involvement of the adenosine a 1 and a 2a receptors in the antidepressant-like effect of zinc in the...
TRANSCRIPT

Available online at www.sciencedirect.com
logical Psychiatry 32 (2008) 994–999www.elsevier.com/locate/pnpbp
Progress in Neuro-Psychopharmacology & Bio
Involvement of the adenosine A1 and A2A receptors in the antidepressant-likeeffect of zinc in the forced swimming test
Kelly R. Lobato a, Ricardo W. Binfaré a, Josiane Budni a, Angelo Oscar Rosa a,Adair Roberto S. Santos b, Ana Lúcia S. Rodrigues a,⁎
a Departamento de Bioquímica, Centro de Ciências Biológicas, Universidade Federal de Santa Catarina,Campus Universitário — Trindade — 88040-900, Florianópolis-SC, Brazil
b Departamento de Ciências Fisiológicas, Centro de Ciências Biológicas, Universidade Federal de Santa Catarina,Campus Universitário — Trindade — 88040-900, Florianópolis-SC, Brazil
Received 19 November 2007; received in revised form 4 January 2008; accepted 15 January 2008Available online 26 January 2008
Abstract
It was previously shown that the acute administration of zinc chloride elicits an antidepressant-like effect in the mouse forced swimming test(FST). We have also shown that the activation of adenosine A1 and A2A receptors produces an antidepressant-like effect in FST. Thus, this studyinvestigated the involvement of adenosine receptors in the antidepressant-like effect of zinc in the FST. The antidepressant-like effect of ZnCl2(30 mg/kg, i.p.) in the FST was prevented by the pretreatment of animals with caffeine (3 mg/kg, i.p., a non-selective adenosine receptorantagonist), DPCPX (2 mg/kg, i.p., a selective adenosine A1 receptor antagonist) or ZM241385 (1 mg/kg, i.p., a selective adenosine A2A receptorantagonist), administered at doses that per se produced no anti-immobility effect. Moreover, the treatment of mice with CHA (0.05 mg/kg, i.p., aselective adenosine A1 receptor agonist), DPMA (0.1 mg/kg, i.p., a selective adenosine A2A receptor agonist) or dipyridamole (0.1 µg/site, i.c.v.,an adenosine transporter inhibitor) was able to potentiate the action of sub-effective doses of ZnCl2. Taken together, the results suggest that theantidepressant-like effect of zinc in the mouse FST might involve a direct or indirect activation of adenosine A1 and A2A receptors.© 2008 Elsevier Inc. All rights reserved.
Keywords: Adenosine; Antidepressant; Depression; Forced swimming test; Zinc
1. Introduction
Zinc is a transition metal that serves as a cellular signalingmolecule, present at high concentrations in the central nervoussystem, particularly in the hippocampus and cerebral cortex(Vallee and Falchuk, 1993; Nowak et al., 2005) where it producesa wide variety of neuromodulatory effects (Choi and Koh, 1998).Several preclinical and clinical studies indicate an important roleof zinc in the pathophysiology and therapy of depression. Clinical
Abbreviations: CHA, N6-cyclohexyladenosine caffeine; DPCPX, 8-cyclo-pentyl-1,3-dipropylxanthine; DPMA, N6-[2-(3,5-Dimethoxyphenyl)-2-(methyl-phenyl)ethyl]adenosine; FST, forced swimming test; i.c.v., intracerebroventricular;i.p., intraperitoneal; NMDA, N-methyl-D-aspartate; ZM241385, 4-(2-[7-amino-2-{2-furyl}{1,2,4}triazolo-{2,3-a}{1,3,5}triazin-5-yl-amino]ethyl)-phenol.⁎ Corresponding author. Tel.: +55 48 37215043; fax: +55 48 37219672.E-mail addresses: [email protected], [email protected]
(A.L.S. Rodrigues).
0278-5846/$ - see front matter © 2008 Elsevier Inc. All rights reserved.doi:10.1016/j.pnpbp.2008.01.012
reports show that human depression might be accompanied withlower serum zinc concentrations (McLoughlin and Hodge, 1990;Maes et al., 1994, 1997, 1999; Nowak et al., 2005). Besides that,the plasma zinc level in depressed patients after recovery wasfound to be higher than in depressed patients (McLoughlin andHodge, 1990; Narang et al., 1991).
