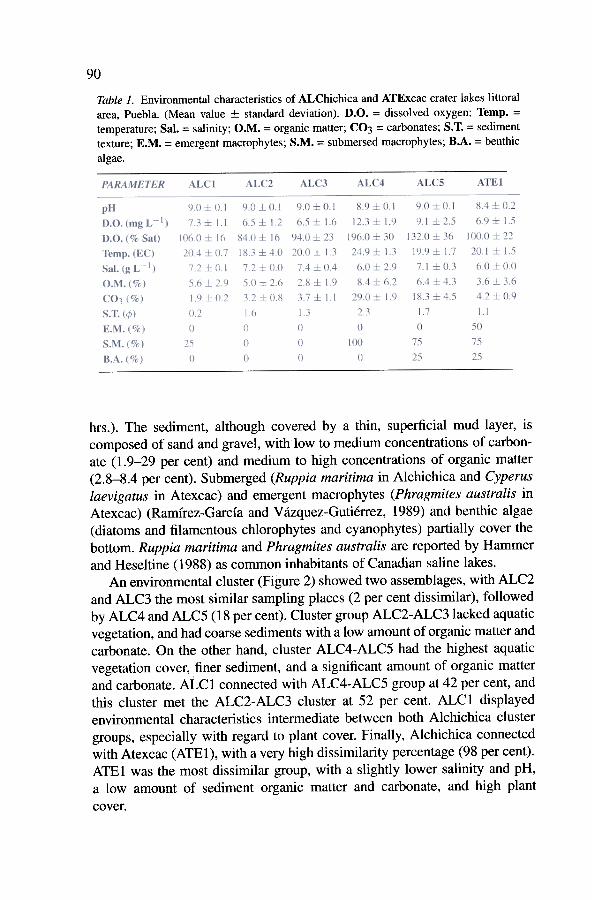
The environmental characteristics of the littoral areas of Alchichica andAtexcac are given on Table 1. Both crater lakes are temperate (> 18 °C),hyposaline (6-7.4 9 L -1), alkaline (pH > 8), and had a high concentration ofdissolved oxygen (> 80 per cent) during sampling hours (i.e., 8:30-17:30
90
Table 1. Environmental characteristics of ALChichica andATExcac crater lakes littoralarea, Puebla. (Mean value ::I: standard deviation). D.O. = dissolved oxygen; Temp. =temperature; Sal. = salinity; O.M. = organic matter; CO3 = carbonates; S. T. = sedimenttexture; E.M. = emergent macrophytes; S.M. = submersed macrophytes; B.A. = benthic
algae.
hrs.). The sediment, although covered by a thin, superficial mud layer, iscomposed of sand and gravel, with low to medium concentrations of carbon-ate (1.9-29 per cent) and medium to high concentrations of organic matter(2.8-8.4 per cent). Submerged (Ruppia maritima in Alchichica and Cyperuslaevigatus in Atexcac) and emergent macrophytes (Phragmites australis inAtexcac) (Ramírez-García and Vázquez-Gutiérrez, 1989) and benthic algae(diatoms and filamentous chlorophytes and cyanophytes) partially cover thebottom. Ruppia maritima and Phragmites australis are reported by Hammerand Heseltine ( 1988) as comrnon inhabitants of Canadian saline lakes.
An environmenta1 cluster (Figure 2) showed two assemblages, with ALC2and ALC3 the most sirnilar sampling places (2 per cent dissimilar), followedby ALC4 and ALC5 (18 per cent). Cluster group ALC2-ALC3lacked aquaticvegetation, and had coarse sediments with a low amount of organic matter andcarbonate. On the other hand, cluster ALC4-ALC5 had the highest aquaticvegetation cover, finer sediment, and a significant amount of organic matterand carbonate. ALCl connected with ALC4-ALC5 group at 42 per cent, andthis cluster met the ALC2-ALC3 cluster at 52 per cent. ALCl displayedenvironmental characteristics intermediate between both Alchichica clustergroups, especially with regard to plant cover. Finally, Alchichica connectedwith Atexcac (ATE1), with a very high dissimilarity percentage (98 per cent).ATEl was the most dissimilar group, with a slightly lower salinity and pH,a low amount of sediment organic matter and carbonate, and high plant
cover.

91
Dissimilarity ( 1-Pearson r}
o 25 50 75 100%
ALC4
ALC2
ALC3
Figure 2. Environmental dissimilarity dendrogram ( I -Pearson r, complete linkage ), showing
the sampling stations of the saline crater lakes of Puebla, Mexico (ALChichica and ATExcac ).
Plant type and cover as well as salinity were the most important envi-ronmental parameters explaining the differences between sampling stationsin Alchichica and Atexcac. The same parameters were found to be the mostsignificant by Alcocer (1995) and Alcocer et al. (1993a) when comparingenvironments in the six crater lakes of Puebla using multivariate analysis(cluster and principal component analysis).
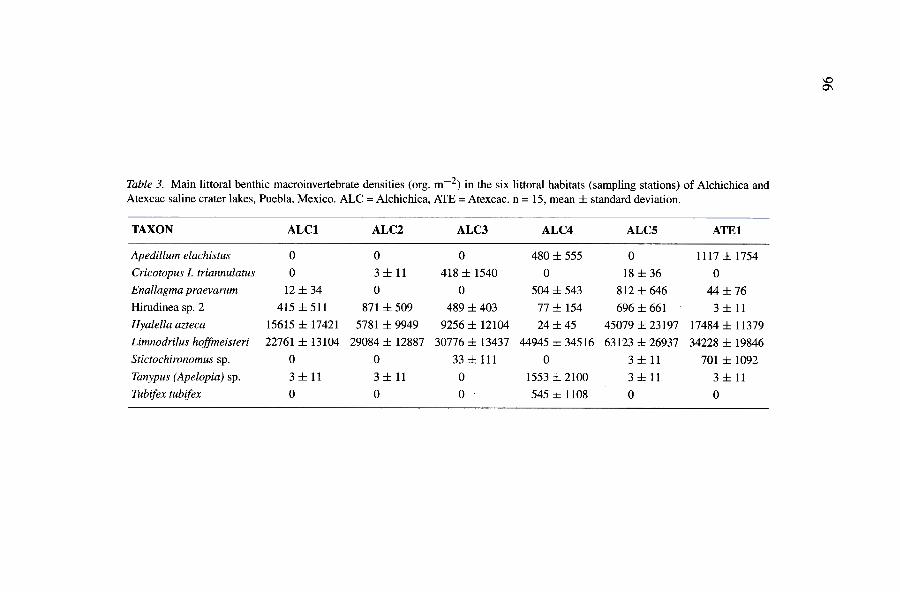
The littoral comrnunity of benthic macroinvertebrates comprised 50 taxa(Table 2), 44 in Alchichica (88 per cent) and 21 in Atexcac (42 per cent).Alchichica and Atexcac have a comparatively rich macroinvertebrate faunadue, mostly, to their relatively low salinity, as found by Hammer et al.(1975) and Williams (1972) in other low-salinity lakes. Four taxa composedup to 99 per cent of the organisms present: oligochaetes (70-73 per cent),amphipods (24-26 per cent), rnidges (1-3 per cent) and leeches (0.1-1 percent). Three oligochaetes (Limnodrilus hoffineisteri, the most significant),one amphipod (Hyalella azteca), 19 chironomid species (Tanypus (Apelopia)sp. and Stictochironomus sp., the most important in Alchichica and Atexcac,respectively), and two leech species were identified.
