isolation and characterization of structural components of bacillus cereus ahu 1356 cell walls
TRANSCRIPT

Eur. J . Biochem. 75, 513-522 (1977)
Isolation and Characterization of Structural Components of Bacillus cereus AHU 1356 Cell Walls
Ken-ichi AMANO, Setsuro HAZAMA, Yoshio ARAKI, and Eiji IT0
Department of Chemistry, Faculty of Science, Hokkaido University, Sapporo
(Received February 1, 1977)
From lysozyme digests of the N-acetylated cell walls of BacilZus cereus AHU 1356, a polysaccharide fraction and a teichoic acid fraction were isolated by ion-exchange chromatography and gel chromatography. The former fraction, accounting for 50 % of the walls, contained N-acetylglucos- amine, N-acetylmannosamine, N-acetylgalactosamine and glucose in a molar ratio of 4: 1 : 1 : 1 together with a small amount of the peptidoglycan constituents. The latter fraction accounted for 5 % of the walls and was composed of N-acetylglucosamine, galactose, glycerol and phosphorus in a molar ratio of 1 : 1.4: 1 : 1 and a small amount of the peptidoglycan constituents. The molecular weight of the polysaccharide fraction was about 33 000 as estimated by gel chromatography. After acid hydrolysis of the polysaccharide fraction, a stoichiometric amount of muramic acid 6-phosphate was detected which was probably involved in the linkage between the polysaccharide and peptidoglycan moieties. The polysaccharide moiety detached from the peptidoglycan components by mild acid hydrolysis was isolated as material with an approximate molecular weight of 28000.
Structural studies on the polysaccharide, involving Smith degradation, Cr03 oxidation and proton magnetic resonance spectrometry of the polysaccharide fraction together with the analysis of oligosaccharides obtained from partial acid hydrolysis of the polysaccharide fraction, led to the tentative formulation of the most likely structure of the repeating unit as follows :
GlcNAc 1
1{6
j: PI6
Glc GlcNAc GlcNAc 1
+3)-GalNAc-cc(l+4)-ManNAc-~(1+3)-GlcNAc-r(1+.
The cell walls of the gram-positive bacteria are known to consist of the basal structure, peptido- glycan, and some of the specific components such as proteins, polysaccharides, teichoic acids and teich- uronic acids. However, our knowledge of the structure of these cell wall polysaccharides is still limited.
Hughes reported that the cell walls of Bacillus cereus N.R.R.L. 596 have three components, peptido- glycan, an acidic polysaccharide and a neutral poly-
Abbreviations. GlcNAc, N-acetylglucosamine; ManNAc, N- acet ylmannosamine; GalNAc, N-acetylgalactosamine.
Enzymes. Lysozyme (EC 3.2.1.17); alkaline phosphatase (EC 3.1.3.1); glycerol kinase (EC 2.7.1.30); glycerol-3-phosphate de- hydrogenase (EC 1.1.1.8); P-glucosidase (EC 3.2.1.21); cc-N-acetyl- galactosaminidase (EC 3.2.1.49); a-glucosidase (EC 3.2.1.20); exo-P-N-acetylhexosaminidase (EC 3.2.1.52).
saccharide composed of N-acetylglucosamine, N- acetylgalactosamine and glucose [I]. The structures of these polysaccharides were not determined. On the other hand, Araki et al. found that in the cell walls from B. cereus AHU 1356 the glucosamine residues in the glycan chain of peptidoglycan are almost entirely N-unsubstituted [2]. From this observation, it was inferred that these cell walls may have some anionic components which can make up for the posi- tive charges on the N-unsubstituted glucosamine residues. In fact, further studies showed the cell walls of this strain to contain mannosamine, galactose, glycerol and phosphorus in addition to the already recognized galactosamine and glucose.
Mannosamine has been found in a few bacterial polysaccharides such as capsular polysaccharides of Diplococcus pneumoniae types IV [3,4] and IX [5,6].

514 Structural Components of Bacillus cereus Cell Walls
The structure of the polysaccharides of D. pneu- moniae type IX was reported. In addition, meningo- coccal group A polysaccharide [7], as well as Bacillus pumilus polysaccharide [8] cross-reactive with the meningococcal group A polysaccharide, was charac- terized as a polymer of N-acetylmannosamine phos- phate linked by 1,6-phospho-diester bond. Recent studies in our laboratory using a cell-free preparation from B. cereus AHU 1356 indicated that N-acetyl- mannosamine was introduced from UDP-ManNAc into a polysaccharide comprising glucosamine, man- nosamine, galactosamine and glucose through inter- mediate formation of lipid-linked oligosaccharides (Yamamori, Murazumi, Araki and Ito, unpublished). These facts prompted us to elucidate the structure of the specific components and the nature of their bind- ing with peptidoglycan in the cell walls of this strain.
The present paper reports the separation and characterization of polysaccharide-linked glycopep- tide from lysozyme digests of the N-acetylated cell walls. The isolation of teichoic-acid-linked glyco- peptide is also described.
MATERIALS AND METHODS
Materials
The cell walls of B. cereus AHU 1356 were prepared as described previously but without trypsin digestion. Hen's egg-white lysozyme (twice crystal- lized) and Escherichia coli alkaline phosphatase were purchased from Sigma Chemicals; Sephadex G-types, dextran T-types, blue dextran 2000 and Sepharose 6B from Pharmacia Fine Chemicals; DEAE-cellulose (Serva) from Seikagaku Kogyo ; Glucostat reagent from Worthington Biochemicals ; glycerol kinase and glycerol-3-phosphate dehydrogenase from Boehringer Mannheim. N-Acetylchito-oligoses were prepared from chitin according to the method of Rupley [9].
Lysozyme Digestion of N-Acetylated Cell Walls
N-Acetylation of cell walls from Bacillus cereus AHU 1356 was carried out by shaking 2.2g of the sample in a solution containing 10 ml acetic anhydride, 16.8 g NaHC03 and 200 ml distilled water overnight at 0 "C as described previously [2]. Then the insoluble material was collected by centrifugation, washed twice with distilled water, and lyophilized (2.4 g). The resulting N-acetylated cell walls (1.8 g) was digested at 37 "C for 48 h in a mixture containing 440 mg hen's egg-white lysozyme, 1 % sodium azide and 50 mM Tris-HC1 pH 7.2 in a final volume of 1.5 1. The lysozyme digest was concentrated to 50 ml and dialyzed against 4 1 distilled water for 1 day. The dialysis was repeated three times, and the nondialyzable fraction was lyophilized (1.3 g).
Isolation of Polysaccharide-Linked and Teichoic- Acid- Linked Glycopep tides
The nondialyzable fraction of the lysozyme digest was dissolved in 10 mlO.05 M NH4HC03 and applied to a Sephadex G-50 column (5.0 x 95 cm). The column was eluted with 0.05 M NH4HC03 at a flow rate of 80 ml/h, and the elution pattern was monitored by determining reducing groups and hexoses (phenol/ sulfuric acid method). The void volume as determined by blue dextran was 700 ml. The fractions (elution volume 640 - 760 ml) that contained polymers ex- cluded from the column, were pooled, dialyzed against distilled water and lyophilized.