Preclinical studies have shown that zinc exhibits antidepres-sant-like effects in tests used for evaluation of antidepressantactivity. Zinc produces antidepressant-like effects in the forcedswimming test (FST), both in mice and rats and in tail suspensiontest in mice (Kroczka et al., 2001, 2002; Nowak et al., 2003b;Rosa et al., 2003). This effect in the FST is mediated, at least inpart, by the inhibition ofN-methyl-D-aspartate (NMDA) receptorsand an interaction with the L-arginine-nitric oxide pathway (Rosaet al., 2003). Additionally, low doses of zinc administeredtogether with ineffective doses of imipramine or citalopram en-hanced antidepressant-like effect in the mouse FST (Kroczka

995K.R. Lobato et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 994–999
et al., 2001; Szewczyk et al., 2002), suggesting that zinc maycontribute for antidepressant action. Moreover, our group hasrecently shown that zinc administration was able to attenuate themalathion induced-depressant-like effects in the FST in rats(Brocardo et al., 2007). Together, the results suggest that zincmaybe a metal with antidepressant properties.
The purine nucleoside adenosine acts as an extracellularsignaling molecule influencing synaptic transmission andmodulating the activity of the nervous system (Ribeiro et al.,2003). The involvement of adenosine in the pathophysiology ofdepression and in antidepressant action is suggested by clinicaldata (Phillis and Wu, 1982; Phillis, 1984; Berk et al., 2001). Inaddition, previous clinical studies have also shown a relationshipbetween the enzyme adenosine deaminase, major depressionand antidepressant activity. A study from Elgün et al. (1999)indicated an inverse relationship between the activity of thisenzyme and the severity of depression. Regarding this issue, arecent study reported that the treatment with selective serotoninreuptake inhibitors increases the adenosine deaminase activity(Herken et al., 2007). Additional evidence for a role of adenosinein the pathophysiology of depression was given in the last yearsin preclinical studies. It was reported that adenosine and theselective A1 and A2A agonists CHA and DPMA produceantidepressant-like effect in the FST through a mechanism thatinvolves the activation of these receptors (Kaster et al., 2004).
Therefore, considering the involvement of zinc in depressiondemonstrated by clinical and preclinical data (McLoughlin andHodge, 1990; Narang et al., 1991; Maes et al., 1994, 1997,1999; Kroczka et al., 2001, 2002; Rosa et al., 2003; Nowaket al., 2003a,b, 2005; Brocardo et al., 2007) and the alreadyknown participation of adenosine and adenosine receptors in thepathophysiology of depression (Phillis and Wu, 1982; Phillis,1984; Berk et al., 2001; El Yacoubi et al., 2001; Kaster et al.,2004, 2005, 2007, Herken et al., 2007), we sought to investigatethe involvement of the adenosine A1 and A2A receptors in theeffect of zinc in the FST, a test widely used to screen newantidepressant drugs and to investigate the mechanisms under-lying the antidepressant action of drugs (Cryan et al., 2005).This study may help to further contribute to the understandingof the mechanisms underlying the antidepressant-like effects ofzinc, which are not fully established.
2. Methods
2.1. Animals
Swiss mice of either sex (30–40 g) were maintained atconstant room temperature (22–24 °C) with free access towater and food, under a 12:12 h light:dark cycle (lights on at07:00 h). All experiments were carried out between 11:00 and17:00 h, with each animal used only once (N=6–8 animals pergroup). The procedures in this study were performed inaccordance with the National Institute of Health Guide for theCare and Use of Laboratory Animals and approved by theEthics Committee of the Institution. All efforts were made tominimize animals suffering and to reduce the number of ani-mals used in the experiments.
2.2. Drugs and treatment
Zinc chloride, N6-cyclohexyladenosine (CHA), caffeine, N6-[2-(3,5-Dimethoxyphenyl)-2-(methylphenyl)ethyl]adenosine(DPMA) (Sigma Chemical Co., USA), 4-(2-[7-amino-2-{2-furyl}{1,2,4}triazolo-{2,3-a}{1,3,5}triazin-5-yl-amino]ethyl)-phenol (ZM241385), 8-cyclopentyl-1,3-dipropylxanthine(DPCPX) and dipyridamole (Tocris Cookson, USA) were used.Dipyridamole was administered by intracerebroventricular (i.c.v.)route, under light ether anesthesia, directly into the lateral ven-tricle, in a volume of 5 μl per mouse, given over 30 s, and thecannula remained in place for another 30 s, as previouslydescribed (Kaster et al., 2007). All other drugs were administeredby intraperitoneal (i.p.) route in a constant volume of 10 ml/kgbody weight. Drugs were dissolved in saline except DPMA,ZM241385 and DPCPX that were dissolved in saline with 5%DMSO. Control animals received appropriate vehicle.