Alchichica and Atexcac have a low number of common taxa (i.e., 15,30 per cent of total species richness). Six species were found exclusively inAtexcac, and 29 in Alchichica. A cluster analysis based on presence-absence(JSI = Jaccard Sirnilarity Index) of the 50 species in each sampling station

92
Table 2. Taxonomic list of the littoral benthic macroinvertebrates of ALChichica and
ATExcac saline crater lakes, Puebla.
Taxa ALC ALC ALC ALC ALC ALC ATE1 2 3 4 5
xx
x x x xx
xx
x x x xx
x x
x
xx
xx
xx
xx
xxx x
x x x x x x x
x x xx
xx
xx
xx
xx
xx
xxx
xxx
xx
xx
xx
x
TURBELLARIA
NEMATODA
OLIGOCHAETA
Limnodrilus hoffineisteri
Tubifex tubifex
Nais variabilis
HIRUDINEA
Hirudinea sp. 1
Hirudinea sp. 2
AMPHIPODA
Hyalella azteca
EPHEMEROPTERA
Callibaetis montanus
ODONATA
Enallagma praevarum
Aeshna dugesi
HEMIPTERA
Buenoa sp.
Ambl)'sus sp.
Krizousacorixa tolteca
TRICHOPTERA
Oecetis sp.
Grensia sp.
Oxyethira sp.
Polycentropus sp.
COLEOPTERA
Stenus sp.
Tropistemus sp.
Berosus sp.
Donacia sp.
Hydroporus sp.
Laccodytes ( ? ) sp.
DIPTERA
Culex sp.
Stratiomys sp.
Ephydra hians
xxxx
x xx
xxx
x xx
x
xx
xxxxxx
xxx
xxxxx
x
xxx
xxx
x

93
Table 2. Continued.
Taxa ALC ALC ALC ALC ALC ALC ATE
1 2 3 4 5
(Figure 3) showed that ALC1 and ALC2 were most similar in biologicalcomposition (JSI = 0.6). Apart from ALC1 and ALC2, the JSI applied toall Alchichica and Atexcac sampling stations indicated low biological resem-blance (i.e., JSI < 0.5). Although ALC4 and ALC5 have more species (9-11 )than ALC1 and ALC2, the latter group has a higher percentage of commonspecies than the former. The most diverse station in Alchichica, ALC4, has aspecies composition that is more similar to Atexcac than to other Alchichicastations; nonetheless, their similarity was low (JSI = 0.324). Note that both
are the least saline (i.e. , 6 9 L -1) of all stations sampled.Oligochaetes (Limnodrilus hoffineisteri, Tubifex tubifex and Nais vari-
abüis) were the most abundant littoral macroinvertebrates in Alchichica and

94
Jaccard Similarity Index
ALC2
Figure 3. Biological similarity dendrogram (Jaccard Similarity Index), showing samplingstations of the saline crater lakes of Puebla, Mexico (ALChichica and ATExcac ).
Atexcac, well above chironomid abundance. Oligochaetes averaged 38,247org. m-2 (up to 73 per cent of total abundance) in Alchichica, and 34,228org. m-2 (up to 70 per cent) in Atexcac. This is unusual given that the sha1lowlittoral areas of athalassohaline lakes are usually dominated by chironomids,and oligochaetes are the dominant group in the deep benthos (e.g. Galat etal., 1981; Turodancea and Harrison, 1988).
Amphipods, represented by Hyalella azteca, were the second most com-mon group, forming up to 24 per cent and 26 per cent of total abundancein Alchicha and Atexcac, respectively, and averaging 15,151 org. m-2 and17,484 org. m-2. According to Colbum (1988), Hyalella azteca is a typicaland comrnon species in low salinity pools (2-8 9 L -1) in the Death Valley,Califomia. We found it to be abundant near the upper limit of this salinity
range.Note that Limnodrilus hoffmeisteri, although an important species in fresh-
water lakes, especially in profunda1 regions, is not often reported from saltlakes. This is a1so the case for Stictochironomus. On the other hand, Hyalellaazteca and other arnphipods are frequently encountered in the littoral zoneof moderately saline lakes. Two uncommon organisms in Alchichica andAtexcac, Physa and Ephydra hians, have been reported from lower (Physa)or higher (Ephydra) salinity environments. Ephydra hians has been reported

95
(Alcocer et al" 1997; A.lcocer etal., unpublished) inhabiting two more salinelakes of the basin of Oriental: Totolcingo (K25 up to 30 mS cm-l) andTecuitlapa Norte (K25 up to 55.7 mS cm-1 ).
Insects predominated in Alchichica and Atexcac, comprising 40 of the 50taxa recognized. By far, the most diverse group of macroinvertebrates of thecrater lakes is the chironomids (Alcocer et al" 1993a), They comprised upto 19 species, 14 in Alchichica (Alcocer et al" 1993b) and lO in Atexcac,Cryptochironomus, a genus of little importance in the crater lakes, togetherwith Procladius and Tanytarsus, was the most common chironomid found byRawson and Moore (1944) in saline Saskatchewan lakes, Probably the muchlower salinity of these lakes (i.e" 0,5 9 L -I or more) explains this, Although
absent in Alchichica and Atexcac, Procladius and Tanytarsus are present insome freshwater crater lakes of Puebla (Alcocer et al., 1993a), Procladiuswas also found by Bayly and Williams (1966) in Australia at 22,3 9 L -I,Other insect groups in Alchichica and Atexcac have also been reported inWestem Australia at salinities greater than in our lakes (i,e., > lO 9 L -I ):
Odonata, Corixidae, Culicidae, Chironomidae, Ephydridae, Stratiomyidaeand Coleoptera (Geddes et al., 1981),
Lake Bullenmerri (Timms, 1981) has a salinity similar to the crater lakes(8 9 L -I) but its biota is quite different, Bullenmeri has only 12 species,with Antipodrilus timmsi and Chironomus duplex dominant, and with abun-dant amphipods in the shallower areas, Nevertheless, the adjacent freshwater(0.4 9 L -I) Lake Purrumbete (Timms, 1981) has 37 species, This number iscomparable to Alchichica and, as in our crater lakes, Limnodrilus hoffineisteriis the dominant taxon. Seven chironomid species were found, with Procladiusvillosimanus the most numerous, The same genus occurs in Atexcac but is ofminor importance there.