The polymer fraction was separated by chromatog- raphy on a DEAE-cellulose column (2.7 x 32 cm) equilibrated with 0.02 M NaCl in 5 mM Tris-HC1 buffer pH 7.2. The column was eluted at a flow rate of 40 ml/h with 400 ml of the same NaCl solution and then with a linear gradient from 0.02 to 0.3 M NaCl in the same buffer (1200 ml). The fractions of 13.7 ml were collected and the elution pattern was monitored by determining reducing groups, hexoses and total phosphorus. A neutral polysaccharide was eluted at 0.06-0.10 M NaCl and an acidic polymer was eluted at 0.16-0.22 M NaC1. The neutral poly- saccharide and the acidic polymer fractions were separately pooled, dialyzed against distilled water and lyophilized.
The neutral polysaccharide fraction (757 mg) dis- solved in 4 ml 0.05 M NH4HC03 was purified by chromatography on a Sephadex G-200 column (2.7 x 96 cm) in 0.05 M NH4HC03. The flow rate was 8 ml/h and the void volume was 124 ml. The fractions containing polysaccharide-linked glycopeptide as indicated by the hexose content (elution volume 166 - 248 ml) were pooled, dialyzed against distilled water and lyophilized.
The acidic polymer fraction (170 mg) obtained from chromatography on the DEAE-cellulose col- umn was dissolved in 2 ml 0.05 M NH4HC03 and subjected to chromatography on a Sepharose 6B column (1.5 x 100 cm) in 0.05 M NH4HC03 at a flow rate of 4 ml/h. The void volume was 64ml. The fractions containing hexoses and phosphorus (elution volume 69- 120 ml) were pooled, dialyzed against distilled water and lyophilized.
Mild A c id Hydrolysis of Polysaccharide-Linked Glycopeptide and Isolation of Polysaccharide
The polysaccharide-linked glycopeptide (5.3 mg) was heated in 1 ml 0.01 M HCl at 100 "C for 30 min and lyophilized. The hydrolysate dissolved in 0.5 ml of 0.01 M NaCl in 5 mM Tris-HC1 buffer pH 7.2 was applied to a DEAE-cellulose column (1.5 x 8 cm)

K . Amano, S. Hazdma, Y. Araki, and E. Ito 515
equilibrated with 0.01 M NaCl in the same buffer. The column was eluted at a flow rate of 8 ml/h succes- sively with 45 ml of the same NaCl solution, a linear gradient from 0.02 to 0.2 M NaCl in 100 ml of the same buffer and finally with 25 ml of 0.5 M NaCl in the same buffer. Hexose-containing material was eluted at 0.01 M NaCl (elution volume 9-21 ml, fraction A), whereas glycopeptide with reducing power was at 0.5 M NaCl (elution volume 153 - 177 ml, fraction B). The hexose-containing neutral polysaccharide frac- tion (A) was concentrated to 0.5 ml, applied to a column of Sephadex G-200 (1 .O x 70 cm) in 0.05 M NH4HC03, and eluted with 0.05 M NH4HC03 (flow rate 3 ml/h, void volume 23 ml). The fractions con- taining polysaccharide (elution volume 32- 43 ml, fraction A-1) and the fractions containing oligo- saccharides (54- 60 ml, fraction A-2) were separately pooled and lyophilized. Desaltation of the glyco- peptide fraction (B) was carried out by gel filtration on a column of Sephadex G-25 (1 .O x 65 cm).
Smith Degradation of Polysaccharide-Linked Glycopeptide
A portion of the polysaccharide-linked glyco- peptide (0.7 mg) was oxidized with 0.03 M NaI04 in 1 ml 0.01 M ammonium acetate buffer pH 4.5 for 48 h in the dark at room temperature according to the procedure described by Goldstein et al. [lo]. The oxidized sample was dialyzed against distilled water and nondialyzable material was lyophilized (sample 1). A portion (two-thirds) of sample 1 was dissolved in 0.5 ml 0.1 M NaBH4, kept for 15 h at room temperature, lyophilized and hydrolyzed in 1 ml 0.2 M HCl for 15 h at room temperature. The hydrolysate was neutralized, concentrated and then applied to a column of Sephadex G-50 (1.0 x 71 cm). The column was eluted with 0.05 M NH4HC03 and the fraction eluted at the void volume (elution volume 23 - 27 ml) was lyophilized after dialysis against distilled water (sample 2). A portion (a half) of sample 2 was oxidized again with NaI04 as described above, giving sample 3.
Partial Acid Hydrolysis of' Polysaccharide- Linked Glycopeptide and Fractionation of Oligosaccharides
The polysaccharide-linked glycopeptide (67 mg) was hydrolyzed in 3 ml 0.5 M HCl at 100 "C for 1 h. After lyophilization, the hydrolysate was N-acetylated in a mixture containing 0.5 ml acetic anhydride and 0.85g NaHC03 in a final volume of 10ml for 24 h at 4 "C. The mixture was then applied to a Dowex 50 W-X8 (H' form) column (1.0 x 13 cm) and the column was washed with 30ml distilled
water. The effluent, concentrated to I ml, was further added to a column of Sephadex G-15 (1.5 x 100 cm) in 0.05 M NH4HC03. The column was eluted with 0.05 M NH4HC03 at a flow rate of 6 ml/h and fractions of 0.9 ml were collected. The elution pattern was monitored by determining reducing groups.
Analytical Methods
Amino acids, amino sugars and muramic acid 6-phosphate were analyzed in a Shibata AA-100 autoanalyzer [2] with samples (50 nmol) hydrolyzed in 4 M HCI for 4 h at 100 "C in sealed tubes. The analyses were also carried out with the hydrolysates of samples (50nmol) which had been reduced with NaBH4 as described by Tipper et a/ . [l 11 or oxidized with 0.01 M NaI04 in 0.1 ml of 10 mM ammonium acetate buffer pH 4.5 for 48 h in the dark at room temperature as described by Goldstein et al. [lo]. D-Glucosamine, D-mannosamine and D-galactosamine were separated from one another by the analyzer as described previously [2], but the column was first eluted for 150 min at 59 "C, instead of being eluted for 100 min at 40 "C, with 0.2 M citrate buffer pH 3.2. Amino alditols resulting from reduction of amino sugars were differentiated from the original amino sugars by the autoanalyzer under the same elution conditions.
Reducing groups were determined by the method of Park and Johnson [I21 with N-acetylglucosamine as a reference standard ; hexoses by the phenol/HzSO4 method [13] with glucose as a reference standard; total phosphorus by the method of Lowry and Roberts [14]; D-glucose by the D-glucose oxidase method [15]. To determine total glycerol, acid hydrolysates of samples were treated with alkaline phosphatase. Glycerol was separated by quantitative paper chro- matography in solvent 1 and determined as formalin after periodate oxidation [16]. Glycerol was also determined enzymatically by the method of Wieland
Molecular weights of polysaccharide-linked glyco- peptide and its polysaccharide moiety were determined by gel chromatography on a column (1 .O x 70 cm) of Sephadex G-200 in 0.05 M NH4HC03 at a flow rate of 2.5 ml/h. Elution was monitored by assaying hexoses with phenol/HzSO4. Dextrans T-20, T-40 and T-70 were used as standards.
Anomeric configuration of glycosidic linkages in polysaccharide and oligosaccharides was determined by the chromic anhydride oxidation method [18,19]. Proton magnetic resonance spectra were measured in a JNM PS-100 (Jeol) instrument (100 MHz) in 2 H 2 0 at 90 "C [20] using samples (40 mg) dissolved in 0.5 ml of 'Hz0 and tetramethylsilane as an external standard.
