in vitro immune functions in thiamine-replete and -depleted lake trout (salvelinus namaycush)
TRANSCRIPT

lable at ScienceDirect
Fish & Shellfish Immunology 38 (2014) 211e220
Contents lists avai
Fish & Shellfish Immunology
journal homepage: www.elsevier .com/locate / fs i
Full length article
In vitro immune functions in thiamine-replete and -depleted laketrout (Salvelinus namaycush)
Christopher A. Ottinger a,*, Dale C. Honeyfield b, Christine L. Densmore a,Luke R. Iwanowicz a
aU.S. Geological Survey, Leetown Science Center, National Fish Health Research Laboratory, Leetown, WV 25430, USAbU.S. Geological Survey, Leetown Science Center, Northern Appalachian Research Laboratory, Wellsboro, PA 16901, USA
a r t i c l e i n f o
Article history:Received 9 January 2014Received in revised form10 March 2014Accepted 15 March 2014Available online 27 March 2014
Keywords:ThiamineLake troutGreat lakesIn vitroOxidative stress
* Corresponding author.E-mail addresses: [email protected]
A. Ottinger).
http://dx.doi.org/10.1016/j.fsi.2014.03.0241050-4648/Published by Elsevier Ltd. This is an open
a b s t r a c t
In this study we examined the impacts of in vivo thiamine deficiency on lake trout leukocyte functionmeasured in vitro. When compared outside the context of individual-specific thiamine concentrations nosignificant differences were observed in leukocyte bactericidal activity or in concanavalin A (Con A), andphytohemagglutinin-P (PHA-P) stimulated leukocyte proliferation. Placing immune functions intocontext with the ratio of in vivo liver thiamine monophosphate (TMP e biologically inactive form) tothiamine pyrophosphate (TPP e biologically active form) proved to be the best indicator of thiaminedepletion impacts as determined using regression modeling. These observed relationships indicateddifferential effects on the immune measures with bactericidal activity exhibiting an inverse relationshipwith TMP to TPP ratios, Con A stimulated mitogenesis exhibiting a positive relationship with TMP to TPPratios and PHA-P stimulated mitogenesis exhibiting no significant relationships. In addition, these re-lationships showed considerable complexity which included the consistent observation of a thiamine-replete subgroup with characteristics similar to those seen in the leukocytes from thiamine-depletedfish. When considered together, our observations indicate that lake trout leukocytes experience cell-type specific impacts as well as an altered physiologic environment when confronted with athiamine-limited state.
Published by Elsevier Ltd. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/3.0/).
1. Introduction
Early mortality syndrome (EMS) results in embryonic mortalityin salmonids (coho salmon (Oncorhynchus kisutch), chinook salmon(Oncorhynchus tshawytscha), steelhead trout (Oncorhynchusmykiss), brown trout (Salmo trutta) and lake trout (Salvelinusnamaycush)). Clinical symptoms of EMS include loss of equilibrium,swimming in a spiral pattern, lethargy, hyper-excitability, andhemorrhage and death occurring between hatch and first feeding[1,2]. Stocks of Atlantic salmon (Salmo salar) from the Finger Lakesand the Baltic Sea exhibit a similar early life-stage mortality, calledCayuga syndrome [3] and M74 [4], respectively. Low egg thiaminelevels and enhanced survival following thiamine treatments arecommon characteristics of EMS, Cayuga Syndrome and M74. Inaddition to the clinical symptoms, visual detail discrimination and
, [email protected] (C.
access article under the CC BY-NC
motion detection in lake trout fed a low thiamine diet has beenshown to be significantly lower than seen in thiamine-replete laketrout fry [5] as has the ability of lake trout fry to avoid a piscinepredator and to forage on Daphnia [6]. Taken together, these find-ings support the suggestion made by Brown et al. [7] that thephysiologic impacts of thiamine deficiency may be far reaching.EMS is a result of thiamine deficiency in the sexually maturecomponent of wild lake trout populations with low egg thiamineconcentrations noted within hatcheries when wild fish are used asbrood stock [7] as well as in eggs obtained from natural spawningbeds [8]. Given that thiamine deficiency is occurring in adult laketrout, and that the physiologic impact could be far reaching, wepreviously have suggested that this deficiency could impact laketrout immune function and thus disease resistance [9].
Diet and nutritional status are known to affect immune functionand disease resistance in fish [10,11]. The immunomodulatory roleof specific vitamins and their effects on disease resistance in fisheshas been examined in some detail with particular attention paid toascorbic acid (vitamin C) [12e15] and the tocopherols (vitamin E)[16,17]. The effect of thiamine (vitamin B1) deficiency in lake trout
-ND license (http://creativecommons.org/licenses/by-nc-nd/3.0/).

C.A. Ottinger et al. / Fish & Shellfish Immunology 38 (2014) 211e220212
immune function has also received limited attention [9]. In thisstudy we continue this work by examining the impacts of in vivothiamine deficiency on lake trout leukocyte functions measuredin vitro and placed in context with in situ thiamine concentrations.We also examine the relationship between fish weight and thia-mine concentrations in an attempt to place our observed in vitroimmune functional-measures into context with broader physio-logic processes.
2. Materials and methods
2.1. Fish, diet, and culture conditions
Seneca strain lake trout were spawned, hatched and reared aspreviously described [9]. A minimum fish size of 200 g was selectedfor use in this study to provide ample tissue for cell isolation. Fishwere reared to approximately 100 g on commercial feed and thenrandomly divided into two groups. Thiamine deficiency was ach-ieved in the thiamine deficient group via delivery of a previouslydescribed casein bacterial thiaminase diet [9,18]. A thiamine defi-cient status was confirmed by skeletal muscle levels in treatmentsubsamples [19]. Once a thiamine-depleted state was confirmed,the fish were fed a similar diet without thiaminase and containing0.4 mg kg�1 feed of dietary thiamine [9,18] which was sufficient toprevent death while retaining the thiamine-depleted state.Thiamine-replete fish were fed through the entire study with apreviously described diet containing 2.0 mg kg�1 feed of dietarythiamine [9,18]. Other than the elevated thiamine concentrations,this feed was identical to that provide the thiamine-depleted fish.The base nutritional requirement for thiamine is 1.0 mg kg�1offeed [20]. In all diets the calculated crude protein content was 45.7%and the dietary lipid (fish oil) was 15%. Fish were fed daily tosatiation.
2.2. In vitro immune study
Immune function studies were conducted at the U.S. GeologicalSurvey, Leetown Science Center, Fish Health Branch (Kearneysville,WV, USA). During these studies the trout were maintained in anindoor flow-through system consisting of 900 L circular tankssupplied with 12 �C oxygenated spring water. Water flow into thetanks was maintained at rate of 10 total volume replacements perday. Tanks were illuminated by fluorescent lighting with photo-period adjusted tomatch local seasonal change. The lake trout wereacclimated to tank conditions for at least two months prior toinitiation of experimental protocols. No morbidity or mortality wasobserved during the acclimation or experimental periods.
