leiomodins: larger members of the tropomodulin (tmod) gene family
TRANSCRIPT

Genomics 73, 127–139 (2001)doi:10.1006/geno.2000.6501, available online at http://www.idealibrary.com on
Leiomodins: Larger Members of the Tropomodulin (Tmod)Gene Family
Catharine A. Conley,*,1 Kimberly L. Fritz-Six,† Angels Almenar-Queralt,†and Velia M. Fowler†
*Space Life Sciences, MS 239-11, NASA Ames Research Center, Moffett Field, California 94035-1000; and †Department of CellBiology, MB24, The Scripps Research Institute, 10550 N. Torrey Pines Road, La Jolla, California 92037
Received June 5, 2000; accepted January 8, 2001
The 64-kDa autoantigen D1 or 1D, first identified asa potential autoantigen in Graves’ disease, is similar tothe tropomodulin (Tmod) family of actin filamentpointed end-capping proteins. A novel gene with sig-nificant similarity to the 64-kDa human autoantigenD1 has been cloned from both humans and mice, andthe genomic sequences of both genes have been iden-tified. These genes form a subfamily closely related tothe Tmods and are here named the Leiomodins(Lmods). Both Lmod genes display a conserved intron–exon structure, as do three Tmod genes, but the in-tron–exon structure of the Lmods and the Tmods isdivergent. mRNA expression analysis indicates thatthe gene formerly known as the 64-kDa autoantigenD1 is most highly expressed in a variety of humantissues that contain smooth muscle, earning it thename smooth muscle Leiomodin (SM-Lmod; HGMW-approved symbol LMOD1). Transcripts encoding thenovel Lmod gene are present exclusively in fetal andadult heart and adult skeletal muscle, and it is herenamed cardiac Leiomodin (C-Lmod; HGMW-approvedsymbol LMOD2). Human C-Lmod is located near thehypertrophic cardiomyopathy locus CMH6 on humanchromosome 7q3, potentially implicating it in this dis-ease. Our data demonstrate that the Lmods are evolu-tionarily related and display tissue-specific patternsof expression distinct from, but overlapping with, theexpression of Tmod isoforms. © 2001 Academic Press
INTRODUCTION
The tropomodulin (Tmod)2 protein family consists offour ;40-kDa protein isoforms that are highly con-served among vertebrates and invertebrates, but showno similarity to other known protein families (Conley et
1 To whom correspondence should be addressed. Telephone: (650)604-0234. Fax: (650) 604-3954. E-mail: [email protected].
2 Abbreviations used: 64kD D1, human 64-kDa autoantigen D1;C-Lmod, cardiac leiomodin; EST, expressed sequence tag; E-Tmod,erythrocyte tropomodulin; N-Tmod, neural tropomodulin; Sk-Tmod,skeletal muscle tropomodulin; SM-Lmod, smooth muscle leiomodin;TM, tropomyosin; Tmod, tropomodulin; U-Tmod, ubiquitous tropo-modulin.
127
al., 1998; Almenar-Queralt et al., 1999; Cox andZoghbi, 2000). The original Tmod was identified as a;40-kDa tropomyosin-binding protein in erythrocytesthat was shown to modulate the binding of tropomyo-sin (TM) to actin filaments (Fowler, 1987, 1990). Sub-sequently, it was found that this Tmod also bound topure actin filaments and capped the slow-growing(pointed) ends of pure actin or TM–actin filaments withmicromolar or picomolar affinity, respectively (Weberet al., 1994, 1999). This capping activity of Tmod isunique and is essential for cellular function in vivo.Disruption of Tmod function in beating cardiac musclecells, by microinjecting antibodies that inhibit the ac-tin-capping activity of Tmod, leads to new actin poly-merization from the thin filament pointed ends andprevents the cells from beating (Gregorio et al., 1995).Transgenic mice overexpressing Tmod in the heart de-velop myofibril disorganization and dilated cardiomy-opathy (Sussman et al., 1998).
All Tmod isoforms are of nearly the same length anddisplay very few gaps and insertions relative to oneanother. However, Tmod isoforms display distinct pat-terns of tissue-specific expression and are differentiallyregulated during development (Watakabe et al., 1996;Almenar-Queralt et al., 1999; Cox and Zoghbi, 2000).The first Tmod isoform identified is termed erythro-cyte-Tmod (E-Tmod) for its initial purification fromhuman erythrocytes (Fowler, 1987). E-Tmod (genename TMOD1) is highly expressed in striated musclesand at lower levels in a number of nonmuscle tissues(Sung et al., 1992; Ito et al., 1995), including the lens ofthe eye (Fischer et al., 2000). Neural-Tmod (N-Tmod;TMOD2) was initially identified in rat brain (Watak-abe et al., 1996) and is expressed almost exclusively inembryonic and adult neural tissue (Watakabe et al.,1996; Cox and Zoghbi, 2000). Skeletal-Tmod (Sk-Tmod;TMOD4) was first purified from chicken breast muscleand is expressed in fast skeletal muscle fibers, lens,and erythrocytes in adult chickens, but is not presentin embryonic skeletal muscle or heart (Almenar-Quer-alt et al., 1999). Ubiquitous-Tmod (U-Tmod; TMOD3)was originally cloned from a set of ESTs conservedbetween humans, mice, and pigs, and it displays broad
0888-7543/01 $35.00Copyright © 2001 by Academic Press
All rights of reproduction in any form reserved.

Cp
tcu
ostams(ifLsrenL(pt
128 CONLEY ET AL.
representation in diverse tissues (Conley et al., 1998;Cox and Zoghbi, 2000). The Tmod proteins are con-served at greater than 80% similarity for the sameisoform in different vertebrate species, but the fourdifferent isoforms from a single species are conservedonly at about 60% similarity. This high level of se-quence conservation for each Tmod isoform betweendiverse species suggests substantial conservation ofbiochemical functions.
The only other known vertebrate protein that showssignificant amino acid sequence similarity to the 40-kDa Tmods is a protein that has been called “the hu-man 64-kDa autoantigen D1” or “1D” (Wall et al., 1993;Sung et al., 1996; Conley and Fowler, 1996; Conley etal., 1998; Fowler and Conley, 1999; Conley, in press),which we refer to as 64kD D1. The 64kD D1 protein issimilar to the 40-kDa Tmods in two regions: the amino-terminus of 64kD D1 shows a high degree of similarityto most of the amino-terminal region of E-Tmod thatpreferentially binds muscle TM, and a central region of64kD D1 is highly similar to the entire carboxy-termi-nal half of the 40-kDa Tmods where the putative actin-capping domain is located (Conley and Fowler, 1996;Sung et al., 1996; reviewed in Fowler, 1997; Fowler and
onley, 1999). 64kD D1 was originally identified as aotential autoantigen in Graves’ disease (Dong et al.,
1991). Initial analyses by Northern blot and quantita-tive PCR indicated that 64kD D1 mRNA was morehighly expressed in thyroid than extraocular muscle orother tissues from dog or human (Dong et al., 1991;Zhang et al., 1996). However, by immunoblot analysis,the 64kD D1 protein is clearly expressed in a subset ofextraocular muscle fibers in rat and rabbit, but 64kDD1 protein is barely detectable in rat or rabbit thyroid(Conley and Fowler, 1999). Patients with thyroid-asso-ciated ophthalmopathy display elevated levels of auto-immune antibodies that recognize 64kD D1 (Krom-minga et al., 1998), but as yet, the relationship betweenhe expression of 64kD D1 protein in extraocular mus-les and thyroid-associated ophthalmopathy is not wellnderstood.In this paper, we report the initial characterization
f a novel gene that is related to 64kD D1 and con-erved between humans and mice. We also report theissue-specific expression, in humans, of this new genend the 64kD D1 gene, in comparison with knownembers of the Tmod family. We have named this
ubfamily of larger Tmod-related proteins “Leiomodin”Lmod), from the Greek “leio-” for “smooth,” and remov-ng the Latin diminutive “-ul-” from “-modulin.” Theormer 64kD D1 will be referred to as smooth muscle-mod (SM-Lmod), based on its predominant expres-ion in smooth muscle, and the novel gene will beeferred to as cardiac-Lmod (C-Lmod), based on itsxpression in heart. SM-Lmod was assigned the geneame LMOD1, and C-Lmod was assigned the nameMOD2, by the HUGO Nomenclature Committee
http://www.gene.ucl.ac.uk/nomenclature/). The dataresented here demonstrate that the Lmods are likelyo be the result of a gene duplication event and that
they are expressed in distinct subsets of tissues over-lapping with those in which Tmods are expressed andbring the total number of mammalian Tmod familymembers to six.