~ 7 1 .
516 Structural Components of Bacillus cereus Cell Walls
Paper Chromatography
Paper chromatography was carried out by the descending method on Toyo no. 50 filter paper in the following solvents : (1) butan-1-ol/pyridine/water (6/4/3, v/v/v); (2) butan-1-ol/acetic acid/water (4/1/5, v/v/v, upper phase); ( 3 ) isobutyric acid/0.5 M NH40H (5/3, v/v). For chromatography in solvent 1, paper was previously dipped into 0.015 M or 0.05 M NazB4- 0 7 adjusted to pH 10 and dried [21]. N-Acetylamino sugars were detected by the method of Sharon and Seifter [22], and reducing sugars with the alkaline silver nitrate reagent [23]. Rates of migration on paper chromatography are given relative to N-acetylglucos- amine ( R G I ~ N A ~ ) .
Enzymatic Hydrolysis of Oligosaccharides
The enzymes used were: Charonia lampas fi-glucosi- dase (5 units/ml) and C. lampas a-N-acetylgalactos- aminidase (0.1 unit/0.2 ml) purchased from Seikagaku Kogyo; yeast a-glucosidase (type I) (38 units/ml) purchased from Sigma Chemicals ; pig epididymis exo-P-N-acetylhexosaminidase (60 units/ml) prepared according to the reported procedure [2].
Oligosaccharides (50 nmol) were incubated at 37 "C overnight with 1 1-11 of exo-fi-N-acetylhexos- aminidase, a-N-acetylgalactosaminidase or fi-glucosi- dase in 100 pl 50 mM sodium citrate buffer pH 4.4. Incubation was also carried out with 1 pl of a- glucosidase in 100 pl 50 mM potassium phosphate buffer pH 6.5. After incubation the reaction mixture was applied to a combination column of Dowex 3 -X2 and Dowex 50W-X8 and the column was eluted with three bed volumes of distilled water. The eluate was lyophilized and subjected to paper chromatography in solvent 1.
RESULTS
Composition of Cell Walls
As reported previously [2], the cell walls of B. cereus AHU 1356 resist acid hydrolysis and lyso- zyme digestion because of the occurrence of N- unsubstituted glucosamine residues in the glycan chain of the peptidoglycan component. Therefore, a cell wall preparation N-acetylated with acetic an- hydride was used for analysis and structural studies. The analyses (Table 1) of the N-acetylated cell walls accounted for 88% of the weight of the preparation. In addition to the peptidoglycan constituents, galactos- amine, mannosamine and glucose were found in an approximate molar ratio of 1 .O : 1 .O : 1 .O. Glucosamine was present in large molar excess over the other amino sugars and peptidoglycan constituents. Moreover, qualitative analysis by paper chromatography indicat- ed the presence of considerable amounts of galactose
Table 1. Composition of N-acetylaied cell wulls, polysucchuride- linked glycopeptide and teichoic-acid-linked glycopepiide fioni B. cereus Analytical methods were those described in Materials and Methods. Values refer to dry weight of material. Molar ratios were calculated taking muramic acid 6-phosphate as 1 .OO. The value of galactose content was calculated by subtracting the value for glucose from the hexose content
Component Amount present in __ ~~~ _ _ _ ~~~ ~~~
N-acetylated poly- teichoic- cell walls saccharide- acid-linked
linked glyco- glycopeptide peptide
nmol/mg (mol/mol) .~ ~ _ _ ~ ~~~~ _-
Muramic acid 6-phosphate 15 (1.0) 23 (1.0) 14 (1.0)
Muramic acid 358 (23.9) 75 (3.2) 56 (4.0) Glutamic acid 358 (23.9) 87 (3.7) 64 (4.6) Alanine 630 (42.0) 157 (6.7) 116 (8.3) Diaminopimelic
acid 363 (24.2) 106 (4.5) 80 (5.7) Glucosamine 1880 (125) 2730 (119) 1440 (103) Mannosamine 364 (24.3) 622 (27.0) 0 (0) Galactosamine 351 (23.4) 629 (27.3) 0 (0) Hexose 517 (34.5) 610 (26.5) 1970 (141) Glucose 368 (24.5) 610 (26.5) 0 (0) Galactose 149 (9.9) 0 (0) 1970 (141)
0 (0) 1080 (77.1) Total phosphorus 100 (5.7) 28 (1.2) 1350 (96.4) Glycerol - (-1
and glycerol. Muramic acid 6-phosphate, as assayed with the autoanalyzer, accounted for 15% of the total phosphorus content. Although the procedure for preparation of the cell walls did not involve protease digestion, amino acids other than the peptidoglycan constituents represented in total less than 1 % of the weight. On mild acid treatment (0.1 M HCI, 60 "C, 24 h) of the cell walls, most of the hexosamine and hexose components, together with majority of glycerol and phosphorus, were released as soluble material [2, 241. These data suggest that the cell walls contained teichoic acid and polysaccharide linked to the peptido- glycan with acid-labile bonds.
Fractionation of Lysozyme Digest of Cell Walls