Anterior kidney leukocytes from 36 thiamine-replete and 36thiamine-depleted lake trout were evaluated for lymphocytemitogenesis and for macrophage bactericidal activity. Prior to tissueharvests, fish were euthanized with an overdose of tricaine meth-anesulfonate (Finquel, Argent Chemical Laboratories, Redmond,WA) immediately upon removal from the tanks, weighed, and bledfrom the caudal vessels. Anterior kidney was collected using aseptictechnique and processed for leukocyte isolations as describedbelow. Liver samples were collected and immediately frozenat �80 �C for tissue thiamine determinations described below.Plasma was separated from whole blood by centrifugation at1000 RCF at 4 �C for 20 min and archived at �80 �C.
Lake trout anterior kidney leukocytes isolation and the leuko-cyte mitogenesis assay were performed as described by Gauthieret al. [21]. All mitogens were diluted in L-15 media containing 5%fetal bovine serum (FBS). The anterior kidney leukocytes at a con-centration of 2 � 107 cells ml�1 were plated at 50 ml well�1 andwere treated at 50 ml well�1 with either concanavalin A (Con A) at
10 mg ml�1, phytohemagglutinin-P (PHA-P) at 20 mg ml�1, poke-weed mitogen (PWM) at 50 mg ml�1, or mitogen-free media (con-trol wells). Mitogen treated and control wells for each thiamine-replete and -depleted fish were replicated in triplicate and werealways used in tandem in the same plate. Leukocytes were treatedwith 25 ml well�1 of sterile-filtered 65 mM bromodeoxyuridine(BrdU) in L-15 (BrdU final concentration¼ 13 mMwell�1), 18 h priorto the evaluation of mitogenesis by BrdU enzyme-linked immu-nosorbent assay (ELISA). Following incubation with BrdU, cellswere washed with 100 ml well�1 Dulbecco’s phosphate bufferedsaline (DPBS) and fixed for 15 min with 1% paraformaldehyde inDPBS (pH 7.2). Cells were washed as above and the leukocytemembranes were porated by a 30 s exposure to 50 ml well�1 of0.01% Tween 20 in DPBS. Cells were again washed and the wellswere then blocked with 250 ml well�1 of DPBS containing 1% bovineserum albumin (blocking buffer). Blocking buffer was replacedwithout washing by 50 ml well�1 of 0.2 U ml�1 anti-bromodeoxyuridine labeled with horseradish peroxidase (mAb,Fab) in blocking buffer and the plates were incubated for 1 h. Cellswere washed 5 times with 250 ml well�1 DPBS and 50 ml well�1 of asubstrate solution (4.8 ml 10 mM citrate buffer (pH 4) with 200 ml18.2 mM 2,20-azino-bis(3-ethylbenzthiazoline-6-sulphonic acid)(ABTS) and 5 ml 30% hydrogen peroxide) was added. The opticaldensity (405 nm) of the solution in each well was determined at5 min increments over a period of 20 min using a V-max KineticMicroplate Reader (Molecular Devices Corporation, Sunnyvale, CA)set for a 405 nm wave length.
Liver thiamine pyrophosphate (TPP), thiamine monophosphate(TMP), and free thiamine content were quantified using the high-performance liquid chromatography (HPLC) method described byBrown et al. [19]. Commercially available authentic standards(Sigma Chemical) were used to identify chromatographic peaks.Data reported are from duplicate analyses of each sample in whichthere was less than ten percent difference in value.
Bactericidal activity of adherent cells was evaluated with amodification of the methods described by Harms et al. [22]. Cellsfrom each fish in L-15/0.1% FBS were loaded as two sets of replicates(quadruplicate and triplicate) at 100 ml well�1 into a 96 well tissue-culture plate and incubated for 2 h. Twelvewells on each plateweretreated the same as the other wells except that cell-free media wasemployed (positive control). Following the incubation, media wasremoved from all wells and replaced with 100 ml well�1 L-15/5%FBS. Adherent cells, including the macrophages, were then incu-bated for another 36 h. Forty-eight hour room temperature culturesof Yersinia ruckeri (National Fish Health Research Laboratory, #11.40) in tryptic soy broth (Becton, Dickinson and Company, Sparks,MD, USA) were washed and suspended in Hanks balanced salt so-lution (HBSS) to a density of 0.05 OD (600 nm). Following incuba-tion, media was removed from the plates, and wells were washedwith 100 ml unsupplemented L-15. L-15 media with 5% FBS and noantibiotics was added at 100 ml well�1, followed by the addition ofthe Y. ruckeri suspension at 25 ml well�1 to well sets replicated inquadruplicate and the twelve-well sets containing no cells. Plateswere then incubated for 4 h, the media in the wells containingbacteria was removed and replaced with filter-sterilized 0.2%polyoxyethylene-sorbitan monolaurate (Tween 20) in tissue cul-ture grade water at 50 ml well�1. Ten-fold dilutions were made ofindividual well contents in tryptic soy broth, and 10 ml aliquots ofthe dilutions were cultured on tryptic soy agar (Becton, Dickinsonand Company). Culture plates were incubated 24 h at 22 �C, and thenumber of colony-forming units of the bacteria was determined foreach corresponding assay well. Following the plating of the bac-terial dilutions, the number of adherent cells in the wells replicatedin triplicate was determined. Bactericidal activity was expressed asthe % colony-forming units (CFU) reduction. The % CFU reduction
C.A. Ottinger et al. / Fish & Shellfish Immunology 38 (2014) 211e220 213
was defined as: ((1� (CFU treated/CFU control))� 100) where “CFUtreated” equaled the mean CFU value for replicate wells withadherent leukocytes and “CFU control” equaled themean CFU valuefor replicate wells with media only.