Some genera present in the saline lakes of the Paroo, New SouthWales (Timms, 1993), were also recorded in Alchichica and Atexcac. Theyinclude Procladius, Cricotopus, Chironomus, Dicrotendipes, Tanytarsus,Culex, Berosus, and Stratiomyidae. Some of them (Cricotopus, Dicro-tendipes, Culex and Stratiomyidae) are reported from less saline waters, butwith Tanytarsus from 11,2 to 25,5 9 L -I,
Williams et al, (1990) listed the taxa in 79 Australian lakes spanning awide salinity range, Some of these also occur in Alchichica and Atexcac, e,g,Chironomus, Procladius, Ephydra, Berosus and Oligochaeta, Nevertheless,Physa, Ischnura, and Hirudinea were found at a salinity lower than occurs inAtexcac and Alchichica. Additionally, although Paratanytarsus (1,5-57.4 9L -I) and Tanytarsus (14,5-102 9 L -I) were reported from higher salinitiesby Williams et al, (1990), they occur in the freshwater crater lakes of Pueblabut not in Alchichica and Atexcac,
96
"O§~u:Eu:Eu~"""'o'V;d~
oo
.-~~
.-;.
"' Q
jO
JJ"O
3]~]
"' ~
"'
:t:.-t1~.'=
§
.D
Qj
.a a
"i§oj9
II;=
: ~
><
.
.-u"' ~
Qj
u
~
><
B.5
~---"
I' ~
a~~¿
uO
.-
~~"'
uQ
j .-
.-~.'=
u
"'-~~
.g II
QjU
;¡
e~.DQ
j .
t:: 8
Qj
.-
~
~
.8 ~
u ¿
~-
a.DQ
j
:EO
:~
"'
]~~-
~2
~-u.5
~~
.-
~~
"'.u
""' ~
~
~.t:)
Qj
~.::E
zo~~ u~< ~u~< ~u~ .,.u~< 11)u~< ...¡.¡
~
"'::! .-
~
"' .
-~
...A"'
::! ::!
.~
."'::!::..
"'~~
';;; ::
~§
.- .-~
;.
"' ~
.-=: .~
"'
N
~
c ~
c
'"' ...~
'"'
"' -k
~
"' ."'
.-=:
c "'
"'
";¡)...;~A
...",::~~
"'~
"' "'
::! ~
~
~::!
oj~~
e,--::!~
Q
~
~
C""S
=~
~"'¡:¡~
k
~,¿
;~:a",C
C~
~~
'"'-2
";;::...;".;.'='
"' .-~
~
~
'"'
:: ~
~0~
:::E:E
'~~
~~
O\\D
.,¡- r--
.,¡- C
'1V
) r')
00 O
\r--
\D--O
\O-
-r ~
~
~~
~~
~
r--0.,¡-r').'¡-00-r')0-.,¡-
00C'10
-.,¡- C
'1 r--
-r-- .,¡-
-r')
r-- r--
O\r')
\D--O
\\D
.'¡-\Dr')\D
--r')
\D
\D
C'1
C'1
--
~~
~~
~~
~O
00
C'1
\D
O\
r') r')
r') O
--0\r--C'1
OO
\DO
-V
) r')
.,¡- \D\D-O
00
V)
r').'¡- V
) 00
V)
.,¡-V)V
).,¡- --
V)
V)-.'¡-r')
C'1-
~
~~
~~
~
~00.,¡-r--.,¡-V
)0r')V)
00 0
r-- C
'1 .,¡-
V)
.,¡-.,¡-
V)
O\
V)V
):;;¡:
-
8 ~
0 r')-.'¡--
.,¡- 0C
'1r')-V
) .,¡
-~~
~~
0~00\\D
\Dr')00
00 00
V)
r-- r')
-.,¡- C
'1 r--
.,¡- O
\ 0r')r--
O\
00O
\ .,¡-
00-00\C
'1 -
-V)O
\- -
~
~~
~
~0r')0--.'¡-0r')0
r-- 00
00oor--o
V)
O\
C'1
Ñ
8-.,¡-
-.,¡--r--r')
-r')V
)-- -
~~
~~
~
00C'1v)v)-0r')0
---\D.,¡-
\D
r--V
) C
'1-C
'1
"O~(,;u:Eu:E.!,¡
<I:o~o.,g~
doO
I).~C
.~
(,;
j~"'"O~
~:;J"O(,;
C.~
(,;
.D
(,; "'
.!::-tj~
C(,;
o Q
j~
8.~-.~
Ir¡.~
-
"' II
] C
C
~.~
u
~~
"' Q
jI 8~
~~<I:
¿uO
I).~~
.!::"'
u"'
.~
(,;.!::8
u
o<
:E
IIQ
jU~
...:¡:0<
1:Q
j .
~
0Q
j .~~
~
.8 ~
u .
(,; (,;
8--fj
u .,
:E~
c "'
:i,~--(,;
Qj
9 ~
.~
-uc Q
j.~
c
(,; .~
~~
"'.u
~
(,;~
u
-~~
Qj
~<
r- r-M
NN
V)r-
r- ;:2;0\V
)0\0-3
08~a;~
8o
000000~
~
~~
~~
~"'"
OO
V)\0M
\0V)
O
r-N--\00
M
MO
Nr-O
\oO
00"'"V
)00
0000...;...;000
OM
r-r-r-V)r-
-"'" "'"
"'" "'"
--O
O\v)v)N
OO
0r-V)-V
)00000...;...;00
~~
~~
~~
~V
)\00000","V)
000","","-0000\00"'"0000O
O\v)N
v)OO
0000Ncr)000
-NV
)MO
O
OO
NV
) O
OO
\N","
V)r-
-\ON
ON
N
M0
\0-0- M
O0
OO
ON
00
~
~~
~~
~
~-M
r-Nr-
O\M
M
0","-V)
000-N
\00- "'"-
0 \O
00r- N
o00000N
000
0\ ","\00N
-ooNM
v)"'"
MO
V)-
O
M\0r-o
0 0000
~
~~
~~
"'" 000v)v)
-000\8
--\0N
O"'"
"'" r-
00000...;000M
r-M
- r-
0 \O
V)o\
-0
NO
\V)
0O
"'" "'"
r- 0
0 000
0
~
~~
~
~-::gooM
V
)
8 M
~
¡:::
80
r-N\0
000000...;000
NO
ON
V)
r-N
oor-oo -
","N\0v)
0O
","oor- 0
0000 0
~~
~~
~
\oo\M-
V)
","r-r-0\ 0
-","r-N
0O
Mr-M
0
00000...;000.,~
.-':3
"' .
-~
...o..,
~
~
.~
."'
~~
.. "'@
:'~.~
':3 ~
.~
.,
.~.s:: .~
"'
N
':3 ~
~
C)
'"' ...~
'"'
"' -k
':3 "'
."' .s::
C)
"' "'
~...;~
0 ,~
~:s
.,~
"'N~
",~~
..~
",':3~
~,--,-~
"' "
Q)
~0'""Q
~"':3.S
::.'