MATERIALS AND METHODS
Identification of novel Lmod and Tmod transcripts. A detaileddescription of the method used to identify novel Tmod isoform tran-scripts is presented elsewhere (Conley et al., 1998; C. A. Conley,manuscript in preparation). Briefly, the protein sequences of knownTmod isoforms were used to search the Database of Expressed Se-quence Tags (dbEST), and ESTs that were similar but not identicalto known Tmod sequences were identified. EST sequences were thencompiled into contigs with the program Sequencher (Gene CodesCorp., Ann Arbor, MI), and cDNAs corresponding to individual ESTswere selected. The human cDNA clone 271180 from the IMAGEConsortium, corresponding to human U-Tmod, EST N34535, and themouse cDNA clones 876920 and 315826 from the IMAGE Consor-tium, corresponding to SM-Lmod, EST AA497562, and C-Lmod, ESTW12756, respectively, were ordered from Genome Systems (St.Louis, MO). Exon 2 of human C-Lmod was obtained by PCR fromhuman genomic DNA (Clontech, Palo Alto, CA) and cloned into pCR4.0 using the TOPO-TA cloning system (Invitrogen Corp., Carlsbad,CA). The full-length open reading frame of mouse C-Lmod wasobtained by RT-PCR from mouse heart total RNA (Clontech) andcloned using the TOPO-TA cloning system. Plasmid DNAs werepurified with the Qiagen (Valencia, CA) Qiaprep miniprep purifica-tion system and sequenced using a Model 377 ABI automated se-quencer. Sequences were assembled and edited using the programSequencher.
Genomic sequences potentially encoding Lmod genes were identi-fied by searching the Genome Survey Sequences (GSS) and HighThroughput Genome Sequencing (HTGS) portions of GenBank usingboth the BLASTN and the TBLASTX algorithms. The human C-Lmod genomic sequence is contained in GenBank entry AC006333(R. H. Waterston, unpublished results), and the partial human SM-Lmod genomic sequence is contained in Genbank entry AC025664(B. Birren et al., unpublished results). The mouse SM-Lmod genomicsequence is contained in GenBank entry AC016814 (B. Birren et al.,unpublished results). There is a gap in the mouse SM-Lmod genomicsequence that falls within intron 1, but all exon sequences arepresent, based on alignments with the human SM-Lmod cDNA se-quence. Shortly before this paper was submitted, three sequencesthat show $99% identity with human C-Lmod, Accession Nos.AX002051–AX002053, were released into GenBank as part of apatent entitled “Heart and Skeleton Muscle Specific Nucleic Acid,the Production and Use Thereof” (M. E. Hoffman and T. Henkel). Thesource organism for these sequences is unidentified, but based on thehigh level of sequence similarity to human C-Lmod, they are likely tobe human in origin.
The human genomic sequences for E-Tmod, N-Tmod, and U-Tmodwere obtained by searching GenBank and HTGS with their respec-tive cDNA sequences. The genomic sequence of human E-Tmod isfound on chromosome 9 clone RP11-244N9, Accession No. AL162385(submitted by the Sanger Centre, Hinxton, UK). Exons of both N-Tmod and U-Tmod are present in human chromosome 15 cloneRP11-56B16, Accession No. AC009679 (submitted by the WhiteheadInstitute, Cambridge, MA), indicating that these genes must belocated within 184 kb of each other on human chromosome 15.N-Tmod genomic sequences are also present on chromosome 15 cloneRP11-414J1, Accession No. AC020685 (submitted by the WhiteheadInstitute), and clone CTD-2308G16, Accession No. AC026770 (sub-mitted by the Multimegabase Sequencing Center, University ofWashington, Seattle, WA). U-Tmod genomic sequences are alsopresent on clone RP11-47K1, Accession No. AC016824 (submitted byGenome Therapeutics Corp., Waltham, MA). Identification of in-tron–exon splice junctions was performed by aligning the genomicclones with ESTs and cDNA sequences, as well as by comparingencoded protein sequences. A number of gaps are present in the

S((bfaTLs
3Dsbtt
NiCEsw
t15ftttoqd
TfkkbhDclscwofPssqhsptgSjcSlcaL
ebsqtsiphoCstAf
129LEIOMODINS: MEMBERS OF THE Tmod GENE FAMILY
introns of the Tmod genomic sequences, but the complete sequencesfor all coding exons are present.
Protein sequence comparison. The cDNA sequences for humanM-Lmod (X54162), human E-Tmod (M77016), human N-Tmod
AF177169), human Sk-Tmod (AF165217), and human U-TmodAF237631) were obtained from GenBank under the accession num-ers given. The partial cDNA sequences of mouse SM-Lmod and theull-length sequence of mouse C-Lmod were submitted to GenBanknd given Accession Nos. AF237627 and AF237628, respectively.he full-length cDNA sequences of human C-Lmod and mouse SM-mod were predicted from genomic sequences and used in furtherequence analysis. Prediction of pI and other protein characteristics
was performed using the ProtParam tool of the Swiss Institute ofBioinformatics at http://expasy.cbr.nrc.ca/tools/protparam.html.Pairwise sequence comparisons were performed using the BLASTalgorithm implemented by the National Center for BiotechnologyInformation (www.ncbi.nlm.nih.gov). All cDNA sequences were en-tered into the program DNAStrider (C. Marck, CEA, Saclay, France)for protein translation, and the translated amino acid sequenceswere multiply aligned using the ClustalW algorithm implemented bythe program SeqPup (D. Gilbert, Indiana University), and align-ments were finalized by manual inspection. Multiple alignmentsused for generating the figures shown in this report were derivedusing standard parameters, but no differences in the alignment ofvertebrate isoforms were observed when parameters were altered.