The cell walls after N-acetylation could easily be digested with lysozyme. The nondialyzable fraction of the lysozyme digest obtained from 1.8 g of the N-acetylated cell walls was passed through a column of Sephadex G-50. The fraction eluted at the void volume accounted for most of the hexoses and phosphorus contained in the starting material (yield : 923 mg dry weight, 139 pmol reducing groups, 776 ymol hexoses and 180 ymol total phosphorus). This polymer fraction was separated by DEAE- cellulose column chromatography into two fractions
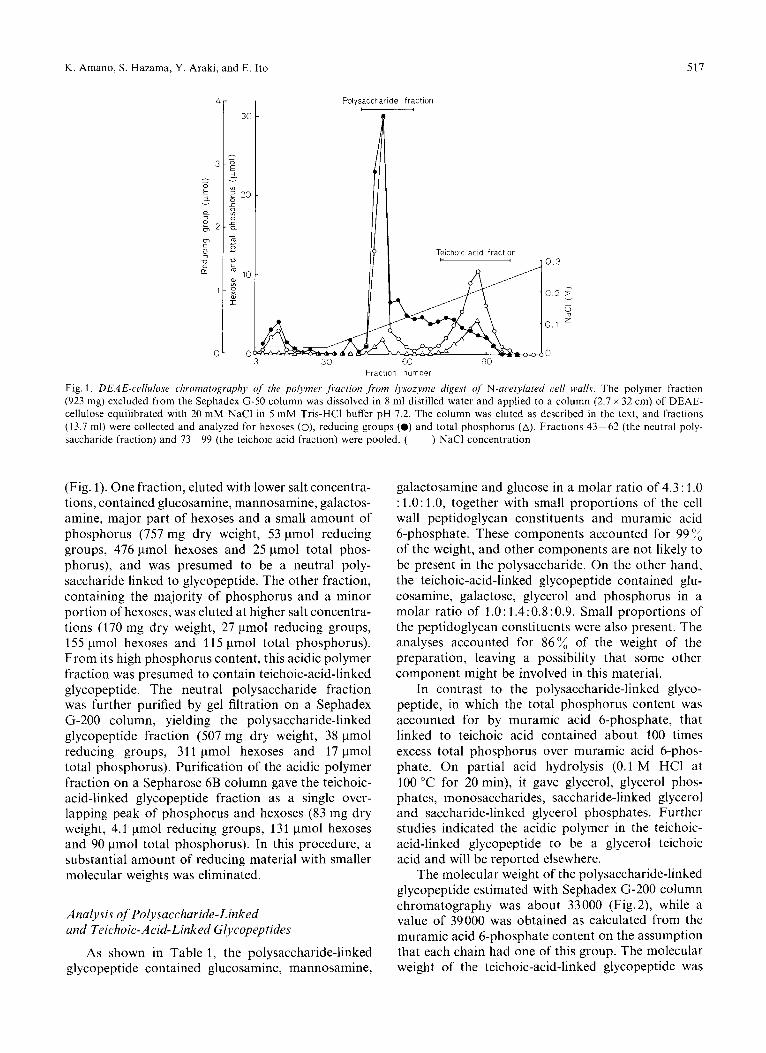
K . Amano, S. Hazama, Y . Araki, and E. Ito 517
Polvsaccharide fraction c
0 3 30
Teichoic acid fraction I 1 0 . 3
Fraction number
Fig. 1. DEAE-cc.llulosc. clironzutogrupliy of the polymer. ,/i.uciinn from Iysozynze digest OJ N-acrtylurrd c,rll walls. The polymer fraction (923 mg) excluded from the Sephadex G-50 column was dissolved in 8 ml distilled water and applied to a column (2.7 x 32 cm) of DEAE- cellulose equilibrated with 20 mM NaCl in 5 mM Tris-HCI buffer pH 7.2. The column was eluted as described in the text, and fractions (13.7 ml) were collected and analyzed for hexoses (O), reducing groups (0) and total phosphorus (A). Fractions 43-62 (the neutral poly- saccharide fraction) and 73-99 (the teichoic acid fraction) were pooled. ( ) NaCl concentration
(Fig. 1). One fraction, eluted with lower salt concentra- tions, contained glucosamine, mannosamine, galactos- amine, major part of hexoses and a small amount of phosphorus (757 mg dry weight, 53 pmol reducing groups, 476 pmol hexoses and 25 pmol total phos- phorus), and was presumed to be a neutral poly- saccharide linked to glycopeptide. The other fraction, containing the majority of phosphorus and a minor portion of hexoses, was eluted at higher salt concentra- tions (170 mg dry weight, 27 pmol reducing groups, 155 pmol hexoses and 11 5 pmol total phosphorus). From its high phosphorus content, this acidic polymer fraction was presumed to contain teichoic-acid-linked glycopeptide. The neutral polysaccharide fraction was further purified by gel filtration on a Sephadex G-200 column, yielding the polysaccharide-linked glycopeptide fraction (507 mg dry weight, 38 pmol reducing groups, 311 pmol hexoses and 17 pmol total phosphorus). Purification of the acidic polymer fraction on a Sepharose 6B column gave the teichoic- acid-linked glycopeptide fraction as a single over- lapping peak of phosphorus and hexoses (83 mg dry weight, 4.1 pmol reducing groups, 131 pmol hexoses and 90 pmol total phosphorus). In this procedure, a substantial amount of reducing material with smaller molecular weights was eliminated.
Analysis of Polysacchauide-Linked and Teichoic-Acid-Linked Glycopeptides
As shown in Table 1, the polysaccharide-linked glycopeptide contained glucosamine, mannosamine,
galactosamine and glucose in a molar ratio of 4.3 : 1 .O : 1.0: 1.0, together with small proportions of the cell wall peptidoglycan constituents and muramic acid 6-phosphate. These components accounted for 99 of the weight, and other components are not likely to be present in the polysaccharide. On the other hand, the teichoic-acid-linked glycopeptide contained glu- cosamine, galactose, glycerol and phosphorus in a molar ratio of 1 .O : 1.4: 0.8 : 0.9. Small proportions of the peptidoglycan constituents were also present. The analyses accounted for 86% of the weight of the preparation, leaving a possibility that some other component might be involved in this material.
In contrast to the polysaccharide-linked glyco- peptide, in which the total phosphorus content was accounted for by muramic acid 6-phosphate, that linked to teichoic acid contained about 100 times excess total phosphorus over muramic acid 6-phos- phate. On partial acid hydrolysis (0.1 M HCI at 100 "C for 20 min), it gave glycerol, glycerol phos- phates, monosaccharides, saccharide-linked glycerol and saccharide-linked glycerol phosphates. Further studies indicated the acidic polymer in the teichoic- acid-linked glycopeptide to be a glycerol teichoic acid and will be reported elsewhere.
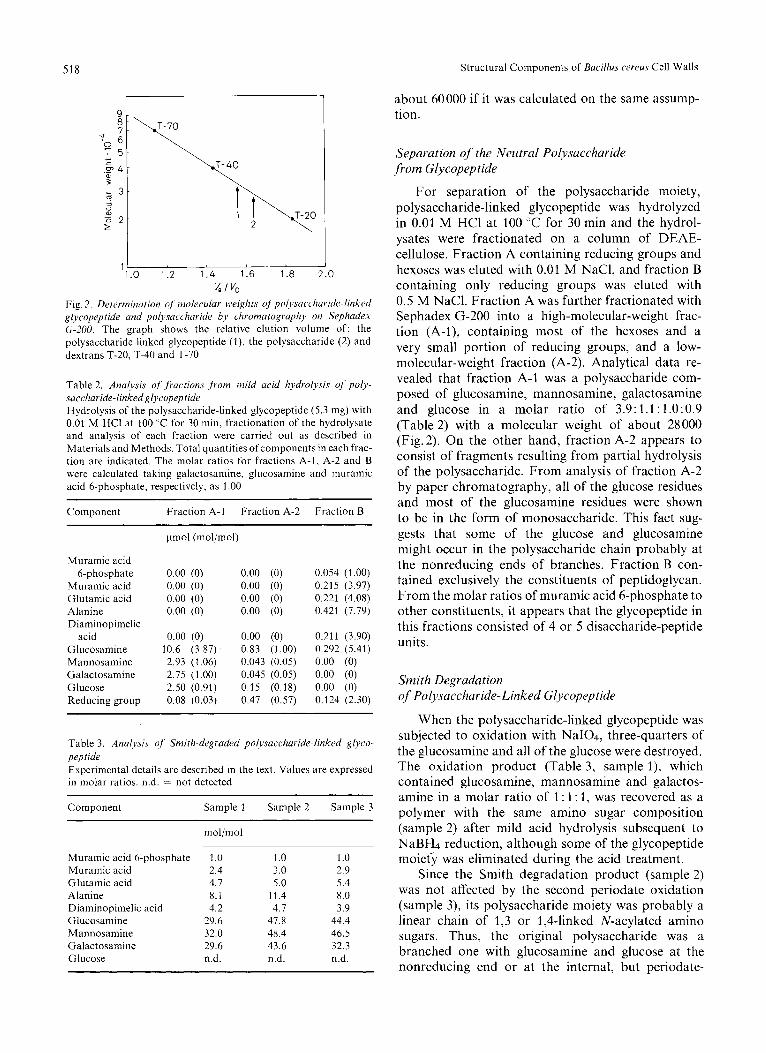
The molecular weight of the polysaccharide-linked glycopeptide estimated with Sephadex G-200 column chromatography was about 33000 (Fig. 2), while a value of 39000 was obtained as calculated from the muramic acid 6-phosphate content on the assumption that each chain had one of this group. The molecular weight of the teichoic-acid-linked glycopeptide was
518 Structural Components of Bacillus cereus Cell Walls
about 60000 if it was calculated on the same assump- tion.