~...':3:a~
C)'"'~
~"':3S
~~
-~S
1;.,;:S"'
.-~
..':3 ~
'"'
~
~
~6~
:E':f:;:j~
~~
97

98
Lake Shala in Ethiopia (Turodancea and Harrison, 1988) is sirnilar toAlchichica and Atexcac in environmental characteristics (i.e., temperature,pH, dissolved oxygen, salinity, sediment texture, etc.) and origin. However,Lake Shala is quite different biologically. Dorninant taxa in that lake are ostra-cods, followed by oligochaetes and chironomids. Some ostracods were alsopresent in the meiobenthic fauna of Alchichica and Atexcac (Alcocer et al.,unpublished), but absent in the macrobenthos. Species richness in our lakes isalso higher than in Lake Shala. Nevertheless, Shala, Alchichica and Atexcacall have high benthic density and biomass (Tables 3, 4) despite differencesin species composition and low phytoplankton productivity. Turodancea andHarrison (1988) explained this for Shala by invoking the absence ofpredators;this is not so in our lakes where there are both vertebrate predators (e.g. silver-sides and salamanders) and invertebrate predators (e.g. Odonata, Hemiptera,Coleoptera) (Escobar Briones et al., unpublished).
Saline lakes in Central Asia, e.g. Balkhash in Kazakhstan, Issyk-Kul inKirghizia and Chany in Novosibirsk, have a high species richness (Aladin andPlotnikov, 1993). Nonetheless, they have few common taxa with Alchichicaand Atexcac. These include some chironomids (e.g. Chironomus, Stictochi-ronomus, Cricotopus) and tubificids (e.g. Tubifex, Nais).
Lakes Lenore (5.2-5.9 9 L -1), Killarney (5.6-5.7 9 L -1), Rabbit (8.5-8.7 9 L -1) and Arthur (8.9-24.7 9 L -1) in the Canadian prairies (Harnmeret al., 1990) have a sirnilar salinity to Alchichica and Atexcac. Hyalellaazteca, Callibaetis, Chironomus, Cricotopus, Cryptochironomus, Procladiusand Enallagma are common inhabitants in both series of lakes. Even so,Dicrotendipes, Psectrocladius and Physa, common in Alchichica and Atex-cac, are reported from less saline Canadian lakes (Hammer et al., 1990).Ephydra hians was found in very low numbers in Alchichica; it is a typicalinhabitant of hypersaline lakes. Lenore is by far the richest lake, with 31species (comparable to Alchichica), followed by Killarney (17), Rabbit (12)and Arthur (11). Other saline lakes in Saskatchewan and Alberta (Timms etal., 1986) have a large number of species in common with Alchichica andAtexcac.
Tectonic Pyramid Lake, Nevada, is similar in ionic composition, pH andsalinity (5.1 9 L -1) to Alchichica and Atexcac. Almost 50 per cent ofthe 35benthic macroinvertebrate genera reported by Galat et al. (1981) are presentin Alchichica and Atexcac. This trend towards common biota is also seen inthe hyposaline lakes of Death Valley, California (Colburn, 1988), and lakesin the Sud Lipez region of Bolivia (Dejoux, 1993).
From the above discussion, greater sirnilarity is evident in species com-position between Alchichica and Atexcac and the saline lakes of North andSouth America, than between these two lakes and the saline lakes of Asia and

99
Australia. This is probably related to the insect-impoverished fauna that ischaracteristic of Australian saline lakes. assuggested by Geddes et al. (1981).Insects dominate the fauna of North America saline lakes. while in Australiachironomids. oligochaetes and molluscs dominate lakes of low salinity. andostracods. isopods and. to a smaller extent. chironomids and ceratopogonidsthose of higher salinity (Timms. 1983). In fact. not only in North America buta1so in the northem Hemisphere as a whole. insects. especially chironomids.dominate (Hammer. 1986; Timms et al., 1986).
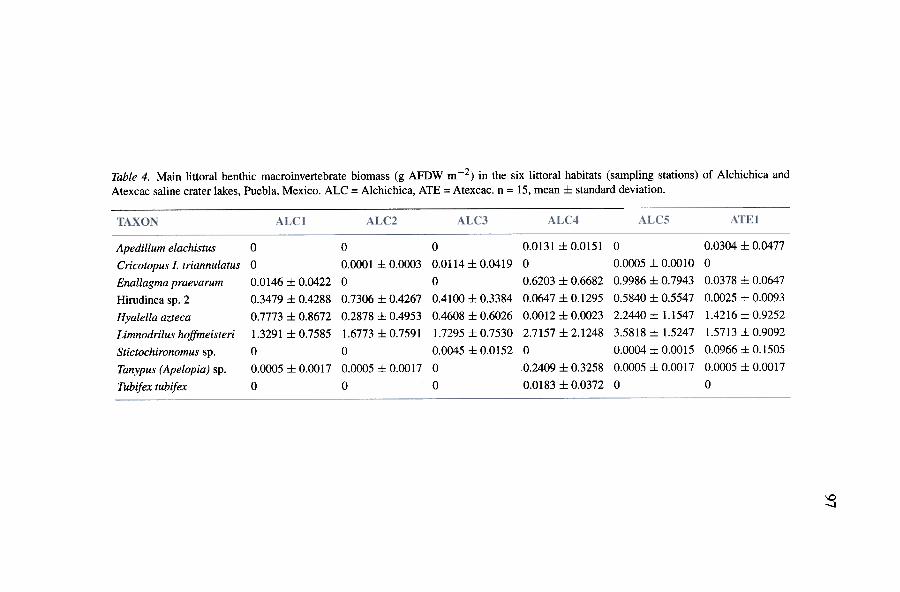
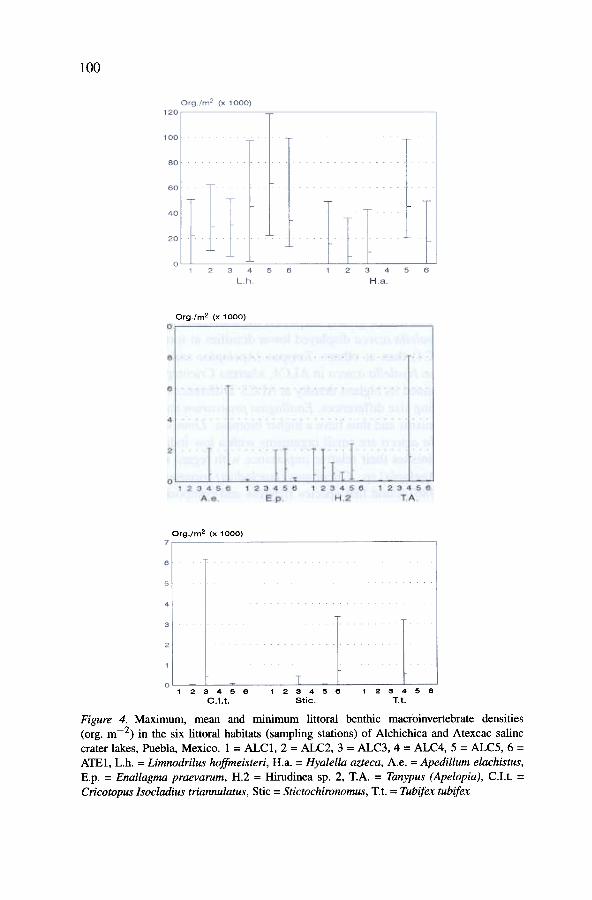