2.3. Data analysis
Data analysis was performed using Systat 11 (SYSTAT SoftwareInc., Richmond, CA) unless otherwise stated. Multiple measuresweremade from the tissue and anterior kidney leukocytes obtainedfrom each fish. Data sets from fish with missing values were notconsidered for analysis. Data for each treatment-specific measurewere evaluated for normality using the ShapiroeWilk NormalityTest. Statistical comparisons between thiamine-replete and-deplete measures were performed using the parametric test,Analysis of Variance (ANOVA), followed by Tukey post-hoc com-parisons (normally distributed data) or the nonparametric test,KruskaleWallis One-Way Analysis of Variance by Ranks (KW),followed by the ManneWhitney U-test for post-hoc comparisons(nonparametric data). For consistency, all paired data sets arepresented as box plots. The statistical methods employed in the
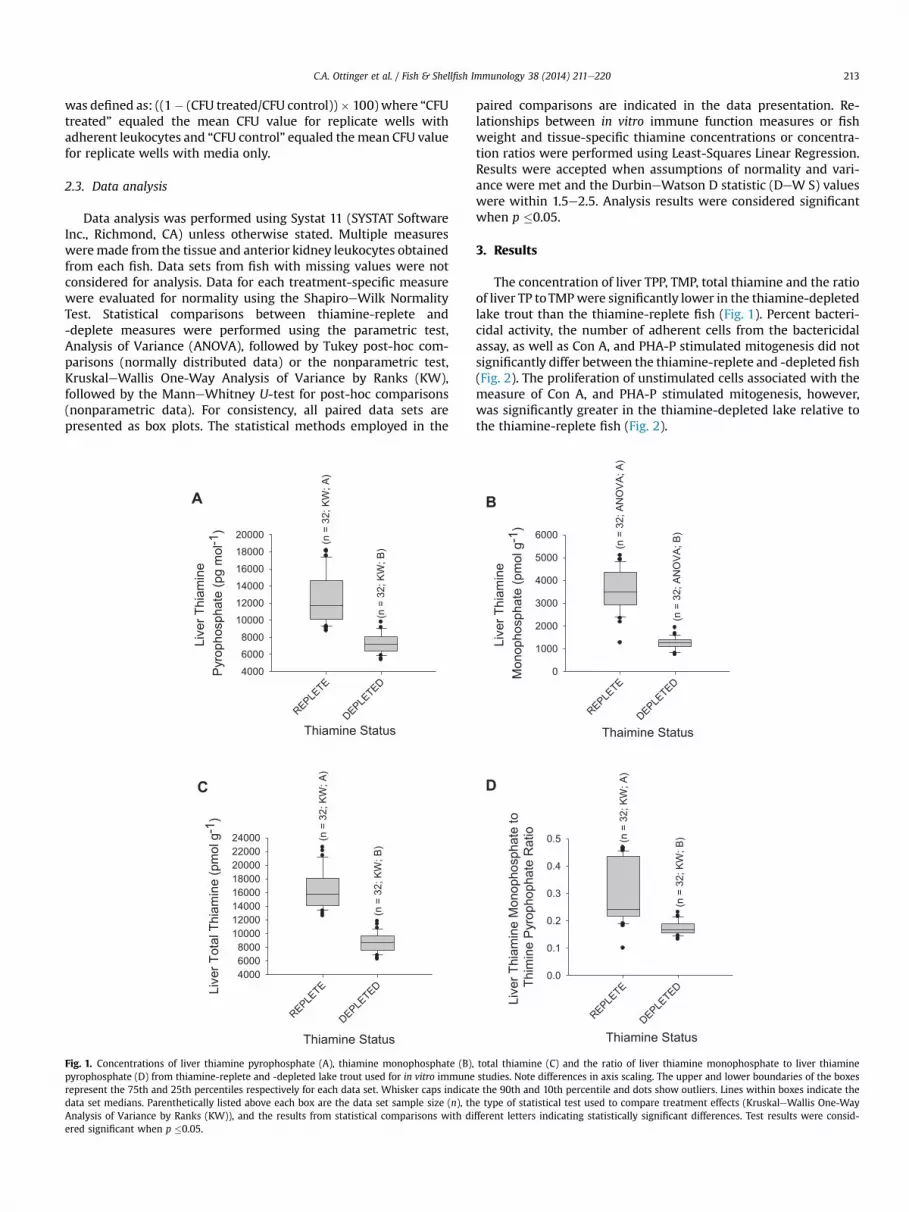
Fig. 1. Concentrations of liver thiamine pyrophosphate (A), thiamine monophosphate (B),pyrophosphate (D) from thiamine-replete and -depleted lake trout used for in vitro immunerepresent the 75th and 25th percentiles respectively for each data set. Whisker caps indicatdata set medians. Parenthetically listed above each box are the data set sample size (n), thAnalysis of Variance by Ranks (KW)), and the results from statistical comparisons with diered significant when p �0.05.
paired comparisons are indicated in the data presentation. Re-lationships between in vitro immune function measures or fishweight and tissue-specific thiamine concentrations or concentra-tion ratios were performed using Least-Squares Linear Regression.Results were accepted when assumptions of normality and vari-ance were met and the DurbineWatson D statistic (DeW S) valueswere within 1.5e2.5. Analysis results were considered significantwhen p �0.05.
3. Results
The concentration of liver TPP, TMP, total thiamine and the ratioof liver TP toTMPwere significantly lower in the thiamine-depletedlake trout than the thiamine-replete fish (Fig. 1). Percent bacteri-cidal activity, the number of adherent cells from the bactericidalassay, as well as Con A, and PHA-P stimulated mitogenesis did notsignificantly differ between the thiamine-replete and -depleted fish(Fig. 2). The proliferation of unstimulated cells associated with themeasure of Con A, and PHA-P stimulated mitogenesis, however,was significantly greater in the thiamine-depleted lake relative tothe thiamine-replete fish (Fig. 2).
total thiamine (C) and the ratio of liver thiamine monophosphate to liver thiaminestudies. Note differences in axis scaling. The upper and lower boundaries of the boxese the 90th and 10th percentile and dots show outliers. Lines within boxes indicate thee type of statistical test used to compare treatment effects (KruskaleWallis One-Wayfferent letters indicating statistically significant differences. Test results were consid-

C.A. Ottinger et al. / Fish & Shellfish Immunology 38 (2014) 211e220214
In thiamine-depleted lake trout weight was negatively relatedto the ratio of liver TMP to TPP (Fig. 3). No significant relationshipsbetween thiamine-replete fish weight and liver TMP to TPP ratioswere observed. When weights of depleted fish were compared tothose of thiamine-replete fish with similar liver TMP to TPP ratios(<0.25) and to those of thiamine-replete fishwith TMP toTPP ratioshigher than those in thiamine-depleted fish (>0.25) no significantdifferences were observed (Fig. 3).
A negative relationship was observed between bactericidal ac-tivity and liver TMP to TPP ratios in thiamine-depleted fish but notin the thiamine-replete fish (Fig. 4). Bactericidal activity inthiamine-replete fish with liver TMP to TPP ratios greater thanthose of the thiamine-depleted fish was significantly lower thanthat of the thiamine-depleted fish but no difference in bactericidalactivity was observed between the thiamine-replete and -depletedfish with similar TMP to TPP ratios or between the two thiamine-replete groups (Fig. 4). No relationships were observed betweenthe number of adherent cells associated with bactericidal activity
Fig. 2. In vitro percent bactericidal activity (A), adherent cell numbers (B), mitogen stimuladetermination of mitogen-specific cell proliferation (D) obtained for anterior kidney leukocyplots and level of statistical significance are as per Fig. 1. Parenthetically listed above each boeffects (Analysis of Variance (ANOVA) KruskaleWallis One-Way Analysis of Variance by Ranstatistically significant differences. Test results were considered significant when p �0.05Phytohemagglutinin type P; OD e Optical Density at 405 nm.
and liver TMP to TPP ratio in either the thiamine-replete or-depleted groups (Fig. 4).