In order to investigate the involvement of the adenosine A1
and A2 receptors in the antidepressant-like effect of zinc in theFST, animals were pretreated with caffeine (3 mg/kg, i.p., a non-selective adenosine receptor antagonist), DPCPX (2 mg/kg, i.p.,a selective adenosine A1 receptor antagonist), ZM241385 (1 mg/kg, i.p., a selective adenosine A2A receptor antagonist) or withappropriate vehicle. After 30 min the animals received zincchloride (30 mg/kg, i.p.) or vehicle before being tested in theFST 30 min later.
In a separate set of experiments, in order to investigate apossible synergistic effect between zinc chloride and adenosiner-gic agonists, animals were treated with a sub-effective dose ofzinc chloride (10 mg/kg, i.p.) or vehicle 20 min after the pre-treatment with CHA (0.05 mg/kg, i.p., a selective adenosine A1
receptor agonist), DPMA (0.1 mg/kg, i.p., a selective adenosineA2A receptor agonist) or vehicle.A further 30minwere allowed toelapse before the animals were tested in the FST. Alternatively,animals were pretreated with dipyridamole (0.1 µg/site, i.c.v., anadenosine transporter inhibitor) 15 min before zinc chloride(10 mg/kg, i.p.) administration, and 20 min later the FST wascarried out. Control animals received appropriate vehicle.
The doses of zinc chloride were chosen based on a previouslyreported dose–response curve of this metal in the mouse forcedswimming test (Rosa et al., 2003). The doses of the adenosinereceptor agonists and antagonists were selected on the basis ofprevious results from our laboratory (Kaster et al., 2004) andthey do not alter locomotor activity in the open-field test (datanot shown).
2.3. Forced swimming test (FST)
Mice were individually forced to swim in an open cylindricalcontainer (diameter 10 cm, height 25 cm), containing 19 cm ofwater at 25±1 °C; the total duration of immobility during a6 min test was scored as described previously (Kaster et al.,2004, 2005). Each mouse was judged to be immobile when itceased struggling and remained floating motionless in the water,making only those movements necessary to keep its head abovewater. A decrease in the duration of immobility is indicative ofan antidepressant-like effect (Porsolt et al., 1977).
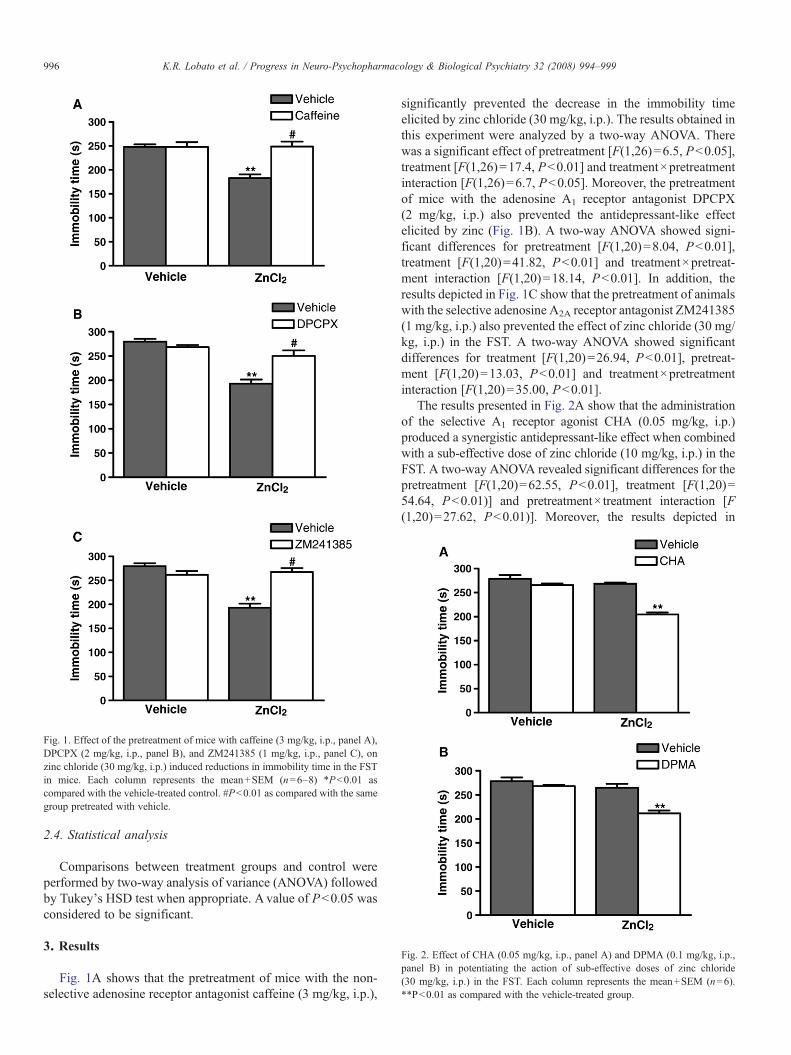
Fig. 1. Effect of the pretreatment of mice with caffeine (3 mg/kg, i.p., panel A),DPCPX (2 mg/kg, i.p., panel B), and ZM241385 (1 mg/kg, i.p., panel C), onzinc chloride (30 mg/kg, i.p.) induced reductions in immobility time in the FSTin mice. Each column represents the mean+SEM (n=6–8) *Pb0.01 ascompared with the vehicle-treated control. #Pb0.01 as compared with the samegroup pretreated with vehicle.