Figures 4 and 5 show the density (Table 3) and biomass (Table 4) ofthe main 9 littoral benthic macroinvertebrate species (they show density orbiomass :::: 1 per cent of total abundance or biomass at one sampling sta-tion at least) in each habitat (i.e.. the 6 sampling stations) of the salinecrater lakes of Puebla. The oligochaete Limnodrilus hoffmeisteri and theamphipod Hyalella azteca greatly surpassed the abundance of other species.Nonetheless. Hyalella azteca displayed lower densities at some stations (e.g.ALC3 and ALC4) than at others. Tanypus (Apelopia) and Tubifex tubifexwere greater than Hyalella azteca in ALC4. whereas Cricotopus (Isocladius)triannulatus attained its highest density at ALC3. Differences were lower forbiomass. reflecting size differences. Enallagma praevarum and Hirudinea sp.2 are large organisms and thus have a higher biomass. Limnodrilus hoffmeis-teri and Hyalella azteca are small organisms with a low individual biomasswhich thus diminishes their relative importance with regard to other species
[e.g. Tanypus (Apelopia) sp.. Cricotopus (Isocladius) triannulatus].Hammer (1986) found that species richness and composition are highly
correlated with salinity over a broad range of salinities. suggesting that salin-ity is an important factor determining the fauna in saline localities. However.Williams et al. (1990) found that at a finer scale (i.e.. within particular salinityranges) the correlation is non-significant at intermediate salinities (> 10 and< 200 9 1-1). Nonetheless. in the lowest part ofthe range (~ lO 9 L -I). thatis at salinities similar to the crater lakes. "the relationship remains highlysignificant (p < 0.001). indicating that salinity is closely associated with thespecies richness of freshwater species... These findings could hardly apply toa comparison of Alchichica and Atexcac since the salinity difference betweenthem is minimal (6-7.4 9 L -1). However. considering species richness in alllow salinity lakes. including Alchichica and Atexcac (Table 5. Figure 6). itseems there is more a trend than a significant relationship as reported byWilliams et a1. (1990).
The six crater lakes of Puebla cover a sa1inity range at the lowest part ofthe salinity spectrum (0.1-74 9 L -1) considered by Williams et al. (1990).Nonetheless. their species richness (Alcocer, 1995; Alcocer et al.. 1993a)do not accord with the findings of Williams et al. (1990). The lowest and
100
Org/m2 (X 1000)
Org./m2 (x 1000)
123456123456123456C.I.t. Stic. T.t.
Figure 4. Maximum, mean and minimum littoral benthic macioinvertebrate densities(org. m-2) in the six littoral habitats (sampling stations) of Alchichica and Atexcac salinecrater lakes, Puebla, Mexico. 1 = ALC1, 2 = ALC2, 3 = ALC3, 4 = ALC4, 5 = ALC5, 6 =ATE1, L.h. = Limnodrilus hoffmeisteri, H.a. = Hyalella azteca, A.e. = Apedillum elachistus,E.p. = Enallagma praevarum, H.2 = Hirudinea sp. 2, T.A. = Tanypus (Apelopia), C.I.t. =Cricotopus Isocladius triannulatus, Stic = Stictochironomus, T.t. = Tubifex tubifex
101
123456123456123456C.I.t. Stic. T.t.
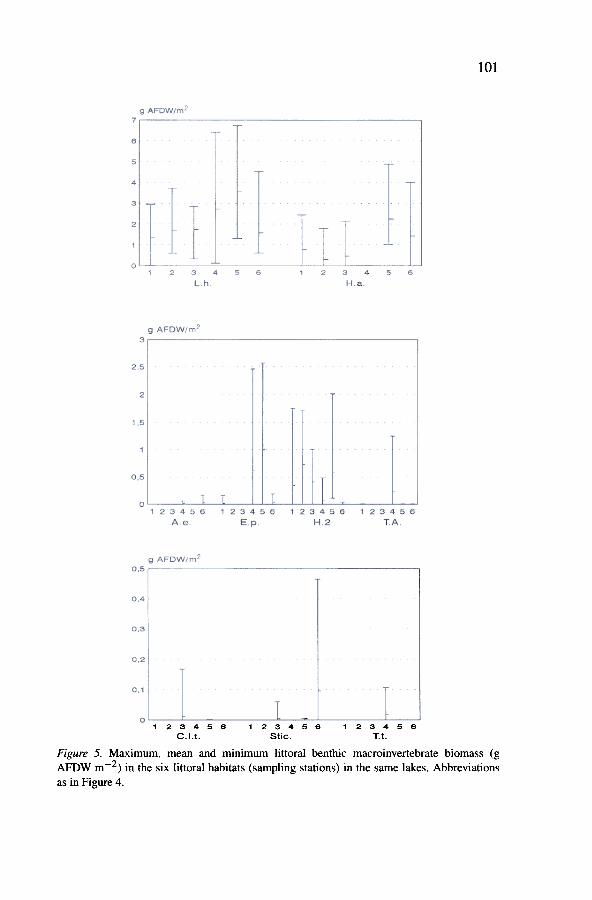
Figure 5. Maximum, mean and minimum littoral benthic macroinvertebrate biomass (gAFDW m-2) in the six littoral habitats (sampling stations) in the same lakes. Abbreviationsas in Figure 4.
102
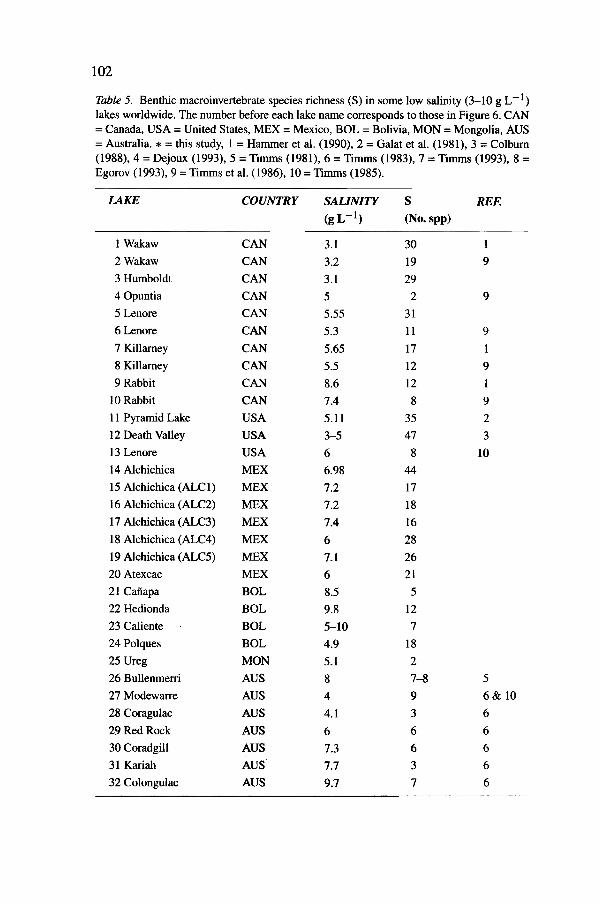
Table 5. Benthic macroinvertebrate species richness (S) in some low salinity (3-10 9 L -I )
lakes worldwide. The number before each lake name corresponds to those in Figure 6. CAN= Canada, USA = United States, MEX = Mexico, BOL = Bolivia, MON = Mongolia, AUS= Australia. * = this study, 1 = Hammer et al. (1990), 2 = Galat et al. (1981), 3 = Colburn(1988),4 = Dejoux (1993),5 = Timms (1981), 6 = Timms (1983),7 = Timms (1993), 8 =Egorov (1993),9 = Timms et al. (1986), 10 = Timms (1985).