Leukocytes from thiamine-replete fish exhibited significantpositive regression results when relationships between liver TMPto TPP ratios and Con A-associated mitogenesis (Fig. 5). No signif-icant relationships were observed between thiamine concentra-tions or ratios and Con A-associatedmitogenesis in leukocytes fromthiamine-depleted fish (Fig. 5) nor were there any potential rela-tionship observed between thiamine concentrations or ratios andPHA-P-associated mitogenesis in leukocytes from either thiamine-replete or -depleted trout (Fig. 5). Con A stimulated mitogenesisexhibited by cells from fishwith liver TMP toTPP ratios greater thanthose observed in the thiamine-depleted fish and thiamine-repletefish with ratios similar to the thiamine-depleted fish were signifi-cantly greater than those of the latter two groups which did notsignificantly differ (Fig. 5). Like Con A stimulated mitogenesis, theproliferation of unstimulated leucocytes from thiamine-replete fishused in the calculation of stimulation index values exhibited a
ted cell proliferation (C), and proliferation measures of unstimulated cells used in thetes from thiamine-replete and -depleted lake trout. Note differences in axis scaling. Boxx are the data set sample size (n), the type of statistical test used to compare treatmentks (KW)), and the results from statistical comparisons with different letters indicating. Abbreviations: REP e Replete; Dep e Depleted; CON A e Concanavalin A; PHA-P e

Fig. 3. Fish-specific relationships between total weight and the ratio of liver thiamine monophosphate to thiamine pyrophosphate in thiamine-replete (closed circle) and thiamine-depleted lake trout (open circle) used as cell sources for in vitro immunoassays. Linear regression results and sample size for the replete and thiamine-depleted treatment groupsshown in the scatter plot are provided above the graph. Inserted box plot provides comparisons between depleted fish, thiamine-replete fish with ratios similar to thiamine-depleted fish (ratio < 0.25), and thiamine-replete fish with ratios greater than thiamine-depleted fish (ratio > 0.25). Box plots and level of statistical significance are as perFig. 1. Statistical test associated with box plots are as per Fig. 2.
C.A. Ottinger et al. / Fish & Shellfish Immunology 38 (2014) 211e220 215
positive relationship with liver TMP to TPP ratios (Fig. 6). The samecomparison for thiamine-depleted fish was not significant. Whenthe optical density values representing the proliferation of unsti-mulated leucocytes from the thiamine-depleted fish werecompared to those of thiamine-replete fish with similar TMP to TPPratios and those with higher ratios, the thiamine-replete fish withTMP to TPP ratios similar to the thiamine-depleted fish exhibitedsignificantly lower optical density values than the thiamine-depleted fish and were also significantly lower than those fromthe thiamine-replete fish with elevated ratios (Fig. 6). The lattertwo groups were not significantly different.
4. Discussion
The relationship between in vivo thiamine concentrations andleukocyte function persists when the cells are removed andcultured in a thiamine-replete environment. How long these re-lationships may last is not known, however, based on our currentfinding we can conclude that the relationships will last for at least48 h. A central component of our observations is the associationsbetween in vitro immune function and in vivo thiamine concen-trations. Male rats fed a thiamine deficient diet exhibited reducedblood TMP concentrations relative to TPP [23] which is consistent
with our observation of reductions of liver TMP to TPP ratios inthiamine-depleted relative to thiamine-replete lake trout (Fig. 1).When in vitro immune functions in thiamine-replete and thiamine-depleted lake trout were compared outside the context ofindividual-specific thiamine concentrations significant differenceswere only observed in the proliferation of untreated cells (Fig. 2).Not until the functional measures were placed in context with liverTMP to TPP ratios did differences emerge (Figs. 3e6). One funda-mental difference observed between the thiamine-replete andthiamine-depleted lake trout was the relatively high heterogeneityin TMP to TPP ratios exhibited by the thiamine-replete fish (Fig. 1).Observed differences did not clearly discriminate between thethiamine-replete and thiamine-depleted fish when fish weight andleukocyte functions were expressed relative to TMP to TPP ratios.Instead, the relationships between the TMP toTPP ratios and in vitroimmune measures split the thiamine-replete group into two seg-ments with one thiamine-replete subgroup exhibiting character-istics very similar to the thiamine-depleted fish. Although thisstudy was not designed to evaluate diet effect on weight of thia-mine-depleted fish (Fig. 3), weight was associatedwith liver TMP toTPP ratios suggesting that the mechanisms involved in the regu-lation of thiamine use and allocation may have broad physiologicalimpacts including howmacronutrients are transformed and stored.

Fig. 4. Fish-specific relationships between bactericidal activity (A), the number of adherent cells associated with observed bactericidal activity and the ratio of liver thiaminemonophosphate to thiamine pyrophosphate in thiamine-replete (closed circle) and thiamine-depleted lake trout (open circle) used as cell sources for in vitro immunoassays. Scatterand box plots as described in Fig. 3.
C.A. Ottinger et al. / Fish & Shellfish Immunology 38 (2014) 211e220216
The teleost dietary recommendations for thiamine provided by theNational Research Council [20] were established based on generalperformance and not specific pathway metabolic requirements.Situation-dependent thiamine metabolic requirements likely varybetween individuals potentially placing some individuals into pe-riodic states resembling thiamine depletion. Such an event wouldexplain our observation of low liver TMP to TPP ratios in some fishwithin the thiamine-replete group. The significant reductions inthiamine tissue concentrations and ratios (Fig. 1) in the thiamine-depleted group are the expected outcome of dietary treatmentsemployed and a mandatory component of our study. Offering anexplanation, however, as to how in vitro immune measures wouldassociate with liver thiamine concentration is a major challenge.Even in mammals, basic studies on the specific roles of thiamine inimmune function are lacking. The potential relationships betweenthiamine and lake trout immune function are extensive andcomplex.
The principal form of thiamine in tissues is TPP (also known asthiamine diphosphate). The role of TPP as a cofactor in energymetabolism and fatty acid synthesis is well described in a review by
Depeint et al. [24] and only salient points will be provided here.Withinmitochondria, TPP is a required cofactor for the tricarboxylicacid cycle enzyme complex alpha-ketoglutarate dehydrogenase,and for the related tricarboxylic acid cycle-associated enzymecomplexes pyruvate dehydrogenase and branched-chain ketoaciddehydrogenase all of which catalyze the oxidative decarboxylationof ketoacids. Thiamine pyrophosphate is also a cofactor for cyto-plasmic transketolase that catalyzes the first and final steps in thepentose phosphate pathway resulting in the production of nico-tinamide adenine dinucleotide phosphate (NADPH) which main-tains cellular reductioneoxidation (redox) condition, glutathionelevels, and protein sulphhydryl groups. NADPH is a required factorfor all nitrous oxide synthases (NOS) [25], NADPH oxidases (NOXfamily) [25] and for microsomal fatty acid elongation [26]. In ste-roidogenesis, NADPH is required for both Type I (mitochondrial)and Type II (cytoplasmic) cytochrome P450 associated synthesis[27]. Thiamine depletion has been shown to significantly reducethe activity of enzymes in Baltic salmon (S. salar) sac fry includingtransketolase, alpha-ketoglutarate dehydrogenase, glucose-6-phosphate dehydrogenase, and cytochrome p450 1A [28].

Fig. 5. Fish-specific relationships between concanavalin A (Con A; A) stimulated mitogenesis, phytohemagglutinin type P (PHA-P; B) stimulated mitogenesis and the ratio of liverthiamine monophosphate to thiamine pyrophosphate in thiamine-replete (closed circle) and thiamine-depleted lake trout (open circle) used as cell sources for in vitro immu-noassays. Scatter and box plots as described in Fig. 3.