RNA blot analysis. Isolation of human E-Tmod and Sk-TmodcDNAs is described elsewhere (Sung et al., 1992; Almenar-Queralt etal., 1999). A 2.6-kb human E-Tmod probe containing the full-lengthcoding region and a partial 39 UTR was excised from pGEX-KG withNcoI and HindIII. A partial 1.1-kb Sk-Tmod probe missing both the59 UTR and nucleotides coding for the first 68 amino acids wasexcised with EcoRI and XhoI from pBluescript SK(2) (IMAGE clone628632, Accession No. AA192195). A 1.4-kb human U-Tmod probecontaining the coding region and portions of the 59 and 39 UTRs wasexcised with EcoRI and NotI from pT7T3D (IMAGE clone 271180,Accession No. N34535). A 1.9-kb fragment of SM-Lmod was ampli-fied by PCR from pET64 (Conley and Fowler, 1999) using the T7 andT3 primers flanking the full-length coding region insert. A 1.3-kbhuman C-Lmod exon 2 probe was excised from pCR 4.0 with EcoRI.Digests were separated by agarose gel electrophoresis and blotted toa DE81 membrane. Insert fragments were eluted overnight at 65°Cin 1 M NaCl in TE (pH 7.5), extracted in phenol–chloroform, andethanol precipitated. 32P-labeled probes were prepared by incubating0 ng of DNA with 50 mCi of [32P]dCTP (DuPont-NEN, Wilmington,E) at room temperature for 1 h using the Prime-a-Gene labeling
ystem (Promega, Madison, WI) as described in the protocol suppliedy the manufacturer. The 32P-labeled probes were then purifiedhrough two Microspin G-50 columns (Amersham–Pharmacia Bio-ech, Piscataway, NJ).
For tissue-level expression analyses, a Human Multiple Tissueorthern Blot and a Human Master Blot array containing RNA
solated from a variety of human tissues were purchased fromlontech. The human RNA blots were prehybridized with 10 ml ofxpressHyb solution (Clontech) containing 1.5 mg of denaturedalmon sperm DNA at 65°C for 4 – 6 h. The blots were hybridizedith 5 ml of ExpressHyb containing 30 mg of Cot-1 DNA, 150 mg of
denatured salmon sperm DNA, 50 ml of 203 SSC, and 1.5 3 107
cpm of 32P-labeled probe overnight at 65°C. After hybridization,he membranes were washed four times with 23 SSC containing% SDS at 65°C and twice with 0.13 SSC containing 0.5% SDS at5°C for 20 min each. The membranes were exposed to X-ray filmor 4 – 48 h at 280°C with an intensifying screen. The blots werehen stripped and reprobed as described in the Clontech Labora-ories protocol (PT3004-1). Relative expression levels were ob-ained from the Human Master Blot by quantifying the intensityf each dot in arbitrary units using NIH Image software. Ouruantitation method was verified using a ubiquitin control asescribed in the Clontech Laboratories protocol.
RESULTS
Identification of Novel Tmod-like Genes
The rapidly growing genome databases and ex-pressed sequence databases provide a rich source foridentifying additional isoforms of proteins found usingconventional molecular biology or biochemical ap-proaches. To determine whether Tmod-related genesthat had not previously been identified were repre-sented in dbEST, we developed a database screeningmethod using the available Tmod-related sequencesand used it to identify additional EST sequences thatencoded potential Tmod isoforms (Conley et al., 1998).
his screen identified a number of distinct EST classesrom a variety of species, most of which encoded 40-Da Tmod proteins. Of those that did not encode 40-Da Tmods, one class of mouse ESTs encoded the car-oxy-terminus of a protein showing 96% identity to theuman gene encoding the human 64-kDa autoantigen1, hereafter referred to as SM-Lmod, so this EST
lass was considered to be the mouse SM-Lmod homo-ogue. None of the clones we sequenced that corre-ponded to this novel EST class carried full-lengthDNAs (Fig. 1). All the mouse SM-Lmod cDNAs thate obtained from the IMAGE consortium containednly partial coding sequences, and efforts to obtain aull-length mouse SM-Lmod cDNA sequence by RT-CR were unsuccessful, perhaps because this tran-cript is predicted to fold into an extensive secondarytructure (data not shown). However, genomic se-uences encoding both mouse and human SM-Lmodave been submitted to GenBank as part of the genomeequencing projects. Therefore, the mouse SM-Lmodrotein sequence we present here is based on a puta-ive open reading frame predicted from the mouseenomic sequence, based on the structure of the humanM-Lmod gene and mouse ESTs that span the splice
unctions. Because Tmod genes in general are highlyonserved between species, and the human and mouseM-Lmod sequences we present are quite similar, it is
ikely that the predicted mouse SM-Lmod sequence isorrect. However, there is a formal possibility that andditional intron may be present in the mouse SM-mod mRNA that we have not discovered.An additional novel class of human and mouse ESTs
ncoded amino acid sequences that were 98% identicaletween the two species. However, these ESTs corre-ponded to partial cDNAs encoding amino acid se-uences that were only 39% identical and 67% similaro the carboxy-terminus of human SM-Lmod andhowed no significant similarity to any other proteinsn GenBank. This EST class was concluded to be theroduct of a novel Lmod-like gene conserved betweenumans and mice, which we have named C-Lmod. Webtained the full-length open reading frame of mouse-Lmod by RT-PCR with primers selected from ESTequences and our mouse partial cDNA, and we usedhat sequence to identify human C-Lmod ESTs (Fig. 1).dditionally, as this paper was being submitted, three
ull-length cDNAs of unspecified origin were released

CGiqL
g(scctaiab7Iitnqcbb
lqth
tPighmhmlh(dtLmasfibas
130 CONLEY ET AL.
in GenBank that show $99% identity with the human-Lmod ESTs we identified (Fig. 1, “patent cDNAs”).enomic sequences for the human C-Lmod gene were
dentified using a search with the C-Lmod protein se-uences; however, genomic sequences for mouse C-mod have not yet been submitted to GenBank.Both the human C-Lmod and the mouse SM-Lmod
enes contain three exons spanning the coding regionFig. 1). Exon 1 contains the 59 UTR and the firsteveral hundred bases of the coding region, exon 2ontains the majority of the coding region, and exon 3ontains the last ;25 bases of the coding region andhe 39 UTR. The DNA sequences encoding the humannd mouse SM-Lmod transcripts are more than 80%dentical over their entire length, as are the humannd mouse C-Lmods, but the most conserved regionetween human SM-Lmod and human C-Lmod is only3% identical, and this region is only 275 bases long.ntron 1 of both human C-Lmod and mouse SM-Lmods greater than 5 kb, and intron 2 of both genes is lesshan 1 kb, although the precise length of one intron isot known because there is a gap in the genomic se-uence. However, the exon–exon splice junctions in theoding regions of mouse SM-Lmod are well defined,ecause our cDNA sequences or ESTs in dbEST spanoth exon splice sites in each gene (Fig. 1). The simi-
FIG. 1. Diagram of Lmod genes and transcripts. (A) SM-Lmodgenomic region, and mouse ESTs. (B) C-Lmod: mouse cDNA, humanmouse ESTs. The primers used to amplify exon 2 of the human C-Lmin A and B are indicated as gray boxes, and the length in nucleotidesbelow the box, and the length of the individual exons is indicated ra
arity in intron–exon structures as well as coding se-uences indicate that SM-Lmod and C-Lmod are likelyo be evolutionarily related genes conserved betweenumans and mice.The human SM-Lmod gene was previously mapped
o chromosome 1q32 as part of the Human Genomeroject (see http://www.ncbi.nlm.nih.gov/Genomes/
ndex.html). Human contig AC006333, containingenomic sequences encoding the human C-Lmod gene,as been mapped to chromosome 7q31–q32. Theouse contig AC016814, containing mouse SM-Lmod,
ad not been mapped at the time this paper was sub-itted; however, we have predicted the chromosomal
ocations for both mouse genes based on large-scaleomologies between mouse and human chromosomesTable 1). The SM-Lmod gene has been assigned theesignation LMOD1 (human) or Lmod1 (mouse), andhe C-Lmod gene has been assigned the designationMOD2/Lmod2, by the Genome Nomenclature Com-ittee (http://www.gene.ucl.ac.uk/nomenclature/). Ex-
mination of these regions using LocusLink identifiedeveral human disease loci with no previously identi-ed candidate genes (see Discussion). Accession num-ers, Unigene ID numbers, chromosomal locations,nd linked human diseases are shown in Table 1. Aummary of information about the Tmod gene family is
man cDNA and partial genomic region, mouse partial cDNA andnomic region, putative human cDNA from patent, and human andene are indicated by small arrows in the ORF. Open reading framesndicated. For genomic sequences, the size of the introns is indicatedr than the length of the entire ORF.