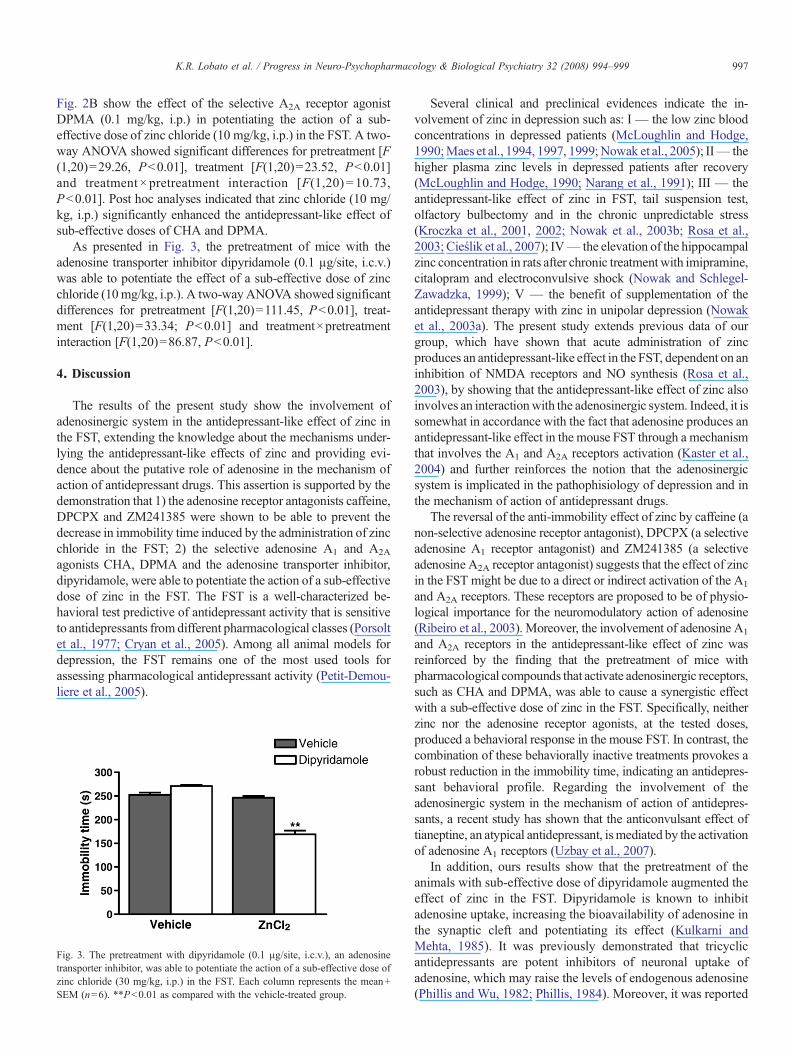
Fig. 2. Effect of CHA (0.05 mg/kg, i.p., panel A) and DPMA (0.1 mg/kg, i.p.,panel B) in potentiating the action of sub-effective doses of zinc chloride(30 mg/kg, i.p.) in the FST. Each column represents the mean+SEM (n=6).**Pb0.01 as compared with the vehicle-treated group.
996 K.R. Lobato et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 994–999
2.4. Statistical analysis
Comparisons between treatment groups and control wereperformed by two-way analysis of variance (ANOVA) followedby Tukey's HSD test when appropriate. A value of Pb0.05 wasconsidered to be significant.