C.A. Ottinger et al. / Fish & Shellfish Immunology 38 (2014) 211e220 217
Thiamine monophosphate is known to be a hydrolysis productderived from TPP and has no known physiologic function. A prin-cipal effect of thiamine depletion is oxidative stress.
The relationship between oxidative stress, thiamine depletion,and immune function has been reported inmammals in associationwith brain pathology [29]. Associations between oxidative stressand thiamine depletion have also been reported in fish [28,30,31].Like mammals, thiamine depletion is associated with tissue pa-thology [32e34] and altered brain function [35]. Diseases inhumans associated with thiamine deficiency have been linked topoorly regulated inflammatory responses signaled through toll-likereceptors (TLR) [29,36e39] which in turn can be linked todemonstrated differences in energy metabolism occurring inmammalian macrophages as well as some T-cells. Redox states areknown to impact and be impacted by cellular metabolic conditionswhich in turn impact cell proliferation and differentiation in a cell-type dependent manner [40]. Mammalian pro-inflammatory M1macrophages and T-helper subtype 17 (TH17) cells primarily pro-duce energy via glycolysis whereas anti-inflammatory M2
macrophages and T-regulatory (Treg) cells produce energy primarilyvia oxidative phosphorylation [41]. Given that thiamine deficiencyand the associated oxidative stress induce suppression of oxidativephosphorylation, a thiamine-limited condition should favor TH17and M1 macrophages phenotypes thus favoring a pro-inflammatory regulatory environment and elevated bactericidalactivity. The hypoxia inducible transcription factor-alpha (HIF1-a)-dependent glycolytic pathway regulates the differentiation ofTprogenitor-cells into TH17 or Treg phenotypes [42,43] and is essentialfor the production of some pro-inflammatory cytokines in M1macrophages [41]. Hypoxia inducible transcription factor-alpha hasalso been directly associated with pathology and mortalityobserved in thiamine-depleted Baltic salmon [44] yolk-sac frywhich gives some indication that the HIF1-a-dependent glycolyticpathway could have a regulatory effect in lake trout Tprogenitor-likecells and macrophages similar to that observed in mammals.
Teleosts possess many of the same antimicrobial mechanismsobserved in mammals [45] indicating the presence of a robustinnate immune system [45e47]. A diverse group of TLRs have been
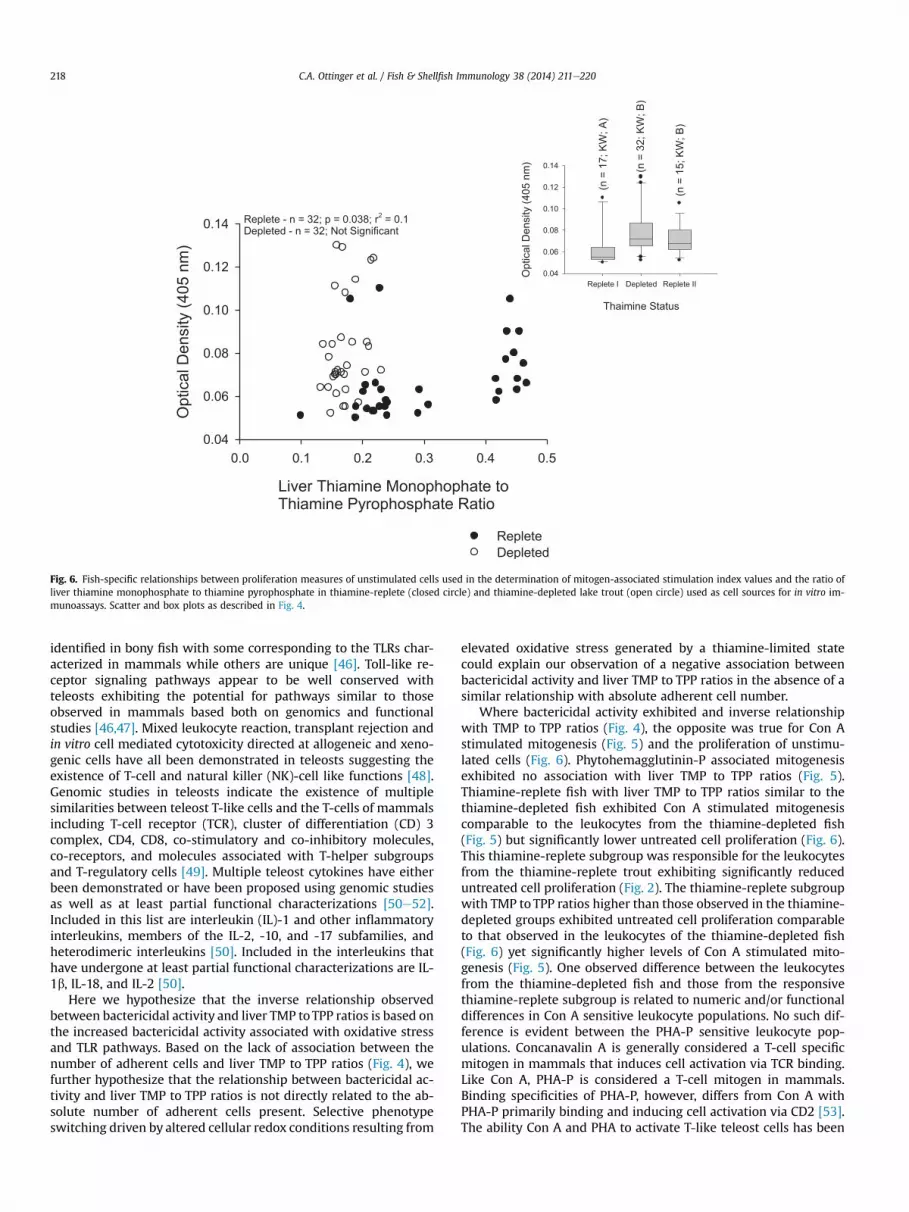
Fig. 6. Fish-specific relationships between proliferation measures of unstimulated cells used in the determination of mitogen-associated stimulation index values and the ratio ofliver thiamine monophosphate to thiamine pyrophosphate in thiamine-replete (closed circle) and thiamine-depleted lake trout (open circle) used as cell sources for in vitro im-munoassays. Scatter and box plots as described in Fig. 4.
C.A. Ottinger et al. / Fish & Shellfish Immunology 38 (2014) 211e220218
identified in bony fish with some corresponding to the TLRs char-acterized in mammals while others are unique [46]. Toll-like re-ceptor signaling pathways appear to be well conserved withteleosts exhibiting the potential for pathways similar to thoseobserved in mammals based both on genomics and functionalstudies [46,47]. Mixed leukocyte reaction, transplant rejection andin vitro cell mediated cytotoxicity directed at allogeneic and xeno-genic cells have all been demonstrated in teleosts suggesting theexistence of T-cell and natural killer (NK)-cell like functions [48].Genomic studies in teleosts indicate the existence of multiplesimilarities between teleost T-like cells and the T-cells of mammalsincluding T-cell receptor (TCR), cluster of differentiation (CD) 3complex, CD4, CD8, co-stimulatory and co-inhibitory molecules,co-receptors, and molecules associated with T-helper subgroupsand T-regulatory cells [49]. Multiple teleost cytokines have eitherbeen demonstrated or have been proposed using genomic studiesas well as at least partial functional characterizations [50e52].Included in this list are interleukin (IL)-1 and other inflammatoryinterleukins, members of the IL-2, -10, and -17 subfamilies, andheterodimeric interleukins [50]. Included in the interleukins thathave undergone at least partial functional characterizations are IL-1b, IL-18, and IL-2 [50].