LAKE COUNTRY SAUNITY
(gL-l)
REF.s
(No. spp)
1 Wakaw
2 Wakaw
3 Humboldt
4 Opuntia
5 Lenore
6 Lenore
7 Killamey
8 Killamey
9 Rabbit
10 Rabbit
11 Pyramid Lake
12 Death Valley
13 Lenore
14 Alchichica
15 Alchichica (ALC1)
16 Alchichica (ALC2)
17 Alchichica (ALC3)
18 Alchichica (ALC4)
19 Alchichica (ALC5)
20 Atexcac
21 Cañapa
22 Hedionda
23 Caliente
24Polques25 Ureg
26 Bullenmerri
27 Modewarre
28 Coragulac
29 Red Rock
30 Coradgill
31 Kariah
32 Colongulac
CANCAN
CANCANCANCAN
CANCAN
CANCAN
USAUSAUSAMEXMEX
MEXMEX
MEX
MEXMEXBOL
BOLBOLBOL
MONAUS
AUS
AUSAUS
AUSAUSAUS
3.1
3.2
3.1
5
5.55
5.3
5.65
5.5
8.6
7.4
5.11
3-5
6
6.98
7.2
7.2
7.4
6
7.1
6
8.5
9.8
5-10
4.9
5.1
8
4
4.1
6
7.3
7.7
9.7
30
19
29
2
31
11
17
12
12
8
35
47
8
44
17
18
16
28
26
21
5
12
7
18
2
7-8
9
3
6
6
3
7
1
9
9
9
I
9
I
9
2
3
10
5
6& 10
6
6
6
6
6
103
Table 5. Continued.
LAKE COUNTRY SAUNITY
(gL-l)
REF.s
(No. spp)
33 Ballyrnere
34 Strathem
35 Mid Bell
36 Sth Nichebulka
37 Lower Bell
38 Rainbar
39 Gidgee
40 Burkanoko
AUSAUS
AUS
AUSAUS
AUSAUS
AUS
3.46
3.63
4.18
4.91
5.42
6
7.56
9.5
36
II
21
21
12
5
30
18
7
7
7
7
7
7
7
7
s50
012
014
40
033011
0530 B1 039018
019
035 036
024
D2020 02 81615 Ch7
]4007
37~8 09 22 D034
10 6]27013
O 29038
10
300 Cb23 026
031
032021
0284IIJ25
o3 6 7
Salinity (g/L
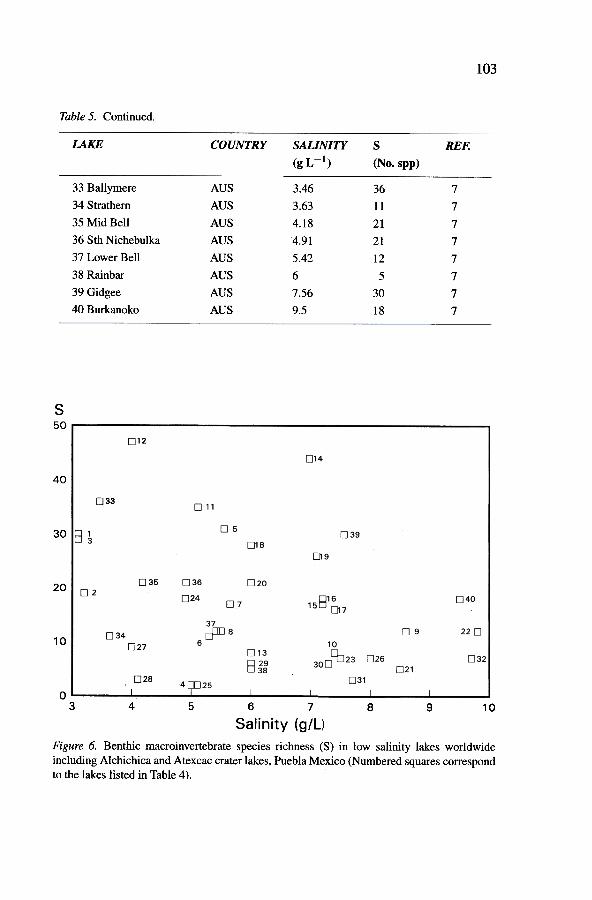
Figure 6. Benthic macroinvertebrate species richness (S) in low salinity lakes worldwideincluding Alchichica and Atexcac crater lakes, Puebla Mexico (Numbered squares correspondto the lakes listed in Table 4).
4 5 8 9 10

104
the highest species richness occur in the sa1ine 1akes Atexcac (21 spp.) andA1chichica (44 spp.), respectively. In between, the freshwater crater lakes(0.1-1.0 9 L -1) show intermediate species richness values (27-30 spp.). Itcan be concluded that salinity does not determine the differences in speciesrichness and composition between these two crater lakes.
As indicated, salinity (0.1- 7.4 9 L -1) is one of the most important featuresdifferentiating the crater lakes of Puebla, a1though it was not correlated withtheir species richness. Habitat heterogeneity and protection given by aquaticvegetation, tufa towers and rocks promoted the largest number of species (i.e.,from 21 to 28 in ALC4, ALC5 and ATE1), whereas exposed substrates andhomogeneity decreased it (i.e., from 15 to 17 in ALC1, ALC2 and ALC3)(Table 2). The main factor controlling species richness and organism densityin Alchichica and Atexcac seems to be the presence of aquatic vegetation.It does this by increasing habitat heterogeneity and providing food and pro-tection against predators in the littoral environment. Note that Alchichica islarger than Atexcac and has a more heterogeneous littora1 area.