1 1.0 1.2 1.4 1.6 1.8 2.0
v, I vo Fig. 2. Determination of moliwdar wvi&.s of'polysriccllaride-linked glycopeptide and polysaccharide by cliromutogruphy on Sephadex G-200. The graph shows the relative elution volume of: the polysaccharide-linked glycopeptide (l), the polysaccharide (2) and dextrans T-20, T-40 and T-70
Table 2. Analysis offructions from mild acid hydrolysis of poly- succharide-linked Rl~copeptide Hydrolysis of the polysaccharide-linked glycopeptide (5.3 mg) with 0.01 M HCI at 100 "C for 30 min, fractionation of the hydrolysate and analysis of each fraction were carried out as described in Materials and Methods. Total quantities of components in each frac- tion are indicated. The molar ratios for fractions A-1, A-2 and B were calculated taking galactosamine, glucosamine and muramic acid 6-phosphate, respectively, as 1.00
~
Component Fraction A-1 Fraction A-2 Fraction B
Muramic acid 6-phosphate
Muramic acid Glutamic acid Alanine Diaminopimelic
acid Glucosamine Mannosamine Galactosamine Glucose Reducing group
0.00 (0) 0.00 (0) 0.00 (0) 0.00 (0) 0.00 (0) 0.00 (0) 0.00 (0) 0.00 (0)
0.00 (0) 0.00 (0) 10.6 (3.87) 0.83 (1.00) 2.93 (1.06) 0.043 (0.05) 2.75 (1.00) 0.045 (0.05) 2.50 (0.91) 0.15 (0.18) 0.08 (0.03) 0.47 (0.57)
0.054 (1.00) 0.215 (3.97) 0.221 (4.08) 0.421 (7.79)
0.211 (3.90) 0.292 (5.41) 0.00 (0) 0.00 (0) 0.00 (0) 0.124 (2.30)
Table 3. Analysis of' Smith-degraded pol~~.Faccharide-linked glyco- p e p tide Experimental details are described in the text. Values are expressed in molar ratios. n.d. = not detected
Component Sample 1 Sample 2 Sample 3
Muramic acid 6-phosphate Muramic acid Glutamic acid Alanine Diaminopimelic acid Glucosamine Mannosamine Galactosamine Glucose
mol/mol ~.
1 .o 2.4 4.7 8.1 4.2
29.6 32.0 29.6 n.d.
~-
I .o 3.0 5.0
11.4 4.7
47.8 48.4 43.6 n.d.
~
1.0 2.9 5.4 8.0 3.9
44.4 46.5 32.3 n.d.
Separation of the Neutral Polysaccharide ,from Glycopeptide
For separation of the polysaccharide moiety, polysaccharide-linked glycopeptide was hydrolyzed in 0.01 M HCl at 100 "C for 30 min and the hydrol- ysates were fractionated on a column of DEAE- cellulose. Fraction A containing reducing groups and hexoses was eluted with 0.01 M NaCl, and fraction B containing only reducing groups was eluted with 0.5 M NaCl. Fraction A was further fractionated with Sephadex G-200 into a high-molecular-weight frac- tion (A-I), containing most of the hexoses and a very small portion of reducing groups, and a low- molecular-weight fraction (A-2). Analytical data re- vealed that fraction A-1 was a polysaccharide com- posed of glucosamine, mannosamine, galactosamine and glucose in a molar ratio of 3.9: 1.1: 1.0:0.9 (Table 2) with a molecular weight of about 28000 (Fig.2). On the other hand, fraction A-2 appears to consist of fragments resulting from partial hydrolysis of the polysaccharide. From analysis of fraction A-2 by paper chromatography, all of the glucose residues and most of the glucosamine residues were shown to be in the form of monosaccharide. This fact sug- gests that some of the glucose and glucosamine might occur in the polysaccharide chain probably at the nonreducing ends of branches. Fraction B con- tained exclusively the constituents of peptidoglycan. From the molar ratios of muramic acid 6-phosphate to other constituents, it appears that the glycopeptide in this fractions consisted of 4 or 5 disaccharide-peptide units.
Smith Degradation of Polysaccharide-Linked Glycopeptide
When the polysaccharide-linked glycopeptide was subjected to oxidation with NaI04, three-quarters of the glucosamine and all of the glucose were destroyed. The oxidation product (Table 3, sample I), which contained glucosamine, mannosamine and galactos- amine in a molar ratio of 1 : 1 : 1, was recovered as a polymer with the same amino sugar composition (sample 2) after mild acid hydrolysis subsequent to NaBH4 reduction, although some of the glycopeptide moieG was eliminated during the acid treatment.
Since the Smith degradation product (sample 2) was not affected by the second periodate oxidation (sample 3), its polysaccharide moiety was probably a linear chain of 1,3 or 1,4-linked N-acylated amino sugars. Thus, the original polysaccharide was a branched one with glucosamine and glucose at the nonreducing end or at the internal, but periodate-
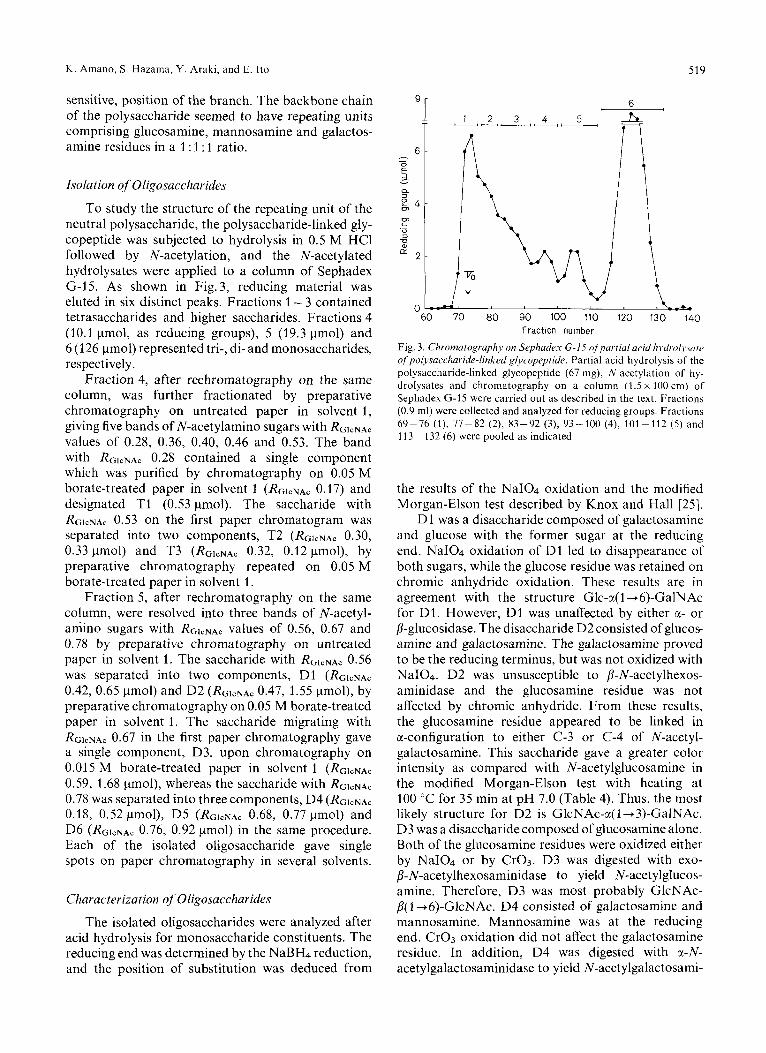
K . Amano, S. Hazama, Y. Araki, and E. Ito 519
sensitive, position of the branch. The backbone chain of the polysaccharide seemed to have repeating units comprising glucosamine, mannosamine and gdlactos- amine residues in a 1 : 1 : 1 ratio.