3. Results
Fig. 1A shows that the pretreatment of mice with the non-selective adenosine receptor antagonist caffeine (3 mg/kg, i.p.),
significantly prevented the decrease in the immobility timeelicited by zinc chloride (30 mg/kg, i.p.). The results obtained inthis experiment were analyzed by a two-way ANOVA. Therewas a significant effect of pretreatment [F(1,26)=6.5, Pb0.05],treatment [F(1,26)=17.4, Pb0.01] and treatment×pretreatmentinteraction [F(1,26)=6.7, Pb0.05]. Moreover, the pretreatmentof mice with the adenosine A1 receptor antagonist DPCPX(2 mg/kg, i.p.) also prevented the antidepressant-like effectelicited by zinc (Fig. 1B). A two-way ANOVA showed signi-ficant differences for pretreatment [F(1,20)=8.04, Pb0.01],treatment [F(1,20)=41.82, Pb0.01] and treatment×pretreat-ment interaction [F(1,20)=18.14, Pb0.01]. In addition, theresults depicted in Fig. 1C show that the pretreatment of animalswith the selective adenosine A2A receptor antagonist ZM241385(1 mg/kg, i.p.) also prevented the effect of zinc chloride (30 mg/kg, i.p.) in the FST. A two-way ANOVA showed significantdifferences for treatment [F(1,20)=26.94, Pb0.01], pretreat-ment [F(1,20)=13.03, Pb0.01] and treatment×pretreatmentinteraction [F(1,20)=35.00, Pb0.01].
The results presented in Fig. 2A show that the administrationof the selective A1 receptor agonist CHA (0.05 mg/kg, i.p.)produced a synergistic antidepressant-like effect when combinedwith a sub-effective dose of zinc chloride (10 mg/kg, i.p.) in theFST. A two-way ANOVA revealed significant differences for thepretreatment [F(1,20)=62.55, Pb0.01], treatment [F(1,20)=54.64, Pb0.01)] and pretreatment× treatment interaction [F(1,20)=27.62, Pb0.01)]. Moreover, the results depicted in
997K.R. Lobato et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 994–999
Fig. 2B show the effect of the selective A2A receptor agonistDPMA (0.1 mg/kg, i.p.) in potentiating the action of a sub-effective dose of zinc chloride (10 mg/kg, i.p.) in the FST. A two-way ANOVA showed significant differences for pretreatment [F(1,20)=29.26, Pb0.01], treatment [F(1,20)=23.52, Pb0.01]and treatment × pretreatment interaction [F(1,20) = 10.73,Pb0.01]. Post hoc analyses indicated that zinc chloride (10 mg/kg, i.p.) significantly enhanced the antidepressant-like effect ofsub-effective doses of CHA and DPMA.
As presented in Fig. 3, the pretreatment of mice with theadenosine transporter inhibitor dipyridamole (0.1 µg/site, i.c.v.)was able to potentiate the effect of a sub-effective dose of zincchloride (10mg/kg, i.p.). A two-wayANOVA showed significantdifferences for pretreatment [F(1,20)=111.45, Pb0.01], treat-ment [F(1,20)=33.34; Pb0.01] and treatment×pretreatmentinteraction [F(1,20)=86.87, Pb0.01].
4. Discussion
The results of the present study show the involvement ofadenosinergic system in the antidepressant-like effect of zinc inthe FST, extending the knowledge about the mechanisms under-lying the antidepressant-like effects of zinc and providing evi-dence about the putative role of adenosine in the mechanism ofaction of antidepressant drugs. This assertion is supported by thedemonstration that 1) the adenosine receptor antagonists caffeine,DPCPX and ZM241385 were shown to be able to prevent thedecrease in immobility time induced by the administration of zincchloride in the FST; 2) the selective adenosine A1 and A2A
agonists CHA, DPMA and the adenosine transporter inhibitor,dipyridamole, were able to potentiate the action of a sub-effectivedose of zinc in the FST. The FST is a well-characterized be-havioral test predictive of antidepressant activity that is sensitiveto antidepressants from different pharmacological classes (Porsoltet al., 1977; Cryan et al., 2005). Among all animal models fordepression, the FST remains one of the most used tools forassessing pharmacological antidepressant activity (Petit-Demou-liere et al., 2005).
Fig. 3. The pretreatment with dipyridamole (0.1 µg/site, i.c.v.), an adenosinetransporter inhibitor, was able to potentiate the action of a sub-effective dose ofzinc chloride (30 mg/kg, i.p.) in the FST. Each column represents the mean+SEM (n=6). **Pb0.01 as compared with the vehicle-treated group.