: huge
od gis ithe

i(etp
ortCf5ip
fcrfrtttmeCbdsdt
C
fifsPtdasrttttfqg
131LEIOMODINS: MEMBERS OF THE Tmod GENE FAMILY
maintained at the Web site http://lifesci.arc.nasa.gov/conley/tmod.
Comparison of Lmod Protein Sequences
The full-length amino acid sequence of mouse SM-Lmod, obtained by translating the predicted cDNA,shows 82% identity and 90% similarity to the pub-lished human SM-Lmod protein sequence (Fig. 2). Thepredicted sequences of full-length human and mouseC-Lmod proteins are 89% identical and 94% similar(Fig. 2). However, the SM-Lmods are only 32% identi-cal and 45% similar to the C-Lmods. Curiously, bothSM-Lmod and C-Lmod display much lower levels ofsimilarity between the mouse and the human se-quences in a region from aa 85 to 305 of the multiplealignment. Human SM-Lmod and mouse SM-Lmod areonly 77% similar in this region, while in human andmouse C-Lmod the region displays 86% amino acidsimilarity. In both isoforms, this region is highlycharged, with human and mouse C-Lmod having apredicted pI of 3.9 over ;103 aa, while the SM-Lmodsoforms display a predicted pI of 9.3 (mouse) or 9.6human) over ;215 aa. This dramatic charge differ-nce, as well as the different lengths of this region inhe two isoforms, strongly implicates this region as aotential site of isoform-specific functional differences.When the SM-Lmods and C-Lmods are compared to
ne another, the SM-Lmods display a large insertionelative to the C-Lmods in the amino-terminal half ofhe protein, at aa 85–305 of the multiple alignment.-Lmod contains both an amino-terminal extension,
rom aa 1 to 30 and a carboxy-terminal insertion at aa26–613 of the multiple alignment (Fig. 2). Intrigu-ngly, both SM-Lmod and C-Lmod contain longolyproline sequences near their carboxy-termini, the
TAB
Gene Names, GenBank Accessionand Linked Diseases fo
SM-Lm
Gene names and Accession Nos.Human LMOD1
cDNA X54162Genomic AC025664
Mouse Lmod1cDNA AF237627 (partial)Genomic AC016814
Unigene ID Hs.79386Locuslink ID 25802Chromosome location
Human D1S2622–D1S3061q32
Mouse (predicted) 1, 69–75 cMHuman disease loci GFND (glomerulop
PPS (popliteal pterCMD1D (dilated ca
Note. The Gene Nomenclature Committee has assigned the gene nnames LMOD2 or Lmod 2 to C-Lmod. The cDNAs listed as human cDNsequences show $99% identity with the human genomic sequence, inbeen mapped in humans, and the map locations in mouse have been
unction of which is currently unclear. Also, the Lmodsontain several small gaps or insertions in the mouseelative to the human sequences, suggesting that theolded structure of these proteins, at least in someegions, may not be highly constrained during evolu-ion. However, when the exon splice sites are located inhe protein sequence, the first junction is located onlyhree amino acids apart in mouse SM-Lmod and hu-an C-Lmod, and the second junction is present at
xactly the same place in mouse SM-Lmod and human-Lmod (triangles in Fig. 2). This conservation of exonoundary locations, in proteins displaying significantifferences in sequence conservation, suggests thatome of the sequence divergence could arise from ran-om drift in portions of the proteins that are not func-ionally constrained.
omparison of Lmod and Tmod Sequences
During the course of our database screen, we identi-ed the human genomic sequences of three Tmod iso-orms, E-Tmod, N-Tmod, and U-Tmod, that had beenubmitted to GenBank as part of the Human Genomeroject. A comparison of the intron–exon structure ofhe Tmods relative to the Lmods reveals significantifferences (Fig. 3). The genomic structures of humannd mouse E-Tmod were previously reported to beimilar (Chu et al., 2000) and are consistent with theesults we obtained. The coding region for each of thehree Tmod genes is carried on nine short exons, andhe size of each coding exon is conserved among allhree Tmod genes. The genomic sequences containinghe Tmod genes were submitted as multiple sequenceragments from the same clone, and gaps in the se-uence fall within many of the introns, although noaps were present in the exon sequences. This makes it
1
umbers, Chromosomal Location,M-Lmod and C-Lmod
C-Lmod
LMOD2AX002051-3 (from patent)AC006333Lmod2AF237628Not availableHs.25595043850
D7S655–D7S6867q316, 3–11 cM
y) CMH6 (hypertrophic cardiomyopathy)um) SPCH1 (speech disorder)omyopathy) DFNB14/17 (recessive deafness)
es LMOD1 (human) and Lmod1 (mouse) to SM-Lmod and the gene(patent) give no source species in the GenBank entry; however, theseating that they are probably of human origin. The Lmod genes haveedicted on the basis of large-scale chromosomal homologies.
LE
Nr S
od
athygirdi
amA
dicpr

scb
ttLwdli
estaoedoT
oa(ttsotTs
hns
132 CONLEY ET AL.
impossible to determine the exact size of the intronsand thus the total length of the human Tmod genomicsequences. However, by adding all the available se-quence fragments, it can be estimated that the lengthof the E-Tmod genomic region is greater than 15 kb,and the length of each of the N-Tmod and U-Tmodgenomic regions is at least 40 kb. The large size and thepresence of nine or more exons in the Tmod genes arein dramatic contrast to the three exons and ;10-kbize of the Lmod genes. These data represent the firstomparison of intron–exon structure between verte-rate Tmod family genes.Alignment of the four human Tmod proteins with the
wo human Lmod proteins (Fig. 4) reveals several in-eresting features of this protein family. Unlike SM-mod, the amino-terminus of C-Lmod aligns very wellith the amino-termini of the 40-kDa Tmods, and in-eed the sequence of C-Lmod aligns with the entireength of the 40-kDa Tmods with only a few smallnsertions, suggesting that C-Lmod could be consid-
FIG. 2. Multiple alignment of predicted amino acid sequencesuman and mouse C-Lmod. Numbers along the top of each row of thumbers to the right of each protein sequence indicate the positionplice junctions. Box threshold is 60% of all sequences conserved at
red a 40-kDa Tmod with a carboxy-terminal exten-ion. The long internal insertion in SM-Lmod relativeo C-Lmod and the 40-kDa Tmods falls within a region,a ;95–;180 of the 40-kDa Tmods, that was previ-usly recognized as divergent in the Tmods (Watakabet al., 1996; Cox and Zoghbi, 2000). This substantialivergence suggests that this region could be a sourcef isoform-specific functional differences within themod family.When the locations of the exon–exon splice junctions
f the Lmods and Tmods are mapped onto the multiplelignment, several additional features can be observedshading in Fig. 3, triangles in Fig. 4). The start me-hionine of SM-Lmod aligns near the first splice junc-ion in the coding region of the Tmods, while the firstplice junction of the Lmods falls within 10 amino acidsf the Tmods’ second splice junction and at the end ofhe conserved amino-terminal putative skeletal muscleM-binding domain. In the Tmods, the fourth exonplice junction falls shortly before the start of the cen-
full-length human and predicted mouse SM-Lmod and full-lengthlignment indicate the position within the multiple alignment, whilehin each sequence. The triangles indicate the location of exon–exont site.