Isolation of Oligosuccharides
To study the structure of the repeating unit of the neutral polysaccharide, the polysaccharide-linked gly- copeptide was subjected to hydrolysis in 0.5 M HCl followed by N-acetylation, and the N-acetylated hydrolysates were applied to a column of Sephadex G-15. As shown in Fig.3, reducing material was eluted in six distinct peaks. Fractions 1 - 3 contained tetrasaccharides and higher saccharides. Fractions 4 (10.1 pmol, as reducing groups), 5 (19.3 pmol) and 6 (126 pmol) represented tri-, di- and monosaccharides, respectively.
Fraction 4, after rechromatography on the same column, was further fractionated by preparative chromatography on untreated paper in solvent 1, giving five bands of N-acetylamino sugars with R G I ~ N A ~ values of 0.28, 0.36, 0.40, 0.46 and 0.53. The band with R G ~ ~ N A ~ 0.28 contained a single component which was purified by chromatography on 0.05 M borate-treated paper in solvent 1 ( R G I ~ N A ~ 0.17) and designated T1 (0.53 pmol). The saccharide with R G ~ ~ N A ~ 0.53 on the first paper chromatogram was separated into two components, T2 ( R G I ~ N A ~ 0.30, 0.33 pmol) and T3 ( R G ~ ~ N A ~ 0.32, 0.12 pmol), by preparative chromatography repeated on 0.05 M borate-treated paper in solvent 1.
Fraction 5, after rechromatography on the same column, were resolved into three bands of N-acetyl- amino sugars with RclcNAc values of 0.56, 0.67 and 0.78 by preparative chromatography on untreated paper in solvent 1. The saccharide with R G ~ ~ N A ~ 0.56 was separated into two components, D1 ( R G I ~ N A ~ 0.42, 0.65 pmol) and D2 ( R G I ~ N A ~ 0.47, 1.55 pmol), by preparative chromatography on 0.05 M borate-treated paper in solvent 1. The saccharide migrating with R G ~ ~ N A ~ 0.67 in the first paper chromatography gave a single component, D3, upon chromatography on 0.015 M borate-treated paper in solvent 1 ( R G I ~ N A ~ 0.59, 1.68 pmol), whereas the saccharide with R G I ~ N A ~ 0.78 was separated into three components, D4 ( R G I ~ N A ~ 0.18, 0.52 pmol), D5 ( R G ~ ~ N A ~ 0.68, 0.77 pmol) and D6 ( R G ~ ~ N A ~ 0.76, 0.92 pmol) in the same procedure. Each of the isolated oligosaccharide gave single spots on paper chromatography in several solvents.
Characterization of Oligosaccharides
The isolated oligosaccharides were analyzed after acid hydrolysis for monosaccharide constituents. The reducing end was determined by the NaBH4 reduction, and the position of substitution was deduced from
0 60 -=J 70 80 ,w, 90 100 110 120 \.- 130 140
m .- 73
u 2
70
1 70
1 60 70 80 90 100 110 120 130 140
Fraction number Fig. 3. Chronzaiography on Sephade.y G-15 q/partiul acid liydrol~~.stirc ofpolysaccharide-linked glycopepride. Partial acid hydrolysis of the polysaccharide-linked glycopeptide (67 mg), N-acetylation of hy- drolysates and chromatography on a column (1.5 x 100 cm) of Sephadex G-15 were carried out as described in the text. Fractions (0.9 ml) were collected and analyzed for reducing groups. Fractions 69-76 (l), 77-82 (2 ) , 83-92 (3), 93-100 (4), 101-112 (5) and 11 3 - 132 (6) were pooled as indicated
the results of the NaI04 oxidation and the modified Morgan-Elson test described by Knox and Hall [25].
D 1 was a disaccharide composed of galactosamine and glucose with the former sugar at the reducing end. NaI04 oxidation of D1 led to disappearance of both sugars, while the glucose residue was retained on chromic anhydride oxidation. These results are in agreement with the structure Glc-a( 1 -+6)-GalNAc for D1. However, D1 was unaffected by either c(- or p-glucosidase. The disaccharide D2 consisted of glucos amine and galactosamine. The galactosamine proved to be the reducing terminus, but was not oxidized with NaI04. D2 was unsusceptible to 0-N-acetylhexos- aminidase and the glucosamine residue was not affected by chromic anhydride. From these results, the glucosamine residue appeared to be linked in a-configuration to either C-3 or C-4 of N-acetyl- galactosamine. This saccharide gave a greater color intensity as compared with N-acetylglucosamine in the modified Morgan-Elson test with heating at 100 "C for 35 min at pH 7.0 (Table 4). Thus, the most likely structure for D2 is GlcNAc-a( 1 + 3)-GalNAc. D3 was a disaccharide composed of glucosamine alone. Both of the glucosamine residues were oxidized either by NaI04 or by Cr03. D3 was digested with exo- p-N-acetylhexosaminidase to yield N-acetylglucos- amine. Therefore, D3 was most probably GlcNAc- p(l +6)-GlcNAc. D4 consisted of galactosamine and mannosamine. Mannosamine was at the reducing end. Cr03 oxidation did not affect the galactosamine residue. In addition, D4 was digested with a-N- acetylgalactosaminidase to yield N-acetylgalactosami-
5 20 Structural Components of Bacillus cereus Cell Walls
Table 4. Rates of the chromogen formation ,from isolated disac- charides in the modified Morgan-Elson iest Samples (20 nmol) were heated at 100 ;C for 35 min in 0.24 ml 0.05 M potassium phosphate buffer pH 7.0 [2S], then chromogens were estimated by the method of Reissig et al. [27]. Absorbance was measured at 585 nm and expressed relative to N-acetyl- glucosamine
Compound Chromogen formation
N - Acetylglucosamine 1 .00 GlcNAc-fi(1+4)-GlcNAc 0.07 D2, GlcNAc-GalNAc 2.11 D4, GalNAc-ManNAc 0.18 D5, ManNAc-GlcNAc 1.66 D6, GlcNAc-ManNAc 7.57
H
I
6 5 4 3
L i, 2