Here we hypothesize that the inverse relationship observedbetween bactericidal activity and liver TMP toTPP ratios is based onthe increased bactericidal activity associated with oxidative stressand TLR pathways. Based on the lack of association between thenumber of adherent cells and liver TMP to TPP ratios (Fig. 4), wefurther hypothesize that the relationship between bactericidal ac-tivity and liver TMP to TPP ratios is not directly related to the ab-solute number of adherent cells present. Selective phenotypeswitching driven by altered cellular redox conditions resulting from
elevated oxidative stress generated by a thiamine-limited statecould explain our observation of a negative association betweenbactericidal activity and liver TMP to TPP ratios in the absence of asimilar relationship with absolute adherent cell number.
Where bactericidal activity exhibited and inverse relationshipwith TMP to TPP ratios (Fig. 4), the opposite was true for Con Astimulated mitogenesis (Fig. 5) and the proliferation of unstimu-lated cells (Fig. 6). Phytohemagglutinin-P associated mitogenesisexhibited no association with liver TMP to TPP ratios (Fig. 5).Thiamine-replete fish with liver TMP to TPP ratios similar to thethiamine-depleted fish exhibited Con A stimulated mitogenesiscomparable to the leukocytes from the thiamine-depleted fish(Fig. 5) but significantly lower untreated cell proliferation (Fig. 6).This thiamine-replete subgroup was responsible for the leukocytesfrom the thiamine-replete trout exhibiting significantly reduceduntreated cell proliferation (Fig. 2). The thiamine-replete subgroupwith TMP to TPP ratios higher than those observed in the thiamine-depleted groups exhibited untreated cell proliferation comparableto that observed in the leukocytes of the thiamine-depleted fish(Fig. 6) yet significantly higher levels of Con A stimulated mito-genesis (Fig. 5). One observed difference between the leukocytesfrom the thiamine-depleted fish and those from the responsivethiamine-replete subgroup is related to numeric and/or functionaldifferences in Con A sensitive leukocyte populations. No such dif-ference is evident between the PHA-P sensitive leukocyte pop-ulations. Concanavalin A is generally considered a T-cell specificmitogen in mammals that induces cell activation via TCR binding.Like Con A, PHA-P is considered a T-cell mitogen in mammals.Binding specificities of PHA-P, however, differs from Con A withPHA-P primarily binding and inducing cell activation via CD2 [53].The ability Con A and PHA to activate T-like teleost cells has been

C.A. Ottinger et al. / Fish & Shellfish Immunology 38 (2014) 211e220 219
demonstrated in studies involving a broad spectrum of teleostspecies [54e58]. The signaling pathway activated by Con A and PHAbinding, however, has not been specifically identified. The simi-larities between mammal and teleost TCR [48,49] suggest that theCon A-associated activation mechanisms may be similar in the twogroups. A CD2-like molecule, however, has yet to be identified inteleosts making the identification of a PHA-P dependent activationpathway difficult. Our observations demonstrating that in vitroimmune measures associate with in vivo thiamine concentrationsindicate that in vivo conditions were impacting in vitro responses.In vitro responses were measured in a thiamine-replete cultureenvironment suggesting that the impact of in vivo condition couldnot be rapidly reversed by simply increasing thiamine availability.Given that redox states are altered when thiamine is limiting andthat redox state impacts cell proliferation in a cell-type dependentmanner [40], we hypothesize that lake trout T-like cells have dif-ferential sensitivities to thiamine-limited conditions and that thedifferential sensitivities are dependent on activation pathway-specific metabolic requirements that may or may not be metwhen thiamine-limiting conditions alters cell metabolism.
In a previous study, using the same cohort of thiamine-repleteand thiamine-depleted lake trout as used here, we demonstratedthat thiamine-deficiency impacted in vivo hapten-specific antibodyproduction [9]. In this study, thiamine-depleted lake trout injectedwith the T-independent antigen trinitrophenol-lipopolysaccharideexhibited significant elevations in hapten-specific antibody activityand a prolonged response duration relative to thiamine-repletefish. In this same study, a parallel comparison using the T-depen-dent antigen trinitrophenol-keyhole limpet hemocyanin was per-formed and no such differences were observed. Placing the resultsof this current study in the context of this in vivo study we hy-pothesize that the deleterious oxidative stress resulting fromthiamine depletion in lake trout results in cell-type specific redoxenvironments that create a predisposition towards an inflamma-tory response. This predisposition is made apparent when leuko-cyte activation occurs following stimulation with a strong pro-inflammatory antigen such as bacterial lipopolysaccharide. Pre-disposition towards an inflammatory response, however, does notimply inhibition of all other immune response mechanisms sincethe observed in vivoT-dependent responsewas not impacted by thethiamine-deficient state. Like the response difference observed inCon A and PHA-P stimulated mitogenesis relative to liver TMP toTPP ratios, our previously reported difference in T-dependent and-independent antibody responses could in-part be explained basedon cell-type specific responses to altered redox states.
Altered redox states provide a useful hypothesis for events thatmay be impacting individual cell-types but it does not necessarilyprovide a complete description of all our observations. The differ-ences we observed in thiamine-replete and -depleted leukocyteunstimulated replication at similar TMP to TPP ratios combinedwith similar Con A stimulation values at these same ratios, how-ever, suggests that other mechanisms may be involved. A morecomplete explanation that accounts for other pathways throughwhich a thiamine-limited state could alter lake trout immunefunction requires a broader consideration of physiologic changeassociated with thiamine deficiency. Although the immune mea-sures and thiamine availability maybe functionally associated usingan oxidative stress model, the full physiologic impact of thiaminedeficiency on lake trout immune responses has the potential ofbeing far more complex with altered redox states associated withoxidative stress being a one potential modulator among several.Our observation that fish weight can be associated with liver TMPto TPP ratios and the demonstrated role of thiamine deficiency indiseases like early mortality syndrome clearly indicated that athiamine-limited state can have broad physiologic effects. The
potential that these broader physiologic effects could also impactimmune pathways must be considered. Thiamine availability andimmune function could be associated through shared physiologicpathway(s) adding a less direct relationship between the two. Forexample, changes in immune-relevant steroid synthesis resultingfrom NADPH depletion [27] alone or in combination with alteredneuronal activity associated with reduced thiamine availability [59]could contribute, along with detrimental oxidative stress, to alteredimmune function. We have previously suggested [9] that the im-pacts of the neuroendocrine axis on cytokine production wouldmake a plausible target for future studies evaluating thiamine andimmune function. Here we modify this statement by suggestingthat to obtain a better understanding of the alterations of immunefunctions associated with thiamine availability and thiamineassociatedmetabolism a broadmultifaceted-approachwill likely berequired.