Multiple correlation analysis (Table 6) of abundance (Table 3 and Fig-ure 4) and biomass (Table 4 and Figure 5) of the 9 most abundant taxa toenvironmental parameters (p < 0.05) exhibited the fol1owing trends. Thegreatest densities and biomass of Limnodrilus hoffmeisteri occurred at ALC4,ALC5 and ATE 1, stations with the highest sediment carbonate content. The1owest density of Hyalella azteca occurred at the warmest ALC4, whereasthe highest biomass developed at ALC5 and ATEl where benthic algae wereabundant. Apedilum elachistus was restricted to ALC4 and ATE1, stationswith a lower salinity and pH, and ATEl only, with emergent macrophytes.The highest densities and biomass of Enallagma praevarum were detected atALC4, ALC5 and ATE1, stations with the highest sediment carbonate con-tent, dissolved oxygen and amount of submerged macrophytes. The 1owestconcentrations of Hirudinea sp. 2 were associated with the low salinity andpH at ALC4 and ATE1, and at ATE1, abundant emergent macrophytes. Warmand well-oxygenated waters and the fine sediment texture characteristics ofALC4 seems to be suitable to Tanypus (Apelopia) and Tubifex tubifex whichwere abundant there. Cricotopus (Isocladius) triannulatus was abundant atALC3, but was not significantly correlated with any environmental parameter.Finally, the highest abundance of Stictochironomus occurred at ATE1, thestation with emergent macrophytes and the lowest pH (i.e., 8.4 :J:: 0.2).
Submerged macrophytes provide Enallagma praevarum larvae a place toclimb and prey on food and this species developed abundantly in such ahabitat. Hyalella azteca was abundant among benthic and epiphytic algaewhere it has food and protection against predators. Emergent macrophytescovered with epiphytic algae are probably an important habitat for the her-

105
Table 6. Multiple correlation ana'ysis (p < 0.05) of abundance (first row) and biomass(second row) of 9 most abundant taxa in relation to some environmental parameters.
Taxon pH DO Temp Sal Text OrgM CO3 EM SM HA
Apedillum elachistus -0.98
-0.89-0.96 0.91
Cricotopus I. triannulatus
Enallagma praevarum 0.83 0.82
0.91
0.92
Hirudinea sp.2 0.86
0.890.96 -{}.91
-0.84Hyalella azteca
0.88
Limnodrilus hoffineisteri 0.88
0.91
Stictochironomus -0.81-0.91 0.85
Tanypus (Apelopia) sp. 0.90 0.89
0.87 0.93 0.84
Tubifex tubifex 0.87 0.93
0.87 0.93
bivorous Apedilum elachistus and Stictochironomus. ALC4 is an iso1ated,sha11ow, warm station with many submerged macrophytes. Large amountsof organic matter accumu1ated in the sediment favor the deve1opment ofthe detritivorous Tubifex tubifex and Limnodrilus hoffmeisteri. This 1ocationa1so favoured carnivorous Tanypus (Apelopia) and, to a sma11er extent, thepredator Enallagma praevarum. A1though ALC4 1acked benthic a1gae, sub-merged macrophytes there provided a suitab1e habitat for Hyalella azteca.Even so, ALC4 is the on1y station where Hyalella azteca was present insma11 numbers. The presence of Hyalella azteca in the digestive tract of thesi1versides Poblana alchichica (A1cocer et al., unpub1ished) and the sa1aman-ders Ambystoma taylorii (Santine11i, 1995) suggests predation by vertebrates(a1so invertebrates such as Tanypus (Apelopia) and Enallagma praevarum)may be an exp1anation. However, carbon stab1e isotope ratios of the 1ittora1

106
community of Alchichica show Hyalella azteca is not a major food item forthese vertebrates (Escobar Briones et al., unpublished).
Seasonal variation in the abundance of the biota shows a winter-springpeak just after the end of the winter circulation and before the early stagesof stratification. The most productive period (algal blooms, mainly diatomsor the cyanobacteria Nodularia spumigena) in the lakes coincides with anincrease in water temperature. Reproduction could be triggered by this risein temperature and food availability. The highest number of oligochaetes wasobserved in fall and winter in Alchichica, and spring in Atexcac, but thendropped off moderately during the rest of the year. The maximum numberof chironornid larvae occurred in winter in Alchichica and Atexcac and justbefore emergence. A similar peak was observed in Pyramid Lake in springfor oligochaetes and Chironomus by Galat et al..(1981).
Although benthic macroinvertebrate density and biomass fluctuatedwidely (see standard deviation in Tables 3 and 4), the littoral areas ofAlchichica and Atexcac may be regarded as highly productive. Mean sea-sonal densities fluctuated from 12,068 org. m-2 (ALC4) to 122,577 org. m-2(ALC5), and biomass from 0.9175 9 AFDW m-2 (ALC1) to 9.1506 9AFDW m-2 (ALC5). Greatest mean seasonal abundance and biomassvalues were reached in ALC5 (97,363-122,577 org. m-2 and 6.7520-9.1506 9 AFDW m-2); conversely, the lowest abundance and biomass valueswere recorded in ALC2 (25,748-44,815 org. m-2 and 1.6326-3.5581 9AFDW m-2).
Protozoa, chlorophyll a and nutrient concentrations indicated that Atexcacis an oligotrophic and Alchichica is an oligo-mesotrophic lake (Lugo et al.,1993). Nonetheless, according to benthic macroinvertebrates, the littoral areaof both crater lakes is more eutrophic (mesosaprobic) in condition. Signif-icant quantities of autochthonous organic matter (benthic algae and aquaticmacrophytes) and allochthonous organic matter (wind-blown terrestrial plantdebris) support a high secondary littoral benthic biomass. This is importantfor the aquatic vertebrates (atherinid fish and ambystomatid salamanders),which sustain a local fishery.
Acknowledgments
We are grateful to Drs. Bohdan Bilyj (Chironomidae), David B. Herbst(Ephydridae), Ken J. Tennessen (Odonata), Robert D. Waltz (Baetidae),Ra1ph O. Brinkhurst and Mercedes Marchese (Oligochaeta) for providingidentifications. This investigation was partially supported by CONACyT(Project 25430-T) and DGAPA, UNAM (Project PAPIIT-IN204597). We area1so indebted for field and laboratory facilities provided by M.R. Sánchez, S.

107
Estrada, M. Ubeda, J.C. Tavera, M.J. MoDtoya, M.M. Chávez, Y. MoliDa aDdL.A. Oseguera. The COmmeDts and suggestioDS of Prof. W.D. Williams are
widely appreciated.
References
Aladin, N.V. and Plotnikov, I.S. 1993. Large saline lakes of the fonner USSR: A summaryreview. Hydrobiologia 267: 1-12.
Alcocer, I. 1995. Análisis holístico de la comunidad de macroinvertebrados bentónicoslitorales de seis lagos-cráter con un gradiente de salinidad. Dr. in Sc. thesis. Facultad deCiencias, Universidad Nacional Autónoma de México. México, 106 pp.
Alcacer, I. and Escobar, E. 1990. The drying up of the Mexican Plateau axalapazcos. Salinet4: 34--36.