Several clinical and preclinical evidences indicate the in-volvement of zinc in depression such as: I— the low zinc bloodconcentrations in depressed patients (McLoughlin and Hodge,1990;Maes et al., 1994, 1997, 1999;Nowak et al., 2005); II— thehigher plasma zinc levels in depressed patients after recovery(McLoughlin and Hodge, 1990; Narang et al., 1991); III — theantidepressant-like effect of zinc in FST, tail suspension test,olfactory bulbectomy and in the chronic unpredictable stress(Kroczka et al., 2001, 2002; Nowak et al., 2003b; Rosa et al.,2003; Cieślik et al., 2007); IV— the elevation of the hippocampalzinc concentration in rats after chronic treatment with imipramine,citalopram and electroconvulsive shock (Nowak and Schlegel-Zawadzka, 1999); V — the benefit of supplementation of theantidepressant therapy with zinc in unipolar depression (Nowaket al., 2003a). The present study extends previous data of ourgroup, which have shown that acute administration of zincproduces an antidepressant-like effect in the FST, dependent on aninhibition of NMDA receptors and NO synthesis (Rosa et al.,2003), by showing that the antidepressant-like effect of zinc alsoinvolves an interactionwith the adenosinergic system. Indeed, it issomewhat in accordance with the fact that adenosine produces anantidepressant-like effect in the mouse FST through a mechanismthat involves the A1 and A2A receptors activation (Kaster et al.,2004) and further reinforces the notion that the adenosinergicsystem is implicated in the pathophisiology of depression and inthe mechanism of action of antidepressant drugs.
The reversal of the anti-immobility effect of zinc by caffeine (anon-selective adenosine receptor antagonist), DPCPX (a selectiveadenosine A1 receptor antagonist) and ZM241385 (a selectiveadenosine A2A receptor antagonist) suggests that the effect of zincin the FST might be due to a direct or indirect activation of the A1
and A2A receptors. These receptors are proposed to be of physio-logical importance for the neuromodulatory action of adenosine(Ribeiro et al., 2003). Moreover, the involvement of adenosine A1
and A2A receptors in the antidepressant-like effect of zinc wasreinforced by the finding that the pretreatment of mice withpharmacological compounds that activate adenosinergic receptors,such as CHA and DPMA, was able to cause a synergistic effectwith a sub-effective dose of zinc in the FST. Specifically, neitherzinc nor the adenosine receptor agonists, at the tested doses,produced a behavioral response in the mouse FST. In contrast, thecombination of these behaviorally inactive treatments provokes arobust reduction in the immobility time, indicating an antidepres-sant behavioral profile. Regarding the involvement of theadenosinergic system in the mechanism of action of antidepres-sants, a recent study has shown that the anticonvulsant effect oftianeptine, an atypical antidepressant, ismediated by the activationof adenosine A1 receptors (Uzbay et al., 2007).
In addition, ours results show that the pretreatment of theanimals with sub-effective dose of dipyridamole augmented theeffect of zinc in the FST. Dipyridamole is known to inhibitadenosine uptake, increasing the bioavailability of adenosine inthe synaptic cleft and potentiating its effect (Kulkarni andMehta, 1985). It was previously demonstrated that tricyclicantidepressants are potent inhibitors of neuronal uptake ofadenosine, which may raise the levels of endogenous adenosine(Phillis and Wu, 1982; Phillis, 1984). Moreover, it was reported

998 K.R. Lobato et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 994–999
that chronic treatment with imipramine in rats enhanced thepresynaptic inhibitory effect of adenosine in the field excitatorypostsynaptic potential in the hippocampus (Zahorodna et al.,2002). The synergistic effect of dypiridamole and zinc in theFST might be related to a possible inhibition of the uptake ofadenosine, thus raising the nucleoside levels in the synaptic cleftand potentiating its activity. Thus, zinc may share the samemechanism of classical antidepressants, interacting with theadenosine transporter and directly and/or indirectly increasingadenosinergic transmission. In line with this, it is interesting tomention that zinc is proposed to play a role as a neuromodulatorby affecting the function of neurotransmitter transporters (Nør-gaard-Nielsen and Gether, 2006). Zinc inhibits the transportersfor glutamate (Vandenberg et al., 1998), dopamine (Norregaardet al., 1998), glycine (Ju et al., 2004) and GABA (Cohen-Kfiret al., 2005). Thus, the hypothesis that zinc also inhibits theadenosine transporter deserves further studies.