fore awittha

akcCCf
otatr
TmLtlfoaSapmLs
TSwpBet
1i1
133LEIOMODINS: MEMBERS OF THE Tmod GENE FAMILY
tral/carboxy-terminal region that is conserved betweenthe Lmods and the Tmods, and the last exon splicejunction within the coding region of the Tmods falls atthe very end of this conserved region. Such a correla-tion between regions of the Tmod and Lmod proteinsand the exon splicing structure of the Tmods suggeststhat clusters of exons may be grouped into functionalcassettes.
mRNA Expression Analysis of the Lmods and SeveralTmod Isoforms
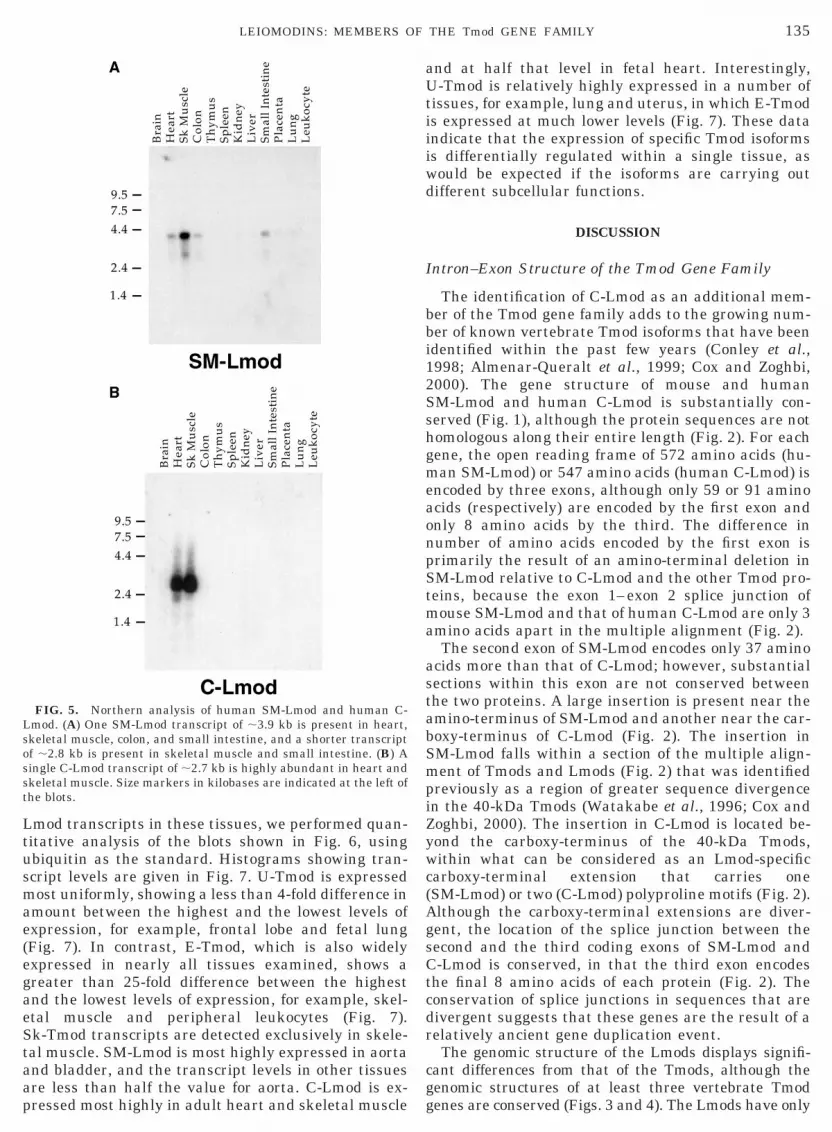
To determine the transcript size classes of the hu-man Lmods, we probed multiple tissue Northern blotscontaining transcripts from a variety of human tissues(Fig. 5). On Northern blots probed for SM-Lmod, onemajor transcript of ;3.9 kb is detected in heart, skel-etal muscle, colon, and small intestine (Fig. 5A). mRNAloaded on these Northern blots was not normalized;thus the relative transcript abundance in differentlanes does not reflect steady-state levels in tissues. TheSM-Lmod transcript size is in agreement with previousreports for SM-Lmod (Dong et al., 1991; Kromminga et
l., 1998). A single human C-Lmod transcript of ;2.7b is expressed exclusively in heart and skeletal mus-le on Northern blots (Fig. 5B). None of the human-Lmod ESTs, nor any of the three putative human-Lmod cDNAs, is longer than 2.2 kb, suggesting that
ull-length human C-Lmod cDNAs have not yet been
FIG. 3. Diagram of Lmod and Tmod exon splice junctions. Exonsin white. Coding regions were given shaded patterns to indicate consexon of SM-Lmods is at least 180 bp, the second exon is 1489 bp, an366 bp, the second exon is 1343 bp, and the third exon is at least 40the exception of the first and last exons (last two exons for U-Tmod).25–75 bp of 59 UTR. The second exon is 155–158 bp, the third is 1109–111 bp, the seventh is 137–142 bp, and the eighth is 145–146 bndividual Tmod isoforms, as well as small shifts in the position of th.3 kb, while the ninth exon of U-Tmod is 583 kb, and the tenth exo
btained. The partial mouse C-Lmod cDNA we ob-ained has a 39 untranslated region of ;500 bases with
poly(A) tail (Fig. 1), which would give a full-lengthranscript of 2.5 kb, suggesting that this clone mayepresent the major full-length transcript in mice.To quantitate the relative levels of Lmod and several
mod transcripts in different tissues, we probed hu-an multiple tissue mRNA dot-blots (Fig. 6). SM-mod is highly expressed in a variety of adult humanissues containing smooth muscle (Fig. 6), as well as atower levels in skeletal muscle, but not detectably inetal tissues. These results are consistent with previ-us reports of SM-Lmod protein in adult rat bladdernd extraocular muscle (Conley and Fowler, 1999) andM-Lmod transcripts in thyroid and muscle (Dong etl., 1991; Zhang et al., 1996). C-Lmod transcripts areresent only in fetal and adult heart and adult skeletaluscle (Fig. 6). These data demonstrate that themods are regulated during development as well as inpecific adult tissues.To compare directly the expression of Lmods and
mods, we used probes specific for human E-Tmod,k-Tmod, and U-Tmod (Fig. 6). [N-Tmod expressionas previously examined in a range of tissues and wasresent only in neural tissue (Watakabe et al., 1996).ecause Lmods are not expressed in brain (Fig. 6), welected not to pursue the expression of N-Tmod fur-her.] Figure 6 shows that, as its name implies,
e indicated by separated boxes, and noncoding regions are indicateded regions in the protein sequences, as indicated in the key. The firste third exon is at least 774 bp. The first exon of C-Lmod is at least
p. The length of the exons is similar for all three Tmod genes, withfirst exon of all Tmods contains 125 bp of coding region and at least25 bp, the fourth is 89–95 bp, the fifth is 128–130 bp, the sixth isith the variability resulting from amino acid insertions/deletions in
plice sites. The ninth exons of E-Tmod and N-Tmod are both at leasts at least 403 bp.
arervd th4 bThe6–1p, we sn i

134 CONLEY ET AL.
U-Tmod is the most widely expressed of all the tran-scripts examined, with detectable expression in everytissue tested, including both fetal and adult tissues.E-Tmod is also expressed in almost every tissue tested;however, it is much more abundant in fetal and adultheart and skeletal muscle. In contrast, Sk-Tmod tran-scripts are detected exclusively in skeletal muscle (Fig.6). This is a more restricted expression pattern than
FIG. 4. Multiple alignment of human Lmods and Tmod. Numbermultiple alignment, while numbers to the right of each protein sequin the Tmod sequences is indicated with black triangles beneath thabove the alignment. The location of the exon 1–exon 2 splice junctioindicated by a horizontal bar linking the two sites. SM-Lmod exhibitswhich covers the region of low similarity for both the Tmods and tpolyproline motifs. Box threshold is 60% of all sequences conserved
was reported previously for chicken Sk-Tmod, which isalso expressed in lens and erythrocytes (Almenar-Queralt et al., 1999). These data greatly expand therange of tissues in which Tmod family members areknown to be expressed (or not) and demonstrate thateach of the Lmod and Tmod proteins has a uniquepattern of expression.