Alcocer, I., Lugo, A., Estrada, S., Ubeda, M. and Escobar, E. 1993a. La macrofauna bentónicade los axalapazcos mexicanos. Actas del VI Congreso Español de Limnología 33: 409-415.
Alcocer, I., Lugo, A., Estrada, S., Ubeda, M. and Escobar, E. 1993b. Littoral chironornids ofa Mexican Plateau athalassohaline lake. Verh. Intemat. Verein. Limnol. 25: 444-447.
Alcocer, I., Lugo, A., Sánchez, M.R., Chávez, M. and Escobar, E. 1998. Threats to the salinelakes of the Oriental basin, Mexico, by human activities. Verh. Intemat. Verein. Limnol.26: 1383-1386.
Alcocer, I., Lugo, A., Escobar, E. and Sánchez, M. 1997. The macrobenthic fauna of a fonnerperennial and now episodically filled Mexican saline lake. Int. I. Salt Lake Res. 5: 261-274.
Alcocer, I. and Williams, W.D. 1993. Lagos salinos Mexicanos. In: S.I. Salazar-Vallejo andN.E. González (Eds) Biodiversidad Marina y Costera de México, pp. 849-865. Com. Nal.Biodiversidad y CIQRO, Mexico.
APHA 1985. Standard Methods for the Exarnination of Water and Wastewater. AmericanPublic Health Assaciation, New York, 1193 pp.
Arredondo-Figueroa, I.L., Borrego-Enríquez, L.E., Castillo-Domínguez, R.M. and Valladolid-Guerrero, M.A. 1983. Batimetría y morfometría de los lagos "maars" de la cuenca deOriental, Puebla, México. Biotica 8(1): 37-47.
Bayly, I.A.E. and Williams, W.D. 1966. Chemical and biological studies on some saline lakesof south-east Australia. Aust. I. Mar. Freshwat. Res. 17: 177-228.
Colbum, E.A. 1988. Factors influencing species diversity in saline waters of Death Valley,USA. Hydrobiologia 158: 215-226.
Dejoux, C. 1993. Benthic invertebrates of some saline lakes of the Sud Lipez region, Bolivia.Hydrobiologia 267: 257-267.
Egorov, A.N. 1993. Mongolian salt lakes: some features of their geography, thennal pattems,chernistry and biology. Hydrobiologia 267: 13-21.
Galat, D., Lider, E.L., Vigg, S. and Robertson, S.R. 1981. Limnology of a large, deep, NorthAmerican terrninallake, pyrarnid Lake, Nevada, U.S.A. Hydrobiologia 82: 281-317.
García, E. 1988. Modificaciones al Sistema de Clasificación Climática de Koppen (paraAdaptarlo a las Condiciones de la República Mexicana). E. García. México, 217 pp.
Gasca, A. 1981. Algunas notas de la génesis de los lagos cráter de la Cuenca de Oriental.Puebla- Tlaxcala- Veracruz. Colección Científica Prehistoria 98. Inst. Nal. Antrop. Hist.Méx. México. 55 DD.

108
,
Geddes, M.C., de Deckker, P., Williams, W.D., Morton, D.W. and Topping, M. 1981. Onthe chernistry and biota of some saline lakes in Western Australia. Hydrobiologia 82:201-222.
Harnrner, U.T. 1986. Saline Lake Ecosystems ofthe World. W.Junk, Dordrecht, 616 pp.Harnrner, U.T., Haynes, R.C. Haseltine, J.M. and Swanson, S.M. 1975. The sa1ine lakes of
Saskatchewan. Verh. Internat. Verein. Limnol. 19: 589-598.Harnrner, U. T. and Heseltine, M. 1988. Aquatic macrophytes in sa1ine lakes of the Canadian
prairies. Hydrobiologia 158: 101-116.Harnrner, U.T., Sheard, J.S. and Kranabetter, J. 1990. Distribution and abundance of littoral
benthic fauna in Canadian prairie saline lakes. Hydrobiologia 197: 173-192.Heine, K. 1973. Variaciones climáticas más importantesdurante los últimos 40,000 años en
México. Comunicaciones Proyecto Puebla- Tlaxca1a 7: 51-57 .Lugo, A., Alcocer, J., Sánchez, M.R. and Escobar, E. 1993. Trophic status of tropicallakes
indicated by littora1 protozoan assemblages. Verh. Intemat. Verein. Limnol. 25: 441-443.Ramírez-García, P. and Vázquez-Gutiérrez, F. 1989. Contribuciones al estudio lirnnobotánico
de la zona litoral de seis lagos cráter del estado de Puebla. An. Inst. Cienc. Mar y Limnol.Univ. Na1. Autón. Méx. 16: 1-16.
Rawson, D.A. and Moore, J.E. 1944. The saline lakes of Saskatchewan. Can. J. Res. 22: 141-201.
Santinel1i, M.A. 1995. Historia natural de la sa1amandra de Alchichica, Puebla (Caudata:Ambystomatidae) Ambystoma taylorii. Undergraduate Biology thesis. Universidad delVa1le de México, Mexico, 125 pp.
Timms, B.V. 1981. Anima1 communities in three Victorian lakes of differing salinity.Hydrobiologia 81: 181-193.
Timms, B.V. 1983. A study ofbenthic communities in some sha1low sa1ine lakes ofWesternVictoria, Austra1ia. Hydrobiologia 105 : 165-177.
Timms, B. V. 1993. Saline lakes of the Paroo, inland New South Wa1es, Austra1ia. Hydrobiolo-gia 267: 269-289.
Timms, B. V. 1985. The structure of macrobenthic communities of Austra1ian lakes. Proc. Ecol.Soc. Aust. 14: 51-59.
Timms, B.V., Harnrner, U.T. and Sheard, J.W. 1986. A study ofbenthic communities in somesa1ine lakes in Saskatchewan and Alberta, Canada. Int. Revue. ges. Hydrobiol. 71: 759-777.
Tudorancea, C. and Harrison, A. 1988. The benthic communities of the saline lakes Abijataand Sha1a (Ethiopia). Hydrobiologia 158: 117-123.
Vilaclara, G., Chávez, M. Lugo, A., González, M. and Gaytán, M. 1993. Comparative descrip-tion of crater-Iakes basic chernistry in Puebla State, Mexico. Verh. Internat. Verein.Lirnnol. 25: 435-440.
Weber, C.I. (ed) 1973. Biological field and laboratory methods for measuring the qua1ity ofsurface waters and effluents. Macroinvertebrates. U.S. Environmenta1 Protection Agency,Cincinnati, 38 pp.
Williams, W.D. 1972. The uniqueness of salt lake ecosystems. In: Z. Kajak and A. Hillbricht-I1kowska (Eds) Productivity Problems of Freshwáters, pp. 349-361. Proceedings of theIBP-UNESCO Symposiurn. Polish Scientific Publishers, Warsow.
Williams, W.D., Boulton, A.J. and Taaffe, R.G. 1990. Salinity as a determinant of sa1t lakefauna: a question of sca1e. Hydrobiologia 197: 257-266.