Zinc might influence a wide range of enzymatic reactions(Nowak and Szewczyk, 2002). In this regard, it may affect theactivity of the enzyme adenosine deaminase, which catalyzes thedeamination of adenosine (or 2''deoxyadenosine) yielding ino-sine (or 2'-deoxyinosine) and ammonia (Cooper et al., 1997). Astudy from Elgün et al. (1999) has shown that serum activity ofadenosine deaminase was decreased in patients with majordepression, as compared to controls, and there is an inverserelationship between the enzyme activity and the severity ofdepression. Therefore, we cannot rule out that the antidepressant-like effect elicited by zinc in this study might be dependent of aninteraction with the enzyme adenosine deaminase. However thishypothesis deserves further investigation.
5. Conclusion
Our study extends literature data regarding the mechanismsunderlying the antidepressant-like effect of zinc in the FST. Herewe show, to our knowledge for the first time, that the antidep-ressant-like effect of zinc depends, at least in part, on an inter-action with the adenosine A1 and A2A receptors. We suggest thatthis effect might be due to a direct effect of zinc at these receptorsand/or indirectly mediated by a zinc-induced augmentation ofbioavailability of adenosine in the synaptic cleft.
References
Berk M, Plein H, Ferreira D, Jersky B. Blunted adenosine A2A receptor functionin platelets in patients with major depression. Eur Neuropsychopharmacol2001;11:183–6.
Brocardo PS, Assini F, Franco JL, Pandolfo P, Müller YM, Takahashi RN, et al.Zinc attenuates malathion-induced depressant-like behavior and confersneuroprotection in the rat brain. Toxicol Sci 2007;97:140–8.
Cieślik K, Klenk-Majewska B, Danilczuk Z, Wróbel A, Łupina T, Ossowska G.Influence of zinc supplementation on imipramine effect in a chronicunpredictable stress (CUS) model in rats. Pharmacol Rep 2007;59:46–52.
Choi DW, Koh JY. Zinc and brain injury. Annu Rev Neurosci 1998;21:347–75.Cohen-Kfir E, Lee W, Eskandari S, Nelson N. Zinc inhibition of gamma-
aminobutyric acid transporter 4 (GAT4) reveals a link between excitatory andinhibitory neurotransmission. Proc Natl Acad Sci U S A 2005;102:6154–9.
Cooper BF, Sideraki V,Wilson DK, Dominguez DY, Clark SW, Quiocho FA, et al.The role of divalent cations in structure and function of murine adenosinedeaminase. Protein Sci 1997;6:1031–7.
Cryan JF, Valentino RJ, Lucki I. Assessing substrates underlying the behavioraleffects of antidepressants using the modified rat forced swimming test.Neurosci Biobehav Rev 2005;29:547–69.
Elgün S, Keskinege A, Kumbasar H. Dipeptidyl peptidase IV and adenosinedeaminase activity. Decrease in depression. Psychoneuroendocrinology1999;24:823–32.
El YacoubiM, Ledent C, Parmentier M, Bertorelli R, Ongini E, Costentin J, et al.Adenosine A2A receptor antagonists are potential antidepressants: evidencebased on pharmacology and A2A receptor knockout mice. Br J Pharmacol2001;134:68–77.
Herken H, Gurel A, Selek S, Armutcu F, Ozen ME, Bulut M, et al. Adenosinedeaminase, nitric oxide, superoxide dismutase, and xanthine oxidase inpatients with major depression: impact of antidepressant treatment. ArchMed Res 2007;38:247–52.
Ju P, Aubrey KR, Vandenberg RJ. Zn2+ inhibits glycine transport by glycinetransporter subtype 1b. J Biol Chem 2004;279:22983–91.
Kaster MP, Budni J, Binfaré RW, Santos ARS, Rodrigues ALS. The inhibition ofdifferent types of potassium channels underlies the antidepressant-like effectof adenosine in the mouse forced swimming test. Prog Neuropsychophar-macol Biol Psychiatry 2007;31:690–6.
Kaster MP, Rosa AO, Rosso MM, Goulart EC, Santos ARS, Rodrigues ALS.Adenosine administration produces an antidepressant-like effect in mice:evidence for the involvement of A1 and A2A receptors. Neurosci Lett2004;355:21–4.
Kaster MP, Rosa AO, Santos ARS, Rodrigues ALS. Involvement of nitric oxide-cGMP pathway in the antidepressant-like effects of adenosine in the forcedswimming test. Int J Neuropsychopharmacol 2005;8:601–6.
Kroczka B, Brański P, Palucha A, Pilc A, NowakG. Antidepressant-like propertiesof zinc in rodent forced swim test. Brain Res Bull 2001;55:297–300.
Kroczka B, Zieba A, Dudek D, Pilc A, Nowak G. Zinc exhibits an anti-depressant-like effect in the forced swimming test in mice. Pol J Pharmacol2002;52:403–6.