To evaluate the steady-state abundance of Tmod and
ong the top of each row of the alignment indicate position within thee indicate the position within each sequence. The location of intronsignment, while the Lmod introns are indicated with gray trianglesn SM-Lmod is several amino acids removed from that in C-Lmod, asong insertion in the second and third rows of the multiple alignment,Lmods. Both Lmods carry carboxy-terminal extensions containingthat site.
s alence aln ia lheat
12SshgmeaonpStma
astabSmpi
135LEIOMODINS: MEMBERS OF THE Tmod GENE FAMILY
Lmod transcripts in these tissues, we performed quan-titative analysis of the blots shown in Fig. 6, usingubiquitin as the standard. Histograms showing tran-script levels are given in Fig. 7. U-Tmod is expressedmost uniformly, showing a less than 4-fold difference inamount between the highest and the lowest levels ofexpression, for example, frontal lobe and fetal lung(Fig. 7). In contrast, E-Tmod, which is also widelyexpressed in nearly all tissues examined, shows agreater than 25-fold difference between the highestand the lowest levels of expression, for example, skel-etal muscle and peripheral leukocytes (Fig. 7).Sk-Tmod transcripts are detected exclusively in skele-tal muscle. SM-Lmod is most highly expressed in aortaand bladder, and the transcript levels in other tissuesare less than half the value for aorta. C-Lmod is ex-pressed most highly in adult heart and skeletal muscle
FIG. 5. Northern analysis of human SM-Lmod and human C-Lmod. (A) One SM-Lmod transcript of ;3.9 kb is present in heart,skeletal muscle, colon, and small intestine, and a shorter transcriptof ;2.8 kb is present in skeletal muscle and small intestine. (B) Asingle C-Lmod transcript of ;2.7 kb is highly abundant in heart andskeletal muscle. Size markers in kilobases are indicated at the left ofthe blots.
and at half that level in fetal heart. Interestingly,U-Tmod is relatively highly expressed in a number oftissues, for example, lung and uterus, in which E-Tmodis expressed at much lower levels (Fig. 7). These dataindicate that the expression of specific Tmod isoformsis differentially regulated within a single tissue, aswould be expected if the isoforms are carrying outdifferent subcellular functions.
DISCUSSION
Intron–Exon Structure of the Tmod Gene Family
The identification of C-Lmod as an additional mem-ber of the Tmod gene family adds to the growing num-ber of known vertebrate Tmod isoforms that have beenidentified within the past few years (Conley et al.,998; Almenar-Queralt et al., 1999; Cox and Zoghbi,000). The gene structure of mouse and humanM-Lmod and human C-Lmod is substantially con-erved (Fig. 1), although the protein sequences are notomologous along their entire length (Fig. 2). For eachene, the open reading frame of 572 amino acids (hu-an SM-Lmod) or 547 amino acids (human C-Lmod) is
ncoded by three exons, although only 59 or 91 aminocids (respectively) are encoded by the first exon andnly 8 amino acids by the third. The difference inumber of amino acids encoded by the first exon isrimarily the result of an amino-terminal deletion inM-Lmod relative to C-Lmod and the other Tmod pro-eins, because the exon 1–exon 2 splice junction ofouse SM-Lmod and that of human C-Lmod are only 3
mino acids apart in the multiple alignment (Fig. 2).The second exon of SM-Lmod encodes only 37 amino
cids more than that of C-Lmod; however, substantialections within this exon are not conserved betweenhe two proteins. A large insertion is present near themino-terminus of SM-Lmod and another near the car-oxy-terminus of C-Lmod (Fig. 2). The insertion inM-Lmod falls within a section of the multiple align-ent of Tmods and Lmods (Fig. 2) that was identified
reviously as a region of greater sequence divergencen the 40-kDa Tmods (Watakabe et al., 1996; Cox and
Zoghbi, 2000). The insertion in C-Lmod is located be-yond the carboxy-terminus of the 40-kDa Tmods,within what can be considered as an Lmod-specificcarboxy-terminal extension that carries one(SM-Lmod) or two (C-Lmod) polyproline motifs (Fig. 2).Although the carboxy-terminal extensions are diver-gent, the location of the splice junction between thesecond and the third coding exons of SM-Lmod andC-Lmod is conserved, in that the third exon encodesthe final 8 amino acids of each protein (Fig. 2). Theconservation of splice junctions in sequences that aredivergent suggests that these genes are the result of arelatively ancient gene duplication event.
The genomic structure of the Lmods displays signifi-cant differences from that of the Tmods, although thegenomic structures of at least three vertebrate Tmodgenes are conserved (Figs. 3 and 4). The Lmods have only

136 CONLEY ET AL.
three coding exons, the second of which is long, while theTmods have nine coding exons, spaced approximatelyequally within the open reading frame. Of all the Tmodsand Lmods, U-Tmod is the only gene that contains asplice junction within the 39 untranslated region: thismay be a site for alternative splicing as only one of thetwo human U-Tmod cDNA sequences available in Gen-Bank carries this additional exon. The difference in in-tron–exon structure between the Tmods and the Lmodssuggests that the ancestral Lmod split from the ancestralTmod prior to the development of the current genomic
FIG. 6. mRNA dot-blot analysis of human Lmods and Tmods. Thedot. SM-Lmod is highly expressed in a number of tissues containingand adult skeletal muscle. U-Tmod is expressed in all tissues, E-Tmoand Sk-Tmod is expressed exclusively in adult skeletal muscle.
structure. Interestingly, the intron–exon structures oftwo invertebrate Tmod homologues are not similar toeither the vertebrate Tmod genes or each other (Chu etal., 2000). Whether the Lmods lost introns or the Tmodsgained introns, relative to the ancestral gene structure,remains to be established.