Disclaimer
Any use of trade, product, or firm names is for descriptive pur-poses only and does not imply endorsement by the U.S.Government.
Acknowledgments
We thank The Great Lakes Fisheries Commission for theirfinancial support of this project and Mr. William Ridge for hisassistance with feed preparation and animal care.
References
[1] McDonald G, Fitzsimons J, Honeyfield DC, editors. Early life stage mortalitysyndrome in fishes of the Great Lakes and Baltic Sea, Symposium 21.Bethesda: American Fisheries Society; 1998.
[2] Marquenski SV, Brown SB. Early mortality syndrome in the Great Lakes. In:Rolland RM, Gilbertson M, Peterson RE, editors. Chemically induced alter-ations in functional development and reproduction of fishes. Pensacola:SETAC Press; 1997.
[3] Fisher JP, Brown SB, Wooster GW, Bowser PR. Maternal blood, egg and larvalthiamin levels correlate with larval survival in landlocked Atlantic salmon(Salmo salar). J Nutr 1996;128:2456e66.
[4] Börjeson H, Norrgren L. M74 syndrome: a review of potential etiologicalfactors. In: Rolland RM, Gilbertson M, Peterson RE, editors. Chemicallyinduced alterations in functional development and reproduction of fishes.Pensacola: SETAC Press; 1997.
[5] Carvalho PSM, Tillitt DE, Zajicek JL, Claunch RC, Honeyfield DC, Fitzsimons JD,et al. Thiamine deficiency effects on the vision and foraging ability of laketrout fry. J Aquat Anim Health 2009;21:315e25.
[6] Fitzsimons JD, Brown SB, Williston B, Williston G, Brown LR, Moore K, et al.Influence of thiamine deficiency on lake trout larval growth, foraging, andpredator avoidance. J Aquat Anim Health 2009;21:302e14.
[7] Brown SB, Honeyfield DC, Hnath JG, Wolgamood M, Marcquenski SV,Fitzsimons JD, et al. Thiamine status in adult salmonines in the Great Lakes. JAquat Anim Health 2005;17:59e64.
[8] Czesny SJ, Rinchard J, Lee B-J, Dabrowski JM, Cao Y. Does spatial variation inegg thiamine and fatty-acid concentration of Lake Michigan lake trout Sal-velinus namaycush lead to differential early mortality syndrome and yolkoedema mortality in offspring? J Fish Biol 2012;80:2475e93.
[9] Ottinger CA, Honeyfield DC, Densmore CL, Iwanowicz LR. Impact of thiaminedeficiency on T-cell dependent and T-cell independent antibody production inLake Trout (Salvelinus namaycush). J Aquat Anim Health 2012;24:258e73.
[10] Balfry SK, Higgs DA. Influence of dietary lipid composition on the immunesystem and disease resistance of fin fish. In: Lim C, Webster CD, editors.Nutrition and fish health. New York: Haworth Press Inc; 2001.
[11] Lall SP. Nutrition and health of fish. In: Cruz-Suarez LE, Ricque-Marie D, Tapia-Salazar M, Olvera-Novoa MA, Civera-Cerecedo R, editors. Avances en Nutri-cion Acuicola V; 2000. Yucatan, Merida.
[12] Tewary A, Patra BC. Use of vitamin C as an immunostimulant. Effect ongrowth, nutritional quality, and immune response of Labeo rohita (Ham.). FishPhysiol Biochem 2008;34:251e9.
[13] Sahoo PK, Mukherjee SC. Immunomodulation by dietary vitamin C in healthyand aflatoxin B1-induced immunocompromised rohu (Labeo rohita). CompImmunol Microbiol Infect Dis 2003;26:65e76.
[14] Dunier M, Vergnet C, Siwicki AK, Verlhac V. Effect of lindane exposure onrainbow trout (Oncorhynchus mykiss) immunity. IV. Prevention of nonspecific

C.A. Ottinger et al. / Fish & Shellfish Immunology 38 (2014) 211e220220
and specific immunosuppression by dietary vitamin C (ascorbate-2-polyphosphate). Ecotoxicol Environ Saf 1995;30:259e68.
[15] Waagbø R, Sandnes K, Glette J, Nilsen ER, Albrektsen S. Dietary vitamin B6 andvitamin C. Influence on immune response and disease resistance in Atlanticsalmon (Salmo salar). Ann N Y Acad Sci 1992;669:379e82.
[16] Puangkaew J, Kiron V, Somamoto T, Okamoto N, Satoh S, Takeuchi T, et al.Nonspecific immune response of rainbow trout (Oncorhynchus mykiss Wall-baum) in relation to different status of vitamin E and highly unsaturated fattyacids. Fish Shellfish Immunol 2004;16:25e39.
[17] McCoy MA, McLoughlin MF, Rice DA, Kennedy DG. Pancreas disease inAtlantic salmon (Salmo salar) and vitamin E supplementation. Comp BiochemPhysiol A 1994;109:905e12.
[18] Honeyfield DC, Hinterkopf JP, Fitzsimons JD, Tillitt DE, Zajicek JL, Brown SB.Development of thiamine deficiencies and early mortality syndrome in laketrout by feeding experimental and feral fish diets containing thiaminase.J Aquat Anim Health 2005;17:4e12.
[19] Brown S, Honeyfield DC, Vandenbyllaardt L. Thiamine analysis in fish egg andtissues by high-pressure liquid chromatography. In: McDonald G, Fitzsimons J,Honeyfield DC, editors. Early life stage mortality syndrome in fishes of theGreat Lakes and Baltic Sea, Symposium 21. Bethesda: American FisheriesSociety; 1998.
[20] National Research Council. Nutrient requirements of fish and shrimp. Com-mittee on Animal Nutrition. Washington DC: National Academies Press; 2011.
[21] Gauthier DT, Cartwright DD, Densmore CL, Blazer VS, Ottinger CA. Measure-ment of in vitro mitogenesis in fish: ELISA based detection of the thymidineanalogue 50-bromo-20-deoxyuridine. Fish Shellfish Immunol 2003;14:279e88.
[22] Harms CA, Ottinger CA, Kennedy-Stoskopf S. Correlation of transforminggrowth factor-b (TGF-b mRNA) expression with cellular immunoassays intriamcinolone-treated captive hybrid striped bass. J Aquat Anim Health2000;12:9e17.
[23] Molina PE, Fan J, Boxer R, Gelato MC, Lang CH, Abumrad NN. Modulation ofinsulin-like growth factor-I: a specific role for vitamin B-1 (thiamine). J NutrBiochem 1996;4:207e13.
[24] Depeint F, Bruce WR, Shangari N, Mehta R, O’Brien PJ. Mitochondrial functionand toxicity: role of the B vitamin family on mitochondrial energy meta-bolism. Chem Biol Interact 2006;163:94e112.
[25] Stuehr DJ. Mammalian nitric oxide synthases. Biochim Biophys 1999;1411:217e30;Tretter L, Adam-Vizi V. Generation of reactive oxygen species in the reactioncatalyzed by a-ketoglutarate dehydrogenase. J Neurosci 2004;24:7771e8.