Kulkarni SK, Mehta AK. Purine nucleoside-mediated immobility in mice:reversal by antidepressants. Psychopharmacology 1985;85:460–3.
Maes M, De Vos N, Demedts P, Wautures A, Neels H. Lower serum zinc inmajor depression in relation to changes in serum acute phase proteins. JAffect Disord 1999;56:189–94.
Maes M, D'Haese PC, Scharpe S, D'Hondt PD, Cosyns P, De Broe ME.Hypozincemia in depression. J Affect Disord 1994;31:135–40.
Maes M, Vandoolaeghe E, Neels H, Demedts P, Wauters A, Meltzer HY, et al.Lower serum zinc in major depression is a sensitive marker of treatmentresistance and of the immune-inflammatory response in that illness. BiolPsychiatry 1997;42:349–58.
McLoughlin IJ, Hodge SJ. Zinc in depressive disorder. Acta Psychiatr Scand1990;82:451–3.
Narang RL, Gupta KR, Narang AP, Singh R. Levels of copper and zinc indepression. Indian J Physiol Pharmacol 1991;35:272–4.
Nørgaard-Nielsen K, Gether U. Zn2+ modulation of neurotransmitter transpor-ters. Handb Exp Pharmacol 2006;175:1–22.
Norregaard L, Frederiksen D, Nielsen EO, Gether U. Delineation of anendogenous zinc-binding site in the human dopamine transporter. EMBO J1998;17:4266–73.
Nowak G, Schlegel-Zawadzka M. Alterations in serum and brain trace elementlevels after antidepressant treatment: part I. Zinc. Biol Trace Elem Res1999;67:85–92.
Nowak G, Szewczyk B. Mechanisms contributing to antidepressant zinc actions.Pol J Pharmacol 2002;54:587–92.
Nowak G, Szewczyk B, Pilc A. Zinc and depression. An update. Pharmacol Rep2005;57:713–8.
Nowak G, Siwek M, Dudek D, Zieba A, Pilc A. Effect of zinc supplementationon antidepressant therapy in unipolar depression: a preliminary placebo-controlled study. Pol J Pharmacol 2003a;55:1143–7.
Nowak G, Szewczyk B, Wierońska JM, Brański P, Palucha A, Pilc A, et al.Antidepressant-like effects of acute and chronic treatment with zinc in forcedswim test and olfactory bulbectomy model in rats. Brain Res Bull2003b;61:159–64.
Petit-Demouliere B, Chenu F, Bourin M. Forced swimming test in mice: areview of antidepressant activity. Psychopharmacology 2005;177:245–55.

999K.R. Lobato et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 994–999
Phillis JW. Potentiation of the action of adenosine on cerebral cortical neuronesby the tricyclic antidepressants. Br J Pharmacol 1984;83:567–75.
Phillis JW,WuPH. The effect of various centrally active drugs on adenosine uptakeby the central nervous system. Comp Biochem Physiol 1982;72C:179–87.
Porsolt RD, Bertin A, Jalfre M. Behavioral despair in mice: a primary screeningtest for antidepressants. Arch Int Pharmacodyn Ther 1977;229:327–36.
Ribeiro JA, Sebastião AM, de Mendonça A. Adenosine receptors in the nervoussystem: pathophysiological implications. Prog Neurobiol 2003;68:377–92.
Rosa AO, Lin J, Calixto JB, Santos ARS, Rodrigues ALS. Involvement ofNMDA receptors and L-arginine-nitric oxide pathway in the antidepressant-like effects of zinc in mice. Behav Brain Res 2003;144:87–93.
Szewczyk B, Brański P, Wierońska JM, Palucha A, Pilc A, Nowak G. Inter-action of zinc with antidepressants in the mouse forced swimming test. Pol JPharmacol 2002;54:681–5.
Uzbay TI, Kayir H, Ceyhan M. Effects of tianeptine on onset time ofpentylenetetrazole-induced seizures in mice: possible role of adenosine A1
receptors. Neuropsychopharmacology 2007;32:412–6.Vallee BL, Falchuk KH. The biochemical basis of zinc physiology. Physiol Rev
1993;73:79–118.Vandenberg RJ, Mitrovic AD, Johnston GA. Molecular basis for differential
inhibition of glutamate transporter subtypes by zinc ions. Mol Pharmacol1998;54:189–96.
Zahorodna A, Bijak M, Hess G. Differential effects of repeated imipramine onhippocampal responsiveness to adenosine and serotonin. Eur Neuropsycho-pharmacol 2002;12:355–60.