Functional Implications
The characteristic biochemical functions of Tmodsare to bind TM and to cap the pointed ends of actin or
agram at the bottom right indicates the source tissue for each mRNAoth muscle, while C-Lmod is expressed only in fetal and adult hearthighly expressed in fetal and adult heart and adult skeletal muscle,
dismod is

137LEIOMODINS: MEMBERS OF THE Tmod GENE FAMILY
TM–actin filaments, and these functions are localizedto different regions of the Tmod molecule. The TM-binding activity of E-Tmod has been localized to twodistinct regions in its amino-terminal portion (Babcockand Fowler, 1994). Skeletal muscle TM binds to theamino-terminal ;90 amino acids of E-Tmod, whilenonmuscle TM binds to a region within amino acids;90 to ;130 (Babcock and Fowler, 1994; Sung et al.,1992). SM-Lmod displays the most divergence from thecanonical 40-kDa Tmod sequences and is missing theentire nonmuscle TM-binding region. Preliminary evi-dence indicates that SM-Lmod does bind skeletal mus-cle TM, although apparently to a lesser extent thanE-Tmod, but SM-Lmod does not bind brain TM (Conleyand Fowler, 1996; Conley, in press). The actin-cappingactivity of E-Tmod has been shown to require a regionin its carboxy-terminal half (Gregorio et al., 1995;Fowler et al., 2000), which is conserved with a centralportion of the Lmods (Fig. 4). We have shown by dif-ferential centrifugation that SM-Lmod cofractionateswith the actin-containing Triton-insoluble cytoskeletonof smooth muscle (Conley and Fowler, 1996; Conley, inpress). Unfortunately, our attempts to assess the actin-capping activity of SM-Lmod were inconclusive due toproblems with protein aggregation (C. A. Conley, R.Kuhl, A. Weber, and V. M. Fowler, unpublished data).
Several interesting correlations can be made be-tween the genomic structure of the Tmod gene familyand the biochemical functions that have been reported.The boundary between exons 2 and 3 of the Tmods andthat between exons 1 and 2 of C-Lmod fall quite nearthe boundary between the skeletal muscle TM anderythrocyte (nonmuscle) TM binding domains ofE-Tmod (shading in Fig. 3 and triangles in Fig. 4), asidentified by Babcock and Fowler (1994) and Sung etal. (1992). The amino-terminal deletion in SM-Lmodmay have been a result of losing the ancestral firstexon, as the SM-Lmod amino-terminus is locatedslightly upstream of the splice junction between thefirst and the second exons of the Tmods (shading in Fig.3 and triangles in Fig. 4). It is tempting to speculatethat the skeletal muscle TM-binding domain of theTmods and Lmods has been added in a “cassette” fash-ion, although it is not clear whether the intron is an-cestral or derived. The third and fourth exons of theTmods cover a region that is highly divergent amongTmod and Lmod proteins (bar in Fig. 2, shading in Fig.3, and triangles in Fig. 4), suggesting that functionalconstraints on this region are low. The conserved cen-
FIG. 7. Histograms showing quantitation of human Tmods andLmods, from the blots in Fig. 6. Quantitative expression levels aremeasured relative to ubiquitin for all blots. U-Tmod is expressed inall tissues while Sk-Tmod is expressed exclusively in skeletal mus-cle. Both E-Tmod and C-Lmod are most highly expressed in fetal andadult heart and skeletal muscle, but only E-Tmod is expressed atlower levels in other tissues as well. SM-Lmod is most highly ex-pressed in aorta and bladder and is present at less than half themaximum level in other tissues. The quantitation is in arbitraryunits, and the labels on the ordinate correspond to the diagram inFig. 6.

1gll
C
C
C
C
C
C
F
138 CONLEY ET AL.
tral/carboxy-terminal section in the multiple align-ment of Lmod and Tmod proteins is carried on thesecond exon of the Lmods but requires exons 5 through8 of the Tmods (shading in Fig. 3 and triangles in Fig.4). Here, it is tempting to speculate that this putativeactin-capping region was originally a single-exon cas-sette, which was modified differently in the Lmods andTmods subsequent to the ancient gene duplicationevent that created them. Determination of whether theintrons found in Tmod family genes are ancestral orderived will require the examination of additionalgenomic sequences from a variety of divergent taxa.
Chromosomal Location of Lmods and Implications forHuman Disease
Recently, it was demonstrated that the four humanTmod genes are located on paralogous sections of chro-mosomes 1, 9, and 15 (Cox and Zoghbi, 2000), suggest-ing that they may have been duplicated as part of alarger block duplication event (Smith et al., 1999). Incontrast, the Lmods do not appear to be located onparalogous chromosome regions (data obtained fromGeneMap ’99). SM-Lmod maps to human chromosome1q32, between markers D1S2622 and D1S306 and neargenes encoding myogenin, a nonreceptor tyrosine phos-phatase, and an unknown cysteine- and glycine-richprotein. C-Lmod maps to human chromosome 7q31,between markers D7S655 and D7S686 and near genesencoding a receptor protein tyrosine phosphatase,sperm adhesion molecule 1, and an unknown proteinwith similarity to reverse transcriptase. These dataindicate that the Lmods were duplicated as part of asmall-scale duplication event, rather than the large-scale event that created the Tmods.
Interestingly, both SM-Lmod and C-Lmod map closeto recognized disease loci (Table 1): SM-Lmod mapsnear the glomerulopathy locus GFND (Vollmer et al.,998), the PPS locus for a syndrome of popliteal ptery-ium that manifests as harelip and webbing in theower extremities (Lees et al., 1999), and the CMD1Docus for dilated cardiomyopathy type 2 (Durand et al.,
1995). However, it is not clear how SM-Lmod mightfunction in the progression of these diseases. C-Lmodmaps near the hypertrophic cardiomyopathy locusCMH6 (MacRae et al., 1995), the human speech disor-der locus SPCH1 at 7q31 (Fisher et al., 1998), and therecessive deafness loci DFNB 14 and 17, which may besynonymous (Mustapha et al., 1998). Because C-Lmodis expressed at detectable levels only in heart andskeletal muscle (Figs. 6 and 7), it seems unlikely thatthis protein would be involved in either a speech dis-order or hereditary deafness. However, it is very at-tractive to hypothesize that defects in C-Lmod mightbe involved in the pathogenesis of hypertrophic cardio-myopathy, given the known functions of other Tmodisoforms. Evaluating CMH6 patients for defects in theC-Lmod gene or protein is a promising area for futureresearch.
ACKNOWLEDGMENTS
The authors thank Dr. Marian Ludgate for generous sharing ofreagents, Rebecca Kuhl for assistance with DNA sequencing, and Dr.David Hoffman and Ingrid Udranszky for the rapid cloning of humanC-Lmod exon 2. This work was supported by grants to V.M.F. fromthe National Institutes of Health (GM-34225) and from the HumanFrontiers in Science Program. C.A.C. was the recipient of a NationalInstitutes of Health (National Research Service Award) postdoctoralfellowship and is currently supported by the NASA FundamentalBiology program.
Note added in proof. After this paper was accepted, a portion ofmouse genomic sequence, Accession No. AC084272, was released inGenbank that encodes some exons of the mouse SK-Tmod gene. Theavailable exons show a similar structure to the other Tmod genes.
REFERENCES
Almenar-Queralt, A., Lee, A., Conley, C. A., and Fowler, V. M.(1999). Identification of a novel tropomodulin isoform, skeletaltropomodulin, that caps actin filament pointed ends in skeletalmuscle. J. Biol. Chem. 274: 28466–28475.
Babcock, G. G., and Fowler, V. M. (1994). Isoform specific interactionof tropomodulin with skeletal muscle and erythrocyte tropomyo-sins. J. Biol. Chem. 269: 27510–27518.
hu, X., Thompson, D., Yee, L. J., and Sung, L. A. (2000). Genomicorganization of mouse and human erythrocyte tropomodulin genesencoding the pointed end capping protein for the actin filaments.Gene 256: 271–281.onley, C. A. (2001). Leiomodin and Tropomodulin in smooth muscle.Genomics, in press.onley, C. A., and Fowler, V. M. (1996). A new family of actinfilament pointed end capping proteins. Mol. Biol. Cell 7(Suppl.):379A.onley, C. A., and Fowler, V. M. (1999). Localization of the human64kD autoantigen D1 to myofibrils in a subset of extraocularmuscle fibers. Curr. Eye Res. 19: 313–322.onley, C. A., Almenar-Queralt, A., and Fowler, V. M. (1998). Iden-tifying novel tropomodulin isoforms. Mol. Biol. Cell 9(Suppl.): 18A.ox, P. R., and Zoghbi, H. Y. (2000). Sequencing, expression analysis,and mapping of three unique human tropomodulin genes and theirmouse orthologs. Genomics 63: 97–107.