d (PPm)
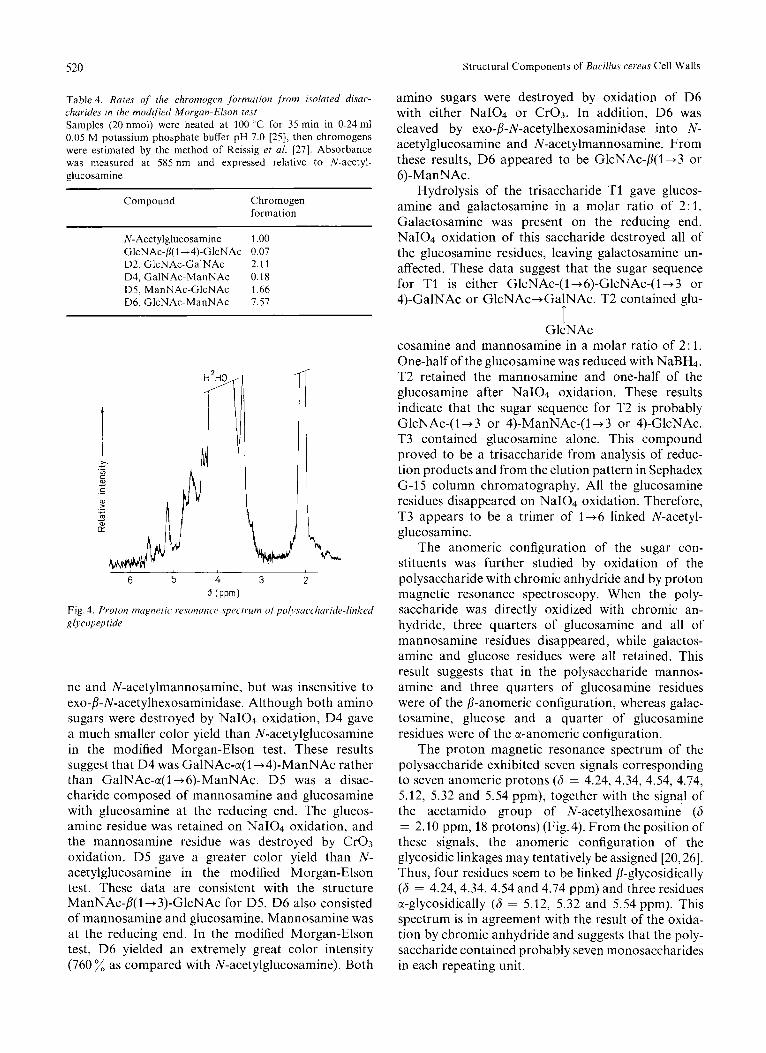
Fig. 4. Proton nicignetic wsonunc'e spectrum ~~fpol~.suc.cl~uride-linked glycopep tide
ne and N-acetylmannosamine, but was insensitive to exo-fi-N-acetylhexosaminidase. Although both amino sugars were destroyed by NaI04 oxidation, D4 gave a much smaller color yield than N-acetylglucosamine in the modified Morgan-Elson test. These results suggest that D4 was GalNAc-a(1+4)-ManNAc rather than GalNAc-a(1+6)-ManNAc. D5 was a disac- charide composed of mannosamine and glucosamine with glucosamine at the reducing end. The glucos- amine residue was retained on NaI04 oxidation, and the mannosamine residue was destroyed by Cr03 oxidation. D5 gave a greater color yield than N- acetylglucosamine in the modified Morgan-Elson test. These data are consistent with the structure ManNAc-fi( 1 + 3)-GlcNAc for D5. D6 also consisted of mannosamine and glucosamine. Mannosamine was at the reducing end. In the modified Morgan-Elson test, D6 yielded an extremely great color intensity (760 as compared with N-acetylglucosamine). Both
amino sugars were destroyed by oxidation of D6 with either NaI04 or Cr03. In addition, D6 was cleaved by exo-fi-N-acetylhexosaminidase into N- acetylglucosamine and N-acetylmannosamine. From these results, D6 appeared to be GlcNAc-P(1+3 or 6)-ManNAc.
Hydrolysis of the trisaccharide T1 gave glucos- amine and galactosamine in a molar ratio of 2: 1. Galactosamine was present on the reducing end. NaI04 oxidation of this saccharide destroyed all of the glucosamine residues, leaving galactosamine un- affected. These data suggest that the sugar sequence for T1 is either GlcNAc-(l+6)-GlcNAc-(l-+3 or 4)-GalNAc or G1cNAc-t GalNAc. T2 contained glu-
T GlcNAc
cosamine and mannosamine in a molar ratio of 2: 1. One-half of the glucosamine was reduced with NaBH4. T2 retained the mannosamine and one-half of the glucosamine after NaI04 oxidation. These results indicate that the sugar sequence for T2 is probably GlcNAc-( 1 -3 or 4)-ManNAc-(l-+3 or 4)-GlcNAc. T3 contained glucosamine alone. This compound proved to be a trisaccharide from analysis of reduc- tion products and from the elution pattern in Sephadex G-15 column chromatography. All the glucosamine residues disappeared on NaI04 oxidation. Therefore, T3 appears to be a trimer of 1+6 linked N-acetyl- glucosamine.
The anomeric configuration of the sugar con- stituents was further studied by oxidation of the polysaccharide with chromic anhydride and by proton magnetic resonance spectroscopy. When the poly- saccharide was directly oxidized with chromic an- hydride, three quarters of glucosamine and all of mannosamine residues disappeared, while galactos- amine and glucose residues were all retained. This result suggests that in the polysaccharide mannos- amine and three quarters of glucosamine residues were of the /I-anomeric configuration, whereas galac- tosamine, glucose and a quarter of glucosamine residues were of the a-anomeric configuration.
The proton magnetic resonance spectrum of the polysaccharide exhibited seven signals corresponding to seven anomeric protons (6 = 4.24, 4.34, 4.54, 4.74, 5.12, 5.32 and 5.54 ppm), together with the signal of the acetamido group of N-acetylhexosamine (6 = 2.10 ppm, 18 protons) (Fig. 4). From the position of these signals, the anomeric configuration of the glycosidic linkages may tentatively be assigned [20,26]. Thus, four residues seem to be linked fi-glycosidically (6 = 4.24,4.34,4.54 and 4.74 ppm) and three residues ct-glycosidically (6 = 5.12, 5.32 and 5.54 ppm). This spectrum is in agreement with the result of the oxida- tion by chromic anhydride and suggests that the poly- saccharide contained probably seven monosaccharides in each repeating unit.

K. Ammo, S. Hazama, Y. Araki, and E. Ito 52 1
DISCUSSION
The studies described in this paper show that the cell walls of B. cereus AHU 1356 are composed of peptidoglycan, polysaccharide and teichoic acid. The polysaccharide and the teichoic acid appear to account approximately for 50 and 5%, respectively, of the weight of the cell walls. Neither amino acids other than the peptidoglycan constituents nor uronic acids were detected in significant amounts in the hydrolysates of the N-acetylated cell walls.
After fractionation of the lysozyme digest of the N-acetylated cell walls, more than 70% of 6-phos- phorylated muramic acid residues of the cell walls was recovered in the two polymer fractions, although only 10 % of total muramic acid residues was present in these fractions. As calculated on the basis of the approximate molecular weight of the polysaccharide- glycopeptide complex (33000), each molecule of the complex appears to contain, on the average, one 6-phosphorylated muramic acid residue. Mild acid hydrolysis cleaved the linkage between the glyco- peptide and the polysaccharide which consisted of glucosamine, mannosamine, galactosamine and glu- cose in a molar ratio of 4: 1 : 1 : 1. The above result suggests that this polysaccharide in the cell walls is bound to the peptidoglycan through an acid-labile phosphodiester linkage involving C-1 of the reducing terminal sugar of the polysaccharide chain and C-6 of the muramic acid residue of the peptidoglycan. A similar type of attachment of the wall polysaccharide to peptidoglycan has been proposed in other cell walls [25,28,29].