[26] Jump DB. Mammalian fatty acid elongases. Methods Mol Biol 2009;579:375e89.
[27] Miller WL. Minireview: regulation of steroidogenesis by electron transfer.Endocrinology 2005;146:2544e50.
[28] Amcoff P, Akerman G, Borjeson H, Tjarnlund U, Norrgren L, Balk L. Hepaticactivities othiamine-dependent enzymes, glucose-6-phosphate dehydroge-nase and cytochrome p4501a in Baltic salmon (Salmo salar) yolk-sac fry afterthiamine treatment. Aquat Toxicol 2000;48:391e402.
[29] Hazell AS, Butterworth RF. Update of cell damage mechanisms in thiaminedeficiency: focus on oxidative stress, excitotoxicity, and inflammation.Alcohol Alcohol 2009;44:141e7.
[30] Lundstrom J, Carney B, Amcoff P, Borjeson H, Forlin L, Norrgren L. Anti-oxidative systems, detoxifying enzymes and thiamine levels in Baltic salmon(Salmo salar) that develop M74. AMBIO 1999;28:24e9.
[31] Vuori KAM, Nikinmaa M. M74 syndrome in Baltic salmon and the possible roleof oxidative stresses in its development: present knowledge and perspectivesfor future studies. AMBIO 2007;36:168e72.
[32] Lundstrom J, Borjeson H, Norrgren L. Ultrastructural pathology of Balticsalmon, Salmo salar, L., yolk-sac fry with M74 syndrome. J Fish Dis 2002;25:143e54.
[33] Fitzsimons JD, Ketola G, Wooster GW, Brown SB. Use of a thiamine antagonistto induce Cayuga-syndrome-like mortalities in larval Atlantic salmon. J AquatAnim Health 2001;13:151e7.
[34] Lundstrum J, Borjeson J, Norrgren L. Histopathological studies of yolk-sac fryof Baltic salmon (Salmo salar) with M74 syndrome. AMBIO 1999;28:16e23.
[35] Amcoff P, Elofsson UOE, Borjeson J, Norrgren L, Nilsson GE. Alterations ofdopaminergic and serotondergic activity in the brain of sea-run Baltic salmonsuffering a thiamine deficiency-related disorder. J Fish Biol 2002;60:1407e16.
[36] Shi Q, Karuppagounder SS, Xu H, Pechman D, Chen H, Gibson GE. Responses ofthe mitochondrial alpha-ketoglutarate dehydrogenase complex to thiaminedeficiency may contribute to regional selective vulnerability. Neurochem Int2007;50:921e31.
[37] Karuppagounder SS, Shi Q, Xu H, Gibson GE. Changes in inflammatory processassociated with selective vulnerability following mild impairment of oxidativemetabolism. Neurobiol Dis 2007;26:353e62.
[38] Gill R, Tsung A, Billian TR. Linking oxidative stress to inflammation: toll-likereceptors. Free Radic Biol Med 2010;48:1121e32.
[39] Mkaddem SB, Bens M, Vandewalle A. Differential activation of Toll-like re-ceptor-mediated apoptosis induced by hypoxia. Oncotarget 2010;1:741e50.
[40] Agathocleous M, Harris WA. Metabolism in physiological cell proliferation anddifferentiation. Trends Cell Biol 2013;23:484e92.
[41] O’Neill LAJ, Hardie DG. Metabolism of inflammation limited by AMPK andpseudo-starvation. Nature 2013;493:346e55.
[42] Shi LZ, Wang R, Huang G, Vogel P, Neale G, Green DR, et al. H1F1a-dependentglycolytic pathway orchestrates a metabolic checkpoint for the differentiationof TH17 and Treg cells. J Exp Med 2011;208:1367e76.
[43] Dang EV, Barbi J, Yang H-Y, Jinasena D, Yu H, Zheng Y, et al. Control of TH17/Treg balance by hypoxia-inducible factor 1. Cell 2011;146:772e84.
[44] Vuori KAM, Soitamo A, Vuorinen PJ, Nikinmaa M. Baltic salmon (Salmo salar)yolk-sac fry mortality is associated with disturbances in the function ofhypoxia-inducible transcription factor (HIF-1 alpha) and consecutive geneexpression. Aquat Toxicol 2004;68:301e13.
[45] Rieger AM, Barreda DR. Antimicrobial mechanisms of fish leukocytes. DevComp Immunol 2011;35:1238e45.
[46] Palti Y. Toll-like receptors in bony fish: from genomics to function. Dev CompImmunol 2011;35:1263e72.
[47] Purcell MK, Smith KD, Hood L, Winton JR, Roach JC. Conservation of Toll-likereceptor signaling pathways in teleost fish. Comp Biochem Physiol D Geno-mics Proteomics 2006;1:77e88.
[48] Fischer U, Koppang EO, Nakanishi T. Teleost T and NK cell immunity. FishShellfish Immunol 2013;35:197e206.
[49] Laing KJ, Hansen JD. Fish T cell: recent advance through genomics. Dev CompImmunol 2011;35:1282e95.
[50] Secombes CJ, Wang T, Bird S. The interleukins of fish. Dev Comp Immunol2011;35:1336e45.
[51] Wiens GD, Glenney GW. Origin and evolution of TNF and TNF receptor su-perfamilies. Dev Comp Immunol 2011;35:1324e35.
[52] Zou J, Secombes CJ. Teleost fish interferons and their role in immunity. DevComp Immunol 2011;35:1376e87.
[53] Dumont C, Déas O, Mollereau B, Hebib C, Giovino-Barry V, Bernard A, et al.Potent apoptotic signaling and subsequent unresponsiveness induced by asingle CD2 mAb (BTI-322) in activated human peripheral T cells. J Immunol1998;160:3797e804.
[54] Kono T, Korenaga H. Cytokine gene expression in CD4 positive cells of theJapanese pufferfish, Takifugu rubripes. PLoS One 2013;8:e66364. http://dx.doi.org/10.1371/journal.pone.066364.
[55] Edholm E-S, Safford JL, Quiniou SM, Waldbieser G, Miller NW, Bengtén E, et al.Channel catfish, Ictalurus punctatus, CD4-like molecules. Dev Comp Immunol2007;31:172e87.
[56] Park H, Zhou H, Bengten E, Wilson M, Chinchar VG, Clem LW, et al. Activationof channel catfish (Ictalurus punctatus) T cells involves NFAT-like transcriptionfactors. Dev Comp Immunol 2002;26:775e84.
[57] Partula S. Surface markers of fish T-cell. Fish Shellfish Immunol 1999;9:241e57.
[58] Siegl E, Nebe B, Blunk H, Rychly J. detection of mitogen induced stimulation ofleukocytes from the rainbow trout (Oncorhynchus mykiss) by flow cytometricanalysis of intracellular calcium. Comp Biochem Physiol A 1998;119:915e23.
[59] Bettendorf L, Wins P. Thiamin diphosphate in biological chemistry: new as-pects of thiamin metabolism, especially triphosphate derivatives acting otherthan as cofactors. FEBS J 2009;276:2917e25.