Dong, Q., Ludgate, M., and Vassart, G. (1991). Cloning and sequenc-ing of a novel 64-kDa autoantigen recognized by patients withautoimmune thyroid disease. J. Clin. Endocrinol. Metabol. 72:1375–1381.
Durand, J.-B., Bachinski, L. L., Bieling, L. C., Czernuszewicz, G. Z.,Abchee, A. B., Yu, Q. T., Tapscott, T., Hill, R., Ifegwu, J., Marian,A. J., Brugada, R., Daiger, S., Gregoritch, J. M., Anderson, J. L.,Quinones, M., Towbin, J. A., and Roberts, R. (1995). Localization ofa gene responsible for familial dilated cardiomyopathy to chromo-some 1q32. Circulation 92: 3387–3389.
Fischer, R. S., Lee, A., and Fowler, V. M. (2000). Tropomodulin andtropomyosin mediate lens cell actin cytoskeleton reorganization invitro. Invest. Ophthalmol. Vis. Sci. 41: 166–174.
Fisher, S. E., Vargha-Khadem, F., Watkins, K. E., Monaco, A. P., andPembrey, M. E. (1998). Localisation of a gene implicated in asevere speech and language disorder. Nat. Genet. 18: 168–170.
Fowler, V. M. (1987). Identification and purification of a novel mr43,000 tropomyosin-binding protein from human erythrocytemembranes. J. Biol. Chem. 22: 12792–12800.
Fowler, V. M. (1990). Tropomodulin: A cytoskeletal protein thatbinds to the end of erythrocyte tropomyosin and inhibits tropomy-osin binding to actin. J. Cell Biol. 111: 471–482.
owler, V. M. (1997). Capping actin filament growth: Tropomodulinin muscle and nonmuscle cells. In “50th Annual Meeting andSymposium of the Society of General Physiologists: Cytoskeletal

F
G
G
I
K
L
M
M
S
S
S
S
V
W
W
W
W
Z
139LEIOMODINS: MEMBERS OF THE Tmod GENE FAMILY
Regulation of Membrane Function,” Vol. 52, pp. 79–89, Rock-efeller Univ. Press, New York.
Fowler, V. M., and Conley, C. A. (1999). Tropomodulin. In “Guide-book to the Cytoskeletal and Motor Proteins,” 2nd ed., pp. 154–158, Oxford Univ. Press, London.
owler, V. M., Moyer, J., Almenar-Queralt, A. A., Fritz-Six, K., andShukla, U. (2000). Identification of a novel conserved actin cappingdomain in the pointed end capping protein, tropomodulin. Mol.Biol. Cell 11(Suppl.): 557A.regorio, C. C., and Fowler, V. M. (1995). Mechanisms of thin fila-ment assembly in embryonic chick cardiac myocytes: Tropomodu-lin requires tropomyosin for assembly. J. Cell Biol. 129: 683–695.regorio, C. C., Weber, A., Bondad, M., Pennise, C. R., and Fowler,V. M. (1995). Requirement of pointed-end capping by tropomodulinto maintain actin filament length in embryonic chick cardiac myo-cytes. Nature 376: 83–86.
to, M., Swanson, B., Sussman, M., Kedes, L., and Lyons, G. (1995).Cloning of tropomodulin cDNA and localization of gene transcriptsduring mouse embryogenesis. Dev. Biol. 167: 317–328.romminga, A., Hagel, C., Arndt, R., and Schuppert, F. (1998).Serological reactivity of recombinant 1D autoantigen and its ex-pression in human thyroid and eye muscle tissue: A possible au-toantigenic link in Graves’ patients. J. Clin. Endocrinol. Metab.83: 2817–2823.
ees, M. M., Winter, R. M., Malcolm, S., Saal, H. M., and Chitty, L.(1999). Popliteal pterygium syndrome: A clinical study of threefamilies and report of linkage to the Van der Woude syndromelocus on 1q32. J. Med. Genet. 36: 888–892.acRae, C. A., Ghaisas, N., Kass, S., Donnelly, S., Basson, C. T.,Watkins, H. C., Anan, R., Thierfelder, L. H., McGarry, K., Row-land, E., et al. (1995). Familial hypertrophic cardiomyopathy withWolff–Parkinson–White syndrome maps to a locus on chromosome7q3. J. Clin. Invest. 96: 1216–1220.ustapha, M., Salem, N., Weil, D., El-Zir, E., Loiselet, J., and Petit,C. (1998). Identification of a locus on chromosome 7q31, DFNB14,responsible for prelingual sensorineural nonsyndromic deafness.Eur. J. Hum. Genet. 6: 548–551.
mith, H. G. C., Knight, R., and Hurst, L. D. (1999). Vertebrategenome evolution: A slow shuffle or a big bang? BioEssays 21:697–703.
ung, L. A., Fowler, V. M., Lambert, K., Sussman, M. A., Karr, D.,and Chien, S. (1992). Molecular cloning and characterization ofhuman fetal liver tropomodulin, a tropomyosin binding protein.J. Biol. Chem. 267: 2616–2621.
ung, L. A., Fan, Y., and Lin, C. C. (1996). Gene assignment, expres-sion, and homology of human tropomodulin. Genomics 34: 92–96.
ussman, M. A., Welch, S., Cambon, N., Klevitsky, R., Hewett, T. E.,Price, R., Witt, S. A., and Kimball, T. R. (1998). J. Clin. Invest. 101:51–61.ollmer, M., Jung, M., Ruschendorf, F., Ruf, R., Wienker, T., Reis, A.,Krapf, R., and Hildebrandt, F. (1998). The gene for human fi-bronectin glomerulopathy maps to 1q32, in the region of the reg-ulation of complement activation gene cluster. Am. J. Hum. Genet.63: 1724–1731.all, J. R., Bernard, N., Boucher, A., Salvi, M., Zhang, Z. G., Ken-nerdell, J., Tyutyunikov, A., and Genovese, C. (1993). Pathogene-sis of thyroid-associated ophthalmopathy: An autoimmune disor-der of the eye muscle associated with Graves’ hyperthyroidism andHashimoto’s thyroiditis. Clin. Immunol. Immunopathol. 68: 1–8.atakabe, A., Kobayashi, R., and Helfman, D. M. (1996). N-tropo-modulin: A novel isoform of tropomodulin identified as the majorbinding protein to brain tropomyosin. J. Cell Sci. 109: 2299–2310.eber, A., Pennise, C. R., Babcock, G. G., and Fowler, V. M. (1994).Tropomodulin caps the pointed ends of actin filaments. J. Cell Biol.127: 1627–1635.eber, A., Pennise, C. A., and Fowler, V. M. (1999). Tropomodulinincreases the critical concentration of barbed end-capped actinfilaments by converting ADP. Pi-actin to ADP-actin at all pointedfilament ends. J. Biol. Chem. 274: 34637–34645.
hang, Z.-G., Wall, J. R., and Bernard, N. F. (1996). Tissue distribu-tion and quantitation of a gene expressing a 64-kDa antigen asso-ciated with thyroid-associated ophthalmopathy. Clin. Immunol.Immunopathol. 80: 236–244.