The result from the Smith degradation shows that the polysaccharide of B. cereus cell walls probably has a backbone chain, composed of 1 : l : l ratio of glucosamine, mannosamine and galactosamine bound to one another through 1,3 or 1,4 linkages, and branches containing glucose and three quarters of the total glucosamine residues. The analysis of the di- and trisaccharides obtained from partial acid hydroly- sis of the polysaccharide-linked glycopeptide led to the tentative formulation of the partial structures shown in Scheme 1. From the structures of D4 and D5, the backbone chain of the polysaccharide appears to possess the following repeating unit : GalNAc-a( 1 +4)- ManNAc-/3( 1 + 3)-GlcNAc. In addition, the indicat- ed partial structures suggest the binding of the other monosaccharide components as the branches to the backbone constituents, that is, the binding of glucose to C-6 of N-acetylgalactosamine and of N-acetyl- glucosamine to C-3 of N-acetylmannosamine. The remaining two glucosamine residues appear to be bound as a disaccharide GlcNAc-( 1 +6)-GlcNAc to C-6 of N-acetylglucosamine of the backbone chain. Other ways of binding of the two glucosamine residues seem unlikely because of failure of explaining the
D1
D2
D3
D4
D5
D6
T1
T2
T3
Glc 1
4 6
PI:
GalNAc
GlcNAc-r(1+3)-GalNAc
GlcNAc
GlcNAc
GalNAc-r(1+4)-ManNAc
ManNAc-p(1+3)-GlcNAc
GlcNAc 1
P 13 or 6
ManNAc
GlcNAc
I: GlcNAc-(l+3 or 4)-GalNAc or
GlcNAc
GlcNAc-GalNAc
GlcNAc
1
I' 3 o r 4 ManNAc-(I +3 or 4)-GlcNAc
GlcNAc I: I: GlcNAc
GlcNAc Scheme 1. Structure of'oligosucchuridcs
formation of fragments T1 and T3. Therefore, the structure of the polysaccharide of B. cereus AHU 1356 cell walls is most likely
GlcNAc
11: Glc GlcNAc GlcNAc
The configuration of glycosidic linkages in the polysaccharide was deduced from enzymic digestion of the disaccharides and from chromic anhydride oxidation of the disaccharides and polysaccharide. The oxidation of the polysaccharide destroyed man- nosamine and three quarters of the glucosamine residues. The proton magnetic resonance spectrum of the polysaccharide also supports this structure. The spectrum indicates the occurrence of seven differ-

522 K. Amano, S. Hazama, Y. Araki, and E. Ito: Structural Components of Bacillus cereus Cell Walls
ent glycosidic linkages of which four seem to be of P-configuration and three of a-configuration. In ad- dition, the spectrum suggests that the amino sugars in the polysaccharide were all N-acetylated. The amino sugar components of the polysaccharide appear to be N-acetylated also in the intact cell walls of this strain, since it has previously been shown that the amino sugars in the cell walls of this strain are all N-substituted except glucosamine in peptidoglycan [2]. The molecular weight of the polysaccharide, estimated from gel filtration (28000) and from the muramic acid 6-phosphate content in the glyco- peptide moiety (34000), suggests the presence of more than 20 repeating units. To establish the linkages of the monosaccharide residues in the polysaccharide, especially of the glucosamine residues in the branches, further work is required which may include the search for enzymes capable of hydrolyzing glycosides of N-acetylmannosamine as well as studies on the chem- istry of derivatives of this amino sugar.
REFERENCES
1. Hughes, R. C. (1971) Biochem. J . 121, 791-802. 2. Araki, Y., Nakatani, T., Nakayama, K. & Ito, E. (1972) J .
3. Higginbotham, J. D., Heidelberger, M . & Gotschlich. E. C.
4. Higginbotham, J . D. & Heidelberger, M. (1972) Curhohj~lr.
5. Higginbotham, J . D., Das, A. & Heidelberger, M. (1972) Bio-
6. Das, A., Higginbotham, J . D. & Heidelberger, M. (1972) Bio-
Biol. Cliem. 247, 6312-6322.
(1970) Proc. Nail Acatl. Sci. U.S.A. 67, 138-142.
R ~ s . 23, 165-173.
chem. J . I26.225-231.
clrc~m. J . 126. 233 - 236.
7. Liu, T.-Y., Gotschlich, E. C., Jonssen, E. K. & Wysocki, J . R.
8. Vann, W. F., Liu, T.-Y. & Robbins, J. B. (1976) Infect. Immun.
9. Rupley, J. A. (1964) Biochim. Biophys. Acta, 83, 245-255. 10. Goldstein, I . J., Hay, G. W., Lewis, B. A. & Smith, F. (1965)
11. Tipper, D. J. & Strominger, J . L. (1966) Biochem. Biophys. Rus.
12. Park, J . T. & Johnson, M. J . (1949) J . Biol. Chem. 181, 149-
13. Dubois, M., Gilles, K. A,, Hamilton, J. K., Robers, P. A. &
14. Lowry, 0. H., Roberts, N. R., Leiner, K. W., Wu, M.-L. &
15. Huggett, A. St. G. & Nixon, D. A . (1957) Biochem. J . 66,
16. Hanahan, D. J. & Olley, J . N. (1958) J . B id . Chem. 231,
17. Wieland, 0. (1974) in Merhod of Enzymaric Analj~sis, 2nd edn (Bergmeyer, H. U., ed.) pp. 1404- 1409, Verlag Chemie, Weinheim/Bergstr.
18. Angyal, S. J. &James, K. (1970) Carbohydr. Rrs. 12, 147- 151. 19. Hoffman, J., Lindberg, B. & Svensson, S. (1972) Acra Chem.
20. Dmitriev, B. A, , Lvov, V. L., Kochetkov, N. K., Jann, B. &
21. Leloir, L. F., Staneloni, R. L., Carminatti, H . & Behrens, H . N.
22. Sharon, N. & Seifter, S. (1964) J . B id . Chern. 239, 2398-2402. 23. Trevelyan, W. E., Procter, D. P. & Harrison, J . C. (1950)
24. Hayashi, H., Araki, Y . & Ito, E. (1973) J . Bacterial. 113.
25. Knox, K. W. & Hall, E. A. (1965) Biochen?. J . Y6, 302-309. 26. Choy, Y. M. & Dutton, G. G. A. (1973) Can. J . Chenz. 51,
27. Reissig, J . L., Strominger, J . L. & Leloir, L. F. (1955) J . Biol.
28. Knox, K. W. & Holmwood, K. J. (1968) Biochem. J . 108,
29. Hughes, R. C. (1970) Biochem. J . 117, 431 -439.
(1971) J . Biol. Chem. 246, 2849-2858.
13, 1654- 1662.
Methods Carhohydr. Chum. 5, 361 - 370.
Commun. 22,48 - 56.
151.
Smith, F. (1956) Anal. Chem. 28, 350-356.
Farr, A. L. (1954) J . Biol. Chem. 207, 1 - 17.
12p.
813 - 828.
Scund. 26, 661 - 666.
Jann, K. (1976) Eur. J . Biochem. 64, 491 -498.
(1973) Biochem. Biophvs. Res. Commun. 52, 1285- 1292.
A'uture (Lond.) 166, 444 - 445.
592-598.
198-207.
Chrm. 217,959-966.
363 - 368.
K. Amano, S. Hazama, Y . Araki, and E. Ito, Department of Chemistry, Faculty or Science, Hokkaido University, Kita-10-jyo, Nishi-8-chome, Kita-ku, Sapporo, Japan 060